IOTEKNOLOGI & IOSAINS - Open Journal System BPPT
125
B p-ISSN 2442 – 2606 e-ISSN 2548 – 611X B JBBI Vol 5 No 1 Hal 1-110 June 2018 ISSN 2442 -2606 IOTEKNOLOGI & IOSAINS VOLUME 5 NOMOR 1 JUNE 2018 KARAKTERISTIK DAN SIFAT KINETIKA ENZIM KITINASE ASAL JAMUR ENTOMOPATOGEN Beauveria bassiana Nunung Eni Elawati, Sri Pujiyanto, Endang Kusdiyantini ANALISIS FILOGENETIK BEBERAPA KLON KARET DENGAN MARKA AFLP (AMPLIFIED FRAGMENT LENGTH POLYMORPHISM) Juniza Firdha Suparningtyas, Okky Dwi Pramudyawardhani, Devit Purwoko, Teuku Tajuddin DEKOLORISASI PEWARNA TEKSTIL SUMIFIX BLUE DAN REACTIVE RED 2 OLEH BAKTERI YANG DIISOLASI DARI LIMBAH INDUSTRI TEKSTIL Intan Permatasari, Rully Adi Nugroho, Vincentia Irene Meitiniarti KARAKTERISTIK FISIK PAKAN IKAN APUNG NON-EKSTRUSI YANG DIBUAT MELALUI FERMENTASI Rhizopus oryzae Asep Badru Zaman, Catur Sriherwanto, Etyn Yunita, Imam Suja'i PERBANYAKAN IN VITRO PISANG KEPOK var. UNTI SAYANG TAHAN PENYAKIT DARAH MELALUI PROLIFERASI TUNAS Maria Imelda, Aida Wulansari, Laela Sari OPTIMASI PROSES UNTUK EKSPRESI GEN ENDOGLUKANASE DARI Bacillus sp. RP1 OLEH Escherichia coli BL21 (DE3)/ egc Hans Victor, Maelita Ramdani Moeis AKTIVITAS Stenotrophomonas rhizophila DAN Trichoderma sp. DALAM MENGHAMBAT PERTUMBUHAN Ganoderma boninense Bedah Rupaedah, Debby Viola Amanda, Reni Indrayanti, Nia Asiani, Bambang Sukmadi, Asep Ali, Abdul Wahid, Taufik Firmansyah, Mahmud Sugianto PEMANFAATAN TUMBUHAN OBAT SECARA EMPIRIS PADA SUKU MANDAILING DI TAMAN NASIONAL BATANG GADIS SUMATERA UTARA Aswarina Nasution, Tatik Chikmawati, Eko Baroto Walujo, Ervizal A.M. Zuhud MULTIPLIKASI IN VITRO ANGGREK HITAM (Coelogyne pandurata Lindl.) PADA PERLAKUAN KOMBINASI NAA DAN BAP Roni Kartiman, Dewi Sukma, Syarifah Iis Aisyah, Agus Purwito ISOLASI DAN ANALISIS GENISTEIN DARI TEMPE BUSUK MENGGUNAKAN METODE KROMATOGRAFI KOLOM Hartati Soetjipto, Yohanes Martono, Zulfa Yuniarti ANALISIS BIOINFORMATIKA BERBASIS WEB PADA SEKUEN GENOM PARSIAL SAGU (Metroxylon sagu Rottb.) Devit Purwoko, Imam Civi Cartealy, Teuku Tajuddin, Diny Dinarti, Sudarsono Sudarsono
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of IOTEKNOLOGI & IOSAINS - Open Journal System BPPT

B p-ISSN 2442 – 2606
e-ISSN 2548 – 611X
B
JBBI Vol 5 No 1 Hal 1-110 June 2018 ISSN 2442 -2606
IOTEKNOLOGI &
IOSAINS VOLUME 5 NOMOR 1 JUNE 2018
KARAKTERISTIK DAN SIFAT KINETIKA ENZIM KITINASE ASAL JAMUR ENTOMOPATOGEN
Beauveria bassiana
Nunung Eni Elawati, Sri Pujiyanto, Endang Kusdiyantini
ANALISIS FILOGENETIK BEBERAPA KLON KARET DENGAN MARKA AFLP (AMPLIFIED FRAGMENT
LENGTH POLYMORPHISM)
Juniza Firdha Suparningtyas, Okky Dwi Pramudyawardhani, Devit Purwoko, Teuku Tajuddin
DEKOLORISASI PEWARNA TEKSTIL SUMIFIX BLUE DAN REACTIVE RED 2 OLEH BAKTERI
YANG DIISOLASI DARI LIMBAH INDUSTRI TEKSTIL
Intan Permatasari, Rully Adi Nugroho, Vincentia Irene Meitiniarti
KARAKTERISTIK FISIK PAKAN IKAN APUNG NON-EKSTRUSI YANG DIBUAT
MELALUI FERMENTASI Rhizopus oryzae
Asep Badru Zaman, Catur Sriherwanto, Etyn Yunita, Imam Suja'i
PERBANYAKAN IN VITRO PISANG KEPOK var. UNTI SAYANG TAHAN PENYAKIT DARAH
MELALUI PROLIFERASI TUNAS
Maria Imelda, Aida Wulansari, Laela Sari
OPTIMASI PROSES UNTUK EKSPRESI GEN ENDOGLUKANASE DARI Bacillus sp. RP1
OLEH Escherichia coli BL21 (DE3)/ egc
Hans Victor, Maelita Ramdani Moeis
AKTIVITAS Stenotrophomonas rhizophila DAN Trichoderma sp. DALAM MENGHAMBAT
PERTUMBUHAN Ganoderma boninense
Bedah Rupaedah, Debby Viola Amanda, Reni Indrayanti, Nia Asiani, Bambang Sukmadi, Asep Ali,
Abdul Wahid, Taufik Firmansyah, Mahmud Sugianto
PEMANFAATAN TUMBUHAN OBAT SECARA EMPIRIS PADA SUKU MANDAILING
DI TAMAN NASIONAL BATANG GADIS SUMATERA UTARA
Aswarina Nasution, Tatik Chikmawati, Eko Baroto Walujo, Ervizal A.M. Zuhud
MULTIPLIKASI IN VITRO ANGGREK HITAM (Coelogyne pandurata Lindl.) PADA PERLAKUAN
KOMBINASI NAA DAN BAP
Roni Kartiman, Dewi Sukma, Syarifah Iis Aisyah, Agus Purwito
ISOLASI DAN ANALISIS GENISTEIN DARI TEMPE BUSUK MENGGUNAKAN
METODE KROMATOGRAFI KOLOM
Hartati Soetjipto, Yohanes Martono, Zulfa Yuniarti
ANALISIS BIOINFORMATIKA BERBASIS WEB PADA SEKUEN GENOM PARSIAL
SAGU (Metroxylon sagu Rottb.)
Devit Purwoko, Imam Civi Cartealy, Teuku Tajuddin, Diny Dinarti, Sudarsono Sudarsono

i
VOLUME 5 NOMOR 1 JUNE 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
KATA PENGANTAR
Alhamdulillah berkat rahmatNya Jurnal Bioteknologi & Biosains Indonesia (JBBI) telah
berhasil mendapatkan Akreditasi B dari Kemenristekdikti pada tanggal 30 Oktober 2017
berdasarkan Surat Keputusan Direktur Jenderal Penguatan Riset dan Pengembangan
Nomor 48a/E/KPT/2017. Kini dengan semangat baru kami menghadirkan JBBI dengan
edisi Volume 4 Nomor 2 Desember 2017.
Rasa syukur yang dalam kami sampaikan ke hadirat Tuhan Yang Maha Esa, karena
berkat rahmat dan hidayahNya, kami dapat menghadirkan Jurnal Bioteknologi &
Biosains Indonesia (JBBI) Volume 5 Nomor 1 Juni 2018. Sertifikat akreditasi yang kami
terima dari Kemenristekdikti sejak tanggal 4 Desember 2017 yang lalu berdasarkan
Surat Keputusan Direktur Jenderal Penguatan Riset dan Pengembangan Nomor
51/E/KPT/2017, telah menjadi titik tolak bagi kami untuk meningkatkan kualitas JBBI.
JBBI telah mulai dikenal oleh berbagai komunitas ilmiah sebagai media komunikasi
yang mempublikasi naskah atau tulisan dalam bidang bioteknologi dan biosains,
mencakup hasil-hasil kerekayasaan dan penelitian mutakhir. Penyaringan naskah-
naskah yang masuk pun kini telah dapat dilakukan dengan lebih ketat.
Pada edisi kali ini JBBI menampilkan 11 tulisan hasil riset dari berbagai institusi dan
perguruan tinggi. Kami menyadari bahwa jurnal ini masih jauh dari sempurna, oleh
karena itu perlu pembenahan. Segala kritik, saran dan himbauan dari semua pihak
yang bersifat membangun selalu kami harapkan demi kesempurnaan penerbitan pada
edisi mendatang.
Redaksi

ii
VOLUME 5 NOMOR 1 JUNE 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
UCAPAN TERIMA KASIH
Ucapan terima kasih dan penghargaan yang setinggi-tingginya kami sampaikan kepada
para Mitra Bestari, yang telah memberikan sumbang sarannya dalam menelaah
naskah-naskah yang masuk. Kesungguhan para Mitra Bestari inilah yang telah
memungkinkan kami meningkatkan kualitas dan menjaga mutu penulisan pada
penerbitan Jurnal Bioteknologi & Biosains Indonesia (JBBI) edisi Bulan Juni 2018 ini:
1. Agustin Krisna Wardani
2. Anis Herliyanti Mahsunah
3. Churiyah
4. Diana Dewi
5. Dr. Dudi Hardianto
6. Dr. Ir. Elok Zubaidah, MP
7. Dr. Erwahyuni Endang Prabandari
8. Juwartina Ida Royani, M.Si
9. Marwan Diapari, PhD
10. Prof. Dra. Netty Widyastuti, M.Si
11. Dr. Ratu Siti Aliah
12. Rinaldi Sjahril
13. Dr. Riza Arief Putranto, DEA
14. Dr. Rofiq Sunaryanto, S.Si, M.Si
15. Prof. Dr. Sismindari, SU., Apt
16. Prof. Dr. Suyanto Pawiroharsono, DEA
17. Tia Setiawati
Kepada Tim Teknis yang telah bekerja keras dalam proses penerbitan, mulai dari
editing tata bahasa, layout halaman, disain cover dan pekerjaan-pekerjaan teknis lain
sehingga edisi ini terwujud juga disampaikan terima kasih dan penghargaan yang
sebesar-besarnya. Tanpa kerja keras mereka upaya penerbitan edisi ini mungkin akan
terkendala. Terakhir kami sampaikan terima kasih kepada penulis yang dengan
komitmen, pemikiran dan kerja kerasnya telah berkontribusi pada edisi Bulan Juni 2018.
Redaksi

iii
VOLUME 5 NOMOR 1 JUNE 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
DAFTAR ISI
KATA PENGANTAR
i
UCAPAN TERIMA KASIH
ii
KARAKTERISTIK DAN SIFAT KINETIKA ENZIM KITINASE ASAL JAMUR
ENTOMOPATOGEN Beauveria bassiana
1-7
Nunung Eni Elawati, Sri Pujiyanto, Endang Kusdiyantini
ANALISIS FILOGENETIK BEBERAPA KLON KARET DENGAN MARKA
AFLP (AMPLIFIED FRAGMENT LENGTH POLYMORPHISM)
8-19
Juniza Firdha Suparningtyas, Okky Dwi Pramudyawardhani, Devit Purwoko, Teuku
Tajuddin
DEKOLORISASI PEWARNA TEKSTIL SUMIFIX BLUE DAN REACTIVE
RED 2 OLEH BAKTERI YANG DIISOLASI DARI LIMBAH INDUSTRI
TEKSTIL
20-26
Intan Permatasari, Rully Adi Nugroho, Vincentia Irene Meitiniarti
KARAKTERISTIK FISIK PAKAN IKAN APUNG NON-EKSTRUSI YANG
DIBUAT MELALUI FERMENTASI Rhizopus oryzae
27-35
Asep Badru Zaman, Catur Sriherwanto, Etyn Yunita, Imam Suja'i
PERBANYAKAN IN VITRO PISANG KEPOK var. UNTI SAYANG TAHAN
PENYAKIT DARAH MELALUI PROLIFERASI TUNAS
36-43
Maria Imelda, Aida Wulansari, Laela Sari
OPTIMASI PROSES UNTUK EKSPRESI GEN ENDOGLUKANASE
DARI Bacillus sp. RP1 OLEH Escherichia coli BL21 (DE3)/ egc
44-52
Hans Victor, Maelita Ramdani Moeis
AKTIVITAS Stenotrophomonas rhizophila DAN Trichoderma sp. DALAM
MENGHAMBAT PERTUMBUHAN Ganoderma boninense
53-63
Bedah Rupaedah, Debby Viola Amanda, Reni Indrayanti, Nia Asiani, Bambang
Sukmadi, Asep Ali, Abdul Wahid, Taufik Firmansyah, Mahmud Sugianto

iv
PEMANFAATAN TUMBUHAN OBAT SECARA EMPIRIS PADA SUKU
MANDAILING DI TAMAN NASIONAL BATANG GADIS SUMATERA
UTARA
64-74
Aswarina Nasution, Tatik Chikmawati, Eko Baroto Walujo, Ervizal A.M. Zuhud
MULTIPLIKASI IN VITRO ANGGREK HITAM (Coelogyne pandurata
Lindl.) PADA PERLAKUAN KOMBINASI NAA DAN BAP
75-87
Roni Kartiman, Dewi Sukma, Syarifah Iis Aisyah, Agus Purwito
ISOLASI DAN ANALISIS GENISTEIN DARI TEMPE BUSUK
MENGGUNAKAN METODE KROMATOGRAFI KOLOM
88-97
Hartati Soetjipto, Yohanes Martono, Zulfa Yuniarti
ANALISIS BIOINFORMATIKA BERBASIS WEB PADA SEKUEN GENOM
PARSIAL SAGU (Metroxylon sagu Rottb.)
98-107
Devit Purwoko, Imam Civi Cartealy, Teuku Tajuddin, Diny Dinarti, Sudarsono
Sudarsono
INDEKS KATA KUNCI
108-109
INDEKS PENGARANG 110

1
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
KARAKTERISTIK DAN SIFAT KINETIKA ENZIM KITINASE ASAL
JAMUR ENTOMOPATOGEN Beauveria bassiana
Characteristics and Kinetics of Chitinase Enzyme from Entomopathogenic Fungus Beauveria bassiana
Nunung Eni Elawati*, Sri Pujiyanto, Endang Kusdiyantini Departemen Biologi, Fakultas Sains dan Matematika, Universitas Diponegoro
Jl. Prof. Soedharto, SH, Tembalang, Semarang 50275, Indonesia *E-mail: [email protected]
ABSTRACT
Beauveria bassiana is one of the entomopathogenic fungi that produces chitinase when
infecting the host. Chitinase is widely used as biocontrol agents because it can degrade
chitin into an environmentally friendly product. This study aims to characterize and test the kinetics of chitinase from B. bassiana. This characterization includes determination of pH and
optimum temperature, enzyme stability and enzyme kinetics test by determining Km and Vmax
value with Lineweaver-Burk equations. The result of experiment showed that the chitinase B.
bassiana had pH and optimum temperature of 5 and 40ºC respectively. This enzyme was
stable until 90 minutes incubation at 40ºC. The Km and Vmax values were 0.181 mg/L and
0.022 mg/L.sec respectively. The Km value is higher than Vmax, which means the affinity of
the enzyme to the lower substrate requiring high substrate concentration to increase the
reaction rate. It can be concluded that the chitinase activity of B. bassiana is still low.
Keywords: Beauveria bassiana, characteristics and kinetics, chitinase enzyme,
entomopathogenic, Lineweaver-Burk
ABSTRAK
Beauveria bassiana merupakan salah satu jamur entomopatogen yang memproduksi
kitinase saat menginfeksi inangnya. Enzim kitinase saat ini banyak digunakan sebagai agen
biokontrol karena dapat mendegradasi kitin menjadi produk yang ramah lingkungan.
Penelitian ini bertujuan untuk mengkarakterisasi dan menguji kinetika enzim kitinase asal jamur B. bassiana. Metode yang digunakan dalam karakterisasi ini mencakup penentuan pH
dan suhu optimum, kestabilan enzim pada suhu optimumnya, dan uji kinetika enzim yang mencakup penentuan nilai Km dan Vmaks dengan persamaan Lineweaver-Burk. Hasil
penelitian karakterisasi menunjukkan bahwa enzim kitinase B. bassiana mempunyai pH dan
suhu optimum masing-masing 5 dan 40ºC. Enzim ini stabil sampai pada 90 menit inkubasi
pada suhu 40ºC. Nilai Km diperoleh 0,181 mg/L dan Vmaks sebesar 0,022 mg/L.detik. Nilai Km
lebih tinggi daripada Vmaks, yang artinya afinitas enzim terhadap substrat rendah sehingga
membutuhkan konsentrasi substrat yang tinggi untuk meningkatkan kecepatan reaksi, maka dapat disimpulkan bahwa aktivitas kitinase dari B. bassiana masih tergolong rendah.
Kata kunci: Beauveria bassiana, entomopatogen, enzim kitinase, karakteristik dan kinetik, Lineweaver-Burk
Received: 12 January 2018 Accepted: 16 March 2018 Published: 05 June 2018

Karakteristik Dan Sifat Kinetika Enzim Kitinase... Elawati et al.
2
PENDAHULUAN
Kitinase merupakan salah satu enzim
yang memiliki nilai ekonomi tinggi. Kitinase mampu menghidrolisis ikatan β-1,4-asetamido-2-deoksi-D-glikosida pada kitin dan oligomer kitin (Jung dan Park 2014). Enzim kitinase saat ini banyak digunakan sebagai agen biokontrol karena dapat mendegradasi kitin menjadi produk yang ramah lingkungan dan dapat digunakan dalam bidang kesehatan, pangan, industri dan lain-lain (Pratiwi et al. 2015). Peranan enzim kitinase yang sangat prospektif terhadap kehidupan masyarakat banyak mendorong ilmuwan dan peneliti melakukan eksplorasi mikroorganisme kitinolitik, yaitu mikroorganisme yang dapat mendegradasi kitin dengan menggunakan enzim kitinase.
Enzim kitinase dihasilkan oleh bakteri, fungi, tanaman, dan hewan (Nguyen et al. 2015). Salah satu mikroorganisme penghasil kitinase berasal dari jamur yaitu Beauveria bassiana (de Carolina Sanchez-Perez et al. 2014). Jamur ini sejak lama diketahui memiliki potensi sebagai agen pengendali hayati yang dapat mengendalikan populasi serangga hama, sehingga banyak dikembangkan sebagai agensia hayati dalam bidang pertanian (Suryadi et al. 2013). B. bassiana dalam menginfeksi serangga dengan menembus kutikula inangnya dengan cara mensekresikan kitinase untuk mendegradasi komponen utama dari kutikula dan dilanjutkan dengan penetrasi hifa (de Carolina Sanchez-Perez et al. 2014).
Berbagai organisme kitinolitik dapat menghasilkan beragam jenis kitinase dengan karakteristik dan spesifitas terhadap substrat yang bervariasi, oleh karena itu perlu pengujian tentang karakteristik kitinase asal B. bassiana serta kinetika enzimnya
agar saat aplikasi di lapangan dapat optimal. Tujuan dari penelitian ini adalah mengkarakterisasi dan menguji kinetika enzim kitinase asal B. bassiana. BAHAN DAN METODE
Bahan yang digunakan dalam penelitian ini adalah: Jamur B. bassiana,
Kitin, Potato Dextrose Agar (PDA), bacto agar, HCl pekat, NaOH, NaCl, NH4Cl, MgSO4·7H2O, K2HPO4, CaCl2,
Na2HPO4·2H2O, (NH4)2SO4, buffer sitrat pH 4 & 5, DNS, buffer fosfat pH 6-8, buffer glisin-NaOH pH 9, yeast extract. Penyiapan jamur B. bassiana
Jamur B. bassiana yang digunakan adalah hasil isolasi dari serangga Helopeltis antonii, yang telah terinfeksi B. bassiana
pasca aplikasi di Perkebunan Kakao Temanggung, kemudian ditumbuhkan pada medium PDA. Peremajaan dilakukan dengan mengambil sebagian kecil area agar yang telah ditumbuhi oleh B. bassiana selanjutnya
diinokulasikan pada stok agar miring. Tabung reaksi diinkubasi pada suhu ruang selama 7 hari. Produksi enzim kitinase
Dua ose kultur jamur B. bassiana
diinokulasikan di dalam medium kitin cair yang mengandung 125 mL koloidal kitin 0,3%, 0,65 g Na2HPO4·2H2O, 1,5 g KH2PO4, 0,25 g NaCl, 0,5 g NH4Cl, 0,12 g MgSO4·7H2O, dan 0,005 g CaCl2. Kultur diinkubasi selama 5 hari di dalam inkubator bergoyang (120 rpm) pada suhu ruang. Sebanyak 10 mL kultur jamur B. bassiana (+5.000 spora/mL) dipipet dan diinokulasikan ke dalam 1.000 mL medium kitin cair dengan teknik aseptik. Kultur diinkubasi di dalam inkubator bergoyang pada suhu ruang selama 5 hari. Kultur disentrifus dengan kecepatan 10.000 rpm selama 5 menit. Supernatan yang dihasilkan adalah ekstrak enzim kitinase (Suryadi et al. 2013). Pengujian aktivitas kitinase
Penentuan aktivitas enzim dilakukan dengan metode Ueda dan Arai (1992), dengan membuat sampel 1 mL 0,3 % larutan koloid kitin (substrat) ditambah 2 mL 0,2 M buffer fosfat pH 7 dan 1 mL supernatan kitinase (enzim) dimasukkan ke dalam tabung reaksi dan diinkubasi pada suhu 37ºC selama 1 jam. Kontrol dibuat dengan komponen sama seperti sampel namun sebelum inkubasi diinaktivasi terlebih dahulu pada suhu 100ºC selama 10 menit. Sampel dan kontrol kemudian diukur absorbansinya menggunakan spektrofotometer pada λ 420 nm. Aktivitas kitinase ditentukan secara turbidimetri. Satu unit aktivitas kitinase didefinisikan sebagai jumlah kitinase yang menyebabkan pengurangan absorbansi sebesar 0,001 campuran reaksi per menit.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
3
Unit aktivitas kitinase dinyatakan dengan rumus:
(U/mL) =
X
Keterangan Abs kontrol: absorbansi kontrol (substrat + enzim
yang diinaktivasi) Abs sampel: absorbansi sampel (substrat + enzim
yang tidak diinaktivasi)
Karakterisasi enzim kitinase
Karakterisasi enzim dilakukan terhadap: suhu dan pH optimum, stabilitas enzim pada suhu optimalnya dan kinetika enzim. Penetapan pH optimum
Penetapan pH optimum dilakukan dengan cara mereaksikan enzim dengan substrat koloid kitin 0,5% pada suhu 37ºC selama 30 menit pada berbagai kondisi pH larutan buffer 4 sampai 9. Buffer pH 4 dan 5 dibuat dari buffer sitrat fosfat, pH 6-8 dari buffer fosfat, dan buffer pH 9 dibuat dari buffer glisin-NaOH. Konsentrasi buffer yang digunakan adalah 50 mM. Penetapan suhu optimum
Penetapan suhu optimum dilakukan dengan cara menginkubasi sistem reaksi enzimatis pada rentang suhu uji (25, 30, 35, 40, 45, 50, 55, dan 60ºC) selama 30 menit. Sedangkan pH yang digunakan adalah hasil pengujian pH optimum. Penetapan waktu inkubasi optimum
Penetapan waktu inkubasi optimum dilakukan dengan mereaksikan enzim pada rentang waktu inkubasi 15, 30, 60, 120, 150 menit. Inkubasi dilakukan pada buffer pH optimal dan suhu optimal. Studi kinetika enzim
Studi kinetika enzim dilakukan dengan mereaksikan enzim, buffer fosfat pH 7 dan substrat kitin dari konsentrasi 0,1 – 1,0% dengan interval 0,2% dalam volume reaksi 600 μL. Campuran kemudian diinkubasi pada suhu 37ºC selama 30 menit. Aktivitas enzim kitinase dilakukan dengan menghubungkan konsentrasi substrat dengan konsentrasi N-asetil-D-glukosamin yang diperoleh dari kurva standar N-asetil-D-glukosamin sehingga didapatkan nilai
kecepatan. Nilai Km dan Vmaks ditentukan berdasarkan persamaan Lineweaver-Burk
yang merupakan grafik hubungan antara 1/[S] dan 1/V. HASIL DAN PEMBAHASAN Hasil kultivasi dan pertumbuhan jamur
Isolasi B. bassiana dilakukan pada
medium PDA yang diinkubasi selama 7 hari. Hasil pengamatan morfologi B. bassiana
disajikan pada Gambar 1. Biakan B. bassiana pada medium PDA
mempunyai miselia dan konidia berwarna putih. Konidium dihasilkan oleh biakan yang telah bersporulasi yang berbentuk bulat, yang membentuk kumpulan seperti tepung. Konidia diproduksi di atas konidiofor yang berbentuk seperti botol. Menurut Kulu et al.
Gambar 1. Morfologi B. bassiana. (A) pertumbuhan B.
bassiana pada medium PDA inkubasi 5 hari. (B) struktur mikroskopis B. bassiana perbesaran 40x10. a: konidia, b: hifa.
A
B
a
b

Karakteristik Dan Sifat Kinetika Enzim Kitinase... Elawati et al.
4
(2015), ciri-ciri B. bassiana secara morfologi
koloni berwarna putih, tekstur lembut seperti serbuk. Karakter mikroskopis B. bassiana
memiliki miselium yang bersekat dan berwarna putih, konidiofor bercabang dan berpola zig-zag. Spora berbentuk bulat, bening (hialin), bersel satu tanpa sekat. Spora muncul dari setiap percabangan konidiofor.
Pengaruh pH
Derajat keasaman (pH) menjadi faktor yang mempengaruhi aktivitas enzim (Purkan et al. 2016). Pengaruh pH terhadap aktivitas enzim dapat dilihat pada Gambar 2.
Aktivitas maksimum enzim kitinase dicapai pada buffer sitrat fosfat pH 5 karena menunjukkan absorbansi yang terbesar dibanding yang lain dengan aktivitas enzim 1,15 (U/mL). Hasil ini sejalan dengan yang dikemukakan oleh Karthik et al. (2014), secara umum jamur bekerja pada pH asam untuk memproduksi kitinase yaitu dibawah pH 6. Lebih lanjut penelitian oleh Wang et al. (2015) melaporkan bahwa enzim kitinase stabil pada pH 5 dengan aktivitas enzim residu sebesar 70%. Aktivitas kitinase yang tinggi pada pH optimum disebabkan oleh terjadinya ionisasi asam-asam amino pada sisi aktif enzim, sehingga terjadi interaksi yang optimum antara enzim dengan substrat (Rahmawati 2016).
Aktivitas kitinase setelah pH optimum mengalami penurunan, hal ini disebabkan adanya perubahan muatan ion pada rantai samping yang terionisasi sehingga mengakibatkan terjadinya denaturasi enzim yang disertai hilangnya aktivitas katalitik enzim. Adanya perubahan struktur tersier menyebabkan kelompok hidrofobik kontak dengan air sehingga solubilitas enzim
menjadi berkurang yang mengakibatkan turunnya aktivitas enzim secara bertahap (Suryadi et al. 2013).
Hasil analisis statistik ANOVA menunjukkan bahwa perlakuan pH berpengaruh nyata (P<0,05) terhadap aktivitas enzim kitinase dari B. bassiana. Setelah dilakukan uji Duncan dengan taraf nyata α 0,05 terhadap pH menunjukkan bahwa pada pH 5 aktivitas kitinase berbeda nyata terhadap perlakuan pH lain yang diujikan. Hal ini dibuktikan dengan nilai p-value >0.05.
Pengaruh suhu
Setiap enzim memiliki kisaran suhu tertentu untuk mencapai aktivitas yang optimum. Rentang suhu yang diujikan dimulai dari 25ºC sampai 60ºC selama 30 menit. Hasil pengaruh suhu terhadap aktivitas kitinase disajikan pada Gambar 3.
Aktivitas kitinase mengalami penurunan setelah mendapatkan kondisi suhu optimum yaitu suhu 40ºC. Hal ini terjadi karena enzim merupakan jenis protein yang dapat mengalami denaturasi pada suhu tinggi. Denaturasi ini menyebabkan perubahan pada konformasi enzim akibat adanya perenggangan ikatan hidrogen yang bersifat reversibel sehingga dapat mempengaruhi sisi aktif enzim untuk berikatan dengan substrat (Haedar et al. 2017). Penelitian Wang et al. (2015) melaporkan bahwa kisaran suhu optimal untuk aktivitas kitinase kisaran 40 – 85ºC.
Aktivitas enzim kitinase menurun setelah suhu 40ºC, sampai dengan suhu 50ºC kemudian pada suhu 55ºC kembali meningkat sedikit, hal ini diduga adanya peningkatan suhu inaktivasi yang meningkat, sehingga aktivitasnya masih terlihat
Gambar 3. Grafik pengaruh suhu terhadap aktivitas
kitinase
b b.c
c
d
b.c
a a
a
0.00
0.10
0.20
0.30
0.40
25 30 35 40 45 50 55 60
Aktivitas e
nzim
(U
/mL)
Suhu (°C)
,
,
,
,
,
Gambar 2. Grafik pengaruh berbagai pH terhadap
aktivitas kitinase
c
e
d
b
a a
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
4 5 6 7 8 9
Aktivitas e
nzim
(U
/mL)
pH
4
6
9
,
,
,
,
,
,
,
,
Buffer sitrat
Buffer fosfat Buffer glisin

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
5
meningkat. Meskipun demikian, berdasarkan pengujian statistik dengan Duncan, pada suhu 50ºC, 55ºC, dan 60ºC tidak memiliki perbedaan yang nyata, sehingga dapat diartikan perlakuan ketiga suhu tersebut sama dan tidak berpengaruh terhadap naik turunnya aktivitas enzim.
Analisis statistik ANOVA menunjukkan bahwa perlakuan suhu berpengaruh nyata (P<0,05) terhadap aktivitas enzim kitinase dari B. bassiana. Setelah dilakukan uji
Duncan dengan taraf nyata α 0,05 terhadap suhu, menunjukkan bahwa pada suhu 40ºC aktivitas kitinase berbeda nyata terhadap perlakuan suhu lain yang diujikan. Hal ini dibuktikan dengan nilai p-value >0,05.
Pengaruh waktu inkubasi
Semua enzim memiliki kemampuan untuk mempertahankan stabilitas sisi aktifnya dalam waktu tertentu. Kemampuan ini bersifat spesifik dan berbeda antara enzim satu dengan enzim yang lain (Noviendri et al. 2008). Pengujian stabilitas waktu inkubasi terhadap aktivitas enzim kitinase dilakukan dari rentang waktu inkubasi 15 hingga 120 menit dengan pH 5 dan suhu 40ºC yang merupakan pH dan suhu optimum. Hasil uji stabilitas enzim kitinase terhadap waktu inkubasi disajikan pada Gambar 4.
Aktivitas enzim kitinase tertinggi dicapai pada waktu inkubasi 90 menit dengan aktivitas enzim kitinase sebesar 0,096 U/mL. Aktivitas enzim meningkat sejak 15 menit inkubasi hingga mencapai aktivitas optimum pada 90 menit inkubasi. Aktivitas kitinase menurun setelah waktu inkubasi optimum yaitu pada waktu inkubasi 120 menit dengan aktivitas enzim sebesar 0,064 U/mL. Waktu inkubasi berpengaruh terhadap jumlah produk hidrolisis enzim
yang dihasilkan. Berdasarkan analisis statistik ANOVA
tampak bahwa perlakuan waktu inkubasi berpengaruh nyata (P<0,05) terhadap aktivitas kitinase dari B. bassiana. Uji
Duncan dengan taraf nyata α 0,05 menunjukkan bahwa pada waktu inkubasi 90
menit aktivitas kitinase berbeda nyata terhadap perlakuan waktu inkubasi lain yang diujikan. Hal ini dibuktikan dengan nilai p-value >0,05.
Kinetika enzim
Aktivitas enzim juga dipengaruhi oleh konsentrasi substrat yang digunakan. Semakin tinggi konsentrasi substrat yang digunakan, maka semakin tinggi aktivitas enzim atau semakin tinggi kecepatan reaksi yang dikatalisis oleh enzim tersebut. Namun pada suatu titik tertentu, yaitu kecepatan maksimum (Vmaks), penambahan konsentrasi substrat dalam jumlah tertentu tidak akan dapat meningkatkan kecepatan reaksi enzim, melainkan dapat menurunkan aktivitas enzim atau kecepatan reaksi. Dalam hal ini, substrat yang ditambahkan tersebut akan menjadi inhibitor dalam reaksi enzimatik (Noviendri et al. 2008).
Kinetika enzim dapat diketahui dengan menentukan nilai Km dan Vmaks. Penelitian ini menggunakan persamaan Lineweaver-Burk untuk menentukan nilai Km dan Vmaks. Penggunaan persamaan Lineweaver-Burk
memiliki keuntungan karena lebih mudah secara matematis. Hasil penentuan kinetika enzim kitinase disajikan dalam bentuk kurva double reciprocal Lineweaver-Burk pada Gambar 5.
Penentuan nilai Km dan Vmaks diperoleh dari persamaan linier y=8,216x+45,194. Persamaan ini dihitung berdasarkan pengukuran konsentrasi N-asetilglukosamin
Gambar 5. Kurva double reciprocal Lineweaver-Burk
-40
-20
0
20
40
60
80
100
120
140
-10 -5 0 5 10 15
1/V
1/S 1/Km
1/Vmaks
Slope=
Km/Vma
ks
Gambar 4. Grafik pengaruh waktu inkubasi terhadap
aktivitas kitinase
a
b a.b
b
c
0.00
0.02
0.04
0.06
0.08
0.10
0.12
15 30 60 90 120
Aktivitas e
nzim
(U
/mL)
Waktu inkubasi (menit)
,
, ,
,
,
,
,

Karakteristik Dan Sifat Kinetika Enzim Kitinase... Elawati et al.
6
(GlcNAc) sebagai produk hidrolisis substrat koloid kitin pada berbagai konsentrasi selama waktu inkubasi 30 menit. Berdasarkan perhitungan persamaan tersebut diperoleh nilai Vmaks=0,022 mg/L.detik dan nilai Km=0,181 mg/L. Nilai Vmaks 0,022 mg/L didefinisikan sebagai suatu kondisi optimum enzim kitinase B. bassiana
dapat mengubah substrat koloid kitin menjadi monomer N-asetil-D-glukosamin sebesar 0,022 mg/L setiap detiknya. Nilai Km yang dihasilkan lebih tinggi daripada Vmaks, hal ini berarti afinitas enzim terhadap substrat rendah sehingga membutuhkan konsentrasi substrat yang tinggi untuk meningkatkan kecepatan reaksi. Hasil penelitian di Cina (Zhang et al. 2004) melaporkan produksi kitinase dari B. bassiana isolat BB 174 pada kondisi
formulasi padat mempunyai nilai Km 0,52 mg/mL dan Vmaks sebesar 0,70 mg/L.detik. Penelitian lain oleh Nguyen et al. (2015) melaporkan nilai Km dan Vmaks kitinase dari Lecanicillium lecanii 43H masing-masing sebesar 0,82 mg/mL dan 4,51 U/mg, kemudian Ma et al. (2012) melaporkan kitinase dari Gliocladium catenulatum HL11 mempunyai nilai Km 0,82
mg/mL dan Vmaks 2,832 mg/mL. Jika dibandingkan dengan penelitian sebelumnya, kinetika enzim yang dihasilkan oleh B. bassiana pada penelitian ini lebih
rendah. Hasil aktivitas kitinase yang didapatkan tidak sesuai dengan harapan. Hal ini dimungkinkan karena beberapa faktor antara lain pH, suhu, konsentrasi substrat, dan komponen medium yang kurang sesuai dengan jenis enzim kitinase yang dihasilkan oleh B. bassiana pada penelitian ini,
mengingat jenis enzim kitinase yang diproduksi oleh jamur terdapat 3 subgrup yaitu grup A, B, C masing-masing memiliki spesifitas yang berbeda (Karthik et al. 2014).
KESIMPULAN
Enzim kitinase yang dihasilkan oleh B. bassiana mempunyai suhu optimum 40ºC
dan pH optimum 5 dalam buffer sitrat fosfat. Enzim ini stabil pada waktu inkubasi 90 menit. Enzim kitinase memiliki nilai Km sebesar 0,181 mg/L dan Vmaks sebesar 0,022 mg/L.detik. Nilai Km lebih tinggi daripada Vmaks, hal ini berarti afinitas enzim terhadap substrat rendah sehingga membutuhkan
konsentrasi substrat yang tinggi untuk meningkatkan kecepatan reaksi. DAFTAR PUSTAKA de Carolina Sanchez-Perez L, Barranco-
Florido JE, Rodriguez-Navarro S, Cervantes-Mayagoitia JF, Ramos-Lopez MA (2014) Enzymes of entomopathogenic fungi, advances and insights. Adv Enzyme Res 2: 65-76. Doi: 10.4236/aer.2014.22007
Haedar N, Natsir H, Fahruddin, Aryanti W (2017) Produksi dan karakterisasi enzim kitinase dari bakteri kitinolitik asal kerang Anadara granosa. J Ilmu
Alam Ling 8: 14-21. Jung WJ, Park RD (2014) Bioproduction of
chitooligosaccharides: Present and perspectives. Mar Drugs 12:5328-5356. Doi: 10.3390/md12115328
Karthik N, Akanksha K, Binod P, Pandey A (2014) Production, purification and properties of fungal chitinase – A Review. Indian J Exp Biol 52: 1025-1035.
Kulu IP, Abadi AL, Afandhi A, Nooraidawati (2015) Morphological and molecular identification of Beauveria bassiana as
entomopathogen agent from Central Kalimantan peatland, Indonesia. Int J ChemTech Res 8:2079-2084.
Ma GZ, Gao HL, Zhang YH, Li SD, Xie BY, Wu SJ (2012) Purification and characterization of chitinase from Gliocladium catenulatum strain HL-1-1.
Afr J Microbiol Res 6: 4377–4383. Doi: 10.5897/AJMR12.605
Nguyen HQ, Quyen DT, Nguyen SLT, Vu VH (2015) An extracellular antifungal chitinase from Lecanicillium lecanii: purification, properties, and application in biocontrol against plant pathogenic fungi. Turk J Biol 39:6-14. Doi: 10.3906/biy-1402-28
Noviendri D, Fawzya YN, Chasanah E (2008) Karakteristik dan sifat kinetika enzim kitinase dari isolat bakteri T5a1 asal terasi. J Pascapanen Bioteknol Kelautan dan Perikanan 3:123-129.
Pratiwi RS, Susanto TE, Wardani YAK, Sutrisno A (2015) Enzim kitinase dan aplikasi di bidang industri: Kajian Pustaka. J Pangan Agroindustri 3: 878-887.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
7
Purkan P, Baktir A, Sayyidah AR (2016) Produksi Enzim kitinase dari Aspergillus niger menggunakan limbah
cangkang rajungan sebagai induser. J Kimia Riset 1: 34 – 41. Doi: 10.20473/jkr.v1i1.2440
Rahmawati H, Purnomo AJ, Umniyatie S, Pramiadi D, Sari N (2016) Identification and characterization of chitinase enzyme producing bacteria from bat guano and its potential to inhibit the growth of fungus Colletotrichum sp. cause anthracnose on the chili by in vitro. Int J Adv Agricultural & Environmental Engg 3: 249-254. Doi: 10.15242/IJAAEE.U0516208
Suryadi Y, Priyatno TP, Susilowati DN, Samudra IM, Yudhistira N, Purwakusumah ED (2013) Isolasi dan
karakterisasi kitinase asal Bacillus cereus 11 UJ. J Biol Indon 9: 51-62.
Doi: 10.14203/jbi.v9i1.146 Ueda M, Arai M (1992) Purification and
some properties of chitinase from Aeromonas sp. No. 10S-24. Biosci
Biotech Biochem 56: 460-464. Doi: 10.1271/bbb.56.460
Wang J, Zhang J, Song F, Gui T, Xiang J (2015) Purification and characterization of chitinases from ridgetail white prawn Exopalaemon carinicauda. Molecules 20:1955-1967.
Doi: 10.3390/molecules20021955 Zhang J, Cai J, Wu K, Jin S, Pan R, Fan M
(2004) Production and properties of chitinase from Beauveria bassiana Bb174 in solid state fermentation. Chin J Appl Ecol 5: 863-866.

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
8
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
ANALISIS FILOGENETIK BEBERAPA KLON KARET DENGAN MARKA
AFLP (AMPLIFIED FRAGMENT LENGTH POLYMORPHISM)
Phylogenetic Analysis of Rubber Tree Clones using AFLP (Amplified Fragment Length Polymorphism) Marker
Juniza Firdha Suparningtyas
1, Okky Dwi Pramudyawardhani
2, Devit Purwoko
3, Teuku Tajuddin
3*
1Departemen Biokimia FMIPA Institut Pertanian Bogor. Jl Meranti, Kampus IPB Dramaga, Bogor 16680
2Bioteknologi FST Universitas Al Azhar Indonesia. Jl Sisingamangaraja Jakarta Selatan 12110
3Balai Bioteknologi BPPT. Gedung 630 Kawasan PUSPIPTEK, Tangerang Selatan, Banten 15314
*Email: [email protected] ABSTRACT National rubber productivity is lower than other rubber producing countries in the world. The DNA marker-based plant breeding program is required to increase latex production and other superior characters. Breeding process by identifying genetic diversity can be done using AFLP marker. The aim of this study was to analyze the phylogenetic six rubber clones. Pre-amplification and amplification process were performed using 64 primer pair combinations followed by electrophoresis in 6% polyacrylamide gel. A total of 2806 AFLP fragments have been detected. Phylogenetic tree of six clones showed 60% of genetic similarity, which was consisted of two groups. GT 1 clone in the first group and was separated from other clones in the second group. Sub-group at the phylogenetic peak contained IRR 104 and RRIM 600 clones with genetic similarity of 74%. The information obtained from this study showed the genetic diversity of the six rubber clones. The unique bands were obtained as marker specific which could be used to identify clones. Keywords: AFLP, DNA marker, phylogenetic tree, polymorphism, rubber clones ABSTRAK Produktivitas karet nasional masih lebih rendah dibanding dengan negara-negara penghasil karet lainnya di dunia. Diperlukan program pemuliaan tanaman karet yang berbasis marka DNA untuk meningkatkan produksi lateks serta karakter unggul lainnya. Proses pemuliaan untuk mengidentifikasi keragaman genetik bisa dilakukan dengan marka AFLP. Tujuan studi ini adalah untuk menganalisis filogenetik enam klon karet berdasarkan produktivitasnya. Proses pre-amplifikasi dan amplifikasi dilakukan dengan menggunakan 64 kombinasi pasangan primer yang dilanjutkan dengan analisis hasil elektroforesis pada gel poliakrilamid 6%. Sebanyak 2806 pita AFLP telah dihasilkan. Kekerabatan keenam klon menunjukkan kemiripan genetik sebesar 60%. Pohon filogenetik membentuk dua kelompok, yang memisahkan GT 1 dengan klon-klon lainnya. Sub-kelompok pada puncak filogenetik terdiri dari klon IRR 104 dan RRIM 600 dengan kemiripan genetik sebesar 74%. Informasi yang diperoleh telah menunjukkan keragaman genetik keenam klon karet yang bersifat polimorfis. Diperoleh pita-pita unik dari pasangan primer tertentu yang dapat berperan sebagai marka spesifik dan dapat digunakan untuk mengidentifikasi klon. Kata Kunci: AFLP, klon karet, marka DNA, pohon filogenetik, polimorfisme
Received: 03 November 2017 Accepted: 02 May 2018 Published: 06 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
9
PENDAHULUAN
Tanaman karet (Hevea brasiliensis
Muell. Arg.) adalah komoditi perkebunan penghasil lateks sebagai sumber bahan baku industri otomotif. Lateks atau karet alam memiliki kualitas yang lebih baik dari pada karet sintetik, karena daya elastisitasnya yang lebih tinggi. Karet alam tidak mudah panas, mempunyai daya aus tinggi, serta bersifat plastisitas sehingga pengolahannya menjadi lebih mudah, khususnya bagi industri ban pesawat udara dan mobil balap. Disamping itu karet alam memiliki daya tahan yang tinggi terhadap keretakan (Bueche 1957; Shan et al. 2011).
Produksi karet alam ikut berkontribusi bagi pertumbuhan ekonomi nasional sebagai sumber devisa nonmigas dan turut serta dalam pelestarian fungsi lingkungan hidup. Indonesia adalah negara penghasil karet alam kedua terbesar di dunia setelah Thailand (Rubber Board 2016). Walaupun Indonesia memiliki luas kebun karet terbesar di dunia (3,6 juta hektar) namun produktivitasnya rendah, hanya 1 ton/ha. Produktivitas ini masih berada di bawah negara-negara penghasil karet lainnya di dunia, seperti Viet Nam (1,7 ton/ha), Thailand (1,6 ton/ha), dan Malaysia (1,3 ton/ha) (Kementerian Pertanian 2015). Inti dari permasalahan ini bermula dari kurangnya perhatian pemerintah terhadap komoditi karet, walaupun sebenarnya karet termasuk dalam daftar 15 komoditi yang menjadi prioritas di Dirjenbun Kementan (Dirjenbun 2015). Selanjutnya juga dikemukakan bahwa 85% perkebunan karet di Indonesia adalah milik rakyat (Kementerian Pertanian 2015). Namun umumnya perkebunan rakyat ini kurang produktif dan hanya setengah dari mereka yang memiliki akses pada teknologi budidaya, penanggulangan penyakit maupun bibit unggul (Damanik et al. 2010). Pada tahun 2012 sampai dengan 2014, peningkatan ekspor karet cukup signifikan dari 2.445 ribu ton meningkat menjadi 2.624 ribu ton. Namun demikian nilai jualnya mengalami penurunan, senilai USD 7.861 miliar pada tahun 2012 menjadi USD 4.742 miliar pada tahun 2014 (Dirjenbun 2016). Demi menyusun program pemuliaan tanaman karet yang berbasis marka DNA untuk meningkatkan produksi lateks
nasional, diperlukan klon-klon karet unggul yang memiliki keunggulan genetik yang tinggi terutama pada karakter yang berkaitan dengan daya produktivitas hasil tanaman dan ketahanan terhadap serangan penyakit.
Keberagaman genotipe plasma nutfah tanaman karet merupakan modal dasar untuk keberhasilan program pemuliaan dalam pengembangan tanaman unggul sebagai upaya meningkatkan kapasitas produksi lateks. Teknik pemuliaan tanaman dalam perolehan bibit unggul yang lebih cepat sangat diperlukan mengingat teknik pemuliaan tanaman secara konvensional memerlukan waktu seleksi yang panjang hingga 20-25 tahun, sejak persilangan sampai menghasilkan produk klon unggul. Sebagai alternatif untuk memanfaatkan keunggulan biologi molekuler adalah pemuliaan tanaman yang dipandu dengan marka DNA yang hanya membutuhkan waktu sepertiganya, yaitu 7-8 tahun (Collard and Mackill 2008; Jiang 2013). Dengan demikian dapat melengkapi pengembangan dalam pemuliaan tanaman karet yang unggul dan potensial secara cepat. Pemilihan marka DNA yang digunakan dalam analisis genetik perlu mempertimbangkan tujuan yang diinginkan, fasilitas laboratorium yang tersedia serta kelebihan dan kekurangan dari berbagai marka DNA tersebut. Marka DNA yang ideal adalah memiliki kemudahan akses, kemudahan prosedur analisis, polimorfisme yang tinggi, ko-dominan yang dapat membedakan antara homozigot dan heterozigot, serta tingkat pengulangannya yang tinggi.
Studi keragaman genetik pada karet telah dilakukan oleh beberapa peneliti dengan menggunakan beragam marka DNA, yaitu: marka intron length polymorphism atau ILP (Li et al. 2013), Random Amplified Polymorphic DNA atau RAPD (Wibowo 2010; Novalina dan Sagala 2011), marka isozim dan mikrosatelit (Gouvea et al. 2010; Le Guen et al. 2011), marka simple sequence repeat atau SSR (Feng et al.
2012; Hamzah 2014), dan marka EST-SSR (Feng et al. 2009; Yu et al. 2010; Perseguini et al. 2012). Menurut Toruan-Mathius et al. (2002) hasil yang diperoleh dari teknik RAPD tidak konsisten. Hal ini disebabkan oleh rendahnya akurasi dalam pengulangan hasil amplifikasi, sedikitnya jumlah fragmen DNA

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
10
yang diperoleh dari primer yang digunakan dan jumlah lokus yang dihasilkan, serta rendahnya tingkat diferensiasi dalam analisis dendrogram klon karet. Dengan demikian hasil setiap kelompok klon karet kurang terpisah dengan baik. Pada penelitian ini digunakan marka AFLP (Amplified Fragment Length Polymorphism) karena memiliki
tingkat pengulangan dengan derajat kekonsistenan hasil yang tinggi, dapat membedakan homozigot dan heterozigot (Wooten et al. 2009), keragaman tanaman dengan pola sidik jari DNA spesifik (Tajuddin et al. 2012) serta biaya awal (start-up cost) yang tidak begitu mahal dan tidak membutuhkan informasi sekuens awal dibanding SSR dan SNP (Jiang 2013). Marka AFLP juga telah digunakan pada tanaman karet (Nurhaimi-Haris et al. 2003; Xiao et al. 2009; Roy et al. 2012; Tang et al. 2013). Menurut Kuck et al. (2012) kelebihan dari marka AFLP dapat mendeteksi variasi dan keragaman genetik antar populasi, individu, serta spesies berdasarkan perbedaan letak basa tertentu dari marka DNA dalam genom tanaman.
Tujuan dari penelitian ini adalah untuk
menganalisis keragaman genetik dari enam
klon tanaman karet dengan menggunakan
marka AFLP. Selain itu juga untuk masing
klon. Manfaat yang diharapkan dari studi ini
adalah untuk dapat menjadi acuan dalam
pemilihan tetua atau induk untuk persilangan
klon selanjutnya, dan primer selektif yang
diperoleh dapat digunakan di lapang untuk
memandu pada tahap seleksi pada program
pemuliaan berikutnya.
BAHAN DAN METODE
Bahan tanaman
Bahan tanaman karet yang digunakan adalah klon GT 1, IRR 104, PB 260, PR 300, RRIC 100 dan RRIM 600, dengan produktivitas yang bervariasi yang berumur lebih dari 15 tahun dan ditanam di lahan kebun koleksi Balai Bioteknologi BPPT.
Analisis kuantitatif dan kualitatif
Isolasi DNA genom karet dilakukan
dengan metode CTAB (Doyle 1990) dengan
beberapa modifikasi menggunakan sampel
daun (Gambar 1). Modifikasi dilakukan pada
penambahan jumlah merkaptoetanol
menjadi 1% dan aplikasinya dipisah dari
buffer ekstraksi. Rasio kemurnian DNA
terhadap kontaminasi protein dan
konsentrasi DNA hasil isolasi diuji
menggunakan spektrofotometer Nanodrop
yang diukur pada perbandingan panjang
gelombang 260 dan 280 nm (Å260/Å280).
Gambar 1. (A). Sampel daun karet dan (B). pemisahan DNA karet dalam tube
A B

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
11
Setiap sampel DNA hasil isolasi diambil
sebanyak 4 µL untuk dilakukan pemisahan
pada elektroforesis gel agarosa konsentrasi
1,5% dalam bufer TE 1X dengan daya 50 V
selama 50 menit dan menggunakan
pewarna SYBR safe (Bakhtiar et al. 2010).
Analisis polimorfisme DNA Metode AFLP yang digunakan
berdasarkan pada metode Vuylsteke et al. (2007). Setelah proses amplifikasi, produk pre-amplifikasi primer tunggal diamplifikasi selektif menggunakan kombinasi pasangan primer (Tabel 1).
Sebanyak 1 µL produk pre-amplifikasi ditambah dengan 2,5 µL buffer PCR 10x, 2 µL dNTP campuran, 2,5 µL 25 mM MgCl2, 0,2 µL Taq polymerase, primer terpilih 10 pmol TruI dan 10 pmol EcoRI masing-
masing 1 µL, dan 14,8 µL ddH2O sampai volume mencapai 25 µL. Larutan kemudian diaduk secara perlahan dan disentrifugasi singkat untuk menurunkan seluruh cairan
yang ada di dinding tabung. Amplifikasi dilakukan dengan menggunakan PCR. Program yang digunakan adalah sebagai berikut: satu siklus 94ºC selama 2 menit, 94ºC selama 30 detik, 65ºC selama 30 detik dan 72ºC selama 2 menit. Tahap berikutnya adalah suhu penempelan (annealing) yang diturunkan 0,7ºC selama 60 detik dan diakhiri dengan suhu 4ºC.
Elektroforesis gel poliakrilamid
Produk amplifikasi PCR dielektroforesis pada gel poliakrilamid 6% (0,1% ammonium persulfat (APS) dan TEMED) dengan ukuran 13 x 11 Cm. Pre-elektroforesis (pemanasan) dilakukan selama 20 menit (110 W) supaya suhu permukaan gel kira-kira 50°C. Hasil amplifikasi PCR dicampur dengan 20 µL loading buffer dan 19 µL H2O, kemudian didenaturasi pada suhu 95ºC selama tiga menit dan segera didinginkan di dalam lemari es. Larutan sampel sebanyak 10 µL, dimasukkan pada setiap sumur gel dan dielektroforesis selama ±40 menit (80 W). Gel poliakrilamid kemudian diwarnai dengan larutan perak nitrat di atas shaker selama 30 menit, dicuci dengan dH2O bebas ion selama 10 detik, kemudian ditambah larutan pengembang (45 g sodium karbonat; 2,25 mL formaldehid 37%; 300 µL sodium tiosulfat dalam 1500 mL ddH2O pada suhu 4–10ºC) dan dibiarkan hingga fragmen-fragmen dapat dilihat sesuai yang diinginkan. Selanjutnya larutan fiksasi dituangkan dan dibiarkan selama dua menit, dan dilapis dengan cellophane membrane backing (BIORAD), serta dibiarkan selama
lebih kurang dua hari sampai kering.
Analisis data Pita-pita DNA diubah ke dalam bentuk
data biner dengan memberi nilai 1 jika munculnya pita DNA dan 0 jika tidak ada pita. Data biner yang diperoleh dari AFLP tersebut selanjutnya digunakan dalam analisis keragaman genetik, kemiripan genetik dibuat dalam bentuk matriks dengan SIMQUAL (similarity for qualitative data).
Berdasarkan nilai kemiripan genetik tersebut dilakukan analisis pengelompokan menggunakan sub program SAHN dengan metode UPGMA (Unweighted Pair Group Method Average) program NTSYS-pc 2.11.
Tabel 1. Pasangan primer amplifikasi selektif
No Primer No Primer
1. 2. 3. 4. 5. 6. 7. 8.
M-CAA E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
33. 34. 35. 36. 37. 38. 39. 40.
M-CTA E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
9. 10. 11. 12. 13. 14. 15. 16.
M-CAC E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
41. 42. 43. 44. 45. 46. 47. 48.
M-CTC E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
17. 18. 19. 20. 21. 22. 23. 24.
M-CAG E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
49. 50. 51. 52. 53. 54. 55. 56.
M-CTG E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
25. 26. 27. 28. 29. 30. 31. 32.
M-CAT E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG
57. 58. 59. 60. 61. 62. 63. 64.
M-CTT E-AAC E-AAG E-ACA E-ACC E-ACG E-ACT E-AGC E-AGG

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
12
HASIL DAN PEMBAHASAN
Isolasi DNA tanaman karet
Konsentrasi dan kemurnian DNA hasil isolasi dari klon-klon karet diperoleh nilai yang sangat baik. Nilai tertinggi diperoleh dari klon PB 260 dengan konsentrasi 4310,4
ng/L, dan yang terendah adalah RRIC 100
dengan konsentrasi 905,8 ng/L (Tabel 2). Rasio absorban pada 260 dan 280 nm digunakan untuk menilai kemurnian DNA. Sampel DNA akan dianggap murni apabila didapati rasio sekitar 1,8 – 2,0. Apabila rasio tersebut lebih tinggi, maka dapat diindikasikan adanya zat pengotor berupa protein, fenol atau kontaminan lain yang kuat serapannya pada 280 nm (Nanodrop Technology Inc 2007). Hal ini terjadi pada klon RRIC 100 dengan rasio kemurnian
2,17 pada gelombang serapan A260/A280, yang muncul smear pada gel hasil
elektroforesis yang diduga merupakan zat pengotor atau kontaminan (gambar tidak ditampilkan). Sedangkan apabila rasio lebih rendah dari kisaran 1,8 – 2,0 maka dapat diindikasikan adanya kontaminan berupa RNA pada sampel yang diuji (Nanodrop Technology Inc 2007). Selanjutnya hasil analisis kualitatif berupa elektroferogram menunjukkan bahwa DNA hasil isolasi memiliki ukuran DNA genom sekitar 10 kb (Gambar 2).
Produk PCR AFLP tanaman karet
Hasil elektroforesis produk ligase pada tahap pre-amplifikasi dilakukan pengenceran 10x dan 1x (tanpa pengenceran). Hasilnya menunjukkan bahwa pita fragmen hasil pre-amplifikasi dengan tanpa pengenceran (1x) lebih jelas dibanding dengan pita fragmen pre-amplifikasi pengenceran 10x yang terlihat samar. Selanjutnya produk pre-amplifikasi yang digunakan pada tahap
Tabel 2. Konsentrasi dan kemurnian DNA tanaman
karet hasil isolasi
Jenis Klon Konsentrasi DNA
maksimum (ng/µL)
Kemurnian
(Å260/Å280)
GT 1 1374,3 1,61 - 2,02
IRR 104 1024,2 1,80 - 1,97
PB 260 4310,4 1,55 - 1,81
PR 300 4127,3 1,60 - 1,78
RRIC 100 905,8 2,05 - 2,17
RRIM 600 1259,5 1,70 - 2,04
Gambar 2. Hasil elektroforesis DNA isolasi (Marka 1
kb, 1-3: DNA klon IRR 104, klon RRIM 600, dan klon PR 300)
Gambar 3. Hasil elektroforesis gel poliakrilamid

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
13
amplifikasi selektif ialah hasil pre-amplifikasi dengan tanpa pengenceran, untuk memperoleh pita-pita AFLP yang optimal.
Dari tahap pre-amplifikasi kemudian dilanjutkan ke tahap amplifikasi menggunakan 64 kombinasi pasangan primer terhadap semua klon tanaman karet. Hasil amplifikasi divisualisasikan pada gel poliakrilamid 6%. Gel poliakrilamid dengan konsentrasi 6% direkomendasikan untuk memisahkan fragmen DNA sebesar 80 – 800 bp (Barril dan Nates 2012). Menurut Nurhaimi-Haris et al. (2003), proses silver staining telah menunjukkan hasil yang lebih baik ketika memvisualisasi fragmen-fragmen AFLP yang berasal dari daun karet melalui gel poliakrilamid. Gambar 3 memperlihatkan pita-pita DNA yang terlihat pada GT 1, PB 260, dan RRIC 100 hasil beberapa pasang primer selektif bersama dengan marka Thermo Scientific Fermentas Gene Ruller
100 bp serta replika yang dibuat secara manual.
Berdasarkan dari pengukuran pita menggunakan marka Thermo Scientific Fermentas Gene Ruller 100 bp dan dengan
aplikasi program komputer PhotocaptMw versi 99.0.2.0, hasil pita yang didapatkan dari 64 pasang primer adalah 2806 pita. Ukuran pita berkisar antara 65 – 3833 bp.
Dari elektroferogram AFLP dihasilkan jumlah pita AFLP terbanyak adalah 24 pita yang berasal dari amplifikasi M-CAT/E-AAG pada klon PR 300 (tidak ditampilkan). Klon
IRR 104 menghasilakn 10 pita oleh primer M-CAC/E-AGG dan M-CAT/E-ACA. Selanjutnya, pita klon RRIM 600 diperoleh sejumlah 23 pita oleh primer M-CAC/E-AAG.
Keragaman genetik klon Karet
Elektroferogram yang dihasilkan dari amplifikasi AFLP diperoleh pita-pita AFLP dengan berbagai variasi ukuran. Untuk pembentukan pohon filogenetik berupa dendrogram, dilakukan pengolahan data biner berdasarkan variasi tersebut. Dendrogram dibuat menggunakan semua pita AFLP.
Dari dendrogram diperoleh koefisien kemiripan genetik antar keenam klon adalah rendah yaitu sebesar 0,60 yang berarti menunjukkan kekerabatan genetik keenam klon sebesar 60% (Gambar 4). Pada koefisien kemiripan 64% tampak dua kelompok kekerabatan. Kelompok 1 hanya terdiri dari klon GT 1. Sedangkan kelompok 2 terdiri dari klon PB 260, IRR 104, RRIM 600, RRIC 100 dan PR 300.
Marka spesifik klon
Dari elektroferogram diketahui ada primer yang menghasilkan pita-pita unik. Pita unik adalah pita yang mempunyai satu ukuran tertentu yang hanya muncul pada satu klon saja, oleh satu pasang primer spesifik. Pasangan primer M-CAA/E-ACA menghasilkan satu pita unik untuk klon GT 1 pada ukuran 168 bp, sedangkan pasangan
Gambar 4. Pohon filogenetik enam klon karet berdasarkan pola pita marka AFLP

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
14
primer M-GAC/E-ACG menghasilkan 2 pita unik pada ukuran 383 dan 138 bp. Untuk klon IRR 104, pita unik ukuran 20 bp diperoleh dari pasangan primer M-CAA/E-AGC. Pita-pita unik dari berbagai pasangan primer untuk klon-klon tertentu beserta ukurannya dapat dilihat pada Tabel 3.
Pembahasan
Keragaman genetik merupakan variasi yang tejadi di dalam genom suatu spesies,
yang disebabkan oleh berbagai macam faktor. Variasi genetik pada tanaman banyak dimanfaatkan pada persilangan guna menghasilkan varietas-varietas unggul. Salah satunya di Indonesia adalah tanaman karet dengan berbagai macam klon telah dihasilkan sejak tahun 1864 (Heru dan Andoko 2008). Enam klon yang digunakan dalam penelitian ini yaitu GT 1, IRR 104, PB 260, PR 300, RRIC 100 dan RRIM 600 adalah klon-klon yang dikembangkan di beberapa negara, termasuk Indonesia.
GT 1 (Gondang Tapen 1) merupakan klon primer yang berasal dari Indonesia yang termasuk sebagai klon tertua. Sedangkan IRR 104 (Indonesian Rubber Research)
adalah klon karet ungul anjuran asal Indonesia berdasarkan Surat Keputusan Menteri Pertanian Tahun 2005. Klon ini berasal dari tetua BPM 101 dengan RRIC 110 (Kementerian Pertanian 2007; Lasminingsih dan Sipayung 2012). Klon PB 260 berasal dari persilangan PB 5/51 dengan PB 49 dan dikembangkan oleh Prang Besar, Malaysia. Berikutnya klon PR 300 (Proefstation Rubber) asal Indonesia
yang berasal dari persilangan PR 226 dan PR 228 yang keduanya juga berasal dari Indonesia. Ketiga klon tersebut merupakan klon penghasil lateks (Daslin 2014). Klon RRIC (Rubber Research Institute of Ceylon) merupakan klon asal Sri Lanka, dan BPM (Balai Penelitian Medan) yang merupakan klon asal Indonesia. Klon RRIM 600 berasal dari Malaysia yang merupakan hasil persilangan TJIR 1 (Tjirandji 1) asal Indonesia dengan PB 86 asal Malaysia (Situmorang et al. 2007; Priyadarshan 2011). Terakhir klon RRIC 100 adalah klon yang berasal dari persilangan RRIC 52 dengan PB 86 yang dikembangkan di Sri Lanka.
Data biner pita DNA AFLP digunakan untuk mengkonstruksi pohon filogenetik atau dendrogram sehingga diketahui tingkat kemiripan genetik keenam klon karet, GT 1, IRR 104, PB 260, PR 300, RRIC 100 dan RRIM 600. Diperoleh hasil bahwa keenam klon membentuk dua kelompok yang menunjukkan hubungan kekerabatan secara fenotipe produktivitas menurut Priyadarshan (2011) sebagai klon unggul penghasil lateks. Daslin (2005) membagi sejarah pemuliaan tanaman karet ke dalam 4 generasi. Klon GT 1 termasuk dalam generasi II dengan produktivitas 704 kg/ha/tahun, sedangkan
Tabel 3. Pasangan primer spesifik dan ukuran unik yang dihasilkan pada masing- masing klon
Klon Pasangan
primer spesifik Ukuran pita unik
GT 1 M-CAA/E-ACA 168 bp
M-CAA/E-ACT 2357 bp
M-CAG/E-ACT 156 bp
M-CAT/E-AAG 437 bp
M-CTG/E-AAG 2416 bp
M-CTC/E-ACA 1445 bp
M-CTG/E-ACG 376 bp
M-GAC/E-ACG 383 bp, 138 bp
M-GAC/E-AGG 138 bp, 86 bp, 70 bp
IRR 104 M-CAA/E-AGC 20 bp
PB 260 M-CAG/E-AGC 2250 bp
M-CTA/E-ACA 187 bp
M-CTC/E-AAC 226 bp
M-CTT/E-ACT 109 bp
M-GAC/E-ACG 1907 bp
M-GAC/E-AGC 1906 bp
RRIC 100 M-CAG/E-AGC 179 bp
M-CAT/E-ACC 1701 bp
M-CTA/E-ACA 2916 bp
M-CTC/E-ACA 3500 bp
M-CTC/E-ACC 418 bp
M-CTG/E-ACC 160 bp
M-GAC/E-AGC 3428 bp
RRIM 600 M-CTC/E-ACG 326 bp
M-CTC/E-ACA 31 bp
M-CAC/E-ACG 333 bp
M-CTA/E-AGG 261 bp
M-CTT/E-ACC 185 bp
PR 300 M-CTC/E-AAC 30 bp, 23 bp
M-CTA/E-AGG 64 bp
M-CAT/E-ACC 72 bp, 61 bp
M-CAT/E-AAG 91 bp, 71 bp, 54 bp
M-CAC/E-ACC 58 bp
M-CTG/E-AGG 51 bp, 43 bp

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
15
PR 300 adalah generasi III dengan produktivitas 1090 kg/ha/tahun. Klon-klon generasi IV termasuk IRR 104, PB 260, RRIC 100 dan RRIM 600 dengan produktivitas 1630 kg/ha/tahun (Daslin 2013; Daslin dan Pasaribu 2015).
Klon RRIM 600 dan RRIC 100 dikembangkan dan berasal dari dua negara yang berbeda, yaitu dari Malaysia dan Sri Lanka. Kedua klon ini memiliki koefisien kemiripan genetik sebesar 66%. Jarak kekerabatan yang dekat ini bisa terjadi karena persilangan keduanya dihasilkan dari satu tetua donor yang sama yang berasal dari Malaysia, yaitu PB 86. Klon IRR 104 dan RRIM 600 membentuk satu sub-kelompok pada puncak pohon filogenetik dengan koefisien kemiripan genetik sebesar 74%. Klon yang berada pada kelompok kekerabatan yang sama berarti bahwa klon tersebut memiliki jarak kekerabatan yang dekat (Gregory 2008; Townsend et al. 2012).
Nurhaimi-Haris et al. (2003) melakukan pengujian marka AFLP pada 10 klon karet berdasarkan tingkat resistensi terhadap cendawan patogen daun Corynespora cassiicola, dengan menggunakan 10 pasang primer selektif. Dari hasil penelitiannya, diperoleh kemiripan genetik pada 10 klon karet sebesar 85,0%. Keragaman genetik dalam spesies merupakan modal dasar bagi suatu tanaman untuk tumbuh, berkembang dan beradaptasi dengan perubahan lingkungan atau penyakit sehingga dapat bertahan hidup dari generasi ke generasi (Woelan et al. 2014). Hal ini bisa terjadi karena lingkungan tempat tumbuh terutama jika lingkungan tersebut diketahui sangat berbeda secara geografisnya sehingga dapat mempengaruhi keragaman genetik yang mengekspresikan variasi secara fenotipenya. Dengan demikian fenotipe merupakan hasil interaksi genotipe dengan lingkungannya (Kubota et al. 1998; Syukur et al. 2012).
Hasil yang diperoleh dari penelitian ini sesuai dengan penelitian Toruan-Mathius et al. (2002). Mereka telah melakukan pengujian keragaman genetik klon-klon karet berdasarkan marka AFLP dan RAPD, yang membedakan dua kelompok klon karet menjadi klon yang bersifat resisten terhadap patogen Corynespora cassiicola dan klon
yang bersifat rentan terhadap jenis patogen tersebut. GT 1 termasuk ke dalam golongan
klon yang rentan terhadap jenis patogen tersebut, sedangkan RRIC 100 termasuk ke dalam golongan resisten.
Uji kuantitatif terhadap DNA hasil isolasi dari daun karet dilakukan dengan menggunakan spektrofotometer Nanodrop pada panjang gelombang 260 dan 280 nm, sedangkan uji secara kualitatif menggunakan gel elektroforesis agarosa 1% dengan pewarna SYBR safe (Gambar 1). DNA termasuk baik dan murni jika memiliki rasio Å260/Å280 antara 1,8 – 2,0 (Yuwono
2007). Dengan nilai kemurnian yang diperoleh, DNA hasil isolasi masih dapat digunakan sebagai cetakan DNA pada PCR untuk marka AFLP karena selama proses PCR tetap akan mengamplifikasi cetakan DNA secara spesifik oleh primer walaupun dengan kemurnian yang rendah (Bintang 2010). Dengan demikian kemurnian DNA hasil isolasi yang diperoleh ini tetap dapat digunakan dalam proses tahapan AFLP.
Konsentrasi DNA hasil isolasi yang berbeda-beda dapat disebabkan oleh rusaknya DNA selama proses isolasi atau adanya kontaminan (Healey et al. 2014). Pada penelitian ini tahapan AFLP digunakan konsentrasi DNA karet yang diencerkan hingga 250 ng/µL, namun dalam penelitian yang lainnya dengan konsentrasi DNA 50 ng/µL saja AFLP dapat dilakukan (Nurhaimi-Haris et al. 2003). Pengenceran ini dimaksudkan agar menghasilkan produk PCR AFLP dengan ketebalan pita yang seragam. Analisis kualitatif menggunakan pewarna SYBR safe. SYBR safe, yang
memiliki nama lain yaitu metil benzen sulfonat dan dilarutkan dalam dimetilsulfoksida (Evenson et al. 2012), membuat pita DNA berpendar setelah dielektroforesis di bawah sinar UV. Hasil kualitatif tersebut telah cukup menunjukkan keberadaan DNA tanaman karet, walaupun tidak secara utuh karena adanya smear
pada masing-masing lajur. Pita tebal yang berada di atas menunjukkan bahwa bobot molekul DNA genom yang diperoleh cukup besar, yaitu sekitar 10 kb. Smear di bawah pita DNA menunjukkan telah terjadinya degradasi atau fragmentasi DNA genom selama proses isolasi (Hamzah 2014).
Enzim restriksi yang digunakan dalam studi ini adalah EcoRI sebagai enzim pemotongan jarang (rare cutter) dan TruI sebagai enzim pemotongan sering (frequent

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
16
cutter). Enzim TruI yang digunakan adalah
enzim yang mengenali situs pemotongan basa yang mirip dengan enzim MseI, hanya
saja suhu optimal pada saat proses pemotongan berlangsung berbeda, yaitu 65oC untuk TruI dan 37oC untuk MseI.
Pemotongan enzim restriksi bertujuan untuk mendapatkan jumlah fragmen DNA yang optimum sehingga pola pita AFLP nantinya dapat diberi nilai dengan baik karena tingkat polimorfisme yang dihasilkan lebih tinggi.
Menurut Reflinur dan Lestari (2015) banyaknya DNA yang terdegradasi sebelum mengalami digesti menyebabkan banyak DNA genom tidak terpotong sempurna oleh kedua enzim restriksi dan tidak menempel dengan baik oleh kedua adapter yang menyebabkan fragmen DNA tidak teramplifikasi dengan sempurna karena tidak memiliki situs penempelan primer. Oleh karena itu, DNA yang baik merupakan faktor penting dalam metode AFLP ini untuk menghindari pemotongan DNA yang tidak sempurna dan dapat menyebabkan kesalahan dalam identifikasi pita polimorfis (Nurhaimi-Haris et al. 2010). Kekeliruan dalam mengidentifikasi pita menyebabkan tidak munculnya ukuran pita yang sesuai pada elektroferogram di tahap pemotongan dan penempelan. Hal ini menyebabkan perbedaan ukuran fragmen DNA pada elektroforegram gel poliakrilamid setelah amplifikasi. Namun, dalam studi ini semua pita yang dihasilkan tetap dianggap mewakili karakter tertentu pada masing-masing klon, maka semua pita digunakan dalam identifikasi keragaman genetik.
Berdasarkan hasil elektroferogram yang diperoleh dan pengolahan data biner, pita terbanyak dihasilkan oleh pasangan primer M-CAT/E-AAG pada klon PR 300, dengan jumlah sebanyak 24 pita. Jika dibandingkan dengan penelitian sebelumnya, jumlah pita AFLP pada studi ini sedikit lebih rendah dibandingkan dengan jumlah dari penelitian sebelumnya yang dapat mencapai 34 – 65 pita AFLP (Nurhaimi-Haris et al. 2010). Jumlah pita yang lebih sedikit kemungkinan disebabkan karena pita yang seharusnya dihasilkan belum tampak ketika pita lain sudah muncul. Hal ini dapat disebabkan karena dalam proses pewarnaan perak nitrat yang kurang optimum.
Pola pita spesifik dapat digunakan untuk mengidentifikasi klon-klon karet karena adanya pola polimorfisme yang unik pada masing-masing klon. Masing-masing primer menghasilkan polimorfisme yang berbeda yang dapat digunakan untuk identifikasi suatu tanaman (Misra et al. 2010; Ghosh et al. 2011). Menurut Chial (2008), polimorfisme yang bersifat unik dapat dijadikan sebagai identitas genotipe atau sidik jari DNA suatu sampel yang diuji. Pola polimorfisme yang dihasilkan dapat digunakan untuk identifikasi klon karet melalui deteksi pita spesifik pada primer yang digunakan. Sebelum pita spesifik ini dapat digunakan sebagai sarana seleksi di lapang, maka diperlukan tahap validasi terlebih dahulu untuk memastikan perannya sebagai pemandu dalam pemuliaan tanaman karet. KESIMPULAN
Marka AFLP berhasil digunakan untuk menganalisis keragaman genetik enam klon karet yang ada di kebun koleksi Balai Bioteknologi, BPPT berdasarkan jumlah pita polimorfis yang muncul dan tingkat kemiripan genetik yang dihasilkan. Analisis keragaman genetik dari enam klon karet yaitu GT 1, IRR 104, PB 260, PR 300, RRIC 100 dan RRIM 600 menggunakan 64 pasang primer dan menghasilkan 2806 pita AFLP dengan ukuran pita yang bervariasi antara 65 hingga 3833 bp dan bersifat polimorfis. Tingkat kemiripan genetik keenam klon karet tersebut adalah sebesar 64%, dan terbagi dalam dua kelompok dengan tingkat kemiripan genetik pada puncak pohon filogenetik yang diperoleh antara IRR 104 dan RRIM 600 adalah 74%. Beberapa pita unik diperoleh sebagai marka spesifik untuk klon-klon tertentu. DAFTAR PUSTAKA
Bakhtiar Y, Yahya S, Sumaryono W, Sinaga MS, Budi SW, Tajuddin T (2010) Isolation and identification of Mycorrhizosphere bacteria and their antagonistic effects towards Ganoderma boninense in vitro.
Microbiol Indones 4:96-102. doi: 10.5454/mi.4.2.9

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
17
Barril P, Nates S (2012) Introduction to agarose and polyacrylamide gel electrophoresis matrices with respect to their detection sensitivities, Gel Electrophoresis – Principles and Basics, Magdeldin S (Ed). ISBN: 978-953-51-0458-2. Intech. doi: 10.5772/38573
Bintang M (2010) Biokimia: Teknik Penelitian. Erlangga, Jakarta
Bueche F (1957) Mechanical properties of natural and synthetic rubbers. J Polym Sci 25: 305-324. doi: 10.1002/pol.1957.1202511005
Chial H (2008) DNA fingerprinting using amplified fragment length polymorphisms (AFLP): No genome sequence required. Nat Educ 1:176
Collard BCY, Mackill DJ (2008) Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Phil Trans R Soc B 363: 557–572. doi: 10.1098/rstb.2007.2170
Damanik S, Syakir M, Tasma M, Siswanto (2010) Budidaya dan pasca panen karet. Pusat Penelitian dan Pengembangan Perkebunan, Bogor
Daslin A (2005) Kemajuan pemuliaan dan seleksi dalam menghasilkan kultivar
karet unggul. Hlm: 2637. Prosiding Lokakarya Nasional Pemuliaan
Tanaman Karet, 2223 November 2005. Balai Penelitian Sungei Putih, Pusat Penelitian Karet, Medan
Daslin A (2013) Produktivitas klon karet pada berbagai kondisi lingkungan di perkebunan. Agrium 18: 1-6
Daslin A (2014) Perkembangan penelitian klon karet unggul IRR seri 100 sebagai penghasil lateks dan kayu. Warta Perkaretan 33:1-10
Daslin A, Pasaribu SA (2015) Uji adaptasi klon karet IRR seri 100 pada agroklimat kering di kebun Sungei Baleh Kabupaten Asahan Sumatera Utara. J Penelitian Karet 33: 25-34
Dirjenbun (2015) Rencana Strategis Direktorat Jenderal Perkebunan Tahun 2015-2019. Direktorat Jenderal Perke-bunan Kementerian Pertanian. Jakarta
Dirjenbun (2016) Statistik Perkebunan Indonesia Karet 2015-2017. Direktorat Jenderal Perkebunan, Kementerian Pertanian, Jakarta
Doyle JJ (1990) Isolation of plant DNA from fresh tissue. Focus 12:13-15
Evenson WE, Boden LM, Muzikar KA, O’Leary DJ (2012) 1H and 13C NMR assignments for the cyanine dyes SYBR safe and thiazole orange. J Org Chem 77:10967-10971. doi: 10.1021/jo3021659
Feng S, Wu Y, Li W, Yu F, Wang J (2012) Analysis of genetic diversity and SSR allelic variation in rubber tree (Hevea brasilensis). The Molecular Basic of
Plant Genetic Diversity. Caliskan M (Ed). 7:136-150. ISBN: 978-953-51-0157-4
Feng SP, Li WG, Huang HS, Wang JY, Wu YT (2009) Development, characterization and cross-species/ genera transferability of EST-SSR markers for rubber tree (Hevea brasiliensis) Mol Breeding 23:85–97.
doi: 10.1007/s11032-008-9216-0 Ghosh S, Majumder PB, Sen Mandi S (2011)
Species-specific AFLP markers for identification of Zingiber officinale, Z. montanum and Z. zerumbet (Zingiberaceae). Genet Mol Res 10:218-29. doi: 10.4238/vol10-1gmr1154
Gouvea LRL, Rubiano LB, Chioratto AF, Zucchi MI, Goncalves PS (2010) Genetic divergence of rubber tree estimated by multivariate techniques and microsatellite markers. Genet Mol Biol 33:308-318. doi: 10.1590/S1415-47572010005000039
Gregory TR (2008) Understanding evolutionary trees. Evo Edu Outreach 1: 121-137. doi: 10.1007/s12052-008-0035-x
Hamzah P (2014) Evaluasi marka Simple Sequence Repeat (SSR) untuk identifikasi genotip klon karet (Hevea brasiliensis). Skripsi, Institut Pertanian Bogor
Healey A, Furtado A, Cooper T, Henry RJ (2014) Protocol: a simple method for extracting next-generation sequencing quality genomic DNA from recalcitrant plant species. Plant Methods 10:21. doi: 10.1186/1746-4811-10-21
Heru DS, Andoko A (2008) Petunjuk Lengkap Budi Daya Karet. Agromedia Pustaka, Jakarta

Analisis Filogenetik Beberapa Klon Karet... Suparningtyas et al.
18
Jiang GL (2013) Molecular markers and marker-assisted breeding in plants Pp 45-83. Andersen SB (Ed.) Plant breeding from Laboratories to Fields. ISBN 978-953-51-1090-3. InTech. doi: 10.5772/52583
Kementerian Pertanian (2007) Klon karet unggul untuk 2006-2010. Tabloid Sinar Tani Edisi 28 Maret-3 April 2007
Kementerian Pertanian (2015) Outlook Karet – Komoditas Pertanian Subsektor Perkebunan. Pusat Data dan Sistem Informasi Pertanian, Sekretariat Jenderal
Kubota C, Tajuddin T, Kozai T (1998) Photoautotrophic micropropagation of mangosteen plantlets. Jap J Trop Agr 42:31-32
Kück P, Greve C, Misof B, Gimnich F (2012) Automated masking of AFLP markers improves reliability of phylogenetic analyses. PLoS One 7:e49119. doi: 10.1371/journal.pone.0049119
Lasminingsih M, Sipayung HH (2012) Petunjuk Praktis Pembibitan Karet. ISBN: 979-006-402-0. AgroMedia Pustaka, Jakarta
Le Guen V, Gay C, Xiong TC, Souza LM, Rodier-Goud M, Seguin M (2011) Development and characterization of 296 new polymorphic microsatellite markers for rubber tree (Hevea brasiliensis). Plant Breeding 130: 294–
296. doi: 10.1111/j.1439-0523.2010.01774.x
Li D, Xia Z, Deng Z, Liu X, Feng F (2013) Development, characterization, genetic diversity and cross-species/genera transferability of ILP markers in rubber tree (Hevea brasiliensis). Genes
Genom 35:719-731. doi: 10.1007/s13258-013-0122-4
Misra A, Shasany AK, Shukla AK, Darokar M, Singh S, Velusamy S, Singh J, Bagchi GD, Jain SP, Saikia D, Khanuja SPS (2010) AFLP markers for identification of Swertia species (Gentianaceae). Genet Mol Res 9:1535-1544. doi: 10.4238/vol9-3gmr785
NanoDrop Technologies Inc (2007) ND-1000 Spechtrophotometer V3.5 user’s manual
Novalina, Sagala AD (2011) Studi segregasi dan pewarisan marka-marka RAPD
pada tanaman karet hasil persilangan PB 260 dengan PN. Biospecies 4:18-26
Nurhaimi-Haris, Aswidinnoor H, Toruan-Mathius N, Purwantara A (2003) Kemiripan genetik klon karet (Hevea brasiliensis Muell. Arg.) berdasarkan metode Amplified Fragment Length Polymorphisms (AFLP). Menara
Perkebunan 71:1-15 Nurhaimi-Haris, Suwanto A, Suhartono MT,
Aswidinnoor H (2010) Isolation and cloning of cDNA fragments from rubber plant with resistant and susceptible characters to Corynespora cassiicola
using cDNA-AFLP method. Menara Perkebunan 78:33-42
Perseguini JMKC, Romao LRC, Brinez B, Junior EJS, Goncalves PS, Benchimol LL (2012) Genetic diversity of cultivated accesions and wild species of rubber tree using EST-SSR markers. Pesq agropec bras 47:1087-1094
Priyadarshan PM (2011) Biology of Hevea
Rubber. CABI, London Reflinur, Lestari P (2015) Penentuan lokus
gen dalam kromosom tanaman dengan bantuan marka DNA. J Litbang Pert 34:177-186
Roy CB, Ravindran M, Saha T (2012) Efficient screening of AFLP primer combinations for evaluating genetic diversity among cultivated rubber (Hevea brasiliensis) clones. Natural
Rubber Research 25:21-30 Rubber Board (2016) Monthly Rubber
Statistical News Vol 7 No 12 May 2016. Statistics & Planning Department, Rubber Board, Kottayam-686 002 Kerala, India
Shan C, Gu Z, Wang L, Li P, Song G, Gao Z, Yang X (2011) Preparation, characterization, and application of NR/SBR/Organoclay nanocomposites in the tire industry. J Appl Polym Sci 119: 1185-1194. doi: 10.1002/app.32773
Situmorang A, Sinaga MS, Suseno R, Hidayat SH, Siswanto, Darussamin A (2007) Virulensi isolat Corynespora cassiicola asal sentra perkebunan
karet Indonesia terhadap beberapa klon karet anjuran. J Penelitian Karet 25:37-58

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
19
Syukur M, Sujiprihati S, Yunianti R (2012) Teknik Pemuliaan Tanaman. Penebar Swadaya, Jakarta
Tajuddin T, Cartealy IC, Safarrida A, Purwoko D, Zulaeha S, Ardiyani M (2012) Identification on Indonesian accessions of Curcuma xanthorrhiza Roxb. using AFLP markers. Biology, Medicine & Natural Product Chemistry 1:33-37
Tang C, Xiao X, Li H, Fan Y, Yang J, Qi J, Li H (2013) Comparative analysis of latex transcriptome reveals putative molecular mechanisms underlying super productivity of Hevea brasiliensis. PLoS One 8(9):e75307.
doi: 10.1371/journal.pone.0075307 Toruan-Mathius N, Lalu Z, Soedarsono,
Aswidinnoor H (2002) Keragaman genetik klon-klon karet (Hevea brasiliensis Muell. Arg.) yang resisten dan rentan terhadap Corynespora casiicola
berdasarkan penanda RAPD dan AFLP. Menara Perkebunan 70:35-49
Townsend JP, Su Z, Tekle YI (2012) Phylogenetic signal and noise: Predicting the power of a data set to resolve phylogeny. Syst Biol 61:835-849. doi: 10.1093/sysbio/sys036
Vuylsteke M, Peleman JD, van Eijk MJ (2007) AFLP-based transcript profiling (cDNA-AFLP) for genome-wide expression analysis. Nat Protoc
2:1399-1413. doi: 10.1038/nprot.2007.174
Wibowo IY (2010) Analisis keragaman genetik tanaman karet hasil persilangan antara RRIM 600 dan PN 1546 dengan menggunakan teknik RAPD. Skripsi, Institut Pertanian Bogor
Woelan S, Sayurandi, Irwansyah E (2014) Keragaman genetik tanaman karet (Hevea brasiliensis Muell. Arg.) dari hasil persilangan interspesifik. J Penelitian Karet 32:109-121
Wooten JA, Tolley-Jordan LR (2009) Validation of phylogenetic signals in amplified fragment length data: testing the utility and reliability in closely related taxa. BMC Res Notes 2:26. doi: 10.1186/1756-0500-2-26
Xiao X, Li H, Tang C (2009) A silver-staining cDNA-AFLP protocol suitable for transcript profiling in the latex of Hevea brasiliensis (para rubber tree). Mol
Biotechnol 42:91-99. doi: 10.1007/s12033-008-9139-3
Yu F, Wang BH, Feng SP, Wang JY, Li WG, Wu YT (2010) Development, characterization, and cross-species/ genera transferability of SSR markers for rubber tree (Hevea brasilien-sis) Plant Cell Rep. 30:335–344
Yuwono T (2007) Teori dan Aplikasi Polymerase Chain Reaction. Andi
Publisher, Yogyakarta

Dekolorisasi Pewarna Tekstil Sumifix Blue... Permatasari et al.
20
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
DEKOLORISASI PEWARNA TEKSTIL SUMIFIX BLUE DAN
REACTIVE RED 2 OLEH MIKROBA YANG DIISOLASI DARI
LIMBAH INDUSTRI TEKSTIL
Decolorization of Sumifix Blue and Reactive Red 2 Textile Dyes by Microbes Isolated from Textile Waste Water
Intan Permatasari, Rully Adi Nugroho, Vincentia Irene Meitiniarti
*
Fakultas Biologi, Univeristas Kristen Satya Wacana, Jln. Diponegoro 52-60 Salatiga 50711 Indonesia.
*Email: [email protected]
ABSTRACT Azo dyes represent the most commonly used group of dyes in textile industry and discharged into industrial effluents worldwide. Aims of this study are to isolate microbe from textile waste water and to determine their ability to decolorize Sumifix Blue and Reactive Red 2 textile dyes. Microbe was isolated from textile effluent of PT Timatex, Salatiga. The activity for decolorization was assayed by inoculating microbial isolates into dye containing medium. Living and nonliving cell were incubated in dye containing medium in order to determine if microbial cells involved in decolorizing dye. Five different microbial isolates have been isolated from textile waste water. Isolates IBLTT_1 and IBLTT_5 showed the highest activity to decolorize Sumifix Blue, and only isolate IBLTT_1 showed the highest capability in decolorizing Reactive Red 2. Both isolates indicated positive potential towards biotreatment of textile waste water. Further results confirmed that decolorization was due to biodegradation, rather than physical adsorption by inactive cells. Keywords: decolorization, microbial isolation, Reactive Red 2, Sumifix Blue, textile effluent
ABSTRAK Pewarna azo mewakili kelompok pewarna yang umum digunakan pada industri tekstil dan banyak dijumpai di buangan limbah industri tekstil. Tujuan dari penelitian ini adalah untuk mendapatkan isolat dari limbah tekstil dan untuk mengetahui kemampuannya dalam mendekolorisasi pewarna tekstil Sumifix Blue dan Reactive Red 2. Sampel diperoleh dari limbah industri tekstil PT Timatex, Salatiga. Uji kemampuan dekolorisasi dilakukan dengan menginokulasikan isolat mikroba ke dalam medium Nutrient Broth yang mengandung pewarna. Untuk mengetahui apakah sel mikroba terlibat dalam dekolorisasi pewarna, maka sel hidup dan mati diinokulasi pada medium tersebut. Lima isolat yang berbeda diperoleh dalam penelitian ini. Isolat IBLTT_1 dan IBLTT_5 merupakan isolat dengan kemampuan dekolorisasi Sumifix Blue tertinggi. Isolat IBLTT_1 juga merupakan isolat dengan kemampuan dekolorisasi Reactive Red 2 tertinggi. Kedua isolat tersebut menunjukkan potensi positif terhadap pengolahan limbah tekstil. Hasil lebih lanjut menegaskan bahwa dekolorisasi Sumifix Blue dan Reactive Red 2 disebabkan oleh proses biodegradasi, bukan diadsorpsi oleh sel yang mati. Kata kunci: dekolorisasi, isolat mikroba, limbah tekstil, Reactive Red 2, Sumifix Blue
Received: 19 February 2018 Accepted: 12 April 2018 Published: 07 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
21
PENDAHULUAN
Industri tekstil merupakan salah satu industri penyumbang pencemaran air. Pencemaran air oleh industri tekstil berasal dari air limbah yang masih mengandung pewarna yang berasal dari aktivitas pewarnaan dan pembilasan. Hal ini disebabkan karena sekitar 10-15% zat warna yang digunakan dalam proses pencelupan tidak mengikat serat sehingga akan tersisa pada air limbah (Naresh et al. 2013).
Pewarna azo merupakan kelompok pewarna yang paling banyak digunakan pada industri tekstil (Brüschweiler et al. 2014; Lalnunhlimi dan Krishnaswamy 2016). Pewarna ini menyusun 60-70% dari seluruh pewarna dalam produksi tekstil, karena mudah disintesis dan murah, serta stabilitas dan ketersediaan berbagai warna dibandingkan dengan pewarna alami. Setidaknya 10-15% dari pewarna yang digunakan dibuang ke badan air sehingga buangan dapat membahayakan lingkungan (Rawat et al. 2016; Abo-State et al. 2017).
Menurut Velmurugan dan Revikumar (2014), senyawa azo merupakan senyawa aromatik dengan satu atau lebih ikatan (-N=N-) yang terikat pada cincin aromatik. Berdasarkan jumlah ikatan azo-nya, pewarna azo digolongkan dalam 4 jenis yaitu monoazo dengan 1 ikatan azo, diazo dengan 2 ikatan azo, triazo dengan 3 ikatan azo dan poliazo dengan lebih dari 3 ikatan azo (Kiernan 2001). Reactive Red 2 merupakan
contoh dari pewarna monoazo (Emilia et al. 2016), sedangkan Sumifix Blue merupakan contoh dari pewarna triazo (Yan 2003).
Menghilangkan atau mengurangi zat warna dan senyawa organik di dalam limbah cair salah satunya dapat dilakukan dengan proses degradasi yang melibatkan dekolorisasi (Abubacker dan Mehala 2014). Menurut Chacko dan Subramaniam (2011), dalam proses dekolorisasi biasanya dimulai dengan dekolorisasi pewarna dengan pemutusan ikatan azo yang dilakukan oleh enzim azoreduktase. Azoreduktase memutuskan ikatan azo (-N=N-) dengan NADH sebagai kofaktor (Leelakriangsak dan Borisut 2012). Perubahan struktur kimia akibat adanya pemutusan ikatan azo secara visual dapat dilihat dari pemudaran atau penurunan kepekatan warna.
Bakteri merupakan mikroorganisme
yang paling sering digunakan sebagai agen dekolorisasi karena pertumbuhannya yang cepat, mudah beradaptasi pada suhu dan salinitas yang ekstrim, dan mudah dikultivasi (Khalid et al. 2008; Shah et al. 2013; Mumtaz et al. 2015). Selain itu, keberadaannya dalam limbah juga banyak (Sriram dan Reetha 2015).
Enterococcus faecalis merupakan
salah satu bakteri yang terbukti dapat digunakan sebagai agen pendekolorisasi zat warna reaktif. Penelitian sebelumnya menyebutkan bahwa E. faecalis dapat mendekolorisasi pewarna azo Acid Red 27 dan Reactive Red 2 dengan efisiensi
dekolorisasi mencapai 95-100% selama 15 jam (Handayani et al. 2007).
Selain E. faecalis, Bacillus subtilis
juga dilaporkan dapat mendekolorisasi pewarna tekstil (Hassan et al. 2013; Kathjkeyan et al. 2016). B. subtilis dapat mendekolorisasi pewarna tekstil Remazol Black B (Shah et al. 2013), Red RR (Shah 2014) dan Reactive Red ME4BL (Velmurugan dan Ravikumar 2014). Jaiswal dan Gomashe (2017) menyatakan bahwa B. subtilis dapat mendekolorisasi pewarna Acid Red 2 hingga 90% selama 72 jam.
Penelitian ini dilakukan untuk mendapatkan dan memanfaatkan isolat dari limbah tekstil dan untuk mengetahui kemampuannya dalam mendekolorisasi pewarna tekstil Sumifix Blue dan Reactive Red 2. BAHAN DAN METODE
Sampel dan pewarna azo
Sampel limbah diperoleh dari unit pengolahan limbah aerob PT Timatex, Salatiga, sedangkan isolat pembanding E. faecalis ID6017 dan B. subtilis diperoleh
dari Laboratorium Mikrobiologi, Fakultas Biologi Universitas Kristen Satya Wacana, Salatiga. Pewarna azo Sumifix Blue dan Reactive Red 2 didapat dari Laboratorium
Ekologi dan Lingkungan, Fakultas Biologi, Universitas Kristen Satya Wacana, Salatiga.
Isolasi dan pemeliharaan
Sampel diencerkan berseri hingga 10-8, kemudian sebanyak 0,2 mL sampel yang sudah diencerkan diinokulasikan ke dalam medium nutrient agar yang mengandung pewarna Sumifix Blue atau Reactive Red 2

Dekolorisasi Pewarna Tekstil Sumifix Blue... Permatasari et al.
22
masing-masing dengan konsentrasi 50 mg L-1 dan diinkubasi pada suhu 37ºC selama 48 jam. Masing-masing koloni tunggal yang tumbuh dimurnikan dan dipelihara dalam medium nutrient agar yang mengandung
pewarna 50 mg L-1.
Uji kemampuan dekolorisasi pewarna
Isolat pendekolorisasi pewarna Sumifix Blue atau Reactive Red 2 dikulturkan terlebih dahulu di dalam 100 mL medium nutrient broth pada suhu 37ºC sela-
ma 48 jam hingga OD kultur pada 600 nm mencapai 0,5 (Spektrofotometer Shimadzu).
Sebanyak 0,2 mL kultur diinokulasikan ke dalam 100 mL medium nutrient broth yang mengandung pewarna Sumifix Blue atau Reactive Red 2 masing-
masing dengan konsentrasi 50 mg L-1 dan diinkubasi secara statis pada suhu 37ºC (Vimala et al. 2015; Maheswari et al. 2016). Pengambilan sampel dilakukan setiap 6 jam selama 48 jam dan absorbansi diukur pada
530 nm untuk medium dengan pewarna Sumifix Blue dan 520 nm untuk medium dengan pewarna Reactive Red 2.
Kemampuan dekolorisasi isolat-isolat mikroba juga dibandingkan dengan E. faecalis dan B. subtilis.
Untuk mengetahui apakah sel mikroba atau absorbsi oleh sel yang sudah mati yang berperan dalam proses dekolorisasi, maka 0,2 mL kultur segar dan kultur yang dimatikan dengan cara sterilisasi (121ºC, 15 menit) dari masing-masing isolat diinokulasikan ke dalam 4 mL
medium nutrient broth yang mengandung 50 mg L-1 pewarna. Kemampuan dekolori-
sasi pewarna oleh sel mati dibandingkan dengan sel hidup (Khehra et al. 2005).
Analisis data
Parameter persentase dekolorisasi, kecepatan dekolorisasi, dan efisiensi dekolorisasi dihitung mengikuti rumus Al-Garni et al. (2013). Oleh karena data yang diperoleh tidak memenuhi asumsi ANOVA, data dianalisis dengan menggunakan uji nonparametrik Kruskal-Wallis 1-way ANOVA diikuti dengan uji posterior Mann-Whitney U untuk mengetahui perbedaan kecepatan dekolorisasi antarisolat. HASIL DAN PEMBAHASAN
Isolat mikroba dari limbah tekstil
Dari penelitian ini diperoleh 5 isolat yang mampu mendekolorisasi pewarna Sumifix Blue dan Reactive Red 2 pada medium nutrient broth. Gambar 1 menunjukkan morfologi (bentuk, ukuran, tepi dan permukaan koloni) masing-masing isolat. Masing-masing isolat hasil isolasi dari limbah industri tekstil mempunyai morfologi yang berbeda. Kemampuan dekolorisasi pewarna
Kelima isolat mampu mendekolori-sasi Sumifix Blue, dengan dekolorisasi
tercepat dilakukan oleh IBLTT_5, diikuti IBLTT_1, IBLTT_3, IBLTT_2 dan IBLTT_4 dalam waktu paling lama 48 jam (Gambar 2a). Isolat-isolat tersebut juga mampu mendekolorisasi Reactive Red 2, dengan
dekolorisasi tercepat dilakukan oleh IBLTT_1, diikuti IBLTT_5, IBLTT_3, IBLTT_2 dan IBLTT 4 dalam waktu paling lama lebih dari 48 jam (Gambar 2.b)
Gambar 2.a dan 2.b menunjukkan
bahwa waktu dekolorisasi kelima isolat yang
diisolasi dari limbah industri tekstil
(IBLTT_1, IBLTT_2, IBLTT_3, IBLTT_4,
IBLTT_5) masih lebih rendah dibandingkan
dengan waktu dekolorisasi E. faecalis yang
hanya memerlukan waktu sekitar 18 jam.
Namun, isolat IBLTT_1 dan IBLTT_5
memerlukan waktu dekolorisasi kedua
pewarna relatif lebih cepat dibandingkan
dengan B. subtilis.
Gambar 1. Morfologi koloni isolat dari limbah industri
tekstil pada perbesaran 40x. (a). Isolat
IBLTT_1, (b). Isolat IBLTT_2, (c). Isolat
IBLTT_3, (d). Isolat IBLTT_4, (e). Isolat
IBLTT_5

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
23
a).
b).
Gambar 2. Dekolorisasi pewarna (a). Sumifix Blue dan (b). Reactive Red 2 oleh isolat yang diisolasi dari limbah industri tekstil
Dekolorisasi 50 mg L-1 pewarna
Sumifix Blue dan Reactive Red 2 tercepat dilakukan oleh E. faecalis dalam waktu 18 jam, sehingga persentase dekolorisasi, kecepatan dekolorisasi dan efisiensi dekolorisasi antarisolat mikroba dihitung dengan waktu inkubasi 18 jam (Tabel 1).
Tabel ini menunjukkan bahwa dari kelima isolat, isolat IBLTT_1 dan IBLTT_5 merupakan isolat-isolat mikroba dengan
kecepatan dan efisiensi dekolorisasi pewarna Sumifix Blue terbesar (P<0,05)
dibandingkan dengan 3 isolat mikroba lain yang juga diisolasi dari limbah tekstil. Isolat IBLTT_1 juga memiliki kecepatan dan efisiensi dekolorisasi pewarna Reactive Red 2 tertinggi (P<0,05) dibandingkan dengan 4
isolat yang lain. Meskipun demikian, kecepatan dan efisiensi dekolorisasi kedua isolat masih lebih rendah dibandingkan
0
10
20
30
40
50
60
0 6 12 18 24 30 36 42 48 54
Konsentr
asi S
um
ifix
Blu
e (
mg L
-1)
Waktu (jam)
IBLTT 1 IBLTT 2 IBLTT 3 IBLTT 4
IBLTT 5 B.subtilis E.faecalis
0
10
20
30
40
50
60
0 6 12 18 24 30 36 42 48 54
Konsentr
asi R
eactive R
ed 2
(m
g L
-1)
Waktu (jam)
IBLTT 1 IBLTT 2 IBLTT 3 IBLTT 4
IBLTT 5 B. subtilis E.faecalis
B. subtilis E. faecalis
E. faecalis

Dekolorisasi Pewarna Tekstil Sumifix Blue... Permatasari et al.
24
dengan kecepatan dan efisiensi dekolorisasi mikroba E. faecalis (P<0,05).
Tabel 1 juga menunjukkan bahwa kecepatan dan efisiensi dekolorisasi pewarna triazo Sumifix Blue oleh kelima
isolat lebih cepat dibandingkan dengan pewarna monoazo Reactive Red 2. Hasil ini berbeda dengan hasil penelitian Sharma et al. (2011) yang melaporkan bahwa pewarna dengan struktur monoazo menunujukkan kecepatan dekolorisasi lebih tinggi. Franciscon et al. (2012) juga mendapatkan hasil bahwa Brevibacterium sp. VN-15
mampu mendekolorisasi pewarna monoazo Reactive Red 198 (satu kelompok dengan Reactive Red 2) lebih cepat yaitu dalam
waktu 10 jam dibandingkan dengan pewarna diazo Reactive Blue 5 dalam waku 24 jam dan pewarna triazo Direct Blue 71 (satu kelompok dengan Sumifix Blue) dalam waktu 48 jam dengan konsentrasi awal pewarna adalah 100 mg L-1. Shah et al. (2013) juga melaporkan bahwa efisiensi dekolorisasi Reactive Red oleh Bacillus sp.ETL 1982 mencapai 95% dalam waktu
24 jam. Namun dalam penelitian Van der Zee et al. (2001) dilaporkan bahwa kecepatan dekolorisasi paling lambat dijumpai pada pewarna reaktif dengan gugus triazin, salah satunya Reactive Red.
Lebih lanjut Khehra et al. (2005) melaporkan bahwa Reactive Red 120 tidak
dapat terdekolorisasi secara signifikan oleh Stenothropomonas acidaminiphila, Pseudomonas putida, Bacillus cereus, dan Pseudomonas fluorescens dengan
kemampuan dekolorisasi maksimum hanya 61% selama 24 jam.
Hasil penelitian ini juga menunjukkan
bahwa kultur yang dimatikan tidak mampu
mendekolorisasi pewarna Sumifix Blue dan
Reactive Red 2 (Gambar 3). Hal ini dapat
dilihat dari hilangnya warna biru dari
pewarna Sumifix Blue dan warna merah dari
Reactive Red 2 hanya terjadi pada kultur
hidup. Menurut Chacko dan Subramaniam
(2011) hilangnya warna ini disebabkan oleh
putusnya ikatan azo pada struktur
pewarnanya. Enzim lacase dan
azoreduktase merupakan enzim yang
berperan dalam proses penghilangan warna
oleh baketri (Athira et al. 2016). Hasil ini
menunjukkan bahwa dekolorisasi tidak
disebabkan adsorpsi fisik oleh kultur yang
tidak aktif.
Peneliti-peneliti lain juga mendapatkan
hasil yang sama, bahwa hanya sel bakteri
yang hidup yang mampu mendekolorisasi
pewarna Remazol Black B (Asad et al. 2007)
dan Reactive Red (Shah et al. 2013). Asad
et al. (2007) juga mendapatkan hasil bahwa
aktivitas dekolorisasi pewarna Remazol
Black B tidak dijumpai di supernatan media
kultur yang telah dipisahkan dari selnya. Hal
ini mengindikasikan bahwa tidak ada enzim
ekstraseluler atau bioproduk lain yang
mungkin berperan dalam dekolorisasi. KESIMPULAN
Telah diperoleh 5 isolat yang mampu
mendekolorisasi pewarna Sumifix Blue dan
Reactive Red 2. Kecepatan dekolorisasi
pewarna Sumifix Blue tertinggi dihasilkan
oleh isolat IBLTT_5, sedangkan dekolorisasi
pewarna Reactive Red 2 tertinggi dihasilkan
oleh isolat IBLTT_1. Dekolorisasi kedua
Tabel 1. Efisiensi isolat mikroba mendekolorisasi 50 mg L-1
pewarna Sumifix Blue atau Reactive Red 2 selama 18 jam inkubasi
Isolat
Dekolorisasi
Kecepatan Dekolorisasi
Efisiensi Dekolorisasi
Sumifix Blue (%)
Reactive Red2 (%)
Sumifix Blue (%)
Reactive Red2 (%)
Sumifix Blue (%)
Reactive Red2 (%)
Isolat 1 54,64 45,73
3,04 2,54
54,70 45,81
Isolat 2 31,55 31,18
1,75 1,73
31,59 31,23
Isolat 3 41,42 30,68
2,30 1,70
41,47 30,72
Isolat 4 27,51 16,46
1,53 0,91
27,54 16,48
Isolat 5 72,41 19,58
4,02 1,09
72,49 19,61
B. subtilis 39,48 33,57
2,19 1,87
39,52 33,63
E. faecalis 99,90 99,84
5,55 5,55
100,00 100,00

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
25
pewarna tidak disebabkan adsorpsi fisik oleh
kultur yang tidak aktif, tetapi dihasilkan oleh
aktivitas sel kultur aktif.
DAFTAR PUSTAKA
Abo-State MAM, Saleh YE, Hazaa HA
(2017) Decolorization of congo red dye
by bacterial isolates. J Eco Heal Env
5:41-48. Doi: 10.12785/jehe/050201
Abubacker MN, Mehala T (2014)
Decolourization of directorange-102
and malachite green by bacterial
consortium. BiolifeJ 2:1293-1300
Al-Garni SM, Ghanem KM, Kabli SA, Biag
AK (2013) Decolorization of crystal
violet by mono and mixed bacterial
culture techniques using optimized
culture conditions. Pol J Environ Stud
22: 1297-1306
Asad S, Amoozegar MA, Pourbabaee AA,
Sarbolouki MN, Dastgheib SMM
(2007) Decolorization of textile azo
dyes by newly isolated halophilic and
halotolerant bacteria. Bioresour
Technol 98:2082-2088.
Doi:10.1016/j.biortech.2006.08.020
Athira S, Gowdhaman D, Ponnusami V
(2016) Molecular characterization and
azoreductase activity of Pseudomonas
stutzeri isolated from textile dye
effluent. Int J Chem Sci 14: 2703-2709
Brüschweiler BJ, Küng S, Bürgi D, Muralt L,
Nyfeler E (2014) Identification of non-
regulated aromatic amines of
toxicological concern which can be
cleaved from azo dyes used in clothing
textiles. Regul Toxicol Pharmacol 69:
263-272. Doi:
10.1016/j.yrtph.2014.04.011
Chacko JT, Subramaniam K (2011) Enzymatic degradation of azo dyes - Areview. Int J Environ Sci 1: 1250-
1260 Emilia TA, Wijaya YA, Mermaliandi F (2016)
Degradation of reactive red 2 by fenton and photo-fenton oxidation processes. J Eng Appl Sci 11:5227-5231.
Franciscon E, Grossman MJ, Paschoal JA, Reyes FG, Durrant LR (2012) Decolorization and biodegradation of reactive sulfonate azo dyes by a newly isolated Brevibacterium sp. strain VN-
15. Springerplus 1: 37-46. Doi: 10.1186/2193-1801-1-37
Handayani W, Meitiniarti VI, Timotius KH (2007) Decolorization of acid red 27 and reactive red 2 by Enterococcus faecalis under a batch system. World J
Microbiol Biotechnol 23: 1239-1244. Doi: 10.1007/s11274-007-9355-1
Hassan MM, Alam MZ, Anwar MN (2013) Biodegradation of textile azo dyes by bacteria isolated from dyeing industry effluent. Int Res J Biological Sci 2: 27-
31 Jaiswal SS, Gomashe AV (2017)
Bioremediation of textile azo dyes by newly isolated Bacillus sp. from dye
contaminated soil. Int J Biotechnol Biochem 13:147-153
Karthikeyan G, Radha T, Ragunathan R,
Johnney J (2016) Degradation and
decolourisation of dye effluents by
using Bacillus subtilis CBNR isolates.
Int Res J Multidisciplinary Sci Technol
1: 142-147
Khehra MS, Saini HS, Sharma DK, Chadha
BS, Chimni SS (2005) Comparative
studies on potential of consortium and
constituent pure bacterial isolates to
decolorize azo dyes. Water Res 39:
5135-5141. Doi:
10.1016/j.watres.2005.09.033.
Khalid A, Arshad M, Crowley DE (2008)
Decolorization of azo dyes by
Shewanella sp. under saline
conditions. Appl Microbiol Biotechnol
79: 1053-1059. Doi: 10.1007/s00253-
008-1498-y Kiernan JA (2001) Classification and naming
of dyes, stains and fluorochromes. Biotech Histochem 76: 261-278. Doi: 10.1080/bih.76.5-6.261.278
Leelakriangsak M, Borisut S (2012)
Characterization of the decolorizing
activity of azo dyes by Bacillus subtilis
azoreductase AzoR1. Songklanakarin
J Sci Technol 34: 509-516
Lalnunhlimi S, Krishnaswamy V (2016)
Decolorization of azo dyes (Direct Blue
151 and Direct Red 31) by moderately
alkaliphilic bacterial consortium. Braz J
Microbiol 47: 39-46. Doi:
10.1016/j.bjm.2015.11.013
Maheswari NU, Sivagami S (2016) Biological
degradation of textile dyes using

Dekolorisasi Pewarna Tekstil Sumifix Blue... Permatasari et al.
26
marine Bacillus species. Int J Pure App
Biosci 4: 123-128. Doi: 10.18782/2320-
7051.2326 Mumtaz S, Malik NH, Naz I, Ahmed S (2015)
Decolorization of textile dye reactive blue 221 by bacteria isolated from anthropogenic dye-contaminated soil. Pol J Environ Stud 24: 1705-1716
Naresh B, Preethi C, Sneha S, Bhagyashree
R, Parizad P (2013) Microbial decolorization of disperse textile dye brown 21 by Enterobacter gergoviae
isolated from textile effluent. Int Res J Environ Sci 2:31-36
Rawat D, Mishra V, Sharma RS (2016)
Detoxification of azo dyes in the
context of environmental processes.
Chemosphere 155: 591-605. Doi:
10.1016/j.chemosphere.2016.04.068
Shah MP (2014) Biodegradation of azo dyes
by three isolated bacterial strains: An
environmental bioremedial approach. J
Microb Biochem Technol S3:007.
Doi: 10.4172/1948-5948.S3-007
Shah MP, Patel KA, Nair SS, Darji AM
(2013) Microbial degradation of textile
dye (Remazol Black B) by Bacillus
spp. ETL-2012. J Bioremed Biodeg 4:
180-184.Doi: 10.4172/2155-6199.1000180
Sharma S, Munjal A, Gupta S (2011)
Comparative studies on decolorization
of textile azo dyes by different bacterial
consortia and pure bacterial isolate. J
Pharm Res 4: 3180-3183
Sriram N dan Reetha D (2015) Isolation and
characterization of dye degrading
bacteria from textile dye effluents.
Cent Euro J Exp Biol 4: 5-10
Van der Zee FP, Lettinga G, Field JA (2001)
Azo dye decolorisation by anaerobic
granular sludge. Chemosphere
44:1169-1176
Velmurugan S, Revikumar R (2014) Biodegradation and decolorization of reacive dye red ME4BL by Bacillus subtilis. Int J of Environ Bioremed Biodeg 2: 250-255
Vimala G, Jeyakumar P, Devi AC, Singh A, Iyer P (2015) Azo dye degrading bacteria from textile effluent. Int J Curr Microbiol App Sci 4: 199-210
Yan MP, Razak A, Bakar Wan AB (2003) Photocatalytic degradation of sumifix blue. Borneo Sci 13: 63-69

27
VOLUME 5 NOMOR 1 JUNI 2017 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI dan BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
KARAKTERISTIK FISIK PAKAN IKAN APUNG NON-EKSTRUSI YANG
DIBUAT MELALUI FERMENTASI Rhizopus oryzae
Physical Characteristics of Non-Extruded Floating Fish Feed Produced through Rhizopus oryzae Fermentation
Asep Badru Zaman
1, Catur Sriherwanto
2,*, Etyn Yunita
1, Imam Suja’i
2
1Dept. Biologi, Fakultas Sains dan Teknologi, Universitas Islam Negeri (UIN) Syarif Hidayatullah Jakarta
2Bitechnology Laboratory, BPPT, Building 630 PUSPIPTEK Area, Tangerang Selatan, Banten 15314
*E-mail: [email protected]
ABSTRACT To reduce the feed production cost of floating fish feed, an innovative method which is cheaper, easier, and simpler compared to the use of extruder machine is needed to provide buoyancy to fish feed. This research was conducted to determine the physical characteristics of floating fish feed prepared through fermention using Rhizopus oryzae on the mixed substrate consisting of commercial sinking fish feed, cassava bagasse, and Lemna minor. The resulting fermented feed was subsequently compared with commercial sinking fish feed in terms of its water stability, absorption, and floatability. The water stability curves of both feed types showed a similar pattern for 20 hours. Both decreased dramatically in the first 0.5 hours, from 100% to 81% for the fermented feed, and to 83% for the commercial feed. The shape of a 40-minute water absorption curve for both feed types was very similar. The fermented feed absorbed water nearly 4 times its initial dry weight, while commercial feed up to about twice as much as its initial dry weight. For three hours, both fermented and commercial floating pellets stayed afloat 100% without sinking. Thus, fermentation using R. oryzae deserves further development as an alternative way to provide buoyancy to fish feed. Keywords: floatability, floating feed, sinking feed, water absorption, water stability ABSTRAK Untuk menekan biaya produksi pakan ikan apung, diperlukan inovasi pengapungan pakan ikan yang lebih murah, mudah, dan sederhana dibandingkan dengan penggunaan mesin ekstruder. Penelitian ini dilakukan untuk mengetahui karakteristik fisik pakan apung hasil fermentasi menggunakan kapang Rhizopus oryzae pada substrat campuran pakan tenggelam komersial, onggok, dan Lemna minor. Pakan fermentasi ini lalu dibandingkan dengan pakan apung komersial dalam hal stabilitas dalam air, absorbsi air, dan daya apung. Stabilitas dalam air kedua jenis pakan memiliki pola yang mirip selama 20 jam. Stabilitas keduanya menurun drastis pada 0,5 jam pertama, dari 100% ke 81% untuk pakan apung fermentasi, dan ke 83% untuk pakan apung komersial. Bentuk kurva absorpsi air selama 40 menit untuk kedua jenis pakan sangatlah mirip. Pakan apung fermentasi menyerap air hampir 4 kali bobot kering awalnya, sedangkan pakan apung komersial hingga sekitar 2 kali bobot kering awalnya. Selama tiga jam, baik pakan apung fermentasi maupun komersial mengapung 100% tanpa tenggelam. Jadi, fermentasi menggunakan R. oryzae layak dikembangkan lebih lanjut sebagai cara pengapungan alternatif pakan ikan.
Kata Kunci: daya apung, daya serap air, stabilitas dalam air, pakan apung, pakan tenggelam
Received: 16 Maret 2018 Accepted: 07 June 2018 Published: 08 June 2018

Karakteristik Fisik Pakan Ikan Apung... Zaman et al.
28
PENDAHULUAN
Lebih dari 50% dari total biaya
produksi budidaya ikan berasal dari pakan (Rana et al. 2009). Pakan ikan di Indonesia dibuat menggunakan bahan baku yang sebagian besarnya masih didatangkan dari negara lain sehingga berdampak pada mahalnya harga pakan (Rasidi dan Haryadi 2016). Tingginya harga pakan ikan inilah yang mendorong digulirkannya program Gerpari (Gerakan Pakan Ikan Mandiri) oleh Direktorat Pakan, Direktorat Jenderal Perikanan Budidaya, Kementerian Kelautan dan Perikanan agar kedaulatan pangan di sektor perikanan dapat diwujudkan, khususnya agar terciptanya kemandirian pakan dengan menggunakan potensi ketersediaan bahan baku pakan lokal (Dirjen Perikanan Budidaya 2016), dan dengan demikian biaya pembelian pakan pun bisa ditekan dan keuntungan pembudidaya ikan dapat ditingkatkan.
Namun pembuatan pakan mandiri oleh pelaku usaha sektor perikanan terkendala sejumlah faktor, di antaranya adalah kualitas fisik pakan yang mudah pecah dan larut dalam air sehingga cepat menurunkan kualitas air; serta pakan yang mudah tenggelam sebelum dikonsumsi ikan (Kurniawan dan Lestariadi 2017). Pakan yang tidak dimakan ikan akan mengotori kolam ikan, dan selanjutnya akan mengalami penguraian secara mikrobiologis yang menghabiskan sejumlah besar oksigen yang terlarut dalam air kolam tersebut. Selain itu proses ini menaikkan kadar amonia dan nitrit hingga konsentrasi yang dapat meracuni ikan dalam waktu singkat (Somerville et al. 2014). Karena itu, diperlukan teknologi pengapungan pakan tepat guna yang tidak memerlukan biaya investasi dan operasional semahal pembuatan pakan komersial skala industri besar yang melibatkan proses ekstrusi menggunakan mesin ekstruder.
Metode non-ekstrusi ini sebenarnya pernah dilakukan sebelumnya menggunakan khamir Saccharomyces cerevisiae. Namun proses ini masih belum praktis karena masih memerlukan pengetahuan dan ketrampilan khusus tentang penambahan zat pengikat (binding agent), serta dibutuhkan tahapan penggorengan dalam minyak (deep frying)
yang membutuhkan energi panas, minyak goreng, dan fasilitas penggorengan (Saputra 2016; Lindasari 2017).
Mempertimbangkan kondisi tersebut, metode lain yang jauh lebih praktis dan sederhana dalam pembuatan pakan ikan apung sangat diperlukan. Laboratorium Bioteknologi Pakan, Balai Bioteknologi-BPPT, kawasan PUSPIPTEK, Tangerang Selatan, telah mengembangkan metode pengapungan pakan ikan fermentatif menggunakan Rhizopus sp. (Sriherwanto et
al. 2017; Leiskayanti et al. 2017). Fermentasi ini tidak membutuhkan sterilisasi alat maupun bahan, tanpa mempertimbangkan jenis serta komposisi zat pengikat (binder),
dilakukan secara non-ekstrusi atau tanpa menggunakan mesin pembuat pakan apung (ekstruder), dan tanpa tahapan deep frying
sehingga lebih hemat energi dan jauh lebih praktis. Penelitian ini dilakukan dalam rangka mengetahui karakteristik fisik pakan apung hasil fermentasi menggunakan kapang R. oryzae pada substrat yang merupakan
campuran dari pakan komersial tenggelam, onggok dan gulma bebek (Lemna minor). Sifat fisik yang diuji meliputi absorpsi air (water absorption), stabilitas dalam air (water stability), dan daya apung (floatability).
BAHAN DAN METODE
Waktu dan lokasi penelitian
Penelitian dilakukan pada bulan Januari-September 2016 di Laboratorium Bioteknologi Pakan, Balai Bioteknologi-BPPT, Setu, Tangerang Selatan, Banten. Penelitian ini menggunakan metode komparatif dengan 2 sampel yang berbeda: pakan apung komersial Hi-Pro-Vite 781 (PT. Central Proteina Prima Tbk dengan kandungan protein, lemak, dan serat masing-masing sekitar 31-33%, 4-6%, dan 3-5%) sebagai kontrol dan pakan apung fermentasi yang dibuat dalam penelitian ini. Penelitian ini meliputi proses pembuatan pakan ikan fermentasi, pengujian karakteris-tik fisik pakan fermentasi yang meliputi sta-bilitas (keutuhan dan kekokohan) dalam air, absorpsi (daya serap) air, dan daya apung.
Fermentasi pakan
Inokulum ragi tempe yang mengandung R. oryzae 1 g (105-106 cfu/g) dicampurkan dengan 50 g substrat yang merupakan campuran dari onggok singkong 30 g, pakan komersial tenggelam P99 (PT. Central Proteina Prima Tbk) 15 g, gulma

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
29
bebek (L. minor) segar 5 g. Setelah diaduk
merata, sebanyak 40 mL larutan mineral yang mengandung KH2PO4 (2,6 g/L), urea (2 g/L), dan (NH4)2SO4 (8,1 g/L) ditambahkan, diaduk lagi hingga merata. Campuran tersebut lalu ditempatkan ke dalam cawan petri kaca (diameter 9 cm, tebal 1 cm), diinkubasi pada suhu 28-30°C (suhu ruang) selama 40 jam. Pakan hasil fermentasi diiris
menggunakan cutter dengan ukuran 0,5 0,5 cm pada kertas grafik, lalu dikeringkan di dalam oven bersuhu 50°C selama 24 jam. Uji stabilitas dalam air
Menggunakan metode Misra et al. (2002), stabilitas dalam air atau kekokohan dan kekompakan dalam air diukur dengan menempatkan 2-3 g sampel dalam saringan teh plastik berukuran ± 200 mesh (Erizal et al. 2016). Sampel dan saringan ini lalu direndam dalam gelas beaker berisi air 500 mL, diberi gelembung udara 3,5 L/menit menggunakan aerator akuarium (Luckiness, L828, Cina). Perendaman dilakukan selama ½, 1, 2, 4, 10, dan 20 jam. Sampel yang telah menyerap air ditiriskan, lalu dioven selama 24 jam pada suhu 60°C hingga kering. Stabilitas dalam air sampel ditentukan menggunakan rumus:
100(%)o
t DW
DWairdalamStabilitas
dimana: DWt : berat kering sesudah perendaman (g) DWo : berat kering sebelum perendaman (g)
Uji absorpsi Air
Uji ini memiliki prosedur sebagaimana uji stabilitas dalam air, bedanya setelah perendaman selama 1, 3, 5, 10, 20 dan 40 menit, sampel ditimbang dalam keadaan basah tanpa pengeringan terlebih dahulu. Absorpsi air dihitung menggunakan rumus berikut (Misra et al. 2002) :
100(%)
o
ot
DW
DWWWairAbsorpsi
dimana: WWt : berat basah setelah direndam air (g) DWo : berat kering sebelum direndam air (g)
Uji daya apung
Daya apung diukur menggunakan metode Cruz et al. (2015) yang telah dimodifikasi, dan dilakukan dalam kondisi tanpa aerator. Sebanyak 10 butir pakan dijatuhkan ke dalam gelas beaker 500 mL yang berisi air 400 mL. Jumlah sampel yang masih mengapung dicatat dalam rentang waktu tertentu, dimulai pada menit ke-0 hingga menit ke-180. Rumus daya apung:
100(%) i
f
F
FapungDaya
dimana: Ff : sisa sampel yang mengapung Fi : jumlah awal sampel (10 butir) Analisis data
Data hasil penelitian disajikan grafik yang bersifat komparatif. Analisis data hasil pengujian fisik dilakukan secara deskriptif serta analisis statistik parametrik (Independent Sample t-test) untuk melihat
perbedaan nyata kedua sampel. HASIL DAN BAHASAN Pengamatan fisik pakan fermentasi
Pencampuran tepung onggok (Gambar 1a), L. minor (Gambar 1b), pakan tenggelam
komersial (Gambar 1c), dan larutan mineral menghasilkan substrat campuran berupa butiran-butiran basah yang terpisah satu sama lain dan tidak rekat dengan warna dominan coklat. Teksturnya mirip seperti dedak padi basah namun tidak berair (Gambar 1d). Setelah fermentasi menggunakan R. oryzae, butiran substrat
yang awalnya terpisah tersebut (Gambar 1d) di akhir fermentasi berubah menjadi satu kesatuan padatan yang menyerupai bentuk cawan petri. Satu butiran substrat dengan yang lainnya diikat oleh jalinan miselia jamur sehingga membentuk tekstur seperti tempe kedelai dan kenyal seperti keju (Gambar 1e).
Saat penyiapan bahan baku komponen pakan, tidak ada pertimbangan penambahan komponen bahan pengikat dan perakat khusus, atau perlakuan mekanis dan termal yang sengaja dilakukan untuk merekatkan dan menyatukan substrat yang bertekstur butiran terpisah tersebut. Namun sesudah fermentasi, butiran-butiran substrat

Karakteristik Fisik Pakan Ikan Apung... Zaman et al.
30
Gambar 1. Bahan pakan onggok (a), L. minor (b), pakan tenggelam komersial (c), dicampur dengan larutan mineral menghasilkan substrat campuran (d) yang, setelah difermentasi oleh R. oryzae, membentuk tekstur menyerupai tempe kedelai (e).
tersatukan membentuk satu kesatuan tekstur yang padat, kompak, dan kokoh. Hal ini menunjukkan bahwa miselia R. oryzae
mampu menggantikan peran zat perekat dan pengikat yang biasanya ditambahkan dalam pembuatan pakan ikan bentuk pelet yang bahan baku awalnya berupa tepung halus. Prosedur mekanis dan termal yang biasa digunakan untuk memunculkan kekompakan, kekokohan, dan daya apung pada pelet juga tidak diperlukan dalam fermentasi ini.
Pakan apung fermentasi dan pakan apung komersial memiliki sejumlah perbedaan fisik (Gambar 2). Pakan apung komersial memiliki warna coklat gelap sedangkan pakan apung fermentasi berwarna lebih muda dan terang. Warna coklat gelap pada pakan apung komersial kemungkinan disebabkan oleh reaksi Maillard, yakni reaksi antara asam amino dengan gula pereduksi pada suhu tinggi (lebih dari 100°C) pada proses ekstrusi (Ljøkjel et al. 2004; Von Rooijen 2016). Hal ini tidak terjadi pada fermentasi pakan yang menggunakan suhu ruangan, yakni sekitar 28-30°C. Pembentukan miselia putih kapang Rhizopus justru menjadikan pakan
fermentasi berubah warna menjadi lebih muda dan keputihan. Pakan fermentasi diperkecil dengan pengirisan pisau cutter
hingga berbentuk balok berukuran panjang
lebar tinggi sekitar 5510 mm, sedangkan pakan apung komersial hampir menyerupai bentuk bulat bola dengan garis tengah sekitar 4 mm. Berbeda dengan pakan tenggelam komersial yang tidak dapat mengapung, pakan fermentasi ini memiliki daya apung yang baik (Gambar 3).
Stabilitas dalam air
Stabilitas dalam air kedua jenis pakan memiliki pola yang sama (Gambar 4) selama 20 jam, dimana keduanya mengalami penurunan drastis pada 0,5 jam pertama yaitu dari 100% turun ke 83% untuk pakan apung komersial dan 81% untuk pakan apung fermentasi. Antara jam ke-0,5 hingga jam ke-10, pakan apung komersial memperlihatkan nilai stabilitas dalam air yang lebih baik dari pakan apung fermentasi, terutama pada jam ke- 2 dimana selisihnya terbesarnya mencapai 4,45%. Secara umum, adanya selisih yang tidak terlalu besar selama 20 jam perendaman dalam air ini menunjukkan bahwa stabilitas dalam air

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
31
Gambar 2. Penampilan fisik pakan apung fermentasi (kiri) dan pakan apung komersial (kanan) dalam kemasan
dengan berat masing-masing 100 g
Gambar 3. Pakan apung fermentasi yang telah dikeringkan mampu mengapung pada permukaan air.
pada pakan apung fermentasi mendekati nilai stabilitas dalam air pada pakan apung komersial. Uji ini memberikan kesimpulan bahwa pakan apung dengan bahan onggok, L. minor dan pakan tenggelam yang difermentasi oleh R. oryzae tanpa
penambahan bahan pengikat khusus dan tanpa menggunakan mesin canggih mampu menyamai tingkat stabilitas dalam air pada pakan apung komersial yang biasa digunakan oleh pembudidaya ikan.
Berbeda dengan pakan untuk ternak, pakan untuk ikan budidaya memerlukan tahapan-tahapan proses tertentu untuk memastikan bahwa pelet yang dihasilkan memiliki stabilitas yang baik dalam air. Stabilitas ini mesti cukup untuk memberi waktu bagi hewan air untuk menyantapnya. Untuk itu, ekstrusi dengan atau tanpa zat pengikat (perekat) yang menyatukan butiran-butiran komponen bahan pakan ikan
diperlukan dalam rangka tetap mengokohkan atau menstabilkan butiran-butiran pakan tersebut agar tidak mudah terurai hancur dalam air, dan memastikan agar sesedikit mungkin zat-zat gizi dalam pakan yang hilang terlarut ke dalam air kolam (Hashim dan Saat 1992; Paolucci et al. 2012).
Zat perekat khusus tidaklah digunakan dalam pembuatan pakan ikan apung dalam eksperimen ini, sebagaimana pada pembuatan pakan komersial yang melibatkan penggunaan zat pengikat yang tergolong karbohidrat seperti agar-agar, alginat, pektin, kitosan, karaginan, karboksimetilselulosa, guargam, maupun dari jenis protein seperti gelatin (Paolucci et al. 2012). Penyatuan butiran-butiran pakan dalam percobaan ini adalah agen hayati berupa miselium kapang tempe, yakni Rhizopus sp., yang sudah dikenal dapat

Karakteristik Fisik Pakan Ikan Apung... Zaman et al.
32
Gambar 4. Stabilitas dalam air dari pakan apung komersial dan pakan apung fermentasi
menyatukan butiran kedelai pada pembuatan tempe kedelai melalui fermentasi pada suhu kamar.
Penggunaan kapang tempe sebagai agen hayati pengikat ini juga menghindari kerusakan nutrisi yang rentan terhadap panas sebagaimana yang terjadi pada proses ekstrusi yang dapat merusak vitamin tertentu (Riaz et al. 2009). Fermentasi Rhizopus bahkan sudah terbukti malah
memperkaya vitamin tertentu pada substrat media tumbuhnya (Sriherwanto 2010).
Absorpsi air
Secara umum, kedua bentuk kurva absorpsi air selama 40 menit untuk kedua jenis pakan sangatlah mirip. Keduanya sangat cepat menyerap air di menit pertama, dimana pakan apung fermentasi sekitar tiga kali lebih kuat menyerap air dibandingkan pakan apung komersial. Proses penyerapan air oleh kedua jenis pakan ini naik tapi dengan kecepatan daya serap yang menurun setelah menit ke-1 hingga menit ke-10. Setelah itu pakan apung komersial tidak lagi mampu menyerap air setelah penyerapannya mencapai sekitar 200% atau 2 kali bobot kering awal. Di menit ke-10, pakan apung fermentasi mampu menyerap air hingga 361%, yakni hampir 4 kali lipat
lebih berat dibandingkan bobot kering pakan tersebut. Namun absorpsi air pakan fermentasi ini menurun di menit ke-20, lalu naik lagi sedikit di menit ke-40. Penurunan ini diduga karena tingkat kekompakan pakan berkurang yang menyebabkan sebagian material pakan tersebut hancur dan meluruh.
Lebih tingginya kemampuan menyerap air pakan apung fermentasi dibandingkan pakan apung komersial mungkin dikarenakan volume total rongga udara yang lebih besar pada pakan fermentasi dibandingkan pada pakan komersial. Besarnya volume total rongga udara ini diduga karena hifa kapang R. oryzae mampu menembus ke dalam butiran substrat sehingga membuat jalinan lorong-lorong udara mikro di dalam substrat. Selain itu, miselium kapang memenuhi rongga antar butiran substrat, sehingga memerangkap udara di celah-celah antar-substrat. Metabolisme nutrisi substrat menjadi senyawa sederhana yang mudah menguap (volatile low molecular weight compounds) juga meninggalkan rongga-
rongga udara mikro dalam substrat. Tersedianya ruang-ruang udara mikro di sela-sela jalinan padat miselium dan di dalam substrat juga memungkinkan air terserap oleh pakan fermentasi secara lebih
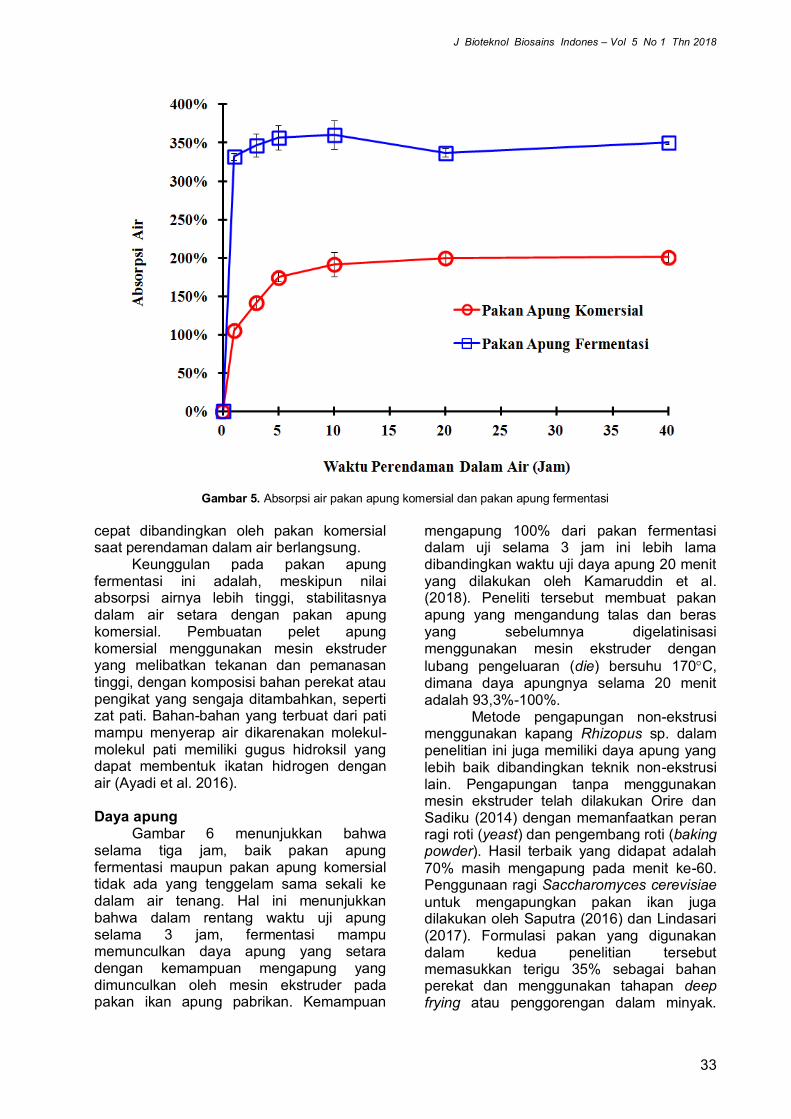
J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
33
Gambar 5. Absorpsi air pakan apung komersial dan pakan apung fermentasi
cepat dibandingkan oleh pakan komersial saat perendaman dalam air berlangsung.
Keunggulan pada pakan apung fermentasi ini adalah, meskipun nilai absorpsi airnya lebih tinggi, stabilitasnya dalam air setara dengan pakan apung komersial. Pembuatan pelet apung komersial menggunakan mesin ekstruder yang melibatkan tekanan dan pemanasan tinggi, dengan komposisi bahan perekat atau pengikat yang sengaja ditambahkan, seperti zat pati. Bahan-bahan yang terbuat dari pati mampu menyerap air dikarenakan molekul-molekul pati memiliki gugus hidroksil yang dapat membentuk ikatan hidrogen dengan air (Ayadi et al. 2016). Daya apung
Gambar 6 menunjukkan bahwa selama tiga jam, baik pakan apung fermentasi maupun pakan apung komersial tidak ada yang tenggelam sama sekali ke dalam air tenang. Hal ini menunjukkan bahwa dalam rentang waktu uji apung selama 3 jam, fermentasi mampu memunculkan daya apung yang setara dengan kemampuan mengapung yang dimunculkan oleh mesin ekstruder pada pakan ikan apung pabrikan. Kemampuan
mengapung 100% dari pakan fermentasi dalam uji selama 3 jam ini lebih lama dibandingkan waktu uji daya apung 20 menit yang dilakukan oleh Kamaruddin et al. (2018). Peneliti tersebut membuat pakan apung yang mengandung talas dan beras yang sebelumnya digelatinisasi menggunakan mesin ekstruder dengan
lubang pengeluaran (die) bersuhu 170C, dimana daya apungnya selama 20 menit adalah 93,3%-100%. Metode pengapungan non-ekstrusi menggunakan kapang Rhizopus sp. dalam penelitian ini juga memiliki daya apung yang lebih baik dibandingkan teknik non-ekstrusi lain. Pengapungan tanpa menggunakan mesin ekstruder telah dilakukan Orire dan Sadiku (2014) dengan memanfaatkan peran ragi roti (yeast) dan pengembang roti (baking powder). Hasil terbaik yang didapat adalah
70% masih mengapung pada menit ke-60. Penggunaan ragi Saccharomyces cerevisiae
untuk mengapungkan pakan ikan juga dilakukan oleh Saputra (2016) dan Lindasari (2017). Formulasi pakan yang digunakan dalam kedua penelitian tersebut memasukkan terigu 35% sebagai bahan perekat dan menggunakan tahapan deep frying atau penggorengan dalam minyak.

Karakteristik Fisik Pakan Ikan Apung... Zaman et al.
34
Gambar 6. Uji daya apung pakan apung komersial dan pakan apung fermentasi tanpa aerator
Pakan ikan yang dihasilkan mampu mengapung selama 6-8 jam. Proses pemunculan sifat apung pada pakan ini kurang efisien dibandingkan dengan yang dilakukan dalam penelitian ini, yang tidak perlu mempertimbangkan penambahan zat perekat, dan tanpa penggorengan dalam minyak yang memerlukan tambahan ongkos energi, bahan, dan sarana produksi lainnya. KESIMPULAN
Fermentasi substrat campuran pakan
tenggelam komersial, onggok, dan gulma bebek menggunakan kapang R. oryzae
memunculkan sifat apung pada produk hasil fermentasinya. Saat dibandingkan dengan pakan apung komersial, pakan apung fermentasi ini memiliki pola kurva stabilitas dalam air yang mirip dengan pakan apung komersial selama 20 jam dalam air. Stabilitas keduanya sama-sama mengalami penurunan drastis pada 30 menit pertama, dari 100% turun hingga ke 81-83%. Demikian pula dengan bentuk kurva absorpsi air selama 40 menit pengujian, kedua jenis pakan tersebut memperlihatkan sifat fisik penyerpan air yang sangat mirip. Pakan apung fermentasi menyerap air
hampir 4 kali berat kering awalnya, sedangkan pakan apung komersial hingga sekitar 2 kali berat kering awalnya. Selama tiga jam pengujian daya apung, kedua jenis pakan mengapung 100% tanpa satu butir pelet pun yang tenggelam. Jadi, fermentasi bahan pakan ikan menggunakan R. oryzae
layak dikembangkan lebih lanjut sebagai cara pengapungan alternatif pakan ikan agar karakteristik fisiknya semakin baik dan mendekati pakan apung ekstrusi.
REFERENSI
Ayadi F, Rosentrater KA, Muthukumarappan
K, Kannadhason S (2016) Effects of Amylose-to-amylopectin ratios on binding capacity of DDGS/soy-based aquafeed blends. J Food Res 5:43. doi: 10.5539/jfr.v5n5p43
De Cruz CR, Kamarudin MS, Saad CR, Ramezani-Fard E (2015) Effects of extruder die temperature on the physical properties of extruded fish pellets containing taro and broken rice starch. Anim Feed Sci Technol 199:137-145. doi: 10.1016/j.anifeedsci.2014.11.010
Dirjen Perikanan Budidaya (2016) Harapan

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
35
pada bantuan pakan mandiri. akuakultur indonesia 24:7
Erizal E, Lana M, Setyo R, Abbas B (2016) Sintesis dan karakterisasi hidrogel superabsorben berbasis asam akrilat hasil iradiasi gamma. J Ilm Apl Isot dan Radiasi 11:27-38. doi: 10.17146/jair.2015.11.1.2697
Hashim R, Saat MAM (1992) The utilization of seaweed meals as binding agents in pelleted feeds for snakehead (Channa striatus) fry and their effects on
growth. Aquaculture 108:299-308. doi: 10.1016/0044-8486(92)90114-Z
Kamarudin MS, de Cruz CR, Saad CR, Romano N, Ramezani-Fard E (2018) Effects of extruder die head temperature and pre-gelatinized taro and broken rice flour level on physical properties of floating fish pellets. Anim Feed Sci Technol 236:122-130. doi: 10.1016/j.anifeedsci.2017.12.007
Kurniawan A, Lestariadi RA (2017) Induction of fish pellet making machine to improve feed community program in catfish farmers in mojokerto regency. J Innov Appl Technol 3:433-438. doi: 10.21776/ub.jiat.2017.003.01.16
Leiskayanti Y, Sriherwanto C, Suja'i I (2017) Fermentasi menggunakan ragi tempe sebagai cara biologis pengapungan pakan ikan. J Bioteknol Biosains Indones 4:54-63. doi: 10.29122/jbbi.v4i2.2503
Ljøkjel K, Sørensen M, Storebakken T, Skrede A (2004) Digestibility of protein, amino acids and starch in mink (Mustela vison) fed diets processed by
different extrusion conditions. Can J Anim Sci 84:673-680. doi: 10.4141/A01-089
Lindasari A (2017) Pembuatan pakan terapung terfermentasi Saccharomyces cerevisiae melalui proses non-ekstrusi. Undergraduate Thesis. Bogor Agricultural University
Misra CK, Sahu NP, Jain KK (2002) Effect of extrusion processing and steam pelleting diets on pellet durability, water absorption and physical response of Macrobrachium rosenbergii. ASIAN Australas J Anim
Sci 15:1354-1358. doi: 10.5713/ajas.2002.1354
Orire AM, Sadiku, SOE (2014) Development of farm made floating feed for aquaculture species. J Int Sci Publ: Agriculture dan Food 2:521-523
Paolucci M, Fabbrocini A, Volpe MG, Varricchio E, Coccia E (2012) Development of biopolymers as binders for feed for farmed aquatic organisms. In: Muchlisin Z (ed) Aquaculture, InTech, Rijeka, Croatia, pp 3-34. http://cdn.intechweb.org/pdfs/ 27101.pdf. Accessed 10 October 2017
Rana KJ, Siriwardena S, Hasan MR (2009) Impact of rising feed ingredient prices on aquafeeds and aquaculture production (No. 541) (p. ix). Food and Agriculture Organization of the United Nations (FAO)
Rasidi R, Haryadi J (2016) Evaluasi kebijakan pengembangan pakan mandiri. In prosiding forum inovasi teknologi akuakultur 1:689-702
Riaz MN, Asif M, Ali R (2009) Stability of vitamins during extrusion. Crit Rev Food Sci Nutr 49:361-368. doi: 10.1080/10408390802067290
Saputra RA (2016) Uji kualitas fisik pakan terapung terfermentasi Saccharomyces cerevisiae melalui proses non-ekstrusi
dan deep frying. Undergraduate Thesis. Bogor Agricultural University
Somerville C, Cohen M, Pantanella E, Stankus A, Lovatelli A (2014) Fish in aquaponics. In: Small-scale aquaponic food production: integrated fish and plant farming, FAO Fisheries and Aquaculture Technical Paper No 589 (pp. 103-121). Food and Agriculture Organization of the United Nations, Rome
Sriherwanto C (2010) Studies on the solid state fermentation of cassava bagasse for animal feed. Doctoral Thesis, Department of Chemistry, University of Hamburg, Germany
Sriherwanto C, Suja'i I, Soraya S (2017) Pemanfaatan kapang Rhizopus sp.
sebagai agen hayati pengapung pakan ikan. J Mikol Indones 1:70-81
Van Rooijen C (2016) Effect of extrusion conditions on the Maillard reaction and in vitro digestibility in two dry dog foods. In: Maillard reaction products in pet foods. PhD Thesis. Wageningen University

Perbanyakan In Vitro Pisang Kepok... Imelda et al.
36
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
PERBANYAKAN IN VITRO PISANG KEPOK var. UNTI SAYANG
TAHAN PENYAKIT DARAH MELALUI PROLIFERASI TUNAS
In Vitro Propagation of Kepok Banana var. Unti Sayang Resistant to Blood Disease through Shoot Proliferation
Maria Imelda, Aida Wulansari*, Laela Sari
Pusat Penelitian Bioteknologi LIPI. Jalan Raya Bogor Km 46, Cibinong 16911, Jawa Barat *E-mail: [email protected]
ABSTRACT Kepok is a popular banana variety but sensitive to blood disease caused by Ralstonia solanacearum (Smith). The discovery of a natural mutant of Kepok banana var. Unti Sayang from Sulawesi which male bud falls naturally, is a shortcut to bypass the chains of the spread of blood disease, since the disease is transmitted by insects through the wounds of the male buds. The superior mutant needs to be mass propagated and disseminated to endemic areas to inhibit the spread of blood disease. To achieve that goal, an efficient and effective techniques of in vitro shoot proliferation needs to be developed. Shoot proliferation was performed by addition of BAP, thidiazuron and adenine sulphate. The results showed that the best medium for shoot multiplication was B2T5A (MS+2 mg/L BAP+0,5 mg/L TDZ+20 mg/L adenine sulphate), and for shoot growth was B4A (MS+4 mg/L BAP+20 mg/L adenine sulphate). Rooting was induced on MS medium without hormones. Acclimatization of plantlets on mixed soil, compost and husks with a ratio of 1:1:1 resulted in 92,35% survival rate.
Keywords: blood disease, in vitro shoot, male budless, natural mutant, var. Unti Sayang ABSTRAK Pisang kepok merupakan varietas yang digemari tetapi sangat peka terhadap penyakit darah yang ditimbulkan oleh bakteri Ralstonia solanacearum (Smith). Ditemukannya mutan alami pisang kepok yang jantungnya gugur secara alami yaitu varietas Unti Sayang dari Sulawesi, merupakan jalan pintas untuk memotong rantai penyebaran penyakit darah, mengingat penyakit ini ditularkan oleh serangga melalui luka bekas bunga jantan pada jantung. Mutan unggul tersebut perlu diperbanyak secara massal dan disebarluaskan ke daerah endemik untuk menghambat penyebaran penyakit darah. Untuk mencapai tujuan tersebut, perlu dikembangkan teknik perbanyakan in vitro pisang kepok Unti Sayang yang efektif dan efisien melalui proliferasi tunas. Proliferasi tunas dilakukan dengan penambahan BAP, thidiazuron dan adenin sulfat. Hasil penelitian ini menunjukkan bahwa media terbaik untuk multiplikasi tunas adalah B2T5A (MS+2 mg/L BAP+0,5 mg/L TDZ+20 mg/L adenin sulfat), media terbaik untuk pertumbuhan tunas adalah B4A (MS+4 mg/L BAP+20 mg/L adenin sulfat). Akar dapat diinduksi pada media MS tanpa hormon. Aklimatisasi planlet pada media campuran tanah, kompos dan sekam dengan perbandingan 1:1:1 menghasilkan 92,35% planlet hidup.
Kata Kunci: penyakit darah, tunas in vitro, tanpa jantung, mutan alami, var. Unti Sayang
Received: 18 January 2018 Accepted: 22 May 2018 Published: 08 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
37
PENDAHULUAN
Pisang merupakan salah satu
komoditas unggulan Indonesia yang berpotensi menunjang ketahanan pangan nasional. Pisang termasuk buah yang banyak digemari, nutrisinya tinggi dan hampir semua bagian tanaman dapat dimanfaatkan. Pisang juga memiliki produktivitas dan daya adaptasi yang tinggi (Suhartanto et al. 2012). Selain itu harga pisang terjangkau dan dapat dikonsumsi baik sebagai buah segar maupun dalam bentuk olahan.
Pisang kepok merupakan salah satu jenis pisang olahan. Pisang kepok memiliki nilai ekonomis yang tinggi karena rasanya yang enak dan manis dengan kadar gula mencapai 300 Brix (Poerba et al. 2012). Salah satu jenis pisang kepok yang dibudidayakan di daerah Sulawesi Selatan yaitu pisang kepok (Musa acuminata x M. balbisiana) var. Unti Sayang atau dikenal juga
dengan nama pisang Amorang. Berdasarkan deskripsi pada SK Mentan Nomor 2048/Kpts/SR.120/5/2010 tentang pelepasan pisang kepok Unti Sayang sebagai varietas unggul disebutkan bahwa pisang ini memiliki keunggulan produktivitas tinggi, tidak memiliki bunga jantan, kulit buah berwarna kuning dan halus, daging buah berwarna kuning dengan rasa manis, buah tidak mudah lepas dari sisirnya serta beradaptasi dengan baik di dataran rendah. Produktivitas pisang kepok var. Unti Sayang mencapai 40 kg per tanaman (Suhartanto et al. 2012).
Pisang kepok var. Unti Sayang sulit diserang penyakit layu bakteri dan penyakit darah karena memiliki pertahanan alami yaitu tidak mempunyai jantung (bunga jantan) sehingga terhindar dari serangan serangga vektor kedua penyakit tersebut (Poerba et al. 2012). Menurut Hermanto dan Emilda (2013) pada percobaan lapang di daerah endemik penyakit darah, pisang kepok yang tidak memiliki bunga jantan atau jantung terbukti lebih tahan terhadap penyakit darah dibandingkan pisang kepok yang berjantung.
Penyebab penyakit darah adalah bakteri Ralstonia solanacearum (Smith) (dahulu dinamakan Pseudomonas solanacearum) (Maramis 2014). Cara
penularannya adalah oleh bakteri yang masuk melalui bekas luka dari bunga jantan
yang gugur. Pada tahap selanjutnya, bakteri dapat ditularkan melalui tanah yang terbawa air dan menginfeksi melalui luka pada akar (Supriati 2010). Selama ini cara mengurangi penyebaran penyakit darah yang paling efektif adalah dengan membuang jantungnya setelah semua bunga betina menjadi buah. Namun, pada areal yang luas cara tersebut tidak efisien karena sulit untuk menghilangkan jantung dari setiap tanaman, selain memakan banyak waktu dan tenaga kerja. Ditemukannya pisang kepok var. Unti Sayang yang jantungnya gugur setelah semua bunga betina menjadi buah (male budless mutant) sangat membantu dalam
memotong rantai penyebaran penyakit darah. Penularan penyakit juga dapat terjadi
melalui penanaman bibit yang sudah terinfeksi, tapi hal ini dapat diatasi dengan penggunaan bibit hasil kultur jaringan yang bersih dari penyakit (Eden-Green 1994). Bibit pisang hasil kultur jaringan telah banyak digunakan untuk menghindari penularan penyakit. Teknik kultur jaringan lebih efektif dan efisien dalam menghasilkan bibit secara massal dalam waktu yang relatif lebih singkat dibandingkan metode konvensional. Bibit tersebut tetap memiliki sifat unggul dari induknya dengan tingkat keseragaman yang cukup tinggi. Selain itu, penyediaan bibit pisang hasil kultur jaringan secara komersial dapat dijadwalkan sesuai kebutuhan (Supriati 2010).
Penggunaan media dasar dan zat pengatur tumbuh (ZPT) yang tepat merupakan faktor penting yang perlu diperhatikan dalam perbanyakan bibit menggunakan teknik kultur jaringan sehingga dapat diperoleh hasil yang optimum. Tujuan penelitian ini adalah untuk mengembangkan teknologi perbanyakan tanaman pisang kepok var. Unti Sayang yang tahan penyakit darah melalui proliferasi tunas in vitro dalam
menunjang penyediaan bibit secara massal untuk disebarluaskan ke daerah endemik sebagai jalan pintas dalam memotong rantai penyebaran penyakit darah. BAHAN DAN METODE Bahan
Bahan tanaman yang digunakan sebagai sumber eksplan adalah anakan pisang kepok Unti Sayang yang sehat berukuran sekitar 40 cm. Bahan-bahan

Perbanyakan In Vitro Pisang Kepok... Imelda et al.
38
untuk pembuatan media dasar MS (Murashige dan Skoog 1962), BAP, TDZ, adenin sulfat, bahan pemadat (agar), sukrosa, aquades, sabun pencuci, larutan desinfektan 30% (bahan aktif NaClO 5,25%) dan aquades steril. Alat-alat yang digunakan adalah timbangan analitik, pH meter, autoklaf, laminar air flow cabinet, botol kultur, cawan
petri, scalpel, pinset, pembakar Bunsen, beaker glass, pipet ukur, magnetic stirrer, kertas saring dan plastic wrap.
Metode Inisiasi kultur
Anakan pisang yang sehat dan berukuran 40 cm dibuang akar dan pucuknya, demikian pula seludang bagian luar dan bonggol bawahnya sampai berukuran +10 cm. Selanjutnya bahan tersebut dicuci bersih dengan air mengalir lalu disterilkan dalam larutan desinfektan 30% (bahan aktif NaClO 5,25%) selama 30
menit dan dibilas beberapa kali dengan aquades steril. Semua pekerjaan tersebut dilakukan dalam laminar air flow cabinet.
Setelah itu seludang luarnya dibuang lagi beberapa lapis, begitu juga bonggol bawah dan pucuknya hingga mencapai ukuran +1,5 cm dan dibelah 4 kemudian ditanam dalam media MS yang telah disiapkan sebelumnya. Semua kultur tersebut diinkubasikan selama 4 minggu dalam ruangan bersuhu 25-26ºC dengan pencahayaan 16 jam per hari.
Proliferasi dan pengakaran tunas
Media yang digunakan untuk proliferasi tunas in vitro adalah media padat
dengan komposisi MS, penambahan sukrosa 20 g dan gelrite 2,5 g/L. Kemasaman media diatur sampai mencapai pH 5,8 dengan penambahan larutan KOH atau HCl. Selanjutnya media diautoklaf pada suhu 121ºC dan tekanan 1 atm selama 15 menit. Perlakuan yang diberikan untuk menginduksi proliferasi tunas adalah penambahan zat pengatur tumbuh BAP sebanyak 2-5 mg/L, baik dengan maupun tanpa thidiazuron (TDZ) 0,5 mg/L dan adenin sulfat 20 mg/L ke dalam media MS (Tabel 1). Semua kultur perlakuan diinkubasikan selama 12 minggu dalam ruangan bersuhu 25-26ºC dengan pencahayaan 16 jam per hari. Parameter yang diamati yaitu jumlah tunas dan tinggi tunas setiap 4 minggu.
Pengakaran tunas dilakukan pada media MS tanpa ZPT. Tunas yang sudah besar dipisahkan dari tunas anakan yang masih kecil, kemudian ditanam pada media tanpa ZPT untuk menginduksi akar.
Aklimatisasi planlet
Planlet yang sudah cukup besar dan berakar banyak dibersihkan dari sisa-sisa agar dengan menambahkan air ke dalam botol kultur sehingga tidak banyak akar yang rusak. Selanjutnya akar planlet tersebut dicuci dengan air mengalir dan dikeringkan. Setelah itu planlet ditanam dalam pot plastik berisi media tanah, kompos dan sekam bakar (1:1:1), yang telah disiram sampai jenuh. Pot disungkup dengan kantung plastik transparan yang telah dilubangi selama ±2 minggu sampai muncul daun baru dan ditempatkan dalam kamar kaca.
Tabel 1. Perlakuan zat pengatur tumbuh (ZPT)
Konsentrasi ZPT (mg/L) Kode media
BAP TDZ Adenin
1 0 0 B1
1 0 20 B1A
1 0,5 0 B1T5
1 0,5 20 B1T5A
2 0 0 B2
2 0 20 B2A
2 0,5 0 B2T5
2 0,5 20 B2T5A
3 0 0 B3
3 0 20 B3A
3 0,5 0 B3T5
3 0,5 20 B3T5A
4 0 0 B4
4 0 20 B4A
4 0,5 0 B4T5
4 0,5 20 B4T5A
5 0 0 B5
5 0 20 B5A
5 0,5 0 B5T5
5 0,5 20 B5T5A
0 0 0 Kontrol

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
39
Analisis data
Penelitian ini menggunakan Rancangan Acak Lengkap. Setiap perlakuan diulang 3 kali (botol) dan setiap botol berisi 3 eksplan. Hasil penelitian dianalisis menggunakan Analysis of Variance (ANOVA) dan dilanjutkan dengan Duncan Multiple Range Test (DMRT) pada taraf
nyata 5%.
HASIL DAN PEMBAHASAN Inisiasi kultur
Eksplan berupa potongan bonggol pisang yang dikulturkan pada media MS tanpa penambahan ZPT mulai tumbuh sekitar 7-10 hari setelah tanam. Pada tahap selanjutnya eksplan yang tidak dipindahkan ke media yang mengandung ZPT menghasilkan tunas tunggal. Proliferasi dan pertumbuhan tunas
Tunas tunggal yang disubkultur ke media MS dengan penambahan ZPT dari kelompok sitokinin (BAP dan TDZ) serta adenin sulfat menghasilkan tunas majemuk (Gambar 1). Pada konsentrasi yang tepat, kombinasi pemberian zat pengatur tumbuh BAP dan TDZ serta adenin sulfat dapat menghasilkan banyak sekali tunas. Tunas terbanyak diperoleh pada media B2T5A (2 mg/L BAP, 0,5 mg/L TDZ dan 20 mg/L adenin sulfat) yang menghasilkan 35,44 tunas pada umur 8 minggu dan 36,81 tunas pada umur 12 minggu. Pada media kontrol, tunas yang dihasilkan hanya 3 pada umur 12 minggu (Tabel 2).
Pengamatan terhadap jumlah tunas yang terbentuk menunjukkan bahwa pada konsentrasi BAP yang rendah (1 atau 2
mg/L), pemberian 0,5 mg/L TDZ dapat meningkatkan jumlah tunas, namun pada konsentrasi BAP yang tinggi (5 mg/L) penambahan 0,5 mg/L TDZ tidak berpengaruh nyata (Tabel 2, Gambar 2). Respon serupa ditunjukkan pada tanaman manggis, kombinasi BAP 2 atau 4 mg/L dengan TDZ 0,05 mg/L menghasilkan multiplikasi tunas terbaik (Lestari et al. 2013). Menurut Guo et al. (2011), interaksi antara BAP dan TDZ bersifat saling melengkapi, karena keduanya dapat terikat pada reseptor yang sama yaitu cytokinin-binding protein (CBP). Reseptor ini yang memiliki 2 sisi aktif pengikatan yaitu satu sisi mengikat sitokinin alami berbasis adenin (BAP) sedangkan sisi yang lainnya mengikat sitokinin berbasis phenylurea (TDZ).
Penggunaan BAP dan TDZ secara bersamaan diduga akan mengaktifkan kedua sisi pengikatan CBP sehingga akan meningkatkan pembentukan tunas.
ZPT dari golongan sitokinin seperti benzyl amino purine (BAP) berperan penting dalam memacu proses pembelahan sel, khususnya didalam proses regenerasi tunas, menstimulasi pertumbuhan tunas lateral dan menghasilkan tunas ganda (Lestari 2011). Selain BAP, penggunaan TDZ juga sangat efektif dalam mendukung pembentukan dan penggandaan tunas in vitro (George et al. 2008). TDZ merupakan
senyawa organik yang banyak digunakan dalam perbanyakan in vitro karena aktivitasnya menyerupai sitokinin. TDZ dapat diberikan bersama-sama dengan ZPT lain, seperti sitokinin atau auksin, dan dapat pula sendiri, namun perlakuan bersama-sama dengan ZPT lain lebih baik hasilnya (Hutchinson et al. 2010).
Gambar 1. Proliferasi tunas in vitro pisang kepok Unti Sayang: (a). Bakal tunas umur 4 minggu, (b). Bakal tunas
umur 8 minggu, (c). Tunas umur 12 minggu
aaa bbb ccc

Perbanyakan In Vitro Pisang Kepok... Imelda et al.
40
Gambar 2. Tunas in vitro pisang kepok Unti Sayang dari berbagai perlakuan umur 6 minggu setelah kultur
Tabel 2. Pengaruh kombinasi BAP, TDZ dan adenin sulfat terhadap jumlah dan tinggi tunas in vitro pisang kepok Unti
Sayang
Media Jumlah tunas (cm) Tinggi tunas (cm)
4 minggu 8 minggu 12 minggu 4 minggu 8 minggu 12 minggu
B1 9,07hij
17,67hij
30,17fgh
2,00cde
3,00cd
2,41bcde
B1A 8,80ghij
17,00ghi
28,00fgh
2,17cde
2,60cd
2,58cde
B1T5 5,60def
21,00ij
32,33gh
0,57a
1,00a
1,00a
B1T5A 6,53efg
14,33fgh
24,00ef
0,67a
1,33ab
1,33ab
B2 8,57ghij
15,36fgh
25,30ef
1,95cde
2,00abc
2,50bcde
B2A 6,74efg
13,69efgh
25,18ef
2,15cde
2,65bcd
3,02def
B2T5 12,70k 25,51
j 33,95
gh 1,30
abc 1,40
ab 1,47
abc
B2T5A 10,98jk 35,44
k 36,81
h 0,97
ab 1,58
a 1,83
abcd
B3 6,80fgh
10,67cdef
18,27de
1,83bcde
2,16abc
2,50bcde
B3A 5,20cdef
9,33bcdef
14,50bcd
2,00cde
2,83cd
2,83de
B3T5 3,23abcd
9,00bcdef
15,17bcd
0,67a
1,17a
1,17a
B3T5A 3,13abc
11,67defg
18,67de
1,17abc
1,83abc
1,83abcd
B4 1,80a
4,67ab
9,00ab
3,00fg
3,40de
3,50ef
B4A 4,27bcde
11,33def
18,00cde
3,60g
4,17e
4,17f
B4T5 4,20bcde
5,67abc
8,50ab
0,50a
1,00a
1,00a
B4T5A 5,73ef
7,00abcd
10,17abc
0,50a
1,00a
1,00a
B5 10,88ijk
17,52ghi
25,45efg
1,98bcd
2,02abc
2,40bcde
B5A 7,62fghi
11,76cdef
24,80ef
2,00cde
2,78cd
3,10ef
B5T5 1,13a
2,07a
3,33a
0,50a
1,00a
1,00a
B5T5A 6,63efg
8,67bcde
12,67bcd
0,50a
1,00a
1,00a
Kontrol 2,60ab
2,68a 3,00
a 2,50
ef 3,00
cd 3,60
ef
Keterangan: Perbedaan huruf menunjukkan berbeda nyata berdasarkan DMRT pada taraf 5%
B1
B2
B3
B4
B5
K
B1T5A
B2T5A
B3T5A
B4T5A
B5T5A
B1T5
B2T5
B3T5
B4T5
B5T5
B1A
B2A
B3A
B4A
B5A

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
41
Penggunaan BAP pada penelitian kultur jaringan pisang kepok telah banyak dilakukan. Pada pisang kultivar kepok merah, multiplikasi tunas tertinggi diperoleh pada media MS yang ditambahkan IAA 0,5 mg/L dan BAP 5 mg/L. Pada media tersebut dihasilkan 6-17 tunas per eksplan dalam waktu 8 hari setelah tanam (Fitramala et al. 2016). Pada pisang kepok Amorang, penggunaan BAP pada konsentrasi 1 mg/L dalam ¼ MS merupakan media terbaik untuk multiplikasi tunas (Supriati 2010). Subkultur berulang pisang kepok var. Unti Sayang pada media MS yang ditambahkan 2 mg/L BAP dan 0,8 mg/L TDZ tidak mampu meningkatkan multiplikasi tunas (Semarayani 2012).
Pemberian adenin sulfat 20 mg/L pada media yang mengandung BAP konsentrasi rendah (1-2 mg/L) dengan atau tanpa TDZ secara statistik tidak berpengaruh nyata terhadap peningkatan jumlah tunas in vitro,
namun nilai rata-rata yang diperoleh lebih tinggi dibandingkan tanpa penambahan adenin sulfat. Adenin termasuk sitokinin alami. Adenin berperan sebagai prekursor pada biosintesis sitokinin. Adenin sering kali menunjukkan pengaruh yang serupa dengan sitokinin yaitu menginduksi pembentukan tunas (Van Staden et al. 2008). Pada beberapa tanaman, penambahan adenin sulfat pada media yang mengandung BAP atau kinetin mampu menghasilkan banyak tunas aksilar, seperti pada tanaman buncis (Arias et al. 2010), Fuchsia hybrida (Wroblewska 2012) dan Stevia rebaudiana
Bertoni (Khan et al. 2014). Respon terbaik pertambahan tinggi
tunas pisang Unti Sayang yaitu pada media B4A (MS+4 mg/L BAP+20 mg/L adenin sulfat). Penambahan adenin sulfat pada media yang mengandung BAP saja menunjukkan pertumbuhan tunas yang lebih tinggi dibandingkan tunas yang ditanam pada media tanpa penambahan adenin sulfat, walaupun secara statistik tidak berbeda nyata. Demikian pula penambahan adenin sulfat pada media yang mengandung BAP dan TDZ menunjukkan tunas yang lebih tinggi dibandingkan tunas yang ditanam pada media tanpa penambahan adenin sulfat. Penambahan TDZ pada media cenderung mengakibatkan terbentuknya banyak tunas namun menghambat pertumbuhan atau pertambahan tinggi tunas
sehingga dihasilkan tunas kerdil atau pendek (Tabel 2, Gambar 2). Untuk menghindari respon seperti itu, maka disarankan perbanyakan tunas in vitro pisang Unti Sayang pada media MS+0,5 mg/L TDZ+2 mg/L BAP tidak lebih dari 8 minggu. Pengakaran tunas in vitro
Pengakaran tunas dapat dengan mudah dilakukan dengan memindahkan tunas yang sudah cukup besar (umur 8 minggu setelah tanam) dari media perlakuan ke media MS tanpa ZPT (Gambar 3B). Akar juga sudah terbentuk pada beberapa perlakuan seperti B2 dan B2A namun jumlahnya terbatas. Oleh sebab itu pengakaran tunas sebaiknya dilakukan pada media kontrol tanpa ZPT. Penambahan auksin seperti IBA atau NAA ke dalam media pengakaran nampaknya tidak diperlukan karena akar yang terbentuk pada media kontrol sudah cukup banyak.
Aklimatisasi planlet
Perbanyakan secara in vitro
dinyatakan berhasil apabila planlet mampu melewati masa aklimatisasi dan dihasilkan bibit yang siap tanam. Aklimatisasi planlet pisang Unti Sayang tidak menemui banyak kendala. Aklimatisasi telah dilakukan terhadap 666 planlet, 617 planlet hidup dan hanya 49 planlet yang mati. Planlet yang berhasil hidup mencapai 92,75%. Penggunaan media untuk aklimatisasi yaitu tanah, kompos dan sekam bakar (1:1:1) serta tahapan aklimatisasi yang dilakukan menunjukkan hasil yang optimal. Seperti pada aklimatisasi pisang Raja Nangka, Kepok dan Mas yang menunjukkan tingkat keberhasilan aklimatisasi mencapai 90% dengan menggunakan media campuran tanah, pasir dan kompos dengan perbandingan 1:1:1 (Avivi et al. 2013). Proses aklimatisasi dapat dilihat pada Gambar 4.
KESIMPULAN
Proliferasi tunas in vitro pisang kepok
var. Unti Sayang menunjukkan hasil terbaik pada media B2T5A (2 mg/L BAP, 0,5 mg/L TDZ dan 20 mg/L adenin sulfat) yang menghasilkan 35,44 tunas pada umur 8 minggu. Berdasarkan penelitian ini, maka untuk metode perbanyakan tunas in vitro

Perbanyakan In Vitro Pisang Kepok... Imelda et al.
42
yang optimal adalah proliferasi tunas in vitro
pada media B2T5A selama tidak lebih dari 8 minggu kemudian dilanjutkan ke media tanpa ZPT untuk pertambahan tinggi dan pengakarannya.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Sdr. Mulyana untuk pemeliharaan kultur serta Sdr. Nana Burhana untuk pemeliharaan tanaman di rumah kaca dan kebun. Penelitian ini didanai oleh Proyek Kompetitif LIPI, Sub-Program Keanekaragaman Sumber Daya Hayati.
DAFTAR PUSTAKA
Arias AMG, Valverde JM, Fonseca PR, Melara MV (2010) In vitro plant
regeneration system for common bean (Phaseolus vulgaris): Effect of N6-
benzylaminopurine and adenine sulphate. Electron J Biotechnol 13 (Issue 1). doi: 10.2225-vol13-issue1-fulltext-7
Avivi S, Soedarmo SH, Prasetyo PA (2013) Multiplikasi tunas dan aklimatisasi tiga varietas pisang: Raja Nangka, Kepok, dan Mas. J Hort Indones 4: 83-89
Eden-Green SJ (1994) Banana blood disease. International Network for the Improvement of Banana and Plantain (INIBAP). Musa Disease Fact Sheet No.3. Montpellier, France
Fitramala E, Khaerunisa E, Djuita NR, Sunarso H, Ratnadewi D (2016) Kultur in vitro pisang (Musa paradisiaca L.) cv. Kepok Merah untuk mikropropagasi cepat. Menara Perkebunan 84: 69-75. doi:10.22302/iribb.jur.mp.v84i2.221
George EF, Hall MA, De Klerk G-J (2008) Plant Propagation by Tissue Culture. Volume 1: The Background. 3rd Edition. Springer-Verlag, New York
Guo B, Abbasi BH, Zeb A, Xu LL, Wei YH (2011) Thidiazuron: A multi-dimentional plant growth regulator. Afr J Biotechnol 10: 8984-9000. doi: 10.5897/AJB11.636
Hermanto C, Emilda ED (2013) Bunch management of banana to control
Gambar 3. Planlet pisang kepok Unti Sayang: (a). Tunas yang baru membentuk akar, (b). Planlet yang sudah
berakar banyak, (c). Tunas siap untuk aklimatisasikan
Gambar 4. Aklimatisasi planlet pisang kepok Unti Sayang: (a). Planlet dalam sungkup plastik, (b). Bibit setelah
sungkupnya dibuka (umur 2 minggu), (c). Bibit umur 4 minggu
a b c
aa bbb ccc

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
43
blood disease. Aust Plant Pathol 42: 653-658
Hutchinson MJ, Onamu R, Kipkosgei L, Obukosia SD (2010) Effect of thidiazuron, NAA and BAP on in vitro propagation of Alstroemeria aurantiaca
cv ‘ROSITA’ from shoot tip explants. J Agric Sci Technol 12: 60-69
Khan MK, Misra P, Sharma T, Shukla PK, Ramteke PW (2014) Effect of adenine sulphate on in vitro mass propagation of Stevia rebaudiana Bertoni. J Med
Plant Res 8: 543-549. doi: 10.5897/JMPR2013.5217
Lestari EG (2011) Peranan zat pengatur tumbuh dalam perbanyakan tanaman melalui kultur jaringan. J AgroBiogen 7: 63-68. doi: 10.21082/jbio.v7n1.2011.p63-68
Lestari EG, Suhartanto MR, Kurniawati A, Rahayu S (2013) Inisiasi tunas ganda tanaman manggis Malinau melalui kultur in vitro untuk perbanyakan
klonal. J Agron Indones 41: 40-46 Maramis RTD (2014) Insidensi penyakit
darah pisang di Kabupaten Minahasa Utara. J Ilmiah Sains 14: 35-45
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol Plant 15: 473-497. doi: 10.1111/j.1399-3054.1962.tb08052.x
Poerba YS, Imelda M, Martanti D (2012) Analisis kestabilan genetik pisang kepok ‘Unti Sayang’ hasil
mikropropagasi dengan marka RAPD dan ISSR. Berita Biologi. 11: 275-282. doi: 10.14203/beritabiologi.v11i2.497
Semarayani CIM (2012) Subkultur berulang tunas in vitro pisang kepok Unti Sayang pada beberapa komposisi media. Skripsi. Fakultas Pertanian. Institut Pertanian Bogor
Suhartanto MR, Sobir, Harti H, Nasution MA, Nurbani (2012) Pengembangan pisang kepok unggul sebagai penopang ketahanan pangan nasional. Hal 444-448. Prosiding Simposium dan Seminar Bersama PERAGI-PERHORTI-PERIPI-HIGI. Mendukung Kedaulatan Pangan dan Energi yang Berkelanjutan. 1-2 Mei 2012, Bogor
Supriati Y (2010) Efisiensi mikropropagasi pisang kepok Amorang melalui modifikasi formula media dan temperatur. J AgroBiogen 6: 91-100. doi: 10.21082/jbio.v6n2.2010.p91-100
Van Staden J, Zazimalova E, George EF (2008) Plant growth regulators II: Cytokinins, their analogues and antagonists. Pp 205-226. In: Plant
Propagation by Tissue Culture. Volume 1: The Background. 3rd Edition. George EF, Hall MA, De Klerk G-J (Eds). Springer, Dordrecht
Wroblewska K (2012) The influence of adenine and benzyladenine on rooting and development of Fuchsia hybrida
cuttings. Acta Agrobotanica 65: 101-108. doi: 10.5586/aa.2012.026

Optimasi Proses Untuk Ekspresi Gen Endoglukanase... Victor dan Moeis
44
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
OPTIMASI PROSES UNTUK EKSPRESI GEN ENDOGLUKANASE DARI
Bacillus sp. RP1 OLEH Escherichia coli BL21 (DE3)/egc
Process Optimization for Endoglucanase Gene Expression Derived from Bacillus sp. RP1 by Escherichia coli BL21 (DE3)/egc
Hans Victor, Maelita Ramdani Moeis*
Sekolah Ilmu dan Teknologi Hayati ITB. Jl. Ganesha No.10, Bandung 40123, Indonesia *Email: [email protected]
ABSTRACT Cellulases are one of the most used enzymes in industrial processes. In an effort to increase production, industries have developed strategies such as isolating new cellulase producing strains, genetic engineering and process optimization since the last 50 years. One endoglucanase producing strain, Bacillus sp. RP1 was isolated from hot springs. The ribosome binding site and coding sequence of the endoglucanase gene (egc) from Bacillus sp. RP1 was cloned into pGEM-T Easy. The recombinant plasmid was used to transform E. coli BL21 (DE3). Cloning was followed by process optimization. Medium composition was selected using Plackett-Burman design. The medium components tested were rice hull, molasses, ammonium chloride, urea and fishmeal. Rice hull and molasses were found to be the factors most influencing enzyme activity and dry cell weight, respectively. The next step involved Box-Behnken method and response surface methodology to optimize the responses against molasses concentration, rice hull concentration and fermentation time. The concentration intervals used to test were 1%, 5.5% and 10% while the fermentation time used were 24, 36 and 48 hours. The conditions which optimized both enzyme activity and dry cell weight were 7.45% molasses, 6.45% rice hull and 39.52 hours of fermentation. Keywords: Bacillus sp. RP1, E. coli BL21 (DE3), egc, Endoglucanase, optimization ABSTRAK Selulase adalah salah satu enzim yang banyak dimanfaatkan dalam berbagai industri. Sebagai upaya untuk memenuhi kebutuhan, 50 tahun terakhir dikembangkan beberapa strategi untuk meningkatkan produksi selulase yang mencakup rekayasa genetika dan optimasi proses. Karena itu, dilakukan kloning gen egc dan RBS yang berasal dari Bacillus sp. RP1 yang diisolasi dari sumber air panas ke dalam vektor pGEM-T Easy. E. coli BL21 (DE3) ditransformasikan dengan vektor yang mengandung gen egc tersebut. Setelah kloning, optimasi proses berupa desain medium turut dilakukan untuk mengoptimalkan ekspresi gen egc. Desain medium diawali dengan seleksi komposisi medium menggunakan metode Plackett-Burman. Komponen medium yang diuji adalah kulit beras, molase, amonium klorida, urea dan tepung ikan. Kulit beras dan molase diperoleh sebagai bahan yang paling berpengaruh terhadap aktivitas enzim dan berat kering sel. Tahap selanjutnya melibatkan metode statistik Box-Behnken dan metodologi respons permukaan yang bertujuan mengoptimalkan respons aktivitas enzim dan berat kering sel terhadap konsentrasi molase, konsentrasi kulit beras dan lama fermentasi. Konsentrasi yang diuji adalah 1%, 5,5% dan 10%, sedangkan lama fermentasi yang diuji adalah 24, 36 dan 48 jam. Konsentrasi optimal molase adalah 7,45% dan konsentrasi optimal kulit beras adalah 6,45% dengan lama fermentasi optimal 39,52 jam.
Kata Kunci: Bacillus sp. RP1, E. coli BL21 (DE3), egc, Endoglukanase, optimasi
Received: 06 April 2018 Accepted: 06 May 2018 Published: 11 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
45
PENDAHULUAN
Selulase adalah keluarga enzim yang
dapat mendegradasi selulosa. Berdasarkan aktivitas katalitiknya, selulosa terbagi menjadi beberapa keluarga, yaitu endo-1,4-β-D-glukan glukanohidrolase (EC 3.2.1.4), (1,4)-β-D-glukan selobiohidrolase (EC 3.2.1.91), 1,4- β-D-oligoglukan selobiohidrolase (EC 3.2.1.74), β-glukosidase (EC 3.2.1.21) dan selobiosa fosforilase (EC 2.4.1.20) (Sadhu dan Maiti 2013).
Aktivitas tersebut dimanfaatkan dalam berbagai industri. Beberapa industri yang memanfaatkan selulase adalah industri agrikultur, industri biokonversi, industri deterjen, industri kertas, industri pangan, industri fermentasi, industri tekstil, dan lain-lain (Kuhad et al. 2011). Tiap industri menghendaki karakteristik selulase yang berbeda, tergantung pada proses yang berlangsung dalam industri tersebut (Kuhad et al. 2011).
Pada umumnya selulase yang berasal dari fungi tidak bersifat termostabil sehingga sulit diaplikasikan dalam industri (Acharya dan Chaudhary 2012). Maka isolasi selulase dilakukan pada mikroba dari lingkungan yang beragam, untuk memperoleh selulase dengan karakteristik yang berpotensi untuk digunakan dalam industri (Maki et al. 2009). Muñoz et al. (2014) mengisolasi sembilan strain bakteri penghasil selulase dari beberapa spesies kerang. Bacillus vallismortis RG-07 diisolasi dari tanah dan
diketahui menghasilkan selulase termostabil dan alkalifilik (Gaur dan Tiwari 2015). Sementara itu, Puspitasari dan Moeis (2008) menganalisis selulase dari spesies Bacillus
sp. RP1 yang diisolasi dari sumber air panas. Bacillus sp. RP1 adalah sumber gen egc yang digunakan dalam optimasi proses
dalam penelitian ini. Selain isolasi mikroba penghasil
selulase, terdapat beberapa strategi lain untuk meningkatkan produksi selulase. Salah satunya adalah meningkatkan produksi melalui optimasi medium atau optimasi proses fermentasi (Parekh et al. 2000). Dalam optimasi medium produksi, kondisi fermentasi (pH, suhu, agitasi, dll.) dan komponen medium (karbon, nitrogen, dll.) diidentifikasi dan dioptimasi (Kuhad et al. 2016; Singh et al. 2017).
BAHAN DAN METODE
Bakteri dan primer
Bakteri yang digunakan sebagai sumber gen egc adalah Bacillus sp. RP1
yang diisolasi dari sumber air panas (Puspitasari dan Moeis 2008) diperoleh dari koleksi Laboratorium Genetika SITH, Institut Teknologi Bandung. Strain E. coli BL21
(DE3) digunakan sebagai sel inang untuk mengekspresikan gen egc menggunakan
promoter dari plasmid dan rbs alami dari egc.
Sekuens primer yang digunakan untuk mengamplifikasi gen egc dan RBSnya adalah primer maju egcF 5’-AGGAGGAAA
AGATCAGATATGAAA-3’ (Lee et al. 2012) dengan modifikasi berupa pengurangan 9 pasang basa pada ujung 3’ dan primer mundur egcR 5’-TCAGTATTTCATCCAC AACGCA-3’ (Moeis et al. 2014). Kloning dan transformasi
Bacillus sp. RP1 ditumbuhkan pada
media LB agar dan diinkubasi pada 37ᴼC
selama 18 jam. Satu koloni Bacillus sp. RP1
yang tumbuh digunakan sebagai sumber DNA dalam proses PCR koloni. Proses PCR koloni dilakukan dengan parameter: 94ºC, 5 menit; 22 siklus 94ºC selama 30 detik, 58,4ºC selama 30 detik, dan 72ºC selama 2 menit; 72ºC selama 7 menit. Produk PCR berukuran 1.568 pasang basa (pb) selanjutnya dipurifikasi menggunakan Gel/PCR DNA Fragments Extraction Kit DF100/DF300 (GeneAid). Gen yang diperoleh diligasikan ke dalam vektor pGEM-T® Easy (Promega) dan ditransformasikan dengan metode kejut panas ke dalam E. coli
BL21 (DE3). Bakteri rekombinan diseleksi dengan
pengujian kualitatif aktivitas endoglukanase yang dilakukan dengan metode pewarnaan agar CMC dengan Gram’s iodine (Gohel et al. 2014). Medium LB/ampisilin/CMC agar dituang ke dalam cawan Petri yang telah dibagi menjadi beberapa daerah. Setiap daerah digoreskan kultur dari satu koloni E. coli BL21 (DE3)/egc hasil transformasi yang
dipilih secara acak dengan jarum ose. Selanjutnya medium diinkubasi pada 37ºC selama 48 jam. Selain diinokulasikan pada medium LB/ampisilin/CMC agar, masing-masing koloni yang dicuplik juga diinokulasikan ke dalam tabung sentrifuga

Optimasi Proses Untuk Ekspresi Gen Endoglukanase... Victor dan Moeis
46
15 mL berisi 5 mL medium LB cair dengan ampisilin 100 μg/mL. Kultur cair selanjutnya diinkubasi pada 37ºC selama 18 jam.
Setelah inkubasi, Gram’s iodine dituangkan pada medium LB/ampisilin/CMC agar hingga terendam seluruhnya. Cawan Petri kemudian dibiarkan pada suhu ruang selama 10 menit. Setelah perendaman, dilakukan pembilasan dengan akuades, dan koloni bakteri yang tumbuh turut dibuang dengan pembilasan.
Kultur cair dari E. coli BL21 (DE3)
transforman yang menunjukkan zona bening disimpan dan digunakan untuk tahap selanjutnya, yaitu konfirmasi dengan isolasi plasmid dan restriksi dengan enzim EcoRI.
Isolasi plasmid dilakukan dengan ATP™ Plasmid Mini Kit (ATP Biotech Inc.). Selanjutnya reaksi restriksi yang terdiri dari 2,0 µL 10x FastDigest Buffer, 2,0 µL DNA plasmid, 1,0 µL EcoRI FastDigest, dan 15,0
µL air bebas nuklease disiapkan. Seluruh komponen dicampurkan dalam tabung mikro 0,2 mL dan diinkubasi selama 15 menit pada 37ºC dan kemudian diinaktivasi pada 80ºC selama 5 menit. Hasil reaksi restriksi dielektroforesis pada gel agarosa 1% selama 25 menit pada tegangan 100 V. Satu koloni yang menghasilkan hasil positif pada uji aktivitas dan pada konfirmasi menggunakan enzim restriksi digunakan untuk tahap selanjutnya dan disebut E. coli BL21 (DE3)/egc.
Seleksi komposisi medium
Seleksi komposisi medium dilakukan dengan desain eksperimen Plackett-Burman dengan respons berupa aktivitas enzim. Sebanyak 20 kombinasi medium yang mengandung satu atau lebih dari kulit beras, molase, tepung ikan, amonium klorida, dan urea, dirancang dengan bantuan perangkat lunak Minitab 17. Medium dibuat sesuai dengan rancangan dari perangkat lunak untuk volume sebesar 10 mL di dalam tabung sentrifuga 50 mL. Selanjutnya medium disterilisasi dengan autoklaf pada 121ºC selama 15 menit. Medium yang telah disterilisasi disuplementasikan dengan ampisilin sebanyak 100 μg/mL dan diinokulasikan dengan 50 µL E. coli BL21 (DE3)/egc dengan OD600 prekultur sebesar 0,5. Selanjutnya medium yang telah diinokulasikan diinkubasi pada 37ºC dengan agitasi 250 rpm selama 24 jam.
Setelah inkubasi, sel dipanen dari 5 mL medium dengan sentrifugasi pada 500g
selama 5 menit untuk memisahkan komponen padat dari medium. Supernatan kemudian dipisahkan ke dalam tabung sentrifuga kosong dan disentrifugasi pada 14.000g selama 2 menit untuk mengendapkan sel. Pelet sel yang diperoleh dibilas sebanyak tiga kali dengan 500 µL buffer lisis. Selanjutnya sel diresuspensi dalam 500 µL buffer lisis (50 mM Tris–Cl, 0,5 mM EDTA) dan dipecahkan dengan bantuan sonikasi. Sonikasi dilakukan menggunakan Ultrasonic Homogenizer FS-150N, 100% output dengan interval 15 detik on, 10 detik off selama 10 menit.
Sel yang telah dilisis melalui proses sonikasi disentrifugasi untuk memisahkan debris sel pada 14.000g selama 2 menit.
Supernatan yang diperoleh digunakan untuk uji aktivitas enzim dengan metode alkali ferisianida. Reaksi untuk pengujian aktivitas enzim dibuat dalam tabung 1,5 mL seperti pada Tabel 1.
Jumlah ekuivalen gula pereduksi yang dilepaskan oleh reaksi antara CMC dengan endoglukanase diperoleh dengan membandingkan absorbansi pada 420 nm dengan kurva standar glukosa. Selanjutnya jumlah ekuivalen gula pereduksi yang diperoleh pada larutan reaksi dikurangi dengan jumlah ekuivalen gula pereduksi yang diperoleh pada blanko substrat dan blanko enzim yang bersesuaian untuk menghilangkan efek gula pereduksi yang mungkin terdapat pada larutan enzim atau larutan substrat. Aktivitas enzim dalam satuan unit didefinisikan sebagai jumlah enzim yang melepaskan satu µmol gula
Tabel 1. Komposisi reaksi uji aktivitas enzim dengan metode alkali ferisianida
Jenis reaksi Buffer (mL)
Substrat (mL)
Enzim (mL)
Blanko total 0,20 0,00 0,00
Blanko enzim* 0,15 0,00 0,05
Blanko substrat 0,05 0,15 0,00
Larutan reaksi* 0,00 0,15 0,05
*Keterangan: dibuat sebanyak 20 kali, satu kali untuk tiap kombinasi medium. Buffer adalah natrium asetat 50 mM pH 5,0. Substrat adalah 1 b/v% CMC dalam buffer natrium asetat 50 mM pH 5,0. Enzim adalah supernatan yang diperoleh pada tahap lisis sel dan sentrifugasi sebelumnya.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
47
pereduksi (ekuivalen glukosa) per jam pada kondisi assay.
Seleksi komposisi medium juga dilakukan dengan respons berupa berat kering sel. Dengan bantuan perangkat lunak Minitab 17, 20 kombinasi medium dibuat dengan bahan berupa kombinasi dari kulit beras, molase, tepung ikan, amonium klorida dan urea. Medium dibuat dengan cara yang sama dengan tahap sebelumnya. Medium yang telah diinkubasi kemudian disentrifugasi dengan kecepatan 500g
selama 5 menit dan supernatan yang diperoleh dipisahkan ke dalam tabung sentrifuga kosong. Supernatan tersebut kembali disentrifugasi dengan kecepatan 14.000g selama 2 menit. Supernatan yang diperoleh dibuang, sedangkan pelet dalam tabung 1,5 mL dikeringkan. Pelet yang telah kering kemudian ditimbang. Optimasi konsentrasi medium
Dua bahan dengan pengaruh paling tinggi terhadap aktivitas enzim dan berat kering dipilih menjadi dua faktor pada tahap optimasi konsentrasi medium. Faktor ketiga yang dipilih adalah lama inkubasi. Faktor-faktor tersebut dikombinasikan dalam desain eksperimen Box-Behnken dengan bantuan Minitab 17. Medium dibuat sesuai dengan hasil pengacakan dari Minitab 17 untuk volume medium sebesar 15 mL. Medium yang telah dibuat disterilisasi dengan autoklaf pada 121ºC selama 15 menit. Lama inkubasi tiap medium disesuaikan dengan hasil pengacakan dari Minitab 17. Setelah inkubasi, tiap medium diambil sebanyak 5 mL dan disentrifugasi pada 500g selama 5
menit untuk memisahkan padatan dalam medium. Supernatan yang diperoleh disentrifugasi pada 14.000g selama 2 menit untuk memperoleh sel. Sel yang diperoleh dibilas dan diresuspensi dalam 500 µL buffer lisis. Suspensi sel dipecahkan dengan metode sonikasi pada kondisi 100% output dengan interval 15 detik on, 10 detik off
selama 10 menit. Sel yang telah dipecahkan
disentrifugasi pada 14.000g selama 2 menit.
Supernatan yang diperoleh dipisahkan dari pelet untuk memperoleh ekstrak kasar enzim. Ekstrak kasar enzim digunakan dalam pengujian aktivitas enzim dengan metode alkali ferisianida.
Medium dengan komposisi yang sama
kembali dibuat, diinokulasikan dan diinkubasikan dengan kondisi yang sama. Setelah inkubasi, tiap medium diambil sebanyak 5 mL dan disentrifugasi pada 500g selama 5 menit dan supernatan dipisahkan. Supernatan yang diperoleh disentrifugasi pada 14.000g selama 2 menit untuk memperoleh sel. Sel yang diperoleh dibilas dan diresuspensi dalam 500 µL buffer lisis. Kemudian sel dikeringkan dalam oven dan ditimbang setelah kering. HASIL DAN PEMBAHASAN
Kloning dan transformasi
Isolasi gen egc dilakukan dengan metode PCR koloni Bacillus sp. RP1 dengan primer egcF dan egcR. Hasil PCR gen egc
menghasilkan pita DNA dengan ukuran ~1,5 kb pada sumur 1 menandakan gen egc berhasil diisolasi dari Bacillus sp. RP1
melalui metode PCR koloni (Gambar 1). Pada elektroforesis hasil restriksi dari
plasmid rekombinan diperoleh satu pita DNA dengan ukuran ~1,5 kb, sesuai dengan ukuran gen egc yang diinsersikan dan satu
pita berukuran ~4,6 kb yang merupakan ukuran gabungan antara pGEM-T® Easy dan gen egc (Gambar 2). Berdasarkan hasil tersebut, gen egc berhasil diinsersikan
ke dalam pGEM-T® Easy.
Seleksi komposisi medium
Seleksi komposisi medium ditentukan berdasarkan efek masing-masing faktor terhadap respons berupa aktivitas enzim dan berat kering sel. Tabel 2 menunjukkan hasil pengujian aktivitas enzim dengan medium yang didesain menurut metode Plackett-Burman, sedangkan Tabel 3 menunjukkan hasil penimbangan berat kering sel dengan medium yang didesain menurut metode Plackett-Burman.
Gambar 3 menunjukkan efek masing-masing faktor terhadap aktivitas enzim dan berat kering sel. Berdasarkan Gambar 3a, dapat dilihat bahwa molase berpengaruh signifikan terhadap aktivitas enzim, sedangkan bahan-bahan lainnya tidak berpengaruh signifikan.
Pengaruh molase disebabkan oleh kandungan galaktosa, salah satu monomer dari rafinosa yang terdapat dalam molase. Keberadaan galaktosa dapat membuat penambahan IPTG tidak diperlukan untuk

Optimasi Proses Untuk Ekspresi Gen Endoglukanase... Victor dan Moeis
48
menginduksi sistem ekspresi pada E. coli BL21 (DE3). Hal ini disebabkan karena
galaktosa dapat menyebabkan auto induksi dari produksi protein pada E. coli BL21 dengan promoter lac (Xu et al. 2012).
Pada strain E. coli dengan mutasi gal-,
induksi oleh galaktosa ekspresi protein dapat mencapai 59-84% dari ekspresi protein yang diinduksi oleh IPTG. Mutasi tersebut menyebabkan E. coli tidak dapat memetabolisme galaktosa sehingga konsentrasi galaktosa intraseluler menjadi
tinggi dan dapat menginduksi promoter lac (Mattanovich et al. 1998). E. coli BL21 (DE3) adalah salah satu strain dengan fenotipe gal- (Studier et al. 2009). Karena itu, ekspresi protein oleh E. coli BL21 (DE3)
dapat terinduksi oleh keberadaan galaktosa. Gambar 3b menunjukkan efek masing-
masing faktor terhadap berat kering sel. Jenis bahan yang berpengaruh signifikan terhadap berat kering sel adalah kulit beras. Hasil ini serupa dengan hasil yang diperoleh Lee et al. (2012) menunjukkan bahwa kulit
Gambar 1. Hasil elektroforesis produk PCR koloni Bacillus sp. RP1 dengan primer egcF dan egcR. 1-4) PCR dengan template koloni Bacillus sp. RP1, 5) GeneRuler 1 kb DNA Ladder
Gambar 2. Hasil restriksi plasmid pGEM-T
® Easy hasil
kloning. A) pGEM-T ®
Easy, B) gen egc
Gambar 3. Grafik Pareto efek komposisi medium terhadap (a). aktivitas enzim endoglukanase dan (b). berat kering sel. (A)
Molase, (B) kulit beras, (C) amonium klorida, (D) urea, (E) tepung ikan. Garis merah adalah garis referensi. Faktor yang nilai efek terstandarisasinya melebihi nilai tersebut adalah faktor yang berpengaruh signifikan

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
49
Tabel 2. Hasil pengujian aktivitas enzim dalam tiap medium yang didesain dengan metode Plackett-Burman .
No. Molase
(% v/v)
Kulit beras
(% b/v)
Amonium
klorida (% b/v)
Urea
(% b/v)
Tepung ikan
(% b/v)
Aktivitas enzim
(U/mL)
1 0 5 1 1 1 0,0466
2 5 5 0 1 1 0,0329
3 5 5 1 0 0 0,1700
4 5 0 1 1 1 0,0446
5 0 5 0 1 1 0,0405
6 5 5 0 0 1 0,0510
7 0 5 1 0 1 0,0911
8 5 5 1 1 0 0,1475
9 5 5 0 0 0 0,1836
10 0 0 1 1 0 0,0467
11 0 0 0 1 0 0,0464
12 5 0 0 0 0 0,0404
13 5 0 1 0 1 0,0523
14 0 0 0 0 0 0,0000
15 0 0 1 0 1 0,0444
16 0 5 0 1 0 0,0117
17 0 5 1 0 0 0,0516
18 5 0 0 1 1 0,1723
19 5 0 1 1 0 0,1490
20 0 0 0 0 1 0,0962
Tabel 3. Hasil penimbangan berat kering sel dalam tiap medium yang didesain dengan metode Plackett-Burman
No. Molase
(% v/v)
Kulit beras
(% b/v)
Amonium
klorida (% b/v)
Urea
(% b/v)
Tepung ikan
(% b/v)
Berat kering
(mg)
1 5 0 0 1 1 3,8
2 5 0 1 0 0 2,7
3 0 0 0 0 1 2,6
4 5 5 1 1 0 5,8
5 5 0 1 0 1 5,1
6 5 0 0 1 0 0,4
7 5 5 1 0 1 7,5
8 0 5 0 0 0 2,5
9 5 0 1 1 1 2,4
10 5 5 0 0 1 6,7
11 0 5 1 1 0 8,2
12 0 0 1 1 0 0,7
13 0 0 0 0 0 0
14 0 0 1 0 0 0
15 0 5 1 1 1 4,0
16 0 5 1 0 1 5,5
17 5 5 0 0 0 3,1
18 5 5 0 1 0 11,3
19 0 5 0 1 1 3,8
20 0 0 0 1 1 3,7
beras adalah sumber karbon yang paling

Optimasi Proses Untuk Ekspresi Gen Endoglukanase... Victor dan Moeis
50
beras adalah sumber karbon yang paling berpengaruh terhadap pertumbuhan dan produksi CMCase oleh E. coli JM109/LBH-10. Optimasi fermentasi
Optimasi konsentrasi medium dan lama fermentasi dilakukan terhadap respons aktivitas enzim dan berat kering sel. Gambar 4 menunjukkan model respons permukaan dari aktiivtas enzim. Sedangkan Gambar 5 menunjukkan model respons permukaan dari berat kering sel.
Model respons permukaan dengan respons aktivitas enzim (y1) dan faktor-faktor konsentrasi molase (x1), konsentrasi kulit beras (x2) dan lama fermentasi (x3) dapat dituliskan dalam Persamaan 1 berikut:
Berdasarkan persamaan tersebut,
aktivitas enzim berada pada titik optimal saat konsentrasi molase dan kulit beras sebanyak 6% dan lama fermentasi 35,15 jam.
Gambar 4 menunjukkan respons
aktivitas enzim memuncak saat nilai ketiga faktor mendekati nilai tengah yang diuji dan menurun ketika mendekati nilai minimal atau maksimal. Bentuk model menyerupai puncak yang hampir simetris menunjukkan interaksi ketiga faktor terhadap aktivitas enzim minimal. Model respons permukaan berat kering sel (y2) dapat dituliskan dalam Persamaan 2 berikut:
Berdasarkan Persamaan 2, titik optimal
berat kering sel terletak saat konsentrasi molase mencapai 10% dan konsentrasi kulit beras sebesar 7,3%. Lama fermentasi yang memberikan berat sel paling tinggi adalah 48 jam. Pada Gambar 5a, ketika konsentrasi kulit beras rendah, peningkatan konsentrasi molase memberi peningkatan berat kering yang lebih signifikan dibandingkan ketika konsentrasi kulit beras tinggi. Pada kondisi konsentrasi kulit beras rendah, molase juga digunakan sebagai sumber karbon,
(%)
Gambar 4. (a). Grafik respons permukaan respons aktivitas enzim terhadap konsentrasi molase dan kulit beras,
(b). konsentrasi molase dan lama fermentasi, (c). konsentrasi kulit beras dan lama fermentasi. Faktor yang tidak tercantum dalam grafik berada pada titik optimalnya
Gambar 5. (a). Grafik respons permukaan respons berat kering sel terhadap konsentrasi molase dan kulit beras,
(b). konsentrasi molase dan lama fermentasi, (c). konsentrasi kulit beras dan lama fermentasi. Faktor yang tidak tercantum dalam grafik berada pada titik optimalnya
(%) (%)
Fermentasi
(%)
a) b) c)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
51
sedangkan pada kondisi konsentrasi kulit beras tinggi, karbon dari kulit beras cukup sehingga molase tidak berpengaruh banyak terhadap pertumbuhan.
Titik optimal dari kedua respons jika disatukan dalam satu model adalah pada konsentrasi molase 7,45%, konsentrasi kulit beras 6,45% dan lama fermentasi 39,52 jam. Jika dibandingkan kedua respons terhadap lama fermentasi, dapat dilihat bahwa aktivitas enzim tidak berbanding lurus dengan berat kering sel. Aktivitas enzim telah mengalami penurunan meskipun berat kering sel masih meningkat. Hasil ini menunjukkan bahwa enzim diproduksi pada awal fase pertumbuhan. Kim et al. (2016) menunjukkan bahwa E. coli JM109 mengekspresikan CMCase pada fase pertumbuhan dan fase stasioner.
Penyebab perbedaan adalah komposisi medium. Pada penelitian ini, rasio C:N dalam medium yang tinggi mengakibatkan sangat mungkin dalam rentang konsentrasi medium yang diuji, E. coli bertumbuh dalam kondisi nitrogen yang terbatas. Dalam kondisi nitrogen yang terbatas, pertumbuhan E. coli dan keluarga Enterobacteriaceae akan berhenti dan berfokus pada ekspresi gen yang berfungsi untuk memperoleh nitrogen semaksimal mungkin (Figueira et al. 2015). Selain itu, proteolisis juga terjadi saat terjadi kekurangan nitrogen (Smith 1995). Proteolisis yang terjadi mungkin mendegradasi protein rekombinan yang diekspresikan, sehingga aktivitas endoglukanase menurun meskipun pertumbuhan masih terjadi. Aktivitas enzim masih dapat lebih dioptimalkan jika ada sumber nitrogen yang ditambahkan.
KESIMPULAN
Berdasarkan hasil penelitian, komposisi
medium yang berpengaruh signifikan pada aktivitas endoglukanase dan berat kering sel dari E. coli BL21 (DE3)/egc adalah dua sumber karbon berupa molase dan kulit beras. Optimasi proses ekspresi endoglukan-ase oleh E. coli BL21 (DE3)/egc dipengaruhi
oleh komposisi medium berupa molase dan kulit beras dan lama fermentasi. Konsentrasi optimal molase adalah 7,45% dan konsentrasi optimal kulit beras adalah 6,45% dengan lama fermentasi optimal 39,52 jam.
DAFTAR PUSTAKA
Acharya S, Chaudhary A (2012)
Optimization of fermentation conditions for cellulases production by Bacillus licheniformis MVS1 and Bacillus sp.
MVS3 isolated from Indian hot spring. Braz Arch Biol Technol 55: 497-503. doi: 10.1590/S1516-89132012000400003
Figueira R, Brown DR, Ferreira D, Eldridge
MG, Burchell L, Pan Z, Helaine S,
Wignesweraraj S (2015) Adaptation to
sustained nitrogen starvation by Escherichia coli requires the
eukaryote-like serine/threonine kinase
YeaG. Sci Rep 5: 1-14. doi:
10.1038/srep17524 Gaur R, Tiwari S (2015) Isolation,
production, purification and
characterization of an organic-solvent-
thermostable alkalophilic cellulase from Bacillus vallismortis RG-07. BMC
Biotechnol 15:19. doi:
10.1186/s12896-015-0129-9
Gohel HR, Contractor CN, Ghosh SK, Braganza VJ (2014) A comparative
study of various staining techniques for
determination of extra cellular cellulase
activity on Carboxy Methyl Cellulose (CMC) agar plates. Int J Curr Microbiol
App Sci 3: 261-266
Kim MH, Kang DU, Lee JW (2016)
Construction of a recombinant Escherichia coli JM109/A-68 for
production of carboxymethylcellulase
and comparison of its production with
its wild type, Bacillus velezensis A-68 in a pilot-scale bioreactor. Biotechnol
Bioprocess Eng 21: 601-611. doi:
10.1007/s12257-016-0468-y
Kuhad RC, Deswal D, Sharma S, Bhattacharya A, Jain KK, Kaur A,
Pletshcke BI, Singh A, Karp M (2016)
Revisiting cellulase production and
redefining current strategies based on major challenges. Renew Sust Energ
Rev 55: 249-272. doi:
10.1016/j.rser.2015.10.132 Kuhad RC, Gupta R, Singh A (2011)
Microbial cellulases and their industrial applications. Enzyme Res 1-10. doi: 10.4061/2011/280696
Lee YJ, Kim HJ, Gao W, Chung CH, Lee JW

Optimasi Proses Untuk Ekspresi Gen Endoglukanase... Victor dan Moeis
52
(2012) Statistical optimization for production of carboxymethylcellulase of Bacillus amyloliquefaciens DL-3 by a recombinant Escherichia coli JM109/DL-3 from rice bran using response surface method. Biotechnol Bioproc Eng 17: 227-235. doi: 10.1007/s12257-011-0258-5
Maki M, Leung KT, Qin W (2009) The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int J Biol Sci 5: 500-516. doi: 10.7150/ijbs.5.500
Mattanovich D, Kramer W, Luttich C, Weik R, Bayer K, Katinger H (1998) Rational design of an improved induction scheme for recombinant Escherichia coli. Biotechnol Bioeng 58: 296-298. doi: 10.1002/(SICI)1097-0290(199804 20)58:2/3<296::AID-BIT26>3.0.CO;2-9
Moeis MR, Natalia D, Ningrum RW, Dwijayanti A (2014) Cloning and expression of endoglucanase gene from thermophilic bacteria Bacillus sp.
RP1. Microbiol Indones 8: 170-176. doi: 10.5454/mi.8.4.4
Muñoz C, Hidalgo C, Zapata M, Jeison D, Riquelme C, Rivas C (2014) Use of cellulolytic marine bacteria for enzymatic pretreatment in microalgal biogas production. Appl Environ Microbiol 80: 4199-4206. doi: 10.1128/AEM.00827-14
Parekh S, Vinci VA, Strobel RJ (2000). Improvement of microbial strains and fermentation process. Appl Microbiol Biotechnol 54: 287-301. doi: 10.1007/s002530000403
Puspitasari IN, Moeis MR (2008) Screening of seven cellulase and xylanase producing Bacillus species and analysis of the cellulase and xylanase expression pattern of the selected isolate. Pp 194-201. Proceedings of the Second International Conference on Mathematics and Natural Sciences (ICMNS) 28-30 October, Bandung
Sadhu S, Maiti TK (2013) Cellulase production by bacteria: A review. Br Microbiol Res J 3: 235-258. doi: 10.5281/zenodo.8687
Singh V, Haque S, Niwas R, Srivastava A, Pasupuleti M, Tripathi CKM (2017). Strategies for fermentation medium optimization: An in-depth review. Front Microbiol 7: 2087. doi: 10.3389/fmicb.2016.02087
Smith A (1995) Gene Expression in
Recombinant Microorganisms. Marcel
Dekker Inc, New York
Studier FW, Daegelen P, Lenski RE, Maslov
S, Kim JF (2009) Understanding the
differences between genome
sequences of Escherichia coli B strains
REL606 and BL21(DE3) and
comparison of the E. coli B and K12
genomes. J Mol Biol 394: 653-680. doi:
10.1016/j.jmb.2009.09.021
Xu J, Banerjee A, Pan SH, Li ZJ (2012)
Galactose can be an inducer for
production of therapeutic proteins by
auto-induction using E. coli BL21
strains. Protein Expr Purif 83: 30-36.
doi: 10.1016/j.pep.2012.02.014

53
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548–611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal:http://ejurnal.bppt.go.id/index.php/JBBI
AKTIVITAS Stenotrophomonas rhizophila DAN Trichoderma sp.
DALAM MENGHAMBAT PERTUMBUHAN Ganoderma boninense
Activities of Stenotrophomonas rhizophila and Trichoderma sp. in Inhibiting the Growth of Ganoderma boninense
Bedah Rupaedah1, Debby Viola Amanda
2, Reni Indrayanti
2, Nia Asiani
1,
Bambang Sukmadi1, Asep Ali
1, Abdul Wahid
1, Taufiq Firmansyah
1, Mahmud Sugianto
1
1Balai Bioteknologi, BPPT. Gedung 630, Kawasan Puspiptek Tangerang Selatan 15314
2Jurusan Biologi FMIPA UNJ. Jl. Rawamangun Muka, Jakarta Timur, DKI Jakarta 13220
*Email: [email protected] ABSTRACT Basal stem rot (BSR) disease in oil palm (Elaeis guineensis Jacq.) due to infection of Ganoderma boninense. Various efforts to overcome BSR disease has been done, such as by utilizing endophytic microbes. The purpose of this research was to determine the activities of Stenotrophomonas rhizophila and Trichoderma sp inhibiting the growth of G. boninense. This research was divided into three stages, namely: stability test of S. rhizophila activity against G. boninense; activity of chitinase and cellulase enzymes produced by S. rhizophila; the effectiveness of S. rhizophila and Trichoderma sp. on G. boninense in a greenhouse. The parameters observed were plant height, leaves number, chlorophyll content, disease incidence and severity. The stability testing of S. rhizophila activity against G. boninense showed 53% of inhibition. Chitinase activity showed negative result. While cellulase index was about 0.46. The effectiveness test showed the significantly different results on plant height, leaves number and chlorophyll content.
Keywords: Chitinase, cellulase, Ganoderma boninense, Stenotrophomonas rhizophila, Trichoderma sp.
ABSTRAK Penyakit busuk pangkal batang (BPB) pada tanaman kelapa sawit (Elaeis guineensis Jacq.) muncul karena diinfeksi oleh Ganoderma boninense. Berbagai upaya penanggulangan penyakit BPB telah dilakukan, diantaranya dengan memanfaatkan mikroba endofit. Tujuan dari penelitian ini adalah untuk mengetahui aktivitas bakteri Stenotrophomonas rhizophila dan Trichoderma sp. dalam menghambat pertumbuhan G. boninense. Penelitian ini dibagi menjadi tiga tahapan, yaitu: pengujian stabilitas aktivitas S. rhizophila terhadap G. boninense; pengujian aktivitas enzim kitinase dan selulase yang dihasilkan oleh S. rhizophila; pengujian efektivitas S. rhizophila dan Trichoderma sp. terhadap G. boninense di rumah kaca. Parameter yang diamati pada pengujian efektivitas berupa tinggi tanaman, jumlah daun, jumlah klorofil, kejadian dan keparahan penyakit. Uji stabilitas aktivitas S. rhizophila terhadap G. boninense menunjukkan adanya penghambatan rata-rata sebesar 53%. Uji aktivitas enzim kitinase pada bakteri S. rhizophila menunjukkan hasil negatif. Sedangkan indeks enzim selulase pada bakteri S. rhizophila sebesar 0.46. Pada uji efektivitas tampak hasil yang berbeda nyata pada parameter tinggi tanaman, jumlah daun dan kandungan klorofil.
Kata Kunci: Kitinase, selulase, Ganoderma boninense, Stenotrophomonas rhizophila, Trichoderma sp.
Received: 20 February 2018 Accepted: 07 May 2018 Published: 11 June 2018

Aktivitas Stenotrophomonas rhizophila dan Trichoderma sp.... Rupaedah et al.
54
PENDAHULUAN
Kelapa sawit (Elaeis guineensis Jacq.)
merupakan salah satu sumber minyak nabati yang menjadi komoditas utama dalam bidang pertanian di Indonesia. Produksi minyak sawit digunakan dalam industri makanan, bahan kimia, dan menjadi pengganti bahan bakar minyak yang saat ini sebagian besar diutamakan untuk minyak bumi. Indonesia merupakan negara penghasil kelapa sawit terbesar di dunia. Lahan sawit Indonesia yang tercatat hingga saat ini seluas 11,67 juta hektar (Dirjen Perkebunan 2015).
Penyakit busuk pangkal batang (BPB) merupakan salah satu masalah yang paling berpengaruh dalam produksi kelapa sawit Indonesia. Penyakit ini muncul pertama kali di Indonesia pada tahun 1931 dan telah menyebabkan 50% kematian pada perkebunan kelapa sawit di PT Perkebunan Nusantara IV Simalungun, Sumatra Utara (Susanto 2011). Penyakit BPB merupakan penyakit tular tanah (soil borne fungi) yang disebabkan oleh jamur Ganoderma boninense. Infeksi G. boninense di lapangan berawal dari adanya persentuhan akar tanaman. Hifa jamur masuk ke dalam jaringan empulur korteks hingga ke dalam jaringan pembuluh (xilem dan floem). Tanaman yang terserang jamur tersebut akan mengalami kebusukan pada bagian pangkal batang dan lama-kelamaan mati.
Mekanisme pengendalian yang biasa dilakukan terhadap penyakit BPB adalah pencabutan, pembersihan lahan dengan pembakaran dan penggunaan fungisida sintetik. Upaya tersebut dinilai belum efektif karena berdampak buruk bagi lingkungan. Upaya yang sekarang banyak dilakukan adalah dengan menggunakan agen hayati seperti mikroba endofit yang bersifat antagonis terhadap G. boninense (Naher et al. 2012; Naher et al. 2013; Paterson 2007).
Agen hayati yang telah dilaporkan mampu menghambat pertumbuhan jamur patogen G. boninense diantaranya adalah Trichoderma (Alexander et al. 2015; Musa
dan Ali 2017; Naher et al. 2012; Naher et al. 2015). Agen hayati Trichoderma spp. dapat digunakan untuk pengendalian jamur patogen Sclerotium rolfsii pada tanaman
bunga matahari (Yaqub dan Shahzad 2011), jamur patogen Phytophthora palmivora pada
tanaman kakao (Umrah et al. 2009); VA mycorrhizae (Glomus spp.) sebagai
pengendali layu akar pada tanaman wijen (Ziedan et al. 2011).
Tujuan dari penelitian ini adalah untuk mengetahui aktivitas mikroba endofit, dalam hal ini bakteri Stenotrophomonas rhizophila dan Trichoderma sp. yang dapat digunakan
sebagai pengendali hayati untuk meng-hambat pertumbuhan jamur patogen G. boninense pada bibit tanaman kelapa sawit.
BAHAN DAN METODE Bahan
Bahan-bahan yang digunakan dalam penelitian ini adalah biakan G. boninense, bakteri S. rhizophila, Trichoderma sp., PDA (Potato Dextrose Agar), NA (Nutrient Agar), Aquades steril, alkohol 70%, kulit udang kering, HCl 1N, NaOH, tanah, pasir, dan balok kayu tanaman karet.
Metode
Penelitian terdiri dari 3 tahapan, yaitu: (1) pengujian stabilitas bakteri S. rhizophila terhadap G. boninense secara in vitro; (2) pengujian aktivitas enzim kitinase dan selulase dari S. rhizophila secara in vitro; (3) pengujian efektivitas bakteri S. rhizophila dan Trichoderma sp. terhadap G. boninense secara in vivo. Metode eksperimen
menggunakan desain rancangan acak lengkap (Tabel 1). Data pengamatan tinggi tanaman, jumlah klorofil, keterjadian penyakit dan keparahan penyakit dicatat dengan menggu-nakan skala linear menurut Abdullah et al. (2003). Data diolah menggunakan ANOVA dua arah, dilanjutkan dengan uji DMRT (Duncan Multiple Range Test) pada taraf 5%.
Ujian stabilitas bakteri S. rhizophila
Percobaan ini bertujuan untuk melihat kemampuan bakteri antagonis S. rhizophila yang telah disubkultur berulang terhadap G. boninense. Media perlakuan pada percobaan ini dibagi menjadi tiga, yaitu media pertumbuhan jamur G. boninense, media pertumbuhan isolat bakteri S. rhizophila, dan media uji antagonis. Bakteri S. rhizophila ditumbuhkan pada media
pertumbuhan NA hingga enam kali subkultur. Bakteri S. rhizophila selanjutnya diuji antagonis terhadap G. boninense. Uji

J Bioteknol Biosains Indones– Vol 5 No 1 Thn 2018
55
antagonis dilakukan dengan menggunakan metode biakan ganda dengan perbandingan 1 : 1 secara in-vitro dalam satu cawan.
Uji aktivitas enzim kitinase S. rhizophila
Tahap awal pengujian aktivitas enzim kitinase dilakukan dengan pembuatan koloidal kitin. Kulit udang sebanyak 20 g ditambahkan 400 mL HCl 37%, diaduk dan diinkubasi selama 2 jam, selanjutnya diinkubasi semalaman pada suhu 4°C, selanjutnya disaring dengan menggunakan glasswol. Residu hasil saringan ditambahkan
aquades dan larutan NaOH 10 N (200 g kristal ditambahkan 500 mL aquades) dan dikocok perlahan dalam keadaan dingin, pH diatur hingga netral, selanjutnya disentrifugasi pada 4800 rpm selama 30 menit. Pelet yang terbentuk dibilas dengan aquades sebanyak dua kali dan digunakan sebagai koloidal kitin.
Pada pengujian ini dibuat 2 jenis media yaitu media chitin broth dan chitin agar. Pembuatan chitin broth dilakukan dengan mencampurkan KH2PO4 0,1%, MgSO4·7H2O 0,05%, yeast extract 0,1%, dan koloidal kitin
1% yang dilarutkan dalam 1 L aquades. Pembuatan chitin agar dilakukan dengan
mencampurkan KH2PO4 0,1%, MgSO4·7H2O 0,05%, yeast extract 0,1%, koloidal kitin 1%, dan
agar 2% yang dilarutkan dalam 1 L aquades. Pengujian aktivitas enzim kitinase
dilakukan dengan menginokulasikan satu ose S. rhizophila ke dalam 20 mL media chitin broth. Setelah dikocok selama 24 jam,
selanjutnya bakteri diuji aktivitasnya dengan cara memasukkan 15 µL suspensi bakteri ke dalam media chitin agar yang sudah dilubangi dengan menggunakan cork borer.
Pengamatan dilakukan sampai terbentuk zona bening di sekitar sumur. Indek kitinase dihitung dengan menggunakan rumus berikut:
dimana
IK : Indeks Kitinase Ø ZBe : Diameter Zona Bening Ø KB : Diameter Koloni Bakteri
Uji aktivitas enzim selulase S. rhizophila
Pada penelitian ini dibuat 2 jenis media yaitu media CMC broth dan CMC agar. Pembuatan CMC broth dilakukan dengan mencampurkan KH2PO4 0,1%, MgSO4·7H2O 0,05%, yeast extract 0,1%, dan CMC 5%
yang dilarutkan dalam 1 L aquades. Pembuatan CMC agar dilakukan dengan
mencampurkan KH2PO4 0,1%, MgSO4·7H2O 0,05%, yeast extract 0,1%, CMC 5%, dan agar 2% yang dilarutkan dalam 1 L aquades.
Pengujian aktivitas enzim selulase dilakukan dengan menginokulasikan satu ose S. rhizophila ke dalam 20 mL media CMC broth. Setelah diinkubasi selama 48
jam, bakteri diuji aktivitasnya dengan cara memasukkan 15 µL suspensi bakteri ke dalam media CMC agar yang sudah dilubangi dengan menggunakan cork borer.
Pengamatan dilakukan pada jam ke-72. Bakteri yang telah diinkubasi ditetesi larutan Congo red 1% dan dibilas dengan menggu-
nakan aquades. Indeks selulase (IS) dihitung dengan menggunakan rumus berikut:
dimana
IS : Indeks Selulase Ø ZBe : Diameter Zona Bening Ø KB : Diameter Koloni Bakteri Uji efektivitas S. rhizophila dan Trichoderma sp.
Pengujian ini dilakukan pada media pertumbuhan kelapa sawit berupa campuran tanah dan pasir dengan perbandingan 1 : 1
di dalam polibag berukuran 30 40 cm. G. boninense yang digunakan terlebih dahulu ditumbuhkan pada batang karet berukuran 6
6 6 cm yang telah dihaluskan dan disterilisasi menggunakan Autoclave pada
Tabel 1. Rancangan percobaan uji antagonis in vivo
Perlakuan Tanpa
Inokulum IIsolat S.
rhizophila (B) Trichoderma sp.
(T) Isolat S. rhizophila +
Tricoderma sp.
Tanpa G. boninense G0B0
15n G0B1
15n G0T1
15n G0B1T1
15n
Inokulasi G. boninense G1B0
15n G1B1
15n G1T1
15n G1B1T1
15n
n= jumlah ulangan

Aktivitas Stenotrophomonas rhizophila dan Trichoderma sp.... Rupaedah et al.
56
suhu 121ºC selama 30 menit. G. boninense
diinokulasikan pada batang karet secara aseptis. Inokulum ditumbuhkan hingga berusia kurang lebih 60 hari.
Pengujian efektivitas dilakukan dengan menumbuhkan benih kelapa sawit pada media uji yang disertai dengan pemberian inokulum bakteri S. rhizophila, Trichoderma sp. dan G. boninense dengan jarak tanam
tertentu. Berikut adalah perlakuan yang diberikan:
Parameter pengamatan dibagi menjadi 2 bagian: (1) parameter pengamatan pertumbuhan vegetatif dengan melihat tinggi tanaman, jumlah daun dan kandungan klorofil yang diamati setiap dua minggu sekali, (2) parameter pengamatan kejadian dan keparahan penyakit. Keparahan penyakit dicatat dengan menggunakan skala linear dari 0 hingga 4 (Abdullah et al. 2003), sebagai berikut: 0 - tanaman yang sehat, 1 – terdapat nekrosis pada tiga helai daun, 2 – terdapat lebih dari tiga helai daun yang mengalami nekrosis, 3 – muncul tubuh buah G. boninense pada pangkal batang, 4 –
tanaman mati. Data kuantitatif diolah dengan menggunakan anova dua arah dan dilanjutkan dengan uji Duncan dengan taraf kepercayaan 95%. Kejadian dan keparahan penyakit dihitung dengan menggunakan rumus:
dimana DI : Kejadian Penyakit
D : Jumlah Tanaman Sakit
H : Jumlah Tanaman Sehat
dimana DS : keparahan penyakit n : jumlah jaringan terserang pada setiap
kategori (skor) V : kategori (skor) serangan Z : nilai kategori serangan tertinggi N : jumlah seluruh tanaman atau bagian
tanaman yang diamati HASIL DAN PEMBAHASAN
Stabilitas aktivitas S. rhizophila
Bakteri antagonis S. rhizophila diuji
kestabilan aktivitasnya dalam menghambat pertumbuhan G. boninense
dengan cara meregenerasi bakteri tersebut sebanyak enam kali, selanjutnya diuji antagonis kembali menggunakan jamur patogen G. boninense secara in vitro. Hasil uji stabilitas menunjukkan
bahwa tidak terdapat perbedaan yang signifikan antar perlakuan uji. Rerataan penghambatan tertinggi terdapat pada biakan keempat dengan panjang miselium Ganoderma sebesar 1,65 ± 0,05 cm dan
persentase penghambatan sebesar 63%, sedangkan untuk rerataan penghambatan terendah terdapat pada biakan kelima sebesar 1,65 ± 0,05 cm dan persentase penghambatan sebesar 53% (Tabel 2). Berdasarkan hasil tersebut ditunjukkan bahwa regenerasi bakteri antagonis setelah beberapa kali tidak berpengaruh terhadap kemampuan bakteri tersebut dalam menghambat pertumbuhan G. boninense.
Tabel 2. Pertumbuhan jamur G. boninense dan persentase penghambatan bakteri S. rhizophila
Biakan N Panjang hifa G. boninense (rerata ±SE cm)
% Hambatan hari 0-5 hari 6-10 hari 11-15
Kontrol 2 2,25 ± 0,25 3,60 ± 0,01 4,50 ± 0,00 0%
F1 2 1,65 ± 0,25 1,80 ± 0,03 1,85 ± 0,25 58,50%
F2 2 1,75 ± 0,25 1,60 ± 0,00 1,75 ± 0,05 61%
F3 2 1,65 ± 0,05 1,85 ± 0,05 1,90 ± 0,00 57%
F4 2 1,65 ± 0,50 1,65 ± 0,15 1,65 ± 0,05 63%
F5 2 1,70 ± 0,00 2,05 ± 0,05 2,10 ± 0,10 53%
F6 2 1,70 ± 0,01 1,90 ± 0,00 1,90 ± 0,00 57%

J Bioteknol Biosains Indones– Vol 5 No 1 Thn 2018
57
Bakteri merupakan organisme prokariot yang mudah bermutasi. Kondisi lingkungan dapat menyebabkan perubahan pada fenotip bakteri. Bakteri yang dibiakkan berulang-ulang pada media yang sama (artificial medium) yang kaya akan nutrisi
akan membuat mikroba tidak perlu lagi berkompetisi untuk mendapatkan nutrisi sehingga akan menurunkan kemampuannya untuk memproduksi senyawa metabolit yang biasa digunakan sebagai proteksi. Akibatnya tingkat virulensi bakteri akan menurun dan mengubah genetik dari bakteri tersebut. Perubahan genetik pada bakteri dapat juga berupa perubahan fisik, baik bentuk, warna, dan ukuran bakteri (Harris dan Theriot 2016). Pada penelitian ini tidak terjadi perubahan kemampuan bakteri dalam menghambat pertumbuhan Ganoderma boninense, sehingga bisa dikatakan bahwa
perubahan yang terjadi pada bakteri selama regenerasi berulang tidak mempengaruhi kemampuannya secara signifikan.
Aktivitas enzim kitinase dan selulasae
Pada pengujian aktivitas enzim
kitinase, sebanyak 64 108 cfu koloni bakteri S. rhizophila diinokulasikan pada media kitin
agar yang telah dilubangi, kemudian diinkubasi hingga terbentuk zona bening. Setelah dilakukan pengamatan hingga 30 hari tidak terdapat zona bening di sekitar koloni bakteri (Gambar 2A). Tidak adanya
zona bening mengindikasikan bahwa tidak terdapat aktivitas enzim kitinase pada bakteri S. rhizophila. Zona bening merupakan tanda
bahwa mikroorganisme kitinolitik seperti bakteri maupun jamur memiliki enzim kitinase yang berfungsi sebagai katalis yang mampu menghidrolisis polimer kitin menjadi kitin oligisakarida atau monomer N-asetilglukosamin. Enzim kitinase mempunyai peranan penting dalam kontrol biologi berbagai jamur patogen dengan mendegradasi senyawa kitin yang ada pada dinding sel jamur patogen (El-Katatny et al. 2001). Kitin merupakan salah satu komponen terbesar penyusun dinding sel Ganoderma. Bakteri yang memilki enzim
kitinase dapat membentuk zona bening, besar kecilnya zona bening yang terbentuk sesuai dengan aktivitas enzim kitinase mikroba tersebut. Zona bening terbentuk karena adanya pemutusan ikatan β-1,4 homopolimer N-asetilglukosamin pada kitin menjadi monomer N-asetilglukosamin.
Pada pengujian enzim selulase,
sebanyak 9 107 cfu bakteri S. rhizophila diinokulasikan ke dalam media CMC agar yang telah dilubangi, kemudian diinkubasi selama 72 jam. Setelah inkubasi, media tersebut diwarnai dengan menggunakan Congo red dan dibilas dengan menggunakan aquades steril. Dari hasil pengamatan terdapat zona bening di sekitar koloni bakteri. Dari perhitungan diameter zona
Gambar 1. Aktivitas bakteri S. rhizophila dalam menghambat pertumbuhan G. boninense; (A) penghambatan tertinggi pada biakan keempat (F4); (B) penghambatan terendah pada biakan kelima (F5)

Aktivitas Stenotrophomonas rhizophila dan Trichoderma sp.... Rupaedah et al.
58
bening dengan diameter koloni bakteri diperoleh indeks selulase sebesar 0.46 (Gambar 2B). Zona bening menunjukkan zona tempat terputusnya ikatan β-1,4-glikosidik yang menghubungkan monomor D-glukosa pada CMC (Haryati et al. 2010). Hal ini menunjukkan bahwa isolat yang memiliki zona bening merupakan isolat yang memiliki kemampuan dalam mendegradasi selulosa. Dinding sel Ganoderma juga
tersusun oleh beberapa polisakarida. Polisakarida penyusun dinding sel Ganoderma diantaranya adalah glukosa,
galaktosa, arabinosa, xylosa dan manosa. Ikatan polisakarida dari Ganoderma berupa β-glukan yang cukup melimpah pada dinding selnya. β-glukan merupakan salah satu penyusun rantai dari selulosa. β-glukan akan terdegradasi oleh enzim selulase, dengan memutus ikatan β-1,4-glikosidik. Penggunaan bakteri antagonis dapat menghambat pertumbuhan miselium Ganoderma karena bakteri antagonis
memproduksi enzim selulase yang mampu memotong ikatan β-glukan pada Ganoderma.
Efektivitas S. rhizophila
Berdasarkan pengamatan terhadap tinggi tanaman kelapa sawit, diperoleh hasil yang berbeda signifikan antar perlakuan. Pertumbuhan kelapa sawit dengan rerataan tertinggi terdapat pada perlakuan tanaman kelapa sawit yang ditumbuhkan tanpa G.
boninense dan dengan Trichoderma (G0T1) sebesar 33,13 ± 1,71 cm dan rerataan terendah pada perlakuan tanaman kelapa sawit yang ditumbuhkan tanpa G. boninense dan dengan bakteri S. rhizophila (G0B1) sebesar 25,15 ± 1,60 cm (Tabel 3).
Pada pengamatan jumlah daun terdapat perbedaan yang signifikan pada semua perlakuan. Jumlah rerataan daun tertinggi terdapat pada perlakuan tanaman kelapa sawit yang ditumbuhkan tanpa Ganoderma dan dengan Trichoderma (G0T1)
sebesar 6,27 ± 0,48 cm dan rerataan terendah pada perlakuan tanaman kelapa sawit dengan Ganoderma dan tanpa bakteri S. rhizophila (G0B1) sebesar 4,71 ± 0,16 cm.
Pertumbuhan daun kelapa sawit pada minggu keenam dan minggu keduabelas tidak menunjukkan perbedaan yang signifikan pada semua perlakuan. Pada minggu terakhir terdapat perbedaan antara perlakuan kelapa sawit yang ditumbuhkan tanpa Ganoderma dan dengan Trichoderma
(G0T1) (Tabel 4). Jumlah klorofil diamati pada minggu
terakhir pengamatan. Pada Tabel 5 terlihat bahwa terdapat perbedaan yang signifikan jumlah klorofil dari setiap perlakuan. Jumlah klorofil tertinggi terdapat pada perlakuan tanaman kelapa sawit yang ditumbuhkan tanpa Ganoderma dan dengan bakteri S. rhizophila (G0B1) dengan rerataan sebesar 28,05 ± 2,95 cm dan jumlah klorofil terendah pada perlakuan tanaman kelapa sawit yang
Gambar 2. Pengujian aktivitas enzim; (A) kitinase yang dilakukan dalam media kitin agar terhadap pembentukan zona bening di sekitar koloni bakteri. (B) enzim selulase terdapat koloni bakteri dan menghasilkan zona bening

J Bioteknol Biosains Indones– Vol 5 No 1 Thn 2018
59
Tabel 3. Rerataan tinggi tanaman kelapa sawit setelah 18 minggu pengamatan
Pengamatan N
Tinggi tanaman (rerata ± SE cm)
G0B0 G0B1 G0T1 G0K1 G1B0 G1B1 G1T1 G1K1
4 minggu 120 6,57ab
± 0,48 5,57a ± 0,34 7,67
c ± 0,34 6,10
ab ± 0,56 7,33
bc ± 0,22 6,40
ab ± 0,35 6,11
ab ± 0,32 7,09
bc ± 0,43
6 minggu 120 12,76b ± 1,15 9,53
a ± 0,98 13,13
b ± 0,72 11,15
ab ± 0,96 12,43
b ± 0,63 11,60
ab ± 0,57 11,46
ab ± 0,73 11,69
ab ± 0,77
8 minggu 120 16,83a ± 1,57 14,28
a ± 1,19 20,95
b ± 0,98 17,54
a ± 0,75 16,93
a ± 0,96 17,13
a ± 0,72 16,68
a ± 0,90 16,66
a ± 1,20
10 minggu 120 18,30b ± 1,66 14,70
a ± 1,16 22,04
c ± 0,81 18,00
b ± 0,97 17,60
ab ± 0,91 18,10
b ± 0,74 17,86
ab ± 0,85 18,27
b ± 1,07
12 minggu 120 20,40ab
± 1,84 17,10a ± 1,24 22,73
b ± 0,82 19,59
ab ± 1,06 20,25
ab ± 0,85 19,73
ab ± 0,84 20,53
ab ± 0,85 20,53
ab ± 1,09
13 minggu 120 23,13ab
± 1,97 19,60a ± 1,43 25,17
b ± 1,04 22,18
ab ± 1,09 22,36
ab ± 1,21 21,17
ab ± 0,94 22,57
ab ± 0,95 22,53
ab ± 1,17
14 minggu 120 25,50ab
± 1,98 21,50a ± 1,46 27,57
b ± 1,23 23,59
ab ± 1,13 23,89
ab ± 1,21 23,67
ab ± 0,89 24,64
ab ± 1,15 24,57
ab ± 1,22
15 minggu 120 26,33ab
± 1,89 22,07a ± 1,39 27,90
b ± 1,23 24,72
ab ± 1,13 25,00
ab ± 1,22 25,20
ab ± 0,86 25,92
ab ± 1,31 25,92
ab ± 1,18
16 minggu 120 28,27ab
± 2,04 23,70a ± 1,44 30,17
b ± 1,53 26,00
ab ± 1,18 26,53
ab ± 1,28 27,00
ab ± 0,91 27,71
ab ± 1,47 27,50
ab ± 1,36
17 minggu 120 29,83b ± 2,12 24,70
a ± 1,56 32,07
b ± 1,71 27,27
ab ± 1,16 27,53
ab ± 1,29 28,80
ab ± 1,02 28,35
ab ± 1,57 28,54
ab ± 1,58
18 minggu 120 30,50bc
± 2,03 25,15a ± 1,60 33,13
c ± 1,71 28,68
b ± 0,97 28,14
b ± 1,29 3 ± 0,10
b ± 0,84 29,57
b ± 1,62 29,19
b ± 1,42
Keterangan:
SE = standard error
*N = jumlah tanaman yang diinokulasi
*G0B0 = tanpa Ganoderma, tanpa mikroba antagonis (kontrol); G0B1= tanpa Ganoderma, dengan S. rhizophila; G0T1= tanpa Ganoderma, dengan Trichoderma;
G0K1 = tanpa Ganoderma, dengan korsorsium mikroba antagonis, G1B0= Ganoderma, tanpa S. rhizophila; G1B1= dengan Ganoderma dan S. rhizophila;
G1T1 = dengan Ganoderma dan Trichoderma; G1K1= dengan Ganoderma dan konsorsium mikroba antagonis
Angka yang diikuti oleh huruf yang sama pada baris yang sama tidak berbeda signifikan.

Aktivitas Stenotrophomonas rhizophila dan Trichoderma sp.... Rupaedah et al.
60
Tabel 4. Jumlah daun kelapa sawit setelah 18 minggu tanam
Pengamatan N
Jumlah daun (rerata ± SE cm)
G0B0 G0B1 G0T1 G0K1 G1B0 G1B1 G1T1 G1K1
4 minggu 120 1,93ab
± 0,11 1,87ab
± 0,09 2,20b ± 0,32 2,00
ab ± 0,13 2,00
ab ± 0,09 1,93
ab ± 0,06 1,86
ab ± 0,09 1,69
a ± 0,12
6 minggu 120 2,33a ± 0,12 2,27
a ± 0,15 2,33
a ± 0,18 2,31
a ± 0,13 2,13
a ± 0,91 2,13
a ± 0,91 2,07
a ± 0,71 2,46
a ± 0,18
8 minggu 120 2,87ab
± 0,13 2,80ab
± 0,10 3,33b ± 0,28 3,82
ab ± 0,18 2,67
a ± 0,12 3,00
ab ± 0,09 3,00
ab ± 0,21 3,08
ab ± 0,28
10 minggu 120 2,93ab
± 0,15 2,73a ± 0,12 3,40
b ± 0,27 3,09
ab ± 0,91 2,87
ab ± 0,16 3,13
ab ± 0,09 3,07
ab ± 0,19 3,23
ab ± 0,23
12 minggu 120 3,27a ± 0,2 3,20
a ± 0,14 3,80
a ± 0,20 3,36
a ± 0,15 3,29
a ± 0,16 3,33
a ± 0,12 3,29
a ± 0,19 3,46
a ± 0,26
13 minggu 120 3,60ab
± 0,21 3,53a ± 0,13 4,27
b ± 0,33 4,00
ab ± 0,13 3,64
ab ± 0,13 3,87
ab ± 0,16 3,64
ab ± 0,16 3,77
ab ± 0,28
14 minggu 120 3,80ab
± 0,22 3,53a ± 0,19 4,40
b ± 0,32 4,18
ab ± 0,18 3,86
ab ± 0,14 4,40
b ± 0,21 3,93
ab ± 0,19 4,31
b ± 0,28
15 minggu 120 4,00ab
± 0,25 3,67a ± 0,15 4,67
b ± 0,33 4,09
ab ± 0,15 4,07
ab ± 0,16 4,47
b ± 0,16 4,14
ab ± 0,23 4,23
ab ± 0,28
16 minggu 120 4,40a ± 0,28 4,27
a ± 0,20 5,27
b ± 0,35 4,91
ab ± 0,10 4,36
a ± 0,16 4,93
ab ± 0,18 4,36
a ± 0,28 4,77
ab ± 0,32
17 minggu 120 4,67a ± 0,28 5,00
ab ± 0,37 5,67
b ± 0,42 5,72
ab ± 0,14 4,93
ab ± 0,19 4,93
ab ± 0,20 5,00
ab ± 0,27 4,92
ab ± 0,34
18 minggu 120 5,00ab
± 0,27 5,27b ± 0,38 6,27
c ± 0,48 5,09
b ± 0,21 4,71
a ± 0,16 5,40
b ± 0,16 5,00
ab ± 0,25 5,31
b ± 0,34
Ket erangan:
SE = standard error
*N = jumlah tanaman yang diinokulasi
*G0B0 = tanpa Ganoderma, tanpa mikroba antagonis (kontrol); G0B1= tanpa Ganoderma, dengan S. rhizophila; G0T1= tanpa Ganoderma, dengan Trichoderma;
G0K1 = tanpa Ganoderma, dengan korsorsium mikroba antagonis, G1B0= Ganoderma, tanpa S. rhizophila; G1B1= dengan Ganoderma dan S. rhizophila;
G1T1 = dengan Ganoderma dan Trichoderma; G1K1= dengan Ganoderma dan konsorsium mikroba antagonis
Angka yang diikuti oleh huruf yang sama pada baris yang sama tidak berbeda signifikan.

J Bioteknol Biosains Indones– Vol 5 No 1 Thn 2018
61
ditumbuhkan tanpa Ganoderma dan dengan Trichoderma (G0T1) dengan rerataan
sebesar 15,03 ± 2,63 cm (Tabel 4). Pengamatan jumlah klorofil berbanding terbalik dengan hasil pengamatan tinggi tanaman dan jumlah daun. Ukuran daun tidak menentukan kuantitas klorofil. Pada perlakuan kelapa sawit yang ditumbuhkan tanpa Ganoderma dan dengan Trichoderma
(G0T1), daun yang dihasilkan memiliki ukuran yang lebih besar namun warna hijau pada daunnya tidak terlalu pekat, sedangkan pada perlakuan kelapa sawit yang ditumbuhkan tanpa Ganoderma dan dengan bakteri S. rhizophila (G0B1) walaupun memiliki daun
dengan ukuran yang relatif kecil, namun memiliki warna hijau yang lebih pekat pada daunnya.
Pengujian keparahan dan kejadian penyakit juga dilakukan untuk melihat seberapa besar kemampuan patogen Ganoderma dalam menyerang tanaman
kelapa sawit. Hingga minggu ke-18 sejak kelapa sawit ditanam, kejadian dan keparahan penyakit tercatat bervariasi pada tiap-tiap perlakuan. Kejadian pernyakit tertinggi terdapat pada perlakuan G1B0 sebesar 28% dan keparahan penyakit sebesar 18%. Skoring tertinggi untuk keparahan penyakit sebesar 2 (lebih dari tiga helai daun klorosis/nekrosis) (Tabel 6). Untuk melihat keparahan penyakit minimal dilakukan hingga usia kelapa sawit mencapai kurang lebih satu tahun dari penanaman. Untuk mencapai tumbuhnya tubuh buah Ganoderma pada batang kelapa
sawit dibutuhkan waktu yang cukup lama, kurang lebih sekitar satu tahun dari
penanaman kelapa sawit. Penggunaan Trichoderma mampu
meningkatkan pertumbuhan tinggi dan jumlah daun tanaman kelapa sawit di rumah kaca dibandingkan perlakuan lainnya. Trichoderma merupakan jamur tanah yang
berperan dalam menguraikan bahan organik tanah, dimana bahan organik tanah ini mengandung beberapa komponen zat seperti N, P, S dan Mg dan unsur hara lain yang dibutuhkan tanaman dalam pertumbuhannya. Trichoderma dapat
menguraikan posfat dari Al, Fe dan Mn. Beberapa mikroba antagonis biasanya
mampu mengatasi efek fungistatis tanah yang berasal dari metabolit yang dihasilkan oleh spesies lain, termasuk tanaman, dan dapat hidup pada kondisi ekstrem pada lingkungan yang tidak menguntungkan. Trichoderma tumbuh dengan cepat di dalam tanah, karena mereka secara alami resisten terhadap berbagai senyawa beracun, termasuk herbisida, fungisida dan insektisida seperti DDT, dan senyawa fenolik.
Inokulasi bakteri S. rhizophila dapat
meningkatkan jumlah klorofil pada tanaman kelapa sawit. Menurut Anggarwulan et al. (2008) bakteri yang dikemas dalam bentuk pupuk hayati lebih efektif dalam meningkatkan pertumbuhan, pembentukan nutrisi, kandungan klorofil dan produksi biomassa. Mikroba antagonis dapat mempengaruhi proses metabolisme sehingga dapat meningkatkan suplai P, dan konsentrasi P yang tinggi akan meningkatkan penambatan nitrogen. Kadar klorofil dapat dijadikan indikator sensitif pada kondisi fisiologis tumbuhan, karena kandungan klorofil berkorelasi positif dengan kandungan nitrogen daun, sehingga dapat Tabel 5. Jumlah klorofil tanaman kelapa sawit pada
minggu ke 18 pasca tanam
No. Perlakuan N Jumlah klorofil
(rerataan ± SE)
1 G0B0 15 18,41ab
± 3,04
2 G0B1 15 28,05c ± 2,95
3 G0T1 15 15,03a ± 2,63
4 G0K1 15 20,58b ± 1,69
5 G1B0 15 22,26b ± 2,83
6 G1B1 15 22,17b ± 2,05
7 G1T1 15 19,91ab
± 2,90
8 G1K1 15 22,59bc
± 3,47
Tabel 6. Indeks kejadian dan keparahan penyakit busuk pangkal batang pada minggu ke-20
No. Perlakuan %
Kejadian Penyakit
% Keparahan Penyakit
1 G0B0 0% 0%
2 G0B1 0% 0%
3 G0T1 0% 0%
4 G0K1 7% 9%
5 G1B0 28% 18%
6 G1B1 13% 13%
7 G1T1 7% 7%
8 G1K1 13% 16%

Aktivitas Stenotrophomonas rhizophila dan Trichoderma sp.... Rupaedah et al.
62
dijadikan indikator laju fotosintesis. Jika pada tanaman mengalami peningkatan laju fotosintesis maka semakin banyak karbohidrat yang terbentuk. Unsur P dalam tanah berperan penting dalam meningkatkan efisiensi kerja kloroplas yang berfungsi sebagai penyerap energi matahari dalam proses fotosintesis, hal tersebut dikarenakan unsur P berperan aktif mentransfer energi dalam sel tanaman. Energi yang dihasilkan dalam proses fotosintesis disebarkan pada jaringan-jaringan tanaman yang mengakibatkan terjadinya pembelahan sel untuk membentuk anakan baru.
Berdasarkan keseluruhan percobaan, perlakuan dengan menggunakan Trichoderma menunjukan hasil yang paling baik dibandingkan perlakuan lainnya. Trichoderma telah diakui sebagai mikroba
yang baik untuk pertumbuhan tanaman dan sebagai biokontrol untuk menghambat pertumbuhan dari mikroba patogen lainnya terutama jamur.
KESIMPULAN
Bakteri S. rhizophila yang diregenerasi
sebanyak 6 kali tidak menurunkan kemampuan bakteri dalam menghambat pertumbuhan G. boninense. Berdasarkan
hasil pengujian, tidak terdeteksi bahwa bakteri S. rhizophila memiliki aktivitas enzim
kitinase, namun memiliki kemampuan menghasilkan enzim selulase dengan indeks selulase sebesar 0,46. Pada pengujian di rumah kaca, bakteri S. rhizophila mampu
meningkatkan jumlah klorofil dan Trichoderma sp. unggul dalam
meningkatkan tinggi tanaman dan jumlah daun tanaman kelapa sawit. DAFTAR PUSTAKA
Abdullah F, Ilias GNM, Nelson M, Izzati
NAMZ, Yusuf UK (2003) Disease assessment and the efficacy of Trichoderma as a biocontrol agent of basal stem rot of oil palms. Res Bul Sci Putra 11: 31-33
Alexander A, Dayou J, Chong KP (2015) Morphological changes of Ganoderma boninense mycelia after challenged by
Tricoderma and Bacillus. AIP Conference Proceedings 1669: 020075
Anggarwulan E, Solichatun, Mudyantini W (2008) Karakter fisiologi kimpul (Xanthosoma sagittifolium (L.) Schott)
pada variasi naungan dan ketersediaan air. Biodiversitas 9: 264-268. doi: 10.13057/biodiv/d090405
Dirjen Perkebunan (2015) Statistik Perkebunan Indonesia Komoditas Kelapa Sawit 2014-2016. Kementerian Pertanian
El-Katatny MH, Gudelj M, Robra KH, Elnaghy MA, Gübitz GM (2001). Characterization of a chitinase and an endo-β-1, 3-glucanase from Trichoderma harzianum Rifai T24
involved in control of the phytopathogen Sclerotium rolfsii. Appl Microbiol Biotechnol 56: 137-143. PMID: 11499921
Harris LK, Theriot JA (2016) Relative rates of surface and volume synthesis set bacterial cell size. Cell 165: 1479-1492. doi: 10.1016/j.cell.2016.05.045
Haryati T, Marbun PA, Purwadaria T (2010) Preservasi xilanase Bacillus pumilus
PU4-2 dengan teknik imobilisasi pada pollard dan penambahan kation. JITV 15: 63-71
Musa H, Ali NBS (2017) Trichoderma spp
consortium and its efficacy as biological control agent of Ganoderma disease of oil palm (Elaeis guineensis
Jacquin). World Academy of Science, Engineering and Technology. Proceeding 19th International Conference on Forest Mycology and Plant Pathology. November 13-14, 2017, Tokyo
Naher L, Yusuf UK, Siddiquee S, Ferdous J, Rahman MA (2012) Effect of media on growth and antagonistic activity of selected Trichoderma strains against Ganoderma. Afr J Microbiol Res 6:
7449-7453. doi: 10.5897/AJMR12.1216
Naher L, Yusuf UK, Ismail A, Tan SG, Mondal MMA (2013) Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis
Jacq.). Aust J Crop Sci 7: 1723-1727 Naher L, Mokhtar SI, Sidek N (2015).
Tricoderma harzianum T32 growth and
antagonistic performance against Ganoderma boninense on different culture media. Pp 33-35. Proceeding

J Bioteknol Biosains Indones– Vol 5 No 1 Thn 2018
63
3rd International Conference on Biological, Chemical and Environmen-tal Sciences (BCES-2015). September 21-22, 2015, Kuala Lumpur Malaysia
Paterson RRM (2007) Ganoderma disease of oil palm: A white rot perspective necessary for integrated control. Crop Protect 26: 1369-1376. doi: 10.1016/ j.cropro.2006.11.009
Susanto A (2011) Ganoderma di perkebunan kelapa sawit dari waktu ke waktu. Simposium Nasional dan Lokakarya Ganoderma: Sebagai
Patogen Penyakit Tanaman dan Bahan Baku Obat Tradisional. 2-3 November 2011, Bogor
Umrah, Anggraeni T, Esyanti RR, Aryantha INP (2009) Antagonisitas dan efektivitas Trichoderma sp. dalam menekan perkembangan Phytophthora palmivora pada buah kakao. Agroland
16: 9-16. doi: 10.22487/J.24077607.2009.v16.i1.211
Yaqub F, Shahzad S (2011) Efficacy and persistence of micobial antagonists against Sclerotium rolfsii under field conditions. Pak J Bot 43: 2627-2634
Ziedan EH, Elewa IS, Mostafa MH, Sahab AF (2011) Application of mycorrhizae for controlling root diseases of sesame. J Plant Protect Res 51: 355-361. doi: 10.2478/v10045-011-0058-0

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
64
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
PEMANFAATAN TUMBUHAN OBAT SECARA EMPIRIS
PADA SUKU MANDAILING DI TAMAN NASIONAL BATANG GADIS SUMATERA UTARA
Empirical Utilization of Medicinal Plant on Mandailing Tribe in
Batang Gadis National Park North Sumatra
Aswarina Nasution1,4
, Tatik Chikmawati1*, Eko Baroto Walujo
2, Ervizal AM Zuhud
3
1Departemen Biologi, FMIPA, Institut Pertanian Bogor. Jl Meranti, Kampus IPB Dramaga, Bogor 16680 2Pusat Penelitian Biologi LIPI. Cibinong Science Center, Jl Raya Jakarta-Bogor Km 46, Bogor 16911
3Departemen KSHE, Fakultas Kehutanan, Jl Lingkar Akademik Kampus IPB Dramaga, Bogor 16680.
4Jurusan Biologi FMIPA, Universitas Negeri Medan. Jl Willem Iskandar Pasar V, Medan 20221
*Email: [email protected] ABSTRACT Mandailing tribe is an indigenous tribe that inhabits the area around Batang Gadis National Park (BGNP), North Sumatra. They have knowledge related to the use of plants for traditional medicine. Nevertheless, the information about this local knowledge is not uncover yet. This study aims to reveal the knowledge of the Mandailing tribe in utilizing plants as a traditional medicine. The research location was in 4 villages around BGNP. Data were collected through interviews with respondents and direct survey in the field. Data were analyzed descriptively qualitative. The results showed that there were about 81 plant species used for treatment covered in 38 families to treat 41 types of diseases. The most widely used medicinal plant species are from the Compositae family. Herbs dominant used by the community as a medicinal plant comprised 50 species of plants. The high diversity of medicinal plants indicated that utilization of plants for health is the main priorities of a Mandailing tribe. Keywords: Biodiversity, disease, local knowledge, Mandailing tribe, traditional medicine ABSTRAK Suku Mandailing merupakan suku asli yang mendiami kawasan di sekitar Taman Nasional Batang Gadis (TNBG), Sumatra Utara. Mereka memiliki pengetahuan terkait pemanfaatan tumbuhan untuk obat tradisional. Namun informasi terkait pengetahuan lokal tersebut belum diungkapkan. Penelitian ini bertujuan untuk mengungkap pengetahuan Suku Mandailing dalam memanfaatkan tumbuhan sebagai obat tradisional. Lokasi penelitian berada di 4 desa di sekitar TNBG. Pengumpulan data melalui wawancara dengan respoden serta survey langsung di lapangan. Data dianalisis secara deskriptif kualitatif. Hasil penelitian menunjukkan ada sekitar 81 spesies tumbuhan yang digunakan untuk pengobatan yang tercakup dalam 38 famili untuk mengobati 41 jenis penyakit. Spesies tumbuhan obat yang paling banyak digunakan berasal dari Famili Compositae. Habitus herba dominan digunakan masyarakat sebagai tumbuhan obat yang meliputi 50 spesies tumbuhan. Tingginya keanekaragaman tumbuhan obat menunjukkan bahwa pemanfaatan tumbuhan untuk kesehatan adalah prioritas utama Suku Mandailing.
Kata Kunci: Biodiversitas, suku Mandailing, obat tradisional, pengetahuan lokal, penyakit
Received: 19 April 2018 Accepted: 19 June 2018 Published: 28 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
65
PENDAHULUAN
Indonesia merupakan negara
megabiodiversitas dengan keanekaragaman tumbuhan yang sangat tinggi. Hutan tropis Indonesia yang terdiri dari berbagai tipe ekosistem merupakan gudang keanekaragam-an hayati lebih dari 239 spesies tumbuhan pangan dan lebih dari 2.039 spesies tumbuhan obat (Zuhud dan Hikmat 2009).
Salah satu kawasan yang memiliki keanekaragaman tumbuhan yang sangat tinggi adalah kawasan Taman Nasional Batang Gadis (TNBG). Secara administratif pemerintahan kawasan TNBG termasuk dalam wilayah Kabupaten Mandailing Natal, Provinsi Sumatera Utara. Secara geografis, TNBG terletak pada kisaran ketinggian 300-2.145 m dpl (BKSDA II Sumatera Utara 2005).
Kawasan TNBG memiliki tipe vegetasi beragam mulai dari hutan rawa dataran tinggi, lahan basah, lembah sungai, hutan gamping, hutan dataran rendah, perbukitan, sampai hutan pegunungan (Kartawinata et al. 2004). Keanekaragaman tipe vegetasi ini mempengaruhi keberadaan flora dan fauna di kawasan tersebut.
Tumbuhan di kawasan TNBG teridentifikasi sekitar 240 spesies yang terdiri dari 47 famili atau sekitar 0,9% dari spesies tumbuhan yang ada di Indonesia (Kartawinata et al. 2004). Sumber daya tumbuhan tersebut digunakan oleh Suku Mandailing yang tinggal di sekitar kawasan ini untuk memenuhi kebutuhan hidup mereka, baik sebagai sumber pangan, obat-obatan, pakan, bahan bangunan dan lain-lain.
Pemanfaatan tumbuhan secara empiris untuk mengobati berbagai jenis penyakit masih menjadi pilihan sebagian penduduk di Indonesia. Data hasil riset kesehatan dasar tahun 2013 (Balitbangkes 2013) menunjukkan sebesar 15,7% rumah tangga di Indonesia masih menggunakan obat tradisional. Penelitian mengenai tumbuhan obat telah banyak dilakukan, seperti penelitian yang dilakukan oleh Indriati (2014) tentang tumbuhan obat yang digunakan Suku Anak Dalam di Desa Tabun Kecamatan VII Koto Kabupaten Tebo Jambi, diperoleh 39 spesies tumbuhan obat yang tergabung dalam 33 famili. Penelitian yang dilakukan oleh Shanthi et al. (2014) tentang
pengobatan tradisional untuk perawatan wanita di masyarakat Keraton Surakarta
Hadiningrat, diperoleh 120 spesies tumbuhan obat yang terbagi dalam 55 famili. Hasil penelitian tersebut menunjukkan bahwa masyarakat Indonesia khususnya masyarakat lokal telah memiliki kemampuan dalam memanfaatkan berbagai spesies tumbuhan untuk pengobatan tradisional.
Suku Mandailing memiliki pengetahuan yang khas dan berbeda dari masyarakat lokal lain terkait dengan pemanfaatan tumbuhan sebagai obat tradisional. Perbedaan tersebut terutama tampak dalam pemilihan spesies tumbuhan serta pengolahan bagian/organ tumbuhan untuk dijadikan sebagai obat. Spesies yang digunakan merupakan spesies yang sering mereka temukan di lingkungan mereka yang belum tentu dapat ditemukan di tempat lain. Pengolahan tumbuhan obat dilakukan secara sederhana, bahkan banyak tumbuhan yang langsung digunakan tanpa proses pengolahan/pemasakan, berbeda dengan daerah lain terutama di Jawa. Masyarakat di Jawa umumnya mengolah tumbuhan obat menjadi produk jamu. Pengetahuan lokal ini merupakan hasil interaksi masyarakat dengan ekosistem hutan dan sumber daya hayati yang terdapat didalamnya yang telah berlangsung lama dan umumnya dipengaruhi oleh tipe ekosistem tempat mereka tinggal, iklim terutama curah hujan, budaya, ekonomi, teknologi dan politik (Walujo 2009). Pengetahuan mengenai pemanfaatan spesies tumbuhan obat banyak diperoleh dari leluhur yang mempunyai kemampuan meramu tumbuhan dan hanya disampaikan melalui lisan dari generasi ke generasi (Anderson et al. 2011), sehingga pengetahuan ini berpotensi hilang dari masyarakat.
Pengetahuan mengenai pemanfaatan tumbuhan sebagai obat tradisional yang dimiliki Suku Mandailing merupakan informasi berharga yang bermanfaat bagi pengembangan obat modern. Oleh karena itu penting untuk melakukan pengungkapan dan dokumentasi terhadap pengetahuan lokal mengenai pemanfaatan tumbuhan sebagai obat tradisional agar pengetahuan lokal ini tidak hilang dan dapat diwariskan ke generasi muda sebelum pandangan, pemikiran dan perilaku mereka terdegradasi oleh pengaruh lingkungan serta intervensi budaya dari luar. Penelitian ini bertujuan untuk mengungkap pengetahuan Suku

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
66
Mandailing dalam memanfaatkan keanekaragaman tumbuhan sebagai obat. BAHAN DAN METODE Lokasi dan waktu penelitian
Penelitian ini dilaksanakan pada bulan November 2015. Lokasi penelitian berada di 4 desa di sekitar TNBG yaitu Desa Sibanggor Jae, Desa Hutabaringin Julu, Desa Pastap Jae dan Desa Botung, Kabupaten Mandailing Natal, Sumatera Utara.
Alat dan bahan
Alat dan bahan yang digunakan dalam penelitian ini meliputi: alat rekam suara, kamera, alat tulis, gunting, parang, tali plastik, kantong plastik berbagai ukuran, amplop sampel, kertas mounting, label gantung, kertas koran, dan sasak. Bahan kimia yang digunakan adalah alkohol 70%.
Metode pengumpulan dan analisis data
Data yang diambil meliputi data keanekaragaman tumbuhan obat, pemanfaatan tumbuhan tersebut secara empiris dalam mengobati penyakit, cara meramu tumbuhan tersebut serta kandungan fitokimia tumbuhan obat.
Data keanekaragaman spesies tumbuhan obat, manfaatnya dalam mengobati penyakit serta cara meramu tumbuhan tersebut dikumpulkan melalui wawancara semi terstruktur. Wawancara dilakukan terhadap responden yaitu tokoh masyarakat, kepala desa serta penduduk yang memiliki pengetahuan yang baik mengenai keanekaragaman tumbuhan obat.
Survei lapangan dilakukan untuk verifikasi spesies dan untuk memperoleh sampel spesies tumbuhan yang dimanfaatkan berdasarkan hasil wawancara dengan responden. Sampel tumbuhan dibuat herbarium. Daun tumbuhan diambil dari lapangan kemudian diletakkan diantara kertas koran dan diapit dengan pengapit yang kuat seperti sasak bambu, kemudian diikat dengan tali. Tumbuhan dibawa ke laboratorium untuk dikeringkan dengan menggunakan oven. Identifikasi tumbuhan obat dilakukan di Herbarium Bogoriense LIPI Bogor.
Analisis fitokimia tiga sampel tumbuhan bermanfaat obat yaitu galunggung (Blumea balsamifera), galah nagori (Eupatorium sp.) serta aren/bargot (Arenga pinnata) menggunakan visualisasi warna
dilakukan di Laboratorium Pusat Studi Biofarmaka LPPM, Institut Pertanian Bogor. Untuk menguji keberadaan alkaloid, 1 gram sampel tumbuhan dihaluskan bersama beberapa tetes NH3, kemudian ditambahkan 5 mL CHCl3 lalu disaring. Filtrat ditambah H2SO4. Sampel diuji dengan tiga pereaksi alkaloid yaitu pereaksi Mayer, Wagner dan Dragendorff. Hasil uji dinyatakan positif jika dengan pereaksi Mayer terbentuk endapan putih, dengan pereaksi Wagner terbentuk endapan coklat dan dengan pereaksi Dragendorff terbentuk endapan jingga.
Kandungan steroid/triterpenoid dapat dideteksi dengan cara menambahkan etanol panas kemudian disaring. Filtrat dipanaskan hingga kering lalu ditambahkan 1 mL dietil eter kemudian dihomogenisasi. Sampel kemudian ditambahkan satu tetes H2SO4 dan CH3COOH anhidrat. Perubahan warna menjadi hijau/biru menunjukkan bahwa sampel positif mengandung steroid, sedangkan warna merah ungu menunjukkan bahwa sampel positif mengandung triterpenoid. Flavonoid dapat dideteksi dengan menambahkan serbuk magnesium, HCl; EtOH (1:1) serta amil alkohol. Terbentuknya warna jingga pada lapisan amil alkohol menunjukkan adanya flavonoid. Saponin dapat dideteksi dengan uji buih setelah dikocok kuat, buih yang stabil selama 30 menit menunjukkan adanya saponin. Tannin dapat dideteksi dengan menambahkan pereaksi FeCl3 10% ke dalam sampel. Terbentuknya warna hitam kehijauan menunjukkan terdapat komponen tannin di dalam sampel (Harborne 1987). HASIL DAN PEMBAHASAN Kekayaan dan keanekaragaman spesies
Berdasarkan hasil wawancara dengan Suku Mandailing, diperoleh 81 spesies tumbuhan obat yang digunakan untuk mengobati 41 jenis penyakit. Tumbuhan tersebut tercakup dalam 38 famili (Tabel lampiran). Spesies tumbuhan obat yang
paling banyak digunakan berasal dari Famili Compositae (Gambar 1).…………………

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
67
Gambar 1. Jumlah spesies tumbuhan obat
berdasarkan famili
Gambar 2. Jumlah spesies tumbuhan obat berdasarkan kategori penyakit
Berbagai spesies tumbuhan dari Famili
Compositae banyak digunakan masyarakat Indonesia sebagai obat. Hasil penelitian Yuniati dan Alwi (2010) menyimpulkan bahwa Compositae merupakan famili tumbuhan obat tertinggi kedua setelah Euphorbiaceae di hutan Desa Pakuli, Sulawesi Tengah. Famili Compositae juga mudah diperoleh di lingkungan masyarakat yang berada di dataran tinggi sehingga banyak digunakan masyarakat sebagai obat.
Selain Compositae, Zingiberaceae juga merupakan famili tumbuhan yang banyak digunakan Suku Mandailing. Famili ini paling banyak digunakan masyarakat Desa Sebangar Kabupaten Bengkalis sebagai obat (Irawan et al. 2013). Tanaman ini sering ditemui di lingkungan masyarakat karena mudah memperbanyak diri melalui perkembangbiakan vegetatif.
Berdasarkan hasil identifikasi spesies
tumbuhan, dari 38 famili yang dimanfaatkan
Suku Mandailing sebagai obat, terdapat 13
famili (31,71%) dari total 41 famili yang
tercakup dalam hasil inventarisasi
tumbuhan yang dilakukan Kartawinata et al.
(2004) di kawasan TNBG. Famili tersebut
meliputi Anacardiaceae, Arecaceae,
Clusiaceae, Compositae, Euphorbiaceae,
Icacinaceae, Lauraceae, Leguminosae,
Melastomataceae, Meliaceae, Moraceae,
Myrtaceae serta Thymelaeaceae.
Hasil ini menunjukkan setidaknya ada 13 famili tumbuhan obat yang diambil masyarakat di hutan TNBG. Meskipun sebagian tumbuhan obat diperoleh dari hutan, namun bukan menjadi ancaman
utama terhadap kelestarian tumbuhan di hutan karena penyebab utama kerusakan hutan adalah penebangan pohon untuk industri kayu (Dick 1991). Kategori penyakit dan jumlah tumbuhan obat
Berdasarkan hasil wawancara, terdapat 41 jenis penyakit yang dapat diobati menggunakan spesies tumbuhan. Jenis-jenis penyakit yang dapat diobati berturut-turut adalah: sakit perut (14 spesies), demam (10 spesies), panas dalam (9 spesies), luka (9 spesies), masuk angin (8 spesies), diabetes (7 spesies), malaria (6 spesies), mata (6 spesies), pengobatan pasca persalinan (5 spesies), gatal-gatal (4 spesies), penyakit kulit (4 spesies), batuk (4 spesies), tekanan darah tinggi (4 spesies), diare (4 spesies), bisul (4 spesies), keropos tulang (3 spesies), terkena racun serangga (3 spesies), digigit serangga (3 spesies), sakit gigi (2 spesies), kurang nafsu makan (2 spesies), asam lambung (2 spesies), rematik (2 spesies), sakit kepala (2 spesies), pencahar/laksatif (membantu mengatasi sembelit) (2 spesies), mabuk jengkol (2 spesies), maag (2 spesies), asam urat (2 spesies), pingsan (2 spesies), pundial (2 spesies), bintua/perut kembung (2 spesies), kurang darah (1 spesies), benjol (1 spesies), cacingan (1 spesies), keracunan bisa ular (1 spesies), kurang gizi (1 spesies), susah buang air kecil (1 spesies), sesak nafas (1 spesies), jerawat (1 spesies), kanker (1 spesies), penambah stamina (1 spesies) serta terkilir (1 spesies) (Gambar 2). Berdasarkan Gambar 2 dapat diketahui bahwa Suku Mandailing memiliki

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
68
Gambar 3. Jumlah spesies berdasarkan bagian
tumbuhan yang digunakan sebagai obat
lebih banyak pengetahuan mengenai pengobatan terhadap penyakit yang sering dialami seperti sakit perut, demam maupun luka dibandingkan dengan penyakit yang jarang dialami. Hal ini tampak dari banyaknya spesies tumbuhan yang dapat digunakan untuk mengobati penyakit-penyakit tersebut. Luka sering dialami ketika masyarakat melakukan aktivitas di hutan maupun lingkungan pertanian. Pengetahuan terkait pengobatan terhadap luka lebih banyak dibandingkan penyakit lain.
Kategori organ tumbuhan yang digunakan Berdasarkan organ tumbuhan yang
digunakan Suku Mandailing untuk pengobatan, dapat dibedakan bagian: akarbatang, daun, buah, biji, rimpang serta kulit buah dan kulit batang. Organ tumbuhan yang paling sering digunakan adalah daun sebanyak 52 spesies, akar 8 spesies, buah 6 spesies, rhizoma 5 spesies, kulit batang 6 spesies, kulit buah 4 spesies, biji 3 spesies, getah batang 2 spesies serta getah buah 1 spesies (Gambar 3).
Organ yang paling banyak digunakan sebagai obat pada Suku Mandailing adalah daun. Hasil penelitian ini relevan dengan hasil penelitian sebelumnya yang dilakukan oleh Irawan et al. (2013) yang mengungkapkan bahwa daun merupakan organ yang paling dominan digunakan masyarakat Desa Sebangar Duri Tiga Belas dan Desa Kesumbo Ampai Duri Kabupaten Bengkalis. Organ daun paling banyak digunakan karena mudah diperoleh masyarakat serta mudah diolah karena teksturnya yang lunak. Selain itu, daun memiliki berbagai senyawa kimia
Gambar 4. Jumlah jenis penyakit yang diobati
berdasarkan cara meramu obat Keterangan:
a. bahan direbus lalu diminum/dimakan, b. bahan dioleskan/digosok/ditempel/dikompres, c. langsung diminum/dimakan, d. bahan dihaluskan/diperas lalu ditempel/gosok/usap, e. bahan dihaluskan/diperas lalu dimakan/
diminum, f. bahan dibakar/diuapkan, g. bahan diremas/dihaluskan lalu dimandikan, h. bahan direbus, lalu dioleskan/dimandikan/
dicelup, i. bahan dimandikan, j. bahan dikeringkan dibuat teh,
k. bahan ditumbuk diikat ke leher, l. bahan direbus lalu disemburkan
itu, daun memiliki berbagai senyawa kimia berkhasiat obat.
Penggunaan daun sebagai obat tradisional berdampak positif bagi kelestarian tumbuhan dibandingkan dengan penggunaan akar karena jika hanya organ daun yang diambil maka tidak akan menyebabkan berkurangnya spesies tumbuhan tertentu, sedangkan jika bagian yang diambil adalah akar tumbuhan maka spesies tumbuhan tersebut akan berkurang jika tidak ada upaya budidaya. Kategori berdasarkan cara meramu
Untuk mengatasi berbagai permasalahan penyakit, Suku Mandailing umumnya menggunakan bahan tumbuhan tunggal, namun ada beberapa penyakit yang menggunakan ramuan dengan menggunakan beberapa spesies tumbuhan. Bahan lain yang sering ditambahkan ke dalam ramuan obat yaitu garam, belerang, minyak goreng, telur ayam kampung serta gula merah.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
69
Gambar 5. Persentase spesies tumbuhan obat
berdasarkan habitus
Cara pengolahan tumbuhan yang dilakukan oleh Suku Mandailing dalam menyiapkan obat yaitu bahan tumbuhan tersebut direbus kemudian diminum air rebusannya, langsung diminum/dimakan, diperas/dihaluskan lalu ditempel, diperas lalu dimandikan, digosok/dioleskan ke bagian yang sakit, dibakar/diuapkan, dihaluskan lalu dimakan, bahan tumbuhan dimandikan, dibakar lalu diikat daun, dikeringkan lalu dibuat teh, ditumbuk lalu diikat ke leher, serta bahan direbus lalu disemburkan ke orang sakit ( Gambar 4). Kategori berdasarkan habitus
Hasil wawancara dan eksplorasi di hutan dan lingkungan Mandailing menunjukkan bahwa habitus tumbuhan yang digunakan sebagai obat adalah pohon, perdu dan herba. Habitus herba dominan digunakan masyarakat sebagai tumbuhan obat yaitu 50 spesies tumbuhan, sedangkan habitus pohon sebanyak 21 spesies tumbuhan. Perdu merupakan tipe habitus yang paling sedikit digunakan masyarakat Mandailing yaitu sebanyak 6 spesies tumbuhan (Gambar 5).
Tipe habitus herba paling banyak digunakan masyarakat karena mudah diperoleh. Selain itu, pemanfaatan tumbuhan dengan habitus tipe herba memberikan dampak positif bagi konservasi hutan dan tumbuhan jika dibandingkan dengan pemanfaatan tumbuhan dengan tipe pohon karena umumnya herba memiliki laju pertumbuhan dan perkembangbiakan yang cepat dan mudah tumbuh di berbagai lokasi. Tumbuhan dengan habitus tipe herba seperti
Gambar 6. Hasil uji fitokimia pada ekstrak (a). daun galunggung (Blumea balsamifera), (b). akar aren/bargot (Arenga pinnata), dan (c). daun galah nagori (Eupatorium sp.)
babadotan (Ageratum conyzoides)
mempunyai daya adaptasi yang tinggi terhadap lingkungannya sehingga mudah tumbuh di berbagai lokasi (Sukamto 2007). Resiko menurunnya populasi spesies tumbuhan dari tipe habitus berupa herba cenderung lebih rendah jika dibandingkan dengan pemanfaatan tumbuhan dengan habitus berupa pohon meskipun tumbuhan tersebut sering diambil untuk dimanfaatkan.
Pengetahuan Suku Mandailing terkait khasiat tumbuhan obat dalam mengatasi penyakit antara lain dapat dibuktikan secara ilmiah melalui identifikasi senyawa metabolit

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
70
sekunder menggunakan teknik analisis visualisasi warna. Berdasarkan hasil penelitian ini diketahui bahwa ekstrak daun tumbuhan galunggung (Blumea balsamifera) positif mengandung flavonoid, tannin, saponin dan steroid. Daun tumbuhan galah nagori (Eupatorium sp.) positif mengandung flavonoid, tannin, saponin, steroid dan triterpenoid. Akar tumbuhan bargot (Arenga pinnata) positif mengandung flavonoid dan saponin. Alkaloid dan quinon menunjukkan hasil negatif pada semua sampel yang diuji (Gambar 6). Hasil uji warna secara lengkap dapat dilihat pada Tabel 1. Galunggung (Blumea balsamifera), galah nagori (Eupatorium sp.) dan bargot (Arenga pinnata) dipilih untuk dianalisis secara fitokimia karena jarang digunakan masyarakat suku lain di Indonesia untuk mengobati penyakit. Penelitian sebelumnya terkait kandungan metabolit sekunder pada ketiga spesies ini juga belum dilakukan sehingga penting untuk melakukan analisis fitokimia pada ketiga spesies ini. Daun
galunggung (Blumea balsamifera) dan daun galah nagori (Eupatorium sp.) banyak
digunakan masyarakat untuk mengobati luka, sedangkan akar bargot/aren (Arenga pinnata) digunakan Suku Mandailing untuk
mengobati keropos tulang. Luka adalah diskontinuitas jaringan
(Barbul dan Efron 2010). Luka tidak dapat dibiarkan sembuh sendiri karena luka yang tidak dirawat dapat menyebabkan komplikasi seperti infeksi dan pendarahan (Syarfati et al. 2011).
Flavonoid, tannin serta saponin dapat membantu proses penyembuhan luka karena berfungsi sebagai antioksidan dan antimikroba yang mempengaruhi penyembuhan luka dan mempercepat epitelisasi (Senthil et al. 2011). Epitelisasi (pembaharuan epitel setelah terjadinya luka), melibatkan proliferasi dan migrasi sel epitel menuju pusat luka dan kontraksi luka disebabkan oleh aksi miofibroblas. Flavonoid telah dibuktikan dapat meningkatkan migrasi dan proliferasi sel epitel, pembentukan
Tabel 1. Hasil identifikasi senyawa kimia ekstrak daun galunggung (Blumea balsamifera), akar bargot (Arenga pinnata) serta daun galah nagori (Eupatorium sp.) dengan teknik visualisasi warna
No. Nama sampel Identitas sampel Senyawa kimia Hasil
1. Galunggung (Blumea balsamifera)
Padatan
Flavonoid Positif
Alkaloid Negatif
Tannin Positif
Saponin Positif
Quinon Negatif
Steroid Positif
Triterpenoid Negatif
2. Bargot (Arenga pinnata)
Padatan
Flavonoid Positif
Alkaloid Negatif
Tannin Negatif
Saponin Positif
Quinon Negatif
Steroid Negatif
Triterpenoid Negatif
3. Galah Nagori (Eupatorium sp)
Padatan
Flavonoid Positif
Alkaloid Negatif
Tannin Positif
Saponin Positif
Quinon Negatif
Steroid Positif
Triterpenoid Positif

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
71
jaringan granulasi, serta meningkatkan migrasi dan aktivitas miofibroblas (Muralidhar et al. 2013). Aktivitas antioksidan yang tinggi pada flavonoid dapat mempercepat penyembuhan luka karena dapat menstimulasi produksi antioksidan endogen pada situs luka dan menyediakan lingkungan yang kondusif untuk terjadinya penyembuhan luka (Ahmed et al. 2012).
Tannin berguna sebagai astringen (menghentikan perdarahan), mempercepat penyembuhan luka dan inflamasi membran mukosa, serta regenerasi jaringan baru (Reddy et al. 2011). Tannin mempercepat penyembuhan luka dengan beberapa mekanisme seluler yaitu membersihkan radikal bebas dan oksigen reaktif, meningkatkan penutupan luka serta meningkatkan pembentukan pembuluh darah kapiler juga fibroblas. Selain itu kandungan tannin mempunyai kemampuan antioksidan dan antibakteri (Lai 2011). Tannin bekerja dengan cara mengendapkan protein dan dapat merusak membran sel sehingga pertumbuhan jamur terhambat. Senyawa tannin merupakan senyawa organik yang aktif menghambat pertumbuhan mikroba dengan mekanisme merusak dinding sel mikroba dan membentuk ikatan dengan protein fungsional sel mikroba. Mekanisme antibakteri yang dimiliki tannin yaitu kemampuannya menghambat sintesis khitin yang digunakan untuk pembentukan dinding sel pada jamur dan merusak membran sel sehingga pertumbuhan jamur terhambat. Tannin juga merupakan senyawa yang bersifat lipofilik sehingga mudah terikat pada dinding sel dan mengakibatkan kerusakan dinding sel (Sudira et al. 2011).
Saponin berpotensi membantu menyembuhkan luka dengan membentuk kolagen pertama yang mempunyai peran dalam proses penyembuhan luka (Astuti et al. 2011). Saponin juga berperan sebagai antibakteri. Mekanisme kerja saponin sebagai antibakteri yaitu dapat menyebabkan kebocoran protein dan enzim dari dalam sel. Saponin dapat menjadi antibakteri karena zat aktif permukaannya mirip detergen, akibatnya saponin akan menurunkan tegangan permukaan dinding sel bakteri dan merusak permeabilitas membran (Madduluri et al. 2013). Rusaknya membran sel ini sangat mengganggu
kelangsungan hidup bakteri. Terpenoid juga diketahui memegang
peranan penting dalam meningkatkan proses penyembuhan luka karena terpenoid diketahui mempunyai efek antimikroba dan antioksidan yang kuat sehingga diduga bertanggungjawab dalam kontraksi luka dan peningkatan kecepatan dari epitelisasi (Saroja dan Annapoorani 2012). Terpenoid mempunyai mekanisme antibakteri dengan cara pengrusakan membran sel bakteri. Kerusakan membran sel dapat terjadi ketika senyawa aktif antibakteri bereaksi dengan sisi aktif dari membran atau dengan melarutkan konstituen lipid dan meningkatkan permeabilitasnya. Membran sel bakteri terdiri dari fosfolipid dan molekul protein. Adanya peningkatan permeabilitas maka senyawa antibakteri dapat masuk ke dalam sel dan dapat melisis membran sel atau mengkoagulasi sitoplasma dari sel bakteri tersebut (Mayanti et al. 2011).
Selain menyembuhkan luka, flavonoid juga berperan dalam mencegah keropos tulang (Sirait 2007). Fitoestrogen merupakan salah satu jenis flavonoid (isoflavon) yang memiliki kemiripan aktivitas dengan estrogen (Urasopon et al. 2008). Fitoestrogen dianggap sebagai zat alternatif yang efektif dalam mencegah keropos tulang yang disebabkan oleh defisiensi estrogen. Flavonoid tersebut dapat menstimulasi proliferasi/perbanyakan sel osteoblast yang berperan dalam pembentukan tulang karena mempunyai aktivitas estrogenik.
Dari uraian diatas diperoleh kesimpulan bahwa Suku Mandailing memiliki pengetahuan terkait pemanfaatan tumbuhan obat mulai dari spesies tumbuhan yang digunakan, penyakit yang dapat diobati, organ tumbuhan yang digunakan untuk pengobatan hingga cara meramu tumbuhan tersebut. Tingginya keanekaragaman tumbuhan obat membuktikan bahwa pemanfaatan tumbuhan untuk kesehatan menjadi prioritas utama suatu kampung yang berlokasi di dekat hutan (Zuhud 2011). Uraian diatas juga menunjukkan bahwa pengetahuan masyarakat mengenai tumbuhan berkhasiat obat dapat dibuktikan secara ilmiah sehingga dapat memberikan kontribusi bagi pengembangan obat tradisional baik obat herbal terstandar atau fitofarmaka, dimana obat herbal terstandar adalah obat tradisional yang sudah

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
72
dilakukan uji preklinik yang meliputi uji efikasi dan uji toksisitas, sedangkan fitofarmaka adalah obat herbal terstandar yang sudah dilakukan uji klinik terbatas.
Identifikasi keanekaragaman spesies
tumbuhan obat pada Suku Mandailing
sangat penting dilakukan sebagai dasar
pengembangan obat tradisional tersebut
diatas. Namun informasi ini juga dapat
memicu terjadinya ekploitasi yang dapat
mengakibatkan penurunan populasi
tumbuhan tersebut. Padahal tingkat
pengetahuan dan pemanfaatan tumbuhan
akan mengalami penurunan seiring dengan
penurunan keanekaragaman spesies
tumbuhan di kawasan tersebut. Oleh karena
itu masyarakat harus memanfaatkan
tumbuhan secara arif dan bijaksana,
sehingga dapat memberikan manfaat
optimal bagi mereka. Untuk menghindari
eksploitasi antara lain dapat dilakukan
domestikasi tanaman obat terpilih untuk
dibudidayakan secara massal.
Pengetahuan mengenai tumbuhan
obat juga harus dijaga, dilestarikan serta
diwariskan kepada generasi muda agar
pengetahuan tersebut tidak hilang. Hal ini
sejalan dengan hasil penelitian Van Sam et
al. (2008) serta Kartawinata (2010) yang
menunjukkan bahwa laju kehilangan spesies
sejajar dengan laju kehilangan pengetahuan
tradisional.
Hasil penelitian ini merupakan tahap awal dari penelitian tentang tumbuhan obat. Oleh karena itu, perlu penelitian lanjutan mengenai aktifitas mikroba dan aktifitas senyawa kimia yang terkandung dalam tumbuhan obat pada Suku Mandailing untuk mengamati efikasinya pada berbagai penyakit sehingga bermanfaat bagi pengembangan obat tradisional yang bisa dipertanggungjawabkan secara ilmiah.
KESIMPULAN
Tumbuhan yang digunakan dalam pengobatan tradisional pada Suku Mandailing berjumlah 81 spesies yang tercakup dalam 38 famili. Spesies yang paling banyak adalah Compositae (11 spesies). Kategori penyakit yang dapat diobati sebanyak 41 jenis. Kategori penyakit yang dapat diobati menggunakan tumbuhan
obat adalah: sakit perut (14 spesies), demam (10 spesies), panas dalam (9 spesies), luka (9 spesies), masuk angin (8 spesies) serta untuk pengobatan lainnya menggunakan antara 1 s/d 7 spesies tumbuhan. Pemanfaatan tumbuhan berkhasiat obat oleh Suku Mandailing menunjukkan bahwa mereka telah memiliki pengetahuan terkait pemanfaatan tumbuhan berkhasiat obat secara empiris. Tingginya keanekaragaman tumbuhan obat membuktikan bahwa kesehatan menjadi prioritas utama Suku Mandailing yang tinggal di sekitar hutan.
UCAPAN TERIMAKASIH
Penelitian ini didanai oleh Direktorat Jendral Pendidikan Tinggi Kementerian Riset dan Teknologi Pendidikan Tinggi melalui Beasiswa Program Pascasarjana Dalam Negeri (BPPDN).
DAFTAR PUSTAKA
Ahmed KA, Abdulla MA, Mahmoud FM (2012) Wound healing potential of Phyllanthus niruri leaf extract in
experimental rats. Middle-East J Sci Res 11: 1614-1618. doi: 10.5829/idosi.mejsr.2012.11.11.1590
Anderson EN, Pearsall D, Hunn E, Turner N (2011) Ethnobiology. ISBN: 978-0-470-54785-4. Willey-Blackwell, New Jersey
Astuti SM, Sakinah M, Andayani R, Risch A (2011) Determination of saponin compound from Anredera cordifolia (Ten) Steenis plant (binahong) to potential treatment for several diseases. J Agric Sci 3: 224-232. doi: 10.5539/jas.v3n4p224
Balitbangkes (2013) Riset Kesehatan Dasar (Riskesdas 2013). Badan Penelitian dan Pengembangan Kesehatan, Kementerian Kesehatan RI, Jakarta.
Barbul A, Efron D (2010) Wound healing In: Schwartz’s Principles of Surgery 9th ed. Pp 210-219. Brunicardi FC, Andersen DK, Billiar TR, Dunn DL, Hunter JG, Matthews JB, Pollock RE (Eds). McGraw-Hill, New York
BKSDA II Sumatera Utara (2005) Rencana pengelolaan Taman Nasional Batang Gadis Kabupaten Madina Provinsi Sumatera Utara. Departemen

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
73
Kehutanan, Medan Dick J (1991) Forest land use, forest use
zonation, and deforestation in Indonesia: A summary and interpretation of existing information. The Ministry of State for Population and Environment (KLH) and the Environmental Impact Management Agency (BAPEDAL), Jakarta
Harborne JB (1987) Metode Fitokimia. Terjemahan dari Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis, oleh Padmawinata K, Soediro I. Penerbit ITB, Bandung
Indriati G (2014) Etnobotani tumbuhan obat yang digunakan Suku Anak Dalam di Desa Tabun Kecamatan VII Koto Kabupaten Tebo Jambi. J Sainstek 6: 52-56
Irawan YR, Fitmawati, Herman (2013) Pengetahuan tumbuhan obat Dukun Sakai Desa Sebangar Duri Tiga Belas dan Desa Kesumbo Ampai Duri Kabupaten Bengkalis. Jurnal Biosaintifika 5: 30-35. doi: 10.15294/biosaintifika.v5i1.2571
Kartawinata K (2010) Dua abad mengungkap kekayaan flora dan ekosistem Indonesia. Bidang
Lingkungan, Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI), Jakarta
Kartawinata K, Samsoedin I, Heriyanto M, Afriastini JJ (2004) A tree species inventory in a one-hectare plot at the Batang Gadis National Park, North Sumatera, Indonesia. Reinwardtia 12: 145-157. doi: http://dx.doi.org/10.14203/reinwardtia.v12i2.60
Lai HY, Lim YY, Kim KH (2011) Potential dermal wound healing agent in Blechnum orientale Linn. BMC
Complement Altern Med 11: 62. doi: 10.1186/1472-6882-11-62
Madduluri S, Rao KB, Sitaram B (2013) In vitro evaluation of antibacterial activity of five indigenous plants extracts against five bacteria pathogens of human. Int J Pharm Pharm Sci 5: 679-684
Mayanti T, Julaeha E, Putri Y (2011) Isolasi dan karakterisasi senyawa antibakteri dari fraksi etil asetat kulit batang Lansium domesticum Corr. cv
kokossan. Pp 1-11. Fakultas MIPA, Universitas Padjajaran, Bandung
Muralidhar A, Babu KS, Sankar TR, Reddanna P, Latha J (2013) Wound healing activity of flavonoid fraction isolated from the stem bark of Butea monosperma (Lam) in albino wistar rats. Eur J Exp Biol 3: 1-6
Reddy BK, Gowda S, Arora AK (2011) Study of wound healing activity of aqueous and alcoholic bark extracts of Acacia catechu on rats. J Pharm Sci 1: 220-
225. doi: 10.5530/rjps.2011.3.8 Saroja M, Annapoorani S (2012) In vitro
antioxidant activity of flavonoid fractions of Cynodon dactylon and Terminalia catappaleaves. Int Res J Pharm 3: 209-211
Senthil P, Kumar AA, Manasa M, Kumar KA, Sravanthi K, Deepa D (2011) Wound healing activity of alcoholic extract of Guazuma ulmifolia leaves on albino
Wistar rats. Int J Pharm Bio Sci 2: 34-38 Shanthi RV, Jumari, Izzati M (2014) Studi
etnobotani pengobatan tradisional untuk perawatan wanita di masyarakat Keraton Surakarta Hadiningrat. Biosaintifika 6: 85-93. doi: 10.15294/biosaintifika.v6i2.3101
Sirait M (2007) Penuntun Fitokimia dalam Farmasi. ISBN: 979-3507-99-3. Penerbit ITB, Bandung
Sudira IW, Merdana IM, Wibawa IPAH (2011) Uji daya hambat ekstrak daun kedondong (Lannea grandis Engl) terhadap pertumbuhan bakteri Erwinia carotovora. Buletin Veteriner Udayana 3: 45-50
Sukamto (2007) Babadotan (Ageratum conyzoides) tanaman multi
fungsi. Warta Puslitbangbun 13(3). Syarfati, Eriani K, Damhoeri A (2011) The
potential of jarak cina (Jatropha multifida L.) secretion in healing new-wounded mice. J Natural 11: 16-19
Urasopon N, Hamada Y, Cherdshewasart W, Malaivijitnond S (2008) Preventive effects of Pueraria mirifica on bone
loss in ovariectomized rats. Maturitas 59: 137–148. doi: 10.1016/j.maturitas.2008.01.001
Van Sam H, Baas P, Kessler PJA (2008) Traditional medicinalplants in Ben En National Park, Vietnam. Blumea 53: 569-601. doi: 10.3767/000651908X607521

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
74
Walujo EB (2009) Etnobotani: Memfasilitasi penghayatan, pemutakhiran pengetahuan dan kearifan lokal dengan menggunakan prinsip-prinsip dasar ilmu pengetahuan. Prosiding Seminar Etnobotani IV: Keanekaragaman Hayati, Budaya dan Ilmu Pengetahuan. Dalam Purwanto Y,
Waluyo EB (Ed.) 12-19. Pusat Penelitian Biologi. LIPI Press, Jakarta
Yuniati E, Alwi M (2010) Etnobotani keanekaragaman jenis tumbuhan obat tradisional dari hutan di Desa Pakuli Kecamatan Gumbasa Kabupaten Donggala, Sulawesi Tengah. Biocelebes 4: 69-75
Zuhud EAM (2011) Pengembangan desa
konservasi hutan keanekaragaman hayati untuk mendukung kedaulatan pangan dan obat keluarga (POGA) Indonesia dalam menghadapi ancaman krisis baru ekonomi dunia di era globalisasi. Orasi Ilmiah Guru Besar 19 November 2011. IPB, Bogor
Zuhud EAM, Hikmat A (2009) Hutan tropika Indonesia sebagai gudang obat bahan alam bagi kesehatan mandiri bangsa. Bunga Rampai Biofarmaka Kehutanan Indonesia: Dari tumbuhan hutan untuk keunggulan bangsa dan negara. Hal. 17-28. Pusat Penelitian dan Pengembangan Hutan Tanaman. Balitbang Kehutanan. Kementerian Kehutanan, Bogor

75
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
MULTIPLIKASI IN VITRO ANGGREK HITAM (Coelogyne pandurata Lindl.)
PADA PERLAKUAN KOMBINASI NAA DAN BAP
In Vitro Multiplication of Black Orchid (Coelogyne pandurata Lindl.) Using the Combination of NAA and BAP
Roni Kartiman
1, Dewi Sukma
2, Syarifah Iis Aisyah
2, Agus Purwito
2*,
1Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pascasarjana, IPB.
Jl. Raya Dramaga ,Bogor 16680 2Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor, Darmaga, Bogor 16680
*Email: [email protected]
ABSTRACT Black orchid is an indigenous plant from Kalimantan, Indonesia. It becomes endangered because of forest over-exploitation and its low natural reproduction rate. Tissue culture is considered to offer a solution to conserve and propagate this species. The aim of this research was to evaluate the effect of Naphtalene Acetic Acid (NAA) and 6-Benzile Amino Purine (BAP) on shoots multiplication of black orchid. The basic medium used was a half of Murashige & Skoog (MS) composition supplemented with 150 mLL
-1 coconut water. Initial
explants used were 6-month-old shoots of germinating seeds. The shoot cultures were incubated for 23 weeks. Results showed that the best combination for shoot multiplication was NAA 0.0 mgL
-1 with BAP 0.2 mgL
-1. Shoot grew better on medium with BAP and without
NAA while roots growth was better on medium without the two plant growth regulators. The addition of BAP up to 0.3 mgL
-1 increased the leaf number, which however decreased at
higher BAP concentration. Keywords: BAP, black orchid, Coelogyne pandurata, multiplication, NAA ABSTRAK Anggrek hitam merupakan flora langka asli Kalimantan, Indonesia. Keberadaa anggrek ini di alam semakin langka akibat eksploitasi berlebihan dan sulitnya perbanyakan secara alami. Kultur jaringan merupakan metode untuk mengatasi kelangkaan anggrek ini. Penelitian ini bertujuan untuk mengetahui pengaruh kombinasi NAA dan BAP terhadap multiplikasi anggrek hitam. Media dasar yang digunakan adalah ½ MS dengan penambahan air kelapa 150 mLL
-1. Eksplan yang digunakan adalah tunas hasil semai biji umur 6 bulan. Kultur tunas
diinkubasi selama 23 minggu. Hasil penelitian menunjukkan bahwa kombinasi terbaik untuk multiplikasi tunas adalah NAA 0 mgL
-1 dengan BAP 0,2 mgL
-1. Tunas tumbuh lebih baik
dalam media dengan penambahan BAP tanpa NAA, sedangkan akar pada media tanpa NAA dan BAP. Penambahan BAP sampai 0.3 mgL
-1 mampu meningkatkan jumlah daun,
namun menurun dengan penambahan di atas konsentrasi tersebut.
Kata Kunci: anggrek hitam, BAP, Coelogyne pandurata, multiplikasi, NAA
Received: 18 May 2018 Accepted: 21 June 2018 Published: 29 June 2018

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
76
PENDAHULUAN
Indonesia merupakan salah satu negara yang memiliki hutan tropis terbesar dan memiliki kekayaan spesies anggrek yang sangat beragam. Tanaman anggrek termasuk ke dalam anggota famili Orchidaceae. Famili ini terdiri atas 800 genus
dan tidak kurang dari 25.000 spesies. Dari jumlah tersebut diperkirakan 5.000 spesies diantaranya berasal dari Indonesia (Adi et al. 2014). Salah satu anggrek alam yang dianggap sebagai ciri khas Indonesia adalah anggrek hitam (Coelogyne pandurata Lindl.).
Anggrek hitam (Gambar 1) merupakan anggrek spesies asli Kalimantan. Nama anggrek hitam diberikan karena bunga anggrek ini memiliki tanda hitam pada bibirnya yang membentang sampai bagian dalam bunga, sedangkan mahkota bunga dan kelopak bunga anggrek hitam berwarna hijau cerah (Gravendeel 2000). Di habitat aslinya, anggrek ini berbunga pada bulan Mei-Juni, dengan bunganya yang berbau harum lembut dan lama mekar bunga sekitar 5-7 hari (Untari 2003).
Populasi anggrek hitam di habitat aslinya hampir punah akibat pengambilan yang berlebihan, kerusakan hutan dan konversi lahan. Selain itu juga periode
berbunga anggrek ini sangat pendek dan sulit untuk disilangkan (Untari 2003).
Perbanyakan anggrek hitam secara konvensional melalui stek batang, pembelahan rumpun, penggunaan pseudobulb atau pemisahan anakan sulit
dilakukan karena keterbatasan tanaman induk di alam (Adi et al. 2014). Demikian juga perkembangbiakan alami anggrek hitam dengan biji di habitat aslinya sangat sulit karena lambatnya pertumbuhan biji yang harus bersimbiosis dengan mikoriza yang ada dan cocok, sehingga laju pertumbuhan dari fase biji sampai tanaman dewasa yang siap berbunga sangat lama (Untari 2003). Perbanyakan konvensional yang sulit dilakukan ini dapat diatasi dengan teknik kultur jaringan (in vitro) sebagai salah satu
usaha konservasi mencegah untuk kepunahan anggrek hitam.
Dalam kultur jaringan anggrek ada beberapa tahapan yang dilalui yaitu sterilisasi bahan tanam (eksplan berupa biji maupun bagian vegetatif tanaman), pengecambahan benih (dalam media hingga membentuk protocorm atau PLBs (Protocorm Likes Bodies), multiplikasi dan regenerasi planlet, pengakaran dan aklimatisasi. Multiplikasi tunas merupakan tahapan yang sangat penting dalam
Gambar 1. Bunga anggrek hitam (C. pandurata Lindl.)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
77
perbanyakan anggrek secara kultur jaringan. Media Murashige dan Skoog (MS) merupakan media yang banyak digunakan dalam kultur in vitro. Teknik kultur jaringan dapat menghasilkan tanaman-tanaman baru anggrek hitam yang secara genetik identik dengan induknya, dalam jumlah banyak dan tidak tergantung musim. Penelitian kultur jaringan anggrek telah banyak dilakukan. Putri (2015) menyatakan bahwa media ½ MS dengan penambahan air kelapa 15% sudah efektif dalam pertumbuhan dan multiplikasi clumps PLBs Phalaenopsis
hibrida. Media ½ MS dengan penambahan 30 g L-1 sukrosa sangat efektif menginduksi kalus dari PLBs anggrek Phalaenopsis var.
Hawaiian Clouds x Carmela’s Dream (Ling et al. 2007). Penelitian Untari (2003) menunjukkan bahwa media dasar VW (Vacint dan Went) dengan penambahan ekstrak pisang ambon 150 g L-1 dan NAA 20 ppm mampu memacu dan menghasilkan rata-rata jumlah tunas tertinggi pada kultur jaringan Coelogyne pandurata Lindl.
Zat pengatur tumbuh (ZPT) auksin dan sitokinin dapat digunakan untuk memacu pertumbuhan. Zat pengatur tumbuh 1-Naphtalene Acetic Acid (NAA) merupakan salah satu ZPT golongan auksin, 6-Benzile Amino Purine (BAP) merupakan golongan
sitokinin. Zat pengatur tumbuh ini memiliki kisaran luas dalam memacu suatu pertumbuhan sehingga kisaran konsentrasi NAA dan BAP yang digunakan tidak menyebabkan terhambatnya pertumbuhan tanaman. NAA pada konsentrasi tertentu berfungsi sebagai inisiasi akar dan batang tanaman, sedangkan BAP berfungsi untuk memacu inisiasi tunas (Restiani et al. 2016). Zat pengatur tumbuh jenis NAA dan BAP mudah di dapat di pasaran dan relatif murah dibandingkan jenis auksin dan sitoninin
lainnya. Tujuan penelitian ini adalah untuk mengetahui pengaruh kombinasi NAA dan BAP pada berbagai konsentrasi terhadap kemampuan multiplikasi tunas dari eksplan C. pandurata Lindl., sehingga diharapkan
diperoleh konsentrasi optimum yang dapat digunakan secara luas untuk perbanyakan anggrek C. pandurata Lindl.
BAHAN DAN METODE Waktu dan tempat penelitian
Penelitian telah dilaksanakan selama 10 bulan (Juli 2016 - April 2017) di Laboratorium Kultur Jaringan 1, Laboratorium Mikroteknik dan Laboratorium Pasca Panen, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor, Dramaga, Bogor, Propinsi Jawa Barat. Bahan dan alat
Bahan tanaman yang digunakan adalah tunas in vitro anggrek C. pandurata
Lindl. yang berumur 6 bulan setelah semai biji, media ½ MS, zat pengatur tumbuh NAA dan BAP, air kelapa muda umur simpan 1 hari di dalam lemari pendingin, alkohol, aquades, spirtus, agar dan gula pasir.
Alat yang digunakan berupa alat-alat laboratorium dan alat-alat tanam yaitu laminar air flow cabinet (LAFC), autoklaf, timbangan analitik, labu Erlenmeyer, gelas
ukur, gelas piala, pH meter, cawan Petri, pipet, pengaduk, jarum injeksi, pinset, scalpel, lampu spirtus, hand sprayer, kompor gas, oven, korek api, alumunium foil, rak
kultur dan botol media. Rancangan percobaan
Penelitian ini menggunakan Rancangan Acak Kelompok (RAK) 2 faktor. Faktor pertama adalah NAA yang terdiri dari
Tabel 1. Kombinasi perlakuan konsentrasi NAA dan BAP
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
N0B0 N1B0 N2B0
0,1 mgL-1
N0B1 N1B1 N2B1
0,2 mgL-1
N0B2 N1B2 N2B2
0,3 mgL-1
N0B3 N1B3 N2B3
0,4 mgL-1
N0B4 N1B4 N2B4
0,5 mgL-1
N0B5 N1B5 N2B5

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
78
3 taraf konsentrasi, yaitu 0,0 (N0); 0,2 (N1) dan 0,5 (N2) mgL-1. Sedangkan faktor kedua adalah BAP yang terdiri dari 5 taraf konsentrasi, yaitu 0,0 (B0); 0,1 (B1); 0,2 (B2); 0,3 (B3); 0,4 (B4) dan 0,5 (B5) mgL-1 sehingga diperoleh 18 kombinasi perlakuan (Tabel 1). Masing-masing perlakuan dilakukan pengulangan sebanyak 4 kali, setiap ulangan terdiri atas 3 botol kultur yang di dalamnya ada 3 tanaman. Sterilisasi
Alat-alat yang digunakan ketika
penanaman, harus dalam keadaan steril.
Alat-alat logam dan gelas disterilkan dalam
oven. Alat-alat tersebut dibungkus dengan
kertas tebal (kertas merang atau kertas kopi)
kemudian dimasukkan ke dalam oven
selama 24 jam dengan suhu 60ºC. Sterilisasi
botol kultur dilakukan dengan pencucian
terlebih dahulu menggunakan deterjen
kemudian dibilas dengan air bersih. Botol-
botol yang sudah bersih dikeringanginkan
dulu beberapa saat sampai airnya kering,
kemudian dimasukan ke dalam autoklaf
pada suhu 121ºC selama 30 menit. Alat-alat
seperti pinset dan scalpel dapat disterilisasi
kembali dengan pemanasan di atas api
spirtus, setelah dicelupkan ke dalam alkohol
96% sebelum dilakukan penanaman.
Media dan aquades disterilisasi dalam
autoklaf sebelum digunakan. Air aquades
disterilisasi dalam botol selai sebanyak 75
mL ditutup dengan tutup botol plastik anti
panas, kemudian dimasukan ke dalam
autoklaf selama 30 menit pada suhu 121ºC.
Media kultur disterilisasi dengan cara yang
sama seperti sterilisasi air aquades.
Kebersihan lingkungan khusus berupa
LAFC dilakukan dengan menyemprot
permukaan tempat kerja dengan alkohol
70%, kemudian dilap dengan tissue. Blower
(peniup udara) dan lampu ultra violet dalam
LAFC dinyalakan selama 15-60 menit
sebelum digunakan dengan tujuan untuk
membersihkan kontaminan di permukaan
tempat kerja. Sehabis kerja, permukaan
tempat kerja dibersihkan kembali dengan
alkohol 70% atau dengan lampu ultra violet
selama 30-60 menit.
Pembuatan media
Langkah awal yang dilakukan dalam
pembuatan media adalah membuat larutan stok yang terdiri atas larutan stok makro NH4NO3, KNO3, CaCl2·2H2O, MgSO4·7H2O dan KH2PO4; larutan stok mikro H3BO3, MnSO4·H2O, ZnSO4·7H2O, KI, Na2MoO4·2H2O, CaCl2·6H2O dan CuSO4·5H2O; larutan stok FE-EDTA yang berasal dari campuran Na2-EDTA (C10H14N2Na2O8·2H2O) dan FeSO4·7H2O serta larutan stok vitamin dan larutan stok NAA dan BAP. Untuk media MS penuh masing-masing stok diambil 10 mLL-1.
Pembuatan media ½ MS dilakukan dengan memasukkan seluruh larutan stok baik makro, mikro, vitamin dan FE-EDTA masing-masing 5 mLL-1. Selanjutnya media ½ MS ini ditambah gula pasir 30 g L-1, agar-agar 8 g L-1 dan air kelapa 150 mLL-1, serta ZPT sesuai dengan kombinasi perlakuan yang telah ditentukan (Tabel 1), media kemudian ditera menjadi satu liter dengan penambahan aquades.
Tingkat kemasaman (pH) larutan diukur dengan menggunakan pH meter. pH diatur hingga mencapai 5,6-5,8 dengan penambahan KOH bila pH<5,6 dan HCl bila pH>5,8. Larutan media selanjutnya dipanaskan sampai larut dan mendidih. Media yang telah mendidih dituangkan ke dalam botol kira-kira sebanyak 30 mL, dan ditutup dengan tutup plastik tahan panas. Media disterilisasi menggunakan autoklaf pada suhu 121ºC selama 30 menit.
Penanaman
Rumpun-rumpun eksplan anggrek hitam dipisah masing-masing menjadi 1 tanaman, kemudian ditanam pada botol media yang telah disiapkan sesuai dengan perlakuan. Setiap botol kultur yang digunakan ditanami tiga eksplan tanaman yang ukurannya seragam (8-20 mm). Proses penanaman dilakukan dalam LAFC.
Pengamatan
Pengamatan dilakukan setiap minggu sekali selama 23 minggu. Parameter/peubah yang diamati terdiri dari: tinggi tanaman (mm) (diukur dari permukaan media sampai ujung tertinggi tanaman), panjang daun maksimum (mm) (diukur dari pangkal hingga ujung daun), lebar daun maksimum (mm) (diukur pada daun terlebar), jumlah daun (buah) (dengan menghitung jumlah daun pada saat pengamatan), jumlah akar (buah)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
79
(dengan menghitung jumlah akar pada saat pengamatan), panjang akar terpanjang (mm) (diukur dari pangkal hingga ujung akar), jumlah PLBs (buah) yang terbentuk (dengan menghitung jumlah PLBs pada saat pengamatan), jumlah tunas baru yang terbentuk (buah) (dengan menghitung jumlah tunas pada saat pengamatan). Parameter-parameter diatas diukur dari luar botol dengan menggunakan penggaris, sedangkan eksplan yang diamati tetap berada di dalam botol.
Selain pengamatan parameter-parameter di atas, diamati juga saat pembentukan PLBs, akar dan tunas pertama serta anatomi stomata yang mengacu metode Gantait et al. (2011) dan kandungan kloroplas dengan metode Sims dan Gamon (2003). Uji anatomi stomata meliputi pengamatan jumlah stomata yang dilakukan pada dua bidang pandang, panjang dan lebar stomata diamati pada tiga bidang pandang yang dipilih secara acak, masing-masing seluas 0,19625 mm2. Pengamatatan stomata dilakukan menggunakan mikroskop Olympus tipe BX 51, kamera tipe DP 25 dengan perangkat lunak program Olympus DP2-BSW. Analisis data
Data dianalisis dengan Analysis of Variance (ANOVA), jika hasil yang didapat
berbeda nyata, dilanjutkan dengan Uji Duncan pada selang kepercayaan 95% (Pramesti 2011). HASIL DAN PEMBAHASAN
Hasil analisis ragam (ANOVA) terhadap respon pertumbuhan in vitro pada
23 minggu setelah tanam (MST) disajikan
pada Tabel 2. Hasil pengamatan menunjukkan bahwa NAA berpengaruh nyata terhadap semua peubah yang diamati, sedangkan BAP hanya berpengaruh nyata terhadap tinggi tanaman dan panjang daun. Interaksi NAA dan BAP berpengaruh nyata terhadap peubah tinggi tanaman, panjang daun, jumlah daun, panjang akar dan jumlah tunas. Pada semua perlakuan dalam percobaan ini, PLBs sama sekali tidak terbentuk sehingga tidak dapat dianalisis.
Tinggi tanaman
Tinggi tanaman pada kombinasi perlakuan NAA dan BAP disajikan pada Tabel 3. Tanaman tertinggi dihasilkan pada media ½ MS tanpa penambahan ZPT sebesar 39,335 mm namun tidak berbeda nyata dengan media tanpa NAA yang ditambahkan BAP 0,1 hingga 0,4 mgL-1. Hasil tersebut menunjukkan bahwa semua perlakuan tanpa NAA memacu tinggi tanaman. Konsentrasi auksin alami dalam tanaman (endogen) diduga sudah mencukupi untuk mendorong pertumbuhan tinggi tanaman. Pertambahan tinggi eksplan disebabkan oleh dua proses yaitu pembelahan dan pemanjangan sel, kedua proses ini terjadi pada jaringan meristem, yaitu pada titik tumbuh batang sehingga tanaman tumbuh semakin besar dan berkorelasi positif dalam menentukan hasil tanaman (Untari 2003). Menurut Untari (2003) pada anggrek hitam yang mendapat perlakuan media dasar VW tanpa penambahan bahan organik dan NAA memberikan rata-rata tinggi planlet terbaik.
Peningkatan konsentrasi NAA dan BAP cenderung menekan tinggi tanaman (Tabel 3). Menurut Untari (2003) bahwa batang dan akar yang sedang memanjang
Tabel 2. Rekapitulasi analisis ragam respon pertumbuhan vegetatif anggrek hitam pada 23 MST
No Peubah NAA BAP NAA * BAP
1 Tinggi tanaman ** ** *
2 Panjang daun ** ** **
3 Jumlah daun ** tn *
4 Lebar daun ** tn tn
5 Panjang akar ** tn **
6 Jumlah akar ** tn tn
7 Jumlah tunas ** tn *
Keterangan: tn : tidak berbeda nyata, *: berbeda nyata pada =5%, **: sangat berbeda nyata pada pada =1%

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
80
tidak memerlukan penambahan sitokinin, walaupun kedua organ tersebut dalam pertumbuhannya memerlukan hormon tersebut untuk proses pemanjangan sel, tetapi kandungan alami sitokinin dalam jaringan tanaman kemungkinan sudah mencukupi. Terkait dengan hal tersebut, kandungan auksin endogen dalam tanaman tunas-tunas in vitro anggrek hitam diduga
sudah cukup tinggi, sehingga penambahan auksin eksogen akan meningkatkan perimbangan total auksin dan sitokinin
dalam kultur dan menekan pertumbuhan tinggi tanaman.
Jumlah, panjang dan lebar daun
Hasil analisis sidik ragam menunjukkan adanya pengaruh interaksi NAA dan BAP terhadap jumlah daun dan panjang daun, sedangkan lebar daun hanya dipengaruhi secara nyata oleh NAA (Tabel 3). Rataan jumlah dan panjang daun tanaman anggrek hitam in vitro disajikan
pada Tabel 4 dan 5.
Tabel 3. Rataan tinggi tanaman (mm) in vitroanggrek hitam pada perlakuan NAA dan BAP pada 23 MST.
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
39,335a 24,918
bc 26,835
bc
0,1 mgL-1
38,113a 21,330
cd 23,918
bc
0,2 mgL-1
35,058a 17,113
d 21,083
cd
0,3 mgL-1
38,113a 23,500
bc 21,585
cd
0,4 mgL-1
33,750a 23,085
bcd 26,948
bc
0,5 mgL-1
28,250b 22,750
bcd 25,470
bc
Keterangan: angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%.
Tabel 4. Rataan jumlahdaun (buah)anggrek hitam in vitro pada perlakuan NAAdanbAP pada 23 MST.
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
27,14bc
29,22abc
13,81f
0,1 mgL-1
23,86cde
24,22cd
15,72def
0,2 mgL-1
36,75a 16,69
def 15,86
def
0,3 mgL-1
37,11a 22,03
cf 14,19
ef
0,4 mgL-1
35,81ab
25,48cd
13,78f
0,5 mgL-1
31,36ab
24,97cd
17,19def
Keterangan: angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%.
Tabel 5. Rataan panjangdaun (mm)anggrek hitam in vitro pada perlakuan NAAdanbAP pada 23 MST
ZPT NAA
Konsentrasi 0 mgL-1 0,2 mgL
-1 0,5 mgL
-1
BAP
0 mgL-1 25,81
a 13,61
de 12,09
def
0,1 mgL-1
24,09ab
9,67ef 11,61
def
0,2 mgL-1
19,81bc
7,49f 9,67
ef
0,3 mgL-1
24,59ab
12,39def
13,19de
0,4 mgL-1
20,33bc
10,08ef 16,64
cd
0,5 mgL-1
15,42cd
11,78def
14,83de
Keterangan: angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
81
Rata-rata jumlah daun tertinggi yaitu pada media dengan penambahan NAA 0,0 mgL-1 dan BAP 0,3 mgL-1 yaitu sebesar 37,11 mm, tetapi tidak berbeda nyata dengan perlakuan NAA 0,0 mgL-1 dan BAP 0,2 mgL-1 sebesar 36,75 mm (Tabel 4), sedangkan rata-rata panjang daun tertinggi terdapat pada media tanpa penambahan ZPT yaitu 25,81 mm (Tabel 5). Auksin dan sitokinin merupakan dua jenis ZPT tanaman yang seringkali digunakan untuk menginduksi morfogenesis tanaman. NAA dan BAP merupakan jenis ZPT yang memiliki kisaran luas dalam memacu dan menghambat suatu pertumbuhan. NAA pada konsentrasi tertentu berfungsi untuk inisiasi akar dan batang tanaman, sedangkan BAP berfungsi untuk memacu inisiasi tunas. Menurut Suhentaka (2010) interaksi auksin dan sitokinin yang sangat nyata menunjukkan bahwa perlakuan sitokinin tidak dapat dilepaskan dari pengaruh auksin, sehingga dalam penggunaan sitokinin, baik efek mendorong maupun efek menghambat proses pembelahan sel tergantung dari adanya fitohormon lainnya. Dalam perlakuan ini sitokinin cenderung meningkatkan jumlah
daun namun menekan panjang daun. Diduga konsentrasi sitokinin (BAP) yang ditambahkan dan auksin endogen sudah memberikan perimbangan yang sesuai untuk memacu pembelahan sel dalam pembentukan organ daun. Terlihat bahwa penambahan BAP sampai konsentrasi 0,3 mgL-1 mampu meningkatkan jumlah daun, kemudian menurun seiring dengan penambahan BAP diatas konsentrasi tersebut (Tabel 4), berbeda pada NAA semakin tinggi konsentrasi NAA yang diberikan (N1 dan N2) maka jumlah daun lebih sedikit dibandingkan N0. Panjang akar
Hasil analisis sidik ragam menunjukkan jumlah akar hanya dipengaruhi oleh perlakuan NAA sedangkan panjang akar dipengaruhi oleh interaksi NAA dan BAP (Tabel 2). Akar mulai tumbuh pada minggu ke-4 setelah tanam yaitu pada media tanpa ZPT eksogen (kontrol), diikuti minggu ke-6 pada perlakuan NAA 0,0 mgL-1 dan BAP 0,1 mgL-1, NAA 0,0 mgL-1 dan BAP 0,2 mgL-1 dan NAA 0,0 mgL-1 dan BAP 0,3 mgL-1 sedangkan pada perlakuan-perlakuan
Tabel 6. Rataan panjang akar (mm) anggrek hitam in vitro pada perlakuan NAA dan BAP pada 23 MST.
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
23,583a 4,028
de 7,503
cde
0,1 mgL-1
21,638ab
1,055e 8,638
cde
0,2 mgL-1
14,503bc
0,890e 7,110
cde
0,3 mgL-1
23,503a 2,610
de 6,388
cde
0,4 mgL-1
11,168cd
2,138de
15,333abc
0,5 mgL-1
11,055cd
7,250cde
11,168cd
Keterangan: Angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%.
Tabel 7. Rataan jumlah tunas (buah) anggrek hitam in vitro pada perlakuan NAA dan BAP pada 23 MST.
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
5,805bcd
6,055a-d 1,723
e
0,1 mgL-1
3,613cde
5,583bcd
2,500e
0,2 mgL-1
8,945a 3,528
cde 2,250
e
0,3 mgL-1
7,778ab
4,475cde
2,195e
0,4 mgL-1
7,778ab
6,475abc
2,028e
0,5 mgL-1
6,418abc
5,643bcd
3,248de
Keterangan: Angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%.

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
82
lainnya akar baru tumbuh pada 7 MST (minggu setelah tanam).
Pada Tabel 6 secara umum terlihat bahwa peningkatan konsentrasi NAA dan BAP menekan panjang akar. Rata-rata panjang akar tertinggi terlihat pada perlakuan NAA 0,0 mgL-1 dan BAP 0,0 mgL-1
(23,583 mm), tetapi tidak berbeda nyata dengan perlakuan NAA 0,0 mgL-1 dan BAP 0,1 mgL-1 (21,638 mm) dan NAA 0,0 mgL-1 dan BAP 0,3 mgL-1 (23,503 mm) (Tabel 6). Hatta et al. (2008) menyatakan bahwa penambahan sitokinin mempengaruhi pembelahan sel dan morfogenesis, sedangkan auksin berperan mengatur pertumbuhan dan pemanjangan sel. Rata-rata panjang akar tertinggi pada perlakuan tanpa penambahan ZPT diduga karena kandungan auksin endogen yang sudah cukup tinggi sehingga mampu menginduksi pertumbuhan akar, akibatnya penambahan auksin eksogen akan menekan pemanjangan akar. Hal ini sesuai dengan pernyataan Kurnianingsih et al. (2009) yang menyatakan bahwa eksplan tunas samping Anthurium yang ditanam pada media kultur menghasilkan auksin endogen dengan konsentrasi cukup tinggi sehingga pertumbuhan eksplan lebih diarahkan pada pemanjangan sel dan pembentukan akar. Jumlah tunas
Jumlah tunas merupakan parameter penting dalam kultur jaringan, karena semakin banyak tunas yang terbentuk akan memberikan peluang didapatkan bibit dalam jumlah banyak pula. Kultur anggrek hitam (C. pandurata Lind.) memperlihatkan
proliferasi pada medium perlakuan (Tabel 7). Penelitian ini memperlihatkan bahwa rata-rata jumlah tunas tertinggi pada perlakuan NAA 0,0 mgL-1 dan BAP 0,2 mgL-1 sebesar 8,945 buah (Tabel 7, Gambar 2c) lebih tinggi dibanding kontrol yang hanya mencapai
5,805 buah (Gambar 2a). Laju multiplikasi sebesar 8,945 yang artinya setiap satu tunas anggrek hitam yang ditanam pada media ½ MS dengan penambahan NAA 0,0 mgL-1 dan BAP 0,2 mgL-1 akan mampu menghasilkan tunas baru sebanyak 8,945 buah. Pertumbuhan dan perkembangan suatu kultur dipengaruhi oleh perimbangan antara hormon yang ditambahkan ke dalam media (hormon eksogen) dan hormon yang dihasilkan oleh tanaman itu sendiri (hormon endogen). Perbandingan antara sitokinin dan auksin yang tinggi maka akan mendorong terbentuknya tunas, sedangkan perbandingan sitokinin dan auksin rendah akan mendorong terbentuknya akar. Menurut Paramartha et al. (2012) meningkatnya auksin di dalam sel merupakan stimulus untuk aktivasi sitokinin. Aktifnya sitokinin diikuti dengan aktifnya enzim yang menaikkan sintesis protein yang merupakan protein pembangun sel sehingga menyebabkan terbentuknya sel-sel baru yang pada akhirnya terdiferensiasi menjadi organ tertentu.
Pengaruh BAP terhadap multiplikasi tunas terlihat juga pada penelitian Fithriyandini et al. (2015) yang melaporkan bahwa penambahan BAP 2,5 ppm pada media ½ MS terhadap anggrek bulan (Phalaenopsis amabilis) memberikan hasil jumlah PLBs, waktu muncul tunas, jumlah tunas dan jumlah daun terbaik. Penelitian pada tanaman anggrek Grammatophyllum scriptum (Lindl.) yang dilakukan Markal et al.
(2015) menunjukkan bahwa waktu terbentuknya tunas tercepat, jumlah tunas terbanyak dan jumlah daun terbanyak terjadi pada media yang ditambahkan BAP 1 mgL-1 dan NAA 0,5 mgL-1. Penelitian Karyanti dan Kartini (2017) pada tanaman Satoimo (Colocasia esculenta (L) Schoot var antiquorum) memperlihatkan hasil bahwa
pemberian sitokinin (TDZ) dan kasein
Gambar 2. Keragaan tunas in vitro pada berbagai media perlakuan pada 23 MST: (a). media kontrol (tanpa zpt), (b).
media perlakuan NAA 0 mgL-1 BAP 0,2 mgL
-1, (c). media perlakuan NAA 0,5 mgL
-1 BAP 0 mgL
-1.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
83
hidrolisat memberikan pengaruh sangat nyata terhadap jumlah tunas tanaman Satoimo. Menurut Arti dan Mukarlina (2017) sitokinin memacu pembelahan sel tunas, dimana rasio auksin endogen dan sitokinin eksogen yang berimbang menyebabkan pertumbuhan sel-sel meristem yang akan membelah dan berkembang menjadi tunas. Harjadi (2009) menyatakan bahwa pemberian BAP memacu sintesis protein sehingga mendorong terjadinya pembelahan sel yang menginduksi terbentuknya tunas.
Rata-rata jumlah tunas terendah terdapat pada perlakuan NAA 0,5 mgL-1 dan BAP 0,0 mgL-1 sebesar 1,723 buah (Gambar 2b) , diduga akibat tingginya kandungan auksin endogen dan auksin eksogen (NAA) yang diberikan sehingga rasio auksin dan sitokinin menjadi tinggi yang berakibat terhambatnya pembentukan tunas.
Waktu muncul tunas merupakan salah satu indikator pertumbuhan yang menunjukkan bahwa tanaman responsif terhadap perlakuan. Waktu muncul tunas
diamati setiap minggu. Tunas pertama muncul pada minggu ke-5 pada perlakuan NAA 0,0 mgL-1 dan BAP 0,0 mgL-1, NAA 0,0 mgL-1 dan BAP 0,4 mgL-1, NAA 0,2 mgL-1 dan BAP 0,3 mgL-1 dan NAA 0,2 mgL-1 dan BAP 0,4 mgL-1. Selanjutnya perlakuan NAA 0,0 mgL-1 dan BAP 0,1 mgL-1, NAA 0,0 mgL-
1 dan BAP 0,2 mgL-1, NAA 0,0 mgL-1 dan BAP 0,3 mgL-1, NAA 0,2 mgL-1 dan BAP 0,2 mgL-1 dan NAA 0,2 mgL-1 dan BAP 0,4 mgL-1 pada minggu ke-6, sedangkan perlakuan lainnya baru menunjukkan keluarnya tunas baru pada minggu ke-7.
Klorofil
Klorofil merupakan pigmen berwarna hijau yang berfungsi sebagai penyerap cahaya dalam kegiatan fotosintesis (Bakti 2009). Hasil uji klorofil pada daun anggrek hitam dengan metode Sims dan Gamon (2003) terlihat pada Tabel 8 yang memperlihatkan bahwa jumlah klorofil total menunjukkan hasil yang sangat berbeda nyata antara beberapa perlakuan.
Tabel 8. Rataan jumlah klorofil (mgg-1) anggrek hitam in vitro pada perlakuan NAA dan BAP pada 23 MST.
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
a b Total a b Total a b Total
BAP
0,0 mgL-1
0,95a 0,41
ab 1,35
a 0,44
cf 0,21
cde 0,64
cde 0,43
cf 0,21
cde 0,64
cde
0,1 mgL-1
0,37def
0,19de
0,55de
0,42cf 0,22
be 0,64
cde 0,35
ef 0,15
de 0,49
de
0,2 mgL-1
0,32f 0,17
de 0,53
de 0,62
af 0,29
ae 0,90
ae 0,66
af 0,31
ae 0,97
ae
0,3 mgL-1
0,71ae
0,34ad
1,05ad
0,31f 0,15
e 0,45
e 0,48
bf 0,25
ae 0,73
be
0,4 mgL-1
0,52bf 0,26
ae 0,78
be 0,73
ad 0,43
a 1,16
abc 0,82
ab 0,39
abc 1,21
ab
0,5 mgL-1
0,47bf 0,21
cde 0,68
be 0,56
bf 0,28
ae 0,84
ae 0,78
abc 0,39
abc 1,18
abc
Keterangan: Angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%
Tabel 9. Rataan panjang dan lebar stomata (µm) anggrek hitam in vitro pada pengaruh NAA dan BAP pada 23 MST
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
P. Stomata L. Stomata P. Stomata L. Stomata P. Stomata L. Stomata
BAP
0,0 mgL-1
38,99a 35,05
ab 39,49
a 35,88
a 36,94
abc 26,66
cd
0,1 mgL-1
38,82a 29,67
abcd 33,89
abcd 28,89
abcd 38,38
ab 32,21
abc
0,2 mgL-1
29,03cd
26,6 cd
31,15abcd
28,77abcd
31,82abcd
26,76cd
0,3 mgL-1
35,8 abc
27,34bcd
32,77abcd
23,95d 30,27
bcd 23,41
d
0,4 mgL-1
32,09abcd
27,34bcd
30,42bcd
23,41d 32,04
abcd 27,07
bcd
0,5 mgL-1
29,95cd
25,57cd
33,09abcd
28,92abcd
26,33d 23,25
d
Keterangan: Angka yang diikuti huruf yang sama menunjukkan tidak berbeda nyata berdasarkan Uji DMRT pada = 5%

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
84
Hasil pengamatan menunjukkan bahwa kandungan total klorofil tertinggi terdapat pada perlakuan tanpa penambahan ZPT NAA dan BAP sebesar 1,35 mgg-1, namun tidak berbeda nyata dengan perlakuan NAA 0,0 mgL-1 dan BAP 0,3 mgL-1 (1,05 mgg-1), NAA 0,2 mgL-1 dan BAP 0,4 mgL-1 (1,16 mgg-1), NAA 0,5 mgL-1 dan BAP 0,4 mgL-1 (1,21 mgg-1) dan NAA 0,5 mgL-1 dan BAP 0,5 mgL-1 (1,18 mgg-1). Peningkatan konsentrasi BAP pada setiap level NAA cenderung meningkatkan jumlah klorofil total, kecuali pada NAA 0,0 mgL-1. Pada konsentrasi NAA 0,2 dan 0,5 mgL-1, klorofil total tertinggi diperoleh pada kombinasi dengan BAP 0,4 mgL-1, yaitu NAA 0,2 mgL-1
dan BAP 0,4 mgL-1 sebesar 1,156 mgg-1 dan NAA 0,5 mgL-1 dan BAP 0,4 mgL-1 sebesar 1,179 mgg-1.
Klorofil sangat berperan penting dalam fotosintesis, jumlah klorofil berkaitan erat dengan hasil yang diperoleh dari proses
yang melibatkan cahaya tersebut. Total klorofil yang tinggi menunjukkan energi yang dihasilkan dari proses fotosintesis tersebut juga tinggi. Energi ini akan digunakan tanaman untuk pertumbuhannya. Pada percobaan ini tanaman dengan perlakuan tanpa penambahan ZPT menunjukkan pertumbuhan yang terbaik terlihat dari tinggi tanaman (39,335 mm) (Tabel 3) dan panjang daun tertinggi (25,81 mm) (Tabel 5) dibandingkan dengan perlakuan lainnya sesuai dengan klorofil total (1,35 mgg-1) yang dimiliki oleh tanaman tersebut. Menurut Thamrin (2008) menyatakan bahwa tinggi rendahnya kandungan klorofil di daun berhubungan dengan kecukupan hara yang diperlukan tanaman, dimana kandungan klorofil yang tinggi mampu mendorong proses fotosintesis tanaman sehingga fotosintat yang dihasilkan akan banyak dan pertumbuhan vegetatif tanaman lebih pesat (dalam hal ini tinggi tanaman paling tinggi
Tabel 10. Rataan jumlah stomata (buah) anggrek hitam in vitro pada pengaruh NAA dan BAP pada 23 MST
ZPT NAA
Konsentrasi 0,0 mgL-1
0,2 mgL-1
0,5 mgL-1
BAP
0,0 mgL-1
11,50cd
12,50bcd
15,00bcd
0,1 mgL-1
9,50cd
15,25bcd
11,50cd
0,2 mgL-1
16,00abcd
18,25abc
18,75abc
0,3 mgL-1
14,75bcd
20,00ab
13,50bcd
0,4 mgL-1
16,00abcd
23,25a 13,25
bcd
0,5 mgL-1
13,25bcd
12,00bcd
18,50abc
Keterangan: Angka pada kolom yang sama diikuti huruf yang sama tidak berbeda nyata berdasarkan Uji DMRT pada = 5%
Gambar 3. Representasi stomata anggrek hitam pada perlakuan kombinasi NAA dan BAP. a) N2B4 (NAA 0,2 dan BAP 0,4 mgL
-1) b) N0B1 (NAA 0,0 dan BAP 0,1 mgL
-1)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
85
(39,335 mm)). Jumlah klorofil yang banyak sangat menguntungkan untuk tanaman in vitro karena menurut Zulkarnain (2009)
sistem perakaran pada planlet yang berasal dari kultur jaringan mudah rusak dan tidak berfungsi sempurna, disamping itu lapisan lilin tipis dan belum berkembang dengan baik akibatnya tanaman akan mati saat aklimatisasi. Pada percobaan ini panjang akar tertinggi 23,583 mm (Tabel 6) dan jumlah klorofil paling banyak (1,35 mgg-1) (Tabel 8) diharapkan dapat membantu persentase tumbuh tanaman anggrek hitam ketika diaklimatisasi (Andarini 2013). Menurut Prasetyo (2017) semakin banyak jumlah klorofil maka daun akan semakin hijau dan laju fotosintesis meningkat. Stomata
Stomata merupakan lubang atau celah yang terdapat pada epidermis organ tumbuhan yang dibatasi oleh sel khusus yang disebut sel penutup (Samosir 2014). Stomata berperan dalam pertukaran CO2 saat fotosintesis dan penguapan air dari tanaman ke lingkungan melalui transpirasi (Arif 2016). Tabel 9 menunjukkan bahwa perlakuan NAA dan BAP mempengaruhi panjang dan lebar stomata. Secara umum terlihat bahwa panjang dan lebar stomata cenderung menurun dengan meningkatkan konsentrasi NAA dan BAP.
Panjang dan lebar stomata terpanjang pada perlakuan NAA 0,2 mgL-1 dan BAP 0,0 mgL-1 sebesar 39,49 dan 35,88 µm. Ukuran stomata berhubungan dengan jumlah kloroplas pada sel penjaga stomata. Peningkatan jumlah kloroplas mengakibatkan perubahan ukuran stomata (panjang dan lebar) menjadi lebih besar. Semakin besar ukuran stomata maka semakin baik menyerap CO2 untuk fotosintesis. Kerapatan stomata yang baik dapat membantu proses metabolisme tanaman. Menurut Megia et al. (2015) ukuran stomata terbagi menjadi tiga bagian, yaitu kurang panjang (< 20 µm), panjang (20-25 µm) dan sangat panjang (> 25 µm) pada penelitian ini semua perlakuan menunjukkan panjang stomata lebih dari 25 µm. Panjang dan lebar stomata sangat penting untuk tanaman in vitro karena tanaman hasil in vitro umumnya memiliki
lapisan lilin yang tipis dan belum berkembang dengan baik, sel-sel palisade
belum berkembang maksimal, jaringan pembuluh dari akar ke pucuk kurang berkembang dan stomata seringkali tidak berfungsi (tidak dapat menutup saat penguapan tinggi) (Andarini 2013).
Perlakuan NAA 0,2 mgL-1 dan BAP 0,4 mgL-1 menunjukkan jumlah stomata paling banyak (Tabel 10, Gambar 3a). Hal ini memperlihatkan kecenderungan bahwa konsentrasi BAP berperan terhadap karakter stomata. Pada NAA 0,0 (N0) dan 0,2 (N1) mgL-1 semakin tinggi konsentrasi BAP yang ditambahkan sampai 0,4 mgL-1 terlihat jumlah stomata makin meningkat, tetapi ketika ditambahkan dengan konsentrasi BAP lebih tinggi jumlah stomata cenderung menurun. Jumlah stomata yang banyak dan perlakuan hardening off (proses penguatan) tanaman in vitro akan membantu
kesuksesan tanaman saat aklimatisasi nanti.
KESIMPULAN
Hasil penelitian menunjukkan bahwa respon pertumbuhan tunas in vitro anggrek
hitam terhadap berbagai kombinasi NAA dan BAP. Media terbaik untuk multiplikasi tunas adalah media tanpa NAA dengan penambahan BAP 0,2 mgL-1. Morfogenesis tunas yang baik dengan jumlah dan panjang daun serta akar terbaik adalah pada media tanpa NAA dan BAP. Dengan demikian tahapan multiplikasi tunas dapat dilakukan pada media tanpa NAA ditambah BAP 0,2 mgL-1, kemudian tunas dipindah ke media tanpa NAA dan BAP untuk penyiapan planlet yang siap untuk diaklimatisasi.
DAFTAR PUSTAKA
Adi NKAP, Astarini IA, Astiti NPA (2014) Aklimatisasi anggrek hitam (Coelogyne pandurata Lindl.) hasil perbanyakan in vitro pada media berbeda. J Simbiosis 2: 203-214
Andarini YN (2013) Respon planlet anggrek Dendrobium spectabile pada pemberian beberapa taraf paclobutrazol selama tahap aklimatisasi. Skripsi, Institut Pertanian Bogor
Arif A (2016) Keragaan planlet Dendrobium lasianthera (JJ Smith) hasil iradiasi
sinar gamma generasi Mv1. Skripsi, Institut Pertanian Bogor

Multiplikasi In Vitro Anggrek Hitam...Kartiman et al.
86
Arti LT, Mukarlina (2017) Multiplikasi anggrek bulan (Dendrobium sp) dengan
penambahan ekstrak taoge dan benzyl amino purine (BAP) secara in vitro. J Protobiont 6: 278-282
Bakti PLW (2009) Analisis kandungan klorofil dan laju fotosintesis tebu transgenik PS-IPB 1 yang ditanam di kebun percobaan PG Djatiroto, Jawa Timur. Skripsi, Institut Pertanian Bogor
Fithriyandini A, Maghfoer MD, Wardiyati T (2015) Pengaruh media dasar dan 6-benzylaminopurine (BAP) terhadap pertumbuhan dan perkembangan nodus tangkai bunga Anggrek Bulan (Phalaenopsis amabilis) dalam perbanyakan secara in vitro. J Produksi Tanaman 3: 43-49
Gantait S, Mandal N, Bhattacharyya S, Das PK (2011) Induction and identification of tetraploids using in vitro colchicine treatment of Gerbera jamesonii Bolus
cv. Sciella. Plant Cell Tiss Organ Cult 106:485-493. doi:10.1007/s11240-011-9947-1
Gravendeel B (2000) Reorganising the orchid genus Coelogyne: A phylogenetic classification based on morphology and molecules. National Herbarium Nederland, Universiteit Leiden, Netherland
Harjadi SS (2009) Zat Pengatur Tumbuh. Penebar Swadaya, Jakarta
Hatta M, Hayati M, Irayani U (2008) Pengaruh IAA dan BAP terhadap pertumbuhan tanaman nilam (Pogestemon cablin Benth) in vitro. J Floratek 3: 56-60
Karyanti, Kartini M (2017) Pengaruh thidiazuron dan hidrolisat kasein terhadap multiplikasi tunas Satoimo (Colocasia esculenta (L) Schott var antiquarum. J Bioteknol Biosains
Indones 4: 70-77. doi: 10.29122/jbbi.v4i2.2535
Kurnianingsih R, Marfuah, Matondang I (2009) Pengaruh pemberian BAP (6-Benzyl Amino Purine) pada media multiplikasi tunas Anthurium hookerii Kuntt. Enum. secara in vitro. Vis Vitalis
02: 23-30 Ling ACK, Yap CP, Shaib JM, Vilasini P
(2007) Induction and morphogenesis of Phalaenopsis callus. J Trop Agric and Fd Sc 35: 147-152
Markal A, Isda MN, Fatonah S (2015) Perbanyakan anggrek Grammatophyllum scriptum (Lindl.) BL. melalui induksi tunas secara in vitro dengan penambahan BAP dan NAA. JOM FMIPA 2: 108-114
Paramartha AI, Ermavitalini D, Nurfadilah S (2012) Pengaruh penambahan kombinasi konsentrasi ZPT NAA dan BAP terhadap pertumbuhan dan perkembangan biji Dendrobium taurulinum J.J Smith secara in vitro. J
Sains dan Seni ITS 1: 40-43 Pramesti G (2011) SPSS 18.0 dalam
Rancangan Percobaan. PT. Elek Media Komputindo, Jakarta
Prasetyo AN (2017) Kajian pemberian giberelin pada fisiologi tanaman iles-iles (Amorphophallus muelleri Blume.)
berbeda umur. Skripsi, Institut Pertanian Bogor
Putri HA (2015) Pengaruh komposisi media dasar dan kitosan terhadap pertumbuhan protocorm like bodies (PLBs) dan planlet Anggrek Phalaenopsis hibrida. Skripsi, Institut
Pertanian Bogor Megia R, Ratnasari, Hadisunarso (2015)
Karakteristik morfologi dan anatomi, serta kandungan klorofil lima kultivar tanaman penyerap polusi udara Sansevieria trifasciata. J Sumberdaya
Hayati 1: 34-40 Restiani R, Semiarti E, Indrianto A (2016)
Konservasi anggrek hitam (Coelogyne pandurata Lindl.) melalui
mikropropagasi pada berbagai medium kultur. Pp 393-404. Prosiding Simposium Nasional Pendidikan Biologi (Symbion 2016) 27 Agustus 2016, UAD Yogyakarta
Samosir IA (2014) Karakter morfologi dan anatomi daun beberapa spesies Sansevieria. Skripsi, Institut Pertanian Bogor
Sims DA, Gamon JA (2003) Estimation of vegetative water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens Environ 84: 526-537. doi: 10.1016/S0034-4257(02)00151-7
Suhentaka EB (2010) Pengaruh konsentrasi BA dan NAA pada tahap multiplikasi

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
87
secara in vitro terhadap keberhasilan aklimatisasi nanas (Ananas comosus
(L) Merr) kultivar Smooth Cayenne. Skripsi, Institut Pertanian Bogor
Thamrin M (2008) Peningkatan pembungaan jeruk Pamelo (Citrus grandis (L.)
Osbeck) “Cikoneng” melalui strangulasi. Tesis, Institut Pertanian Bogor
Untari R (2003) Pengaruh jenis media organik dan NAA terhadap pertumbuhan anggrek hitam (Coelogyne pandurata Lindl.) di dalam kultur in vitro. Skripsi, Institut Pertanian
Bogor Zulkarnain (2009) Kultur Jaringan Tanaman:
Solusi Perbanyakan Tanaman Budidaya. Bumi Aksara, Jakarta

88
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
ISOLASI DAN ANALISA GENISTEIN DARI TEMPE BUSUK
MENGGUNAKAN METODE KROMATOGRAFI KOLOM
Isolation and Analysis of Genistein of Overripe Tempe using Column Chromatography Method
Hartati Soetjipto*, Yohanes Martono, dan Zulfa Yuniarti
Program Studi Kimia, Fakultas Sains dan Matematika, Universitas Kristen Satya Wacana, Jl. Diponegoro
No. 52-60 Salatiga 50711 Jawa Tengah
*Email: [email protected]
ABSTRACT Genistein is one of the aglycone isoflavone compounds in tempe that has various biochemical activities, including anticancer, antitumor, and antioxidants. Commonly used isoflavone extraction methods resulted in isoflavone crude extract. The aim of this study was to isolate the genistein of overripe tempe through determining the appropriate combination of mobile phases in genistein isolation and the determination of genistein content in both crude extract and isolate. The overripe tempe was first extracted, then genistein was isolated from the crude extract using column chromatography method. The determination of mobile phase combination was done by Thin Layer Chromatography while the genistein content was quantitatively determined by using High Performance Liquid Chromatography. The results showed that the appropriate combination of mobile phase for genistein isolation was chloroform : methanol (15 : 1, v/v). Genistein content in the crude extract and isolates were 4737.50 and 31.36 μg/g extract, respectively. The genistein purity in the isolates was 63.80%, while the purity in the isoflavone extract was 31.98%. Keywords: genistein, HPLC, isoflavone, overripe tempe, TLC ABSTRAK Genistein merupakan salah satu senyawa isoflavon aglikon dalam tempe yang memiliki bermacam-macam aktivitas biokimia, diantaranya antikanker, antitumor, dan antioksidan. Metode ekstraksi isoflavon yang umum diterapkan, menghasilkan ekstrak kasar isoflavon yang masih berupa campuran. Tujuan dari penelitian ini adalah untuk mengisolasi genistein dari tempe busuk melalui tahap penentuan kombinasi fase gerak yang tepat dalam isolasi genistein serta penentuan kandungan genistein baik dalam ekstrak kasar maupun isolat. Tempe busuk mula-mula diekstrak, selanjutnya genistein diisolasi dari ekstrak kasar menggunakan metode kromatografi kolom. Penentuan kombinasi fase gerak dilakukan secara Kromatografi Lapis Tipis, sedangkan kandungan genistein secara kuantitatif ditentukan dengan menggunakan Kromatografi Cair Kinerja Tinggi. Hasil penelitian menunjukkan bahwa kombinasi fase gerak yang tepat untuk isolasi genistein adalah kloroform : metanol (15 : 1, v/v). Kandungan genistein dalam ekstrak kasar dan isolat genistein berturut-turut sebesar 4737,50 dan 31,36 μg/g ekstrak. Kemurnian genistein dalam isolat adalah sebesar 63,80%, sedangkan kemurniannya dalam ekstrak isoflavon adalah sebesar 31,98%.
Kata Kunci: genistein, HPLC, isoflavon, tempe busuk, KLT
Received: 18 April 2018 Accepted: 12 June 2018 Published: 29 June 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
89
PENDAHULUAN
Isoflavon merupakan senyawa
polifenol yang termasuk dalam golongan flavonoid. Bahan pangan yang menempati urutan pertama kaitannya dengan kandungan senyawa isoflavon dan derivatnya adalah kedelai. Kandungan total isoflavon pada kedelai berkisar antara 34,39 hingga 48,95 mg/100 g (Bhagwat et al. 2008). Terdapat empat bentuk isoflavon di dalam kedelai, yaitu glikosida, asetil glikosida, malonil glikosida, dan aglikon, yang masing-masing memiliki tiga isomer. Isomer dari isoflavon aglikon yaitu genistein, daidzein, dan glisitein. Menurut Astuti (2008) genistein dan daidzein merupakan isoflavon utama dalam kedelai, diikuti turunan β-glikosida-nya yaitu genistin dan daidzin.
Kandungan isoflavon aglikon dalam kedelai meningkat dengan adanya proses fermentasi (Hong et al. 2012). Tempe merupakan produk fermentasi kedelai yang paling banyak dikonsumsi masyarakat dibandingkan produk fermentasi lainnya. Kandungan genistein dalam tempe mencapai lebih dari dua kali lipat dibandingkan kandungannya dalam kedelai yang belum diolah. Menurut Ahmad et al. (2015), kandungan genistein pada kedelai sebesar 11,10 mg/100 g sedangkan pada tempe sebesar 24,03 mg/100 g. Kandungan isoflavon aglikon dalam tempe diketahui masih meningkat dengan bertambahnya waktu pemeraman. Menurut Riyanto dan Soetjipto (2017), tempe pemeraman hari keempat mengandung isoflavon genistein tertinggi diantara pemeraman hari ke-0 hingga ke-8 yaitu sebesar 26,199 ± 25,146 mg/g.
Berbagai penelitian tentang genistein terus dilakukan sehubungan dengan bioaktivitas genistein, diantaranya antikanker, antitumor, dan antioksidan (Ariani dan Hastuti, 2009). Metode ekstraksi isoflavon yang umum diterapkan, menghasilkan ekstrak kasar isoflavon yang berupa campuran antara berbagai senyawa isoflavon baik dalam bentuk glikosida maupun aglikon. Menurut Sartini dkk. (2014), ekstraksi isoflavon kedelai dengan metode maserasi menggunakan rotary shaker
menghasilkan ekstrak isoflavon yang terdiri dari daidzein, genistein, daidzin, dan genistin. Sama halnya dengan penelitian
tersebut, Kuligowski dkk. (2016) menyatakan ekstraksi kedelai terfermentasi dengan teknik yang berbeda yaitu menggunakan pengadukan (stirring) juga menghasilkan hasil ekstrak dengan kandungan serupa.
Dalam penerapannya sebagai senyawa antikanker, antitumor, dan antioksidan, diharapkan diperoleh genistein yang lebih murni. Sebab adanya senyawa lain dapat memberikan kontribusi pada bioaktivitas genistein (Singh-Gupta et al. 2010). Oleh sebab itu, pada penelitian ini akan diisolasi isoflavon genistein dari tempe busuk pemeraman hari keempat. Pemurnian genistein dari ekstrak kasar akan dilakukan secara kromatografi kolom, sedangkan konsentrasi genistein baik dalam ekstrak maupun isolat ditentukan dengan menggunakan Kromatografi Cair Kinerja Tinggi (High Performance Liquid Chromatography / HPLC).
BAHAN DAN METODE Bahan dan alat
Bahan baku dalam penelitian ini adalah tempe yang diperoleh dari pengrajin tempe di Bugel, Salatiga, Jawa Tengah. Senyawa standar yang digunakan adalah genistein (Sigma Chemical Co, Amerika
Serikat). Pelarut kimia yang digunakan adalah n-heksana, etanol, kloroform, dan metanol dengan derajat pro analisis produk E-Merck, Jerman.
Peralatan yang digunakan antara lain neraca analitis dengan ketelitian 0,0001 g (Ohaus PA214), grinder (Philiphs HR-2108), drying cabinet, moisture analyzer (OHAUS
MB 25), peralatan ekstraksi (Soxhlet), centrifuge, ultrasonikator (Krisbow Ultrasonic
Gambar 1. Sampel tempe busuk pemeraman hari ke-4

Isolasi dan Analisa Genistein dari Tempe Busuk... Soetjipto et al.
90
cleaner DSA50-GL2-2.5L), rotary evaporator
(Buchi R-114), plat KLT silika gel 60 F254, chamber KLT, seperangkat alat kromatografi kolom, dan HPLC (Knauer Smartline 5000, Smartline pump 1000). Preparasi sampel
Sampel tempe yang digunakan adalah tempe pemeraman hari ke-4 (Gambar 1). Sampel dipotong tipis-tipis lalu dikeringkan pada drying cabinet dengan suhu 50°C
selama 2 hari. Sampel selanjutnya dihaluskan dengan grinder dan disimpan dalam wadah kering untuk analisis lebih lanjut. Pengukuran kadar air
Sebanyak kurang lebih 1 g sampel dimasukkan ke dalam alat moisture analyzer. Kadar air sampel akan tertera pada layar monitor alat terhitung sebagai persentase air yang hilang saat proses pemanasan sampel. Ekstraksi isoflavon
Sampel tempe didefatisasi menggunakan pelarut n-heksana (1:5, b/v) selama 3 jam pada alat Soxhlet (Yunindarwati dkk. 2016 yang dimodifikasi). Simplisia yang telah bebas lemak tersebut selanjutnya dikeringkan. Sebanyak 25 g simplisia kering tersebut diekstraksi dengan cara ultrasonikasi menggunakan pelarut etanol 70% (1:6, b/v) selama 1 jam. Ekstrak cair selanjutnya dipisahkan dengan cara sentrifugasi kecepatan rendah (800 rpm) selama 10 menit. Residu yang tersisa diekstraksi kembali sebanyak 2 kali menggunakan pelarut baru. Seluruh ekstrak cair dikumpulkan dan dipekatkan di rotary evaporator hingga didapatkan ekstrak kental. Selanjutnya ekstrak kental yang diperoleh ditimbang kemudian dihitung rendemennya berdasarkan persamaan berikut:
%100)%()(
Re
airkadarsampelberatsampelberat
ekstrakberat
ndemen
Kromatografi lapis tipis
Analisis Kromatografi Lapis Tipis (KLT) (Hessler et al. 1997 yang dimodifikasi) dilakukan dengan menggunakan fase diam plat silika gel 60 F254 dan fase gerak berupa
campuran kloroform : metanol berbagai komposisi. Pengamatan dilakukan di bawah sinar ultra violet dengan panjang gelombang 254 nm. Nilai Rf diukur, kemudian dibandingkan dengan nilai Rf senyawa standar genistein. Perhitungan nilai Rf dihitung dengan persamaan berikut:
pelarutpermukaanolehdigerakkanyangjarak
senyawaolehdigerakkanyangjarakRf
Kromatografi kolom
Fase diam yang digunakan pada kromatografi kolom (Asih 2009 yang dimodifikasi) adalah silika gel, sedangkan fase gerak yang digunakan adalah kloroform : metanol pada komposisi yang memberikan pemisahan terbaik pada KLT. Perbandingan antara sampel dengan fase diam adalah 1 : 10 (b/b). Eluat ditampung pada botol penampung fraksi setiap ± 0,5 mL, kemudian keseluruhan fraksi yang dihasilkan dianalisis menggunakan KLT. Fraksi hasil KLT yang mempunyai pola pemisahan sama (harga Rf sama) digabungkan, kemudian diuapkan. Penentuan kandungan isoflavon genistein
Identifikasi isoflavon menggunakan metode HPLC dilakukan dengan pengkondisian instrumen HPLC dan pembuatan larutan sampel (César et al. 2006 yang dimodifikasi). Larutan sampel dibuat dengan melarutkan ekstrak dalam 5 mL metanol. Selanjutnya sebanyak 20 μL larutan sampel diinjeksikan ke HPLC setelah pengkondisian HPLC selesai. Kromatogram HPLC dianalisis dengan menggunakan pembanding kromatogram isoflavon genistein standar.
Kondisi operasional instrumen HPLC meliputi fase diam Vertex, Euroshper 100-5
C18, 250 4,6 mm (WL 75) dan fase gerak menggunakan campuran metanol : asam asetat 0,1% dengan perbandingan 48:52 (v/v). Kecepatan alir yang digunakan sebesar 1,2 mL/min dengan volum injeksi 20 μL dan menggunakan detektor UV 254 nm. Analisis data
Analisis kuantitatif genistein secara HPLC dilakukan dengan metode standar eksternal. Konsentrasi genistein dalam sampel dapat diketahui dengan memplotkan area puncak analit pada kurva baku standar.

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
91
HASIL DAN PEMBAHASAN
Pengukuran kadar air
Berdasarkan proses pengukuran, kadar air dari serbuk tempe busuk yang
sudah kering sebesar 6,51%. Dibandingkan dengan penelitian Riyanto dan Soetjipto (2017), dimana kadar air serbuk tempe hari ke-4 yang sudah dikeringkan sebesar 4,566%, penelitian ini menghasilkan nilai
(10:1) (15:1) (20:1) (25:1) (50:1)
Gambar 2. Hasil analisis KLT perbandingan standar genistein dengan ekstrak isoflavon tempe. (Keterangan: Fase
gerak =CHCl3 : CH3OH (v/v); fase diam = plat silika gel; noda kiri = standar genistein; noda kanan = ekstrak isoflavon tempe)
Tabel 1. Hasil KLT ekstrak isoflavon tempe dan standar genistein pada berbagai perbandingan fase gerak
Nilai Rf Fase gerak CHCl3 : CH3OH (v/v)
10 : 1 15 : 1 20 : 1 25 : 1 50 : 1
Standar Genistein 0,50 0,45 0,33 0,28 0,12
Ekstrak Isoflavon
Noda 1 0,50 0,45 0,35 0,28 0,12
Noda 2 0,43 0,35 0,27 0,22 0,07
Noda 3 0,33 0,27 0,2 0,17 —
Noda 4 0,25 0,18 — — —

Isolasi dan Analisa Genistein dari Tempe Busuk... Soetjipto et al.
92
kadar air yang lebih tinggi. Menurut Rosida dkk. (2009), kadar air sampel dipengaruhi oleh kadar air awal bahan bakunya. Selain itu, perbedaan ukuran sampel juga sangat mempengaruhi nilai kadar air sampel tersebut (Purnama dkk. 2012). Nilai kadar air yang diperoleh digunakan untuk menentukan berat kering dari sampel pada perhitungan rendemen ekstrak.
Rendemen ekstrak isoflavon
Ekstrak isoflavon diperoleh melalui ekstraksi tempe fermentasi hari ke-4, dengan metode ultrasonikasi menggunakan pelarut etanol 70% yang sebelumnya telah melewati tahap defatisasi menggunakan pelarut n-heksana. Rendemen ekstrak isoflavon yang diperoleh sebesar 24,87% (b/b). Lewidharti dkk. (2015) melaporkan bahwa tempe fermentasi hari ke-4 yang diekstrak secara maserasi dengan pelarut metanol 80%, menghasilkan rendemen sebesar 30,31% (b/b). Dibandingkan dengan hasil penelitian tersebut, penelitian ini menghasilkan rendemen ekstrak yang lebih rendah. Hal ini disebabkan karena efisiensi ekstraksi secara ultrasonikasi juga dipengaruhi oleh frekuensi gelombang ultrasonik selain dipengaruhi pemilihan pelarut, waktu ekstraksi, dan suhu ekstraksi.
Paparan gelombang ultrasonik yang terlalu lama dapat mengganggu stabilitas senyawa isoflavon. Paparan tersebut menimbulkan adanya disosiasi ultrasonik sehingga terbentuk senyawa radikal. Kehadiran senyawa radikal (spesies berenergi tinggi) tersebut menyebabkan terjadinya reaksi oksidatif pada proses ekstraksi yang mempengaruhi stabilitas senyawa (Rostagno et al. 2009). Perbedaan perlakuan pada sampel tempe, termasuk sumber bahan baku pembuatan tempe (varietas, kematangan, dan tempat tumbuh kedelai), dan cara pengolahan tempe yang digunakan juga berpengaruh terhadap rendemen ekstrak yang dihasilkan (Suharto dkk. 2017). Kromatografi lapis tipis
Hasil analisis kromatografi lapis tipis (KLT) ekstrak isoflavon pada berbagai perbandingan fase gerak dilihat di bawah sinar UV 254 nm (Gambar 2). Ekstrak yang di-KLT dibandingkan dengan standar genistein pada setiap perbandingan fase gerak.
Rf ekstrak yang relatif bernilai sama dengan Rf standar membuktikan bahwa noda yang muncul adalah genistein (Tabel 1). Perbandingan kloroform : metanol (10 : 1, v/v) dan (15 : 1, v/v) menghasilkan pemisahan 4 noda, kloroform : metanol (20 : 1, v/v) dan (25 : 1, v/v) 3 noda, dan kloroform : metanol (50 : 1, v/v) 2 noda. Semakin banyak perbandingan kloroform yang digunakan, Rf yang dihasilkan semakin rendah dan noda yang muncul semakin sedikit. Hal ini disebabkan karena senyawa yang akan dipisahkan semakin tertahan pada fase diam karena pengaruh polaritas baik dari senyawa, fase diam, maupun fase gerak. Rf yang beragam merupakan hasil dari berbagai tingkat afinitas antara komponen campuran dengan fase diam dan gerak.
Hasil KLT pada fase gerak kloroform : metanol (10 : 1, v/v) sesuai dengan penelitian Jyoti et al. (2015) dimana Rf dari genistein standar sebesar 0,50. Noda pemisahan yang terbentuk pada fase gerak klorofom : metanol (20 : 1, v/v) menunjukkan tailing, dimana noda
cenderung miring ke arah kanan. Hal ini diduga disebabkan karena kondisi chamber
yang belum jenuh sehingga mempengaruhi migrasi fase gerak menjadi tidak lurus. Menurut Sherma dan Fried (2003), parameter-parameter yang menyebabkan tailing adalah volume sampel yang
berlebihan saat diaplikasikan ke plat KLT, keaktifan plat KLT, jarak pengembang, dan kejenuhan chamber.
Noda pemisahan yang berhimpit pada fase gerak kloroform : metanol (50 : 1, v/v) disebabkan karena polaritas fase gerak yang kurang sesuai sehingga tidak mampu memisahkan komponen senyawa secara maksimal. Noda yang berhimpit juga dapat disebabkan oleh volume sampel yang berlebihan saat diaplikasikan ke KLT (Bele dan Khale 2011).
Hasil yang diperoleh menunjukkan fase gerak kloroform : metanol (10 : 1, v/v) memberikan pemisahan yang paling baik dilihat dari nilai Rf yang dihasilkan. Senyawa isoflavon yang dipisahkan dengan fase gerak tersebut, memenuhi parameter optimum yaitu nilai Rf berada pada pendistribusian di wilayah 0,2-0,8 (Shewiyo et al. 2012). Namun untuk fase gerak yang digunakan pada kromatografi kolom,

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
93
diharapkan jarak pemisahan antar noda lebih panjang (terutama untuk noda genistein dengan noda berikutnya) sehingga nantinya dapat diperoleh senyawa yang lebih murni. Sehingga pada kromato-grafi kolom digunakan fase gerak kloroform : metanol dengan perbandingan 15 : 1 (v/v).
Hasil KLT yang diperoleh tidak hanya digunakan untuk menentukan kombinasi fase gerak yang tepat untuk isolasi, namun juga dapat digunakan untuk memperkirakan berapa senyawa yang terpisahkan berdasarkan jumlah noda pemisahan yang muncul. Oleh sebab itu, tahapan optimasi fase gerak untuk isolasi senyawa penting dilakukan sebelum tahapan isolasi itu sendiri.
Kromatografi kolom
Terhadap 0,305 g ekstrak isoflavon tempe, dilakukan proses pemisahan dengan menggunakan fase diam silika gel 60 (70-100 Mesh) sebanyak 3 g (panjang kolom 20 cm, diameter 1 cm) menggunakan fase gerak kloroform : metanol (15 : 1, v/v). Hasil kromatografi kolom adalah 66 fraksi (tiap fraksi ± 0,5 mL). Selanjutnya terhadap fraksi-fraksi tersebut dilakukan KLT gabungan dimana tiap fraksi dianalisis menggunakan KLT, fase gerak kloroform : metanol (10 : 1, v/v). Fraksi-fraksi yang
membentuk noda dengan nilai Rf relatif sama, digabungkan menjadi satu kelompok fraksi. Hasil dari KLT gabungan, membentuk 4 kelompok fraksi yaitu fraksi A, B, C, dan D (Gambar 3).
Fraksi A merupakan gabungan dari fraksi ke-22 hingga 33, fraksi B dari fraksi ke-34 hingga 40, fraksi C dari fraksi ke-41 hingga 45, sedangkan fraksi D dari fraksi ke-46 hingga 60. Fraksi pertama hingga fraksi ke-21 dan fraksi ke-61 hingga 66 tidak memperlihatkan adanya noda pemisahan yang menunjukkan bahwa pada fraksi tersebut hanya terdapat fase gerak.
Fraksi A dan B menunjukkan positif genistein karena terbentuk noda dengan nilai Rf yang sama dibandingkan dengan Rf standar genistein, yaitu sebesar 0,47. Fraksi A dinilai relatif lebih murni sebagai isolat genistein secara KLT karena menghasilkan noda tunggal, sedangkan fraksi B menghasilkan dua noda dimana noda kedua diduga masih termasuk senyawa golongan isoflavon yang terkandung dalam tempe. Fraksi A inilah yang selanjutnya disebut isolat genistein dan digunakan untuk analisis lanjutan. Fraksi B, C, dan D menghasilkan noda sangat tipis. Hal ini diduga karena kadar senyawa isoflavon dalam fraksi tersebut sangat rendah.
Gambar 3. Hasil analisis KLT perbandingan standar genistein dengan fraksi-fraksi hasil kromatografi kolom. (Keterangan: fase gerak = CHCl3 : CH3OH (10 : 1, v/v); fase diam = plat silika gel; noda kiri ke kanan = standar genistein, fraksi A, fraksi B, fraksi C, dan fraksi D)

Isolasi dan Analisa Genistein dari Tempe Busuk... Soetjipto et al.
94
Kadar isoflavon genistein
Kadar isoflavon genistein baik dalam ekstrak kasar isoflavon tempe maupun isolat genistein hasil kromatografi kolom ditentukan secara kuantitatif menggunakan HPLC. Analisis kuantitatif genistein
dilakukan dengan menggunakan metode standar eksternal dengan memplotkan area puncak analit pada kurva baku standar. Kromatogram HPLC ekstrak isoflavon tempe dan isolat genistein ditunjukkan pada Gambar 4. Puncak genistein ekstrak isoflavon dan isolat genistein berturut-turut
A
a.
b.
Gambar 4. Kromatogram HPLC (a). ekstrak isoflavon tempe dan (b). isolat genistein
Gambar 5. Reaksi hidrolisis genistin menjadi genistein (Rostagno et al. 2009)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
95
muncul pada waktu retensi 27,000 dan 27,683 menit. Puncak-puncak lain yang terdeteksi diduga merupakan senyawa golongan isoflavon, salah satu diantaranya yang dominan diduga merupakan daidzein (tR = 15,800 menit).
Kadar genistein dari hasil analisis dengan HPLC pada ekstrak isoflavon sebesar 4737,50 μg/g ekstrak. Dibandingkan dengan hasil penelitian Yunindarwati dkk.(2016), penelitian ini menghasilkan ekstrak dengan kadar genistein yang lebih tinggi. Yunindarwati dkk. (2016) melaporkan bahwa ekstrak kedelai terfermentasi Aspergillus oryzae
hari ke-4 mengandung genistein sebesar 54,373 ± 1,755 μg/g ekstrak.
Salah satu penyebab perbedaan kadar genistein dalam ekstrak, diduga disebabkan oleh jenis kapang yang digunakan dalam proses pembuatan tempe. Cheng et al. (2010) menyatakan bahwa jenis kapang yang berbeda, memiliki aktivitas biotransformasi isoflavon yang berbeda pula. Menurut Lee dan Chou (2006), Rhizopus sp. no. 2 menunjukkan
peningkatan tertinggi isoflavon aglikon dalam fermentasi koji kacang hitam sebesar 2,9 hingga 58,9% setelah fermentasi, dibandingkan dengan Aspergillus awamori, Aspergillus oryzae, Aspergilllus sojae, dan Rhizopus azygosporus yang hanya berkisar
18,9 – 38,9% setelah fermentasi. Biotransformasi isoflavon glikosida
(genistin) menjadi isoflavn aglikon (genistein) terjadi akibat reaksi hidrolisis oleh aktivitas enzim β-glukosidase. Reaksi hidrolisis genistin menjadi genistein ditunjukkan pada Gambar 5.
Kadar genistein dalam isolat terukur sebesar 31,36 μg/g ekstrak. Bila dibandingkan dengan kadar genistein dalam ekstrak kasar yang diisolasi (yang digunakan untuk kromatografi kolom), yaitu sebesar 248,66 μg/g ekstrak, maka dapat ditentukan persen perolehan kembali (% recovery) yaitu sebesar 12,61%. Rendahnya % recovery yang diperoleh
diduga disebabkan karena ada sebagian genistein yang belum terpisah dengan senyawa lain (masih dalam bentuk campuran) (Gambar 3), selain itu terdapat keterbatasan pada sistem kromatografi kolom yang mempengaruhi hasil pemisahan. Menurut Beller dan Hilleary
(1976), kolom yang digunakan untuk kromatografi kolom harus dielusi dengan campuran pelarut yang setidaknya memiliki polaritas seperti pelarut yang digunakan untuk melarutkan sampel. Keterbatasan tersebut sering menyebabkan pemisahan dan goresan komponen menjadi tidak baik.
Bila dibandingkan antara isolat (31,36 μg/g ekstrak) dengan ekstrak isoflavon (4737,50 μg/g ekstrak), genistein yang terisolasi dalam isolat relatif lebih rendah. Hal ini menunjukkan metode isolasi yang dilakukan dengan menggunakan fase diam silika gel 60 (70-100 Mesh) dan fase gerak kloroform : metanol (15 : 1, v/v) masih perlu dikembangkan lebih lanjut agar % recovery
genistein dapat lebih sempurna. Kemurnian genistein dihitung
berdasarkan area puncak kromatogram (Gambar 4), yaitu dengan membagi area puncak genistein dengan total area puncak-puncak yang ada. Sehingga kemurnian genistein dalam isolat sebesar 63,80%. Dibandingkan dengan kemurniannya dalam ekstrak isoflavon sebesar 31,98%, kemurnian genistein dalam isolat lebih besar
hampir mencapai 2 lipat. Namun, kemurnian tersebut dinilai masih belum cukup besar sehingga seperti yang sudah disebutkan sebelumnya, metode yang diterapkan pada penelitian ini masih perlu dikembangkan lebih lanjut.
KESIMPULAN
Kombinasi fase gerak yang tepat
dalam isolasi genistein adalah kloroform : metanol (15 : 1, v/v). Kandungan genistein dalam ekstrak kasar isoflavon tempe busuk hari ke-4 dan isolat genistein berturut-turut sebesar 4737,50 dan 31,36 μg/g ekstrak. DAFTAR PUSTAKA
Ahmad A, Ramasamy K, Majeed ABA, Mani
V (2015) Enhancement of β-secretase inhibition and antioxidant activities of tempeh, a fermented soybean cake through enrichment of bioactive aglycones. Pharm Biol 53:758–766. doi: 10.3109/13880209.2014.942791
Ariani SRD, Hastuti W (2009) Analisis isoflavon dan uji aktivitas antioksidan pada tempe dengan variasi lama waktu fermentasi dan metode ekstraksi.

Isolasi dan Analisa Genistein dari Tempe Busuk... Soetjipto et al.
96
Prosiding Seminar Nasional Kimia dan Pendidikan Kimia, Pp 568–580. 18 Maret 2009, Surakarta
Asih IARA (2009) Isolasi dan identifikasi senyawa isoflavon dari kacang kedelai (Glycine max). J Kimia 3:33–40
Astuti S (2008) Isoflavon kedelai dan potensinya sebagai penangkap radikal bebas. J Tek Industri dan Hasil Pertanian 13:126–136
Bele AA, Khale A (2011) An overview on thin layer chromatography. Int J Pharma Sci Res 2:256-267. doi: 10.13040/IJPSR.0975-8232.2(2).256-67
Beller NR, Hilleary CJ (1976) A sample preparation technique for column chromatography. J Chem Educ 53: 498. doi: 10.1021/ed053p498
Bhagwat S, Haytowitz DB, Holden JM (2008) USDA database for the isoflavone content of selected foods: Release 2.0. Nutrient Data Laboratory, United State of America
Cheng KC, Lin JT, Wu JY, Liu WH (2010) Isoflavone conversion of black soybean by immobilized Rhizopus spp. Food
Biotechnol 24: 312-331. doi: 10.1080/08905436.2010.524459
César IC, Braga FC, Soares CD, Nunan EA, Pianetti GA, Condessa FA, Barbosa TA, Campos LM (2006) Development and validation of a RP-HPLC method for quantification of isoflavone aglycones in hydrolyzed soy dry extracts. J Chromatogr B 836:74–78. doi: 10.1016/j.jchromb.2006.03.030
Hessler PE, Larsen PE, Constantinou AI, Schram KH, Weber JM (1997) Isolation of isoflavones from soy-based fermentations of the erythromycin-producing bacterium Saccharopolyspora erythraea. Appl
Microbiol Biotechnol 47:398–404. doi: 10.1007/s002530050947
Hong GE, Mandal PK, Lim KW, Lee CH (2012) Fermentation increases isoflavone aglycone contents in black soybean pulp. Asian J Animal Vet Adv 7:502-511. doi: 10.3923/ajava.2012.502.511
Jyoti, Agrawal SS, Saxena S, Sharma A (2015) Phytoestrogen "genistein": Its extraction and isolation from soybean seeds. Int J Phramacogn Phytochem Res 7:1121-1126
Kuligowski M, Pawłowska K, Jasińska-Kuligowska I, Nowak J (2016) Isoflavone composition, polyphenols content and antioxidative activity of soybean seeds during tempeh fermentation. CyTA J Food 15:27-33. doi: 10.1080/19476337.2016.1197316
Lee IH, Chou CC (2006) Distribution profiles of isoflavone isomers in black bean kojis prepared with various filamentous fungi. J Agric Food Chem 54:1309-1314. doi: 10.1021/jf058139m
Lewidharti RS, Soetjipto H, Andini S (2015) Dinamika konsentrasi genistein dalam proses pembusukan tempe kedelai. Prosiding Seminar Nasional Kimia dan Pendidikan Kimia VII. 18 April 2015, Surakarta
Purnama FA, Dewi L, Hastuti SP (2012) Kadar air, abu, protein dan karbohidrat pada tahap pembuatan tempe. Skripsi, Universitas Kristen Satya Wacana
Riyanto CA, Soetjipto H (2017) Solvent optimization for genistein isolation of “rotten tempe” by high performance liquid chromatography method. J Eksakta: 17:111-118
Rosida DF, Sudaryati HP, Constantia F (2009) Kajian peran angkak pada kualitas tempe kedelai-lamtoro gung (Leucaena leucocephala). J Rekapangan 6:64-72
Rostagno MA, Villares A, Guillamon E, Garcia-Lafuente A, Martinez JA (2009) Sample preparation for the analysis of isoflavones from soybeans and soy foods. J Chromatogr A 1216:2–29. doi: 10.1016/j.chroma.2008.11.035
Sartini, Djide MN, Permana AD, Ismail (2014) Ekstraksi isoflavon kedelai dan penentuan kadarnya secara Ultra Fast Liquid Chromatografi (UFLC). J Sainsmat 3:130-134. doi: 10.2685/sainsmat3211202014
Sherma J, Fried B (2003) Handbook of Thin-Layer Chromatography, Third Edition. Marcel Dekker Inc, New York
Shewiyo DH, Kaale E, Risha PG, Dejaegher B, Smeyers-Verbeke J, Vander-Heyden Y (2012) Optimization of a reversed-phase-high-performance thin-layer chromatography method for the separation of isoniazid, ethambutol, rifampicin and pyrazinamide in fixed-dose combination antituberculosis

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
97
tablets. J Chromatogr A 1260:232–238. doi: 10.1016/j.chroma.2012.08.044
Singh-Gupta V, Zhang H, Yunker CK, Ahmad Z, Zwier D, Sarkar FH, Hillman GG (2010) Daidzen effect on hormone refractory prostate cancer in vitro and in vivo compared to genistein and soy extract: Potentiation of radiotherapy. Pharm Res 27:1115-1127. doi: 10.1007/s11095-010-0107-9
Suharto KF, Soetjipto H, Martono Y (2017)
Pengaruh lama fermentasi tempe terhadap kandungan total senyawa fenolik dan isoflavon genistein. ALCHEMY J Penelitian Kimia 13:230-240. doi: 10.20961/alchemy.v13i2.5094
Yunindarwati E, Ulfa EU, Puspitasari E, Hidayat MA (2016) Penentuan kadar genistein dan aktivitas hambatan tirosinase kedelai (Glycine max) terfermentasi Aspergillus oryzae. J Ilmu Kefarmasian Indones 14:1–7

98
VOLUME 5 NOMOR 1 JUNI 2018 ISSN 2548 – 611X
JURNAL BIOTEKNOLOGI & BIOSAINS INDONESIA
Homepage Jurnal: http://ejurnal.bppt.go.id/index.php/JBBI
ANALISIS BIOINFORMATIKA BERBASIS WEB PADA SEKUEN GENOM PARSIAL SAGU (Metroxylon sagu Rottb.)
WEB-based bioinformatic analysis on partial genome sequence of Sago (Metroxylon sagu Rottb.)
Devit Purwoko
1*, Imam Civi Cartealy
1, Teuku Tajuddin
1, Diny Dinarti
2, Sudarsono
2
1Balai Bioteknologi, BPPT. Gedung 630 Kawasan Puspiptek, Tangerang Selatan, Banten 15314
2 Lab Biologi Molekuler Tanaman, Departmen Agronomi dan Hortikultura, IPB. Darmaga, Bogor 16680.
*Email: [email protected]
ABSTRACT Sago genome sequencing analysis is still very limited. This study is a preliminary study of sago sequence analysis obtained from NGS technology to understand and identify new genetic sequences that have homology to genes in the NCBI database. Sequences were analyzed using Blast2Go to determine the genetic function annotation, putative gene identification was performed on the Arabidopsis database using the BLASTx program with a 10
-3 e-value limit on The Arabidopsis Information Resource (TAIR)
(http://www.arabidopsis.org/index.jsp). Gene interactions were analyzed using DAVID and GeneMania programs. Based on sequence analysis with Blast2Go, 33 sequences with Blastx hit consisting of: 29 sequences had a high homology. The sago sequences with a similarity of ≥ 90% are glutamate decarboxylase and HT1-like serine threonine kinase with hit number 10. The distribution of interactions between genes from GeneMania analysis is known to be mostly interconnected in the 65.13% protein domain, predicted 19.83%, genes with 14.47% shared expression and the remaining 0.57% had localization together. Keywords: bioinformatics, gene annotation, gene ontology, genome sequence, Metroxylon
sagu
ABSTRAK Kajian analisis sekuen genom sagu hingga saat ini masih amat terbatas. Penelitian ini merupakan riset pendahuluan analisis sekuen sagu yang diperoleh dari teknologi NGS untuk mengetahui dan mengidentifikasi sekuen gen baru yang memiliki homologi dengan gen pada database NCBI. Sekuen dianalisis menggunakan perangkat Blast2Go untuk mengetahui anotasi fungsional gen, identifikasi gen putatif dilakukan terhadap database Arabidopsis menggunakan program BLASTx dengan batas e-value 10
-3 pada The Arabidopsis
Information Resource (TAIR). Interaksi gen dianalisis menggunakan program DAVID dan GeneMania. Berdasarkan analisis sekuen dengan Blast2Go, diperoleh 33 sekuen dengan Blastx hit yang terdiri atas: 29 sekuen memiliki homologi yang tinggi. Gen dengan rataan kemiripan ≥ 90% adalah glutamate decarboxylase dan serine threonine-kinase HT1-like dengan jumlah hit 10. Persebaran interaksi antar gen hasil analisis GeneMania diketahui sebagian besar saling terkait pada domain protein 65,13%, koneksi yang berhasil diprediksi 19,83%, gen dengan ekspresi bersama 14,47% dan sisanya 0,57% memiliki peranan bersama. Kata Kunci: anotasi gen, bioinformatika, Metroxylon sagu, ontologi gen, sekuen genome
Received: 18 April 2018 Accepted: 07 Juni 2018 Published: 29 Juni 2018

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
99
PENDAHULUAN
Sagu (Metroxylon sagu Rottb.)
merupakan salah satu tanaman Palmae penghasil pati yang mampu mengakumulasi pati pada batangnya 244 sampai 310 kg/pohon (Ehara et al. 2016). Luas areal hutan sagu dunia lebih dari 50% berada di Indonesia dengan potensi 20-40 ton pati sagu/ha/tahun (Bintoro 2011). Dalam 100 gram tepung sagu mengandung nutrisi sebagai berikut: kalori 285, air 77 g, protein 0,2 g, kalsium 30 mg, karbohidrat 71 g, besi 0,7 mg dan serat 0,3 g (Johnson 2010).
Tanaman sagu memiliki potensi ekonomi, energi dan genetik yang tinggi untuk dikembangkan di Indonesia. Secara ekonomi tanaman sagu memberikan keuntungan yang cukup besar dari pati yang dihasilkan persatuan luas dan waktu. Yamamoto (2010) melaporkan bahwa produksi pati kering dari tanaman sagu rata-rata mencapai 337 kg/pohon sedangkan Bintoro (2011) menyatakan potensi tanaman sagu menghasilkan pati 100-900 kg/pohon.
Jika jarak tanam 9 m 9 m akan terdapat 123 pohon/ha maka akan diperoleh 42 ton pati sagu/ha/tahun (Abbas 2006). Potensi produksi pati Indonesia diperkirakan 5 juta ton pati kering sagu/tahun sedangkan konsumsi pati dalam negeri baru mencapai 210 ton (Sumaryono 2007) dan ekspor baru mencapai 9.680,908 ton pada tahun 2015 (Ditjen Perkebunan 2016). Berdasarkan perhitungan tersebut masih terdapat potensi sekitar 4,5 juta ton pati sagu yang belum dimanfaatkan atau setara dengan 607,5 juta US$ jika diekspor mentah dan bernilai lebih besar lagi jika sudah dalam produk industri.
Potensi sagu yang tidak kalah penting dari sektor energi, jika 4,5 juta ton pati sagu yang belum dimanfaatkan digunakan sebagai bioetanol dengan perhitungan 25 ton/ha/tahun setara dengan 15 kiloliter etanol (Sumaryono 2007) maka akan diperoleh 2,7 juta kiloliter etanol. Tanaman sagu juga memiliki potensi genetik yang belum banyak dikembangkan. Tanaman sagu memiliki mekanisme genetik yang belum banyak dimengerti berkaitan dengan daya adaptasinya pada lingkungan yang tergenang dan pH rendah. Potensi genetik ini dapat dikembangkan untuk mencari gen-gen potensial terhadap cekaman abiotik.
Informasi berkenaan dengan karakter
berduri maupun potensi sagu secara morfologi untuk kandungan pati cukup banyak dikaji (Novero et al. 2012; Kjaer et al. 2004; Karim et al. 2008). Kajian budidaya dan perbanyakan bibit baik secara in-vitro (Riyadi et al. 2016) dan ex-vitro ( Karyanti et
al. 2016) juga telah dilaporkan. Namun Perkembangan kemajuan informasi genomik pada tanaman sagu masih terbatas terutama untuk marka DNA. Kajian molekuler berkenaan dengan keragaman dan kekerabatan sagu di Indonesia menggunakan marka juga belum banyak dilaporkan. Studi penanda DNA tanaman sagu hingga saat ini diantaranya penanda RAPD (Abbas et al. 2009), AFLP (Kjaer et al. 2004) dan DNA kloroplas (cpDNA) (Abbas et al. 2010).
Kajian analisis sekuen genom sagu hingga saat ini masih amat terbatas. Hingga saat ini baru terdeposit 111 sekuen genom dan 413 sekuen EST pada database National Center for Biotechnology Information (NCBI, data di akses Maret
2018). Analisis sekuen sagu telah dilaporkan oleh Wee dan Roslan (2012) dengan memanfaatkan EST dari daun muda sagu bahwa terdapat beberapa sekuen sagu yang belum memiliki homologi dengan sekuen pada database NCBI. Analisis sekuen untuk identifikasi gen ADP-Glucose Phyrophosphorylase (Budiani et al. 2015) dan gen GA 20-Oxidase (Jamel et al. 2011).
Analisis sekuen secara tradisional dengan kloning sekuen kemudian dilanjutkan perunutan dengan metode Sanger membutuhkan tenaga dan biaya. Analisis sekuen memanfaatkan teknologi perunutan DNA terkini yaitu Next Generation Sequencing (NGS) berbasis bioinformatika
mampu menghasilkan informasi yang efektif dan efisien. Hal ini dapat dilihat pada pertambahan data sekuen di NCBI yang dihasilkan melalui teknologi NGS. Hingga tahun 2016 data yang dihasilkan melalui teknologi ini sebesar 3.051,43 TB dengan total basa 5.005,71 Tb (Raza dan Ahmad 2017).
Analisis sekuen dengan bantuan bioinformatika dapat dilakukan dengan dua pendekatan yaitu berbasis web dan berbasis
algoritma bahasa pemograman. Bagi pemula pemanfaatan bioinformatika berbasis web akan lebih mudah dibandingkan dengan algoritma bahasa pemograman. Analisis bioinformatika berbasis web dapat digunakan dalam mencari anotasi

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
100
(penamaan), pemetaan genom, dan analisis sekuen lanjut seperti identifikasi gen homologi serta interaksi antar gen terkait. Analisis bioinformatik berbasis web dapat dijalankan secara daring (dalam jaringan) melalui program yang tersedia dengan akses publik di web. Keunggulan metode tersebut adalah hemat dan dapat menjadi penelitian pendahuluan sebelum percobaan secara nyata dilakukan (Narita et al. 2012). Namun untuk analisis data sekuen yang besar dibutuhkan algoritma bahasa pemograman yang ditunjang oleh super komputer agar dapat berjalan secara simultan.
Penelitian ini merupakan riset pendahuluan analisis sekuen sagu yang diperoleh dari teknologi NGS untuk mengetahui dan mengidentifikasi sekuen gen baru yang memiliki homologi dengan gen yang sudah ada pada basis data NCBI. Analisis interaksi gen potensial juga dilakukan untuk memperoleh informasi gen fungsional yang terkait pada jalur biosintesis suatu protein. Analisis bioinformatik sekuen sagu berbasis web diharapkan dapat
memberi landasan awal untuk diperolehnya gen-gen potensial dari tanaman sagu terkait ketahanan biotik dan abiotik.
BAHAN DAN METODE
Sampel sekuen yang digunakan pada penelitian ini merupakan 85 sekuen sagu yang telah dideposit pada database NCBI. Sekuen tersebut merupakan hasil sekuensing NGS menggunakan metode paired end pada mesin Illumina GAIIX dan di assembly menggunakan perangkat lunak
Ray. Untuk mengidentifikasi gen putatif, sekuen dibandingkan dengan database protein non-redundant NCBI menggunakan program BLASTx dengan nilai ambang 10-3. Hasil BLASTx dengan kemiripan rata-rata sama dengan atau kurang dari 60% dikategorikan sebagai homologi rendah sedangkan yang rata-rata lebih dari 60% dikategorikan sebagai homologi tinggi (Wee dan Roslan 2012). Sementara itu, sekuen tanpa kecocokan diklasifikasikan sebagai sekuen tanpa blast. Sekuen dengan hasil
analisis BLASTx kemudian dipetakan ke Gene Ontology (GO) menggunakan program
BLAST2GO dan dikategorikan berdasarkan fungsi molekuler, proses biologis dan komponen seluler (Wee dan Rooslan 2012).
Analisis sekuen dilakukan untuk mengetahui gen potensial menggunakan program Blastx dengan membandingkan protein non-redundant pada database NCBI. Sekuen juga dibandingkan dengan Arabidopsis database pada The Arabidopsis Information Resource (TAIR) (http://www.arabidopsis.org/index.jsp) menggunakan program BLASTx dengan batas e-value 10-3. Hasil blast kemudian dipetakan untuk mengetahui anotasi ontology gen yang meliputi kategori fungsi molekuler, proses biologi dan komponen seluler dengan menggunakan identifikasi lokus pada Bulk Retrieval System of TAIR
(http://www.arabidopsis.org/tools/bulk/go/index.jsp).
Gen-gen yang dianotasi dengan Arabidopsis dilanjutkan dengan analisis jalur
fungsional menggunakan aplikasi Online DAVID (Huang et al. 2009) (DAVID v6.7; http://david.abcc.ncifcrf.gov/; Diakses Agustus 2017). Aplikasi ini memiliki kelebihan karena mengandalkan algoritma anotasi fungsional sesungguhnya dan mampu mengelompokkan kumpulan banyak gen ke dalam kelompok fungsional. Penggunaan DAVID dimaksudkan untuk menentukan jalur (pathway) mana yang
paling banyak dipengaruhi oleh gen yang diprediksi sebelumnya. Prosesnya dengan memasukan daftar gen yang telah diprediksi sebelumnya ke dalam web aplikasi functional annotation tool DAVID dan identifikasi gen Arabidobsis dipilih sebagai
daftar gen. Idealnya, hanya kelompok anotasi dengan nilai enrichment 1,3 atau lebih tinggi yang dianggap signifikan namun kelompok anotasi dengan nilai enrichment
rendah (kurang dari 1,3) masih dapat dipertimbangkan untuk dianalisis lebih lanjut karena ada kemungkinan memiliki gen yang penting (Huang et al. 2009).
Analisis jaringan (network) interaksi antar gen dilakukan menggunakan GeneMania (Warde-Farley et al. 2010) (http://www.genemania.org/; Diakses Agustus 2017) untuk menentukan apakah ada prediksi pathway yang sesuai dalam network tertentu. GeneMania merupakan
aplikasi berbasis web yang tersedia secara publik untuk mengidentifikasi gen tambahan dari gen prediksi dan hubungan interaksinya dalam bentuk jaringannya. Gen yang diperoleh dari analisis pathway

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
101
menggunakan DAVID diunggah ke GeneMania dengan memilih database Arabidopsis. Semua parameter dibiarkan default untuk analisis. Hasil analisis dengan nilai false discovery rate (FDR) < 0,05 saja
yang dilaporkan. Alur proses analisis sekuen dapat dilihat pada Gambar 1. HASIL DAN PEMBAHASAN
Pada penelitian ini, analisis dilakukan pada 85 sekuen sagu yang diperoleh menggunakan teknologi Next Generation Sequencing (NGS) dan telah dideposit pada database NCBI dengan nomor identitas sekuen mulai dari MG904300 hingga MG904384. Sekuen dianalisis menggunakan BLASTX menggunakan database NCBI pada perangkat Blast2Go dengan nilai ambang 10-3 (Tabel 1). Sebanyak 33 sekuen yang dianalisis diketahui memiliki Blast hit, sedangkan 52 sekuen lainnya tanpa Blast hit. Berdasarkan analisis Blast diperoleh informasi bahwa 17 sekuen memiliki ontologi gen dan berhasil dipetakan. Sekuen yang memiliki kecocokan hanya pada InterProScan yaitu sc-7446. Namun yang memiliki kecocokan dengan database KEGG hanya 2 sekuen dengan 3 enzim yang sudah diketahui yaitu sekuen sc-678470 dan sc-16071 (Lampiran 1). Ketiga enzim tersebut yaitu Adenosinetriphosphatase (EC:3.6.1.3), Nucleoside-triphosphate phosphatase (EC:3.6.1.15) dan Glutamate decarboxylase
(EC:4.1.1.15). Berdasarkan analisis jalur KEGG EC:4.1.1.15 berada dalam jalur metabolism alanine, aspartate dan glutamate sedangkan EC:3.6.1.3 dan EC:3.6.1.15 berada dalam jalur metabolism purine.
Berdasarkan analisis sekuen dengan Blast2Go dapat dilihat bahwa terdapat 33 sekuen dengan Blastx hit yang terdiri atas: 29 sekuen memiliki homologi yang tinggi dengan rataan kemiripan ≥ 60% dan hanya 4 sekuen yang < 60%. Gen yang paling banyak ditemukan pada sekuen sagu dengan rataan kemiripan ≥ 90% adalah glutamate decarboxylase dan serine threonine-kinase HT1-like dengan jumlah hit
10. Glutamate decarboxylase (GAD)
merupakan gen dengan sekuen yang terkonservasi dan diketahui memiliki peranan dalam mengkatalisis dekarboksilasi glutamat untuk menghasilkan asam γ-aminobutirat (GABA). Pada tanaman, aktivitasnya dimodulasi oleh pH dan kalsium. Pengaturan aktivitas GAD oleh kalsium adalah melalui domain pengikat calmodulin yang tampaknya spesifik untuk tanaman dan dihasilkan dari proses peristiwa evolusi. GAD pada tanaman biasanya disandikan oleh beberapa gen sehingga menunjukkan pola ekspresi yang berbeda, termasuk spesifisitas jaringan dan respon terhadap stres, yang menunjukkan bahwa ada fenomena spesialisasi dalam kelompok gen ini. Meskipun peran GABA sebagai
Tabel 1. Distribusi sekuen sagu berdasarkan analisis dengan Blast2Go
Kriteria analisis sekuen Jumlah sekuen
Dianalisis dengan Blast2Go* 85
Memiliki ontologi gen 17
Memiliki Blastx hit 33
Tanpa Blastx hit 52
Berhasil dipetakan 17
Hanya memiliki InterProScan 1
Memiliki hit spesies terbanyak Elaeis guineensis
76
Memiliki anotasi KEGG 2
Anotasi dengan enzim yang sudah diketahui 3
*Nilai ambang ≥10
-3.
Gambar 1. Bagan alur analisis sekuen

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
102
neurotransmitter sangat dikenal pada hewan, beberapa peran untuk GABA dan metabolismenya telah diusulkan pada tanaman selama beberapa dekade terakhir. Peran GABA pada tanaman termasuk berperan sebagai biokimia pH-stat, dalam
penyimpanan nitrogen sementara, sebagai osmolyte yang kompatibel, dalam pertahanan melawan stres biotik, dalam mengendalikan spesies oksigen reaktif (Reactive Oxygen Species / ROS) dan
dalam metabolisme intermediet fotorespirasi. Selain itu, GABA dianggap sebagai sinyal yang mengendalikan ekspresi gen pengatur. Namun, informasi yang tersedia yang mendukung keterlibatan GAD dan / atau GABA dalam banyak proses sangat terbatas, begitu pula kontribusi metabolisme poliamina ke GABA pada tingkat sel (Molina-Rueda et al. 2015). Serine threonine-kinase HT1-like diketahui memiliki peranan penting
untuk regulasi gerakan stomata dan fungsinya lebih sebagai respon terhadap CO2 daripada terhadap ABA atau cahaya (Hashimoto et al. 2006). Berdasarkan analisis sekuen dengan Blast2Go, 52 sekuen belum memiliki kecocokan dengan database pada NCBI. Hal ini menunjukkan bahwa sekuen-sekuen tersebut memiliki potensi untuk dianalisis lebih lanjut dan berpeluang besar dalam memperoleh gen-gen baru yang hanya ada pada tanaman sagu.
Analisis BLAST 85 sekuen terhadap spesies pada database NCBI diperoleh informasi kecocokan beberapa spesies tanaman (Gambar 2), yaitu: delapan (40%)
tanaman monokotil (Oryza sativa Indica Group, Oryza sativa Japonica Group, Nelumbo nucifera, Asparagus officinalis, Musa acuminata subsp. Malaccensis, Ananas comosus, Phoenix dactylifera, Elaeis guineensis) dan dua belas (60%) tanaman dikotil (Aquilegia coerulea, Vigna angularis var. Singularis, Vitis vinifera, Sesamum indium, Macleaya cordata, Gossypium raimondii, Vigna singularis, Phalaenopsis equestris, Boea hygrometrica, Corchorus capsularis, Dendrobium officinale, Glycine max). Sekuen sagu terdistribusi paling
banyak pada tanaman dikotil dibandingkan tanaman monokotil. Hal yang berbeda diperoleh pada EST daun muda sagu bahwa sekuen sagu terdistribusi hampir sama antara tanaman monokotil dan dikotil yaitu 41,94% dan 39,24% (Wee dan Roslan 2012). Berdasarkan analisis sekuen hit terbanyak diperoleh pada Elaeis guineensis (76) dan Phoenix dactylifera (71) yang
merupakan tanaman Palma. Hal ini menunjukkan bahwa primer yang disintesis dari genom sagu dapat digunakan pula pada kedua tanaman tersebut.
Ontologi gen adalah klasifikasi gen sesuai kriteria yang telah ditentukan dengan menghubungkan semua gen yang tersedia ke dalam gen tertentu. Klasifikasi dilakukan dengan menghimpun serangkaian gen dalam tiga kelompok kategori: Celluler Componen (CC), Biological Process (BP) dan Molecular Function (MF). Program BLASTx pada TAIR digunakan untuk mencari anotasi ontologi gen. Hasil anotasi ontologi gen sekuen produk primer tanaman
Gambar 2. Frekuensi 20 spesies tanaman dengan hit terbanyak

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
103
sagu dengan nilai homologi yang tinggi digambarkan pada peta kromosom Arabidopsis thaliana (Gambar 3).
Sekuen tanaman sagu yang memiliki anotasi dengan gen Arabidopsis thaliana
tersebar pada lima kromosom dengan penyebaran terbanyak pada kromosom pertama. AT1G01510 merupakan GO ID yang terbanyak untuk kategori BP, gen ini berperan dalam morfogenesis daun, bunga dan buah. AT5G58300 merupakan GO ID terbanyak untuk kategori MF, gen ini berperan dalam mengikat ATP dan aktivitas protein kinase. AT5G50850 merupakan GO ID yang terbanyak untuk kategori CC, gen ini berperan dalam komponen sel mitokondria.
Hasil anotasi ontologi gen dan kategori fungsional berdasarkan identifikasi lokus dapat dilihat pada Gambar 4. Berdasarkan hasil anotasi sekuen, total 41 ontologi gen dapat ditentukan dan terdistribusi kepada tiga kategori: fungsi molekuler (60), proses biologi (93) dan komponen seluler (103). Jumlah total ketiga kategori tersebut lebih banyak dari jumlah total ontologi gennya (41) karena beberapa ontologi gen yang dipetakan berjumlah satu atau lebih dari masing-masing kategori.
Berdasarkan klasifikasi fungsional ontologi gen diperoleh bahwa kategori komponen seluler (Gambar 4A) membentuk 14 kelompok fungsional yang didominasi oleh komponen intraseluler (18,4%) dibandingkan ekstraseluler (3,9%). Beberapa gen juga diketahui berada pada bagian kloroplas dan mitokondria sekitar 6-7%. Sebagian kecil gen berada pada lokasi plastid sitosol dan retikulum endoplasma berkisar antara 1-2% saja.
Kategori proses biologi (Gambar 4B) membentuk 13 kelompok fungsional dengan proses seluler (26,9%) mendominasi, diikuti proses metabolisme (21,5%). Gen yang berkaitan dengan proses perkembangan dan organisasi sel juga relatif banyak dijumpai. Hal ini dimungkinkan karena sekuen diperoleh dari daun muda yang masih mengalami pertumbuhan dan perkembangan. Gen-gen terkait respon terhadap cekaman biotik dan abiotik juga ditemukan, sebagaimana diketahui bahwa tanaman sagu memiliki ketahanan terhadap cekaman lingkungan seperti lingkungan dengan pH rendah (Harsanto 1986).
Pada kategori fungsi molekuler (Gambar 4C) membentuk 13 kelompok fungsional dimana gen-gen terkait dengan aktivitas faktor transkripsi dan pengikat asam nukleat relatif sedikit dijumpai. Sedangkan gen terkait dengan pengikat RNA dan DNA banyak dijumpai, lebih dari 10%. Analisis sekuen pada ketiga kategori menunjukkan adanya sekuen yang tidak ada kecocokan sama sekali dengan sekuen pada database NCBI (unknown cellular components, unknown biological processes, unkown molecular functions). Hal ini dapat diduga
bahwa sekuen yang tidak diketahui atau belum terklasifikasikan tersebut merupakan sekuen dengan gen-gen yang hanya terdapat pada tanaman sagu saja.
Sudah diketahui umum bahwa gen tidak terekspresi sendiri tetapi ada gen-gen lain yang saling berinteraksi. Maka itu analisis interaksi antar gen juga dilakukan menggunakan GeneMANIA (www.genemania.org) berdasarkan bobot co-expression untuk mengetahui seberapa besar interaksi antar gen yang dianalisis. Metode pembobotan jaringan secara otomatis dipilih untuk menampilkan 20 gen terkait dan paling banyak 10 atribut terkait. Sebelumnya sebanyak 30 gen hasil anotasi ontologi gen pada TAIR dianalisis lebih lanjut menggunakan DAVID untuk mengetahui gen fungsional terkait.
Gambar 3. Penyebaran GO ID hasil ontologi gen pada kromosom Arabidopsis thaliana

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
104
Empat kluster yang diperoleh melalui
a.
b.
c.
Gambar 4 Pengelompokan fungsional genom sagu pada 85 sekuen A: Komponen seluler, B: Proses biologi, C:
Fungsi molekuler

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
105
aplikasi DAVID dipergunakan sebagai kumpulan gen untuk analisis lebih lanjut. Keempat kluster ini memiliki nilai Enrichment
antara 0,32 hingga 1,56 (Lampiran 2). Analisis gen fungsional dari DAVID yang berjumlah 30 gen kemudian dianalisis pada GeneMania sehingga diperoleh gambaran hubungan interaksi berdasarkan co-expression masing-masing gen (Gambar 5).
Dari 30 gen yang disubmit terdapat 20 gen
yang saling berinteraksi dari total 50 gen dengan jumlah 158 koneksi antar gen yang diprediksi oleh GeneMania. Persebaran interaksi antar gen hasil analisis GeneMania diketahui sebagian besar saling terkait pada domain protein 65,13%, koneksi yang berhasil diprediksi 19,83%, yang memiliki ekspresi bersama 14,47% dan sisanya 0,57% memiliki lokalisasi bersama.
Peranan interaksi gen dapat dilihat dari
Gen prediksi dari GeneMania
Garis hubung interaksi
Gen yang dicari
Gambar 5. Hubungan interaksi antar gen terkait hasil analisis ontologi sekuen sagu dengan gen pada Arabidopsis melalui GeneMania. Ukuran node menunjukkan besarnya peranan suatu gen dalam hubungan interaksi antar gen dari sagu.

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
106
ukuran node, makin besar node menandakan besarnya peranan gen pada hubungan interaksi tersebut. Berdasarkan besar peranan 20 gen yang saling berinterksi, gen phosphoribosyl-aminoimidazole carboxylase berada pada peringkat pertama dengan bentuk node yang paling besar. Gen ini diketahui berperan dalam biosintesis purin (https://www.ncbi.nlm.nih.gov/gene/10606). Peringkat ke-2 hingga ke-7 diketahui merupakan kelompok defensin-like protein, peringkat ke-9 hingga ke-14 diketahui merupakan kelompok Yip1 protein, peringkat ke-16 hingga ke-20 merupakan kelompok DUF679: protein yang belum diketahui fungsinya. Sedangkan peringkat ke-8 merupakan gen Apoptosis Inhibitory 5 yang berkaitan dengan kematian sel dan berperan sebagai penghambat endogen Caspase-2 dalam mengatur autophagy, stabilitas genomik dan penuaan sel (Imre et al. 2017). GEX2 pada peringkat 15 berperan pada faktor perkembangan reproduksi (Mori et al. 2014).
KESIMPULAN
Analisis bioinformatika sekuen sagu berbasis web mampu memberikan informasi homologi dan anotasi gen potensial. Sebanyak 33 sekuen sagu memiliki homologi pada database NCBI. Gen yang paling banyak ditemukan pada sekuen sagu dengan rataan kemiripan ≥ 90% adalah glutamate decarboxylase dan serine threonine- kinase HT1-like dengan jumlah hit 10. UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada BPPT atas beasiswa pada program pascasarjana IPB 2014.
DAFTAR PUSTAKA Abbas B (2006) Keragaman genetik
tanaman sagu di Indonesia berdasarkan marka molekuler genom kloroplas dan genom inti [disertasi]. Bogor (ID): Institut Pertanian Bogor
Abbas B, Bintoro MH, Sudarsono, Surahman M, Ehara H (2009) Genetic relationship of sago palm (Metroxylon sagu Rottb.)
in Indonesia based on RAPD markers. Biodiversitas. 10: 168–174. doi: 10.13057/biodiv/d100402
Abbas B, Yanuarius R, Bintoro MH, Sudarsono, Surahman M, Ehara H (2010) Genetic diversity of sago palm in Indonesia based on chloroplast DNA (cpDNA) Markers. Biodiversitas. 11: 112–117. doi: 10.13057/biodiv/d110302
Bintoro MH (2011) Proggress of sago research in Indonesia. The 10th International Sago Symposium. Institut Pertanian Bogor Press, Bogor
Budiani A, Putranto RA, Minarsi H, Riyadi I, Sumaryono, Abbas B (2015) Expression and cloning of gene encoding ADP- Glucose Phyrophosphorylase from sago palm (Metroxylon sagu Rottb). Menara Perkebunan 83: 76-85
Dirjenbun (2016) Statistik Perkebunan Indonesia 2015-2017: Sagu. Direktorat Jenderal Perkebunan, Kementerian Pertanian, Jakarta
Ehara H, Naito H, Mishima T, Toyoda Y, Mizota C, Susanto S, Bintoro MH, Pasolon YB, Abbas B, Suwignyo RA, Munandar (2016) Comparison of growth parameters and yield components of sago palms grown in the islands in Southeast Asia and Melanisia. Seminar Ilmiah dan Lokakarya Nasional Sagu 9-10 November 2016, Bogor, Indonesia
Harsanto PB (1986) Budidaya dan Pengelolaan Sagu. Kanisius, Jakarta
Hashimoto M, Negi J, Young J, Israelsson M, Schroeder JI, Iba K (2006) Arabidopsis HT1 kinase controls stomatal movements in response to CO2. Nat Cell Biol 8: 391-397. doi:10.1038/ncb1387
Huang DW, Sherman BT, Lempicki RA (2009) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 4: 44-57. doi: 10.1038/nprot.2008.211
Imre G, Berthelet J, Heering J, Kehrloesser S, Melzer IM, Lee BI, Thiede B, Dötsch V, Rajalingam (2017) Apoptosis inhibitor 5 is an endogenous inhibitor of
caspase‐2. EMBO Rep 18: 733-744. doi:10.15252/embr.201643744

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
107
Jamel B, Hussain MH, Salleh MA, Busri N (2011) Isolation and characterization of the GA 20-oxidase cDNA from sago palm (Metroxylon sagu Rottb.). AsPac J Mol Biol Biotechnol 19: 83-93
Johnson DV (2010) Tropical Palms. Food and Agriculture Organization (FAO) of The United Nations, Rome
Karim AA, Tie APL, Manan DMA, Zaidul ISM (2008) Starch from the sago (Metroxylon sagu) palm tree–
properties, prospects, and challenges as a new industrial source for food and other uses. Comprehensive Reviews in Food Science and Food Safety 7: 215-228. doi: 10.1111/j.1541-4337.2008.00042.x
Karyanti, Sigit Y, Tajuddin T, Erwinda, Minaldi, Haska N (2016) Penanganan anakan muda pada kultur ex vitro untuk menghasilkan bibit sagu (Metroxylon sagu Rottb.) siap tanam. J
Bioteknol Biosains Indones 3: 13-19. doi: 10.29122/jbbi.v3i1.20
Kjær A, Barfod AS, Asmussen CB, Seberg O (2004) Investigation of genetic and morphological variation in the sago palm (Metroxylon sagu; Arecaceae) in
Papua New Guinea. Ann Bot 94: 109–117. doi: 10.1093/aob/mch112
Molina-Rueda JJ, Garrido-Aranda A, Gallardo F (2015) Glutamate decarboxylase. In: Amino Acids in Higher Plants. Chapter 7: 129. Spain. doi: 10.1079/9781780642635.0129
Mori T, Iqawa T, Tamiya G, Miyaqishima SY, Berger F (2014) Gamete attachment requires GEX2 for successful fertilization in Arabidopsis. Curr Biol
24: 170-175. doi: 10.1016/j.cub.2013.11.030
Narita V, Arum AL, Isnaeni S, Fawzya NY (2012) Analisis bioinformatika berbasis web untuk eksplorasi enzim kitosanase
berdasarkan kemiripan sekuens. J Al-Azhar Indones 1: 197-203
Novero AU, Mabras MB, Esteban HJ (2012) Epigenetic inheritance of spine formation in sago palm (Metroxylon sagu Rottb.). Plant Omics J 5: 559-566
Raza K, Ahmad S (2017) Recent advancement in next generation sequencing techniques and its computational analysis. Tersedia pada: https://arxiv.org/ftp/arxiv/papers/1606/1606.05254.pdf
Riyadi I, Efendi D, Purwoko BS, Santoso D (2016) Embriogenesis somatik tidak langsung pada tanaman sagu (Metroxylon sagu Rottb.)
menggunakan sistem kultur suspensi, perendaman sesaat, dan media padat. J AgroBiogen 12: 37–44
Sumaryono (2007) Tanaman sagu sebagai sumber energi alternatif. Warta Penelitian dan Pengembangan Pertanian 29: 4
Warde-Farley D, Donaldson SL, Gomes O, Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, KaziF, Lopes CT, Maitland A, Mostafavi S, Montojo J, Shao Q, Wright G, Bader GD, Morris Q (2010) The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res 38: W214 -W220. doi: 10.1093/nar/gkq537
Wee CC, Roslan HA (2012) Expressed sequence tags (ESTs) from young leaves of Metroxylon sagu. 3 Biotech. 2: 211–218. doi: 10.1007/s13205-012-0048-6
Yamamoto Y (2010) Starch Productivity In Sago Palm. In: Sago Palm As The
Resource Crop In The 21st Century. Pp 218-232. The society of sago palm studies ed. Kyoto University press, Kyoto

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
108
INDEKS KATA KUNCI
6
6-Benzile Amino Purine...................... 82, 84
A
AFLP 8, 9, 11, 12, 14, 15, 16, 17, 18, 19, 20,
21, 109
anggrek hitam 82, 83, 84, 86, 87, 88, 89, 90,
91, 93, 94, 95
anotasi gen ................................... 108, 117
B
Bacillus sp. RP1 ...................... 45, 46, 48, 56
BAP ... 36, 37, 39, 40, 42, 43, 44, 82, 83, 84,
85, 86, 87, 88, 89, 90, 91, 92, 93, 94
Beauveria bassiana ..................... 1, 2, 3, 8
Biodiversity .............................................. 70
bioinformatics ................................. 107, 117
bioinformatika .............. 108, 109, 117, 118
black orchid.............................................. 82
blood disease ......................... 36, 37, 43, 44
C
cellulase ................................. 45, 55, 56, 57
chitinase ................................ 1, 7, 8, 57, 68
Chitinase .............................................. 1, 57
Coelogyne pandurata . 82, 83, 84, 93, 94, 95
D
daya apung ........... 29, 30, 31, 32, 33, 25, 35
daya serap air ........................................... 30
decolorization ............................... 20, 27, 28
dekolorisasi .......... 20, 21, 22, 23, 24, 25, 26
disease ............................ 36, 57, 68, 69, 70
DNA marker ............................................... 8
E
E. coli BL21 (DE3)....... 45, 46, 47, 49, 50, 55
egc ............................. 45, 46, 47, 48, 49, 55
Endoglucanase ......................................... 45
Endoglukanase ......................................... 46
entomopathogenic ................................. 1, 7
entomopatogen ...................................... 1, 2
enzim .....1, 2, 3, 4, 5, 6, 7, 8, 17, 18, 22, 25,
26, 45, 46, 47, 48, 49, 54, 55, 58, 59, 60,
62, 63, 68, 78, 90, 104, 111, 118
enzyme ............................................ 1, 8, 45
F
floatability ........................................... 29, 31
floating feed ....................................... 29, 35
G
G. boninense .. 57, 58, 59, 60, 61, 62, 63, 68
Ganoderma ... 19, 57, 58, 59, 62, 63, 64, 67,
68, 69
Ganoderma boninense .... 19, 57, 58, 59, 62,
68, 69
gene annotation ..................................... 107
gene ontology ........................................ 107
genistein.96, 97, 98, 99, 100, 101, 102, 103,
104, 105, 106
genome sequence ............................ 19, 107
H
HPLC ......96, 97, 98, 99, 100, 103, 104, 105
I
in vitro shoot ....................................... 36, 37
isoflavon .. 78, 96, 97, 98, 99, 101, 102, 103,
104, 105, 106
isoflavone ......................................... 96, 105
K
karet . 8, 9, 10, 11, 13, 14, 16, 17, 18, 19, 20,
21, 59, 61
keanekaragaman ....... 71, 72, 73, 78, 79, 81
kinetics .......................................................1
kinetik .........................................................2
kitinase ...1, 2, 3, 4, 5, 6, 7, 8, 58, 59, 60, 62,
68
Kitinase .......................................... 3, 58, 60
L
limbah tekstil .................... 20, 21, 22, 23, 25
Lineweaver-Burk ........................ 1, 2, 4, 6, 7

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
109
local knowledge ....................................... 70
M
male budless ..................................... 37, 38
Mandailing tribe ....................................... 70
marka DNA ......................... 8, 9, 10, 20, 109
Metroxylon sagu ..... 107, 108, 109, 117, 118
multiplication ..................................... 36, 82
multiplikasi ..... 37, 40, 42, 82, 83, 84, 85, 90,
93, 94, 95
mutan alami ............................................. 37
N
NAA ... 42, 44, 82, 83, 84, 85, 86, 87, 88, 89,
90, 91, 92, 93, 94, 95
Naphtalene Acetic Acid ..................... 82, 84
natural mutant ................................... 36, 37
O
obat tradisional ................. 70, 71, 72, 75, 79
ontologi gen .................. 108, 111, 113, 114
optimization ................................. 45, 56, 105
overripe tempe ........................................ 96
P
pakan apung ..................... 29, 30, 31, 33, 25
pakan tenggelam ............... 29, 30, 32, 33, 25
pengetahuan lokal ........................70, 71, 72
penyakit ... 10, 17, 37, 38, 44, 57, 59, 61, 67,
70, 71, 72, 73, 74, 75, 77, 78, 79
penyakit darah ..............................37, 38, 44
phylogenetic tree ....................................... 8
pohon filogenetik ................ 9, 15, 16, 17, 18
polimorfisme ............................. 9, 10, 12, 18
polymorphism ...................................... 8, 10
R
Reactive Red 2 ..... 20, 21, 22, 23, 24, 25, 26
rubber ...................................... 8, 19, 20, 21
S
S. rhizophila ... 57, 58, 59, 60, 61, 62, 63, 64,
67, 68
sekuen genome ................................... 108
selulase ................ 45, 46, 58, 59, 60, 63, 68
sinking feed ............................................. 29
stabilitas dalam air .............29, 30, 31, 32, 25
Stenotrophomonas rhizophila ...... 57, 58, 59
suku Mandailing ....................................... 71
Sumifix Blue ............... 20, 21, 22, 23, 25, 26
T
tanpa jantung ........................................... 37
tempe busuk ................. 96, 97, 98, 100, 104
textile effluent .............................. 20, 27, 28
TLC ......................................................... 96
traditional medicine .................................. 70
Trichoderma sp. .... 57, 58, 59, 60, 61, 68, 69
tumbuhan obat..... 70, 71, 72, 73, 74, 75, 77,
78, 79, 80, 81
tunas in vitro .. 37, 38, 39, 40, 42, 43, 44, 85,
88, 93
V
var. Unti Sayang ...............36, 37, 38, 42, 43
W
water absorption .......................... 29, 31, 35
water stability ..................................... 29, 31

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
110
INDEKS PENGARANG
A
Abdul Wahid ............................................. 57
Agus Purwito ............................................. 82
Aida Wulansari .......................................... 36
Asep Ali .................................................... 57
Asep Badru Zaman ................................... 29
Aswarina Nasution .................................... 70
B
Bambang Sukmadi .................................... 57
Bedah Rupaedah ...................................... 57
C
Catur Sriherwanto ..................................... 29
D
Debby Viola Amanda ................................ 57
Devit Purwoko ..................................... 8, 107
Dewi Sukma .............................................. 82
Diny Dinarti ............................................. 107
E
Eko Baroto Walujo .................................... 70
Endang Kusdiyantini ................................... 1
Ervizal AM Zuhud...................................... 70
Etyn Yunita ............................................... 29
H
Hans Victor ................................................ 45
Hartati Soetjipto ........................................ 96
I
Imam Civi Cartealy .................................. 107
Imam Suja’i ............................................... 29
Intan Permatasari...................................... 20
J
Juniza Firdha Suparningtyas ....................... 8
L
Laela Sari ..................................................36
M
Maelita Ramdani Moeis ..............................45
Mahmud Sugianto .....................................57
Maria Imelda .............................................36
N
Nia Asiani ..................................................57
Nunung Eni Elawati .....................................1
O
Okky Dwi Pramudyawardhani ......................8
R
Reni Indrayanti ..........................................57
Roni Kartiman............................................82
Rully Adi Nugroho .....................................20
S
Sri Pujiyanto ................................................1
Sudarsono ....................................... 107, 117
Syarifah Iis Aisyah .....................................82
T
Tatik Chikmawati .......................................70
Taufiq Firmansyah .....................................57
Teuku Tajuddin.................................... 8, 107
V
Vincentia Irene Meitiniarti ..........................20
Y
Yohanes Martono ......................................96
Z
Zulfa Yuniarti .............................................96

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
i
Tabel lampiran. Spesies Tumbuhan Obat yang Digunakan Masyarakat Mandailing
No. Famili Nama ilmiah Nama Lokal Kegunaan Cara meramu
1. Amaranthaceae Amaranthus sp Siarum Jerawat Daun + belerang +
minyak goreng + akar + batang + daun direbus
2. Amaryllidaceae Zephyranthes candida
(Lindl.) Herb. Bungo hujan Demam anak
Daun diremas + air dingin dimandikan
Crinum asiaticum L. Ompu-ompu Terkilir Daun dibakar + minyak dioles ke tangan ikat
dengan daun
3. Anacardiaceae Mangifera indica L. Manggak Asam lambung Kulit buah mangga + kulit lancat/langsat
direbus
4. Arecaceae Areca catechu L. Pining (nama lokal pinang)
Sakit perut Akar digiling lalu
dimakan
Diabetes Buah dikunyah
Gigi Buah + sirih dikunyah
Keropos tulang Akar pinang + kelapa +
aren direbus
Digigit
serangga Pinang muda dikunyah
lalu ditempel
Arenga pinnata (Wurmb.) Merr.
Bargot Keropos tulang Akar aren + akar pinang + akar kelapa direbus
diminum
Malaria Akar + kulit
lancat/langsat + garam direbus diminum
Cocos nucifera L. Harambir Keropos tulang Akar kelapa + akar pinang + akar aren direbus diminum
5. Balsaminaceae Impatiens balsamina L. Atirangga/bunga
pancur Malaria
Daun dihaluskan dimakan
6. Basselaceae Anrederia cordifolia
(Tenore) Binahong Kanker Daun direbus
7. Begoniaceae Begonia sp Bungo kombang Demam anak Daun dimandikan
8. Bromeliaceae Ananas sp Onas ubat Panas dalam Buah diperas diminum
9. Campanulaceae Hippobroma longifolia
(L.) G. Don. Duhut paet Mata katarak Daun direbus diminum
10. Cannaceae Canna indica L. Bungo terompet Demam Daun dimandikan
11. Capparaceae Cleome sp Ancimun Demam anak Daun dimadikan
12. Caricaceae Carica papaya L. Botik Sakit perut Daun + telur ayam kampung digosok
Panas dalam Buah dimakan
Masuk angin Daun diperas + bawang
diusap ke perut
Malaria Daun + daun senduduk
direbus diminum

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
ii
No. Famili Nama ilmiah Nama Lokal Kegunaan Cara meramu
13. Clusiaceae Garcinia x
mangostanaL. Manggis Mabuk jengkol
Kulit buah diperas diminum
14. Compositae Ageratumconyzoides
(L).L.
Angur-angur/rumput
nasi Masuk angin
Daun digosok oles ke perut
Blumea balsamifera
(L.) DC. Galunggung Luka
Daun diperas ditempel ke bagian luka
Mengeluar-kan darah kotor
pasca persalinan
Daun diuapkan
Dichro cephala latifolia
DC Tete babi Demam Daun direbus
Eupatorium perfoliatum
L. Angur-angur
(S.Jae) Luka
Daun dikunyah tempel ke luka
Eupatorium sp Galah nagori Luka Daun diperas ditempel
ke luka
Eupatorium sp Hayu sigodang Digigit
serangga Daun digosok
Luka Daun digosok
Mikania
micranthaKunth. Siroppaspara
Masuk anginLuka
Daun direbusPucuk daun digiling+ kemiri
ditempel
Oldenlandia sp Hayu gota Pencahar Daun direbus lalu
diminum
Soncus oleraceus L. Sijungkut Mata Daun dilalap
Eupatorium sp Silonggom
banua Masuk angin
bayi Daun direbus
Darah tinggi Kulit batang direbus
Diabetes Daun diminum
15. Convolvulaceae Ipomoea sp Andor siang Pencahar Daun diminum
16. Crassulaceae Kalanchoe pinnata
(Lam.) Pers. Dingin-dingin Demam anak Daun dikompres
17. Cucurbitaceae Cucurbita moschata
Duchesne Seno godang Maag Buah direbus bulat-bulat
Sechium edule
(Jacq)Sw. Seno ropa
Tekanan darah tinggi
Buah diparut lalu diminum
18. Cyperaceae Kylinga monocepala
Rottb. Ria-ria Luka Akar ditempel
19. Euphorbiaceae Aleurites
moluccanus(L.) Willd. Tanaon Benjol Biji ditempel
Euphorbia hirta L. Simargalagala Sakit perut Daun digosok dioles ke
perut
Jatropha curcas L. Dulang-dulang Panas dalam Getah buah dioles
Phyllanthus niruri L. Bunga
belimbing/ sidukung anak
Masuk angin Daun digosok dioles ke
perut
Perut kram Akar, batang, daun direbus diminum
Bisul Daun dihaluskan ditempel ke bisul

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
iii
No. Famili Nama ilmiah Nama Lokal Kegunaan Cara meramu
20. Icacinaceae Platea latifolia Blume Sitopu Mata Daun direbus
21. Lamiaceae Clerodendrum sp Sarang banua Sakit perut Daun ditempel
Orthosiphon stamineus
Benth. Kumis kucing Sakit perut Daun direbus
Susah buang
air kecil Daun dikeringkan
diseduh menjadi teh
22. Lauraceae Cinnamomum
burmanni (Nees & T.Nees) Blume
Kulit manis Penghangat
tubuh Kulit manis direbus
Penambah
nafsu makan Kulit manis dikunyah
Mengeluarkan darah kotor
pasca melahirkan
Daun dibakar asap diuapkan
Litsea resinosa Blume Modang lendir Panas dalam Getah batang dioles
Litsea sp Modang sauh Sakit perut Daun direbus
23. Leguminos Erythrina variegata L. Dapdap Kurap Kulit batang digosok
Leucaena glauca (L.)
Benth Potai Mata Daun muda dilalap
Mimosa invisa Colla Sirput Diabetes Daun direbus
Bisul Daun ditumbuk ditempel
24. Liliaceae Allium fistulosum L. Bawang prei Pundial ulat
bulu Daun digosok
Allium sp Gangra Masuk angin Daun dioles ke badan
25. Malvaceae Hibiscus rosa sinensis
L. Bungo raya Demam anak
Daun diremas+air dingin dimandikan
Sida rhombifolia L. Sibaguri Rematik Batang + kulit lancat direbus daun dilalap
Sorghum propinguum
(Kunth) Hitchc. Panggu Mabuk jengkol
Akar direbus lalu diminum
Urena lobata L. Rumput
sampiluhut Demam Daun dimandikan
26. Melastomatacea Melastoma
malabathricumL. Senduduk Luka Daun digosok dan dioles
Malaria Daun +daun papaya
direbus
Digigit pacet Daun digosok
27. Meliaceae Lansium domesticum Corr. var domesticum
Lancat Gatal-gatal Kulit kayu digosok
Malaria Kulit buah + batang
nangka + aren + garam direbus
Asam lambung Kulit buah + kulit mangga direbus
Perut kembung Kulit batang direbus
Sakit perut Daun direbus
28. Moraceae Artocarpusheterophyllu
sLam Sibodak Malaria
Daun direbus lalu dimandikan
29. Musaceae Musa sp Pisang sibosi
Menghilangkan darah kotor
pasca persalinan
Batang dibuat perapian
Musa sp Pisang sililit Panas dalam Air batang pisang
diminum
Musa acuminata x Pisang sitabar Sakit kepala Daun diiris ditempel

Pemanfaatan Tumbuhan Obat Secara Empiris... Nasution et al.
iv
No. Famili Nama ilmiah Nama Lokal Kegunaan Cara meramu
balbisiana cv. Saba
Musa sp Pisang
simanisan Menghilangkan
bekas cacar Kulit buah ditempel
30. Myrtaceae Melaleuca leucadendra
(L.) L. Kayu putih Gatal-gatal Kulit kayu digosok
Syzigium aromaticum
(L.) Merr. & Perry Cengkeh Batuk
Biji dihaluskan + air panas diminum
Gigi berlubang Langsung ditempel
Stamina Daun direbus + kayu
manis direbus dimandikan
Syzigium aqueum (Burm.F.) Alston
Jambu aek Diare Daun direbus lalu
diminum
31. Piperaceae Piper aduncum L. Hayu sirih Diare Daun direbus
Rematik Daun direbus
Piper betle L. Burangir Penambah
darah Daun dikunyah
Batuk Daun direbus diminum
Mata Mata dicelupkan ke
rebusan daun
32. Poaceae Chrysopogon
zizanioides (L.) Roberty
Minyak wangi/ sinauskus/
tomus-tomus Sakit perut
Daun digosok akar +poga (Physalis
angulata) direbus
Digigit
serangga Daun dihaluskan lalu
dioles
33. Polygonaceae Calligonum sp Rumput Fatimah Diabetes Daun direbus
34. Phyllanthaceae Bischofia javanica
Blume Singkam Diabetes Pucuk daun dikunyah
35. Solanaceae Capsicum annuum L Lasiak Pingsan Daun + kunyit + beras + air hangat dimandikan
Capsicum frutescens L. Lasiak cino/suri Sakit perut Daun direbusDaun
ditempel
Solanum torvum Sw. Inggir-inggir/
rimbang MataKeracunan
Daun dilalapDaun direbus
Racun
serangga Buah matang dioles ke bagian yang kena racun
Centella asiatica (L.)
Urb Paga-paga Kurang gizi
Daun diperas lalu ditempel
Asam urat Daun direbus
Panas dalam Daun ditempel
Physalis angulata L Pultak-
pultak/poga Diabetes
Daun dikeringkan dibuat the
36. Thymelaeaceae Phaleria macrocarpa
(Scheff.) Boerl. Mahkota dewa Asam urat
Biji matang direbus diminum airnya
37. Umbelliferae Apium graveolens L. Daun sop Darah tinggi Umbi direbus dimakan
38. Zingiberaceae Alpinia galanga (L.)
Willd. Alas Gatal kulit
Rhizoma dihaluskan dan ditempel
Curcuma domestica
Vahl. Nagorsing/
hunik Sakit perut
Rhizoma ditumbuk lalu diperas airnya
Pingsan Rhizoma diiris lalu
disiram dan diminum
Diare Rhizoma dikunyah
Sakit kepala Rhizoma + gula merah
digiling langsung dimakan

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
v
No. Famili Nama ilmiah Nama Lokal Kegunaan Cara meramu
Kaempferia galanga L. Hasihor Batuk anak Rhizoma ditumbuk diikat ke leher menggunakan
kain
Batuk dewasa Rhizoma langsung
dimakan
Zingiber officinale
Roscoe. Pege Batuk Rhizoma direbus
Zingiber purpureum
Roxb. Hunik bungle
Mengeluarkan darah kotor
pasca persalinan
Rhizoma +Silinjuang (Cordyline fruticosa) dimasukkan dalam tanah lalu dibakar
Zingiber zerumbet (L.)
Roscoe ex Sm. Lempuyang
Pundial setan (darah
tersumbat)
Daun direbus lalu ditulbas (istilah lokal
Mandailing untuk disembur)

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
i
LAMPIRAN 2. ANALISIS BIOINFORMATIKA BERBASIS WEB PADA SEKUEN GENOM PARSIAL SAGU (Metroxylon sagu Rottb.)
Lampiran 2.1. Tabel rangkuman analisis sekuen genom sagu dengan Blast2Go
No. Sekuen Deskripsi Hit E-value Rataan Kemiripan (%)
GO ID Kode Enzim
Nama Enzim InterPro GO ID
1 sc-678470 D -box ATP-dependent RNA helicase D 1 isoform X2
10 3.67E-11 85.4 F:GO:0003676; F:GO:0005524; F:GO:0008026
EC:3.6.1.3; EC:3.6.1.15
Adenosinetriphosphatase; Nucleoside-triphosphate phosphatase
no IPS match
2 sc-16071 glutamate decarboxylase 10 2.67E-71 99.6 F:GO:0030170; P:GO:0006536; F:GO:0004351
EC:4.1.1.15 Glutamate decarboxylase
F:GO:0030170; P:GO:0006536; F:GO:0003824; F:GO:0004351; P:GO:0019752; F:GO:0016831
3 sc-400785 E3 ubiquitin- ligase RF298 10 1.39E-13 80.8 C:GO:0016020
no GO terms
4 sc-7173 reverse transcriptase 10 1.22E-47 63
no GO terms
5 sc-7844 probable glutamate carboxypeptidase 2
10 6.59E-61 88.8
no GO terms
6 sc-2006 E3 ubiquitin- ligase ATL4-like
10 1.04E-48 69 F:GO:0046872; C:GO:0016020
no GO terms
7 sc-646 probable WRKY transcription factor 49
10 6.56E-67 69.5 F:GO:0003700; P:GO:0006355; F:GO:0043565
F:GO:0003700; P:GO:0006355; F:GO:0043565
8 sc-328094 probable histidine kinase 4 10 1.71E-72 88.6
P:GO:0016310; F:GO:0016772
9 sc-732306 probable CCR4-associated factor 1 homolog 7
10 5.63E-74 75.6 F:GO:0003676; C:GO:0005634
F:GO:0003676
10 sc-72 PREDICTED: uncharacterized protein LOC103712229
10 0 76.8
no GO terms

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
ii
No. Sekuen Deskripsi Hit E-value Rataan Kemiripan (%)
GO ID Kode Enzim
Nama Enzim InterPro GO ID
11 sc-25114 clathrin heavy chain 2-like 10 1.06E-34 89.6
P:GO:0016192; C:GO:0030130; C:GO:0030132; F:GO:0005198; P:GO:0006886
F:GO:0005488
12 sc-1642 prostatic spermine-binding -like isoform X2
10 1.06E-18 83.8
no GO terms
13 sc-44990 OTU domain-containing DDB_G0284757
10 7.37E-13 84
no GO terms
14 sc-296883 pentatricopeptide repeat-containing chloroplastic-like
10 2.74E-60 65.5
no GO terms
15 sc-109470 pyruvate dehydrogenase E1 component subunit beta- mitochondrial isoform X2
10 1.15E-08 45.1
P:GO:0006086; F:GO:0003824; F:GO:0004739; P:GO:0008152; P:GO:0055114
no IPS match
16 sc-76763 NAC domain-containing 30-like
10 3.67E-28 73.1
F:GO:0003677; C:GO:0005634; P:GO:0006351; P:GO:0006355
no IPS match
17 sc-9317 NAC transcription factor 29-like
10 3.76E-23 63.4
F:GO:0003677; C:GO:0005634; P:GO:0006351; P:GO:0006355
no IPS match
18 sc-2590 reverse transcriptase 10 4.69E-36 53.4 F:GO:0003676; P:GO:0015074
no IPS match
19 sc-114 Retrovirus-related Pol poly from transposon TNT 1-94
10 1.10E-08 60 F:GO:0046872
no IPS match
20 sc-445809 retrotransposon unclassified 10 3.86E-10 59.8 F:GO:0003676
no IPS match
21 sc-3836 PREDICTED: uncharacterized protein LOC103715697
10 2.35E-92 49.5
no IPS match

J Bioteknol Biosains Indones – Vol 5 No 1 Thn 2018
iii
No. Sekuen Deskripsi Hit E-value Rataan Kemiripan (%)
GO ID Kode Enzim
Nama Enzim InterPro GO ID
22 sc-4442 SUPPRESSOR OF ABI3-5 isoform X7
10 1.11E-37 87
no IPS match
23 sc-2283 nuclear transcription factor Y subunit B-2 isoform X2
10 6.50E-50 81.6
C:GO:0005634; F:GO:0046982; P:GO:0006355; F:GO:0043565
F:GO:0046982
24 sc-7313 ECERIFERUM 3-like 10 2.64E-49 63.1
F:GO:0000254; F:GO:0005506; F:GO:0016491; C:GO:0016020; C:GO:0016021; P:GO:0055114; P:GO:0006633
no GO terms
25 sc-887427 serine threonine- kinase HT1-like
10 1.92E-38 92.2 F:GO:0005524; F:GO:0004672; P:GO:0006468
no GO terms
26 sc-5356 chaperone dnaJ 16 10 6.97E-08 83.7
no GO terms
27 sc-2587 BURP domain-containing 5-like
9 7.82E-06 71.67
no IPS match
28 sc-2223 PREDICTED: uncharacterized protein LOC108511316
6 9.38E-05 68.5
no IPS match
29 sc-250939 auxin-responsive IAA6 isoform X1
5 1.11E-23 70.8
C:GO:0005634; P:GO:0006355
30 sc-198453 GPI-anchored 58 isoform X2 4 7.21E-06 82.25
no IPS match
31 sc-5732 zinc finger CCCH domain-containing 34-like
2 1.19E-07 71 F:GO:0046872
no IPS match
32 sc-100242 glutathione S-transferase F11-like
2 6.26E-05 90.5
no IPS match
33 sc-20759 PREDICTED: uncharacterized protein LOC105056183
2 4.02E-14 85
no IPS match

Analisis Bioinformatika Berbasis Web Pada Sekuen Genom Parsial Sagu...Purwoko et al.
iv
Lampiran 2.2. Grafik anotasi fungsional dari DAVID Bioinformatics Sources 6.8