
Impacts of trichomonosis epidemics on Greenfinch Chloris chloris and Chaffinch Fringilla coelebs...
10
Impacts of trichomonosis epidemics on Greenfinch Chloris chloris and Chaffinch Fringilla coelebs populations in Finland ALEKSI LEHIKOINEN, 1 * ESA LEHIKOINEN, 2 JARI VALKAMA, 1 RISTO A. V € AIS € ANEN 1 & MARJA ISOMURSU 3 1 Finnish Museum of Natural History, Zoology, University of Helsinki, Helsinki, Finland 2 Section of Ecology, University of Turku, Turku, Finland 3 Finnish Food Safety Authority, Production Animal and Wildlife Health Research Unit, Oulu, Finland Population consequences of parasites in wild birds are rarely documented. One exception is the decline of British finch populations due to an epidemic caused by the protozoan parasite Trichomonas gallinae. Finch trichomonosis has recently spread from the UK to northern Europe, but its consequences for local finch populations have not been studied. We assessed the extent to which the trichomonosis epidemic affected the Finnish popu- lation sizes of European Greenfinch Chloris chloris, Common Chaffinch Fringilla coelebs and a control species, Great Tit Parus major, and the body condition of Greenfinches. The disease was first documented in Finland in 2008 and epidemics were observed mainly in southwestern Finland. Greenfinches showed a significant decline of 47% in breeding numbers and 65% in wintering numbers in southern Finland during 2006– 2010. Breeding Chaffinch numbers showed a slight decline (4%) during the same study period that was significant only in central Finland. Great Tit did not show a significant change in breeding numbers. During the initial disease epidemic the body condition of all demographic groups of Greenfinches decreased equally, which suggests that the dis- ease was not selective in respect of age or sex. There were no encounters of Finnish- ringed Greenfinches or Chaffinches in the UK, which could indicate that the parasite has not necessarily been transferred directly from the UK, but perhaps by migrants from Sweden and Germany. Keywords: disease, European finches, mortality, parasites, population fluctuation, population monitoring, Trichmonas gallinae. Diseases and parasites are common in wild animals, but their effects at the population level are poorly known (Newton 1998). Sporadic mass mortality caused by diseases has occasionally been docu- mented, especially in waterfowl and other water- birds (e.g. avian cholera, botulism and avian influenza; Newton 1998, Liu et al. 2005). In Finland, a mortality event in Common Eiders Soma- teria mollissima in the Baltic Sea was attributed at least partly to adenovirus infection (Hollm en et al. 2003). Epidemics have been much more rarely documented in terrestrial bird species (Dhondt et al. 1998, Newton 1998). However, in Red Grouse Lagopus lagopus scoticus managed at high population densities for shooting, population fluctuations are known to be partly regulated by parasites (Redpath et al. 2006). Trichomonosis is a disease caused by the flagel- late protozoan parasite Trichomonas gallinae, which is well known historically in pigeons and raptors (Villanua et al. 2006, Stromberg et al. 2008, San- sano-Maestre et al. 2009) and can cause high mor- tality (Haugen 1952, Hofle et al. 2004). High rates of infection have been recorded in the Rock Pigeon Columba livia, which can also act as an aclinical carrier and reservoir of T. gallinae (Atkin- son et al. 2008). The parasite inhabits the upper digestive tract of the bird and spreads in saliva. The consequent disease in columbids is typically a *Corresponding author. Email: aleksi.lehikoinen@helsinki.fi © 2013 British Ornithologists’ Union Ibis (2013), 155, 357–366
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Impacts of trichomonosis epidemics on Greenfinch Chloris chloris and Chaffinch Fringilla coelebs...

Impacts of trichomonosis epidemics on GreenfinchChloris chloris and Chaffinch Fringilla coelebs
populations in FinlandALEKSI LEHIKOINEN,1* ESA LEHIKOINEN,2 JARI VALKAMA,1 RISTO A. V€AIS€ANEN1 & MARJA ISOMURSU3
1Finnish Museum of Natural History, Zoology, University of Helsinki, Helsinki, Finland2Section of Ecology, University of Turku, Turku, Finland
3Finnish Food Safety Authority, Production Animal and Wildlife Health Research Unit, Oulu, Finland
Population consequences of parasites in wild birds are rarely documented. One exceptionis the decline of British finch populations due to an epidemic caused by the protozoanparasite Trichomonas gallinae. Finch trichomonosis has recently spread from the UK tonorthern Europe, but its consequences for local finch populations have not been studied.We assessed the extent to which the trichomonosis epidemic affected the Finnish popu-lation sizes of European Greenfinch Chloris chloris, Common Chaffinch Fringilla coelebsand a control species, Great Tit Parus major, and the body condition of Greenfinches.The disease was first documented in Finland in 2008 and epidemics were observedmainly in southwestern Finland. Greenfinches showed a significant decline of 47% inbreeding numbers and 65% in wintering numbers in southern Finland during 2006–2010. Breeding Chaffinch numbers showed a slight decline (4%) during the same studyperiod that was significant only in central Finland. Great Tit did not show a significantchange in breeding numbers. During the initial disease epidemic the body condition ofall demographic groups of Greenfinches decreased equally, which suggests that the dis-ease was not selective in respect of age or sex. There were no encounters of Finnish-ringed Greenfinches or Chaffinches in the UK, which could indicate that the parasite hasnot necessarily been transferred directly from the UK, but perhaps by migrants fromSweden and Germany.
Keywords: disease, European finches, mortality, parasites, population fluctuation, populationmonitoring, Trichmonas gallinae.
Diseases and parasites are common in wild animals,but their effects at the population level are poorlyknown (Newton 1998). Sporadic mass mortalitycaused by diseases has occasionally been docu-mented, especially in waterfowl and other water-birds (e.g. avian cholera, botulism and avianinfluenza; Newton 1998, Liu et al. 2005). InFinland, a mortality event in Common Eiders Soma-teria mollissima in the Baltic Sea was attributed atleast partly to adenovirus infection (Hollm�en et al.2003). Epidemics have been much more rarelydocumented in terrestrial bird species (Dhondtet al. 1998, Newton 1998). However, in Red
Grouse Lagopus lagopus scoticus managed at highpopulation densities for shooting, populationfluctuations are known to be partly regulated byparasites (Redpath et al. 2006).
Trichomonosis is a disease caused by the flagel-late protozoan parasite Trichomonas gallinae, whichis well known historically in pigeons and raptors(Villanua et al. 2006, Stromberg et al. 2008, San-sano-Maestre et al. 2009) and can cause high mor-tality (Haugen 1952, Hofle et al. 2004). High ratesof infection have been recorded in the RockPigeon Columba livia, which can also act as anaclinical carrier and reservoir of T. gallinae (Atkin-son et al. 2008). The parasite inhabits the upperdigestive tract of the bird and spreads in saliva.The consequent disease in columbids is typically a
*Corresponding author.Email: [email protected]
© 2013 British Ornithologists’ Union
Ibis (2013), 155, 357–366

necrotic inflammation of the oropharynx thateventually inhibits swallowing and leads to starva-tion and dehydration (Atkinson et al. 2008). Inpasserine birds, necrotic lesions are most pro-nounced in the oesophagus and crop (Pennycottet al. 2005, Neimanis et al. 2010).
Trichomonosis has been identified as a risk forsome endangered bird populations such as Bonel-li’s Eagle Aquila fasciata in the Pyrenees (in whichit is the most important mortality factor for chicks;Real et al. 2000) and Mauritian Pink Pigeon Neseo-enas mayeri (Bunbury et al. 2007, 2008). Tricho-monosis has been found sporadically in RockPigeons in Finland: four cases were recorded in2001–2005, all in the city of Oulu (Archives ofthe Finnish Food Safety Authority Evira). In recentyears, the disease has emerged in passerine birds,particularly finches, across Europe (Pennycott et al.2005, Neimanis et al. 2010) and North America(Forz�an et al. 2010).
In 2005, an epidemic of trichomonosis in pas-serine birds was observed in the UK, especially inEuropean Greenfinches Chloris chloris and Com-mon Chaffinches Fringilla coelebs (Robinson et al.2010). Epidemics in finches were then observed inFennoscandia in 2008 (Neimanis et al. 2010) andin northern Germany in 2009 (Peters et al. 2009),which suggests that the disease may have spreadquickly over a wide area. Finch trichomonosisreached southern Finland in 2008, and as in othercountries the main species affected was the Green-finch (83% of the 17 diagnosed individuals, two ofwhich were confirmed by PCR; Neimanis et al.2010), with epidemics coinciding with the latebreeding and early migration periods. Lawson et al.(2011) showed that the PCR sequences of parasiteisolates from the UK and Fennoscandia were iden-tical and presented evidence of the disease spread-ing from the UK to Fennoscandia in 2008 throughmigrating Chaffinches.
Our aim is to document how a new emergingdisease, finch trichomonosis, affected the largeFinnish Greenfinch and Chaffinch populationsusing census data from both breeding and non-breeding seasons.
Finnish Greenfinches are partial migrants (e.g.Lehikoinen et al. 2008) and are common birdsat garden bird feeders (V€ais€anen 2008). TheGreenfinch population has increased greatly dur-ing recent decades, being six times higher in2005 than in 1983 (V€ais€anen 2006) and consist-ing of approximately 370 000 pairs before the
epidemic (R.A. V€ais€anen unpubl. data). There isa strong geographical gradient in Greenfinchbreeding density in Finland, with the highestdensities and occupancy probabilities in thesouthwest of the country, and much lower den-sities in the north and east (V€ais€anen et al.1998, Valkama et al. 2011).
The Chaffinch is a migratory species, and onlyvery small numbers winter in the southern part ofthe country. The species also visits feeding stationsduring the spring migration, but numbers are smallcompared with the very large breeding populationsize (Lehikoinen & V€ah€atalo 2000, V€ais€anen 2008,Valkama et al. 2011). The Chaffinch is the secondmost numerous breeding bird species in Finland(7–9 million pairs), and its population has beenstable in recent decades (Valkama et al. 2011).
In addition, we used data for Great Tit Parusmajor, which is a partial migrant and, like Green-finch, an abundant species at garden bird feeders(Nowakowski & V€ah€atalo 2003, V€ais€anen 2008),as a control to rule out other external factors. Thepopulation estimate of Great Tit is 1.5–2.0 millionpairs (V€ais€anen et al. 2011). Based on currentknowledge, trichomonosis has been found in GreatTits only rarely (Lawson et al. 2011) and never inFinland (see Results).
Because of the strong gradient in finch popula-tion densities and previous evidence that the dis-ease has spread to the country from the southwest(Lawson et al. 2011), we examined populationchange in three different geographical areas. Inaddition, we assessed whether differences betweenyears in the body condition of trapped birds couldbe related to the epidemic.
We also document the wintering sites of FinnishGreenfinches and Chaffinches based on ringingdata in order to help understand whether directspread from the UK by wintering Finnish finchesseems most plausible or whether the disease wasmore likely first transferred to Scandinavia andthen to Finland by migrants from the southwest.
METHODS
Diagnosing parasitized birds
Passive surveillance of wildlife diseases (currentlyby the Finnish Food Safety Authority Evira) hasbeen undertaken in Finland for decades. Any citi-zen as well as public health officials can deliverdead wild animals for post-mortem examination
© 2013 British Ornithologists’ Union
358 A. Lehikoinen et al.

free of charge. The specimens included in thisstudy were collected through this system.
Surveillance data have been recorded in a digitaldatabase since 2000. Between 2000 and 2011,3206 wild birds (on average 267 per year) havebeen examined. Wild birds are examined in thecity of Oulu, in central Finland. Submissions havecome from all provinces of the country (16 intotal). In 2006–2011, c. 39% of samples were fromsouthern Finland, 51% from central Finland and10% from northern Finland (regional boundariesshown in Fig. 1).
The most common small passerine species inthe database was the Great Tit (112 individuals).The Greenfinch was the second most common,with 104 individuals. Before the emergence offinch trichomonosis in September 2008, 18 Green-finches had been examined, and 56% of thesewere cases of salmonellosis occurring between Jan-uary and April. Trauma was the second most com-mon diagnosis (22% of cases). Only fiveChaffinches were examined during 2000–2011,three of which were cases of trauma and enteritis.One was reported at the beginning of the tricho-monosis epidemic in 2008 but could not be exam-ined due to post-mortem autolysis.
Birds were usually examined within 24–72 hafter death. A routine necropsy was performed,including bacteriology and, when the specimenwas fresh enough, histology. Direct wet mountsfrom the crop or mouth were also examined undera light microscope (4009 magnification) for thepresence of motile flagellate protozoans with anappearance consistent with trichomonads. Samplesswabbed or cut from the inner wall of the crop ormouth were cultured in Trichomonas media no. 2(Oxoid Ltd, Cambridge, UK) and incubated at37 °C for up to 7 days. The culture was firstchecked after 2–3 days of incubation and, if neces-sary, again after 5–7 days under a light microscopeas with the wet mounts.
A case of trichomonosis was diagnosed whentypical macroscopic and/or histological lesions(necrotic ingluvitis, pharyngitis and/or stomatitis)were observed and trichomonads were found inculture or a direct wet mount (40 cases). Tricho-monas gallinae is a labile parasite and dies rela-tively soon after host death (Erwin et al. 2000).We therefore diagnosed trichomonosis without thefinding of live parasites if typical pathologicallesions were present in the oesophagus and/ororopharynx and no Salmonella infection, a com-mon cause of ingluvitis in passerines (Thomaset al. 2007), was found in bacterial culture (42cases). Birds sent by one person from a single sitein one year were considered a single incident oftrichomonosis.
Telephone reports of Greenfinch mortality wererecorded in the summer (June–August) of 2009. Atrichomonosis incident was defined as a locally andtemporally limited mortality event of severalGreenfinches with observations of typical symp-toms (lethargy, difficulties in swallowing andbreathing, unclean beak) in sick birds.
Breeding season data
Annual population indices for Finnish breedingbirds were calculated from census areas (includingline transects, point census routes and territorymaps) that had been repeated in at least twosummers. The census period was June and thebreeding pair was the census unit (Koskimies &V€ais€anen 1991). We took 1984 as the starting yearbecause since then all three study species have hadpopulation trends estimated from more than 100pairs per year. Mean annual sample sizes were720, 4116 and 268 pairs for Great Tit, Chaffinch
Figure 1. Map of Finland showing the areas of South, Central(between 62°10′ and 65°45′N) and North Finland used in theanalyses. The black dot shows the long-term standardizedtrapping site of Turku, where the body condition data werecollected.
© 2013 British Ornithologists’ Union
Trichomonas-epidemic and Finnish Greenfinches 359
and Greenfinch, respectively. We also divided thedata into three geographical areas during pre-epidemic (1984–2005) and epidemic years (2006–2010): South, Central and North Finland (Fig. 1,Table 1). Mean annual sample sizes for these threeareas were 1025, 505 and 65 pairs for Great Tit,4336, 3564 and 327 pairs for Chaffinch, 447, 166and 23 pairs for Greenfinch, respectively. We usedlog-linear Poisson regression (program TRIM, Panne-koek & van Strien 2004) for estimating annualpopulation indices.
Non-breeding season data
Finnish winter bird censuses started in the winterof 1956/57. Since then, the annual abundance ofwintering bird populations has been monitoredalong a combined total of 3800 transect routes,the average length of which is about 10 km.Observers walk along the same routes each yearand count all birds (Koskimies & V€ais€anen 1991).About 500 routes have been counted in each win-ter during the last 50 years. The majority of thedata come from 330 routes that have beenrepeated at least 50 times. There are three censusperiods (early winter 1–15 November since 1975,mid-winter 25 December to 7 January since 1956,and late winter 21 February to 6 March since1967), but for simplicity we used only the mid-winter counts, which have the best survey effortamong the three census periods. As for the breed-ing season analysis, we used data for 1983/84–2010/2011, but considered only data for Great Tit(from 5150 to 94 100 birds per winter, median39 400) and Greenfinch (from 4940 to 37 710birds per winter, median 19 250) because data forChaffinch were too few (from 175 to 2300 birdsper winter, median 500). As in the breeding analy-ses, we used program TRIM to estimate annual
population indices and divided the data into thesame three geographical areas.
Ringing data
In total, 196 434 Greenfinches and 165 775 Chaf-finches were ringed in Finland during 1913–2010,of which 96% of the Greenfinches and 92% of theChaffinches were fully grown birds. There were23 065 encounters of Finnish-ringed Greenfinchesand 4331 of Chaffinches. Over 90% of these wererecaptures made by ringers, with most of theremainder being recovery reports of dead birdssent to the ringing centre by members of thepublic. Here we examined foreign recaptures andrecoveries since 1991 to assess wintering areas andmigration routes.
Body condition
The body mass of ringed birds has been measuredin Turku, southwest Finland (Fig. 1) since the win-ter of 1974/75. We used data from September andOctober because this fits the timing of trichomo-nosis outbreaks, based on observations duringregular trapping.
We used two approaches to model body massvariation in relation to the epidemics. First, wedetermined the body mass limit that separated the0.2% lowest weight full-grown birds in each sex-age category, across the whole dataset. From fieldobservations, this threshold distinguishes birds thatare unable to show rapid behavioural responsesand may show poor flight ability or other signs ofillness. This threshold was 24.11 g for adult males,23.77 g for first-year males, 23.09 g for adultfemales and 23.18 g for first-year females. Wethen assessed the proportion of birds at theseexceptionally low weights for each year from 1974
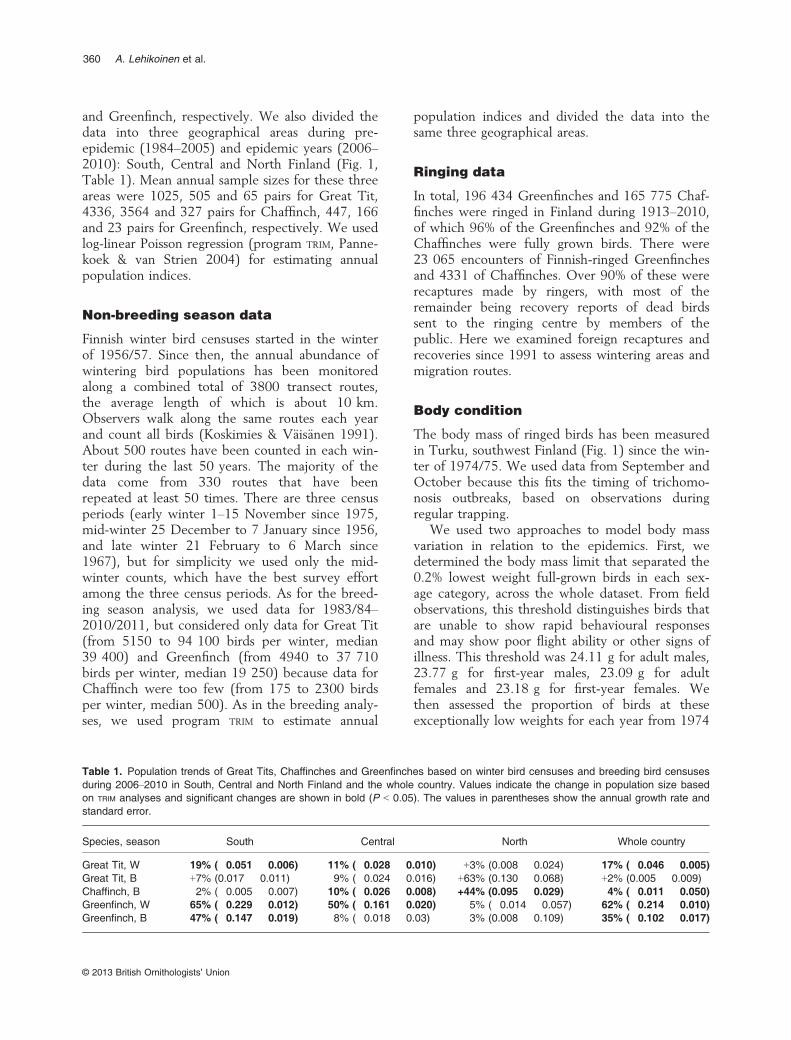
Table 1. Population trends of Great Tits, Chaffinches and Greenfinches based on winter bird censuses and breeding bird censusesduring 2006–2010 in South, Central and North Finland and the whole country. Values indicate the change in population size basedon TRIM analyses and significant changes are shown in bold (P < 0.05). The values in parentheses show the annual growth rate andstandard error.
Species, season South Central North Whole country
Great Tit, W �19% (�0.051 � 0.006) �11% (�0.028 � 0.010) +3% (0.008 � 0.024) �17% (�0.046 � 0.005)Great Tit, B +7% (0.017 � 0.011) �9% (�0.024 � 0.016) +63% (0.130 � 0.068) +2% (0.005 � 0.009)Chaffinch, B �2% (�0.005 � 0.007) �10% (�0.026 � 0.008) +44% (0.095 � 0.029) �4% (�0.011 � 0.050)Greenfinch, W �65% (�0.229 � 0.012) �50% (�0.161 � 0.020) �5% (�0.014 � 0.057) �62% (�0.214 � 0.010)Greenfinch, B �47% (�0.147 � 0.019) �8% (�0.018 � 0.03) �3% (0.008 � 0.109) �35% (�0.102 � 0.017)
© 2013 British Ornithologists’ Union
360 A. Lehikoinen et al.

to 2011. Secondly, we assessed how the body massof Greenfinches and Great Tits (data for Chaffinchwere too few) varied between years in differentage and sex classes during 2001–2011 (omitting2004, when the number of individuals measuredwas low in some age-sex categories). All birdstrapped for ringing were included in body massanalyses irrespective of symptoms of illness.
We used general linear models (PROC GLM, SAS
9.2) to estimate body mass variation in Septemberto October separately for each sex-age category, asa function of year, species and their interaction.
RESULTS
Diagnosed cases and reports oftrichomonosis
The first observations of unusual Greenfinch andChaffinch mortality in Finland were made on thesouthwest coast in August 2008 and reported bytelephone in September 2008 (Fig. 2). TwoGreenfinches and a Chaffinch with lesions consis-tent with trichomonosis were received for post-mortem examination in late September 2008. Thepeak of the epidemic in terms of the number ofdiagnosed cases per month was in July 2009, when16 cases from eight different municipalities wereconfirmed. In total, 82 trichomonosis cases (73Greenfinches, one Chaffinch, one Hawfinch Cocco-thraustes coccothraustes, one Eurasian Siskin Cardu-
elis spinus, one Blue Tit Cyanistes caeruleus, oneHouse Sparrow Passer domesticus, one Eurasian JayGarrulus glandarius, one Eurasian Magpie Pica picaand two Common Wood Pigeons Columba palum-bus) from 33 municipalities had been recorded byDecember 2011 (Fig. 2). The monthly distributionof incidents was: April = 2, June = 5, July = 10,August = 4, September = 5, October = 4, Novem-ber = 6 and December = 5. In addition, telephonereports of incidents were recorded in the summerof 2009 (from June to August). A total of 17 inci-dents from 17 different municipalities werereported, with five in June, eight in July and fourin August.
The samples and telephone reports revealed asteady spread of the disease, which appeared tocome to a halt in 2010, but continued again in thesummer of 2011, spreading from the southwest ofthe country towards the north and east (Fig. 2).
Breeding population trends
The breeding population of Greenfinches increasedfivefold during 1984–1999 and then remained sta-ble at a high level until 2009 (Fig. 3). The popula-tion fell by 47% between 2009 and 2010 in SouthFinland, with non-significant falls of 8 and 3% inCentral and North Finland, respectively. Acrossthe country as a whole, Greenfinch abundance fellto the level of the late 1990s (overall reduction35% during 2006–2010; Table 1), and had not
1 2
1 2
Greenfinch Other species
2008: 4/2 2009: 29/3 2010: 0/3 2011: 10/0
Figure 2. Finch trichomonosis incidents confirmed or reportedby the Finnish Food Safety Authority on Greenfinches (circles)and other species (triangles) in Finland in 2008–2011. Inci-dents from the same municipalities are combined annually.Annual numbers of incidents are shown after the year (Green-finches/other species). In 2010, the three incidents includedsamples of Common Wood Pigeon, House Sparrow andEurasian Siskin.
Figure 3. Annual breeding population indices of Great Tits inFinland from 1979 to 2011 (solid light grey line), Chaffinchesfrom 1975 to 2011 (dashed dark grey line) and Greenfinchesfrom 1984 to 2011 (solid black line). The mean index values ofyears 2006 and 2007 have been scaled to 100 in all species.In the Great Tit and Greenfinch, annual linear populationgrowth rates for the entire surveillance period were +2.1% and+6.3% (both species P < 0.001), respectively, but in theChaffinch there was no significant trend (–0.1%, P > 0.05).
© 2013 British Ornithologists’ Union
Trichomonas-epidemic and Finnish Greenfinches 361

experienced such a severe decline since the start ofthe study period in 1984 (Fig. 3). There was nofurther decline between 2010 and 2011 (Fig. 3).
Breeding numbers of Chaffinch remained rela-tively stable in Finland after 1984 (Fig. 3).Between 2006 and 2010, trends varied betweenregions, with no significant change in SouthFinland, but a 10% decline in Central Finland anda 44% increase in North Finland (Table 1). Acrossthe whole country, Chaffinches showed a signifi-cant 4% decline (Table 1).
Great Tit numbers almost doubled after 1984(Fig. 3) but showed no significant trends eithernationally or regionally between 2006 and 2010(Fig. 3, Table 1).
Non-breeding population changes
The wintering Greenfinch population declined by65% in South Finland and 50% in Central Finlandduring 2006–2010 (Table 1), especially between2008 and 2010 (Fig. 4). However, in NorthFinland the population fluctuated with no overalltrend (Table 1; Fig. 4a). Wintering Great Tit num-bers showed a slight but significant decline inSouth and Middle Finland (Table 1), and nochange in North Finland (Fig. 4b, Table 1).
Wintering and migration routes
In all, 116 Greenfinches ringed in Finland werefound outside Finland during 1991–2010. Encoun-ter sites were situated around the Baltic Sea, andthe majority of the winter encounters were insouthern Sweden and northern Germany (Fig. 5).The distribution of encounters also shows that atleast a few birds ringed in northern Finland over-winter in northern Sweden and Norway.
A total of 26 Chaffinches ringed in Finlandwere found outside Finland during 1991–2010.
Encounter sites were situated to the southwest ofFinland (Sweden, Germany, Belgium, France). Norecoveries of either species were from the UK(Figs 5 and 6).
Body condition
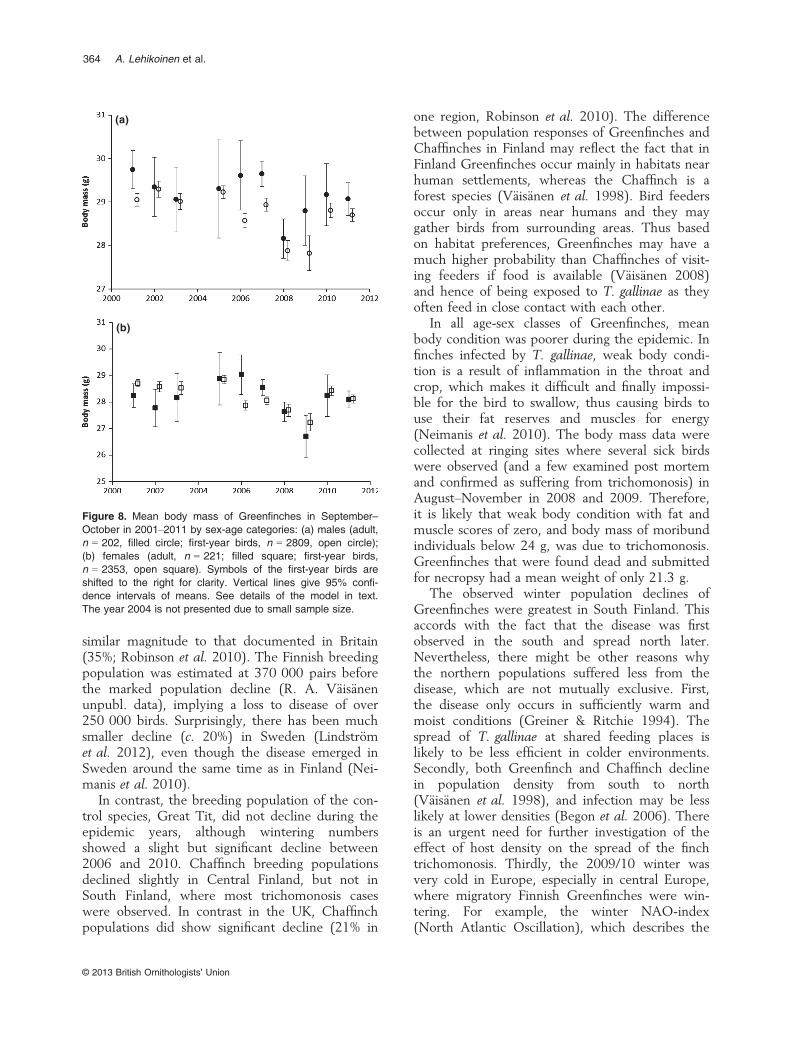
The annual proportion of lean Greenfinches overall age-sex classes was 0.3% (95% confidence limits0.0–1.1%) in 1975–1999 (n = 7283), but theirproportion increased to 2.9 and 3.6% in theautumns of 2008 and 2009 (2008: v21 = 206.1,P < 0.001, 2009: v21 = 101.1, P < 0.001; Fig. 7).We carried out posterior pairwise comparisons(corrected for multiple comparisons) between allpairs of years (2000–2011) in all sex-age categories(Fig. 8). In both first-year males and first-yearfemales, 22 significant differences were found in66 comparisons. These were significantly morelikely to occur in comparisons between an epi-demic and non-epidemic year (in males 15 againstexpected 7.3; v21 = 12.2, P < 0.001, in females 12against expected 7.3; v21 = 4.53, P = 0.046). Inadults, significant differences were scarce becauseof smaller sample sizes, but all significant differ-ences (two in males and three in females) werebetween an epidemic and a non-epidemic year.
DISCUSSION
The abundance of Greenfinches wintering in Fin-land started to decline in the second half of 2008,at the same time that finch trichomonosis was firstconfirmed. The decline was especially marked inmid-winter numbers. The epidemic and declinecontinued during 2009, and the disease spreadnorthwards. High mortality and poor breeding suc-cess in 2009 led to a severe decline in breedingpopulation densities in 2010, even though massmortality was not detected in 2010. However, the
(a) (b)
Figure 4. Abundance of (a) Greenfinches and (b) Great Tits in the winter bird censuses in South, Central and North Finland in win-ters 1984–2011 (1984 = winter 1983/84).
© 2013 British Ornithologists’ Union
362 A. Lehikoinen et al.

disease was documented again in many locationsincluding in northern Finland in 2011. There maybe spatial biases in documented disease cases dueto lower human population density in the north,with consequent reduced likelihood of observingsick birds. However, we believe that with theexception of possible under-reporting of disease inNorth Finland, the observed temporal patternreflects the true changes in the general distributionof the disease.
Our results confirm that trichomonosis is a ‘sum-mer’ disease in Finland, occurring mainly duringwarm months. Breeding season surveys indicatehow the population has been affected by the diseaseduring the previous breeding and post-breedingseason. Both the body condition analyses and thenumber of confirmed disease cases strongly point tothe peak epidemic years of 2008–2009, when sam-ples of ringed birds were also in weaker conditionthan in other years in the 2000s.
The observed population decline of Greenfinch-es in Finland (at least 35% in the whole country,but as great as 47% in South Finland) was of
Figure 5. Foreign encounters of Greenfinches ringed inFinland in 1991–2010 divided into four seasons. Blacksquares = December–February (n = 28), grey triangles pointingupwards = March–April (n = 15), white circles = May–August(n = 7) and grey triangles pointing downwards = September–November (n = 66).
Figure 6. Foreign encounters of chaffinches ringed in Finlandin 1991–2010 divided in four seasons. Black squares =December–February (n = 7), grey triangles pointing upwards =March–April (n = 7), white circles = May–August (n = 2) andgrey triangles pointing downwards = September–November(n = 10).
Figure 7. Proportion of lean Greenfinches (bars and left axis)by years in 1975–2011 and numbers of birds measured (lineand right axis) in September–October in southwest Finland.Trapping effort has been fairly constant in 2001–2011. Notethe fall in numbers trapped during the trichomonosis epidemicin the autumn of 2008 and 2009.
© 2013 British Ornithologists’ Union
Trichomonas-epidemic and Finnish Greenfinches 363
similar magnitude to that documented in Britain(35%; Robinson et al. 2010). The Finnish breedingpopulation was estimated at 370 000 pairs beforethe marked population decline (R. A. V€ais€anenunpubl. data), implying a loss to disease of over250 000 birds. Surprisingly, there has been muchsmaller decline (c. 20%) in Sweden (Lindstr€omet al. 2012), even though the disease emerged inSweden around the same time as in Finland (Nei-manis et al. 2010).
In contrast, the breeding population of the con-trol species, Great Tit, did not decline during theepidemic years, although wintering numbersshowed a slight but significant decline between2006 and 2010. Chaffinch breeding populationsdeclined slightly in Central Finland, but not inSouth Finland, where most trichomonosis caseswere observed. In contrast in the UK, Chaffinchpopulations did show significant decline (21% in
one region, Robinson et al. 2010). The differencebetween population responses of Greenfinches andChaffinches in Finland may reflect the fact that inFinland Greenfinches occur mainly in habitats nearhuman settlements, whereas the Chaffinch is aforest species (V€ais€anen et al. 1998). Bird feedersoccur only in areas near humans and they maygather birds from surrounding areas. Thus basedon habitat preferences, Greenfinches may have amuch higher probability than Chaffinches of visit-ing feeders if food is available (V€ais€anen 2008)and hence of being exposed to T. gallinae as theyoften feed in close contact with each other.
In all age-sex classes of Greenfinches, meanbody condition was poorer during the epidemic. Infinches infected by T. gallinae, weak body condi-tion is a result of inflammation in the throat andcrop, which makes it difficult and finally impossi-ble for the bird to swallow, thus causing birds touse their fat reserves and muscles for energy(Neimanis et al. 2010). The body mass data werecollected at ringing sites where several sick birdswere observed (and a few examined post mortemand confirmed as suffering from trichomonosis) inAugust–November in 2008 and 2009. Therefore,it is likely that weak body condition with fat andmuscle scores of zero, and body mass of moribundindividuals below 24 g, was due to trichomonosis.Greenfinches that were found dead and submittedfor necropsy had a mean weight of only 21.3 g.
The observed winter population declines ofGreenfinches were greatest in South Finland. Thisaccords with the fact that the disease was firstobserved in the south and spread north later.Nevertheless, there might be other reasons whythe northern populations suffered less from thedisease, which are not mutually exclusive. First,the disease only occurs in sufficiently warm andmoist conditions (Greiner & Ritchie 1994). Thespread of T. gallinae at shared feeding places islikely to be less efficient in colder environments.Secondly, both Greenfinch and Chaffinch declinein population density from south to north(V€ais€anen et al. 1998), and infection may be lesslikely at lower densities (Begon et al. 2006). Thereis an urgent need for further investigation of theeffect of host density on the spread of the finchtrichomonosis. Thirdly, the 2009/10 winter wasvery cold in Europe, especially in central Europe,where migratory Finnish Greenfinches were win-tering. For example, the winter NAO-index(North Atlantic Oscillation), which describes the
(a)
(b)
Figure 8. Mean body mass of Greenfinches in September–October in 2001–2011 by sex-age categories: (a) males (adult,n = 202, filled circle; first-year birds, n = 2809, open circle);(b) females (adult, n = 221; filled square; first-year birds,n = 2353, open square). Symbols of the first-year birds areshifted to the right for clarity. Vertical lines give 95% confi-dence intervals of means. See details of the model in text.The year 2004 is not presented due to small sample size.
© 2013 British Ornithologists’ Union
364 A. Lehikoinen et al.

general severity of the winter (positive valuesmean warm winters, negative cold; Hurrell 1995)was the lowest since the 1960s (–4.64; http://www.cgd.ucar.edu/cas/jhurrell/indices.html). Thismay have inhibited the spread of Trichomonas atshared food sources in the wintering areas. In sup-port of this, no Greenfinch infections were docu-mented in Finland during the breeding season of2010. Thus the disease may have become rarerjust before its spread north. This seems to havebeen only a temporary delay, as in the summerand autumn of 2011 incidents of finch trichomo-nosis were observed further north than recordedpreviously.
The cold 2009/10 winter could itself havereduced the Greenfinch population size in 2010,and the wintering numbers of the control species,Great Tit, showed a slight decline during 2006–2010, although this was much less marked thanthat of Greenfinches, and was not reflected in anybreeding population decline. According to Finnishringing data, the majority of the migratory FinnishGreenfinches were wintering in the area (southernSweden, northern Germany), where the diseasewas observed in 2008 (Peters et al. 2009, Neiman-is et al. 2010). Chaffinches winter mainly furthersouthwest, but also migrate through this area. It isinteresting that there were no encounters of Finn-ish ringed Chaffinches or Greenfinches in the UK,but the UK data show that at least some Chaf-finches ringed in the UK have been found in Fin-land (Lawson et al. 2011). Nevertheless, there aremany observations showing that both speciesmigrate between the UK and Scandinavia (Swedenand Norway) (Wernham et al. 2002, Bakken et al.2006, Fransson & Hall-Karlsson 2008, Lawsonet al. 2011). Thus, it may be more likely that thedisease spread first from the UK to Scandinavia,and from there to Finland by migrant birds.
In conclusion, the Finnish Greenfinch popula-tion has faced rapid and severe population declinedue to an epidemic of finch trichomonosis, but theeffect on Chaffinches has been minor. The declineof Greenfinches has been greater than in neigh-bouring Sweden and of the same order as in UK,where the epidemic was first observed (Robinsonet al. 2010). We emphasize the importance ofannual monitoring schemes now and especially inthe future in order to detect rapid populationchanges in common bird species that may becaused by disease epidemics. We encourage fur-ther investigations to study the spread and impact
of finch trichomonosis across European finchpopulations, and recognize a need to evaluatefactors (such as density, feeding practices andclimate) that vary between countries and mayexplain differences in the impacts observed.
We thank the numerous volunteers who helped to col-lect data in the field including censuses, ringing data andinformation on infected birds. Marko Kakko from Kaari-na was first to report observations of the disease. Com-ments of two anonymous referees improved the clarityof the manuscript.
REFERENCES
Atkinson, C.T., Thomas, N.J. & Hunter, D.B. (eds) 2008.Parasitic Diseases of Wild Birds. New Delhi: Wiley-Blackwell.
Bakken, V., Runde, O. & Tjørve, E. 2006. NorskRingmerkingsatlas, Vol. 2. Stavanger: Stavanger Museum.
Begon, M., Townsend, C.R. & Harper, J.L. 2006. Ecology,Individuals, Populations and Communities, 4th edn. Boston:Blackwell.
Bunbury, N., Jones, C.G., Greenwood, A.G. & Bell, D.J. 2007.Trichomonas gallinae in Mauritian columbids: implications foran endangered endemic. J. Wildl. Dis. 43: 399–407.
Bunbury, N., Jones, C.G., Greenwood, A.G. & Bell, D.J.2008. Epidemiology and conservation implications ofTrichomonas gallinae infection in the endangered Mauritianpink pigeon. Biol. Conserv. 141: 153–161.
Dhondt, A.A., Tessaglia, D.L. & Slothower, R.L. 1998.Epidemic mycoplasmal conjunctivitis in house finches fromEastern North America. J. Wildl. Dis. 34: 265–280.
Erwin, K.G., Kloss, C., Lyles, J., Felderhoff, J., Fedynich,A.M., Henke, S.E. & Roberson, J.A. 2000. Survival ofTrichomonas gallinae in white-winged dove carcasses.J. Wildl. Dis. 36: 551–554.
Forz�an, M.J., Vanderstichel, R., Melekhovets, Y.F. &McBurney, S. 2010. Trichomoniasis in finches from theCanadian Maritime provinces – an emerging disease. Can.Vet. J. 51: 391–396.
Fransson, T. & Hall-Karlsson, S. 2008.Svensk Ringm€arknings-atlas, Vol. 3. Stockholm: SOF.
Greiner, E.C. & Ritchie, B.W. 1994. Parasites. In Ritchie,B.W., Harrison, G.J. & Harrison, L.R. (eds) Avian Medicine:Principles and Application: 1007–1029. Lake Worth, FL:Wingers Publishing.
Haugen, A.O. 1952. Trichomoniasis in Alabama mourningdoves. J. Wildl. Manage. 16: 164–169.
Hofle, U., Gortazar, C., Ortiz, J.A., Knispel, B. & Kaleta,E.F. 2004. Outbreak of trichomoniasis in a woodpigeon(Columba palumbus) wintering roost. Eur. J. Wildl. Res. 50:73–77.
Hollm�en, T.E., Franson, J.C., Kilpi, M., Docherty, D.E. &Myllys, V. 2003. An adenovirus associated with intestinalimpaction and mortality of male common eiders (Somateriamollissima) in the Baltic Sea. J. Wildl. Dis. 39: 114–120.
Hurrell, J.W. 1995. Decadal trends in the North AtlanticOscillation: regional temperatures and precipitation. Science296: 676–679.
© 2013 British Ornithologists’ Union
Trichomonas-epidemic and Finnish Greenfinches 365

Koskimies, P. & V€ais€anen, R.A. (eds) 1991. Monitoring BirdPopulations. Helsinki: Zoological Museum, Finnish Museum ofNatural History, University of Helsinki. Available at: www.luomus.fi/seurannat/methods.htm (accessed 14 September2012).
Lawson, B., Robinson, R.A., Neimanis, A., Handland, K.,Isomursu, M., Agren, E.O., Hamnes, I.S., Tyler, K.M.,Chantrey, J., Hugnes, L.A., Pennycott, T.W., Simpson,V.R., John, S.K., Peck, K.M., Toms, M.P., Bennett, M.,Kirkwood, J.K. & Cunningham, A. 2011 Evidence ofspread of the emerging infectious disease, finchtrichomonosis, by migrating birds. EcoHealth 8: 143–153.
Lehikoinen, A. & V€ah€atalo, A. 2000. Phenology of birdmigration at the Hanko Bird Observatory, Finland, in 1979–1999. Tringa 27: 150–224 (in Finnish).
Lehikoinen, A., Ekroos, J., Jaatinen, K., Lehikoinen, P.,Lind�en, A., Piha, M. & V€ah€atalo, A. 2008. Bird populationtrends based on the data of Hanko Bird Observatory(Finland) during 1979–2007. Tringa 35: 146–209 (inFinnish).
Lindstr€om, �A., Green, M. & Ottvall, R. 2012. €Overvakning avF�aglarnas Populationsutveckling. �Arsrapport f€or 2011. Lund:Ekologiska institutionen, Lunds Universitet.
Liu, J., Xiao, H., Lei, F., Zhu, Q., Qin, K. & Zhang, X.W.2005. Highly pathogenic H5N1 influenza virus infection inmigratory birds. Science 309: 1206.
Neimanis, A.S., Handeland, K., Isomursu, M., �Agren, E.,Hamnes, I.S., Bergsjø, B. & Hirvel€a-Koski, V. 2010. Firstreport of epizootic trichomoniasis in wild finches (FamilyFringillidae) in southern Fennoscandia. Avian Dis. 54: 136–141.
Newton, I. 1998. Population Limitation in Birds. London:Academic Press.
Nowakowski, J.K. & V€ah€atalo, A. 2003. Is the great tit Parusmajor an irruptive migrant in North-East Europe? Ardea 91:231–244.
Pannekoek, J. & van Strien, A. 2004. TRIM 3 Manual(TRends & Indices for Monitoring Data). Voorburg: StatisticsNetherlands.
Pennycott, T., Lawson, B., Cunningham, A., Simpson, V. &Chantrey, J. 2005. Necrotic ingluvitis in wild finches. Vet.Rec. 157: 360.
Peters, M., Kilwinski, J., Reckling, D. & Henning, K. 2009.Geh€aufte Todesf€alle von wild lebenden Gru nfinken anFutterstellen infolge Trichomonas-gallinae-Infektionen – einaktuelles problem in Norddeutschland. Kleintierpraxis 54:433–438 (in German).
Real, J., Manosa, S. & Munoz, E. 2000. Trichomoniasis in aBonelli’s eagle population in Spain. J. Wildl. Dis. 36: 64–70.
Redpath, S.M., Mougeot, F., Leckie, F.M., Elston, D.A. &Hudson, P.J. 2006. Testing the role of parasites in drivingthe cyclic population dynamics of a gamebird. Ecol. Lett. 9:410–418.
Robinson, R.A., Lawson, B., Toms, M.P., Peck, K.M.,Kirkwood, J.K., Chantrey, J., Clatworthy, I.R., Evans,A.D., Hughes, L.A., Hutchinson, O.C., John, S.K.,Pennycott, T.W., Perkins, M.W., Rowley, P.S., Simpson,V.R., Tyler, K.M. & Cunningham, A.A. 2010. Emerginginfectious disease leads to rapid population declines ofcommon British birds. PLoS ONE 5: e12215.
Sansano-Maestre, J., Garijo-Toledo, M.M. & Gornez-Munoz,M.T. 2009. Prevalence and genotyping of Trichomonasgallinae in pigeons and birds of prey. Avian Pathol. 38: 201–207.
Stromberg, M.R., Koenig, W.D., Walters, E.L. &Schweisinger, J. 2008. Estimate of Trichomonas gallinae-induced mortality in band-tailed pigeons, upper CarmelValley, California, winter 2006–2007. Wilson J. Ornithol. 120:603–606.
Thomas, N.J., Hunter, D.B. & Atkinson, C.T. (eds) 2007.Infectious Diseases of Wild Birds. Oxford: BlackwellPublishing Ltd.
V€ais€anen, R.A. 2006. Monitoring population changes of 86land bird species breeding in Finland in 1983–2005. Linnut-vuosikirja 2005: 83–98 (in Finnish).
V€ais€anen, R.A. 2008. Changes in frequency and abundanceof 63 bird species at winter feeding sites in Finland during19 winters 1988/1989–2006/2007. Linnut-vuosikirja 2007:60–79 (in Finnish).
V€ais€anen, R.A., Lammi, E. & Koskimies, P. 1998.Distribution, Numbers and Population Changes of FinnishBreeding Birds. Helsinki: Otava (in Finnish).
V€ais€anen, R.A., Hario, M. & Saurola, P. 2011. Populationestimates of Finnish birds. In Valkama, J., Veps€al€ainen, V. &Lehikoinen, A. (eds) The Third Finnish Breeding Bird Atlas.Finnish Museum of Natural History and Ministry ofEnvironment. Available at: http://atlas3.lintuatlas.fi/english(accessed 7 May 2012).
Valkama, J., Veps€al€ainen, V. & Lehikoinen, A. 2011. TheThird Finnish Breeding Bird Atlas. Finnish Museum ofNatural History and Ministry of Environment. Available at:http://atlas3.lintuatlas.fi/english (accessed 29 November2011).
Villanua, D., Hofle, U., Perez-Rodriguez, L. & Gortazar, C.2006. Trichomonas gallinae in wintering Common WoodPigeons Columba palumbus in Spain. Ibis 148: 641–648.
Wernham, C., Toms, M., Marchant, J., Clark, J.,Siriwardena, G. & Baillie, S. 2002. The Migration Atlas:Movements of the Birds of Britain and Ireland. London: T. &A.D. Poyser.
Received 24 January 2012;revision accepted 27 December 2012.
Associate Editor: Luc Lens.
© 2013 British Ornithologists’ Union
366 A. Lehikoinen et al.





![GÉOPOLITIQUE ET POPULATIONS AU TCHAD [Geopolitics and populations in Chad]](https://static.fdokumen.com/doc/165x107/631378e5fc260b71020f1c3f/geopolitique-et-populations-au-tchad-geopolitics-and-populations-in-chad.jpg)














