
Isolation and prevalidation of an Escherichia coli tester strain for the use in mechanistic and...
11
ELSEVIER Mutation Research 312 (1994) 99-109 EnvironmentalMutagenesis Isolation and prevalidation of an Escherichia coli tester strain for the use in mechanistic and metabolic studies of genotoxins * Michel Kranendonk a, Margarida Ruas a, Antdnio Laires a,b, Jos6 Rueff a'* a Department of Genetics, Faculty of Medical Sciences, UNL, R. de Junqueira 96, P-1300Lisbon, Portugal, b Faculty of Sciences and Technology, UNL, Monte de Caparica, Portugal Received 10 September 1993;revision received 16 November 1993;accepted 19 November 1993 Abstract We have isolated an Escherichia coli tester strain for the use in mechanistic and metabolic studies of genotoxins. We started with one of the more used and better characterized E. coli K-12 laboratory strains, ABl157. We isolated a lipopolysaccharide defective mutant of strain AB1886 which is an excision repair deficient derivative of ABl157 and introduced a newly constructed plasmid pKRll, encoding mucAB, resulting in strain MR2101/pKRll. A genotoxicity assay was designed, monitoring the reversion to arginine prototrophy and a preliminary validation was carried out against Ames tester strain TA100 with a set of diagnostic compounds. The results seem to indicate that strain MR2101/pKRll is an adequate tester strain which can be a useful tool in mechanistic studies. Moreover, this strain can serve as mother strain to isolate improved and more specialized tester strains. Key words: E. coli; Tester strain 1. Introduction Despite the apparent simplicity compared with eukaryotic organisms, bacteria possess elaborate mechanisms for responding to DNA damage (Walker, 1984; Sedgwick and Goodwin, 1985; Sedgwick et al., 1991a). Bacteria are therefore widely used in fundamental studies of mecha- nisms involved in the biological response to DNA damage and in short-term genotoxicity tests. In the last decades various bacterial short-term as- * Corresponding author. * Dedicated to the memoryof Professor F.H. Sobels. says have been developed for the detection of chemical carcinogens as mutagens, e.g., the Ames test (Maron and Ames, 1983), WP2 test (Venitt et al., 1984), Inductest (Moureau et al., 1976), E. coli K12/343/113 test (Mohn and Ellenberger, 1977), SOS chromotest (Quillardet et al., 1985) L-ara test (Ruiz-Vazquez et al., 1978) and the Umu test (Oda et al., 1985). Beside the routine screening for genotoxins, these assays have been used in studies delineating mechanisms by which these chemicals exert their deleterious effect (Hofnung and Quillardet, 1986; Quillardet and Hofnung, 1988). The majority of these tests are based on the detection of specific mutational events. A mutation can be the result of the con- 0165-1161/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0165-1161(93)E0049-G
-
Upload
puc-rio-br -
Category
Documents
-
view
3 -
download
0
Transcript of Isolation and prevalidation of an Escherichia coli tester strain for the use in mechanistic and...

E L S E V I E R Mutation Research 312 (1994) 99-109 Environmental Mutagenesis
Isolation and prevalidation of an Escherichia coli tester strain for the use in mechanistic and metabolic studies of genotoxins *
Michel Kranendonk a, Margarida Ruas a, Antdnio Laires a,b, Jos6 Rueff a ' *
a Department of Genetics, Faculty of Medical Sciences, UNL, R. de Junqueira 96, P-1300 Lisbon, Portugal, b Faculty of Sciences and Technology, UNL, Monte de Caparica, Portugal
Received 10 September 1993; revision received 16 November 1993; accepted 19 November 1993
Abstract
We have isolated an Escherichia coli tester strain for the use in mechanistic and metabolic studies of genotoxins. We started with one of the more used and better characterized E. coli K-12 laboratory strains, ABl157. We isolated a lipopolysaccharide defective mutant of strain AB1886 which is an excision repair deficient derivative of ABl157 and introduced a newly constructed plasmid pKRll, encoding mucAB, resulting in strain MR2101/pKRll. A genotoxicity assay was designed, monitoring the reversion to arginine prototrophy and a preliminary validation was carried out against Ames tester strain TA100 with a set of diagnostic compounds. The results seem to indicate that strain MR2101/pKRll is an adequate tester strain which can be a useful tool in mechanistic studies. Moreover, this strain can serve as mother strain to isolate improved and more specialized tester strains.
Key words: E. coli; Tester strain
1. Introduction
Despite the apparent simplicity compared with eukaryotic organisms, bacteria possess elaborate mechanisms for responding to D N A damage (Walker, 1984; Sedgwick and Goodwin, 1985; Sedgwick et al., 1991a). Bacteria are therefore widely used in fundamental studies of mecha- nisms involved in the biological response to D N A damage and in short-term genotoxicity tests. In the last decades various bacterial short-term as-
* Corresponding author. * Dedicated to the memory of Professor F.H. Sobels.
says have been developed for the detection of chemical carcinogens as mutagens, e.g., the Ames test (Maron and Ames, 1983), WP2 test (Venitt et al., 1984), Inductest (Moureau et al., 1976), E. coli K12/343 /113 test (Mohn and Ellenberger, 1977), SOS chromotest (Quillardet et al., 1985) L-ara test (Ruiz-Vazquez et al., 1978) and the Umu test (Oda et al., 1985). Beside the routine screening for genotoxins, these assays have been used in studies delineating mechanisms by which these chemicals exert their deleterious effect (Hofnung and Quillardet, 1986; Quillardet and Hofnung, 1988). The majority of these tests are based on the detection of specific mutational events. A mutation can be the result of the con-
0165-1161/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0165-1161(93)E0049-G

100 M. Kranendonk et al. / Mutation Research 312 (1994) 99-109
version of a genetic lesion in at least two different ways: (i) by miscoding, directed by the lesion and (ii) by SOS mutagenesis, a process involving the induction of specific cellular functions: the SOS functions (Walker, 1984).
We are currently studying the mechanisms and metabolism of genotoxins (Rueff et al., 1992a,b). For this purpose we are interested in a tester bacterium which can assist in such studies, beside the application of the routinely used tester bacte- ria.
The test most frequently used, and therefore best validated, is the Ames assay (Kier et al., 1986). This assay, as well as the L-ara test and the Umu test, uses Salmonella typhimurium as target bacterium. S. typhimurium is considered to be poorly responsive to DNA damage when com- pared to E. coli (Sedgwick and Goodwin, 1985). This poor mutability is despite the fact that it has a DNA damage inducible SOS regulon that re- sembles that of E. coli and contains at least two umuDC-like sequences, which are considered to be required for mutagenic DNA repair (Sedgwick and Goodman, 1985; Orrega and Eisenstad, 1987; Nohmi et al., 1991, 1992; Sedgwick et al., 1991a,b; Woodgate et al., 1991; Sommer et al., 1993). The major part of the success of the Ames Salmonella test is due to the introduction of an E. coli plasmid (pKM101), containing the mucAB operon (a plasmid analogue of the umuDC operon), in the Salmonella tester strains (McCann et al., 1975; Walker, 1979). The SOS response is better under- stood in E. coli than in S. typhimurium (Sedgwick et al., 1991a). Although the mentioned Salmonella assays are characterized and validated and are useful for routine screening, E. coli assays seem to be more appropriate to study mechanisms leading to mutagenic events and the role of DNA repair. The activity of SOS regulated E. coli gene products for certain types of mutations has been shown, e.g., for different frameshift mutations (Koffel-Schwartz et al., 1984; Koffel-Schwartz and Fuchs, 1989; Bintz and Fuchs, 1990).
The involvement of bacterial metabolism in the mutagenic effect of some compounds has been shown (Callen, 1978; Mohn et al., 1984; Snyderwine et al., 1988)).
The characterization of loci of genes whose
products could be involved in such metabolism seems to be more complete for E. coli (compare linkage map of E. coli K12 (Bachmann, 1990) with linkage map of S. typhimurium (Sanderson and Roth, 1988 and references in both)).
An assay applying an E. coli strain, which is genetically well characterized for SOS mutagene- sis as well as for its metabolic activities, could therefore be a powerful tool in studies of the mechanisms and metabolisms by which specific mutagens exert their genotoxic effect.
For such studies, we opted for the develop- ment of a new E. coli tester bacterium as an additional tool beside the routinely used E. coli tester bacteria, starting with one of the more used and better characterized E. coli laboratory strains, ABl157. This strain, originally isolated and used by Howard-Flanders (1966) in one of his first DNA excision repair studies, has been shown to be one of the more mutable E. coli strains (Sedgwick et al., 1991a). This strain has been extensively used in studies of the SOS re- sponse, especially of SOS mutagenesis (Langer et al., 1981, 1985; Sedgwick and Goodwin, 1985; Donnelly and Walker, 1989, 1992; Sedgwick et al., 1991a; Sommer et al., 1993 and references within) and is therefore largely characterized for its response to genotoxic challenge. Moreover, ABl157 has a convenient genetic target to be monitored in mutagenicity testing, namely the reversion from arginine auxotrophy, due to the ArgE3 mutation, to arginine prototrophy by sub- stitution mutations (Todd et al., 1979). We iso- lated a lipopolysaccharide defective mutant of an excision repair deficient mutant of ABl157, namely AB1886 (uvrA6) and introduced a newly constructed mucAB encoding plasmid (pKRll), resulting in strain MR2101/pKRl l . Subse- quently, a mutagenicity assay was developed to monitor the Arg reverse mutation. We show the importance of the deficiency in DNA excision repair and of the defective lipopolysaccharide barrier as well as the presence of the mucAB operon in the tester strain for a high response upon mutagenic treatment. A preliminary valida- tion of MR2101/pKRll was performed against Ames tester strain TA100 with a set of diagnostic compounds.

M. Kranendonk et al. / Mutation Research 312 (1994) 99-109 101
2. Materials and methods
2.1. Reagents, chemicals and enzymes
L-histidine (L-his), D-glucose monohydrate, crystal violet, sodium azide (NaN 3) and 2-nitro- fluorene (2NF) were obtained from Merck (Darmstadt)• The amino acids L-proline (L-pro), L-leucine (L-leu), L-threonine (L-thr) and L- arginine (L-arg) as well as sodium deoxycholate, ethidium bromide, 7,12-dimethylbenz[a]anthra- cene (DMBA), pyrene, quercetin (Q) and 4- nitroquinoline-l-oxide (4NQO) were purchased from Sigma• Ampicillin (sodium salt), tetracycline • HCI, ADNA and the restriction endonuclease HindIII were from NBL. Bacto agar, bacto tryp- tone, bacto yeast extract and bacto MacConkey agar were from Difco, dimethyl sulfoxide (DMSO) (Riedel-de Hahn), nutrient broth (NB) from Fluka, thiamine (diphosphate form: triphospho- neurina) (Biofranco), aflatoxin B1 (AFB1) from Aldrich. The antibiotic kanamycin sulfate was obtained from Boeringher-Mannheim. The re- striction endonucleases BamHI and SalI were obtained from Biolabs and Amersham respec- tively. Klenow's DNA polymerase I, T4 DNA
ligase as well as ATP were from Pharmacia and dTTP, dCTP, dGTP and dATP from Perkin- Elmer Cetus. Benz[a]pyrene (B(a)P) was a gift from Pasteur Institute (Paris).
2.2. Microbiological media
For Escherichia coli: LB medium (Sambrook et al., 1989) was used as nutrient-rich medium and when necessary supplemented with ampicillin (for LPS ÷ strains: 50 /xg/ml, for LPS d strains: 25 /xg/ml), tetracycline (15 /zg/ml) and kanamycin (for LPS ÷ strains: 25/xg/ml, for LPS d strains: 15 /zg/ml). M9 minimal glucose medium (Sambrook et al., 1989) was used as selective minimal medium in other than the standard MR2101/pKRll mu- tagenicity assay. Bacteria were plated on M9 plates (25 ml) in M9 top agar (2 ml) and supple- mented with thiamine (1 /zg/ml), L-pro, L-thre, L-leu and L-his (100 /zg/ml). For the Arg ÷ re- version determination the M9 plates contained 1 /xg/ml L-arg, for survival determinations 100 /xg/ml. All mentioned concentrations are final plate concentrations. For tester plates and tester tops for MR2101/pKR1 see section 2.6.
Table 1 Bacterial strains and plasmids
Strains Genotype Origin
E. coli ABl157
MR1001 AB1886 NK6033 MR2000 PQ33
MR2100 MR2101 DH1
thr-1, ara-14, leuB 6, A( gpt-proA )62, lacY1, tsx-33, qsr-, supE 44, galK 2, A-, rac- hisG4, rfbD1, mgl-51, rpsL31, kdgK51, xyl-5, rntl-1, argE3, thi-1 ABl157, LPS defective ABl157, uvrA6 A(gpt-lac)5, nadA50::TnlO, relA1, spoT1, thi-1, h - AB1886, nadA 50::Tn 10 sfi::Mud(Ap lac)cts, IacAU169, uvrA, galE, galY, PhoC, thr, leu, his, pyrD, thi, trp::Muc +, srl3OO::TnlO, rpoB AB1886, galE MR2100, LPS defective supE44, hsdR17, recA1, endA1, gyrA96, thi-1, relA1
B. Bachmann
This paper (U3 r selection on ABl157) B. Bachmann B. Bachmann This paper (Pl[NK6033] x AB1886 to Tet r) M. Hofnung and P. Quillardet
This paper (Pl[PQ33] x MR2000 to NAD + This paper (C21 r selection on Mr2100) B. Bachmann
S. typhimurium TA100 hisG46, A(gal-uvrB), rfa-lOO1/pKMl01 B.N. Ames
Plasmids Relevant genetic markers Origin
pKM101 mucAB +, Amp r, B.N. Ames pGW270 rnucAB +, Ampr, Kanr G.C. Walker pKRl l mucAB +, Kanr This paper (A[SalI-BamHI] X pGW270)

102 M. Kranendonk et al. / Mutation Research 312 (1994) 99-109
For Salmonella typhimurium (TA100): all me- dia were according to Maron and Ames (1983).
2.3. Bacterial strains and transductions
All transductions were performed according to Miller (1972) with bacteriophage P1 vir. The geno- type and the origin of the different strains are shown in Table 1.
2.4. Isolation of lipopolysaccharide defective (LPS d) mutants
A LPS d mutant of strain ABl157 (MR1001) was isolated by the bacteriophage U3 ~ selection method as described by Ellenberger (1982). We isolated a stable (LPS d) mutant of MR2100 by the bacteriophage C21 r selection method, de- scribed by Ames et al. (1973). The more perme- able LPS d mutants of both ABl157 and MR2100 (designated MR1001 and MR2101 respectively) were isolated by selection of the mutant with the highest sensitivity towards crystal violet, sodium deoxycholate and ethidium bromide.
2.5. Plasmid pKRl l
Plasmid pKRl l was constructed by deletion of the bla (Amp r) gene of plasmid pGW270, a dele- tion derivative of plasmid pKM101 (Langer et al., 1981). pGW270 was cleaved with BamHI and Sail, 3'-recessed termini were filled in by Klenow's DNA polymerase I and ligated with T4 DNA ligase (Sambrook et al., 1989). A portion of the ligation mixture was transformed to DH1 by the CaCI z method (Sambrook et al., 1989). A number of kanamycin resistant transformants were picked up and checked for their plasmid content: minipreparations of plasmid DNA were isolated (Sambrook et al., 1989) and verified for the loss of the BamHI and Sail sites, cleaved with HindlII (a unique restriction site of pGW270 and pKRll ) to verify the deletion of the -~ 2.5-kb (BamHI-SalI) fragment (data not shown). Band intensities of pGW270 and pKRl l were com- pared to check for change in copy number of pKRl l compared to pGW270, pKRl l was trans- formed to the different tester strains by the CaCl2 method.
2.6. Mutagenicity assays
(a) Comparison of pGW270 and pKRl l in SOS mutagenesis. We compared the effect of pGW270 and pKR11 on SOS mutagenesis in strain ABl157, using UV and 4NQO, measuring Arg to Arg + reversion as described before (Langer et al., 1985) with minor modifications: (i) UV mutagenesis: 50 ml of mid-exponential growing cells (2-4 × 108 cfu/ml) of strains ABl157, AB1157/pGW270 and ABl157/pKR11 were harvested, washed and resuspended in the original volume with ice cold saline solution (0.85%). 3.6 ml was placed into a petri dish (diameter: 90 mm) and irradiated for a fixed time with UV light at a UV fluence of 1 J / m 2 per s. All subsequent manipulations were carried in semi-dark conditions. Aliquots of irradiated sus- pensions were plated either directly for Arg + reversion determination or appropriately diluted for survival determination. The plates were incu- bated for 72 h before counting survivors and revertants. (ii) 4NQO mutagenesis: = 2 × 108 cells of a sta- tionary culture of strains ABl157, ABl157/ pGW270 and ABl157 /pKRl l were plated in the presence of different dose levels of 4NQO. Arg ÷ revertants were counted after 48 h of incubation.
(b) MR2101/pKR11. The manipulations de- scribed below of the tester strain MR2101/ pKRl l were to ascertain that an optimal and standardized way of working was achieved. (i) Storage of tester strain: aliquots of a station- ary culture of MR2101 / pKR11 in LB, containing kanamycin (15/xg/ml) were stored in 15% (v/v) glycerol after the characteristics of the isolated strain were checked (LPSd: growth sensitivity on MacConkey agar plates; pKRll : kanamycin resis- tance (15/xg/ml); uvrA: UV sensitivity and Arg auxotrophy: growth and no growth on M9 plates with or without L-Arg, respectively). (ii) Test plate: 30 ml Vogel-Bonner E plates (VB) as used in the Ames assay (Maron and Ames, 1983). (iii) Test top: 2 ml, 0.5% NaCI, 0.6% agar sup- plemented with L-pro, L-thre, L-leu, L-his: 100 /xg/ml and thiamine: 1 /xg/ml (all final plate concentrations). Test tops were liquefied at 100°C

M. Kranendonk et a l . / Mutation Research 312 (1994) 99-109 103
and afterwards maintained at 45-48°C. L-arg (3 /xg/ml, final plate concentration, as determined by L-arg optimization) was added to the test tops just before use. (iv) Cultures of MR2101 /pKR11:20 /xl of a defrosted stock was used to inoculate 5 ml of LB (without kanamycin) and was grown during 14-16 h. In these conditions a cell density of = 1 x 109 c fu /ml was reached. (Other nutrient-rich media were experimented with but LB gave the highest cell density). (v) Test: 200 /xl (= 2 x 108 cells) of a stationary culture was plated in the presence of the test compound (with 500 /zl of $9 mix (10%) when indicated) in test top on test plate. Test plates were incubated for 48 h before Arg + revertants were counted. 10% $9 mix was prepared accord- ing to Maron and Ames (1983). Spontaneous Arg + reversion was determined by plating bacte- ria without test compound in each test.
(c) TAIO0. The assays with tester strain TA100 were performed as described by Maron and Ames (1983).
At least three independent experiments were carried out for each dose level. Mutagenic activi- ties were determined from the slope as the least square line of the linear portion of the dose-re- sponse curve.
permeability to many compounds and have been shown to be useful for testing carcinogenic sub- stances (Ames et al., 1973; Quillardet and Hof- nung, 1985). We used the bacteriophage U3 resis- tance selection method (Ellenberger, 1982) for the isolation of a LPS d mutant of strain ABl157, designated MR1001. The same method was ap- plied for strain AB1886 for the isolation of a highly permeable mutant. Upon storage at low temperature, the AB1886 U3 r mutants acquired a mucoid phenotype with the loss of the increased permeability. The induction of the mucoid phe- notype by low temperature has been shown be- fore (Stout and Gottesman, 1990; Parker et al., 1992). The mucoid appearance is due to the production of colanic acid polysaccharide, form- ing a capsule exterior to the outer membrane (Markowitz, 1977). To prevent the formation of this extra external barrier, we inactivated galE, a gene whose product is essential for colanic acid synthesis (Trisler and Gottesman, 1984), resulting in MR2100. Subsequently, we used bacteriophage C21 for the isolation of LPS d mutants (Ames et al., 1973). Non-mucoids were obtained and the most permeable mutant was selected showing a permeability similar to that found for MR1001 (data not shown).
3.2. The mucAB operon
3. Results
For the isolation of an E. coli tester strain to be used in mechanistic and metabolic studies of genotoxins, we started with one of the more used and better characterized E. coli K12 laboratory strains, ABl157. AB1886, a strain isogenic to ABl157 except for uvrA, was used in the isola- tion as a large number of genotoxins are more readily detected in strains with an excision repair deficient background (Quillardet and Hofnung, 1988). The different strains used and isolated are described in Table 1.
3.1. Isolation of lipopolysaccharide deficient ( LPS d) mutants
Bacteria defective in LPS, the major compo- nent of the outer membrane, have an increased
pKM101, a mucAB (Amp r) encoding plasmid, has been introduced into a number of different bacterial tester strains for its ability to increase the susceptibility of cells to chemical mutagenesis (Langer et al., 1981). For this reason we have introduced a newly constructed mucAB carrying plasmid (pKRll ) into our isolated tester strains. For future mechanistic and metabolic studies of genotoxins, it was necessary to introduce a mu- cAB plasmid without conveying ampicillin resis- tance to the cells as we want to introduce a second (Amp r) plasmid. This second plasmid could either be an expression vector for heterolo- gous expression of mammalian drug metabolizing enzymes or a plasmid carrying a specific genetic target, pKRl l was obtained by deleting a 2.5-kb fragment, containing the Amp r encoding region (bla), of plasmid pGW270. Plasmid pGW270 is a
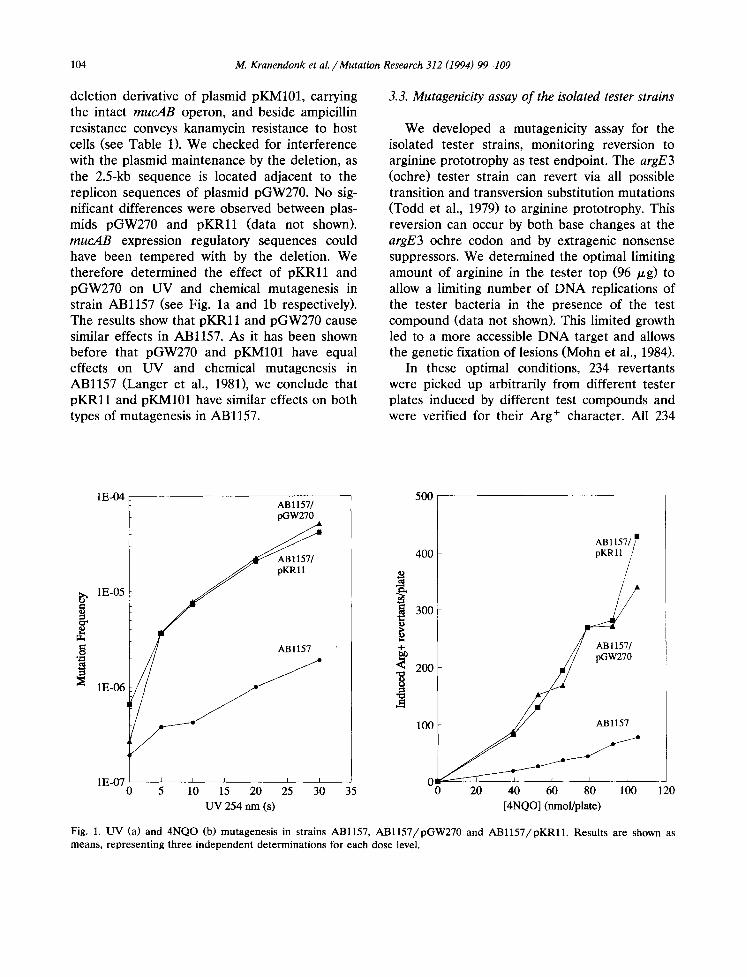
104 M. Kranendonk et al. /Mutation Research 312 (1994) 99-109
deletion derivative of plasmid pKM101, carrying the intact mucAB operon, and beside ampicillin resistance conveys kanamycin resistance to host cells (see Table 1). We checked for interference with the plasmid maintenance by the deletion, as the 2.5-kb sequence is located adjacent to the replicon sequences of plasmid pGW270. No sig- nificant differences were observed between plas- mids pGW270 and pKRl l (data not shown). mucAB expression regulatory sequences could have been tempered with by the deletion. We therefore determined the effect of pKRl l and pGW270 on UV and chemical mutagenesis in strain ABl157 (see Fig. la and lb respectively). The results show that pKRl l and pGW270 cause similar effects in ABl157. As it has been shown before that pGW270 and pKM101 have equal effects on UV and chemical mutagenesis in ABl157 (Langer et al., 1981), we conclude that pKRl l and pKM101 have similar effects on both types of mutagenesis in ABl157.
3.3. Mutagenicity assay of the isolated tester strains
We developed a mutagenicity assay for the isolated tester strains, monitoring reversion to arginine prototrophy as test endpoint. The argE3 (ochre) tester strain can revert via all possible transition and transversion substitution mutations (Todd et al., 1979) to arginine prototrophy. This reversion can occur by both base changes at the argE3 ochre codon and by extragenic nonsense suppressors. We determined the optimal limiting amount of arginine in the tester top (96 /xg) to allow a limiting number of DNA replications of the tester bacteria in the presence of the test compound (data not shown). This limited growth led to a more accessible DNA target and allows the genetic fixation of lesions (Mohn et al., 1984).
In these optimal conditions, 234 revertants were picked up arbitrarily from different tester plates induced by different test compounds and were verified for their Arg + character. All 234
1E-04
i 1E-05
1E-06
ABl157/ pGW270
p K R I I ~
500
-4-
400
300
200
100
I I
1E-°70 5 ,0 20 30 35 °0 120 UV 254 nm (s)
I I AB 1157//-
20 40 60 80 100 [4NQO] (nmol]plate)
Fig. 1. UV (a) and 4NQO (b) mutagenesis in strains ABl157, AB1157/pGW270 and AB1157/pKRll. Results are shown as means, representing three independent determinations for each dose level.
M. Kranendonk et al. / Mutation Research 312 (1994) 99-109 105
1200
uvrA, LPS d mucAB +
1000
800
! 6 0 0 . - I -
400
/ ~ uvrA, LPS d, mucAB-
0 ~ - " - ~" uvrA + LPS d, mucAB +
I I i I I I
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
[AFB 1] (nmol/plate)
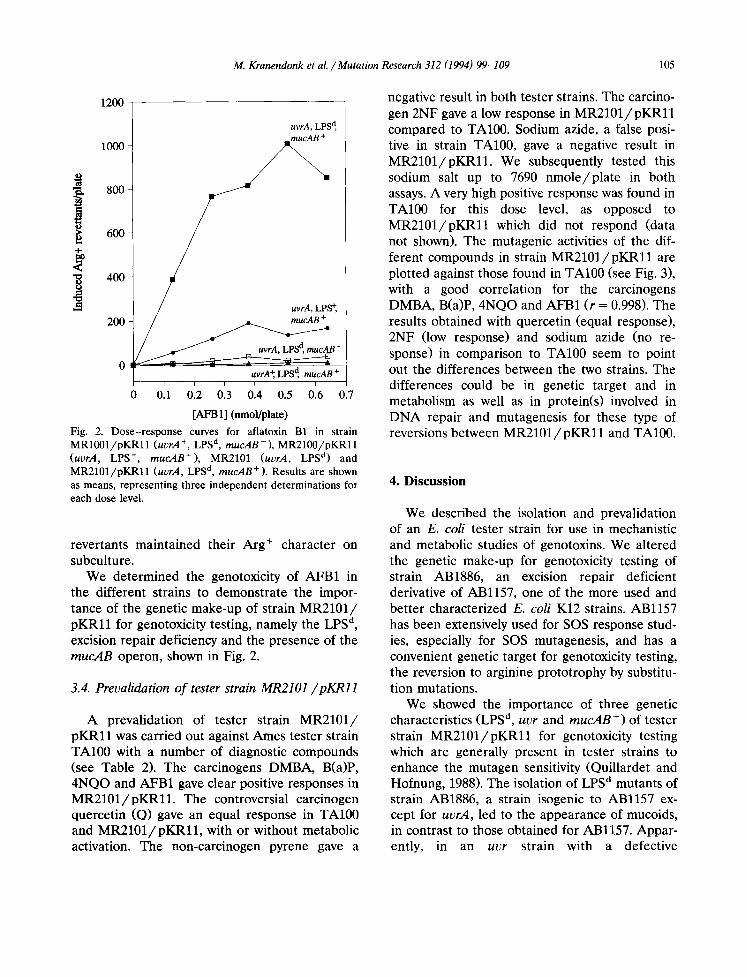
Fig. 2. Dose-response curves for aflatoxin B1 in strain MR1001/pKRll (uvrA ÷, LPS d, raucAB +), MR2100/pKRll (uvrA, LPS ÷, mucAB+), MR2101 (uvrA, LPS ~) and MR2101/pKRll (uvrA, LPS d, mucAB + ). Results are shown as means, representing three independent determinations for each dose level.
revertants maintained their Arg + character on subculture.
We determined the genotoxicity of AFB1 in the different strains to demonstrate the impor- tance of the genetic make-up of strain MR2101/ pKRl l for genotoxicity testing, namely the LPS d, excision repair deficiency and the presence of the mucAB operon, shown in Fig. 2.
3.4. Prevalidation o f tester strain M R 2 1 0 1 / p K R l l
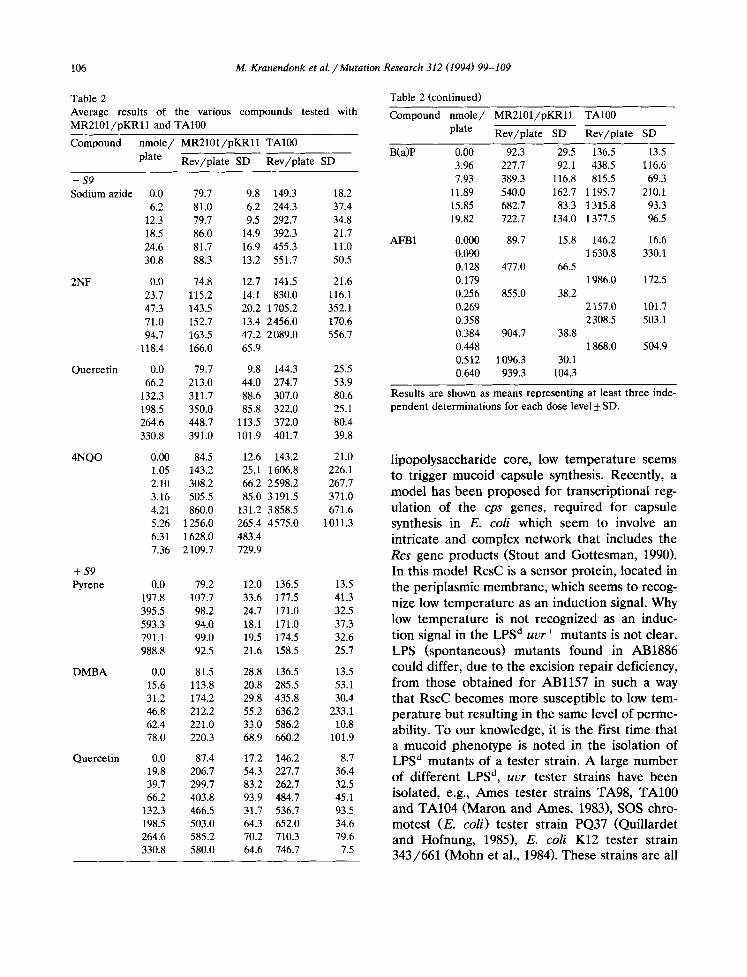
A prevalidation of tester strain MR2101/ pKRl l was carried out against Ames tester strain TA100 with a number of diagnostic compounds (see Table 2). The carcinogens DMBA, B(a)P, 4NQO and AFB1 gave clear positive responses in MR2101/pKRll . The controversial carcinogen quercetin (Q) gave an equal response in TA100 and MR2101/pKRll , with or without metabolic activation. The non-carcinogen pyrene gave a
negative result in both tester strains. The carcino- gen 2NF gave a low response in MR2101/pKRll compared to TA100. Sodium azide, a false posi- tive in strain TA100, gave a negative result in MR2101/pKRll . We subsequently tested this sodium salt up to 7690 nmole/plate in both assays. A very high positive response was found in TA100 for this dose level, as opposed to MR2101/pKRll which did not respond (data not shown). The mutagenic activities of the dif- ferent compounds in strain MR2101/pKRll are plotted against those found in TA100 (see Fig. 3), with a good correlation for the carcinogens DMBA, B(a)P, 4NQO and AFB1 (r = 0.998). The results obtained with quercetin (equal response), 2NF (low response) and sodium azide (no re- sponse) in comparison to TA100 seem to point out the differences between the two strains. The differences could be in genetic target and in metabolism as well as in protein(s) involved in DNA repair and mutagenesis for these type of reversions between MR2101/pKRll and TA100.
4. Discussion
We described the isolation and prevalidation of an E. coli tester strain for use in mechanistic and metabolic studies of genotoxins. We altered the genetic make-up for genotoxicity testing of strain AB1886, an excision repair deficient derivative of ABl157, one of the more used and better characterized E. coli K12 strains. ABl157 has been extensively used for SOS response stud- ies, especially for SOS mutagenesis, and has a convenient genetic target for genotoxicity testing, the reversion to arginine prototrophy by substitu- tion mutations.
We showed the importance of three genetic characteristics (LPS d, uvr and mucAB +) of tester strain MR2101/pKRll for genotoxicity testing which are generally present in tester strains to enhance the mutagen sensitivity (Quillardet and Hofnung, 1988). The isolation of LPS d mutants of strain AB1886, a strain isogenic to ABl157 ex- cept for uvrA, led to the appearance of mucoids, in contrast to those obtained for ABl157. Appar- ently, in an uvr strain with a defective
106 M. Kranendonk et al. / Mutation Research 312 (1994) 99-109
Table 2 Average results of the various compounds tested with MR2101/pKRll and TA100
Compound nmole/ MR2101/pKRll TA100
plate Rev/plate SD Rev/plate SD
- $9 Sodium azide
2NF
Quercetin
4NQO
+ $9 Pyrene
DMBA
Quercetin
0.0 79.7 9.8 149.3 18.2 6.2 81.0 6.2 244.3 37.4
12.3 79.7 9.5 292.7 34.8 18.5 86.0 14.9 392.3 21.7 24.6 81,7 16.9 455.3 11.0 30.8 88.3 13.2 551.7 50.5
0.0 74.8 12.7 141.5 21.6 23.7 115.2 14.1 830.0 116.1 47.3 143.5 20.2 1705.2 352.1 71.0 152.7 13.4 2456.0 170.6 94.7 163.5 47.2 2089.0 556.7
118.4 166.0 65.9
0.0 79.7 9.8 144.3 25.5 66.2 213.0 44.0 274.7 53.9
132.3 311.7 88.6 307.0 80.6 198,5 350.0 85.8 322.0 25.1 264.6 448.7 113.5 372.0 80.4 330.8 391.0 101.9 401.7 39.8
0.00 84.5 12.6 143.2 21.0 1.05 143.2 25.1 1606.8 226.1 2.10 308.2 66.2 2598.2 267.7 3.16 505.5 85.0 3191.5 371.0 4.21 860.0 131.2 3858.5 671.6 5.26 1256.0 265.4 4575.0 1011.3 6.31 1628.0 483.4 7.36 2109.7 729.9
0.0 79.2 12.0 136.5 13.5 197.8 107.7 33.6 177.5 41.3 395.5 98.2 24.7 171.0 32.5 593.3 94.0 18.1 171.0 37.3 791.1 99.0 19.5 174.5 32.6 988.8 92.5 21.6 158.5 25.7
0.0 81.5 28.8 136.5 13.5 15.6 113.8 20.8 285.5 53.1 31.2 174.2 29.8 435.8 30.4 46.8 212.2 55.2 636.2 233.1 62.4 221,0 33.0 586.2 10.8 78.0 220.3 68.9 660.2 101.9
0.0 87.4 17.2 146.2 8.7 19.8 206.7 54.3 227.7 36.4 39.7 299.7 83.2 262.7 32.5 66.2 403.8 93.9 484.7 45.1
132.3 466.5 31.7 536.7 93.5 198.5 503.0 64.3 652.0 34.6 264.6 585.2 70.2 710.3 79.6 330.8 580.0 64.6 746.7 7.5
Table 2 (continued)
Compound nmole/ MR2101/pKRll TA100
plate Rev/plate SD Rev/plate SD
B(a)P
AFB1
0.00 92.3 29.5 136.5 13.5 3.96 227.7 92.1 438.5 116.6 7.93 389.3 116.8 815.5 69.3
11.89 540.0 162.7 1 195.7 210.1 15.85 682.7 83.3 1315.8 93.3 19.82 722.7 134.0 1377.5 96.5
0.000 89.7 15.8 146.2 16.6 0.090 1630.8 330.1 0.128 477.0 66.5 0.179 1986.0 172.5 0.256 855.0 38.2 0.269 2157.0 101.7 0.358 2308.5 503.1 0.384 904.7 38.8 0.448 1868.0 504.9 0.512 1096.3 30.1 0.640 939.3 104.3
Results are shown as means representing at least three inde- pendent determinations for each dose level :t: SD.
l ipopolysacchar ide core, low t e m p e r a t u r e seems
to t r igger mucoid capsule synthesis. Recent ly , a
mode l has b e e n p roposed for t ranscr ipt ional reg-
u la t ion of the cps genes, r equ i red for capsule
synthesis in E. col i which seem to involve an
intr icate and complex ne twork that includes the
R c s gene products (Stout and Got tesman , 1990).
In this mode l RcsC is a sensor protein , loca ted in
the per ip lasmic m e m b r a n e , which seems to recog-
nize low t e m p e r a t u r e as an induct ion signal. Why
low t e m p e r a t u r e is not r ecogn ized as an induc-
t ion signal in the LPS a u v r + mutants is not clear.
LPS (spontaneous) mutan t s found in AB1886
could differ, due to the excision repa i r deficiency,
f rom those ob ta ined for A B l 1 5 7 in such a way
that RscC becomes m o r e susceptible to low tem-
pe ra tu re but resul t ing in the same level of pe rme-
ability. To our knowledge , it is the first t ime that
a muco id pheno type is no ted in the isolat ion of
LPS a mutants o f a tes ter strain. A large n u m b e r
of d i f ferent LPS a, u v r tes ter strains have b e e n
isolated, e.g., A m e s tes ter strains TA98, TA100
and TA104 ( M a r o n and Ames , 1983), SOS chro-
motes t (E . co l i ) tes ter strain PQ37 (Qui l la rde t
and Hofnung , 1985), E. col i K12 tes ter strain 343//661 (Mohn et al., 1984). These strains are all

M. Kranendonk et al. / Mutation Research 312 (1994) 99-109 107
100000
10000
1000
100
10
AFBI
• 2NF / / ~ ( a ) P
,4/DMBA * Q+S9
Q-S9
1 I ] ] I I I~1110 I I I [ . . . . . . . . 1 ~ I . . . . . l ~ I I I I i I I I ~ 0
MR2101/pKR11 (rev/nmol)
Fig. 3. Correlation of mutagenic activities of strain MR2101/ pKRl l against strain TA100 for seven compounds. Mutagenic activities were determined as the least square line of the linear portion of the dose-response curve.
ins. One of the more promising approaches is the direct introduction of genes/cDNAs of mam- malian drug metabolizing enzymes in the assay's target cell (see Gonzalez et al., 1991; Langenbach et al., 1992). A number of different expression vectors have been developed for the expression of mammalian genes in bacteria. They contain in- ducible E. coli promoters, the majority based on the lactose operon repressor LacI. The activity of this promoter can be chemically controlled by isopropyl fl-o-thiogalactopyranoside (IPTG). The controllability of this type of promoter is lost upon introduction into S. typhimurium, due to the lack of the lac operon in this bacterium (Lampel and Riley, 1982). Strain MR2101/ pKRl l with minor modifications becomes a suit- able host for these LacI expression vectors, re- suiting in a tester bacterium in which controllable expression of mammalian or prokaryotic drug metabolizing enzymes can be obtained. Such a tester bacterium can be a powerful tool for metabolic studies of genotoxins.
5. Acknowledgements
gal due to the uvrB deletion, co-deleting the gal operon. LPS a non-mucoid mutants were isolated due to the lack of the gale gene product required for capsule synthesis.
We constructed a new mucAB carrying plas- mid pKRl l to allow the introduction of Amp r conveying plasmids beside a mucAB carrying plasmid, pKRl l seems to have equal effects on SOS mutagenesis as pGW270 and thus seems to equal pKMIO1.
The obtained results seem to characterize MR2101/pKR11 as an adequate strain for mech- anistic studies. This strain could serve as a mother strain for the isolation of specialized tester strains by, e.g., the introduction of specific plasmid-borne or genomic genetic targets and can therefore be an additional tool for the study of mutagenic pathways of specific genotoxins and the involve- ment of DNA repair.
In the last few years, a lot of effort has been put into the unraveling of mammalian metabolic pathways involved in the metabolism of genotox-
We are grateful to Dr. B.J. Bachmann, Prof. Dr. M. Hofnung and Dr. P. Quillardet for supply- ing the E. coli strains, to Dr. G.C. Walker for giving the plasmid pGW270, to Dr. R. Fuchs for the phages U3 and P1 vir, Dr. M. Llagostera and Dr. B. Clerch for supplying phage C21. This work was supported by the CEC (STEP Programme) and the CIENCIA Programme.
6. References
Ames, B.N., F.D. Lee and W.E. Durton (1973) An improved bacterial test system for the detection and classification of mutagens and carcinogens, Proc. Natl. Acad. Sci. USA, 70, 782-786.
Bachmann, B.J. (1990) Linkage map of Escherichia coli K-12, edn. 8, Microbiol. Rev., 54, 130-197.
Bintz, R., and R.P.P. Fuchs (1990) Induction of - 2 frameshift mutations within alternating GC sequences by carcinogens that bind to the C8 position of guanine residues: Develop- ment of a specific mutation assay, Mol. Gen. Genet., 221, 331-338.

108 M. Kranendonk et al. / Mutation Research 312 (1994) 99-109
Callen, D.F. (1978) A review of the metabolism of xenobiotics by microorganisms with relation to short-term systems for environmental carcinogens, Mutation Res., 55, 153-163.
Donnelly, C.E., and G.C. Walker (1989) groE mutants of Escherichia coli are defective in umuDC-dependent UV mutagenesis, J. Bacteriol., 171, 6117-6125.
Donnelly, C.E., and G.C. Walker (1992) Coexpression of umuD' with umuC suppresses the UV mutagenesis defi- ciency of groE mutants, J. Bacteriol., 174, 3133-3139.
Ellenberger, J. (1982) Increased sensitivity of Escherichia coli K12 to certain mutagens as a consequence of a mutation leading to phage U 3 resistance, Mutation Res., 104, 55-60.
Gonzalez, F.J., C.L. Crespi and H.V. Gelboin (1991) cDNA- expressed human cytochrome P450s: a new age of molecu- lar toxicology and human risk assessment, Mutation Res., 247, 113-127.
Hofnung, M., and P. Quillardet (1986) Recent developments in bacterial short-term tests for the detection of genotoxic agents, Mutagenesis, 1, 319-330.
Howard-Flanders, P., R.P. Boyce and L. Theriot (1966) Three loci in Escherichia coli K-12 that control the excision of pyrimidine dimers and certain other mutagen products from DNA, Genetics, 53, 1119-1136,
Kier, L.E., D.J. Brusick, A.E. Auletta, E.S. Von Halle, M.M. Brown, V.F. Simmon, V. Dunkel, J. McCann, K. Moertel- mans, M. Prival, T.K. Roa and V. Ray (1986) The Salmonella typhimurium/mammalian microsomal assay. A report of the US Environmental Protection Agency Gene- Tox Program, Mutation Res., 168, 69-240.
Koffel-Schwartz, N., and R.P.P. Fuchs (1989) Genetic control of 2-AAF induced mutagenesis at alternating GC se- quences: an additional role for RecA, Mol. Gen. Genet., 215, 306-311.
Koffel-Schwartz, N., J.M. Verdier, M. Bichara, A.M. Freund, M.P. Daune and R.P.P. Fuchs (1984) Carcinogen induced mutation spectrum in wildtype, uvrA and umuC strains of E. coli: strain specificity and mutation prone sequences, J. Mol. Biol., 177, 33-51.
Lampel, K.L., and M. Riley (1982) Discontinuity of homology of Escherichia coli and Salmonella typhimurium DNA in the lac region, Mol. Gen. Genet., 186, 82-86.
Langenbach, R., P.B. Smith and C. Crespi (1992) Recombi- nant DNA approaches for the development of metabolic systems used in in vitro toxicology, Mutation Res., 277, 251-275.
Langer, P.J., W.G. Shanabruch and G.C. Walker (1981) Func- tional organization of plasmid pKM101, J. Bacteriol., 145, 1310-1316.
Langer, P.J., K.L. Perry and G.C. Walker (1985) Complemen- tation of a pKM101 derivative that decreases resistance to UV killing but increases susceptibility to mutagenesis, Mutation Res., 150, 147-158.
Markowitz, A. (1977) Genetics and regulation of bacterial capsular polysaccharide synthesis and radiation sensitivity, in: Sutherland (Ed.), Surface Carbohydrates of the Prokaryotic Cell, Academic Press, London, pp. 415-462.
Maron, D.M., and B.N. Ames (1983) Revised methods for the Salmonella mutagenicity test, Mutation Res., 113, 173-215.
McCann, J., N.E. Springarn, J. Kobori and B.N. Ames (1975) Detection of carcinogens as mutagens: bacterial tester strains with R factor plasmids, Proc. Natl. Acad. Sci. USA, 72, 979-983.
Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Mohn, G., and J. Ellenberger (1977) The use of Escherichia coli, K12/343/113 (~.) as a multi-purpose indicator strain in various mutagenicity testing procedures, in: B.J. Kilbey, M. Legator, W. Nichols and C. Ramel (Eds.), Handbook of Mutagenicity Test"Procedures, Elsevier, Amsterdam, pp. 95-118.
Mohn, G.R., P.R.M. Kerklaan, A.A. Zeeland, J. Ellenberger, R.A. Baan, P.H.M. Lohman and F.-W. Ports (1984) Methodologies for the determination of various genetic effects in permeable strains of E. coli K-12 differing in DNA repair capacity, Mutation Res., 125, 153-184.
Moureau, P., A. Bailone and R. Devoret (1976) Prophage ~t induction in Escherichia coli K12 uvrA, uvrB: a highly sensitive test for potential carcinogens, Proc. Natl. Acad. Sci. USA, 73, 3700-3704.
Nohmi, T., A. Hakura, Y. Nakai, M. Watanabe, S.Y. Mu- rayama and T. Sofuni (1991) Salmonella typhimurium has two homologous but different umuDC operons: Cloning of a new umuDC-like operon (samAB) present in a 60-Mega- dalton cryptic plasmid of S. typhimurium, J. Bacteriol., 173, 1051-1063.
Nohmi, T., M. Yamada, Watanabe, M, S.Y. Murayama and T. Sofuni (1992) Roles of Salmonella typhimurium umuDC and samAB in UV mutagenesis and UV sensitivity, J. Bacteriol., 174, 6948-6955.
Oda, Y., S. Nakamura, I. Oki, T. Kato and H. Shinagawa (1985) Evaluation of the new system (umu-test) for the detection of environmental mutagens and carcinogens, Mutation Res., 147, 219-229.
Orrego, C., and E. Eisenstadt (1987) An inducible pathway is required for mutagenesis in Salmonella typhimurium LT2, J. Bacteriol., 169, 2885-2888.
Parker, C.T., A.W. Kloser, C.A. Schnaitman, M.A. Stein, S. Gottesman and B.W. Gibson (1992) Role of the rfaG and rfaP genes in determining the lipopolysaccharide core structure and cell surface properties of Escherichia coli, J. Bacteriol., 174, 2525-2538.
Quillardet, P., and M. Hofnung (1988) The screening, diagno- sis and evaluation of genotoxic agents with batteries of bacterial tests, Mutation Res., 205, 107-118.
Quillardet, P., O. Huisman, R. D'Ari and M. Hofnung (1985) The SOS chromotest, a colorimetric bacterial assay for genotoxins: validation study with 83 compounds, Mutation Res., 147, 79-95.
Rueff, J., A. Laires, J. Gaspar, H. Borba and A. Rodrigues (1992a) Oxygen species and the genotoxicity of quercetin, Mutation Res., 265, 75-81.
Rueff, J., A. Rodrigues, A. Laires and J. Gaspar (1992b)

M. Kranendonk et a l . / Mutation Research 312 (1994) 99-109 109
Activation of promutagens by porphyrinic biometic sys- tems, Mutation Res., 269, 243-250.
Ruiz-Vazquez, R., C. Pueyo and E. Cerda-Almedo (1978) A mutagen assay detecting forward mutations in an arabi- nose-sensitive strain of Salmonella typhimurium, Mutation Res., 54, 121-129.
Sambrook, J., E.F. Fritsch and T. Maniatis (1989) Molecular Cloning: A Laboratory Manual, 2nd edn., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Sanderson, K.E., and J.R. Roth (1988) Linkage map of Salmonella typhimuriurn, edn. VII, Microbiol. Rev., 52, 485-532.
Sedgwick, S.G., and P.A. Goodwin (1985) Differences in mutagenic and recombinational DNA repair in enterobac- teria, Proc. Natl. Acad. Sci. USA, 82, 4172-4176.
Sedgwick, S.G., C. Ho and R. Woodgate (1991a) Mutagenic DNA repair in enterobacteria, J. Bacteriol., 173, 5604- 5611.
Sedgwick, S.G., D. Lodwick, N. Doyle, H. Crowne and P. Strike (1991b) Functional complementation between chro- mosomal and plasmid mutagenic repair genes in bacteria, Mol. Gen. Genet., 229, 428-436.
Snyderwine, E.G., P.J. Wirth, P.P. Roller, R.H. Adamson, S. Sato and S.S. Thorgeirsson (1988) Mutagenicity and in vitro covalent DNA binding of 2-hydroxyamino-3-methyl- imidazolo[4,5-f]quinoline, Carcinogenesis, 9, 411-418.
Sommer, S., J. Knezevic, A. Bailone and R. Devoret (1993)
Induction of only one SOS operon, umuDC, is required for SOS mutagenesis in Escherichia coli, Mol. Gen. Genet., 239, 137-144.
Stout, V., and S. Gottesman (1990) RcsB and RscC: a two component regulator of capsule synthesis in Escherichia coli, J. Bacteriol., 172, 659-669.
Todd, P.A., N. Monti-Bragadi and B.W. Glickman (1979) MMS mutagenesis in strains of Escherichia coli carrying the R46 mutagenic enhancing plasmid: phenotypic analysis of arg + revertants, Mutation Res., 62, 227-237.
Trisler, P., and S. Gottesman (1984) Transcriptional regula- tion of genes necessary for capsular polysaccharide synthe- sis in Escherichia coli K-12, J. Bacteriol., 160, 184-191.
Venitt, S., C. Crofton-Sleigh and R. Forster (1984) Bacterial mutation assays using reverse mutation, in: S. Venitt and J.M. Parry (Eds.), Mutagenicity Testing, A Practical Ap- proach, Chapter 3, IRL Press, Oxford, pp. 45-98.
Walker, G.C. (1979) Mutagenesis and repair enhancing activi- ties associated with the plasmid pKM101, Cold Spring Harbor Symp. Quant. Biol., 43, 893-896.
Walker, G.C. (1984) Mutagenesis and inducible responses to deoxyribonuleic acid damage in Escherichia coli, Micro- biol. Rev., 48, 60-93.
Woodgate, R., A.S. Levine, W.H. Koch, T.A. Cebula and E. Eisenstadt (1991) Induction and cleavage of Salmonella typhimurium UmuD protein, Mol. Gen. Genet., 229, 81-85.




















