
Pollen concentrations in central italy (ascoli piceno and siena
Upload
independentCategory
view
0download
0
RESEARCH ARTICLE 2221
Development 139, 2221-2233 (2012) doi:10.1242/dev.081224© 2012. Published by The Company of Biologists Ltd
INTRODUCTIONPollen dormancy and germination are crucial for reproductivegrowth of flowering plants. In Arabidopsis, mature pollen grainsare released from anthers in a partially dehydrated and dormantstate. Once they fall onto stigma, rehydration of desiccated pollengrains breaks dormancy and turns on pollen germination(McCormick, 2004). Inappropriate pollen germination willinevitably affect plant fertility, and thus studies on pollen dormancyand germination are of great significance to agricultural production.Precocious pollen germination in anthers was first observed in theArabidopsis gametophytic mutant raring-to-go (rtg), in whichpollen grain development is affected as early as the bicellular stageand affected pollen grains are unable to desiccate completely(Johnson and McCormick, 2001). In addition, rtg pollen tubes weredramatically elongated within anthers under high humidityconditions. These findings indicate the importance ofdehydration/rehydration in pollen dormancy and germination; todate, dehydration is considered to be the main factor in maintainingpollen dormancy and preventing precocious germination.
Abscisic acid (ABA) promotes and maintains seed dormancy(Finkelstein et al., 2008). Deficiencies in ABA synthesis or signalingfrequently result in reduced dormancy, seed precocious germinationor vivipary (Koornneef and Karssen, 1994; McCarty, 1995; Bewley,
1997). Supportive evidence comes from the studies of maize vp(viviparous) (Neill et al., 1986), tomato sitiens (Hilhorst and Downie,1996), rice phs (pre-harvest sprouting) (Fang et al., 2008), andArabidopsis ABA-deficient (aba) and ABA-insensitive (abi) mutants(Koornneef et al., 1989). Moreover, recent studies show that theABA content in the lily pollen is increased before anthesis, whichcorrelates with the pollen dehydration stage (Hsu et al., 2010). Aunique desiccation-associated ABA signaling transduction has beenreported, through which a Rop (Rho GTPase of plants) gene isregulated during the stage of pollen dehydration (Hsu et al., 2010).These results indicate the importance of ABA-mediated dehydrationfor pollen dormancy.
Both hydration/humidity and Ca2+ are essential for pollengermination. In Arabidopsis, pollen germination starts with therehydration of pollen grains on the stigma. During this process,Ca2+ flows into the pollen grains and triggers cytoplasmicreorganization. Subsequently, a cytoplasmic gradient of Ca2+ isformed with the highest [Ca2+] at the germination site, which iscrucial for polar tip growth of prospective pollen tubes (Heslop-Harrison and Heslop-Harrison, 1992a; Heslop-Harrison andHeslop-Harrison, 1992b; Franklin-Tong, 1999). Dynamic analysisof cytosolic [Ca2+] ([Ca2+]cyt) in Arabidopsis pollen grains revealsan increase in [Ca2+]cyt at the potential germination site soon afterrehydration; this high Ca2+ concentration is maintained until theonset of pollen germination (Iwano et al., 2004). In hydrated pollenof Nicotiana tabacum L., a high level of calmodulin (CaM) isconsistently present in the region of the germination apertures andis associated with the plasma membrane of the germination bubbleand with the onset of germination (Tirlapur et al., 1994). Highenvironmental humidity and exogenous Ca2+ are also requisites forpollen germination in vitro (Taylor et al., 1997).
National Key Laboratory of Plant Molecular Genetics, Shanghai Institute of PlantPhysiology and Ecology, Chinese Academy of Sciences, 300 Fenglin Road, Shanghai200032, China.
*These authors contribute equally to this work‡Author for correspondence ([email protected])
Accepted 26 March 2012
SUMMARYAppropriate pollen germination is crucial for plant reproduction. Previous studies have revealed the importance of dehydration inmaintaining pollen dormancy; here, we show that phosphatidylinositol pathway-controlled Ins(1,4,5)P3/Ca2+ levels are crucial formaintaining pollen dormancy in Arabidopsis thaliana. An interesting phenotype, precocious pollen germination within anthers,results from a disruption of inositol polyphosphate 5-phosphatase 12 (5PT12). The knockout mutant 5pt12 has normal earlypollen development and pollen dehydration, and exhibits hypersensitive ABA responses, indicating that precocious pollengermination is not caused either by abnormal dehydration or by suppressed ABA signaling. Deficiency of 5PT13 (a close paralogof 5PT12) synergistically enhances precocious pollen germination. Both basal Ins(1,4,5)P3 levels and endogenous Ca2+ levels areelevated in pollen from 5pt12 mutants, and 5pt12 5pt13 double mutants show an even higher precocious germination rate alongwith much higher levels of Ins(1,4,5)P3/Ca2+. Strikingly, exogenous Ca2+ stimulates the germination of wild-type pollen at floralstage 12, even in very low humidity, both in vitro and in vivo, and treatment with BAPTA, a [Ca2+]cyt inhibitor, reduces theprecocious pollen germination rates of 5pt12, 5pt13 and 5pt12 5pt13 mutants. These results indicate that the increase in thelevels of Ins(1,4,5)P3/Ca2+ caused by deficiency of inositol polyphosphate 5-phosphatases is sufficient to break pollen dormancyand to trigger early germination. The study reveals that independent of dehydration, the control of Ins(1,4,5)P3/Ca2+ levels byInositol polyphosphate 5-phosphatases is crucial for maintaining pollen dormancy.
KEY WORDS: Inositol polyphosphate 5-phosphatase (5PT), Cytosolic Ca2+, Pollen dormancy, Inositol 1,4,5-trisphosphate [Ins(1,4,5)P3],Arabidopsis thaliana
Inositol polyphosphate 5-phosphatase-controlledIns(1,4,5)P3/Ca2+ is crucial for maintaining pollen dormancyand regulating early germination of pollenYuan Wang*, Yu-Jia Chu* and Hong-Wei Xue‡
DEVELO
PMENT

2222
Inositol polyphosphate 5-phosphatase (5PT), a key enzyme ofthe phosphatidylinositol (PI) signaling pathway, functions ingrowth, development and responses to stress in plant, yeast andmammals (Stevenson et al., 2000; Astle et al., 2006; Xue et al.,2007; Xue et al., 2009). 5PT removes the phosphate at the 5-position of the inositol ring of specifically phosphorylated (5-position) phosphoinositides [PtdIns(4,5)P2, PtdIns(3,4,5)P3 and/orPtdIns(3,5)P2], and inositol 1,4,5-trisphosphate [Ins(1,4,5)P3] orinositol 1,3,4,5-tetrakisphosphate [Ins(1,3,4,5)P4].
There are 15 5PTs in Arabidopsis (Berdy et al., 2001; Ercetinand Gillaspy, 2004; Zhong and Ye, 2004) that have diverse roles inplant growth and development, and phytohormone signaling.5PT13 modulates cotyledon vein development through theregulation of auxin homeostasis (Lin et al., 2005). FRAGILEFIBER 3 (FRA3) and COTYLEDON VASCULAR PATTERN 2(CVP2) are two 5PTs containing the 5PTase domain. FRA3 isrequired for secondary wall synthesis and actin organization infiber cells (Zhong et al., 2004), and CVP2-mediated Ins(1,4,5)P3
signal transduction is essential for closed venation patterns ofArabidopsis foliar organs (Carland and Nelson, 2004). In addition,5PTs participate in seedling growth (Gunesekera et al., 2007;Ercetin et al., 2008) and blue-light signaling via the regulation ofIns(1,4,5)P3/[Ca2+]cyt (Chen et al., 2008), and are importantregulators of Ins(1,4,5)P3-mediated ABA signaling (Sanchez et al.,2001; Burnette et al., 2003; Lin et al., 2005; Gunesekera et al.,2007). However, no 5PTs have been reported to function in theregulation of reproductive development.
Here, through the functional characterization of Arabidopsis5PT12 and 5PT13, we show that the increase of 5PT-controlledIns(1,4,5)P3/Ca2+ is sufficient for breaking pollen dormancy andtriggering early germination, and further confirm that,independently of dehydration, 5PT-controlled Ins(1,4,5)P3/Ca2+ iscrucial for pollen dormancy.
MATERIALS AND METHODSPlant materials and growth conditionsThe Arabidopsis Columbia ecotype was used. The 5pt13 mutant has beendescribed by Lin et al. (Lin et al., 2005). Seedlings were grown in aphytotron at 22°C with a 16-hour light/8-hour dark photoperiod.
Identification of 5pt12 and 5pt14 knockout mutantThe 5pt12 mutant was obtained from the Arabidopsis Biological ResourceCenter (SALK_065920, http://www.arabidopsis.org/). 5pt12 mutant seedswere screened on MS medium containing 30 mg/l kanamycin forsegregation ratio analysis and a PCR-based approach was employed toconfirm the T-DNA insertion and to identify the homozygous mutant.Genotyping primers 5PT12-F (5�-CACAGCCTCGACGACATTC-3�),5PT12-R (5�-TTGCCAAGCTAGACCTTCC-3�) and LBa1 (5�-TGGTTCACGTAGTGGG CCATCG-3�) were used.
The 5pt14 mutant was obtained from the Arabidopsis BiologicalResource Center (GK-309D03, http://www.arabidopsis.org/). Genotypingprimers 5PT14-F (5�-GGTGGTGTAAGAGGATGGT-3�), 5PT14-R (5�-CGGCACTTCCTTCAACCC-3�) and GK-LB (5�-ATATTGA -CCATCATACTCATTGC-3�) were used to confirm the T-DNA insertionand to identify the homozygous mutant.
Quantitative real-time RT-PCR (qRT-PCR) analysisqRT-PCR was performed to test the expression patterns of 5PT12, 5PT13and 5PT14 in various tissues, the transcription levels of 5PT12 and 5PT14in mutant or wild-type plants, and the expression of ABA-responsive genesin seedlings or flower tissues.
For analyzing the expression pattern of 5PT12, total RNAs wereextracted from 4-day-old seedlings, roots, 4-week-old rosette leaves,flowers and stems. For analyzing the expression pattern of 5PT12, 5PT13and 5PT14 at different floral stages, total RNAs were extracted from
flowers at stages 12, 13 and 14. For testing the transcriptional levels of5PT12 and 5PT14 in mutant or wild-type plants, total RNAs were extractedfrom flowers of 4-week-old plants. For examining the expressions of ABA-responsive genes in flower tissues or seedlings, flowers of 4-week-oldplants at floral stages 12-14 or an earlier stage were collected andsubmerged in ABA (100 M) solution for 0, 1 or 3 hours, and 8-day-oldwild-type and 5pt12 seedlings were submerged in ABA (100 M) solutionfor 0, 0.5, 1 or 2 hours.
Total RNAs were extracted using Trizol reagent (Invitrogen) and thenreversely transcribed according to the manufacturer’s instructions(ReverTra Ace kit, Toyobo). RNA samples were incubated for 30 minutesat 37°C with RNase-free DNase I (Takara), and then precipitated for furtheranalysis.
RotorGene RG-3000A (Corbett Research) and a SYBR green detectionkit (SYBR Green Realtime PCR Master Mix kit; TOYOBO) were used,with a program of denaturation (5 minutes, 95°C), 40 cycles ofdenaturation (95°C, 10 seconds), annealing (55°C, 15 seconds) andelongation (72°C, 20 seconds). Fluorescence is monitored during eachannealing and denaturation phase. The product amounts were determinedby the method of comparative Delta-Delta Ct (Fleige et al., 2006). Primersused were as follows: 5PT12 (5�-AGAATCGGTAGGAGGAAACG-3� and5�-TTCTCAGATTCTTCCTCACC-3�), 5PT13 (5�-CCGAGGAC -AAAATCTAAGTAACG-3� and 5�-CAGCGCCGGTGCTTGGAATAG-3�), 5PT14 (5�-GGTGGTGTAAGAGGATGGT-3� and 5�-CGGCACTTCCTTCAACCC-3�), ABA-responsive genes Rd22 (5�-TTACCAAACACTCCCATTCC-3� and 5�-CGTACACC TCCCT -TTCCAAC-3�), COR47 (5�-GAACAAGCCTAGTGTCATCG-3� and 5�-TCTTGTCGTGGTGTCCTGG-3�), KIN2 (5�-ACCAACAA -GAATGCCTTCCA-3� and 5�-ACTGCCGCATCCGATATACT-3�) andCOR15 (5�-CTCAGTTCGTCGTCGTTTC-3� and 5�-CATC -TGCTAATGCCTCTT T-3�). Arabidopsis ACTIN gene (At5g09810) wasamplified with primers (5�-CCGGTATTGTGCTCGATTCTG-3� and 5�-TTCCCGTTCTGCGGTAGTGG-3�) and used as positive internal control.Primers of 5PT13 have been previously described (Chen et al., 2008). Allthe experiments were repeated three times.
Promoter-reporter gene fusion studiesThe genomic DNA region in front of the translation initiation site ATG wasPCR amplified with primers 5PT12p-1 (5�-CCCAAGCTTTAAGGCAGTGTCTGTCGAGG-3�, added HindIII siteunderlined) and 5PT12p-2 (5�-CGGGATCCGGAAGAGAGA -ATGAGAGAAATG-3�, added BamHI site underlined). The amplified1216 bp DNA fragment was confirmed by sequencing and then subclonedinto modified pCAMBIA1301 (Liu et al., 2003), resulting in the constructcontaining 5PT12 promoter-GUS (-glucuronidase) fusion. The resultantconstruct was transferred into Agrobacterium tumefaciens strain GV3101and then transformed into Arabidopsis using the floral dip method. T1transgenic seedlings were screened on MS medium supplemented with 30mg/l hygromycin and the T-DNA integration was confirmed by PCRanalysis using primers annealing to the hygromycin resistance gene (hyg-s, 5�-GCTTCTGCGGGCGATTTGTGT-3� and hyg-a, 5�-GGTCGC -GGAGGCTATGGATGC-3�). Seedlings of T3 homozygous plants wereanalyzed for GUS activities and observed using a SMZ 800 stereoscope(Nikon).
The transgenic seedling harboring a 5PT13 promoter-GUS reporter geneconstruct has been described previously (Lin et al., 2005).
Complementary expression of 5PT12 in 5pt12 mutantA 7.4 kb fragment of genomic DNA containing the full-length 5PT12 geneand the promoter region was cut from a BAC clone F6E13 (from ABRC,http://www.arabidopsis.org) and subcloned into modified pCAMBIA1301vector (Liu et al., 2003). The resultant construct was transferred into the5pt12 mutant by Agrobacterium-mediated transformation through the floraldipping method. Complemented expression of 5PT12 was confirmedthrough qRT-PCR using primers (5�-AGTACTTCTCCCACCACTTG-3�and 5�-TCTGAACATGACCACAAATCG-3�). T2 homozygous plantswere used for complementation studies.
RESEARCH ARTICLE Development 139 (12)
DEVELO
PMENT

Genetic cross and characterization of 5pt12 5pt13 double mutantTo generate 5pt12 5pt13 double mutant by genetic cross, 5pt13homozygotes (carrying Basta-resistant gene) were used as female parent,and 5pt12 homozygotes (carrying kanamycin-resistant gene) were used asmale parent. The cross was carried out by removing the petals, sepals andandroecia from large green buds in 5pt13 plants, followed by artificialfecundation with the 5pt12 pollen grains on the next day. F1 crossprogenies were selected on kanamycin-containing MS plates and positiveplants were used for harvesting F2 seeds. F2 generation plants were usedfor PCR analysis to confirm the T-DNA insertions in 5PT12 gene loci and5PT13 gene loci. Identified homozygotes were grown for one additionalgeneration and harvested seeds were used for further experiments.
Seed germination assaySurface-sterilized seeds of wild-type or 5pt12 plants were sown onto MSmedium containing different concentrations of ABA (0, 0.1, 0.3 and 0.5M). Following stratification in 4°C for 2 days, seeds were grown at 22°Cfor 7 days and germination rates were calculated. One-hundred seeds wereused for each line, and all the assays were performed in triplicate. Assayswere repeated at least twice.
Scanning electron microscope analysisDehiscent anthers of wild-type and 5pt12 plants were harvested and fixedby FAA, followed by dehydration with a graded ethanol series. Specimenswere dried at crucial points and coated with gold before examination witha Scanning Electron Microscope (JSM-6360LV, Tokyo, Japan).
Observation and measurement of dehydrated pollen grainsDehydrated pollen grains were stripped out from dehiscent anthers of wild-type and 5pt12 plants and covered with type N immersion oil to retain theiroriginal shapes. Image J software was used to measure major and minoraxes of dehydrated pollen grains, and the ratio of minor axes/major axeswas calculated. More than 50 pollen grains of each genotype wereanalyzed, and the assays were repeated twice.
Measurement of pollen viability and DAPI stainingFlower buds prior to opening were harvested, and dispersed pollen grainswere treated with FDA (fluorescein diacetate, diluted with acetone to aconcentration of 100 mg/l) for 30 minutes, and then observed in ultravioletlight with a microscope (Leica).
Pollen harvested from open flowers was treated with DAPI solution(final concentration of 2 mg/l dissolved in 7% sucrose, w/v) for 5 minutesand imaged with a confocal laser scanning microscope (FITC488, ZeissLSM500).
Pollen germination in vitro and in vivoOpen flowers from wild-type and 5pt12 mutant plants were dehydrated ina desiccator at room temperature for at least 2 hours. Pollen germinationwas performed as described by Li et al. (Li et al., 1999) and 2 mM Ca2+
(with an equal molar ratio of CaCl2 and Ca[NO3]2) was added into basalmedium to obtain the optimal pollen germination rate. Soaked filter paperwas used to provide environment humidity. After incubation for 6 hours at25°C, germinated pollen were observed and photographed.
To examine the effect of extracellular Ca2+ on pollen germination, pollenwas cultured in basal agar medium supplemented with differentconcentrations of Ca2+ [with an equal molar ratio of CaCl2 and Ca(NO3)2].For the humidity assay, dry filter paper was used to provide a coarsegradient of environmental humidity conditions by adding different volumesof distilled water onto filter papers. 0, 2.3, 4.6 or 7 ml of distilled waterwas added to five pieces of dry filter paper to provide very low (~0-10%),low (~25-35%), middle (~55-65%) or saturation (high humidity, ~85-95%).Pollen grains (>200) collected from more than three individual plants wereused for each genotype, and each assay was performed in triplicate. Assayswere repeated at least twice.
Different concentrations of Ca2+ [with an equal molar ratio of CaCl2 andCa(NO3)2] were applied to anthers at floral stage 11 to examine the pollengermination in anthers. BAPTA (2 mM, Sigma-Aldrich) was used to block
the cytosolic Ca2+ in anthers of 5pt12, 5pt13 and 5pt12 5pt13 mutants. Atlate floral stage 12, Ca2+- or BAPTA-treated anthers were harvested forpollen tube staining by DAB assay.
Pollen tube staining with Aniline BlueDehiscent anthers or dispersed pollen grains from wild-type and 5pt12,5pt13, 5pt12 5pt13, 5pt14 and 5pt12 plants with complementary expressionof 5PT12 were treated with 0.1% decolorized Aniline Blue (DAB) for 5minutes followed by observation in ultraviolet light with a microscope(Leica).
Quantification of ABA contentAnthers at floral stage 12 or anthers at earlier stage (pollen were too smallto collect) were collected for ABA measurement using liquidchromatography-mass spectrometry (LC-MS) (Welsch et al., 2008).
Measurement of Ins(1,4,5)P3 contentStamens at floral stages 12-14 from wild-type, 5pt12, 5pt13, 5pt12 5pt13and 5pt12 plants with complementary expression of 5PT12 were harvestedfor measuring Ins(1,4,5)P3 content. Extraction of Ins(1,4,5)P3 wasperformed according to a previously published method (Burnette et al.,2003). Ins(1,4,5)P3 content was measured using the D-myo-inositol-1,4,5-trisphosphate [3H] assay kit (GE healthcare) and determined from astandard curve generated with known amounts of Ins(1,4,5)P3.
For inositol phosphates (IPs) control assay, flower tissues at floral stages12-14 of wild-type, 5pt12, 5pt13, and 5pt12 5pt13 plants were collectedand used for the assay. A mixture of unlabelled PtdIns(4,5)P2, Ins(1,3,4)P3
and InsP6 were added into assays. As carried out previously (Perera et al.,2008), PtdIns(4,5)P2, InsP6 and Ins(1,3,4)P3 were added, respectively, at18�, 10� and 1� the concentration of Ins(1,4,5)P3 found in wild-typestamen tissues. Assays were performed in triplicate and experiments wererepeated twice.
Measurement of calcium content using ICP-MSAnthers at floral stages 11-13 from wild-type, 5pt12, 5pt13 and 5pt125pt13 plants were harvested, following addition of 1 ml 100% HNO3 andkept overnight. The samples were kept at 99°C for 1 hour and diluted to14 ml with ultra-pure water. Samples were analyzed using an ELAN DRC-e ICP-MS system (PerkinElmer) and Multi-Element calibration Standard-2A (Agilent) samples were used as standards. Assays were performed intriplicate.
Indo-1 staining and examination of [Ca2+]cyt in microspores, pollengrains and root hair cellsMicrospores or pollen grains at different floral stages from wild-type,5pt12, 5pt13, and 5pt12 5pt13 plants were stained with 20 M Indo-1(57180, Sigma) on slides in the dark. Two hours after loading and washingof the dye, fluorescence was observed with a confocal SMZ510 under UVlight excitation (364-nm excision, and 400- to 435-nm and 480 nmemission) and imaged. [Ca2+]cyt was pseudocolored according to the scale.Pseudocolor ratio images of the [Ca2+]cyt distribution was calculated withOlympus Fluoview Ver. 1.7a Viewer tool (Olympus) to measure thefluorescence intensity. White indicates approximately twice the [Ca2+]cyt ofthat indicated by the blue and 1.5 times the [Ca2+]cyt indicated by theyellow. For the root hair plasmolysis, 7-day-old seedlings of wild-type,5pt12, 5pt13, and 5pt12 5pt13 plants were stained with Indo-1, followedby treatment with 500 mM mannitol for 10 minutes.
A pollen protoplast staining assay was performed as describedpreviously (Wu et al., 2011). Pollen grains at different floral stages fromwild-type, 5pt12, 5pt13, and 5pt12 5pt13 plants were harvested for theassay.
Accession numbersSequence data from this article can be found in the GenBank/EMBL datalibraries under the following accession numbers: 5PT12 (At2g43900),5PT13 (At1g05630), 5PT14 (At2g31830), KIN2 (At5g15970), COR15(At2g42540), RD22 (At5g25610) and COR47 (At1g20440).
2223RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
DEVELO
PMENT

2224
RESULTSThe Arabidopsis 5PT12 gene is preferentiallyexpressed in pollen grains at floral stages 12-14Our previous studies have shown that inositol polyphosphate 5-phosphatase 13 (5PT13) plays crucial roles in auxin-regulatedcotyledon vein development (Lin et al., 2005), blue light-mediatedphotomorphogenesis (Chen et al., 2008) and root gravitropism(Wang et al., 2009). To further characterize the 5PT functions inArabidopsis, we focused on 5PT12, a close paralog of 5PT13(because the mutant of the closest paralog, 5PT14, was notavailable when we started). Based on domain prediction, 5PT12has a conserved 5-phosphatase domain (5PTase domain), which isrequired for 5-phosphatase activity towards inositol phosphates andphosphoinositides, and three WD40 repeats (supplementarymaterial Fig. S1A). Phylogenetic analysis of 15 Arabidopsis 5PTsand representative yeast and mammalian 5PTs showed that 5PT12is most closely related to 5PT13, 5PT14 and FRA3 (supplementarymaterial Fig. S1B).
The gene expression pattern of 5PT12 in various tissues wasanalyzed using quantitative real-time RT-PCR (qRT-PCR) and theresults showed that 5PT12 was mostly expressed in leaves andflowers, whereas it was only weakly expressed in roots, stem andyoung seedlings (Fig. 1A). To investigate the detailed expressionpattern, a construct containing the 1.2 kb 5PT12 promoter regionfused to the Escherichia coli -glucuronidase (GUS)-coding genewas generated and transformed into Arabidopsis. Consistent resultsfrom five independent transgenic lines showed that 5PT12 istranscribed in cotyledon tips and the hydathode of leaves (Fig.1Ba,b).
Interestingly, 5PT12 is preferentially expressed in pollen grainsat floral stages (Smyth et al., 1990; Sanders et al., 1999) 12 to 14(Fig. 1Bc-f). At stage 13, the filaments have elongated to the heightof the stigma, mature pollen and pollen tubes (Fig. 1Bg,h),suggesting a potential role for 5PT12 in pollen development andgermination. qRT-PCR analysis indicated the gradually increasedexpression of 5PT12 as floral development proceeded (from floralstage 12 to 14, Fig. 1C), which is consistent with the microarraydata (https://www.genevestigator.ethz.ch/) showing that the 5PT12gene is largely expressed in anthers, with high transcript abundancein tri-cellular pollen and mature pollen, and low abundance in uni-cellular microspore and bi-cellular pollen. Furthermore, 5PT12,5PT13, 5PT14 and 5PT5 are also highly expressed in pollen grains,based on the microarray data (supplementary material Table S1;qRT-PCR analysis confirmed relatively less expression of 5PT13in comparison with 5PT12, Fig. 1C). As the 5pt14 mutant was notavailable and 5PT5 is not a close paralog to 5PT12, 5PT13 and5PT14, we first focused on the function of 5PT12 in pollendevelopment.
Deficiency of 5PT12 results in precocious pollengermination within anthersA putative T-DNA insertion mutant of 5PT12 was obtained fromSALK, and PCR analysis confirmed the T-DNA insertion at thefirst exon of the 5PT12 (Fig. 1E). The homozygote wascharacterized and designated as a 5pt12 mutant. Both forward andreverse gene-specific genotyping primers could amplify the PCRproduct when combined with T-DNA border primer LBa1 (Fig.1E), indicating that two tandem T-DNA insertions are located back-to-back in 5PT12. This was further confirmed by the correspondingT-DNA flanking sequences. Back-cross between 5pt12 and wild-type plants was performed (supplementary material Fig. S2A) andsegregation analysis of F2 progenies of a single 5PT12/5pt12 F1
plants by PCR amplification of NPTII (selectable marker locatedin T-DNA for resistance to kanamycin) showed that 30 out of 41progenies carried NPTII (supplementary material Fig. S2B). Inaddition, scoring for kanamycin (Kan) resistance showed that 212F2 seedlings were Kan resistant and 72 were Kan sensitive. A chi-square test for both assays confirmed the 3:1 segregation ratio.These results validated that the two T-DNA insertions are closelylinked without segregation. Transcription analysis by qRT-PCRshowed that the 5PT12 gene was completely disrupted (Fig. 1F),indicating that 5pt12 is a knockout mutant.
Owing to the predominant expression of 5PT12 in pollengrains at floral stages 12-14, in mature pollen and in pollentubes, the function of 5PT12 in pollen development andgermination was specifically examined. Phenotypic observationshowed that, unlike wild-type dehiscent anthers, in which veryfew pollen grains remained (floral stage 13), many more pollengrains were left in the 5pt12 dehiscent anthers, and they hadtube-like structures (Fig. 1G). Pollen grains were washed off andtube-like structures could be easily distinguished (Fig. 1H).Further staining with decolored Aniline Blue (DAB), a widelyused dye for pollen tube observations (Escobar-Restrepo et al.,2007), confirmed that the tube-like structures are indeed pollentubes (Fig. 1I).
The precocious pollen germination rates of 5PT12/5pt12heterozygotes were 20-30% and those of 5pt12 homozygotesreached 40-70% in comparison with the rate of 1-3% in wild-typeanthers (Fig. 1J), indicating that 5PT12 deficiency resulted inprecocious pollen germination within the anther. This is a veryinteresting finding, because, in Arabidopsis, only two mutants(raring-to-go and callose synthase 9) and one transgenic lineoverexpressing callose synthase 5 have been reported so far toshow a similar phenotype with unknown mechanism (Johnson andMcCormick, 2001; Xie et al., 2010).
The link between T-DNA insertion in 5PT12 gene and theprecocious germination phenotype was further tested usingrandomly selected 40 progenies of one single 5pt12 plant. PCRanalysis using the genotyping primers confirmed that all the 40progenies were homozygotes (supplementary material Fig. S2C)and all exhibited precocious pollen germination at a rate of 40-70%. Complementation studies were performed by transferringa 7.4 kb fragment of genomic DNA containing the entire 5PT12gene and promoter region into the 5pt12 mutant (Fig. 1F). Theresults showed that the Aniline Blue fluorescence was barelydetectable in complementation lines (Fig. 1I) and the precociouspollen germination rates of complementation lines (T2homozygous lines) were also similar to those in wild type (Fig.1J), confirming that 5PT12 deficiency is responsible forprecocious pollen germination.
Normal pollen development and unaffectedfertility in 5pt12 mutantTo investigate whether there are developmental defects in 5pt12that cause precocious pollen germination, pollen development of5pt12 was examined. The observations showed the normaldevelopment of 5pt12 pollen, and the DAPI staining assay revealedthat precociously germinated pollen grains within 5pt12 anthershave regular tri-cellular structures, similar to wild-type pollengrains (Fig. 2A, left panel). Further analysis of pollen viability withFDA staining showed that 5pt12 pollen grains wereindistinguishable from wild type (Fig. 2A, right panel). Besides,mature pollen grains of 5pt12 that did not germinate precociouslywithin the anther could germinate well in vitro (Fig. 2B), and the
RESEARCH ARTICLE Development 139 (12)
DEVELO
PMENT

2225RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
Fig. 1. 5PT12 is preferentially expressed in leaf and mature pollen grains, and knockout expression of 5PT12 results in precociousgerminated pollen grains within anthers. (A)Quantitative real-time RT-PCR (qRT-PCR) assay shows that 5PT12 is expressed in different tissues,preferentially in the leaves and flowers. (B)Temporal and spatial expression analysis of 5PT12 by promoter-GUS fusion studies reveals that 5PT12 istranscribed in seedlings (a), leaves (b), pollen in anthers at different floral stages (c-f, images d-f represent floral stages 12-14), mature pollen (g),and germinating pollen and pollen tubes (h). Scale bars: 1 mm in a-c; 200m in d-f; 50m in g; 20m in h. (C)qRT-PCR assay shows that 5PT12and 5PT13 are expressed in flowers at different floral stages. (D)Promoter-GUS fusion studies reveal the expression of Arabidopsis 5PT13 in flowers(a), pollen at different floral stages (b-e, representing floral stages 12, 13, 14, and later than 14, respectively), germinating pollen (f) and pollentubes (g). Scale bars: 1 mm in a; 200m in b-e; 20m in g,h. (E)Schematic structure of 5PT12 (At2g43900, boxes represent exons). The position ofthe T-DNA insertion and primers used for genotyping are indicated (top panel). Analysis of heterozygous (5PT12/5pt12), homozygous (5pt12) andwild-type plants is shown in the bottom panel. (F)qRT-PCR assay revealed that expression of 5PT12 is deficient in the 5pt12 mutant, which isrecovered by complementary expression of 5PT12 driven by its native promoter. RNA extracted from flowers was used for analysis, and wild-typeexpression of 5PT12 was set as 1.0. (G)Scanning electron microscope (SEM) analyses of flowers at floral stage 13 of wild-type and 5pt12 plantsshow that pollen grains in 5pt12 anthers have tube-shaped structures (arrows, a representative image is shown in the right panel). (H)Tube-shapedstructures of 5pt12 pollen grains at floral stage 12. Pollen grains were washed off the anthers of flowers at floral stage 12 of both wild-type and5pt12 plants. Scale bar: 20m. (I)The tube-shaped structures in 5p12 anthers are precociously germinated pollen tubes. Anthers of flowers at floralstage 13 are stained with decolored Aniline Blue solution. Complementary expression of 5PT12 rescues normal pollen germination. Fluorescence isdetected under UV light. Scale bars: 50m. (J)Measurement of precocious germination incidence confirms the precocious pollen germination ofhomozygous 5pt12 and heterozygous 5PT12/5pt12 mutants, whereas complementary expression of 5PT12 rescues normal pollen germination.Experiments are repeated three times, and more than 150 pollen grains are observed and measured. Data are presented as average±s.d. D
EVELO
PMENT

2226
seed set rate of 5pt12 was not affected, despite the precociouspollen germination (at the rate of 40-70%, Fig. 2C). All theseresults confirm the normal development of 5pt12 pollen grains.
Unaltered dehydration of 5PT12-deficient pollengrainsPrevious studies have demonstrated the significance ofdehydration, which is probably mediated by ABA signaling, inpollen dormancy and germination (Hsu et al., 2010). Ourobservations showed that precociously germinated pollen grainsin 5pt12 anthers keep their dehydrated shape (Fig. 3A). The axislengths of pollen grains from dehiscent anthers were furthermeasured and there was no statistical difference in the ratio ofminor axis:major axis between wild-type and 5pt12 pollen,suggesting that the mature pollen grains of 5pt12, including theprecocious germinated ones, undergo normal dehydrationprocess.
Deficiencies in ABA synthesis or signaling lead to seedprecocious germination or vivipary, and pollen dehydration isprobably mediated by ABA signaling. As disruption oroverexpression of several characterized Arabidopsis 5PTs hasbeen reported to alter ABA responses, we therefore testedwhether 5pt12 has altered ABA content or ABA signaling. Wild-
RESEARCH ARTICLE Development 139 (12)
Fig. 2. The 5pt12 mutant exhibits normal early pollendevelopment and fertility. (A)Mature pollen grains of the 5pt12mutant have regular tri-cellular structures similar to those of wild-typeplants, as revealed by DAPI staining (left panel). In addition, viabilityanalysis by FDA indicates normal viability of 5pt12 pollen grains.Mature pollen grains at floral stage 14 are used for analyses. Scale bars:10m (left); 100m (right). (B)Pollen grains that do not germinate inanthers germinate and grow normally in vitro. Scale bars: 20m.(C)Fertility is not altered under 5PT12 deficiency. Well-developed seedsin mature siliques of both wild-type and 5pt12 plants are calculated,and the data are presented as the average±s.d. (more than 20independent plants of each line are analyzed).
Fig. 3. Normal dehydration of precocious germinated pollen grainsand hypersensitive ABA responses of 5p12 mutant. (A)Precociouslygerminated pollen grains from dehiscent anthers of 5pt12 mutant havesimilar shapes to those of wild-type plants (left panel). Statistical analysisof the ratio of minor axis to major axis of pollen grains from dehiscentanthers further confirms the similar dehydration of 5pt12 mutants andwild-type pollen grains (right panel). More than 50 pollen grains of wildtype or 5pt12 mutants are analyzed, and the assays are repeated threetimes. Arrowheads indicate the pollen tubes. Scale bars: 20m. (B)LC-MS analysis of anthers of wild type and 5pt12 mutant revealed nochanges of the ABA level in 5pt12 mutants at floral stage 12 or earlierstages. (C)qRT-PCR analysis of the ABA signaling-related genes KIN2,COR15, RD22 and COR47 indicate the enhanced ABA responses of5pt12 seedlings. Eight-day-old seedlings are used for ABA treatments (0,0.5, 1 or 2 hours). (D)Seed germination of the 5pt12 mutant was moresensitive to ABA treatment, further confirming the enhanced ABAresponse under 5PT12 deficiency. Seed germination percentage wascalculated after 7 days ABA treatment (0, 0.1, 0.3 or 0.5M) of wild-typeand 5pt12 seeds. Data are presented as average±s.d. (E)Pollengermination percentages under different relative humidity conditions.Wild-type pollen grains at floral stage 12 are used for in vitro germinationassay on germination media without additional Ca2+. Data are presentedas average±s.d. D
EVELO
PMENT

type or 5pt12 anthers at different floral stages were collected forABA measurements. There was no significant difference in ABAcontent between wild type and 5pt12 either at floral stage 12 orat earlier stages (Fig. 3B). Further analysis of the ABA-inducedexpression of ABA-responsive genes (KIN2, COR15, RD22 andCOR47) in seedlings showed a much enhanced induction ofthese genes under 5PT12 deficiency (Fig. 3C), indicatingstimulated ABA signaling in 5pt12 mutant. Consistently, the5pt12 mutant was more sensitive to exogenous ABA in seedgermination assays (Fig. 3D). The expression patterns of ABA-responsive genes in floral tissues were also tested. However,ABA treatment could not induce uniform and consistentexpression of all the tested ABA-responsive genes, althoughsome expression was induced at specific floral stages(supplementary material Fig. S3). The reason might be that ABAcannot effectively penetrate into the pollen grains owing to thickanther walls (no similar ABA treatment has been reportedpreviously). Taken together, these results indicate unaffectedABA synthesis and stimulated ABA signaling in the 5pt12mutant; thus, precocious pollen germination seen in 5pt12mutants is not due to altered ABA responses.
We also examined the germination rate of pollen (at early floralstage 12) in response to different humidity conditions. Thegermination rate of wild-type stage 12 pollen grains increased by2.8-fold (from 18% to 52%) as the environmental humidity rose,whereas that of 5pt12 early stage 12 pollen grains (at this time thepollen grains are not germinated) showed a moderate increase of1.6-fold (from 46% to 76%, Fig. 3E). Compared with wild-typepollen grains, 5pt12 pollen grains showed a higher germination rateunder low humidity conditions and a significantly highergermination percentage at high environmental humidity (76%versus 45% of wild type). This ruled out the possibility that thealtered environmental humidity caused the precocious pollengermination in 5pt12 mutant.
Taken together, these results strongly suggested that theprecocious pollen germination in the 5pt12 mutant is not caused byalterations in dehydration/rehydration or by the ABA response, andindicated the existence of another key regulator in pollen dormancyand germination, which is possibly triggered in the 5pt12 mutant.Ca2+ is the potential candidate, based on the fact that Ca2+ is crucialfor pollen development and germination, and that Ins(1,4,5)P3
stimulates the cytosolic Ca2+.
5PT13 deficiency synergistically enhancesprecocious pollen germination, and Ins(1,4,5)P3basal level/[Ca2+]cyt is increased in 5pt12 and 5pt125pt13 mutantsBased on microarray data (https://www.genevestigator.ethz.ch/),5PT13, a close paralog of 5PT12, is weakly expressed in pollengrains. qPT-PCR analysis confirmed the expression of 5PT13 atfloral stages 12 to 14 (Fig. 1C), with a similar pattern to that of5PT12, and a detailed expression analysis by promoter-reportergene fusion studies shows that the 5PT13 gene is largelyexpressed in pollen grains at floral stages 13 and 14, as well asin mature pollen and pollen tubes (Fig. 1D). Although theexpression level of 5PT13 is lower than 5PT12 at floral stage 12to 14, especially at floral stage 12, the 5PT13 knockout mutant(5pt13) also exhibits precocious pollen germination in dehiscentanthers (Fig. 4A,B) at a low incidence. Double mutant 5pt125pt13 has a much higher incidence of precocious pollengermination (~60-70%) than 5pt12 or 5pt13 single mutants (Fig.4C). In addition, abnormal pollen tubes were occasionally
observed in precociously germinated pollen grains, includingmore than one pollen tube formed in one pollen grain or aberrantpollen tube structures (Fig. 4D).
Previous studies have shown that in vitro, both 5PT12 and5PT13 exhibit phosphatase activity towards only Ins(1,4,5)P3,rather than towards inositol lipids (Zhong and Ye, 2004). Thus,Ins(1,4,5)P3 is suggested to be involved in the regulation of pollendormancy and germination. Ins(1,4,5)P3 receptor binding proteinwas employed specifically to determine the intrinsic Ins(1,4,5)P3
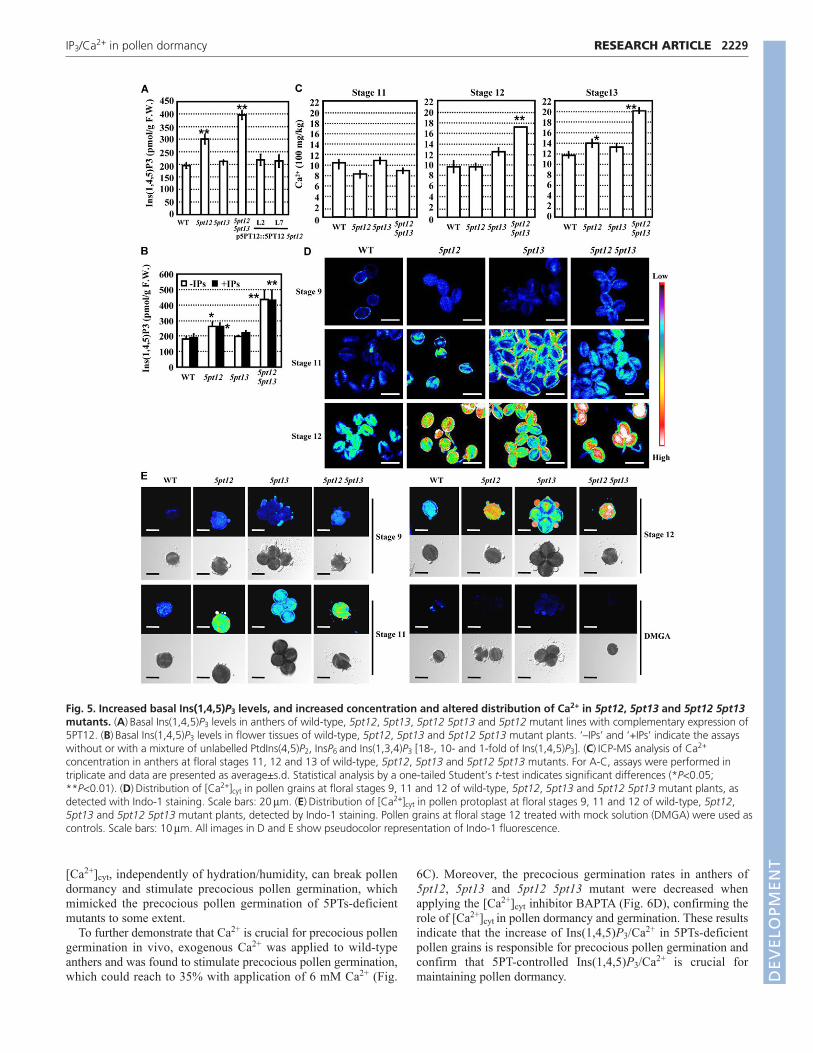
content by using the D-myo-inositol-1,4,5-trisphosphate [3H] assaykit. The basal level of Ins(1,4,5)P3 was significantly increased in5pt12 anthers and was even more increased in 5pt12 5pt13 doublemutants in comparison with wild type, while the complementaryexpression of 5PT12 restored Ins(1,4,5)P3 levels (Fig. 5A). BasalIns(1,4,5)P3 levels corresponded to the precocious germinationincidences in each genotype.
It has been reported that the basal level of InsP2 or InsP6 ishigher than Ins(1,4,5)P3 in Arabidopsis (Perera et al., 2008). Toexclude the potential influence caused by non-specific binding,unlabelled PtdIns(4,5)P2, InsP6 and Ins(1,3,4)P3 [Ins(1,4,5)P3
stereoisomer] were added into the samples and the results showedthat the addition of these inositol phosphates and phospholipids didnot affect the measurement (Fig. 5B), consistent with the specificitydescribed by the manufacturer (the cross-reactivity for otherinositol phosphates is lower than 1%,http://www.gehealthcare.com/lifesciences). These results, thus,indicate a close correlation between Ins(1,4,5)P3 content andprecocious pollen germination, and indicate that Ins(1,4,5)P3 isinvolved in controlling pollen grain dormancy in anthers.
As Ca2+ is crucial for pollen germination on stigma or in vitro,total Ca2+ contents in anthers of wild type, 5pt12, 5pt13 and 5pt125pt13 double mutant were measured using ICP-MS (inductivelycoupled plasma mass spectrometric analysis) (Chen et al., 2003).The results showed that the Ca2+content in 5pt12 mutant anthers(stage 13) was higher than that in wild-type anthers, and the 5pt125pt13 double mutant had an even higher Ca2+ content in stage 12and 13 anthers (Fig. 5C). No significant difference in Ca2+ contentwas detected between the mutants and wild-type stage 11 anthers,where 5PT12 and 5PT13 have very low expression.
Although the Ins(1,4,5)P3 receptor has not been identified inhigher plants, it is assumed that Ins(1,4,5)P3 can stimulate the Ca2+
release from the internal Ca2+ store. The concentration anddistribution of [Ca2+]cyt in 5pt12, 5pt13 or 5pt12 5pt13 pollen werefurther detected by Indo-1 (an indicator of [Ca2+]cyt) staining.[Ca2+]cyt was gradually increased along with the pollendevelopment, and reached its highest level in mature tricellularpollen grains at floral stage 12 (Fig. 5D). At floral stage 9, nodifferences in Indo-1-indicated [Ca2+]cyt were detected betweenwild type and 5pt mutants, whereas, at stage 12, significantlyhigher [Ca2+]cyt was detected in 5pt12 pollen grains, consistent withthe expression pattern of 5PT12. Furthermore, the distribution of[Ca2+]cyt showed a diffuse pattern rather than accumulation ingermination furrows (high concentration of [Ca2+]cyt isaccumulated in germination furrows of wild-type pollen grains forsubsequent germination of pollen tubes, Fig. 5D). The alterationsof [Ca2+]cyt were much more significant in the 5pt12 5pt13 doublemutant (Fig. 5D).
To rule out the possible effect of pollen coat on Indo-1 staining,pollen protoplast was used for detecting Indo-1-indicated [Ca2+]cyt
and similar results were obtained (Fig. 5E). Observation ofplasmolyzed root hair cells stained with Indo-1 further indicatedthat Indo-1 fluorescence came from cytoplasm rather than the cell
2227RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
DEVELO
PMENT

2228
wall (supplementary material Fig. S4), confirming that Ca2+
associated with the pollen coat or cell wall is not labeled by Indo-1.
These results indicate that the Ins(1,4,5)P3-mediated [Ca2+]cyt
increase is responsible for the precocious pollen germination in5pt12 and 5pt12 5pt13 mutants, and suggest that 5PTs andIns(1,4,5)P3/Ca2+ are crucial for maintaining pollen dormancy.
[Ca2+]cyt breaks pollen dormancy and stimulatesprecocious pollen germinationAlthough Ca2+ is known to be required for pollen germination andpollen tube growth, the effect of Ca2+ on pollen dormancy has notbeen reported before. We therefore tested the influence ofexogenous Ca2+ on pollen grains at floral late stage 11. [Ca2+]cyt in5pt12, 5pt13 and 5pt12 5pt13 pollen at floral late stage 11 was onlyslightly higher than that in wild-type pollen (Fig. 5D), and all thesemutants did not show the precocious pollen germination withinanthers at this stage. However, in vitro pollen germination assaysshowed that under 100% humidity, wild-type pollen at floral latestage 11 could germinate in a low percentage (~26%, Fig. 6A),indicating the low germination potential of pollen at late floral
stage 11. As exogenous Ca2+ concentration rose, the precociousgermination rate of wild-type pollen increased, especially atconcentrations of 1 or 1.5 mM (Fig. 6A), indicating that exogenousCa2+ can break the dormancy and promote precocious germinationof wild-type pollen at late floral stage 11 in a dose-dependentmanner. As expected, 5pt12 and 5pt12 5pt13 mutants showedhigher precocious pollen germination rates than wild type whensupplemented with 1 or 1.5 mM Ca2+ (Fig. 6A), consistent with theslightly increased [Ca2+]cyt in pollen of these mutants at floral latestage 11. There was no difference in precocious germination rateunder high concentrations of exogenous Ca2+ (2 mM,supplementary material Fig. S5).
At floral stage 12, pollen grains are in enclosed anthers. Aspollen dehydration is supposed to occur at late stage 12, the insideof the anther should have very low environmental humidity.Therefore, we further tested the effect of Ca2+ on pollengermination in very low relative humidity (0-10%). In the absenceof additional Ca2+, wild-type pollen grains (floral stage 12)germinated at a very low rate, whereas supplementation withexternal Ca2+ significantly stimulated the germination (4 mM Ca2+
produces a 44% germination rate, Fig. 6B). This indicated that
RESEARCH ARTICLE Development 139 (12)
Fig. 4. Both 5pt13 and 5pt12 5pt13had precocious germination ofpollen grains within anthers.(A)Scanning electron microscopy (SEM)analyses of flowers at floral stage 13 ofwild-type, 5pt12, 5pt13 and 5pt125pt13 mutant plants showed that pollengrains in 5pt13 and 5pt12 5pt13mutant anthers have tube-shapedstructures (outlined region is enlarged inthe bottom panel). Scale bars: Top row,left to right, 20, 50, 20, 20 m; bottomrow, 10 m. (B)The tube-shapedstructures in 5pt12, 5pt13 and 5pt125pt13 anthers are precociouslygerminated pollen tubes (outlinedregions are enlarged in the bottompanel). Anthers of flowers at floral stage13 are stained with decolored AnilineBlue solution and fluorescence isdetected under UV light. Scale bars:100m (top panel); 30m (bottompanel). (C)Analysis of precociousgermination incidence confirms theprecocious pollen germination in 5pt13mutants and the 5pt12 5pt13 doublemutant has a much higher incidence ofprecocious germination in comparisonwith 5pt12 or 5pt13 single mutant.More than 300 pollen grains areobserved and calculated. Data arepresented as average±s.d. (D)Theprecociously germinated pollen grains in5pt12 5pt13 mutant anthersoccasionally have distorted pollen tubesor more than one pollen tube on asingle pollen grain (right panel).Arrowhead indicates the abnormalpollen tube. Scale bars: 20m.
DEVELO
PMENT
[Ca2+]cyt, independently of hydration/humidity, can break pollendormancy and stimulate precocious pollen germination, whichmimicked the precocious pollen germination of 5PTs-deficientmutants to some extent.
To further demonstrate that Ca2+ is crucial for precocious pollengermination in vivo, exogenous Ca2+ was applied to wild-typeanthers and was found to stimulate precocious pollen germination,which could reach to 35% with application of 6 mM Ca2+ (Fig.
6C). Moreover, the precocious germination rates in anthers of5pt12, 5pt13 and 5pt12 5pt13 mutant were decreased whenapplying the [Ca2+]cyt inhibitor BAPTA (Fig. 6D), confirming therole of [Ca2+]cyt in pollen dormancy and germination. These resultsindicate that the increase of Ins(1,4,5)P3/Ca2+ in 5PTs-deficientpollen grains is responsible for precocious pollen germination andconfirm that 5PT-controlled Ins(1,4,5)P3/Ca2+ is crucial formaintaining pollen dormancy.
2229RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
Fig. 5. Increased basal Ins(1,4,5)P3 levels, and increased concentration and altered distribution of Ca2+ in 5pt12, 5pt13 and 5pt12 5pt13mutants. (A)Basal Ins(1,4,5)P3 levels in anthers of wild-type, 5pt12, 5pt13, 5pt12 5pt13 and 5pt12 mutant lines with complementary expression of5PT12. (B)Basal Ins(1,4,5)P3 levels in flower tissues of wild-type, 5pt12, 5pt13 and 5pt12 5pt13 mutant plants. ‘–IPs’ and ‘+IPs’ indicate the assayswithout or with a mixture of unlabelled PtdIns(4,5)P2, InsP6 and Ins(1,3,4)P3 [18-, 10- and 1-fold of Ins(1,4,5)P3]. (C)ICP-MS analysis of Ca2+
concentration in anthers at floral stages 11, 12 and 13 of wild-type, 5pt12, 5pt13 and 5pt12 5pt13 mutants. For A-C, assays were performed intriplicate and data are presented as average±s.d. Statistical analysis by a one-tailed Student’s t-test indicates significant differences (*P<0.05;**P<0.01). (D)Distribution of [Ca2+]cyt in pollen grains at floral stages 9, 11 and 12 of wild-type, 5pt12, 5pt13 and 5pt12 5pt13 mutant plants, asdetected with Indo-1 staining. Scale bars: 20m. (E)Distribution of [Ca2+]cyt in pollen protoplast at floral stages 9, 11 and 12 of wild-type, 5pt12,5pt13 and 5pt12 5pt13 mutant plants, detected by Indo-1 staining. Pollen grains at floral stage 12 treated with mock solution (DMGA) were used ascontrols. Scale bars: 10m. All images in D and E show pseudocolor representation of Indo-1 fluorescence.
DEVELO
PMENT

2230 RESEARCH ARTICLE Development 139 (12)
Fig. 6. [Ca2+]cyt breaks pollen dormancy and stimulates precocious pollen germination. (A)Under the 100% humidity condition, exogenousCa2+ (0.5-1.5 mM) promotes in vitro precocious germination of pollen at floral late stage 11, especially those of 5pt12 and 5pt12 5pt13 mutants.Data are presented as average±s.d. (n150-300). (B)Under very low relative humidity, exogenous Ca2+ significantly promotes in vitro precociousgermination of pollen at floral stage 12. Data are presented as average±s.d. (n>200). Representative images highlight germinated pollen grainswhen supplemented with exogenous Ca2+ (right panel). Scale bars: 50m. (C)Observation (left panels) and statistical analysis (right panel) showedthat exogenous Ca2+ significantly promotes in vivo precocious germination of pollen at floral stage 12. More than 300 pollen grains are observedand calculated. Data are presented as average±s.d. Scale bars: 100m. (D)Observation (left panels) and statistical analysis (right panel) confirm thatBAPTA treatment blocks the in vivo precocious germination of pollen at floral stage 12 of 5pt12, 5pt13 and 5pt12 5pt13 mutant plants (left panel).More than 300 pollen grains are observed and calculated. Data are presented as average±s.d. Scale bar: 100m. (E)Model of the role for 5PT-controlled Ins(1,4,5)P3/[Ca2+]cyt in maintaining pollen dormancy. In wild-type plants, 5PT12, 5PT13 and 5PT14 are synergistically highly expressed intri-cellular pollen at floral stage 12, to restrain the level of Ins(1,4,5)P3/[Ca2+]cyt in tri-cellular pollen and maintain the pollen in dormant state.[Ca2+]cyt is mainly controlled by intrinsic Ins(1,4,5)P3 levels to prevent pollen grains from germinating. This prevents the precocious pollengermination within anthers. The dehydration process at late stage 12 further maintains this state. Once the dehydrated pollen grains fall on pistilsand rehydrate, Ca2+ levels will increase and [Ca2+]cyt aggregates in the germination hole, resulting in the broken dormancy and pollen germination.Deficiency of 5PT12 or 5PT13 results in increased intrinsic Ins(1,4,5)P3/[Ca2+]cyt in tri-cellular pollen at floral stage 12 or earlier stages, preventing themaintenance of dormancy and resulting in the precocious germination of pollen grains within anthers. D
EVELO
PMENT

DISCUSSION5PT-controlled Ins(1,4,5)P3/Ca2+ is crucial formaintaining pollen dormancyAfter maturation, pollen grains undergo dehydration and are thenreleased from anthers in a partially dehydrated and dormant state.Pollen germination on the stigma starts with the rehydration that issubject to stringently spatiotemporal control (Heslop-Harrison,1979). Inappropriate dehydration/rehydration results in prematureor earlier germination within the anther, or germination on thewrong surface (Lolle and Cheung, 1993; Lolle et al., 1998).Therefore, pollen dehydration is considered to be the majordeterminant that controls pollen dormancy. Here, our study firstreveal that, in addition to dehydration, 5PTs and Ins(1,4,5)P3/Ca2+
play crucial roles in pollen dormancy and uncover a novelmechanism for controlling it.
Deficiency of 5PT12, 5PT13 leads to precocious pollengermination within anthers, suggesting the essential roles of these5PTs in pollen dormancy. However, the precociously germinatedpollen grains in 5pt12 anthers could dehydrate normally andpollens of the 5pt12 mutant is dehydrated before release from theanthers (Fig. 3A), suggesting that humidity or altered sensitivity toenvironmental humidity are not responsible for precociousgermination, As the 5pt12 mutant exhibits hypersensitive ABAresponses, further excluding the influence of dehydration/hydrationor ABA signaling in altered pollen dormancy and germination ofthe 5pt12 mutant.
As speculated, the basal levels of Ins(1,4,5)P3, the commonsubstrate of 5PT12 and 5PT13, are elevated significantly in the5pt12 or 5pt12 5pt13 mutant, and an increased [Ca2+]cyt
concentration is detected in pollen grains of the 5pt12 mutant or5pt12 5pt13 double mutant (more significantly) at floral stage 12.Previous studies have shown that a 1.6-fold increase in Ins(1,4,5)P3
did affect the seedling growth (Ercetin et al., 2008) and here anapproximate doubling of Ins(1,4,5)P3 in the 5pt12 5pt13 doublemutant is sufficient to lead to precocious pollen germination. Inaddition, preventing the increase in basal Ins(1,4,5)P3 levels in the5pt12 mutant by complementary expression of 5PT12 resulted inthe normal pollen germination rate, further confirming the role ofincreased Ins(1,4,5)P3 levels in precocious pollen germination. Nosignificantly increase in Ins(1,4,5)P3 or total Ca2+ content wasdetected in 5pt13 mutant anthers, which may be because wholeanthers were used as the material for Ins(1,4,5)P3 and total Ca2+
measurement (because pollen grains are too small to obtain enoughmaterial for measurement), and the minor alterations in 5pt13mutant pollen grains might be covered up. However, the increasedCa2+ in the 5pt13 mutant pollen grains can be clearly observed bythe Indo-1 staining assay, which is consistent with precociouspollen germination in the 5pt13 mutant. These findings indicatethat precocious pollen germination, a higher basal level ofIns(1,4,5)P3 and increased [Ca2+]cyt in pollen grains are closelycorrelated.
A more diffuse [Ca2+]cyt distribution pattern was observed in thepollen of the 5pt12 5pt13 double mutant (distinguished from theaggregated pattern in wild-type pollen grains prior to germinationon stigma) (Fig. 5D,E), which may explain the emergence of morethan one pollen tube. The concentration and distribution of [Ca2+]cyt
is supposed to be stringently controlled during pollen dormancyand 5PTs/Ins(1,4,5)P3 may be the regulators.
In vitro pollen germination assays showed that exogenous Ca2+
could break dormancy and stimulate the early germination of wild-type pollen grains at floral late stage 11, which confirms the crucialroles of Ca2+ in pollen dormancy. In addition, exogenous Ca2+ can
break dormancy and trigger early germination of wild-type pollengrains at floral stage 12 under very low humidity (Fig. 6B).BAPTA treatment resulted in the decreased pollen germinationrates in anther of 5pt12, 5pt13 and 5pt12 5pt13 mutants,confirming that the precocious pollen germination in 5PT mutantsis caused by increased [Ca2+]cyt (Fig. 6D). In addition, 5PT14, aclose paralog of 5PT12 and 5PT13, is also expressed at floral stage12 to 14, and the expression level of 5PT14 is higher than 5PT12and 5PT13 at these stages (Fig. 1C, supplementary material Fig.S6A). Recently, we identified a T-DNA insertion mutant of 5PT14,which also exhibited precocious pollen germination in dehiscentanthers (~15-30%) (supplementary material Fig. S6B,C,D).Although all 5PT12, 5PT13 and 5PT14 are expressed in pollengrain, each single mutant exhibits the phenotype of precociouspollen germination and the 5pt12 5pt13 double mutant with muchhigher percentage of the of precocious pollen germination,suggesting the dose effect of 5PTs in regulating the pollengermination. A further study with mutation of all 5PT12, 5PT13and 5PT14 is on the way.
Take together, these findings indicate that 5PT-controlledIns(1,4,5)P3/Ca2+ has a crucial role in maintaining pollen dormancyand the increase of 5PT-controlled Ins(1,4,5)P3/Ca2+ is sufficient tobreak pollen dormancy and trigger early germination, independentof hydration/humidity. This study sheds light on the mechanismthat controls pollen dormancy and germination (Fig. 6E). In wild-type plants, 5PT12, 5PT13 and 5PT14 are highly expressed in tri-cellular pollen at floral stage 12 in order to restrain the level ofIns(1,4,5)P3/[Ca2+]cyt in tri-cellular pollen. The pollen at stage 12is then maintained in a dormant state, and prevented fromprecocious germinating. At late stage 12, tri-cellular pollenundergoes dehydration to further maintain the dormant state. Oncethe dehydrated pollen grains fall on pistils and rehydrate, Ca2+ willflow into pollen grains and aggregates in the germination pore,breaking dormancy and stimulating pollen early germination.Deficiency of 5PT genes will result in the increased intrinsicIns(1,4,5)P3/[Ca2+]cyt in tri-cellular pollen at floral stage 12, whichsufficiently breaks the pollen dormancy and results in theprecocious germination of pollen grains in anthers.
Ins(1,4,5)P3 can be converted into InsP6 (inositolhexakisphosphate) and InsP6 was reported to mobilize intracellularCa2+ stores (Lemtiri-Chlieh et al., 2003) in guard cells. Althoughthere is no evidence that InsP6 is involved in breaking pollendormancy, it is possible that the InsP6 content in 5PT mutants isalso increased and, to some extent, contributes to the alteration of[Ca2+]cyt in 5PT mutants.
Biological significance and engineering prospectof precocious pollen germinationHere, we report a striking phenotype in Arabidopsis: precociouspollen germination within anthers at an incidence of 40-70%,caused by 5PT12 deficiency. Pollen grains in wild-typeArabidopsis do not germinate until falling onto stigma. In fact,pollen grains in almost all the flowering plants will not germinateprior to pollination. The only exceptions are some cleistogamousspecies, in which pollen germinates inside the anther, and thepollen tubes extend via the anther stomium to reach to the stigmafor fertilization (Lord, 1981; Trent, 1942), e.g. Cardaminechenopodifolia (Lord, 1981). It is noteworthy that C.chenopodifolia is a Cruciferae plant, similar to Arabidopsis. Inspecies in which the anther sac fails to open, germinated pollentubes go through the anther walls to the stigma (Hanson, 1943;Lord, 1981). However, little is known about the genetic
2231RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
DEVELO
PMENT

2232
mechanism. Although a similar phenotype has been reported in theraring-to-go (rtg) mutant, the pollen grains of which do notdesiccate completely and thus germinate precociously (Johnson andMcCormick, 2001), the responsible gene and genetic mechanismare still unknown. In our study, 5pt12 and 5pt12 5pt13 mutantsshowed precocious pollen germination within anthers, similar tothese cleistogamous plants, providing convincing evidence of thecrucial roles of 5PTs and Ins(1,4,5)P3/[Ca2+]cyt in pollen dormancy,and revealing a novel mechanism for keeping pollen dormancy.These results provide insights into the mechanisms of pollengermination in such cleistogamous species. It is speculated thatIns(1,4,5)P3/[Ca2+]cyt might also function in breaking dormancyand stimulating pollen germination in these cleistogamous plants,and 5PTs or other enzymes controlling Ins(1,4,5)P3 metabolism inthe pollen of these species are expressed at low levels or aredeficient, which promotes pollen germination in anthers.
Increased Ins(1,4,5)P3/Ca2+ is sufficient to break pollendormancy and trigger early germination, suggesting an engineeringprospect of 5PTs in altering dormancy. The mechanism controllingseed dormancy is very likely to be similar to pollen dormancy andthereby 5PTs may be applied to alter seed dormancy. Recently, twolines with altered expression of callose synthases were reported toshow precocious pollen germination, but at a very low incidence(4-8%) (Xie et al., 2010). However, in contrast to the 5pt12 mutant,all the reported lines, including rtg mutant, have abnormal pollendevelopment as early as the bi-cellular stage. Thus, 5PT12 may bea better candidate for genetic engineering in crops for improvingfertility and sterility, especially under environmental stress. Forcereal crops, seed dormancy before harvest is a desired trait. Insome cultivars, mature grains germinate on maternal plantsfollowing high temperature or rains, causing large economic losses.This phenomenon is characterized as pre-harvest sprouting orvivipary. It is worth testing whether the vivipary phenotype couldbe somehow rescued by overexpression of 5PT12. On the contrary,some crops, such as barley, exhibit excessive dormancy at harvestthat prevent grains from rapid and uniform germination, which isrequired in the malting process. Using a transgenic strategy (usingantisense fragment of 5PT12 under the control of seed-specificpromoter) to specifically silence 5PT orthologs in mature seeds ofthese crops might inhibit seed dormancy. Thus, our researchprovides possibilities of genetic engineering for improving croptraits and agricultural production.
FundingThe study was supported by the National Science Foundation of China[31130060, 30740006] and Shanghai Institutes for Biological Sciences[SIBS2008004].
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material available online athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.081224/-/DC1
ReferencesAstle, M. V., Seaton, G., Davies, E. M., Fedele, C. G., Rahman, P., Arsala, L.
and Mitchell, C. A. (2006). Regulation of phosphoinositide signaling by theinositol polyphosphate 5-phosphatases. IUBMB Life 58, 451-456.
Berdy, S. E., Kudla, J., Gruissem, W. and Gillaspy, G. E. (2001). Molecularcharacterization of At5PTase1, an inositol phosphatase capable of terminatinginositol trisphosphate signaling. Plant Physiol. 126, 801-810.
Bewley, J. D. (1997). Seed germination and dormancy. Plant Cell 9, 1055-1066.Burnette, R. N., Gunesekera, B. M. and Gillaspy, G. E. (2003). An Arabidopsis
inositol 5-phosphatase gain-of-function alters abscisic acid signaling. PlantPhysiol. 132, 1011-1019.
Carland, F. M. and Nelson, T. (2004). COTYLEDON VASCULAR PATTERN2-mediated inositol (1,4,5) trisphosphate signal transduction is essential for closedvenation patterns of Arabidopsis foliar organs. Plant Cell 16, 1263-1275.
Chen, X., Lin, W. H., Wang, Y., Luan, S. and Xue, H. W. (2008). An inositolpolyphosphate 5-phosphatase functions in PHOTOTROPIN1 signaling inArabidopsis by altering cytosolic Ca2+. Plant Cell 20, 353-366.
Chen, Z., Griffin, I. J., Kriseman, Y. L., Liang, L. K. and Abrams, S. A. (2003).Inductively coupled plasma mass spectrometric analysis of calcium isotopes inhuman serum: a low-sample-volume acid-equilibration method. Clin. Chem. 49,2050-2055.
Ercetin, M. E. and Gillaspy, G. E. (2004). Molecular characterization of anArabidopsis gene encoding a phospholipid-specific inositol polyphosphate 5-phosphatase. Plant. Physiol. 135, 938-946.
Ercetin, M. E., Ananieva, E. A., Safaee, N. M., Torabinejad, J., Robinson, J. Y.and Gillaspy, G. E. (2008). A phosphatidylinositol phosphate-specific myo-inositol polyphosphate 5-phosphatase required for seedling growth. Plant Mol.Biol. 67, 375-388.
Escobar-Restrepo, J. M., Huck, N., Kessler, S., Gagliardini, V., Gheyselinck, J.,Yang, W. C. and Grossniklaus, U. (2007). The FERONIA receptor-like kinasemediates male-female interactions during pollen tube reception. Science 317,656-660.
Fang, J., Chai, C., Qian, Q., Li, C., Tang, J., Sun, L., Huang, Z., Guo, X., Sun,C., Liu, M. et al. (2008). Mutations of genes in synthesis of the carotenoidprecursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice.Plant J. 54, 177-189.
Finkelstein, R., Reeves, W., Ariizumi, T. and Steber, C. (2008). Molecularaspects of seed dormancy. Annu. Rev. Plant Biol. 59, 387-415.
Fleige, S., Walf, V., Huch, S., Prgomet, C., Sehm, J. and Pfaffl, M. W. (2006).Comparison of relative mRNA quantification models and the impact of RNAintegrity in quantitative real-time RT-PCR. Biotechnol. Lett. 28, 1601-1613.
Franklin-Tong, V. E. (1999). Signaling and the modulation of pollen tube growth.Plant Cell 11, 727-738.
Gunesekera, B., Torabinejad, J., Robinson, J. and Gillaspy, G. E. (2007).Inositol polyphosphate 5-phosphatases 1 and 2 are required for regulatingseedling growth. Plant Physiol. 143, 1408-1417.
Hanson, C. H. (1943). Cleistogamy and the development of the embryo sac inLespedeza stipulacea. J. Agric. Res. 67, 265-272.
Heslop-Harrison, J. (1979). An interpretation of the hydrodynamics of pollen.Am. J. Bot. 66, 737-743.
Heslop-Harrison, J. and Heslop-Harrison, Y. (1992a). Germination ofmonocolpate angiosperm pollen: effects of inhibitory factors and the Ca2+-channel blocker nifedipine. Ann. Bot. 69, 395-403.
Heslop-Harrison, Y. and Heslop-Harrison, J. (1992b). Germination ofmonocolpate angiosperm pollen: evolution of the actin cytoskeleton and wallduring hydration, activation and tube emergence. Ann. Bot. 69, 385-394.
Hilhorst, H. W. M. and Downie, B. (1996). Primary dormancy in tomato(Lycopersicon esculentum cv. Moneymaker): Studies with the sitiens mutant. J.Exp. Bot. 47, 89-97.
Hsu, S. W., Cheng, C. L., Tzen, T. C. and Wang, C. S. (2010). Rop GTPase and itstarget Cdc42/Rac-interactive-binding motif-containing protein genes respond todesiccation during pollen maturation. Plant Cell Physiol. 51, 1197-1209.
Iwano, M., Shiba, H., Miwa, T., Che, F. S., Takayama, S., Nagai, T., Miyawaki,A. and Isogai, A. (2004). Ca2+ dynamics in a pollen grain and papilla cell duringpollination of Arabidopsis. Plant Physiol. 136, 3562-3571.
Johnson, S. A. and McCormick, S. (2001). Pollen germinates precociously in theanthers of raring-to-go, an Arabidopsis gametophytic mutant. Plant Physiol.126, 685-695.
Koornneef, M. and Karssen, C. M. (1994). In Seed Dormancy and Germinationin Arabidopsis (ed. E. M. Meyerowitz and C. R. Somerville), pp. 313-334. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press.
Koornneef, M., Hanhart, C. J., Hilhorst, H. W. M. and Karssen, C. M. (1989).In vivo inhibition of seed development and reserve protein accumulation inrecombinants of abscisic acid biosynthesis and responsiveness mutants inArabidopsis thaliana (L.) Heynh. Theor. Appl. Genet. 58, 257-263.
Lemtiri-Chlieh, F., MacRobbie, E. A., Webb, A. A., Manison, N. F., Brownlee,C., Skepper, J. N., Chen, J., Prestwich, G. D. and Brearley, C. A. (2003).Inositol hexakisphosphate mobilizes an endomembrane store of calcium inguard cells. Proc. Natl. Acad. Sci. USA 100, 10091-10095.
Li, H., Lin, Y., Heath, R. M., Zhu, M. X. and Yang, Z. (1999). Control of pollentube tip growth by a Rop GTPase-dependent pathway that leads to tip-localizedcalcium influx. Plant Cell 11, 1731-1742.
Lin, W. H., Wang, Y., Mueller-Roeber, B., Brearley, C. A., Xu, Z. H. and Xue,H. W. (2005). At5PTase13 modulates cotyledon vein development throughregulating auxin homeostasis. Plant. Physiol. 139, 1677-1691.
Liu, W., Xu, Z. H., Luo, D. and Xue, H. W. (2003). Roles of OsCKI1, a rice caseinkinase I, in root development and plant hormone sensitivity. Plant J. 36, 189-202.
Lolle, S. J. and Cheung, A. Y. (1993). Promiscuous germination and growth ofwild type pollen from Arabidopsis and related species on the shoot of theArabidopsis mutant, fiddlehead. Dev. Biol. 155, 250-258.
RESEARCH ARTICLE Development 139 (12)
DEVELO
PMENT

Lolle, S. J., Hsu, W. and Pruitt, R. E. (1998). Genetic analysis of organ fusion inArabidopsis thaliana. Genetics 149, 607-619.
Lord, E. M. (1981). Cleistogamy: A tool for the study of floral morphogenesis,function and evolution. Bot. Rev. 47, 421-450.
McCarty, D. R. (1995). Genetic control and integration of maturation andgermination pathways in seed development. Annu. Rev. Plant Physiol. Plant Mol.Biol. 46, 71-93.
McCormick, S. (2004). Control of male gametophyte development. Plant Cell 16,142-153.
Neill, S. J., Horgan, R. and Parry, A. D. (1986). The carotenoid and abscisic acidcontent of viviparous kernels and seedlings of Zea mays L. Planta 169, 87-96.
Perera, I. Y., Hung, C. Y., Moore, C. D., Stevenson-Paulik, J. and Boss, W. F.(2008). Transgenic Arabidopsis plants expressing the type I inositol 5-phosphatase exhibit increased drought tolerance and altered abscisic acidsignaling. Plant Cell 20, 2876-2893.
Sanchez, J. P. and Chua, N. H. (2001). Arabidopsis PLC1 is required for secondaryresponses to abscisic acid signals. Plant Cell 13, 1143-1154.
Sanders, P. M., Bui, A. Q., Weterings, K., McIntire, K. N., Hsu, Y. C., Lee, P. Y.,Truong, M. T., Beals, T. P., Goldberg, R. B. (1999). Anther developmentaldefects in Arabidopsis thaliana male-sterile mutants. Sex. Plant Reprod. 11, 297-322.
Smyth, D. R., Bowman, J. L. and Meyerowitz, E. M. (1990). Early flowerdevelopment in Arabidopsis. Plant Cell 2, 755-767.
Stevenson, J. M., Perera, I. Y., Heilmann, I., Persson, S. and Boss, W. F.(2000). Inositol signaling and plant growth. Trends Plant Sci. 5, 252-258.
Taylor, L. P. and Hepler, P. K. (1997). Pollen germination and tube growth. Ann.Rev. Plant Phys. Plant Mol. Biol. 48, 461-491.
Tirlapur, U. K., Scali, M., Moscatelli, A., Delcasino, C., Cai, G., Tiezzi, A. andCresti, M. (1994). Confocal image analysis of spatial variations in
immunocytochemically identified calmodulin during pollen hydration,germination and pollen tube tip growth in Nicotiana tabacum L. Zygote 2, 63-68.
Trent, J. A. (1942). Life history of Specglaria perfoliata in reference to cleistogamy.Trans. Kansas Acad. Sci. 45, 152-160.
Wang, Y., Lin, W. H., Chen, X. and Xue, H. W. (2009). Arabidopsis 5PTase13 isinvolved in root gravitropism through modulating vesicle trafficking. Cell Res.19, 1191-1204.
Welsch, R., Wust, F., Bar, C., Al-Babili, S. and Beyer, P. (2008). A third phytoenesynthase is devoted to abiotic stress-induced abscisic acid formation in rice anddefines functional diversification of phytoene synthase genes. Plant Physiol. 147,367-380.
Wu, J. Y., Wang, S., Gu, Y. C., Zhang, S. L., Publicover, S. J. and Franklin-Tong, V. E. (2011). Self-incompatibility in Papaver rhoeas activates nonspecificcation conductance permeable to Ca2+ and K+. Plant Physiol. 155, 963-973.
Xie, B., Wang, X and Hong, Z. (2010). Precocious pollen germination inArabidopsis plants with altered callose deposition during microsporogenesis.Planta 231, 809-823.
Xue, H. W., Chen, X. and Li, G. (2007). Involvement of phospholipid signaling inplant growth and hormone effects. Curr. Opin. Plant Biol. 10, 483-489.
Xue, H. W., Chen, X. and Mei, Y. (2009). Function and regulation ofphospholipid signaling in plants. Biochem. J. 421, 145-156.
Zhong, R. and Ye, Z. H. (2004). Molecular and biochemical characterization ofthree WD-repeat-domain-containing inositol polyphosphate 5-phosphatases inArabidopsis thaliana. Plant Cell Physiol. 45, 1720-1728.
Zhong, R., Burk, D. H., Morrison, W. H. and Ye, Z. H. (2004). FRAGILE FIBER3,an Arabidopsis gene encoding a type II inositol polyphosphate 5-phosphatase, isrequired for secondary wall synthesis and actin organization in fiber cells. PlantCell 16, 3242-3259.
2233RESEARCH ARTICLEIP3/Ca2+ in pollen dormancy
DEVELO
PMENT
Copyright © 2022 FDOKUMEN




















