
Induction of phospholipase- and flagellar synthesis in Serratia liquefaciens is controlled by...
11
Molecular Microbiology (1995) 15(3), 445-454 Induction of phospholipase- and f Serratia iiquefaciens is controlled flagellar master operon flhD lesis m ission of the Michael GJvskov,'* Leo Eberl,^ Gunna Christiansen.^ Michael J. Benedlk^ and Soren Molin' ^ Department of Microbiology, BuHding 221, The Tedmica! University of Dentvark. DK-2800 Lyngby, Denmark. ^Institute of Medical Microbiology. The Bartholin Building. University of Aarhus, DK-8000 Arhus C. Denmark. ^Department of Biochemical and Biophysical Sciences, University of Houston. Houston. Texas 77204-5934, USA. Summary When a liquid culture of Serratia spp. reaches the tast part of the logarithmic phase of growth it induces the synthesis of several extracellular hydrolytic enzymes. In this communication we show that synthesis and secretion ofthe extracellular phospholipase is coupled to expression of flagella. Expression of flagella is demonstrated to follow a growth-phase-dependent pattern. Cloning, complementation studies and DNA- sequencing analysis has identified a genetic region in Serratia //guefac/ens which exhibits extensive homol- ogy to the Escherichia coli flhD flagellar master operon. Interruption of the chromosomal flhD operon in S. lique- faciens results in non-flagellated and phospholipase- negative cells, but the synthesis of other exoenzymes is not affected. By placing the flhD operon under the control of a foreign inducible promoter we have shown that increased transcription through the flhD operon leads to induction of flagellar synthesis and phospholipase expression. Introduction In the Serratia spp,, synthesis of a number of hydrolytic extracellular enzymes (nuciease, lipase, phospholipase, proteases and chitinases) is induced in the lafe log phase and takes place during entry into the stationary phase (Heller, 1979), Although expression of extracellular proteins is often related to the growth phase of the bacterial culture there is at the present no evidence of a common regulator of gene expression. Induction of the nuclease, chifinases and the lipase is controlled by the recA, lexA Received 3 May, 1994; revised 5 October, 1994; accepted 7 October, 1994, *For correspondence, Tel, 45933422; Fax 45932809. SOS system which contrasts with the situation for metallo- and serineproteasGS (Ball et ai, 1990) and the phospholi- pase (M, J, Benedik and M- Givskov. unpublished). Cloned phospholipase and metalloprotease follow a growth-phase- dependent expression pattern even when expressed in Escherichia coH (G\\/sko\f etai, 1988; Letoffe etai. 1991), In the case of phospholipase synthesis this growth-phase- dependent regulation is mainly the result of the activafion of the pA promoter, one of two promoters positioned upstream of the phospholipase operon (Givskov and Molin, 1992). By the use of different E. coti strains we have identified two giobai regulators, fihD and cAMP- CRP, as components involved in fhe transcriptional con- trol of phospholipase expression (Givskov and Molin, 1992). An additional feature of phosphoiipase expression is the contribution from readthrough transcription of an upstream gene, orfX, which stimulates expression under conditions of low oxygen content and iiigh growth tempera- tures (Givskov and Molin, 1992). From studies on oloned metalloprotease and prt~lacZ fusions in E. coli, we have found that inactivating mutations in cya, cip or flhD do not affect protease expression (M. J. Benedik and M- Givs- kov, unpublished). The flhD operon, which consists of the two genes flhD and flhC, constitutes a master regulator system respons- ible for expression of the entire flagellar/chemotactic regu- lon in E. co//and in Sa!nionel!a typhimurium. tt has been speculafed that the gene products FlhD and FlhC act in combination as a specialized sigma factor which, in turn, controls expression of flagellin, chemotaxis and molilify genes via the action of a specialized flagellar sigma factor, a'' (Komeda, 1982; Bartett et al... 1988: Helmann, 1991; Macnab, 1992). In E, coli, GAMP-CRP binding is required for expression of the master operon. which makes the whole flagellar regulon sensitive to catabolite repression (Silverman and Simon, 1977: Shi et ai, 1992), Further- more, expression of the master operon is also dependent on the presence of the proteins DnaK, DnaJ and GrpE (Shi ef ai, 1992) and it has been suggested by these authors that inhibition of mofility at 42 C is caused by induction of the heat-shock response. In order fo investigate if synthesis of extracellular phos- pholipase and flagella in Serratia liquefaciens is controlled by the global regulators equivalent fo those identified in
Transcript of Induction of phospholipase- and flagellar synthesis in Serratia liquefaciens is controlled by...

Molecular Microbiology (1995) 15(3), 445-454
Induction of phospholipase- and fSerratia iiquefaciens is controlledflagellar master operon flhD
lesis mission of the
Michael GJvskov,'* Leo Eberl,̂ Gunna Christiansen.^Michael J. Benedlk^ and Soren Molin'^ Department of Microbiology, BuHding 221, The Tedmica!University of Dentvark. DK-2800 Lyngby, Denmark.^Institute of Medical Microbiology. The Bartholin Building.University of Aarhus, DK-8000 Arhus C. Denmark.^Department of Biochemical and Biophysical Sciences,University of Houston. Houston. Texas 77204-5934, USA.
Summary
When a liquid culture of Serratia spp. reaches the tastpart of the logarithmic phase of growth it induces thesynthesis of several extracellular hydrolytic enzymes.In this communication we show that synthesis andsecretion ofthe extracellular phospholipase is coupledto expression of flagella. Expression of flagella isdemonstrated to follow a growth-phase-dependentpattern. Cloning, complementation studies and DNA-sequencing analysis has identified a genetic regionin Serratia //guefac/ens which exhibits extensive homol-ogy to the Escherichia coli flhD flagellar master operon.Interruption of the chromosomal flhD operon in S. lique-faciens results in non-flagellated and phospholipase-negative cells, but the synthesis of other exoenzymesis not affected. By placing the flhD operon under thecontrol of a foreign inducible promoter we haveshown that increased transcription through the flhDoperon leads to induction of flagellar synthesis andphospholipase expression.
Introduction
In the Serratia spp,, synthesis of a number of hydrolyticextracellular enzymes (nuciease, lipase, phospholipase,proteases and chitinases) is induced in the lafe logphase and takes place during entry into the stationaryphase (Heller, 1979), Although expression of extracellularproteins is often related to the growth phase of the bacterialculture there is at the present no evidence of a commonregulator of gene expression. Induction of the nuclease,chifinases and the lipase is controlled by the recA, lexA
Received 3 May, 1994; revised 5 October, 1994; accepted 7 October,1994, *For correspondence, Tel, 45933422; Fax 45932809.
SOS system which contrasts with the situation for metallo-and serineproteasGS (Ball et ai, 1990) and the phospholi-pase (M, J, Benedik and M- Givskov. unpublished). Clonedphospholipase and metalloprotease follow a growth-phase-dependent expression pattern even when expressed inEscherichia coH (G\\/sko\f etai, 1988; Letoffe etai. 1991),In the case of phospholipase synthesis this growth-phase-dependent regulation is mainly the result of the activafionof the pA promoter, one of two promoters positionedupstream of the phospholipase operon (Givskov andMolin, 1992). By the use of different E. coti strains wehave identified two giobai regulators, fihD and cAMP-CRP, as components involved in fhe transcriptional con-trol of phospholipase expression (Givskov and Molin,1992). An additional feature of phosphoiipase expressionis the contribution from readthrough transcription of anupstream gene, orfX, which stimulates expression underconditions of low oxygen content and iiigh growth tempera-tures (Givskov and Molin, 1992). From studies on olonedmetalloprotease and prt~lacZ fusions in E. coli, we havefound that inactivating mutations in cya, cip or flhD donot affect protease expression (M. J. Benedik and M- Givs-kov, unpublished).
The flhD operon, which consists of the two genes flhDand flhC, constitutes a master regulator system respons-ible for expression of the entire flagellar/chemotactic regu-lon in E. co//and in Sa!nionel!a typhimurium. tt has beenspeculafed that the gene products FlhD and FlhC act incombination as a specialized sigma factor which, in turn,controls expression of flagellin, chemotaxis and molilifygenes via the action of a specialized flagellar sigma factor,a'' (Komeda, 1982; Bartett et al... 1988: Helmann, 1991;Macnab, 1992). In E, coli, GAMP-CRP binding is requiredfor expression of the master operon. which makes thewhole flagellar regulon sensitive to catabolite repression(Silverman and Simon, 1977: Shi et ai, 1992), Further-more, expression of the master operon is also dependenton the presence of the proteins DnaK, DnaJ and GrpE(Shi ef ai, 1992) and it has been suggested by theseauthors that inhibition of mofility at 42 C is caused byinduction of the heat-shock response.
In order fo investigate if synthesis of extracellular phos-pholipase and flagella in Serratia liquefaciens is controlledby the global regulators equivalent fo those identified in

446 M. Givskov et ai.
£, colL we cloned and sequenced fhe flhD operon fromS. liquefaciens. This enabled us to inacfivafe the chromo-somal operon present in S. Hquefaciens. By placing theoperon under the transcriptional control of a foreign induc-ibie promoter we were ahle to study the effect increasedexpression of this operon had on phospholipase- and fia-geliar synthesis as weil as on motility.
Results
Cioning and sequencing of the S. liquefaciens flhDoperon
It has been shown that in E. coH the f!hD operon controlsexpression and secretion of phospholipase from cells har-bouring fhe S. Hquefacietis phospholipase operon on aplasmid (Givskov and Molin, 1992:1993). Thus, a fihDs\mw\such as MC4100 will, even if it carries a phospholipase-encoding plasmid such as pMG30, appear to be com-pletely phosphoiipase negative (Phi ) when tested onegg-yolk indicator plates,
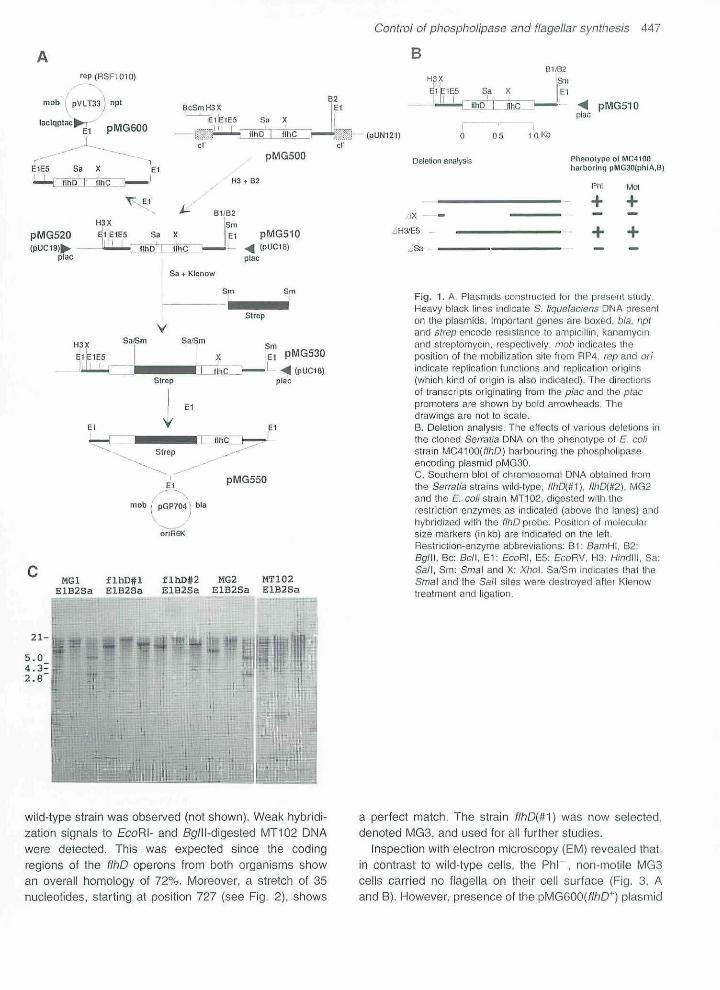
MC4100 cells carrying plasmid pMG30 were trans-formed with a genomic bank derived from S. liguefa-ciens. Formation of Phi*" colonies would be expected toresult from the presence of a plasmid carrying chromoso-mal DNA complementing the fihD locus of the host. Onesuch Phi' ciono was isolated (see Fig. IA) and desig-nated pMG500, Restriction analysis of this plasmidrevealed that the cloning vector pUN121 (Nilsson et a!.,1983) carried two EcoRI fraginents of 0.2 kb and 1,5 kb,respectively. Following subcloning into pUC18 and 19(Fig, 1A) coupled with elimination of the unique internaiSa/i site, it was found thaf the piasmid no longer inducedexpression of phosphoiipase from the MC4100 cells carry-ing pMGSO (Fig, 18), On the other hand, deletion of theH/nd!li-EcoRV fragment had no influence on stimulationof phospholipase expression. A direct link between phos-pholipase expression and motility was observed in thatpMG510 restored the motiiity-negative phenotype ofMC4100 while the 5a/l-de!eted derivative failed to comple-ment this host mutation.
The DNA sequence of the cloned Setratia DNA inpUN121 exhibited significant homology to the flhD operonof E. coH (Fig. 2), The E. coll FlhD protein (119 aminoacids) showed 73% homology to the deduced SerratiaFihD polypeptide (119 amino acids) and the E. coll FlhCprotein (192 amino acids) showed 80% homology to thededuced Serratia FlhC polypeptide (201 amino acids). TheDNA sequence upstream from flhD exhibits two dyad sym-metries and sequences with weak homology to E. coHcAMP-CRP-binding sites (TGTGA,N6,TCACA), There isno obvious match to cr'*̂ promoter recognition sequences,but the sequence CATC,N14,TACGCTCG (boxed inFig. 2) shows partial homology to CATC,N15.ACCGCTAA,
a sequence suggested fo be the promoter for the E, ooliflhD opemn (Barilett et ai, 1988; Macnab, 1992), Exceptfor the above, the DNA sequences upstream and down-stream from the fIbD operon do not show homology tothe corresponding E. ooli sequences.
Inactivation ofthe chromosoman\hD gene
The pMG550 suicide vector canying a streptomycin-resistance-encoding region inserted in the flhD gene wasintroduced into S. Hquefaciens wild-type cells (Fig. 1A),This enabled selection of streptomycin-resistant cloneswhich incorporated the streptomycin marker fragmentinto the chromosomal flhD gene by the process of homolo-gous recombination. The selection and screening was per-formed directly on egg-yolk indioator plates whichidentified Setratia clones expressing the two differenf phe-notypes. Phi' and Phi", Ten clones of each were furthertested for motiiity- All Phi" colonies appeared non-motile, whereas the Phi' colonies were all motile.
The Serratia f!hD operon was cloned into the broad-host-range vector pVLT33 (de Lorenzo et a!., 1993), inan orientation allowing transcription of the HhD operonfrom the vector !acl'^-ptac expression system (Fig, IA).This plasmid, denoted pMG600, was established in threeof the Phi non-motile Serratia slrains- All kanamycin-resistant colonies turned out to be Phi"' and motile. Twostrains in which the phospholipase and motility pheno-types could he complemented by introduction of pMG600were selected and denoted WiD(#1) and f!hD{¥2) and sub-jected to a Southern blot analysis.
The 1-5kbp ScoRI fragment from pMG510, carrying theentire flhDope\or\, was used as a probe against chromoso-mal DNA obtained from the wild-type strain, flhD{m) andf!hD{#2), MG2 (phlA) and the E coli strain MT102(Fig, 1C)- EcoRI-digested wild type and MG2 DNArevealed the presence of homologous DNA in thosestrains. However, the size of the EcoRI fragmentappeared to be much bigger, in the range of 10 kb, thanthe corresponding probe fragment, indicating that thecloned DNA had been subject to either nuclease attack(the Setratia strain produces a powerful nuclease),mechanical shearing or EcoRI star activity, Sa/i-digestedwild-type and MG2 DNA revealed the presence of fwo frag-ments with homology to the probe, as would be expectedfrom the DNA sequence (Fig, 2)- The two flhD strainsshowed that the EcoRI fragment with homology to theprobe was larger than the corresponding wild-type frag-menf and that the Sa!\ site in the flhD gene had been inter-rupted by insertion, BglW and Sa/I digests indicated thatonly the streptomycin marker had been inserted and notthe entire plasmid pMG550, since this plasmid carriessingle sites for both enzymes. When the two f!hD strainswere probed for the phIA gene no difference from the
Control of phosphoHpase and f!age!!ar synthesis 447
Brep(R5F10i0)
tob • pVLT33 ; npl
clqp,ac>^^ PMG600
BMB2H3X
BcSmH3X
' "E1JE1E5 Sa X
B2El
placpMG510
pl\/IG500E1E5 Sa X 61
(pUNl2i) 0 0,5 1-0 Kb
Dele lion analysis
flhD (IliC
T.E1
pMG510
pMG530
{pUC16)
Strep
pMG550
MGl flhD#l flhD#2 MG2 HT102ElB2Sa ElB2Sa ElB2Sa E-lB2Sa ElB2Sa
PhonolypE ol MCdiOOharboring pMG3D(plilA,B)
AX — •
j S a
Fig, 1. A, Plasmids constructed for Ihe present study.Heavy black lines indicate S. Hquefaciens DNA presenton tfie plasmids. Important genes are boxed, bla. nptand strep encode resistance to ampicillin, kanamycinand streptomycin, respeelively. mob indicates theposition of tiie mot)ilizalien site from RP4. rep and oriindicate replication functions and replicslion origins(which kind of origin is also indicated). The directionsoi transcripts originating trom the piac and the ptacpromoters are shown by bold arrowheads. Thedrawings are not to scale,
B- Deletion analysis. The effects of various deletions ihthe cloned Sermtia DNA on the phenotype of E. collstrain MC4100('"/'iD) harbouring the phospholipaseencoding plasmid pMG30,C. Southern b id oE chromosomal DNA obtained tromthe Serrada strains wiid-type, ///JD(#1), f!hD{t,2). MG2and tiie E. co//strain MT102; digested with iherestriction enzymes as indicated (above tiie lanes) andhybridized with the flhD probe- Position of molficularsize markers (inkb) are indicated on the left.Restriction-enzyme abbrevialions: B l ; BamH\. B2:BglW. Be; Bcl\. E l ; EcoRI, E5; EcoRV, H3; H/ndlll, Sa;Sa/I, Sm; Smai and X: Xho\. Sa/Sm indicates that lheSmai and the Sa/I sites were destroyed afier Klenowtreatment and iigation.
2 1 -
wild-type strain was observed (not shown). Weak hybridi-zation signals to EcoRI- and Be;/11-digested MT102 DNAwere detected. This was expected since the codingregions of the flhD operons from both organisms showan overall homology of 72%, Moreover, a stretch of 35nucleotides, starting at position 727 (see Fig, 2), shows
a perfect match. The strain fthD{lh) was now selected,denoted MG3, and used for ali further studies.
Inspection with electron microscopy (EM) revealed that,in contrast to wild-type cells, the Phi , non-moti!e MG3cells carried no flagelia on their cell surface (Fig. 3, Aand B). However, presence of the pMG600{fthD*) plasmid

448 M. Givskove\ al.
1 GCCGAATGCGTGTAbATCbATACACGTTGGTTJTACGCliCGTAGCAAATACTGAAGAGTGG
61 TCG^ATTTAGCGATTGTCG;^A^ATTAT.a-GCAGTCGTTTTTG.r\GCGATTAATTTGCCAACT
121 TTGGGAA-SiC.AACCCAAGTCACGGGCGATAAATTTATCACCCAGCGTATGTTTTTA.AAC AT- #18 >
l S l AGTCTGTCGGGATGGGGA.n.TATGGGTACGTCTGAATTACTTAAGCATATTTATGACATC,Aflhl>—^^ M G H M G T S E L L K H I Y D I N
I II
?.4"L ATCTGTCAT,sTTTACTTTTAGCACAGCGTTTGATTAACGATGA,A_riAAGCGTCCGCGATGTL S Y L L L A Q R L I N D E K A S A M F
V Q D
301 TCCGCC'-FTGGTATTGATGAAACCATGGCCGATGCGCTGGCACAGCTGACTTTGCCGCA-a_£\R L G I D E T M A D A L A Q L T L P Q M
N E T T A
3 6 1 TGG T T/LAGC T GGCCGAGACC.AA.ee A GCTCCTTTGCC AC TTCCGCTTT-A.ACGAA_:̂ .GCC AGAV K L A E T N Q L L C H P R F N E S Q T
V D 'J, l-i• - • - t sLd- S a i l - i t s t d - >
42] CCATCGAACGCC'rGACC;'VAAGAr,TCGCGCGTCGACGATCTGC.3>ACAMiTCCATACCGGCAI E R L T K E S R V D D L Q Q I H T G l
T Q Q P< #16
481 TTTTGCTATCAAGCCATTTACTGCPvAGAGCTATCGAGTAAi'vG.ACGCTAGCCCGACGAAGAL L S S H L L Q E L S S K D A S P T K KM '!:• R M D V N Q P E E A L R
541 A , , \ A C ; A G C C T G A T G A T G G T T G A G A ? A A G T A T C G T T C - ' \ A G - 2 I A G C C A A G G A C A T T C A G C T C G CR A * M M V E K S I V Q E A K D I Q L A
601 CATGGAGCTGATTTCGCTGGGAGCCCGATTGCAGATGCTGGAGAGCGAAACTCAGCTCAGM E L I S L G A R L Q M L E S E T Q L S
T
661 CCGTGGCGGCTTGATT.AA.iVCTTTACAAGGAGTTAGGGGGGCAGCCGCCCGCAAAAGGAATR G G L I K L Y K E L R G Q P P A K G M
R S P721 GTTACCGTTCTCAACCGACTGGTTTATGACCTGGGAACAAAATATTCACTCGTCGATG'i'T
L P F S T D W F M T W E Q N I H S S M FV A
781 TTAGAAGGCCTACCTGTTTCTGATG.AAAJVGCGGGCAGTGCAGCGCGGTGG.^GGCCGTGATY N A Y L F L M K S G Q C S A V E A V IC m Q h T L N G D
< S15A fll5 >X;ioI8 41 CAJiAGCCTATCGCCTCTATCTCGAGC.AGTGTCCGCAGCAGGCCGGCGAGCGCCGCTTGCT
K A Y R L Y L E Q C P Q Q A G E R R L LE G D
901 GGCCGTGACCCGTGCCTGGACGCTGGTGCGCTTCGTCGATAGCGGCATGTTGCAGCTCTCA V T R A W T L V R F V D S G M L Q L S
L E I.
961 TGCCTGCAGCTGCTGTGGCGGCGCATTTATCACTCATGCGCATCAGCCGCTGA.ATGGTTTA C S C C G G A F I T H A H Q P L N G PS N N D V G S
1021 C A T C T G C . A G T T T A T G C C . A A C C C C C A T C G C G C G C C G T A J \ A A A G A C G T A A A C T T T C G C C G C AI C S L C Q P P S R A V K R R K L S P QA Q N
< — f.2Q--lli9 >1 0 8 1 ACTGCCCGATATTATTCCTCAA-CTGCTGGACGAGCAGGTCAAACGCGCATTGAGCGCACA
L A D I I P Q L L D E Q V K R A L S A UP R V Q V -•
:i 1 4 1 TGCGCTCTCGCAATGAATA L S Q *•
Fig. 2. DNA nucleotide sequence of part ofthe cloned S. liquefaciens DNA trom the lefiEcoRV site and downstream through Ihe flhDand the flhC genes, Oligos (wilh numbers)used lo obtain DNA sequence are shown ontop of the corresponding sequence; Ihearrowhead indicates Ihe 5' to 3' direction,'<tstd' means that the sequence was obtain&dby the use of the universal sequencing primer.Regions with potential secondary structuresare indicated wiih heavy arrows over thesequence. Sequences with partial homologylo cAMP-CRP-binding sites are highlighted, Aputative promoter sequence is boxed. Thededuced amino acid sequences of the flhD-and ff/iC-encoded polypeptides are .shownbelow the DNA sequence in bold one-letternotation. Below this, posilions are shownwhere the corresponding E. co//sequence(Bartietl et ai. 1988) differs and thecorresponding amino acid is shown, A dashindicates a missing amino acid and anasterisk indicates the position of atransiational stop codon.
in MG3 lead lo flagellated cells (Fig. 3C). Addition of IPTGto the growth medium resulted in cells carrying even moreflagella than the wild-type strain (Fig. 3D).
On the basis of these analyses we conclude that thePhi and non-motile MG3 strain of S. Hquefaciens carryan inactivating insertion of fhe streptomycin marker inthe flhD gene, which controls expression of flagella andphospholipase.
To investigate whether strain MG3 was able fo mediatesecretion of phospholipase we introduced plasmidpMG330XK into the strain. As this plasmid contains thephlA''\ phlB^ genes under the control of the temperature-inducible 'kpR promoter, an increase in temperature willlead to phospholipase expression. Analogous to E. coH
MC4100f///7DJ, S. liquefaoiens MG3 harbouring this plas-mid did not secrete phospholipase upon a temperatureupshift (not shown). On the other hand, whenpMG330XK was introduced into S. liquefaciens MG2(Givskov and Molin, 1993), a strain which carries an inac-tivating insertion in the chromosomal pWAS operon. induc-tion of the A,pfl promoter led to the production ofextracellular phospholipase (not shown).
The MG3 {flhD) strain was further analysed quantita-tively for production of nuclease, protease and lipase activ-ities as compared fo their production from MG2(phlAB)and the wiid-type strain MGl, Inactivation of the flhDlocus affected neither synthesis nor secretion of theseexoenzymes (not shown).

Control of phospholipase and flagellar synthesis 449
Fig. 3, EM inspeclion of cellular flagellation. Cells were grown on LB plates overnight prior to preparation for EM as described in theExperimental procedures. A, wild-type cell of S. liquefaciens strain MGl : B, cell of strain MG3-{flhD): C, ceil of r=train MG3 harbouringpMG600(flt>D'y, D. same as (C) but the medium was suppiemented with 0,1 mM IPTG- The bar represents 1 i-im.
Expression of the flhD operon controls Itiduotion ofphospholipase synthesis
In wild-type cells, expression of phospholipase is inducedin the late log phase. Aerobic cultured cells present atlow densities in Luria-Bertani (LB) medium (below anOD450 of 0-3) do not produce a detectable amount of theenzyme. Induction of phospholipase synthesis usuallytakes place when the growing culture reaches an OD450value of approximately 0,4. At fhe same time the cultureshifts to a siightly lower growth rate. The specific enzy-matic activhy normally peaks when the culture reachesan OD450 of 2 (see Fig. 2 in Givskov and Molin, 1992),
In order to test if induction of phospholipase synthesisreflects increased flhD expression, we used S, liquefa-ciens strain MG3 carrying the intact flhD operon in transon the pMG600 plasmid, A special feature of this strain isthat expression of the flhD operon can be boosted by theaddition of IPTG. As shown in Fig, 4. there was detect-able growth-phase-dependent phospholipase synthesisin the absence of IPTG, We noted, however, that theamount of enzyme produced per ceil was low compared
to the amount produced by the wild-type strain. WhenIPTG was added, the cells responded by inducing phos-phoiipase synthesis regardless of whether the OD-,,so ofthe culture was 0.3, 0,04, or 0.004 at the time of addition
Densily (OD450)
Fig. 4. Induction of phospholipase synthesis. The Serratia MG3{fihD) strain carried the plasmid pMG600(W)D*), 0,1 mM IPTG wasadded to log-phase cells at the following culture densities: • ,
. OD45a = 0,04; n . OD,i5o = 0.2; X, no addilion.

450 M. Givskov e\ al.
Fig. 5. bM inspection of cellular flagellation, A and B, log-phasecells of the wild-type strain MGl (OD.ie;ii = 0.2); C, late-iog-phasecells of MGl (ODj;^y = 3,0); D, log-phase cells (OD,,r,o = 0,2) ot MG3{flhD) harbouring pMGf>OO{flhD') in the presence of 0,1 mM IPTG,The bar represents 1 iim.
(Fig, 3), The fully induced cultures produced almost thesame amount of enzymatic activity per cell (in all casesthe specific activity reached a maximum value of approxi-mately 0.02),
Synthesis of flagella is induced in the late log phase
We then investigated if the number of flagella per cellwas related to the growth phase of the bacterium andwhether it would be responsive to IPTG induction of theprac-controlled flhD'' operon present on pMG600,
Wiid-type log-phase cells, which had been grown forseveral generations below an OD.150 of 0-2, were collectedby centrifugafion and inspected for flagella by EM analysis.This showed that approximately 95% of the analysed cellscarried one or two flagella (Fig. 5A) and that the remaining5% had between 3 and 10 flagella (Fig, 5B), The culturewas further grown to a density of OD450 - 3.0, The EM ana-lysis showed that each ceil now carried between 10 and 20flagelia (Fig. 5C),
In a similar experiment, log-phase ceils of S. liquefa-ciens MG3 harbouring pMGSOO were grown for several
generations at low densities- At an 004^0 of 0,025, IPTGwas added and the cells were grown for three more gen-erations. The cells were then han/ested at OD^̂ ^o^O-Sand analysed by EM (Fig, 5D). All cells carried at least10 flageila.
Speed of motility can be controlled by expression ofthe flhD operon
Motiiity agar plates were used as described in the Experi-mental procedures to estimate the speed of bacterialmovement. Such experiments were repeated severaltimes and a typical result is presented in Fig, 6. As canbe seen, the motility of S. liquefaciens MG3 harbouringpMG600 was clearly dependent on the amount of supi^le-mented IPTG, Concentrations of inducer up to 1 tiM had noeffect on motility. However, increasing concentrationsresulted in increased motiiity which reached a maximumin the presence of 20 pM and 50 pM IPTG (not shown).Concentrations higher than 100|.iM were found to besomewhat inhibitory.
Discussion
The data presented in this communication show that con-trol over expression of the flagellar regulon in S. liquefa-oiens shares similarities with the situation in E. coH.Cloning of the S. liquefaciens flhD'' operon cotiipiemeti-ted fhe defective operon present in E. coli MC4100strain, and the DNA sequence analysis revealed exlen-sive homology between the coding regions of the flhD
100
Fig. 6. The speed of motiiity is controlled by expression of the flhDoperon- The Serratia MG3 strain {flhD) carried the plasmidpWlQGOQ{flhD''). The soft-agar plates were suppiemented with IPTG(1, 5, 20 and 100 nM) as indicated, A, 12h of incubation; B, 40h ofincubation, incubation was performed at room temperature-

Control of phospholipase and flagellar synthesis 451
operons from the two organisms. Insertional inactivation ofthe chromosomal f//iD locus produced non-motile and non-flagellated S, liquefaciens, and complementation studiesillustrated fhat fiagellar synthesis and motility in this strainalso require expression of the master operon.
Expression of phospholipase activity in S- liquefaciens \scieariy dependent on the presence of a functional flhDoperon. The phospholipase pA promoter exhibits a greatdegree of similarity to o-'̂ -controlied promoters expressingthird-level flageilar E. coti genes encoding proteinsinvolved in chemotaxis and the ftagellar subunit compo-nent, flagellin (Givskov and Moiin, 1992; Macnab, 1992).Interruption of nucieotides in the —10 sequence of thepA promoter indicated that this promoter was truly a'̂dependent (Givskov and Molin, 1993), The phospholipaseoperon is therefore considered to be a member of lhe fla-gellar reguion, unlike other exoenzymes such as nuclease,metalloprofease and lipase produced by this strain. Asphospholipase can be secreted from E- coH strains in theabsence of any additional cloned Serratia genes and assecretion in both E. co//and S, liquefaciens requires thepresence of an intact flhD operon (this study and Givskovand Molin, 1993), it appears that the enzyme is transportedthrough the flageliar export apparatus or, alternatively, bymeans of another putative export mechanism present inboth strains and controlled by the flhD master operon.
Transcription from the phospholipase pA promoter isinduced when cultured celis enter the last part of the logphase (Givskov and Molin, 1992). It was thereforeexpected that synthesis of flagella in S. liquefacienswould be induced in a similar way. The EM analysisdemonstrated that exponentially growing cells expressedonly one or a few flagella in contrast to cells present inthe late log phase, A similar finding was recently reportedfor£co//(Amslerefa/., 1993), Furthermore, these authorshave demonstrated that there is a direct correlationbetween the amount of fiageilin per cell and the speed ofmotility- By placing the cloned flhD operon under the con-trol of the IPTG-inducible ptac promoter, increasingamounts of IPTG lead to higher motiiity of the Serratiacells. As the EM analysis showed that the number of fla-gella per cell increased in the presence of IPTG, it is likelythat the speed of motility and the amount of flagella per eel!can be correlated.
At present, an interesting question is whether inductionof the flagellar regulon takes place in response to nutrientdeprivation or accumulation of a signal molecule reflectingthe density of the growing culture. The siightly reducedgrowth rate following the onset ot phospholipase induction(see Fig. 2 in Givskov and Molin, 1992) oould be a result ofnutrient limitation. Alternatively, it may be caused by induc-tion of costly fiagellar synthesis. Macnab (1992) states thata non-flagellated E. coH strain has an approximately 2%growth-rate advantage over a flagellated strain. At present
we have experimental indications that a mechanism whichsenses Ihe increasing density of the growing culture bymeans of an extracellular transmitter molecule operatesin S. liguefaciens. Investigation of the relationship betweenexpression of the flageliar regulon and a densily-signallingsystem is currently in progress.
Which part of the regulatory system first reoeives thestimulus leading to induction of the flagellar regulon? Thedata presented here show that transcriptionai stimulationof the flhD master operon is the soie signal .required toinduce expression of the regulon. We suggest that induc-tion of fhe flagellar regulon in the late log phase takesplace as a result of transcriptional stimulation of the masteroperon, although this remains unproved. At the present wecannot exclude the possibility that an additional gene orupstream c/s-acting sequence, working in order to enhanceexpression of the fthD operon (but missing from the clonedflhD'' fragment present on pMGGOO), is needed for fullexpression. Our data obtained with the MG3 strain showedfhat in the absence of IPTG addition, the presence of thepMGBOO piasmid did not restore phospholipase expres-sion, flagellation and motility to the level of the wiid-typestrain.
Experimental procedures
Media
Solid and liquid media contained LB (Bertani, 1951), Anti-biotics were dissolved in the media af the foilowing concentra-tions: ampicillin lOOjigml '; chloramphenicoi and kanamycin30|,jgmr ', streptomycin 50 i-ig ml" ';tetracycline, 10 fig ml '.Solid media for detection of phospholipase activity contained2 ml of egg yolk in 0.9% NaCI (50:50) per 20 ml of LB agar.
Motlfity assays
Soft agar plates for detection of motility contained LB with0.3% agarose for S. liquefaciens and 0,25% agarose lor E.col! cells- Log-phase cells were concentrated approximately100-foid by centrifugation and then added to small cylindricalholes present in the centre of the soft agar plates. After incu-bation for several hours the size of the zone resulting fromdense growth of motile bacteria could be determined.
Measurement of enzyme activities from liquid cultures
Measurement of phospholipase activities from Serratia cul-tures has been described previously (Givskov and Molin,1993)- Measurement of nuclease activity and protease activ-ity was performed as diffusion assays on overnight culturesas described by Ball ef ai (1990). Lipase activity was deter-mined from the supernatant of overnight cultured cells ashydrolysis of the colomefric subsfrate p-nitrophenyl palmitate(Winkler and Timmis, 1979).

452 M. Givs!<over al.
Table 1, Bacterial strains used.Sirain
E. coli
CSH50MT102S-17.1S-17 {Xp//)CCn8{/-pir)
De sec ip?/on
jS(!ac-pro) rpsL thiarsD139 (ara-leu}7697pro recA iisdR hsdM'S17,1 lysogenized with
Mac thi hsdRthi
Ipir
Saurce/Hefereiics
Miller (1972)Lab, slockSimon etal. (1983)de Lorenzo etal. (1993)
A(ara-leu) araO MacX74 galE qa!k phoA20 Herrero et ai (1990}thi rpsE fpoB argE(Am) recA 1 l.pir phagelysogen
MC'llOO RraD139 A(arg-lacU169) relA rpsLISOpstF2S rbsR deoC thi f!hD5301
S. liquefaciens
MG1 Wild-type Ap" Tet'̂MG2 MGl Smf̂ (phIA- phlB" )MG3 MGl Sm'"-' {flhO")
Lange and HenggH-Aronis (1991)
Givskov et al. (1986)Givskov and Molin (1992)This worl<
Construction of plasmids and strains
Bacterial strains used are listed in Table 1.The phospholipase-encoding plasmid pMG30 was con-
structed in the following way; a purified Dra\-Sca\ restrictionfragment from pMG330 (Givskov et ai, 1988), carryingXcl857/pR, pNA and phlB, was cloned into the EcoRV siteof pACYC184, giving rise to pMG30,
A genomic bank of S, Hquefaciens MG1 DNA consisted ofEcoRI-cut chromosomal DNA cloned into fhe EcoH\ site ofthe vector PUN121 (Nilsson ef a/,, 1983), Plasmid DNA wasisolated from pooled tetraclycine-resistant clones in the E coHstrain MT102 and transformed info strain MC4100(pMG30) byselection for resistance to chloramphenicol and to ampiciliin.Inserted Serratia DNA of plasmid pMG500 was subcioned intoHin6\\\-, SamHI-cut pUC18 and pUC19 by the use of theunique Hin6\\\ site in the vectors and the unique BglW site pre-sent ai the extreme right of the Serratia DNA, giving rise topMG510andpMG520(seeFig. 1A),
A purified Snial fragment obtained from plasmid mini-Tn5Sm/Sp (de Lorenzo eta!.. 1990) carrying the streptomycin-resistance gene was cloned into the unique Sa/I site ot fhef!hD'*' gene present on pMG510, giving rise to pMG530,An EcoRI restriction fragment containing the streptomycin-resistance gene flanked by Serratia DNA was cloned intothe EcoRI site of the delivery vector pGP704 (Miller andMekalanos, 1988), giving rise to pMG550. This R6K-derivedplasmid will oniy replicate in special strains such asCC118(Xp/r) and S17(Xp/r), in which the p/r gene is presentin trans. Transformation to CCM8{ApJr) enabled selectionfor plasmids cariying the streptomycin marker. pMG550 wasthen established in strain Si7(/vp/f), which then mediated con-jugaiive transfer to S, liquefaciens MGl , and selection wasmade for tetracycline- and streptomycin resistance.
The EcoRI fragment of pMG510 carrying the f!hD' operonwas cloned into the BcoRi site of ihe broad-host-range vectorpVLT33 (de Lorenzo etai, 1993) (Fig, IA), This plasmid con-ferred resistance to kanamycin. A recombinant plasmid,designated pMG600, in which the fihD' operon could be tran-scribed from the ptac promoter present in the vector, waschosen. This plasmid was transferred from the S17.1 donorstrain by conjugation to the Phi" , non-motiie S. iiquefaciensstrains, and plasmid-carrying cells were established by selec-tion for tetracyciine, streptomycin and kanamycin.
For the deletion analysis presented in Fig, 18 (he Xhoifragment present in the Serratia DNA of plasmid pMG510was deleted- The Sa/t site present in the Serratia DNA ofpMG,510 was destroyed by Sa/I digestion, nucleotide fillingof the sticky ends by the action of Klenow polymerase, andfinally leligated- In order to delete the lefi part of the clonedSerratia DNA, pMG510 and pMG520 were cut with /^/ndllland EcoRV, treated with Klenow polymerase to remove thesticky ends and finally religafed.
The plasmid pMG330XK was constructed in the followingway: the BglW fragment from plasmid BIO (Simon et ai.1989) cari"yi[ig the nm gene for kanamycin resistance andthe RP4 moi) site was purified, treated with Klenow poiyrner-ase and cloned into the unique Seal restriction site of thebla gene of pMG330X (Givskov and Molin, 1993), This plas-mid carries inactivating mutations in the phospholipase pApromoter, therefore expression of phospholipase is controlledby the >,c/857/pR temperafure-inducible repressor/promotersystem. This plasmid was transferred from the S17,1 donorstrain by conjugation to the S. liquefaciens strains MG2 andMG3, and plasmid-carrying cells were established by selec-tion for tefracycline, streptomycin and kanamycin.
DNA nudeotide sequencing
The X/iol fragment was isolated from pMG510, cut with Sa/I,and the resultant fragments cloned into the M13 vector mp18.The universal primer was used to obtain DNA sequence. TheEcoRV-HindHi deletion derivative of pMG5'IO was used toobtain DNA sequence from the EcoRV site into the fHiDoperon by the use of the universal primer. New oligos forfurther sequencing were designed trom the obtainedsequence information. We then used M13mp18 and 19clones carrying the entire Sg/ll-H/ndlll fragment frompMG500 for sequencing of the fihO operon. Both strandswere sequenced (see Fig, 2), Preparation of M13 phagesand dideoxy chain termination DNA sequencing were per-formed according to the sequenase method of USB.
Southern blotting
Chromosomal DNA was prepared from 40 ml of exponentially

Contra! of phospholipase and f!agei!ar synthesis 453
growing cells with an OD450 of 1,0, The har^^ested cells wereresuspended in 1 ml ice-cold 0,15M NaCI, 0-001°/o gelatine,lOOmM EDTA and transferred to 125MI pre-heated (68 C)0-5M Tris-HCI (pH6.8), 20inM EDTA, 10% SDS and incu-bated for 3 min; 125 ul of 3 M sodium acetate was added andfhe nucleic acids were extracted five times in a mixture ofphenol:CHCl3:isoamylalcohol (25:24:1). The nucleic acidswere precipitated in EtOiH at - 2 0 C, harvested by a 15 mincenfrifugation in a cooled microcentrifuge and washed twiceat room temperature with 80% EtOH. The pellet vi/as driedand resuspended in 200 jil of waler.
The EcoRI fragment from pMG510 was used as a probeagainst flhD and the Sa/I fragment from pMG323 (Givskovef a/,, 1988) was used as a probe for ph!A. Bofh fragmentswere purified from a gel and labelled in a random primed pro-cess using Klenow poiymerase and digoxigenin-dUTP DNAlabelling and detection kit in accordance with the manufac-turer Boehringer Mannheim's instructions. Hybridizationswere performed at 68 C overnight and washed under condi-tions of high stringency (68 C. 0.1 -, SSC (15 IT IM NaCI,1,5mM sodium citrate), 0 ,1% SDS), Buffers and conditionswere according to Boehringer Mannheim, Hybrids weredetected in an enzyme-linked immunoassay using antidigoxi-genin alkaline phosphatase conjugate, 5-bromo-4-chloro-3-indolyl phosphate and nitroblue tetrazolium sait.
Electron microscopy
Preparation of ceiis for EM was done as described by Kiemm
and Christiansen (1987), A 25 l̂i aiiquot of the bacteria!
culture was placed on top of pariodion carbon-coated glov^-
discharged 300-mesh copper grids for 30 s, The grids were
washed with three drops of phosphate-buffered sailne foi-
lowed by staining with three drops of 1% phosphotungstic
acid (pH7,0) and blotted dry. The grids were examined in a
JEOL 100B electron microscope-
Acknowledgement
This work was supported by the Danish Centre for MicrobialEcology and a grant from the EU, We thank Linda Stabelifor exceilent technical assistance. We thank Dr Victor deLorenzo for sending us the pUT plasmids and the S17 and00118 strains. We also thank Dr Gordon S- A, B- Stewartand Dr Michael K, Winson for helpful suggestions and stimu-lating discussions.
References
Amsler, 0-D., Oho. M., and Matsumara, P, (1993) Multipiefactors underiying the maximum motiiity of Escherichia coHas cuitures enter post-exponentiai growth, J Bacteriol 175:6238-6244,
Bali, T.K,, Wasmuth, O.R,, Braunagei, S.C, and Benedik,M.J, (1990) Expression of Serratia marcescens extra-ceiiuiar proteins requires recA. J Bacterio! 172; 342-349.
Bartlett, D.H,. Frantz, B.B,, and Matzumara, P. (1988)Fiageiiar franscriptionai activators FibB and Flal: genesequences and 5' consensus sequences of operons underFlbB and Flai controi, J Bacterio! 170: 1575-1581.
Bertani, G, (1951) Studies of lysogenesis, i, Tfie mode ofphage liberation by lysogenic Eschericia coH. J Bacterioi62: 293-300,
Givskov, M,, and Molin, S, (1992) Expression of extracellularphospholipase from Serratia liquefaciens is growth-phasedependent, cataboilte repressed and regulated by anaero-biosis, Mol Microbiol 6: 1363-1374,
Givskov, M., and Molin, S, (1993) Secretion of Serratialiquefaciens phospholipase from Escherichia coH. Mo!Microbio! 8: 229-242,
Givskov, M,, Olsen, L,, and Molin, S, (1988) Cloning andexpression in Eschericia co//of the gene for an extracellularphospholipase from Serratia liquefaciens. J Bacteriol 170:5855-5862,
Helmann, J,D, (t991) Alternative sigma factors and theregulation of fiagellar gene expression. Mol Microhioi 5:2875-2882,
Heller, K-B, (1979) Lipolytic activity copurified with the outermembrane of Serratia marcesoens. J Bacterio! 140: 1120-1122,
Herrero, M., de Lorenzo, V,, and Timmis, K.N, (1990)Transposon vectors containing non-antibiotic selectionmarkers for cloning and stable chromosomal insertion offoreign DNA in Gram-negative bacteria, J Bacterio! 172;6557-6667,
Klemm, P-, and Christiansen, G, (1987) Three fim genesrequired for the regulation of iength and mediation ofadhesion of Escherichia coH type 1 fimbriae, Mol GenGenet 208: 439-445,
Komeda, Y, (1982) Fusion of flageilar operons to lactosegenes on a Mu /ac bacteriophage, J Bacteriol ^ 50: 16-26,
Lange, R,. and Hengge-Ai'onis, R- (1991) Growth phaseregulated expression of bolA and morphology of stationaryphase Escherichia coH cells are controlled by a novelSigma factor a^. J Bacterioi ^73: 4474-4481-
Letoffe, S., Delepelaire, P,, and Wandersman, 8, (1991)Oloning and expression in Fscherichia coH ot the Serratiamarcescens metalloprotease gene: secretion of lheprotease from Escherichia coH in the presence of theErwinia chrysanthemi protease secretion functions. JBacterioi 173: 2160-2166.
de Lorenzo. V., Herrero, M,, Jakubzik, U., and Timmis, K,N,(1990) Mini-Tn5 transposon derivatives for insertionmutagenesis, promoter probing, and chromosomal inser-tion of cioned DNA in Gram-negative eubacteria- JBacteriol 172: 6568-6572,
de Lorenzo, V,, Eltis, L,, Kessler, B., and Timmis, K,N. (1993)Analysis of Pseudomonas gene products using lacf^lf^iip-lac plasmids and transposons that confer conditlonaiphenotypes. Gene 123: 17-24,
IVIacnab, R,M, (1992) Genetics and i^iogenesis of t:)acferialflageiia, Annu Rev Genet 26: 131-158,
Miiier, J.H- (1972) Experiments in Molecular Genetics. ColdSpring Harbor, New York: Cold Spring Harbor LaboratoryPress,
Miiier, V,L., and Mei<aianos, J,J, (1988) A novel suicide vectorand its use in construction of insertion mutations:osmoreguiation of outer membrane proteins and virulencedeterminants in Vibrio chlolerae requires toxR. J Bacterio!170: 2575-2583,
Niisson, B,, Uhien, M,. Josephsson, S,, Batenbeck, S,, and

454 M. Givskov et ai.
Phiiipson, L. (1983) An improved positive seiection plasmidvector constructed by oiigonucleotide mediated muta-genesis- Nuc! Acids Res 11: 8019-8030.
Shi, W,, Zhou, Y,, Wiid, J,, Adler, J,, and Gross, CA, (1992)DnaK, DnaJ and GrpE are required for flageflum synthesisin Escherichia coH. J Bacteriol 174: 6256-6263,
Siiverman, M,, and Simon, M, (1977) Bacteriai flagelia, AnnuRev Microbiol 31: 397-419,
Simon, R,, Priefer, U., and Puhier, A. (1983) A broad hostrange mobilization system for in vivo genetic engineering:
Transposon mutagenesis in Gram-negative bacteria. Bio/Technology-\: 784-790,
Simon, R,, Quandt, J,, and Klipp, W- (1989) New derivativesof fransposon Tn5 for mobilization of replicons, generationof operon fusions and induction of genes in Gram-negativebacteria- Gene 80: 161-169,
Winkler, U., and Timmis, K, (1979) Glycogen, hyaluronate,and some polysaccharides greatiy enhance fhe formationof exoiipase by Serratia matcescens. JBacterioliSB: 663-670.





















