Termite (Isoptera) assemblages in some regions of the Goiás State, Brazil
Upload
independentCategory
view
2download
0
HOUSEHOLD AND STRUCTURAL INSECTS
Influence of Environmental Factors on Activity Patterns of Incisitermesminor (Isoptera: Kalotermitidae) in Naturally Infested Logs
VERNARD R. LEWIS,1 SHAWN LEIGHTON, ROBIN TABUCHI, JAMES A. BALDWIN,2
AND MICHAEL I. HAVERTY
Division of Organisms and the Environment, Department of Environmental Science, Policy, and Management, College ofNatural Resources, University of California/Berkeley, 1301 South 46th Street, Building 478, Richmond, CA 94804!4698
J. Econ. Entomol. 106(1): 338!346 (2013); DOI: http://dx.doi.org/10.1603/EC12050
ABSTRACT Acoustic emission (AE) activity patterns were measured from seven loquat [Eriobotryajaponica (Thunb.) Lindl.] logs, "ve containing live western drywood termite [Incisitermes minor(Hagen)] infestations, and two without an active drywood termite infestation. AE activity, as well astemperature, were monitored every 3 min under unrestricted ambient conditions in a small woodenbuilding, under unrestricted ambient conditions but in constant darkness, or in a temperature-controlled cabined under constant darkness. Logs with active drywood termite infestations displayedsimilar diurnal cycles of AE activity that closely followed temperature with a peak of AE activity latein the afternoon (1700!1800 hours). When light was excluded from the building, a circadian patterncontinued and apparently was driven by temperature. When the seven logs were kept at a relativelyconstant temperature (!23 " 0.9#C) and constant darkness, the pattern of activity was closelycorrelated with temperature, even with minimal changes in temperature. Temperature is the primarydriver of activity of these drywood termites, but the effects are different when temperature is increasingor decreasing. At constant temperature, AE activity was highly correlated with the number of termites inthe logs. The possible implications of these "ndings on our understanding of drywood termite biology andhow this information may affect inspections and posttreatment evaluations are discussed.
KEY WORDS acoustic emissions, drywood termites, feeding periodicity, temperature effects
Much of the success achieved by termites in becomingdominant terrestrial organisms has been attributed totheir ability to exploit the rhythmic pattern of theappearance and disappearance of cellulose resourcesde"ned by the environmental parameters of temper-ature, humidity, precipitation, and biotic factors suchas limb- and tree-killing insects. Daily patterns in bi-ology have been coined circadian rhythms and arecommon across many phyla of plants, microbes, andanimals (Beck 1968, Schotland and Sehgal 2001). Cir-cadian (meaning approximately daily) cycles can beendogenous and set by photoperiod, or cycles of lightand dark, within the day. Circadian cycles of insectactivity may also be simply driven by exogenous fac-tors such as temperature, humidity, and light intensity(Beck 1968). Once the exogenous cue is removed oraltered, the circadian nature of the cycle disappears.These circadian cycles can be nested within the sea-sonal cycle.
Hebrant (1970) discovered an endogenous circa-dian rhythm in oxygen consumption in freshly col-lected whole colonies of the termite Cubitermes ex-
iguusMathot that were maintained at a constant 30#Cfor 3 d. He observed two maxima, one in the eveningand one in the morning, and two minima, the "rst atmidnight and the second, more expressed, in the earlyafternoon. He ascribed this variation in oxygen con-sumption to the periods of activity of the mound dur-ing the day. Relative humidity and temperatureseemed to be the main factors driving this activity, aslight did not act as a direct stimulus to C. exiguus. It isimportant to understand that this circadian cycle(rhythm) was sustained even though the phase-set-ting stimuli (or Zeitgebers), temperature and relativehumidity, were no longer inßuential.
For subterranean termites, their foraging habitshave been well studied especially for desert, Medi-terranean, and temperate climates (Haverty et al.1974, 1975, 1999, 2000; La Fage et al. 1976; Esenther1980; Su and Scheffrahn 1988; Grace et al. 1989; Jones1990; French 1991; Haagsma and Rust 1995; Forschlerand Townsend 1996; Baker and Haverty 2007). Usingvarious topical and in-ground monitors, temperatureprobes, and measures of precipitation, foraging andwood consumption were found to be highly rhythmicseasonally and closely followed changes in tempera-ture and were modi"ed by rainfall or increases in soil
1 Corresponding author, e-mail: [email protected] Paci"c Southwest Research Station, USDA Forest Service, P.O.
Box 245, Berkeley, CA 94701.
0022-0493/13/0338!0346$04.00/0 ! 2013 Entomological Society of America

moisture (Haverty et al. 1974, 1999; La Fage et al.1976).
Haverty and Nutting (1974) investigated the inßu-ence of temperature on feeding of the termite Para-neotermes simplicicornis (Banks) and the termite Het-erotermes aureus (Snyder). They found that woodconsumption, measured as milligrams of wood con-sumed per hour per gram dry weight of termite, from16 to 36#C is described by a polynomial equation of theform a $ bX % cX2. Feeding was highest for P. sim-plicicornis at 28#C and highest for H. aureus at 32#C.
Because drywood termites (family Kalotermitidae)are single piece nesters and can live deep in wood,there are few reports of seasonal or daily cycles offeeding or foraging behavior. Seasonal activity pat-terns of drywood termites are very important for theirdetection and treatment. A common seasonal activityfor drywood termites is swarming. In California, thewestern drywood termite Incisitermesminor (Hagen),swarms during the day, starting in summer and con-tinuing into fall (Ebeling 1978). Feeding and foragingare important drywood termite activities; however,little is known about when they occur or what drivesthem.Thecrypticbehaviorofdrywood termites insidewood hinders studies that explore their normal feed-ing and foraging behavior. Acoustic emission (AE)technology has been used to detect the presence ofdrywood and dampwood termites in wood (Lewis andLemaster 1991, Lemaster et al. 1997) and recently hasbeen used to explore seasonal and daily patterns for-aging and feeding activity of I. minor (Lewis et al.2011).
Lemaster et al. (1997) found no periodicity in feed-ing of I. minor in a 24-h day. However, the investiga-tion only ran for 1 wk and was conducted underconstant temperature conditions. Indrayani et al.(2006) also used AE technology to monitor I. minorfeeding when affected by different laboratory tem-peratures and relative humidity; however, they did notreport diurnal or seasonal AE activity. Using AE mon-itoring during an investigation of local chemical treat-ments in a "eld study in southern California, activityof I. minor infestations in untreated locations of struc-tures declined during winter months (V. Lewis, un-published data). Because this"eld study involved onlyfour posttreatment inspection dates, it was not possi-ble to make de"nitive statements on seasonal foragingof drywood termites.
Little research has been conducted on the move-ment patterns of drywood termites. Currently, onlythe speed of locomotion of I. minor (1.4 cm/s) inresponse to temperature and light is known (Cabreraand Rust 1994, 1996, 2000). Rust et al. (1979) inferredmovement of the drywood termite Incisitermes fruti-cavus Rust from studies that measured daily changesin temperature inside galleries for the Jojoba shrub,Simmondsia chinensis (Link); however, the daily orseasonality of movement of drywood termites withinstructures in California remains poorly understood.
Lewis et al. (2011) investigated activity of coloniesof I. minorwith acoustic emission (AE) technology innaturally infested logs. Activity, whether it was feed-
ing, excavation, or movement, was monitored for 11mo under ambient conditions in a small wooden struc-ture. AE, temperature, and humidity data were mea-sured in 3-min increments. Termite activity wasgreater during the warmer summer months comparedwith the cooler winter months. Termites in all logsdisplayed a similar daily cycle of activity, peaking inthe late afternoon. Seasonal and daily ßuctuations intermite activity were signi"cantly associated withtemperature, whereas humidity had a less noticeableeffect on termite activity.
The purpose of this study was to investigate thenature of the circadian cycle of activity in I. minor: therole or inßuence of temperature and light on patternsof feeding and foraging as measured by vibrationalactivity in naturally infested wood. By knowing whendrywood termites are most active and whether phys-ical features of their environment affect their activity,inspections and posttreatment evaluations of remedialtreatments could be greatly improved.
Materials and Methods
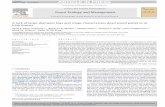
Selection and Preparation of Naturally InfestedWood. Logs from a large loquat tree [Eriobotrya ja-ponica (Thunb.) Lindl.] were collected during thesummer of 2007 from Granada Hills, CA, and broughtto our laboratory. The logs were similar in size andrevealed visual signs of drywood termite activity. Toverify that candidate logs had an active drywood ter-mite infestation, three 1-min recordings of AE activitywere taken from the center of the log (Lewis et al.2004) by using a hand-held device (Tracker, DuneganEngineering, Midland, TX). Five logs averaging !300AE counts/min were randomly selected for use inthese experiments. Two additional logs were ran-domly selected as controls. The control logs were putinto an oven (Isotemp model 655 F; Fisher, Pittsburgh,PA) at 105#C for 24 h to kill all termites within the logs(Lewis et al. 2011).MultichannelAEMonitoringEquipment.All seven
logs had a subsurface sensor installed into their longcenter by drilling a 2.4-mm-diameter hole and insert-ing the sensorprobe1.2cmdeep intowood.All logandsensor assignments were randomly chosen. A 3-m-long cable from each of the seven sensors was con-nected to a port in an AE Smart Device (Dunegan2005) and dedicated computer (Dell Corporation,Austin, TX) that stored all of the data. All logs wereplaced onto a small wooden table, and the AE SmartDevice, dedicated computer, and backup power sup-ply were placed on a wooden desk near the table (Fig.1). To prevent ants [Argentine ants, Linpethemia hu-milie (Mayr)] from attacking the termites in the logson the table, the legs of the table were placed intosmall plastic saucers (15 cm in diameter by 3 cm deep)containing a 1-cm-deep band of motor oil (SAE 30;Ace Hardware Corporation, Oak Brook, IL). Addi-tional ant prevention measures included applying a15-cm-long band of petroleum jelly (Longs Drugs,Walnut Creek, CA) to the cable end leading into themain AE Smart Device controller box. The entire
February 2013 LEWIS ET AL.: ENVIRONMENT AND ACTIVITY PATTERNS OF I. minor 339
system was visited at least weekly to check on theoperating status.Infested Logs and AE Equipment Storage. All logs,
AE, and temperature equipment were stored in a
small, wooden, 50-yr-old building at the University ofCalifornia Richmond Field Station, Richmond, CA.The building was 8.2 by 4.9 m (40.2 m2), one story inheight, and had a slab foundation. The construction
Fig. 1. Seven logs from a large loquat tree [Eriobotrya japonica (Thunb.) Lindl.] used for the study. The logs were similarin maximum diameter, length, and age. The seven small white boxes seen in the center of the photograph are AE SmartMax-modules and are used in conjunction with software to control the random order of sensor selection for AE activity duringdata collection.
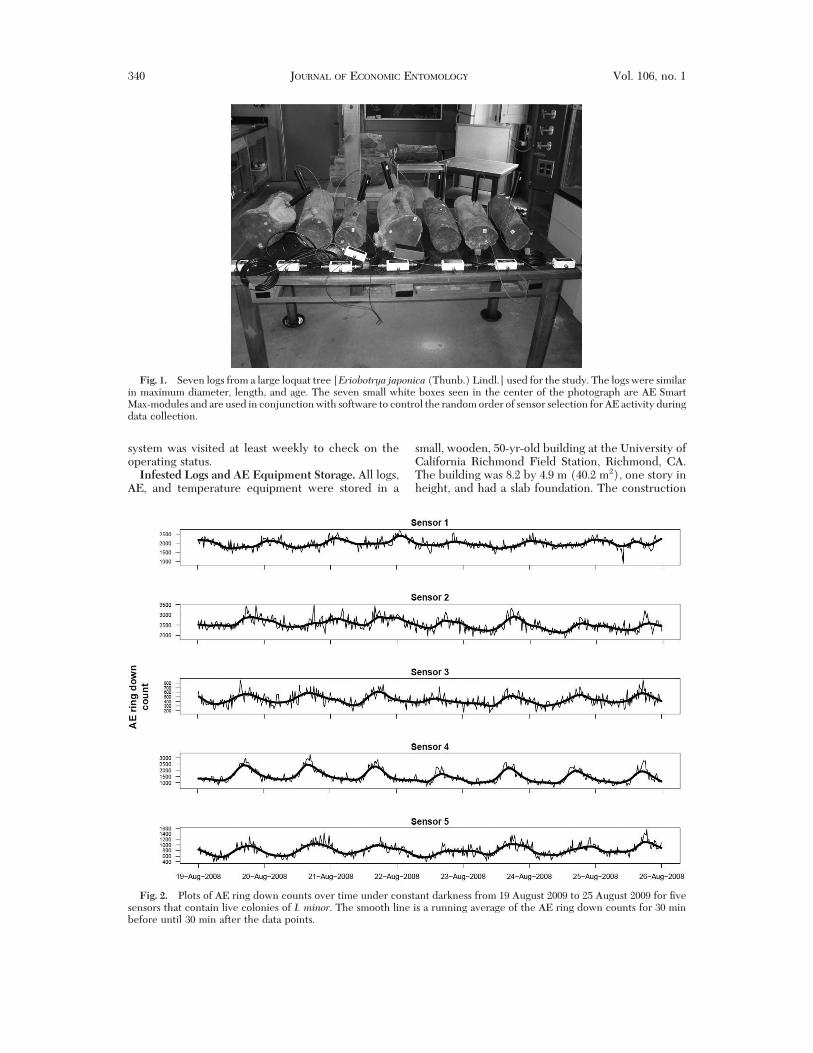
Fig. 2. Plots of AE ring down counts over time under constant darkness from 19 August 2009 to 25 August 2009 for "vesensors that contain live colonies of I. minor. The smooth line is a running average of the AE ring down counts for 30 minbefore until 30 min after the data points.
340 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 106, no. 1
type was double wall with Douglas-"r [Pseudotsugamenziesii (Mirbel) Franco V.] studs, redwood interiorwalls, and ceiling paneling. The exterior was plywoodsiding. The building had"ve windows for natural light.Three windows were on the northeast side (91.4 by60.9 cm in size) and two on the southeast side of thebuilding (121.9 by 111.8 cm and 122 by 91.4 cm in size).There was no air conditioning or heating in the build-ing; environmental conditions were strictly ambientand unrestricted. The roof was tar and shingle and wasin good condition. To minimize human traf"c into thebuilding, a sign up log was created requesting the date,time, name of individuals, and af"xed to the exteriordoor of the building before entry. A warning sign wasposted on the door to noti"ed individuals before en-tering that a vibration-sensitive research project wasbeing conducted.ResponseVariables,Data Summaries, andAnalyses.
All data were downloaded onto a high capacity (256MB), portable storage device (Kingston Data Traveler,Fountain Valley, CA) for long-term storage and futureanalysis. Twenty 3-min recordings were randomly takenamong the seven sensors for each 60-min period duringthe study. No sensor was used more than three times orless than twotimesduringa60-minperiod, and theorderof the readings were randomized. In addition, air tem-perature (Omega Engineering, Stamford, CT) was re-
corded for each 3-min AE recording and saved to anelectronic spread sheet (Excel Corporation, Lubbock,TX). A backup battery power supply (Back-ups; APCCorporate, W. Kingston, RI) was installed in the event ofunexpected power outages.
The entire AE system as described above was runfor 11 mo (15 June 2008!15 May 2009). Lewis et al.(2011) reported results and conclusions from that11-mo study. The same seven logs were used for ad-ditional experiments to determine whether circadianactivity cycles were endogenously or exogenouslycontrolled. Throughout the various studies the vari-ables measured were AE ring down counts and airtemperature. AE ring down counts were proxies forfeeding and other activity of the termites and consti-tuted the response variables of interest.AEActivityUnderConstantDarkness.To ascertain
whether cyclical activity patterns were endogenouscircadian rhythms affected by photoperiod, the win-dows were covered with black plastic sheeting to ex-clude external light. Recordings under conditions oftotal darkness were made from 18 August 2009 to 25August 2009. For each sensor, including those in thecontrol logs, the individual 3-min readings were usedfor the 7-d period from 19 August 2009 until 25 August2009 for display purposes. The resulting smooth line iscomposed of a running average of the AE ring down
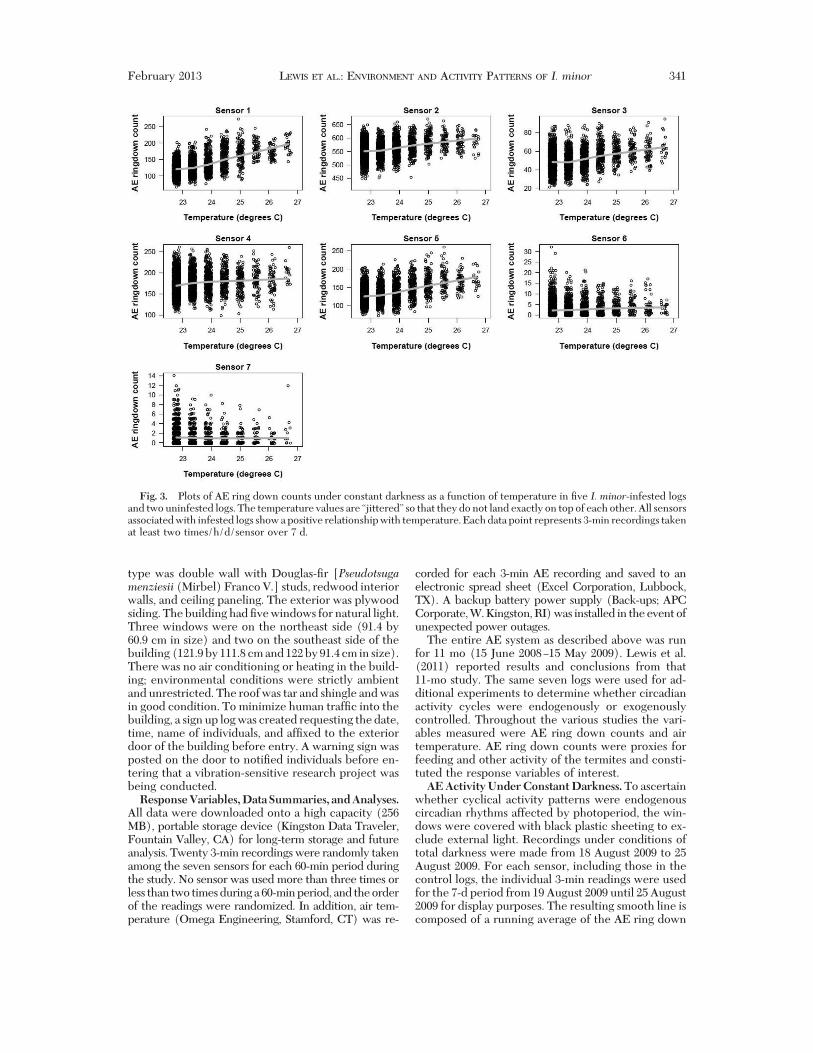
Fig. 3. Plots of AE ring down counts under constant darkness as a function of temperature in "ve I. minor-infested logsand two uninfested logs. The temperature values are “jittered” so that they do not land exactly on top of each other. All sensorsassociated with infested logs show a positive relationship with temperature. Each data point represents 3-min recordings takenat least two times/h/d/sensor over 7 d.
February 2013 LEWIS ET AL.: ENVIRONMENT AND ACTIVITY PATTERNS OF I. minor 341
counts for 30 min before until 30 min after individualdata points. The effect of temperature on AE ringdown counts was displayed for each sensor.AE Activity Under Constant Temperature. To de-
termine whether cyclical activity patterns would con-tinue “to be manifested as circadian rhythms even inthe absence of an entraining photoperiod or thermo-period” (Beck 1968), the seven logs were placed in aconstant-temperature cabinet (Precision Low Tem-
perature Incubator, model 815; 677 mm by 1,448 by 508mm, 566 liter capacity; Geneva Scienti"c LLC, Fon-tana, WI) that also excluded light. Recordings underconstant temperature conditions were made from 24November 2009 until 26 January 2010. The constant-temperature cabinet had a small opening on the topside that allowed for all wiring for AE and temperaturerecording equipment to pass into the interior. AEactivity and temperature were recorded within the cab-inet as they were under the unrestricted and constantdarkness conditions. For each sensor, including those inthe control logs, the individual 3-min readings were usedfor the63-dperiod.Theeffectof temperatureonAEringdown counts was displayed for each sensor.Effect of Termite Numbers on AE Ring DownCounts.At the conclusion of experimentation, the num-ber of surviving termites in each caste was counted afterdissection of all seven logs by using a mallet and woodchisel from15June2010until23June2010.ThemeanAEring down count resulting form the constant tempera-ture experiment was used as the dependent variable andregressed against the total number of termites (R De-velopment Core Team 2004). Data were transformed tothe natural log by using the following model:
ln (AE ring down count) & a $ b *
ln (number of termites remaining) $ error
to make the normality and constant variances of theresiduals more readily apparent.
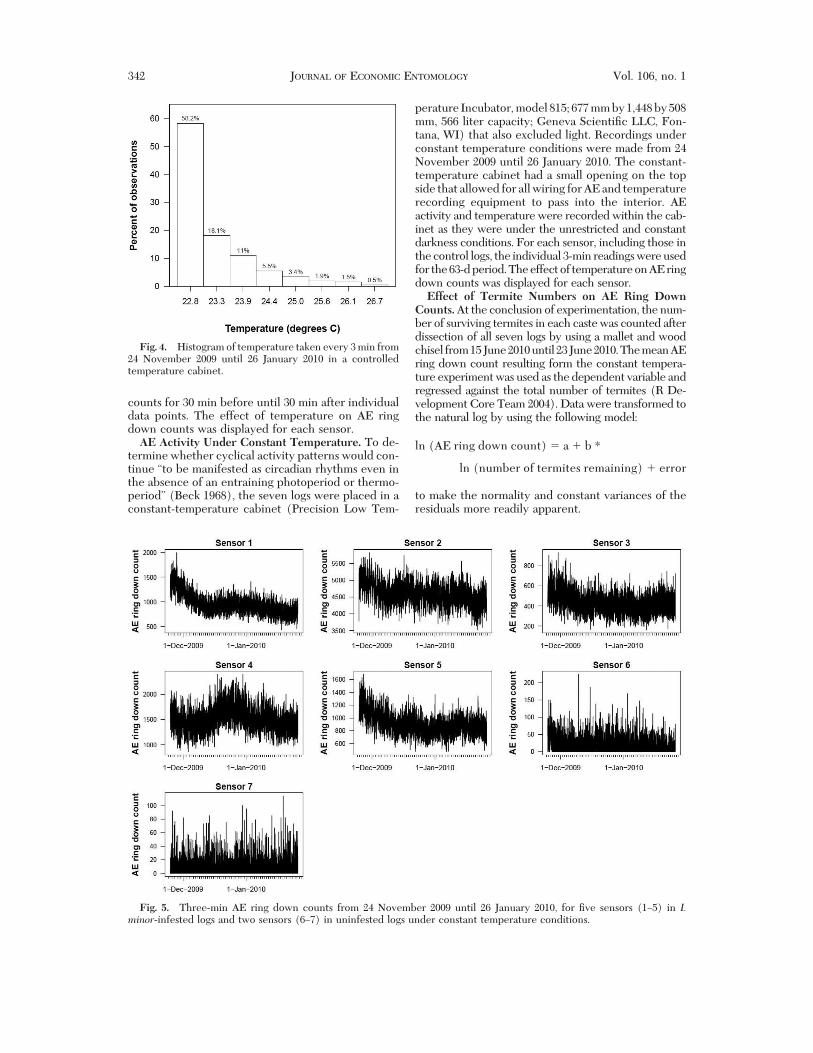
Fig. 4. Histogram of temperature taken every 3 min from24 November 2009 until 26 January 2010 in a controlledtemperature cabinet.
Fig. 5. Three-min AE ring down counts from 24 November 2009 until 26 January 2010, for "ve sensors (1!5) in I.minor-infested logs and two sensors (6!7) in uninfested logs under constant temperature conditions.
342 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 106, no. 1
Results and Discussion
Lewis et al. (2011) reported that seasonal activity,in terms of AE ring down counts, displayed a patternof increasing and decreasing values each day, primar-ily associated with daily changes in temperature. AEactivity was, in general, highest during the warmer latespring, summer, and early fall months (late April!mid-October) and lowest during the late fall through earlyspring (mid-October!mid-April). An increase in day-time temperature, or a sudden heat wave, even inJanuary and February of 2009, resulted in a burst ofincreased AE activity (Lewis et al. 2011).
Furthermore, Lewis et al. (2011) found that withinan average 24-h day, AE activity displayed a nonlinearpattern of activity. This pattern was sinusoidal in shapeand was lowest during the morning (!1,000), in-creased in the afternoon, and peaked in late afternoon(between 1,600 and 1,800), then declined until mid-morning clearly following the temperature pattern.AE Activity Under Constant Darkness. To measure
the inßuence of photoperiod on daily activity, weexcluded light from the building. AE activity remainedcyclical with apparent daily cycles of feeding activity(Fig. 2). Ambient temperature during this 1-wk periodonly varied from 22.5 to 26.5#C. The AE ring downcounts for the "ve sensors in infested logs showed apositive relationship with temperature, whereas ac-tivity in the uninfested logs was at least 10' lower than
that in the infested logs, served as an indication ofbackground AE activity, and was not positively asso-ciated with temperature (Fig. 3).AE Activity Under Constant Temperature. Tem-
peraturewas regulatedsuchthat itwas targeted for23#C,but did rise to 26.7#C on a few occasions. A histogram ofthetemperaturesthatweretakenatapproximately3-minintervals showed that 92.6% of the time the temperatureranged from 22.8 to 24.4#C) (Fig. 4). This temperatureregime resulted in AE ring down counts (Fig. 5) thatwere quantitatively very similar to those observed underthe constant darkness conditions (Fig. 2).
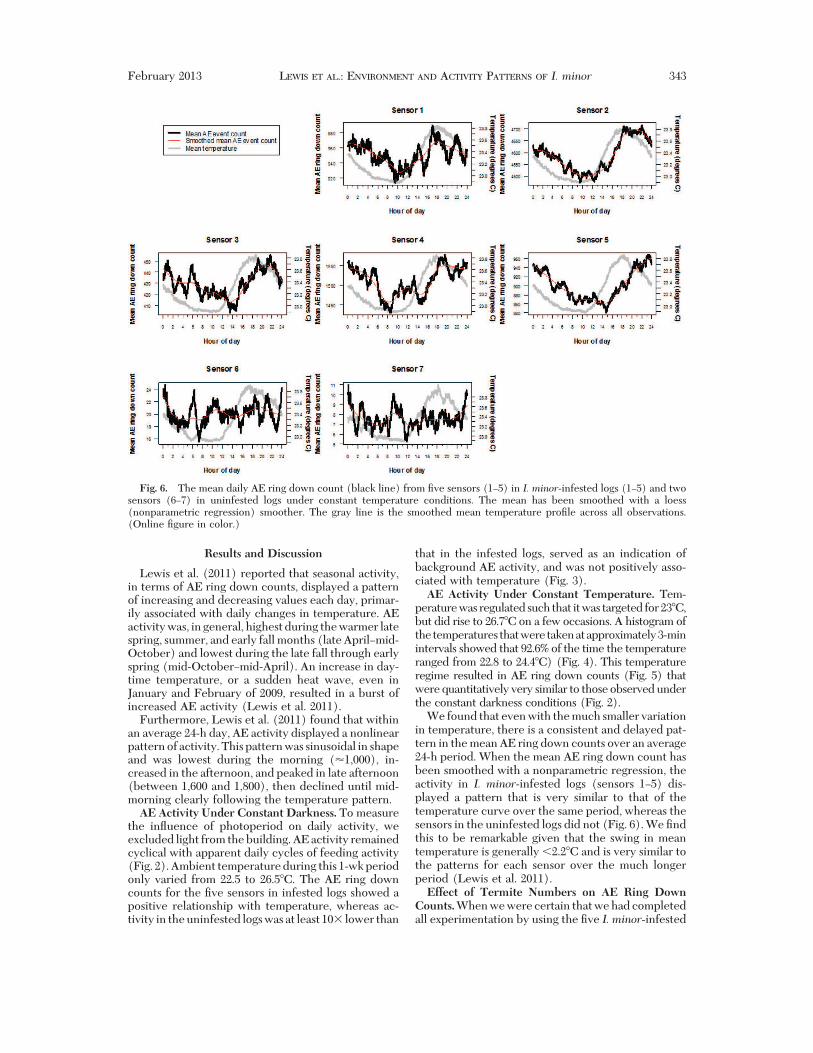
We found that even with the much smaller variationin temperature, there is a consistent and delayed pat-tern in the mean AE ring down counts over an average24-h period. When the mean AE ring down count hasbeen smoothed with a nonparametric regression, theactivity in I. minor-infested logs (sensors 1!5) dis-played a pattern that is very similar to that of thetemperature curve over the same period, whereas thesensors in the uninfested logs did not (Fig. 6). We "ndthis to be remarkable given that the swing in meantemperature is generally (2.2#C and is very similar tothe patterns for each sensor over the much longerperiod (Lewis et al. 2011).Effect of Termite Numbers on AE Ring DownCounts.When we were certain that we had completedall experimentation by using the "ve I. minor-infested
Fig. 6. The mean daily AE ring down count (black line) from "ve sensors (1!5) in I. minor-infested logs (1!5) and twosensors (6!7) in uninfested logs under constant temperature conditions. The mean has been smoothed with a loess(nonparametric regression) smoother. The gray line is the smoothed mean temperature pro"le across all observations.(Online "gure in color.)
February 2013 LEWIS ET AL.: ENVIRONMENT AND ACTIVITY PATTERNS OF I. minor 343
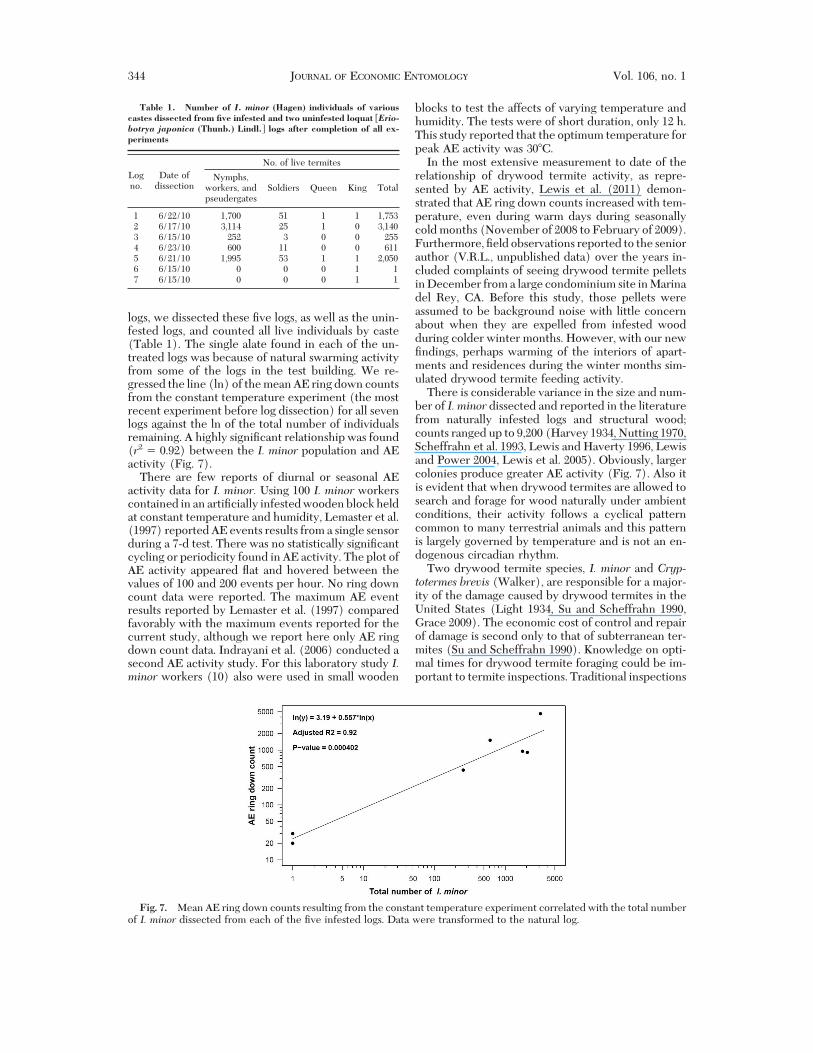
logs, we dissected these "ve logs, as well as the unin-fested logs, and counted all live individuals by caste(Table 1). The single alate found in each of the un-treated logs was because of natural swarming activityfrom some of the logs in the test building. We re-gressed the line (ln) of the mean AE ring down countsfrom the constant temperature experiment (the mostrecent experiment before log dissection) for all sevenlogs against the ln of the total number of individualsremaining. A highly signi"cant relationship was found(r2 & 0.92) between the I. minor population and AEactivity (Fig. 7).
There are few reports of diurnal or seasonal AEactivity data for I. minor. Using 100 I. minor workerscontained in an arti"cially infested wooden block heldat constant temperature and humidity, Lemaster et al.(1997) reported AE events results from a single sensorduring a 7-d test. There was no statistically signi"cantcycling or periodicity found in AE activity. The plot ofAE activity appeared ßat and hovered between thevalues of 100 and 200 events per hour. No ring downcount data were reported. The maximum AE eventresults reported by Lemaster et al. (1997) comparedfavorably with the maximum events reported for thecurrent study, although we report here only AE ringdown count data. Indrayani et al. (2006) conducted asecond AE activity study. For this laboratory study I.minor workers (10) also were used in small wooden
blocks to test the affects of varying temperature andhumidity. The tests were of short duration, only 12 h.This study reported that the optimum temperature forpeak AE activity was 30#C.
In the most extensive measurement to date of therelationship of drywood termite activity, as repre-sented by AE activity, Lewis et al. (2011) demon-strated that AE ring down counts increased with tem-perature, even during warm days during seasonallycold months (November of 2008 to February of 2009).Furthermore,"eld observations reported to the seniorauthor (V.R.L., unpublished data) over the years in-cluded complaints of seeing drywood termite pelletsin December from a large condominium site in Marinadel Rey, CA. Before this study, those pellets wereassumed to be background noise with little concernabout when they are expelled from infested woodduring colder winter months. However, with our new"ndings, perhaps warming of the interiors of apart-ments and residences during the winter months sim-ulated drywood termite feeding activity.
There is considerable variance in the size and num-ber of I. minor dissected and reported in the literaturefrom naturally infested logs and structural wood;counts ranged up to 9,200 (Harvey 1934, Nutting 1970,Scheffrahn et al. 1993, Lewis and Haverty 1996, Lewisand Power 2004, Lewis et al. 2005). Obviously, largercolonies produce greater AE activity (Fig. 7). Also itis evident that when drywood termites are allowed tosearch and forage for wood naturally under ambientconditions, their activity follows a cyclical patterncommon to many terrestrial animals and this patternis largely governed by temperature and is not an en-dogenous circadian rhythm.
Two drywood termite species, I. minor and Cryp-totermes brevis (Walker), are responsible for a major-ity of the damage caused by drywood termites in theUnited States (Light 1934, Su and Scheffrahn 1990,Grace 2009). The economic cost of control and repairof damage is second only to that of subterranean ter-mites (Su and Scheffrahn 1990). Knowledge on opti-mal times for drywood termite foraging could be im-portant to termite inspections. Traditional inspections
Table 1. Number of I. minor (Hagen) individuals of variouscastes dissected from five infested and two uninfested loquat )Erio-botrya japonica (Thunb.) Lindl.* logs after completion of all ex-periments
Logno.
Date ofdissection
No. of live termites
Nymphs,workers, andpseudergates
Soldiers Queen King Total
1 6/22/10 1,700 51 1 1 1,7532 6/17/10 3,114 25 1 0 3,1403 6/15/10 252 3 0 0 2554 6/23/10 600 11 0 0 6115 6/21/10 1,995 53 1 1 2,0506 6/15/10 0 0 0 1 17 6/15/10 0 0 0 1 1
Fig. 7. Mean AE ring down counts resulting from the constant temperature experiment correlated with the total numberof I. minor dissected from each of the "ve infested logs. Data were transformed to the natural log.
344 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 106, no. 1

are visual and based on visual searches for damagedwood or pellets. The results from this study suggestsearches for pellets and active drywood termite infes-tations could be enhance by heating the wood to atleast 25#C before inspection to simulate foraging andfeeding, even in winter, and using AE technology todelimit infestations. Calibration of AE recordings fortemperature and the numbers of termites could ben-e"t determinations of the location(s) and extent of adrywood termite infestation.
Acknowledgments
We thank H. Dunegan and J. Farrow for their technicalassistance in designing and customizing the AE Smart deviceand related hardware to accommodate our research needs.We also thank G. Briseno for developing the software for theautomated collection of AE and temperature data into elec-tronic spreadsheets. We extend sincere thanks to theLeighton family who provided us with the infested logs usedto collect the drywood termite pellets. This research wasmade possible, in part, by contract 084-2856-5 to V.R.L. by theCalifornia Structural Pest Control Board, Department of Pes-ticide Regulation (formerly the Department of ConsumerAffairs), Sacramento, CA.
References Cited
Baker, P. B., and M. I. Haverty. 2007. Foraging populationsand distances of the desert subterranean termite, Het-erotermes aureus (Isoptera: Rhinotermitidae), associatedwith structures in southern Arizona. J. Econ. Entomol.100: 1381!1390.
Beck, S. D. 1968. Insect photoperiodism. Academic, NewYork and London, United Kingdom.
Cabrera, B. J., and M. K. Rust. 1994. The effect of tempera-ture and relative humidity on the survival and wood con-sumption of the western drywood termite, Incisitermes mi-nor (Isoptera: Kalotermitidae). Sociobiology 24: 95!113.
Cabrera, B. J., andM.K. Rust. 1996. Behavioral responses tolight and thermal gradients by the western drywood ter-mite (Isoptera: Kalotermitidae). Environ. Entomol. 25:436!445.
Cabrera, B. J., andM.K. Rust. 2000. Behavioral responses toheat in arti"cial galleries by the western drywood termite(Isoptera: Kalotermitidae). J. Agric. Urban Entomol. 17:157!171.
Dunegan,H. L. 2005. Detection of movement of termites inwood by acoustic emission techniques, US Patent6,883,375 B2, 14 pp. Assignee: H. L. Dunegan, LagunaNiguel, CA.
Ebeling,W. 1978. Urban entomology. University of Califor-nia Press, Berkeley, CA.
Esenther, G. R. 1980. Estimating the size of subterraneantermite colonies by a release-recapture technique, pp.1!5. In Proceedings, 11th Annual Meeting of the Inter-national Research Group on Wood Preservation, 5!9 May1980, Raleigh, NC (IRG Document No. IRG/WP/1112).IRG Secretariat, Stockholm, Sweden.
Forschler, B. T., and M. L. Townsend. 1996. Mark-release-recapture estimates forReticulitermes spp. (Isoptera: Rhi-notermitidae) colony foraging populations from Georgia,U.S.A. Environ. Entomol. 25: 952!962.
French, J.R.J. 1991. Baits and foraging behavior of Austra-lian species of Coptotermes. Sociobiology 19: 171!186.
Grace, J. K. 2009. What can fecal pellets tell us about crypticdrywood termites (Isoptera: Kalotermitidae)? pp. 1!12.
InProceedings, 40th Annual Meeting of the InternationalResearch Group on Wood Protection, 24!28 May 2009,Beijing, China (IRG Document No. IRG/WP/20407).IRG Secretariat, Stockholm, Sweden.
Grace, J. K., A. Abdallay, and K. R. Farr. 1989. Eastern sub-terranean termite (Isoptera: Rhinotermitidae) foragingterritories andpopulations inToronto.Can.Entomol. 121:551!556.
Haagsma, K. A., andM.K. Rust. 1995. Colony size estimates,foraging trends, and physiological characteristics of thewestern subterranean termite (Isoptera: Rhinotermiti-dae). Environ. Entomol. 24: 1520!1528.
Harvey, P. A. 1934. Life history of Kalotermes minor, pp. 208!224. In C. A. Kofoid, S. F. Light, A. C. Horner, M. Randall,W. B. Herms, and E. E. Bowe (eds.), Termites and termitecontrol. University of California Press, Berkeley, CA.
Haverty, M. I., and W. L. Nutting. 1974. Natural wood-consumption rates and survival of a dry-wood and a sub-terranean termite at constant temperatures. Ann. Ento-mol. Soc. Am. 67: 153!157.
Haverty, M. I., J. P. La Fage, and W. L. Nutting. 1974. Sea-sonal activity and environmental control of foraging ofthe subterranean termite, Heterotermes aureus (Sny-der), in a desert grassland. Life Sci. 15: 1091!1101.
Haverty, M. I., W. L. Nutting, and J. P. La Fage. 1975. Den-sity of colonies and spatial distribution of foraging terri-tories of the desert subterranean termite, Heterotermesaureus (Snyder). Environ. Entomol. 4: 105!109.
Haverty, M. I., G. M. Getty, K. A. Copren, and V. R. Lewis.1999. Seasonal foraging and feeding behavior of Reticu-litermes spp. (Isoptera: Rhinotermitidae) in a wildlandand a residential location in northern California. Environ.Entomol. 28: 1077!1084.
Haverty,M.I.,G.M.Getty,K.A.Copren,andV.R.Lewis. 2000.Size and dispersion of colonies of Reticulitermes (Isoptera:Rhinotermitidae) in a wildland and a residential location innorthern California. Environ. Entomol. 29: 241!249.
Hebrant, F. 1970. Circadian rhythm of respiratory metab-olism in whole colonies of the termite, Cubitermes ex-iguus. J. Insect Physiol. 16: 1229!1235.
Indrayani, Y., T. Yoshimura, Y. Yanase, Y. Fujii, and Y. Ima-mura. 2006. Evaluation of the temperature and relativehumidity preferences of the western dry-wood termiteIncisitermesminor (Hagen) using acoustic emission (AE)monitoring. J. Wood Sci. 52: 1!4.
Jones, S. C. 1990. Delineation of Heterotermes aureus(Isoptera: Rhinotermitidae) foraging territories in a So-noran desert grassland. Environ. Entomol. 19: 1047!1054.
La Fage, J. P., M. I. Haverty, and W. L. Nutting. 1976. En-vironmental factors correlated with the foraging behaviorof a desert subterranean termite, Gnathamitermes per-plexus (Banks) (Isoptera: Termitidae). Sociobiology 2:155!169.
Lemaster, R. L., F. C. Beall, and V. R. Lewis. 1997. Detectionof termites with acoustic emission. For. Prod. J. 47: 75!79.
Lewis, V. R., and M. I. Haverty. 1996. Evaluation of sixtechniques for control of the western drywood termite(Isoptera: Kalotermitidae) in structures. J. Econ. Ento-mol. 89: 922!934.
Lewis, V. R., and R. L. Lemaster. 1991. The potential ofusing acoustical emission to detect termites within wood,pp. 34!37. InM. I. Haverty and W. W. Wilcox (technicalcoordinators), Proceedings, symposium on current re-search on wood-destroying organisms and future pros-pects for protecting wood in use. U.S. Dep. Agric. ForestService Gen. Tech. Rep. PSW-128.
Lewis, V. R., and A. B. Power. 2004. Thiamethoxam trialupdate. Pest Control Technol. 9: 74, 76, 78, and 80!81.
February 2013 LEWIS ET AL.: ENVIRONMENT AND ACTIVITY PATTERNS OF I. minor 345

Lewis, V. R., A. B. Power, and M. I. Haverty. 2004. Surfaceand subsurface sensor performance in acoustically de-tecting the western drywood termite in naturally infestedboards. For. Prod. J. 54: 57!62.
Lewis, V. R., A. B. Power, and G. M. Getty. 2005. Fieldevaluation of Thiamethoxam 2SC for the management ofthe western drywood termite (Isoptera: Kalotermitidae),pp. 331!336. In: C. Y. Lee and W. H. Robinson (eds.),Proceedings of the 5th International Conference on Ur-ban Pests, 10!13 July 2005, Suntec, Singapore. PerniagaanPhÕng@P & Y Desing, Network, Malaysia.
Lewis, V., S. Leighton, R. Tabuchi, andM.Haverty. 2011. Sea-sonal and daily patterns in activity of the western drywoodtermite, Incisitermes minor (Hagen). Insects 2: 555!563.
Light, S. F. 1934. The distribution and biology of the com-mon dry-wood termite,Kalotermes minor, pp. 201!207. InC. A. Kofoid, S. F. Light, A. C. Horner, M. Randall, W. B.Herms, and E. E. Bowe (eds.), Termites and termitecontrol. University of California Press, Berkeley, CA.
Nutting, W. L. 1970. Composition and size of some termitecolonies in Arizona and Mexico. Ann. Entomol. Soc. Am.63: 1105!1110.
R Development Core Team. 2004. A language and environ-ment for statistical computing. R Foundation for StatisticalComputing, Vienna, Austria. (http:/www.R-project.org).
Rust, M. E., D. A. Reierson, and R. H. Scheffrahn. 1979.Comparative habits, host utilization and xeric adaptationsof the southwestern drywood termites, Incisitermes fru-ticavus Rust and Incisitermes minor (Hagen) (Isoptera:Kalotermitidae). Sociobiology 4: 239!255.
Scheffrahn,R.H.,W.P.Robbins, P. Busey,N.-Y. Su, andR.K.Mueller. 1993. Evaluation of a novel, hand-held, acous-tic emissions detector to monitor termites (Isoptera: Ka-lotermitidae, Rhinotermitidae) in wood. J. Econ. Ento-mol. 86: 1720!1729.
Schotland, P., and A. Sehgal. 2001. Molecular control ofDrosophila circadian rhythms, pp. 15!30. In D. L. Den-linger, J. M. Giebultowicz, and D. S. Saunders (eds.),Insect timing: circadian rhythmicity to seasonality.Elsevier, Amsterdam, the Netherlands.
Su, N.-Y., and R. H. Scheffrahn. 1988. Foraging populationand territory of the Formosan subterranean termite(Isoptera: Rhinotermitidae) in an urban environment.Sociobiology 14: 353!359.
Su, N.-Y., and R. H. Scheffrahn. 1990. Economically impor-tant termites in the United States and their control. So-ciobiology 17: 77!94.
Received 1 February 2012; accepted 6 December 2012.
346 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 106, no. 1
Copyright © 2022 FDOKUMEN