
In VitroBiosynthesis of Androgens in the Australian Lungfish, Neoceratodus forsteri
8
In Vitro Biosynthesis of Androgens in the Australian Lungfish, Neoceratodus forsteri Jean M. P. Joss,* Ashley Edwards,* and David E. Kime² *School of Biological Sciences, Macquarie University, Sydney 2109, Australia; and ²Department of Animal and Plant Sciences, The University of Sheffield, Sheffield, United Kingdom Accepted September 8, 1995 The serum concentration of testosterone was estimated from a population of wild lungfish over 6–7 years of sampling. Male lungfish were found to have high circu- lating levels of testosterone (z50 ng/ml) which varied seasonally and could be correlated with spermatogenesis as judged by testis histology. Incubation of testis tissue slices with [ 3 H]progesterone, [ 3 H]17-hydroxyprogester- one, or [ 3 H]testosterone confirmed that testosterone is the major androgen in Neoceratodus. Not even trace amounts of 11-keto- or 11b-hydroxytestosterone or 5a- dihydrotestosterone could be identified by TLC separa- tions. There was little or no conjugation of steroids by the testes, except during the spawning season, when glucuronides of androstenedione and testosterone were produced. © 1996 Academic Press, Inc. Testosterone appears to be an almost ubiquitous an- drogen formed by the testes of all vertebrates. In many groups it may be converted to derivatives that are more biologically active in the species in which they are formed. Among the fish, there is good information on androgen production by the testis of teleosts, in which testosterone is commonly converted to 11- ketotestosterone and/or 11-hydroxytestosterone (Kime, 1987, 1993). For the few elasmobranchs that have been investigated, testicular androgens seem to be similar to those of mammals, in that testosterone is predominant (Sourdaine et al., 1990). In both these fish groups testicular conjugation of steroids is not uncom- mon in contrast to mammals where conjugation is generally carried out by the liver as a means of deac- tivation and solubilization for excretion (Kime, 1993). Among amphibians, urodeles have some features in common with teleost fish, in that they commonly pro- duce 11-ketotestosterone, whereas anurans lack 11b- hydroxysteroid dehydrogenase but do have an active testicular 5a-reductase which converts testosterone to the more biologically active 5a-dihydrotestosterone (Muller, 1977; Kime and Hews, 1978a). In birds and mammals this latter enzyme is either weak or absent in the testes but may be present peripherally for con- version in target tissues (Wilson, 1975). The Dipnoi (lungfishes) are generally believed to lie phylogenetically between the fish and the tetrapods (Forey, 1986). The most recent molecular evidence confirms this position (Yokobori et al., 1994; Meyer, 1995). There is currently no information on dipnoan testicular androgen production. This present paper describes testicular androgen synthesis in vitro for the Australian lungfish, Neoceratodus forsteri, and the sea- sonal changes in circulating androgen, as these relate to stages of spermatogenesis. METHODS AND MATERIALS Animals Over a period of 6–7 years, adult lungfish were col- lected by gill netting from a tributary of the Mary General and Comparative Endocrinology 101, 256–263 (1996) Article No. 0028 0016-6480/96 $18.00 Copyright © 1996 by Academic Press, Inc. All rights of reproduction in any form reserved. 256
Transcript of In VitroBiosynthesis of Androgens in the Australian Lungfish, Neoceratodus forsteri

JOBNAME: GCE 101#3 96 PAGE: 1 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
In Vitro Biosynthesis of Androgens in the AustralianLungfish, Neoceratodus forsteri
Jean M. P. Joss,* Ashley Edwards,* and David E. Kime†*School of Biological Sciences, Macquarie University, Sydney 2109, Australia; and †Department of Animal andPlant Sciences, The University of Sheffield, Sheffield, United Kingdom
Accepted September 8, 1995
The serum concentration of testosterone was estimatedfrom a population of wild lungfish over 6–7 years ofsampling. Male lungfish were found to have high circu-lating levels of testosterone (z50 ng/ml) which variedseasonally and could be correlated with spermatogenesisas judged by testis histology. Incubation of testis tissueslices with [3H]progesterone, [3H]17-hydroxyprogester-one, or [3H]testosterone confirmed that testosterone isthe major androgen in Neoceratodus. Not even traceamounts of 11-keto- or 11b-hydroxytestosterone or 5a-dihydrotestosterone could be identified by TLC separa-tions. There was little or no conjugation of steroids bythe testes, except during the spawning season, whenglucuronides of androstenedione and testosterone wereproduced. © 1996 Academic Press, Inc.
Testosterone appears to be an almost ubiquitous an-drogen formed by the testes of all vertebrates. In manygroups it may be converted to derivatives that aremore biologically active in the species in which theyare formed. Among the fish, there is good informationon androgen production by the testis of teleosts, inwhich testosterone is commonly converted to 11-ketotestosterone and/or 11-hydroxytestosterone(Kime, 1987, 1993). For the few elasmobranchs thathave been investigated, testicular androgens seem tobe similar to those of mammals, in that testosterone ispredominant (Sourdaine et al., 1990). In both these fishgroups testicular conjugation of steroids is not uncom-mon in contrast to mammals where conjugation is
generally carried out by the liver as a means of deac-tivation and solubilization for excretion (Kime, 1993).Among amphibians, urodeles have some features in
common with teleost fish, in that they commonly pro-duce 11-ketotestosterone, whereas anurans lack 11b-hydroxysteroid dehydrogenase but do have an activetesticular 5a-reductase which converts testosterone tothe more biologically active 5a-dihydrotestosterone(Muller, 1977; Kime and Hews, 1978a). In birds andmammals this latter enzyme is either weak or absentin the testes but may be present peripherally for con-version in target tissues (Wilson, 1975).The Dipnoi (lungfishes) are generally believed to lie
phylogenetically between the fish and the tetrapods(Forey, 1986). The most recent molecular evidenceconfirms this position (Yokobori et al., 1994; Meyer,1995). There is currently no information on dipnoantesticular androgen production. This present paperdescribes testicular androgen synthesis in vitro for theAustralian lungfish, Neoceratodus forsteri, and the sea-sonal changes in circulating androgen, as these relateto stages of spermatogenesis.
METHODS AND MATERIALS
Animals
Over a period of 6–7 years, adult lungfish were col-lected by gill netting from a tributary of the Mary
General and Comparative Endocrinology 101, 256–263 (1996)Article No. 0028
0016-6480/96 $18.00Copyright © 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.256

JOBNAME: GCE 101#3 96 PAGE: 2 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
River in southeastern Queensland. Within 30 min ofcapture, the fish were weighed, measured, and ablood sample was taken without anesthesia by cardiacpuncture. They were then tagged, with microchipsembedded in the muscle behind the head, and re-leased back into the stream as part of an ongoingstudy on this population of lungfish. The bloodsamples were spun in a field centrifuge and the serumwas stored in liquid nitrogen for return to the labora-tory.In November 1992 one adult male lungfish, weigh-
ing 4 kg, was transported back to the laboratory whereit was bled, anesthetised in MS222, and killed to pro-vide fresh testis tissue for identification of steroid hor-mones. A further three adult male lungfish were simi-larly sacrificed, one each in February 1993 (6 kg), May1994 (3.5 kg), and August 1994 (5 kg), to ascertainwhether there are any seasonal differences in testicu-lar steroidogenesis.
Chemicals
[1,2,6,7-3H]Progesterone (sp act 3.68 TBq/mmol)and [1,2,6,7-3H]testosterone (sp act 3.03 TBq/mmol)were purchased from Amersham International. Hy-droxyprogesterone, 17-[1,2-3H(N)] (sp act 1.48 TBq/mmol), was purchased from New England Nuclear.All other steroids were purchased from Sigma Chemi-cal Co. except the 11-ketotestosterone which was pro-vided by David Kime.
Incubations
Immediately following killing of the lungfish in No-vember, February, and May, three replicates of 100 mgof testis (taken from the midportion of the testis) werefinely sliced and incubated in 25-ml glass conicalflasks with 370 kBq [3H]progesterone in 2 ml 0.025 MHepes buffered Leibovitz culture medium, pH 7.6, atroom temperature (z24°–slightly higher than meantemperature of 23° for spring and autumn in lungfishnatural habitat), with gentle shaking for 3 hr. In Au-gust, the progesterone substrate was replaced with370 kBq [3H]17-hydroxy progesterone. A similar threereplicates of testis tissue were incubated with 370 kBq[3H]testosterone for all four of the lungfish.On completion of incubation, the medium was sepa-
rated from the tissue and extracted twice in 5× volumeof dichloromethane. The dichloromethane extracts
were allowed to evaporate in a fume hood, and thesteroids were redissolved in 2 ml methanol and storedat −80° for further analysis. A 100-ml aliquot of each ofthe aqueous residues from the dichloromethane ex-tractions was counted on a Packard, automatic Tri-Carb spectrometer. From the progesterone and testos-terone incubates of the lungfish sacrificed in Novem-ber, February, and May, the aqueous layerrepresented <4% of the total radioactivity. Since thiswas low, no extraction of conjugated steroids was at-tempted from these samples. In August, the percent-age of counts in the aqueous layer after extraction ofthe testis incubates with both 17-hydroxyprogesteroneand testosterone as substrate was >15%. Accordingly,these aqueous residues were incubated for 48 hr at 37°with b-glucuronidase (Boehringer-Mannheim), 50mlbuffered to pH 4.8 with 1 ml 0.2 M acetate buffer.Following incubation, the medium was again ex-tracted twice in 5× volume of dichloromethane. Thistime the aqueous layer contained <4% of the radioac-tivity and so no further digestion of conjugated ste-roids (e.g., with sulphatase) was attempted.
Chromatography
Thin-layer chromatography (TLC) was carried outon precoated plastic sheets (Merck Keiselgel 60 F254).The TLC systems used were TLC-I, chloroform:methanol (98:2), and TLC-VI, chloroform:methanol(95:5). Twenty micrograms of the following “cold”standard steroids were added to the steroid fractionsbefore chromatography on TLC-VI: androstenedione,17-hydroxyprogesterone, testosterone, 17,20a-dihydroxy-4-pregnen-3-one, 17,20b-dihydroxy-4-pregnen-3-one (17,20bP), 11b-hydroxytestosterone,11-ketotestosterone, and 5a-dihydrotestosterone.Standards were detected by UV absorption at 254
nm, or by iodine vapor, and radioactive peaks weredetected using a Packard Model 7220 TLC scanner andeluted with 5 ml methanol and rechromatographed inTLC-J.
Microchemical Reactions and Identification
Microchemical reactions were carried out as previ-ously described (Kime, 1990). Presumptive identifica-tion, in accord with the criteria of Sandor and Idler(1972), involved identity of chromatographic proper-ties of the steroid and at least one chemical derivative
Androgens in Lungfish 257
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
JOBNAME: GCE 101#3 96 PAGE: 3 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
and crystallization to constant specific activity of thesteroid with authentic standard (Table 1). For crystal-lization, the following solvent combinations wereused: acetone–water, methanol–water, ethanol–water,and acetone.(i) 17-Hydroxyprogesterone—isopolar with authen-
tic steroid in TLC-I and TLC-VI; oxidized by chro-mium trioxide to a product isopolar with 4-andro-stene-3,17-dione in TLC-VI and reduced with 20b-hydroxysteroid dehydrogenase to a product isopolarwith 17,20bP in TLC-VI; crystallized to constant spe-cific activity with authentic material.Identification criteria for individual steroids were
the following:(ii) Testosterone—isopolar with authentic steroid in
TLC-I and TLC-VI; oxidized by chromium trioxide toa product isopolar with 4-androstene-3,17-dione inTLC-VI and acetylated to a product isopolar with tes-tosterone acetate in TLC-I; crystallized to constant spe-cific activity with authentic testosterone.(iii) 4-Androstene-3,17-dione, following incubation
with testosterone—isopolar with authentic steroid inTLC-I and TLC-VI; reduced by sodium borohydride at−20° to a product isopolar with testosterone in TLC-VI; crystallized to constant specific activity with au-thentic androstenedione.
Radioimmunoassay
A highly specific direct radioimmunoassay kit (Far-mos Diagnostica) for testosterone was validated for
lungfish serum by measuring samples that had beenserially diluted with steroid-free lungfish serum. Stan-dards (0.2–50 ng/ml) were prepared in steroid-freelungfish serum, using reference standard testosteronepurchased from Sigma Chemical Co. The serially di-luted lungfish serum samples all fell on, or very closeto, the standard curve. The cross-reactions at the 50%displacement level for the testosterone antiserumweregiven by the manufacturer as 5a-dihydrotestosterone,10%; 5b-dihydrotestosterone, 15%; 5a-androstane-3b,17b-diol, 5%; other steroids (epitestosterone, 4-an-drostene-3,17-dione, 5-androstene-3a,17b-diol, estra-diol-17b, androsterone, dehydroepiandrosterone, andcortisol), <1.0%.In addition to the change in standards, assays were
modified from the procedure suggested by the kitmanufacturer as follows: As the sex of each fish wasunknown prior to assay, serum samples assayed fortestosterone were 25 ml (as suggested in the kit) and 5ml made up to 25 ml with steroid-free lungfish serum,i.e., each sample was assayed in duplicate at two con-centrations. Incubation between antiserum and anti-gen was carried out at 4° overnight rather than at RTfor 2 hr. The longer incubation time decreased theintra-assay variability, calculated from the samesample placed at the beginning, middle, and end ofeach assay, to <5%.
Histology of Testis
Over the period of study, whenever a lungfish waskilled in order to collect tissues for study, if it was a
TABLE 1
Identification of Metabolites by Crystallization to Constant Specific Activity
Specific activity (dpm/mg)/crystallization
Original 1st 2nd 3rd 4th
From [3H]progesteroneProgesterone 7896 7282 7496 7550 728817-Hydroxyprogesterone 3214 2872 2520 2440 2471Androstenedione 10,497 7323 6145 5031 4162Testosterone 3471 2767 2666 2678 2614
From [3H]17-hydroxyprogesteroneAndrostenedione 8302 8204 7701 6605 7104
Following glucuronidase digestionAndrostenedione 675 650 638 639 583
From [3H]testosteroneTestosterone 25,476 26,430 22,739 23,461 22,224Androstenedione 3711 3131 2932 2764 2831
Following glucuronidase digestionTestosterone 760 712 714 722 664
Joss, Edwards, and Kime258
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

JOBNAME: GCE 101#3 96 PAGE: 4 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
male, samples of testis (from several regions, e.g.,fore-, mid-, and hindtestis) and kidney (also represent-ing fore-, mid-, and hindkidney) were placed inBouins fixative. These tissues were routinely embed-ded in wax, sectioned, and stained with hematoxylinand eosin in order to determine the stage of spermato-genesis.
RESULTS
Testicular Androgens
For the three lungfish testes incubated with [3H]pro-gesterone, chromatography in TLC-VI gave four peaksof radioactive material. The peak closest to the solventfront (peak 1) contained an average of 75% of the ra-dioactivity and coeluted with the androstenedionestandard; peak 2 contained 6% and coeluted with the17-hydroxyprogesterone standard; peak 3 contained18% and coeluted with the testosterone standard; andpeak 4, which encompassed the origin, contained 1%of the radioactivity. This latter peak was excludedfrom further analyses. There was no radioactivity inthe areas of the TLC corresponding to 11b-hydroxyand 11-ketotestosterone (area between peaks 3 and 4).The material in peak 1, when rechromatographed inTLC-I partially separated into two overlapping peaks(peak furthest from solvent front—peak 29—wasmuch larger than peak 19) which resisted any furtherattempts to achieve complete separation. It was as-sumed from the percentage of radioactivity containedwithin this peak that it represented the original sub-strate, progesterone (peak 29 in TLC-I), with some me-tabolite, androstenedione (peak 19 in TLC-I). Peak 29
crystallized with authentic progesterone to constantspecific activity (Table 1) but peak 19 did not crystal-lize to constant specific activity with authentic andro-stenedione. Thus, androstenedione was not identifiedpositively as a metabolite of progesterone in lungfishtestis. Both 17-hydroxyprogesterone and testosterone,however, were identified positively as metabolitesboth by microchemical analyses, as given in the meth-ods, and by crystallizations (Table 1). When [3H]17-hydroxyprogesterone was used as substrate instead ofprogesterone with the August lungfish testis, the peak
corresponding to androstenedione did not separateinto two peaks on further chromatography and waspositively identified as androstenedione both by mi-crochemical analyses and by crystallization with au-thentic steroid (Table 1).Incubation with [3H]testosterone for all four lung-
fish testes gave three peaks of radioactivity whenchromatographed in TLC-VI. The peak closest to thesolvent front coeluted with androstenedione standard,was reduced by sodium borohydride to a substancethat coeluted with testosterone in TLC-VI, and con-tained 3% of the radioactivity. The second peak con-tained 90% of the radioactivity, coeluted with testos-terone standard in TLC-VI, and was identified posi-tively as testosterone by microchemical reactions andcrystallization to constant specific activity with au-thentic testosterone. The third peak overlapped theorigin and contained 7% of the radioactivity. This lat-ter peak did not coelute with any of the standard ste-roid metabolites and remains unidentified. There wasno activity in the TLC areas expected for 11b-hydroxyand ketotestosterone or 5a- and 5b-dihydrotestoster-one.Following b-glucuronidase digestion of the aqueous
fractions and subsequent extraction of free steroidsfrom both the [3H]testosterone and [3H]17-OH-progesterone incubation of testis from the Augustlungfish, two peaks were obtained when chromato-graphed in TLC-IV. These corresponded to andro-stenedione (4 and 40%) and testosterone (80 and 25%from T and OH-P, respectively).
Plasma Testosterone
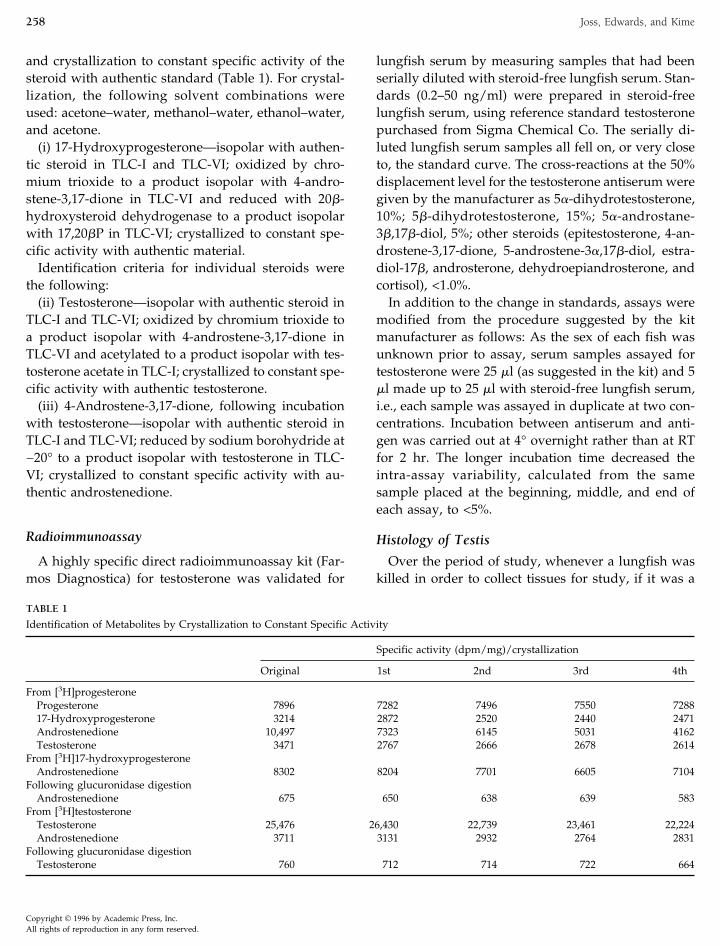
Plasma testosterone levels were measured in bloodsamples taken from wild-caught lungfish in January1985 (nine fish), 1987 (two fish), March 1986 (ninefish), April 1991 (six fish), May 1985 (six fish), 1986(two fish), September 1984 (seven fish), 1986 (fivefish), 1988 (six fish), October 1987 (three fish), 1989(eight fish), and November 1990 (five fish). A seasonalprofile of changes in plasma testosterone in adult malelungfish is shown in Fig. 1. Immature male lungfish,regardless of the time of year, had plasma testosteronelevels ranging from 1.2 to 3.7 ng/ml and adult femalelungfish, from 0.2 to 1.35 ng/ml.
Androgens in Lungfish 259
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
JOBNAME: GCE 101#3 96 PAGE: 5 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
Testis Histology
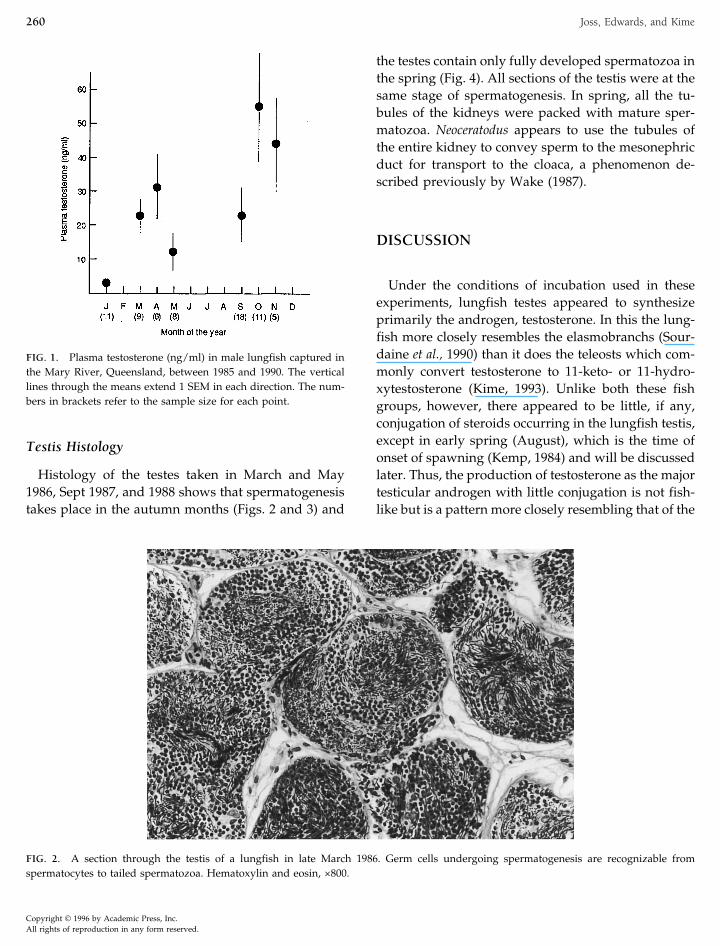
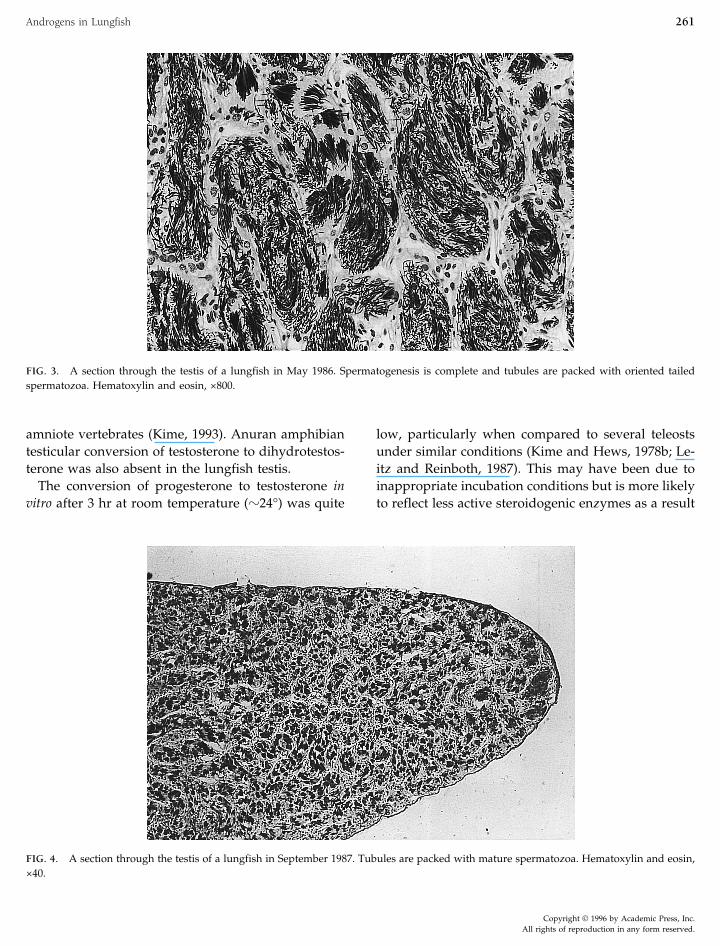
Histology of the testes taken in March and May1986, Sept 1987, and 1988 shows that spermatogenesistakes place in the autumn months (Figs. 2 and 3) and
the testes contain only fully developed spermatozoa inthe spring (Fig. 4). All sections of the testis were at thesame stage of spermatogenesis. In spring, all the tu-bules of the kidneys were packed with mature sper-matozoa. Neoceratodus appears to use the tubules ofthe entire kidney to convey sperm to the mesonephricduct for transport to the cloaca, a phenomenon de-scribed previously by Wake (1987).
DISCUSSION
Under the conditions of incubation used in theseexperiments, lungfish testes appeared to synthesizeprimarily the androgen, testosterone. In this the lung-fish more closely resembles the elasmobranchs (Sour-daine et al., 1990) than it does the teleosts which com-monly convert testosterone to 11-keto- or 11-hydro-xytestosterone (Kime, 1993). Unlike both these fishgroups, however, there appeared to be little, if any,conjugation of steroids occurring in the lungfish testis,except in early spring (August), which is the time ofonset of spawning (Kemp, 1984) and will be discussedlater. Thus, the production of testosterone as the majortesticular androgen with little conjugation is not fish-like but is a pattern more closely resembling that of the
FIG. 1. Plasma testosterone (ng/ml) in male lungfish captured inthe Mary River, Queensland, between 1985 and 1990. The verticallines through the means extend 1 SEM in each direction. The num-bers in brackets refer to the sample size for each point.
FIG. 2. A section through the testis of a lungfish in late March 1986. Germ cells undergoing spermatogenesis are recognizable fromspermatocytes to tailed spermatozoa. Hematoxylin and eosin, ×800.
Joss, Edwards, and Kime260
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
JOBNAME: GCE 101#3 96 PAGE: 6 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
amniote vertebrates (Kime, 1993). Anuran amphibiantesticular conversion of testosterone to dihydrotestos-terone was also absent in the lungfish testis.The conversion of progesterone to testosterone in
vitro after 3 hr at room temperature (z24°) was quite
low, particularly when compared to several teleostsunder similar conditions (Kime and Hews, 1978b; Le-itz and Reinboth, 1987). This may have been due toinappropriate incubation conditions but is more likelyto reflect less active steroidogenic enzymes as a result
FIG. 4. A section through the testis of a lungfish in September 1987. Tubules are packed with mature spermatozoa. Hematoxylin and eosin,×40.
FIG. 3. A section through the testis of a lungfish in May 1986. Spermatogenesis is complete and tubules are packed with oriented tailedspermatozoa. Hematoxylin and eosin, ×800.
Androgens in Lungfish 261
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

JOBNAME: GCE 101#3 96 PAGE: 7 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
of a preference for the delta-5 pathway. An earlierstudy (Joss and Borchers, unpublished observations)using [3H]pregnenolone as substrate and similar incu-bation conditions found by tentative identificationonly (i.e., coelution with standard steroids in morethan one chromatography system) that 80% of the sub-strate had been converted to dehyroepiandrosterone,androstenedione, and testosterone in 2 hr. The possi-bility of a preference for the delta-5 pathway in lung-fish testis warrants further verification by comparingthe conversion to delta-4 and delta-5 metabolites in apulse chase experiment involving incubation with tri-tiated cholesterol.Significant conjugation of androgens in the testis at
the beginning of the spawning season strongly sug-gests that these glucuronides may be playing a phero-monal role at this time. Several studies on teleostshave found that conjugated steroids can act as phero-mones when released into the water with milt or urine(e.g., Scott and Vermierssen, 1994; Sherwood et al.,1991; Schoonen and Lambert, 1986; Stacey and Sor-rensen, 1986). Another possible function is that ofparacrine intratesticular regulation as has been pro-posed by Cuevus et al. (1992) for conjugated andro-gens in dogfish testis. We cannot at present distin-guish between these two or any other possible func-tions for seasonal testicular androgen conjugation inlungfish.The plasma levels of testosterone are quite high dur-
ing March–April and again in September–November,if compared with those of mammals and birds whichhave plasma levels around 0.1–20 ng/ml. High levelsof circulating testosterone are, however, quite com-mon for non-mammalian/avian vertebrates, many ofwhich have much higher recorded values than thosefound in lungfish (Norris, 1985). The seasonal patternof circulating testosterone correlates with spermato-genesis in the testes. Although no male lungfish werekilled in January to provide histological investigationof the testes at that time, the comparatively low levelsof circulating testosterone in male fish in January(mid-summer), taken together with the early stages ofactive spermatogenesis seen in late March, would sug-gest that the testes are nonspermatogenic during thesummer months. Environmental conditions where thelungfish live are quite severe in the summer, with airtemperatures generally exceeding 40° and correspond-ing water temperatures in the low to mid-30°s. Such
anoxic conditions require the lungfish to use theirlungs to obtain oxygen which, in this species, is not thepreferred mode of respiration, their gills being muchbetter developed than in either the African or theSouth American lungfish (Burggren and Johansen,1986). It is, therefore, not surprising that they are re-productively quiescent in the summer. However, asthe temperatures cool in the autumn, the circulatingtestosterone levels rise and the testes become veryspermatogenically active. By May, the month preced-ing winter, all of the spermatogenic tubules within thetestes contain tailed spermatozoa. At this time, thetestosterone levels in the blood drop. They rise againin the spring, when release of the ripe sperm into thetubules of the kidneys occurs prior to mating. Thispattern of summer quiescence, autumn spermatogen-esis, overwintering of mature sperm, and spring mat-ing is fairly typical of anamniote vertebrates living intemperate to subtropical climates (Lofts, 1987) and is,therefore, not unexpected for lungfish.
ACKNOWLEDGMENTS
This research was supported by an ARC grant to J.M.P.J. We alsoexpress our appreciation to the British Council which generouslycontributed to the travel costs of J.M.P.J. under their AcademicLinks and Interchange Scheme. The cheerful donation of manyschool holidays to lungfish collecting trips by Greg, Michael, andTom Joss is gratefully acknowledged.
REFERENCES
Burggren, W. W., and Johansen, K. (1986). Circulation and respira-tion in lungfishes (Dipnoi). In “The Biology and Evolution ofLungfishes” (W. E. Bemis, W. W. Burggren, and N. E. Kemp,Eds.), pp. 217–236. A. R. Liss, New York.
Cuevas, M. E., Millar, W., and Callard, G. (1992). Sulfo-conjugationof steroids and the vascular pathway of communication in dog-fish testis. J. Exp. Zool. 264, 119–129.
Forey, P. L. (1986). Relationships of lungfishes. In “The Biology andEvolution of Lungfishes” (W. E. Bemis, W. W. Burggren, andN. E. Kemp, Eds.), pp. 75–92. A. R. Liss, New York.
Kemp, A. (1984). Spawning of the Australian lungfish, Neoceratodusforsteri (Kreft) in the Brisbane River and in Enoggera Reservoir,Queensland. Mem. Queensl. Mus. 21, 391–399.
Kime, D. E. (1987). The steroids. In “Fundamentals of Comparative
Joss, Edwards, and Kime262
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

JOBNAME: GCE 101#3 96 PAGE: 8 SESS: 21 OUTPUT: Sat Apr 27 09:03:03 1996/xypage/worksmart/tsp000/68045k/22pu
Vertebrate Endocrinology” (I. Chester-Jones, P. M. Ingleton, andJ. G. Phillips, Eds.), pp. 3–56. Plenum, New York/London.
Kime, D. E. (1990). In vitro metabolism of progesterone, 17-hydroxyprogesterone, and 17,20b-dihydroxy-4-pregnen-3-one byovaries of the common carp, Cyprinus carpio: Production rates ofpolar metabolites. Gen. Comp. Endocrinol. 79, 406–414.
Kime, D. E. (1993). “Classical” and “non-classical” reproductive ste-roids in fish. Rev. Fish Biol. Fish. 3, 160–180.
Kime, D. E., and Hews, E. A. (1978a). Androgen biosynthesis in vitroby testes from Amphibia. Gen. Comp. Endocrinol. 35, 280–288.
Kime, D. E., and Hews, E. A. (1978b). In vitro biosynthesis of 11b-hydroxy- and 11-oxotestosterone by the testes of the pike (Esoxlucias) and the perch (Perca fluviatilis). Gen. Comp. Endocrinol. 36,604–608.
Leitz, T., and Reinboth, R. (1987). The biosynthesis of 11-ketotestosterone by the testis of the Siamese fighting fish Bettasplendens Regan (Anabantoidei, Belontiidae). Gen. Comp. Endocri-nol. 66, 145–157.
Lofts, B. (1987). Testicular function. In “Hormones and Reproduc-tion in Fishes, Amphibians, and Reptiles” (D. O. Norris and R. E.Jones, Eds.), pp. 283–326. Plenum, New York/London.
Meyer, A. (1995). Molecular evidence on the origin of tetrapods andthe relationships of the coelacanth. Trends Ecol. Evol. 10, 111–116.
Muller, C. H. (1977). Plasma 5a-dihydrotestosterone and testoster-one in the bullfrog, Rana catesbeiana: Stimulation by bullfrog L.H.Gen. Comp. Endocrinol. 33, 122–132.
Norris, D. O. (1985). “Vertebrate Endocrinology.” Lea and Febiger,Philadelphia.
Sandor, T., and Idler, D. R. (1972). Steroid methodology. In “Steroidsin Nonmammalian Vertebrates” (D. R. Idler, Ed.), pp. 6–36. Aca-demic Press, New York/London.
Schoonen, W. E. G. J., and Lambert, J. G. D. (1986). Steroid metabo-
lism in the testes of the African catfish, Clarias gariepinus(Burchell), during spawning season, under natural conditionsand kept in ponds. Gen. Comp. Endocrinol. 64, 40–52.
Scott, A. P., and Vermierssen, E. L. M. (1994). Production of conju-gated steroids by teleost gonads and their role as pheromones. In“Perspectives in Comparative Endocrinology” (K. D. Davey, R. E.Peter, and S. S. Tobe, Eds.), pp. 645–654. National Research Coun-cil of Canada, Ottawa.
Sherwood, N. M., Kyle, A. L., Kreiberg, H., Warby, C. M., Magnus,T. H., Carolsfield, J., and Price, W. S. (1991). Partial characterisa-tion of a spawning pheromone in the herring Clupea harenguspallasi. Can. J. Zool. 69, 91–103.
Sourdaine, P., Gamier, D. H., and Jegou, B. (1990). The adult dogfish(Scyliorhinus canicula L.) testis: A model to study stage-dependentchanges in steroid levels during spermatogenesis. J. Endocrinol.127, 451–460.
Stacey, N. E., and Sorensen, P. W. (1986). 17a,20b-dihydroxy-4-pregnen-3-one: A steroidal primer pheromone increasing miltvolume in the goldfish, Carassius auratus. Can. J. Zool. 64, 2412–2417.
Wake, M. H. (1987). Urogenital morphology of dipnoans, with com-parisons to other fishes and to amphibians. J. Morphol. Suppl. 1,199–216.
Wilson, J. D. (1975). Metabolism of testicular androgens. In “Hand-book of Physiology,” Sect. 7, “Endocrinology,” Vol. V, “Male Re-productive System” (R. O. Greep and E. B. Astwood, Eds.), pp.491–508. Am. Physiol. Soc., Washington, DC.
Yokobori, S., Hasegawa, M., Ueda, T., Okada, N., Nishikawa, K.,and Watanabe, K. (1994). Relationship among coelacanths, lung-fishes, and tetrapods: A phylogenetic analysis based on mito-chondrial cytochrome oxidase I gene sequences. J. Mol. Evol. 38,602–609.
Androgens in Lungfish 263
Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.




















