
Impact of imazethapyr on the microbial community structure in agricultural soils
7
Impact of imazethapyr on the microbial community structure in agricultural soils Changpeng Zhang a,b,1 , Jun Xu a,1 , Xingang Liu a , Fengshou Dong a , Zhiqiang Kong a , Yu Sheng a , Yongquan Zheng a, * a Key Laboratory of Pesticide Chemistry and Application, Ministry of Agriculture, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China b Institute of Quality and Standard for Agro-products, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China article info Article history: Received 25 March 2010 Received in revised form 24 June 2010 Accepted 29 June 2010 Available online 24 July 2010 Keywords: Microbial community structure PLFA Herbicide Imazethapyr abstract Large amounts of imazethapyr were applied for weed control in cultivation fields in China, but their effects on the soil microbial community remains unclear. In this study, two agricultural soils, a silty loam (HS) and a loamy soil (QL), were spiked with imazethapyr (CK, 0.1, 1 and 10 mg kg 1 ) and incubated for 1, 15, 30, 60, 90 and 120 d. In addition, untreated controls received only water. The soil microbial commu- nity structures were characterized by investigating the phospholipid fatty acids (PLFA) and microbial bio- mass C. Soil microbial biomass C and total concentration of PLFA were variable with incubation time, which were also reduced by the addition of imazethapyr. Imazethapyr addition also decreased the ratios of GN/GP and fungi/bacteria. A larger stress level, measured as the ratio of PLFA (cyc17:0 + cyc19:0)/ (16:1x7c + 18:1x7c), was found in the high concentration (1 and 10 mg kg 1 ) herbicide treatment groups. The effects of imazethapyr at the field application on soil microbial biomass and microbial com- munity were minor. Principal component analysis (PCA) of the PLFA clearly separated the treatments and incubation times. Both soils showed different total PLFA concentrations and ratios of GN/GP and fungi/ bacteria, but similar changes in the PLFA pattern upon soil treatment. The soil microbial community structure was shifted by the addition of imazethapyr, which recovered after 60 d. In addition, the dissi- pation of imazethapyr was slow in both soils. Our results demonstrated that the addition of imazethapyr shifted the microbial community structure, but that it recovered after a period of incubation. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Imazethapyr is an imidazolinone herbicide that is currently used for broad-spectrum weed control in soybean and other le- gume crops (Perucci and Scarponi, 1994). Imazethapyr inhibits the biosynthesis of branched chain amino acids by blocking aceto- lactate synthase (ALS), which is the first common enzyme of the pathway (Zabalza et al., 2007). In China, 150–300 tons of imazeth- apyr are used each year to control weeds in soybean fields (Li et al., 2008). Imazethapyr adsorption was high, increased with decreas- ing soil solution pH in Sharpsburg silty clay loam and Holdrege silt loam (Stougaard et al., 1990). In addition imazethapyr has been shown to be less mobile and to persist for a long time in soils (Stougaard et al., 1990; Loux and Reese, 1993). After 52 weeks in the field, 20–40% of the applied imazethapyr remained in the soil (Goetz et al., 1990). Imazethapyr has also been detected in ground- water supplies and surface water courses (Battaglin et al., 2000), where they represent a threat to human health and the environ- ment. The intensive use of imazethapyr has caused environmental concern because of contamination of non-target sites, especially unforeseen consequences on soil microbes. Soil microorganisms are extremely important among non-target organisms since they are a key component of soil ecosystems, dom- inating the cycling of nutrient elements and playing a major role in maintaining soil quality (Wang et al., 2008). A number of studies have attempted to determine the effects of imazethapyr and other herbicides on soil microbes. The herbicide imazethapyr, at 10-fold and 100-fold higher rates of the recommended field rate for soybean weeding, decreased the biomass C content and dehydrogenase activ- ity and increased the hydrolytic capacity and protease and catalase activity (Perucci and Scarponi 1994). The herbicides rimsulfuron and imazethapyr exerted a detrimental effect on soil microbial bio- mass, and phosphatase activity was enhanced by 10-fold when com- pared to the field rates in response to treatment with these two herbicides (Perucci et al., 2000). In a greenhouse experiment, imaz- ethapyr reduced the soil microbial biomass C and microbial diversity (Lupwayi et al., 2004). However, few studies have examined the ef- fects of the herbicide imazethapyr on the soil microbial community structure in agricultural soils in China. PLFAs are considered to be derived mainly from living organisms, and signature fatty acids can be used as indicators for biomass belonging to certain microbial groups and the ratios between them. The sum of the microbial signature fatty acids in a given sample can 0045-6535/$ - see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2010.06.079 * Corresponding author. Tel./fax: +86 10 62815908. E-mail address: [email protected] (Y. Zheng). 1 These authors made the equivalent contribution to this paper. Chemosphere 81 (2010) 800–806 Contents lists available at ScienceDirect Chemosphere journal homepage: www.elsevier.com/locate/chemosphere
Transcript of Impact of imazethapyr on the microbial community structure in agricultural soils

Chemosphere 81 (2010) 800–806
Contents lists available at ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Impact of imazethapyr on the microbial community structure in agricultural soils
Changpeng Zhang a,b,1, Jun Xu a,1, Xingang Liu a, Fengshou Dong a, Zhiqiang Kong a, Yu Sheng a,Yongquan Zheng a,*
a Key Laboratory of Pesticide Chemistry and Application, Ministry of Agriculture, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, Chinab Institute of Quality and Standard for Agro-products, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
a r t i c l e i n f o a b s t r a c t
Article history:Received 25 March 2010Received in revised form 24 June 2010Accepted 29 June 2010Available online 24 July 2010
Keywords:Microbial community structurePLFAHerbicideImazethapyr
0045-6535/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.chemosphere.2010.06.079
* Corresponding author. Tel./fax: +86 10 62815908E-mail address: [email protected] (Y. Zheng).
1 These authors made the equivalent contribution to
Large amounts of imazethapyr were applied for weed control in cultivation fields in China, but theireffects on the soil microbial community remains unclear. In this study, two agricultural soils, a silty loam(HS) and a loamy soil (QL), were spiked with imazethapyr (CK, 0.1, 1 and 10 mg kg�1) and incubated for 1,15, 30, 60, 90 and 120 d. In addition, untreated controls received only water. The soil microbial commu-nity structures were characterized by investigating the phospholipid fatty acids (PLFA) and microbial bio-mass C. Soil microbial biomass C and total concentration of PLFA were variable with incubation time,which were also reduced by the addition of imazethapyr. Imazethapyr addition also decreased the ratiosof GN/GP and fungi/bacteria. A larger stress level, measured as the ratio of PLFA (cyc17:0 + cyc19:0)/(16:1x7c + 18:1x7c), was found in the high concentration (1 and 10 mg kg�1) herbicide treatmentgroups. The effects of imazethapyr at the field application on soil microbial biomass and microbial com-munity were minor. Principal component analysis (PCA) of the PLFA clearly separated the treatments andincubation times. Both soils showed different total PLFA concentrations and ratios of GN/GP and fungi/bacteria, but similar changes in the PLFA pattern upon soil treatment. The soil microbial communitystructure was shifted by the addition of imazethapyr, which recovered after 60 d. In addition, the dissi-pation of imazethapyr was slow in both soils. Our results demonstrated that the addition of imazethapyrshifted the microbial community structure, but that it recovered after a period of incubation.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction concern because of contamination of non-target sites, especially
Imazethapyr is an imidazolinone herbicide that is currentlyused for broad-spectrum weed control in soybean and other le-gume crops (Perucci and Scarponi, 1994). Imazethapyr inhibitsthe biosynthesis of branched chain amino acids by blocking aceto-lactate synthase (ALS), which is the first common enzyme of thepathway (Zabalza et al., 2007). In China, 150–300 tons of imazeth-apyr are used each year to control weeds in soybean fields (Li et al.,2008). Imazethapyr adsorption was high, increased with decreas-ing soil solution pH in Sharpsburg silty clay loam and Holdrege siltloam (Stougaard et al., 1990). In addition imazethapyr has beenshown to be less mobile and to persist for a long time in soils(Stougaard et al., 1990; Loux and Reese, 1993). After 52 weeks inthe field, 20–40% of the applied imazethapyr remained in the soil(Goetz et al., 1990). Imazethapyr has also been detected in ground-water supplies and surface water courses (Battaglin et al., 2000),where they represent a threat to human health and the environ-ment. The intensive use of imazethapyr has caused environmental
ll rights reserved.
.
this paper.
unforeseen consequences on soil microbes.Soil microorganisms are extremely important among non-target
organisms since they are a key component of soil ecosystems, dom-inating the cycling of nutrient elements and playing a major role inmaintaining soil quality (Wang et al., 2008). A number of studieshave attempted to determine the effects of imazethapyr and otherherbicides on soil microbes. The herbicide imazethapyr, at 10-foldand 100-fold higher rates of the recommended field rate for soybeanweeding, decreased the biomass C content and dehydrogenase activ-ity and increased the hydrolytic capacity and protease and catalaseactivity (Perucci and Scarponi 1994). The herbicides rimsulfuronand imazethapyr exerted a detrimental effect on soil microbial bio-mass, and phosphatase activity was enhanced by 10-fold when com-pared to the field rates in response to treatment with these twoherbicides (Perucci et al., 2000). In a greenhouse experiment, imaz-ethapyr reduced the soil microbial biomass C and microbial diversity(Lupwayi et al., 2004). However, few studies have examined the ef-fects of the herbicide imazethapyr on the soil microbial communitystructure in agricultural soils in China.
PLFAs are considered to be derived mainly from living organisms,and signature fatty acids can be used as indicators for biomassbelonging to certain microbial groups and the ratios between them.The sum of the microbial signature fatty acids in a given sample can

C. Zhang et al. / Chemosphere 81 (2010) 800–806 801
be used as a measure of microbial biomass (Frostegard et al., 1993);therefore, the fatty acid profiles can reflect the impact of imazetha-pyr on the general microbial community composition.
The aims of this study were to examine the effects of the herbi-cide imazethapyr on the total amount of soil microbial biomass(microbial biomass C and total PLFAs) and community structure(GN/GP, fungi/bacteria, the stress level, etc.) in two agriculturalsoils. The microbial biomass C and microbial community structurewere studied by the chloroform fumigation–extraction methodand by the culture-independent method of PLFA analysis. The find-ings are useful to understand the interactions between herbicideand microbes in agricultural soils.
2. Materials and methods
2.1. Soil samples
Two agricultural field soils typical for China were sampled forthe incubation experiments and characterized based on selectedsoil properties (Table 1). Soil 1 (HS) was collected from an experi-mental plot near Langfang (N39�820, E116�700), in Hebei Province.HS was a silty loam that had not been tilled or planted with cropsin the last 2 years. The average temperatures in the region are�1.0 �C in winter and 24.3 �C in summer, and the area receivesapproximately 500–600 mm of rain per year. Soil 2 (QL) was col-lected from the soybean field of Laolai Farm near the City of Nehe(N48�290, E124�510) in Heilongjiang Province. QL was a loamy soilthat had been subjected to conventional-tillage and soybean plant-ing over the last 2 years. The average temperatures in the regionwere �7.5 �C in winter and 22.7 �C in summer, with rainfall ofapproximately 400–550 mm per year. The two soils were not trea-ted with pesticides during the last 2 years.
Soil samples from the top 0–15 cm were collected with a stain-less steel soil tube drill with a diameter of 3 cm. Duplicate compos-ite samples were taken to the laboratory in coolers with ice bags.The samples were then mixed, sieved with 2 mm mesh to removethe plant tissue, and preserved at low temperature before use.
A series of pots (7 � 10 cm) containing 100 g of dried soil wereincubated with distilled water in the dark at 25 �C for 14 d, afterwhich imazethapyr at 0.1, 1 or 10 mg kg�1 soil was added to the trea-ted pots. Control pots received distilled water only. Each experimentwas conducted in triplicate. The pots were sealed and placed in anenvironmental chamber set at 25 �C and 50% humidity. Aerationwas conducted once a day for 1 h and the soil moisture was main-tained at 60% of the water-holding capacity with deionized water.The pots were removed from the environmental chamber at randomafter various incubation time intervals (1, 15, 30, 60, 90 or 120 d), andthe soils were then analyzed for microbial biomass C, communitystructure, and concentration of imazethapyr as described below.
2.2. Determination and analysis of imazethapyr
Imazethapyr was determined by a sequential extraction proce-dure (Ma et al., 2007). Briefly, a 20 g soil sample was weighed intoa 50 mL tube, and 25 mL of 0.1 M NH3–NH4Cl buffer (pH 10) wereadded. The mixture was then swirled for 1 min, extracted for
Table 1Characteristics of the two soils.
Soil Clay(%)
Slit(%)
Sand(%)
Organic matter(g kg�1)
pH Total N (gkg�1)
Total Pkg�1)
HS (siltyloamsoil)
20.63 62.13 17.24 22.13 6.92 0.61 5.12
QL (loamysoil)
23.71 34.83 41.46 56.51 8.48 3.12 1.64
20 min by ultrasonication and centrifuged for 3 min at 6000 rpm.Supernatants of 10 mL were then removed and acidified to pH2.0. The residue was purified on a C18 cartridge (Cleanert C18,500 mg/6 mL, Agela Technologies Inc.) that had been precondi-tioned with 5 mL hexane, 5 mL CH2Cl2, 5 mL CH3OH, 5 mL waterand 5 mL 0.01 M HCl before the addition of 5 mL of acidified super-natant. The cartridge was then washed with 5 mL of water and5 mL of hexane, after which it was eluted with 10 mL CH2Cl2. Next,the filtrate was dried under N2 and resuspended in 2 mL of meth-anol. The residue was then filtered through a 0.22 lm nylon syr-inge filter for chromatographic injection.
The Waters Acquity UPLC system (USA) consisted of a binarysolvent manager and an Acuity column heater equipped with aWaters Acquity UPLC BEH C18 column (2.1 mm � 50 mm, 1.7 lmparticle size) (Milford, MA, USA). Gradient UPLC elution was per-formed using methanol (LC grade) as mobile phase A and 0.2% for-mic acid in ultrapure water as mobile phase B. Separation of theanalytes was performed at 45 �C at a flow rate of 0.3 mL min�1
with an elution gradient of 90–10% B in 2.5 min, then 90% B in0.1 min, holding at 90% B for 2 min. The temperature in the auto-sampler was set at 10 �C and the sample volume injected was 3 lL.
A triple-quadrupole mass spectrometer (Waters Corp., USA)equipped with an electrospray ionization source was used forimazethapyr detection. MS/MS detection was performed in posi-tive mode, and the monitoring conditions were optimized forimazethapyr. Acquisition parameters were a capillary voltage of3 kV, source temperature of 120 �C and desolvation temperatureof 350 �C. The cone and desolvation gas flows were 50 L h�1 and400 L h�1 nitrogen, respectively. The precursor ion was 290 (m/z)and its product quantitative ion was 245 (m/z) when the collisionenergy was set to 22 V. The multiple reaction monitoring (MRM)was conducted in scan mode.
2.3. Determination and analysis of MBC
The MBC was determined by the chloroform fumigation–extrac-tion method (Wang et al., 2007; Lin et al., 1999), with brief modi-fication. Each soil sample was divided into two groups thatcontained 20.0 g of dry soil. One group was fumigated with etha-nol-free chloroform for 24 h in the dark at 25 �C, while anothergroup was untreated. Both fumigated and unfumigated soils wereextracted with 0.5 M K2SO4 solution for 30 min on a shaker [1:4soil/solution (w/w)]. The supernatant was then filtered, after which10.00 mL of filtrate was mixed with 5.00 mL 0.2 M K2Cr2O7 and5 mL H2SO4. The mixture with zeolite was boiled for 10 min at170–180 �C before cooling and titrating with 0.05 M FeSO4
C2H8N2 as an indicator. When the solution changed from yellowto cyan to brownish-red, titration was stopped. The MBC was cal-culated by MBC = Ec/Kc, where Ec = (C extracted from fumigatedsoil) � (C extracted from nonfumigated soil), with C representingcarbon and Kc being a calibration factor of 0.38 (Wu et al., 2006).
2.4. Determination and analysis of PLFA
Phospholipid fatty acid (PLFA) analysis was based on Bossioet al. (1998) and Frostegard et al. (1993). Briefly, a 5 g sample of
(g Total K (gkg�1)
Available N(mg kg�1)
Available P(mg kg�1)
Available K(mg kg�1)
CEC (cmolkg�1)
32.53 234.26 16.38 234.00 12.50
30.20 115.97 97.05 890.30 95.91
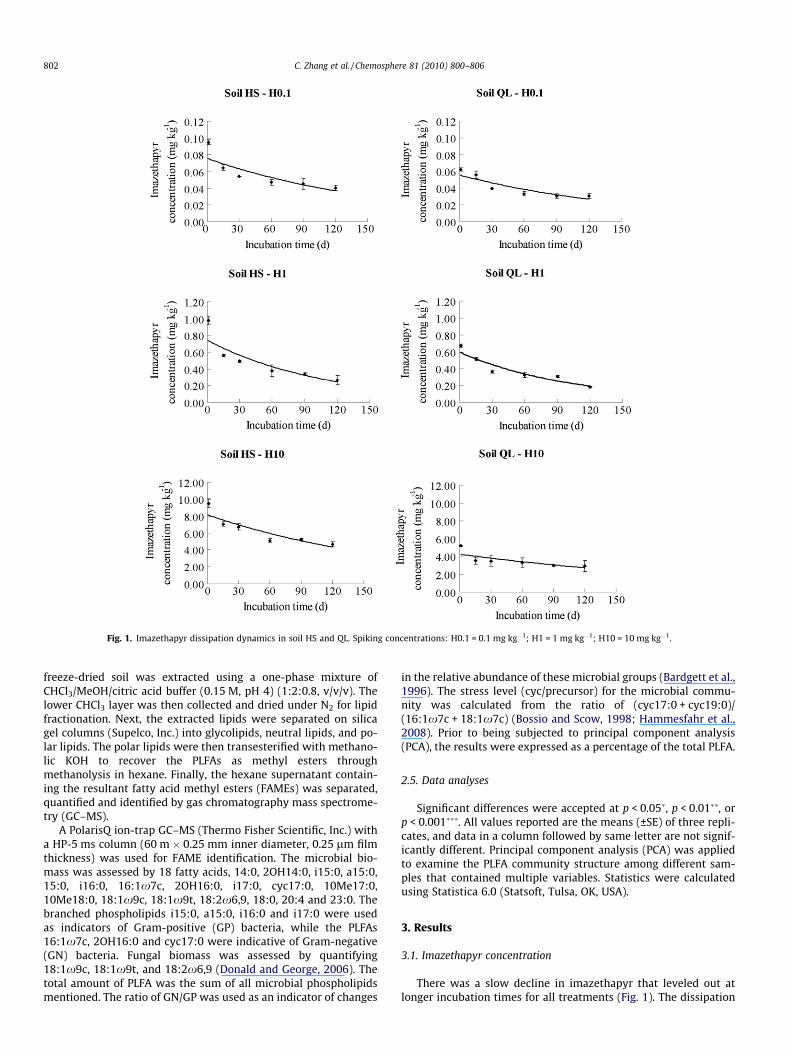
Fig. 1. Imazethapyr dissipation dynamics in soil HS and QL. Spiking concentrations: H0.1 = 0.1 mg kg�1; H1 = 1 mg kg�1; H10 = 10 mg kg�1.
802 C. Zhang et al. / Chemosphere 81 (2010) 800–806
freeze-dried soil was extracted using a one-phase mixture ofCHCl3/MeOH/citric acid buffer (0.15 M, pH 4) (1:2:0.8, v/v/v). Thelower CHCl3 layer was then collected and dried under N2 for lipidfractionation. Next, the extracted lipids were separated on silicagel columns (Supelco, Inc.) into glycolipids, neutral lipids, and po-lar lipids. The polar lipids were then transesterified with methano-lic KOH to recover the PLFAs as methyl esters throughmethanolysis in hexane. Finally, the hexane supernatant contain-ing the resultant fatty acid methyl esters (FAMEs) was separated,quantified and identified by gas chromatography mass spectrome-try (GC–MS).
A PolarisQ ion-trap GC–MS (Thermo Fisher Scientific, Inc.) witha HP-5 ms column (60 m � 0.25 mm inner diameter, 0.25 lm filmthickness) was used for FAME identification. The microbial bio-mass was assessed by 18 fatty acids, 14:0, 2OH14:0, i15:0, a15:0,15:0, i16:0, 16:1x7c, 2OH16:0, i17:0, cyc17:0, 10Me17:0,10Me18:0, 18:1x9c, 18:1x9t, 18:2x6,9, 18:0, 20:4 and 23:0. Thebranched phospholipids i15:0, a15:0, i16:0 and i17:0 were usedas indicators of Gram-positive (GP) bacteria, while the PLFAs16:1x7c, 2OH16:0 and cyc17:0 were indicative of Gram-negative(GN) bacteria. Fungal biomass was assessed by quantifying18:1x9c, 18:1x9t, and 18:2x6,9 (Donald and George, 2006). Thetotal amount of PLFA was the sum of all microbial phospholipidsmentioned. The ratio of GN/GP was used as an indicator of changes
in the relative abundance of these microbial groups (Bardgett et al.,1996). The stress level (cyc/precursor) for the microbial commu-nity was calculated from the ratio of (cyc17:0 + cyc19:0)/(16:1x7c + 18:1x7c) (Bossio and Scow, 1998; Hammesfahr et al.,2008). Prior to being subjected to principal component analysis(PCA), the results were expressed as a percentage of the total PLFA.
2.5. Data analyses
Significant differences were accepted at p < 0.05�, p < 0.01��, orp < 0.001���. All values reported are the means (±SE) of three repli-cates, and data in a column followed by same letter are not signif-icantly different. Principal component analysis (PCA) was appliedto examine the PLFA community structure among different sam-ples that contained multiple variables. Statistics were calculatedusing Statistica 6.0 (Statsoft, Tulsa, OK, USA).
3. Results
3.1. Imazethapyr concentration
There was a slow decline in imazethapyr that leveled out atlonger incubation times for all treatments (Fig. 1). The dissipation
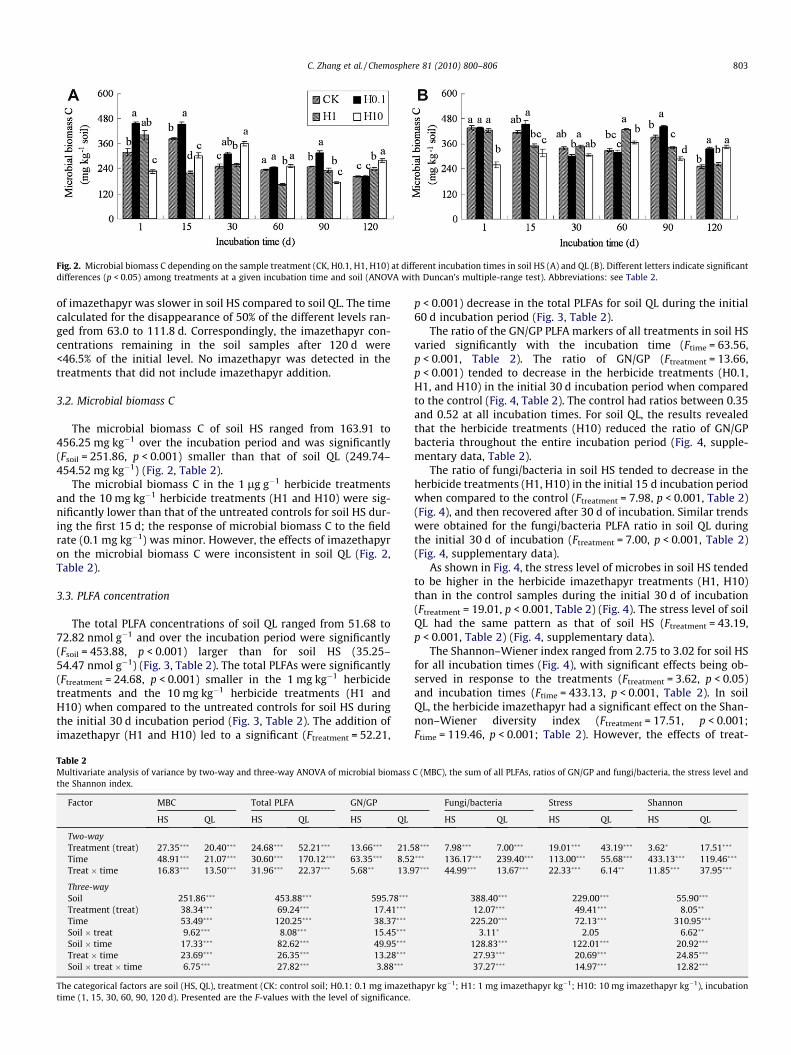
Fig. 2. Microbial biomass C depending on the sample treatment (CK, H0.1, H1, H10) at different incubation times in soil HS (A) and QL (B). Different letters indicate significantdifferences (p < 0.05) among treatments at a given incubation time and soil (ANOVA with Duncan’s multiple-range test). Abbreviations: see Table 2.
C. Zhang et al. / Chemosphere 81 (2010) 800–806 803
of imazethapyr was slower in soil HS compared to soil QL. The timecalculated for the disappearance of 50% of the different levels ran-ged from 63.0 to 111.8 d. Correspondingly, the imazethapyr con-centrations remaining in the soil samples after 120 d were<46.5% of the initial level. No imazethapyr was detected in thetreatments that did not include imazethapyr addition.
3.2. Microbial biomass C
The microbial biomass C of soil HS ranged from 163.91 to456.25 mg kg�1 over the incubation period and was significantly(Fsoil = 251.86, p < 0.001) smaller than that of soil QL (249.74–454.52 mg kg�1) (Fig. 2, Table 2).
The microbial biomass C in the 1 lg g�1 herbicide treatmentsand the 10 mg kg�1 herbicide treatments (H1 and H10) were sig-nificantly lower than that of the untreated controls for soil HS dur-ing the first 15 d; the response of microbial biomass C to the fieldrate (0.1 mg kg�1) was minor. However, the effects of imazethapyron the microbial biomass C were inconsistent in soil QL (Fig. 2,Table 2).
3.3. PLFA concentration
The total PLFA concentrations of soil QL ranged from 51.68 to72.82 nmol g�1 and over the incubation period were significantly(Fsoil = 453.88, p < 0.001) larger than for soil HS (35.25–54.47 nmol g�1) (Fig. 3, Table 2). The total PLFAs were significantly(Ftreatment = 24.68, p < 0.001) smaller in the 1 mg kg�1 herbicidetreatments and the 10 mg kg�1 herbicide treatments (H1 andH10) when compared to the untreated controls for soil HS duringthe initial 30 d incubation period (Fig. 3, Table 2). The addition ofimazethapyr (H1 and H10) led to a significant (Ftreatment = 52.21,
Table 2Multivariate analysis of variance by two-way and three-way ANOVA of microbial biomassthe Shannon index.
Factor MBC Total PLFA GN/GP
HS QL HS QL HS QL
Two-wayTreatment (treat) 27.35��� 20.40��� 24.68��� 52.21��� 13.66��� 21.5Time 48.91��� 21.07��� 30.60��� 170.12��� 63.35��� 8.52Treat � time 16.83��� 13.50��� 31.96��� 22.37��� 5.68�� 13.9
Three-waySoil 251.86��� 453.88��� 595.78���
Treatment (treat) 38.34��� 69.24��� 17.41���
Time 53.49��� 120.25��� 38.37���
Soil � treat 9.62��� 8.08��� 15.45���
Soil � time 17.33��� 82.62��� 49.95���
Treat � time 23.69��� 26.35��� 13.28���
Soil � treat � time 6.75��� 27.82��� 3.88���
The categorical factors are soil (HS, QL), treatment (CK: control soil; H0.1: 0.1 mg imazettime (1, 15, 30, 60, 90, 120 d). Presented are the F-values with the level of significance.
p < 0.001) decrease in the total PLFAs for soil QL during the initial60 d incubation period (Fig. 3, Table 2).
The ratio of the GN/GP PLFA markers of all treatments in soil HSvaried significantly with the incubation time (Ftime = 63.56,p < 0.001, Table 2). The ratio of GN/GP (Ftreatment = 13.66,p < 0.001) tended to decrease in the herbicide treatments (H0.1,H1, and H10) in the initial 30 d incubation period when comparedto the control (Fig. 4, Table 2). The control had ratios between 0.35and 0.52 at all incubation times. For soil QL, the results revealedthat the herbicide treatments (H10) reduced the ratio of GN/GPbacteria throughout the entire incubation period (Fig. 4, supple-mentary data, Table 2).
The ratio of fungi/bacteria in soil HS tended to decrease in theherbicide treatments (H1, H10) in the initial 15 d incubation periodwhen compared to the control (Ftreatment = 7.98, p < 0.001, Table 2)(Fig. 4), and then recovered after 30 d of incubation. Similar trendswere obtained for the fungi/bacteria PLFA ratio in soil QL duringthe initial 30 d of incubation (Ftreatment = 7.00, p < 0.001, Table 2)(Fig. 4, supplementary data).
As shown in Fig. 4, the stress level of microbes in soil HS tendedto be higher in the herbicide imazethapyr treatments (H1, H10)than in the control samples during the initial 30 d of incubation(Ftreatment = 19.01, p < 0.001, Table 2) (Fig. 4). The stress level of soilQL had the same pattern as that of soil HS (Ftreatment = 43.19,p < 0.001, Table 2) (Fig. 4, supplementary data).
The Shannon–Wiener index ranged from 2.75 to 3.02 for soil HSfor all incubation times (Fig. 4), with significant effects being ob-served in response to the treatments (Ftreatment = 3.62, p < 0.05)and incubation times (Ftime = 433.13, p < 0.001, Table 2). In soilQL, the herbicide imazethapyr had a significant effect on the Shan-non–Wiener diversity index (Ftreatment = 17.51, p < 0.001;Ftime = 119.46, p < 0.001; Table 2). However, the effects of treat-
C (MBC), the sum of all PLFAs, ratios of GN/GP and fungi/bacteria, the stress level and
Fungi/bacteria Stress Shannon
HS QL HS QL HS QL
8��� 7.98��� 7.00��� 19.01��� 43.19��� 3.62� 17.51������ 136.17��� 239.40��� 113.00��� 55.68��� 433.13��� 119.46���
7��� 44.99��� 13.67��� 22.33��� 6.14�� 11.85��� 37.95���
388.40��� 229.00��� 55.90���
12.07��� 49.41��� 8.05��
225.20��� 72.13��� 310.95���
3.11� 2.05 6.62��
128.83��� 122.01��� 20.92���
27.93��� 20.69��� 24.85���
37.27��� 14.97��� 12.82���
hapyr kg�1; H1: 1 mg imazethapyr kg�1; H10: 10 mg imazethapyr kg�1), incubation
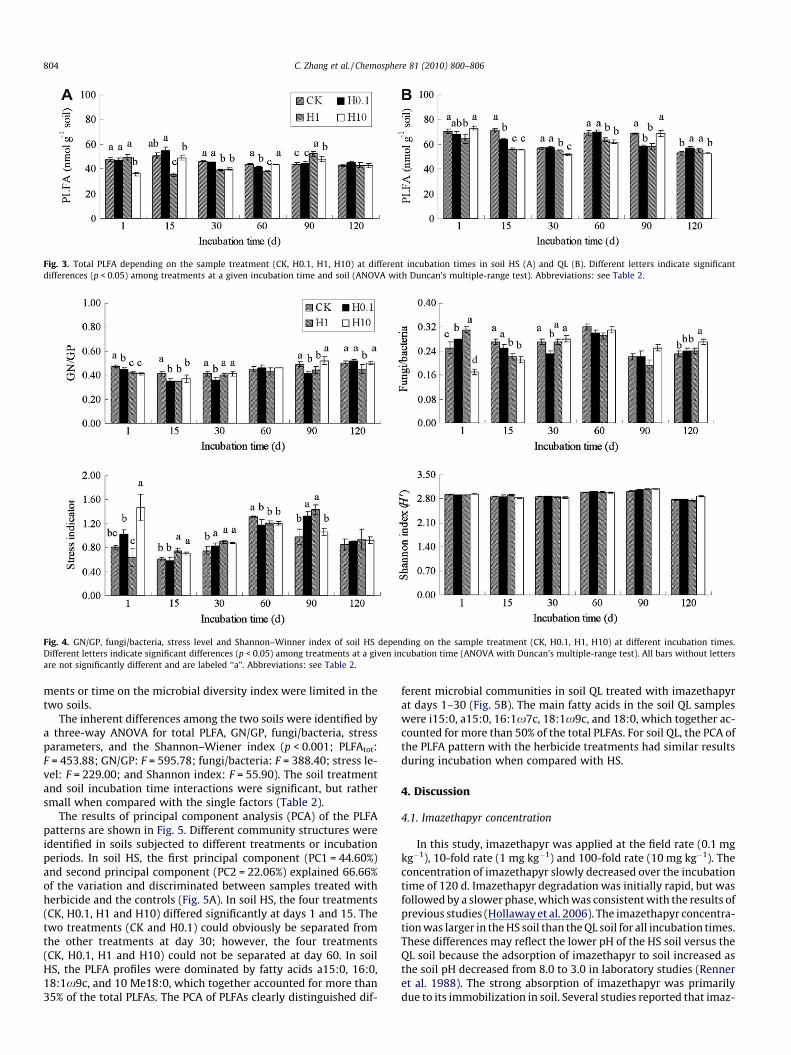
Fig. 3. Total PLFA depending on the sample treatment (CK, H0.1, H1, H10) at different incubation times in soil HS (A) and QL (B). Different letters indicate significantdifferences (p < 0.05) among treatments at a given incubation time and soil (ANOVA with Duncan’s multiple-range test). Abbreviations: see Table 2.
Fig. 4. GN/GP, fungi/bacteria, stress level and Shannon–Winner index of soil HS depending on the sample treatment (CK, H0.1, H1, H10) at different incubation times.Different letters indicate significant differences (p < 0.05) among treatments at a given incubation time (ANOVA with Duncan’s multiple-range test). All bars without lettersare not significantly different and are labeled ‘‘a”. Abbreviations: see Table 2.
804 C. Zhang et al. / Chemosphere 81 (2010) 800–806
ments or time on the microbial diversity index were limited in thetwo soils.
The inherent differences among the two soils were identified bya three-way ANOVA for total PLFA, GN/GP, fungi/bacteria, stressparameters, and the Shannon–Wiener index (p < 0.001; PLFAtot:F = 453.88; GN/GP: F = 595.78; fungi/bacteria: F = 388.40; stress le-vel: F = 229.00; and Shannon index: F = 55.90). The soil treatmentand soil incubation time interactions were significant, but rathersmall when compared with the single factors (Table 2).
The results of principal component analysis (PCA) of the PLFApatterns are shown in Fig. 5. Different community structures wereidentified in soils subjected to different treatments or incubationperiods. In soil HS, the first principal component (PC1 = 44.60%)and second principal component (PC2 = 22.06%) explained 66.66%of the variation and discriminated between samples treated withherbicide and the controls (Fig. 5A). In soil HS, the four treatments(CK, H0.1, H1 and H10) differed significantly at days 1 and 15. Thetwo treatments (CK and H0.1) could obviously be separated fromthe other treatments at day 30; however, the four treatments(CK, H0.1, H1 and H10) could not be separated at day 60. In soilHS, the PLFA profiles were dominated by fatty acids a15:0, 16:0,18:1x9c, and 10 Me18:0, which together accounted for more than35% of the total PLFAs. The PCA of PLFAs clearly distinguished dif-
ferent microbial communities in soil QL treated with imazethapyrat days 1–30 (Fig. 5B). The main fatty acids in the soil QL sampleswere i15:0, a15:0, 16:1x7c, 18:1x9c, and 18:0, which together ac-counted for more than 50% of the total PLFAs. For soil QL, the PCA ofthe PLFA pattern with the herbicide treatments had similar resultsduring incubation when compared with HS.
4. Discussion
4.1. Imazethapyr concentration
In this study, imazethapyr was applied at the field rate (0.1 mgkg�1), 10-fold rate (1 mg kg�1) and 100-fold rate (10 mg kg�1). Theconcentration of imazethapyr slowly decreased over the incubationtime of 120 d. Imazethapyr degradation was initially rapid, but wasfollowed by a slower phase, which was consistent with the results ofprevious studies (Hollaway et al. 2006). The imazethapyr concentra-tion was larger in the HS soil than the QL soil for all incubation times.These differences may reflect the lower pH of the HS soil versus theQL soil because the adsorption of imazethapyr to soil increased asthe soil pH decreased from 8.0 to 3.0 in laboratory studies (Renneret al. 1988). The strong absorption of imazethapyr was primarilydue to its immobilization in soil. Several studies reported that imaz-
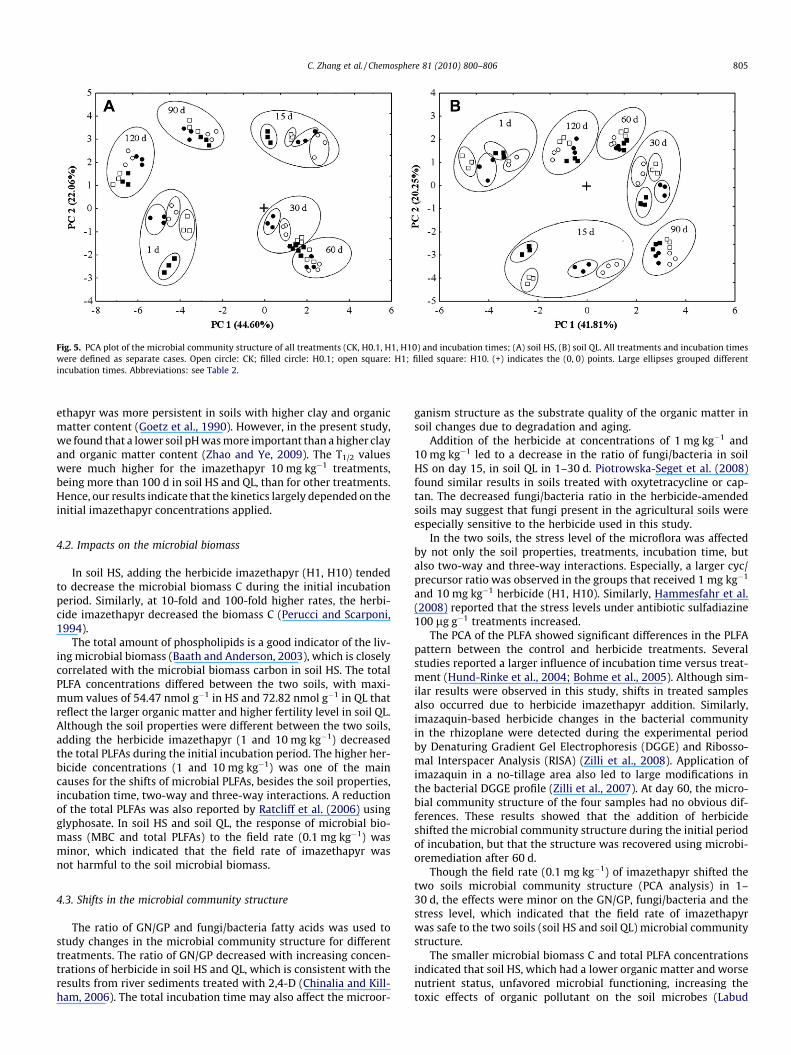
Fig. 5. PCA plot of the microbial community structure of all treatments (CK, H0.1, H1, H10) and incubation times; (A) soil HS, (B) soil QL. All treatments and incubation timeswere defined as separate cases. Open circle: CK; filled circle: H0.1; open square: H1; filled square: H10. (+) indicates the (0, 0) points. Large ellipses grouped differentincubation times. Abbreviations: see Table 2.
C. Zhang et al. / Chemosphere 81 (2010) 800–806 805
ethapyr was more persistent in soils with higher clay and organicmatter content (Goetz et al., 1990). However, in the present study,we found that a lower soil pH was more important than a higher clayand organic matter content (Zhao and Ye, 2009). The T1/2 valueswere much higher for the imazethapyr 10 mg kg�1 treatments,being more than 100 d in soil HS and QL, than for other treatments.Hence, our results indicate that the kinetics largely depended on theinitial imazethapyr concentrations applied.
4.2. Impacts on the microbial biomass
In soil HS, adding the herbicide imazethapyr (H1, H10) tendedto decrease the microbial biomass C during the initial incubationperiod. Similarly, at 10-fold and 100-fold higher rates, the herbi-cide imazethapyr decreased the biomass C (Perucci and Scarponi,1994).
The total amount of phospholipids is a good indicator of the liv-ing microbial biomass (Baath and Anderson, 2003), which is closelycorrelated with the microbial biomass carbon in soil HS. The totalPLFA concentrations differed between the two soils, with maxi-mum values of 54.47 nmol g�1 in HS and 72.82 nmol g�1 in QL thatreflect the larger organic matter and higher fertility level in soil QL.Although the soil properties were different between the two soils,adding the herbicide imazethapyr (1 and 10 mg kg�1) decreasedthe total PLFAs during the initial incubation period. The higher her-bicide concentrations (1 and 10 mg kg�1) was one of the maincauses for the shifts of microbial PLFAs, besides the soil properties,incubation time, two-way and three-way interactions. A reductionof the total PLFAs was also reported by Ratcliff et al. (2006) usingglyphosate. In soil HS and soil QL, the response of microbial bio-mass (MBC and total PLFAs) to the field rate (0.1 mg kg�1) wasminor, which indicated that the field rate of imazethapyr wasnot harmful to the soil microbial biomass.
4.3. Shifts in the microbial community structure
The ratio of GN/GP and fungi/bacteria fatty acids was used tostudy changes in the microbial community structure for differenttreatments. The ratio of GN/GP decreased with increasing concen-trations of herbicide in soil HS and QL, which is consistent with theresults from river sediments treated with 2,4-D (Chinalia and Kill-ham, 2006). The total incubation time may also affect the microor-
ganism structure as the substrate quality of the organic matter insoil changes due to degradation and aging.
Addition of the herbicide at concentrations of 1 mg kg�1 and10 mg kg�1 led to a decrease in the ratio of fungi/bacteria in soilHS on day 15, in soil QL in 1–30 d. Piotrowska-Seget et al. (2008)found similar results in soils treated with oxytetracycline or cap-tan. The decreased fungi/bacteria ratio in the herbicide-amendedsoils may suggest that fungi present in the agricultural soils wereespecially sensitive to the herbicide used in this study.
In the two soils, the stress level of the microflora was affectedby not only the soil properties, treatments, incubation time, butalso two-way and three-way interactions. Especially, a larger cyc/precursor ratio was observed in the groups that received 1 mg kg�1
and 10 mg kg�1 herbicide (H1, H10). Similarly, Hammesfahr et al.(2008) reported that the stress levels under antibiotic sulfadiazine100 lg g�1 treatments increased.
The PCA of the PLFA showed significant differences in the PLFApattern between the control and herbicide treatments. Severalstudies reported a larger influence of incubation time versus treat-ment (Hund-Rinke et al., 2004; Bohme et al., 2005). Although sim-ilar results were observed in this study, shifts in treated samplesalso occurred due to herbicide imazethapyr addition. Similarly,imazaquin-based herbicide changes in the bacterial communityin the rhizoplane were detected during the experimental periodby Denaturing Gradient Gel Electrophoresis (DGGE) and Ribosso-mal Interspacer Analysis (RISA) (Zilli et al., 2008). Application ofimazaquin in a no-tillage area also led to large modifications inthe bacterial DGGE profile (Zilli et al., 2007). At day 60, the micro-bial community structure of the four samples had no obvious dif-ferences. These results showed that the addition of herbicideshifted the microbial community structure during the initial periodof incubation, but that the structure was recovered using microbi-oremediation after 60 d.
Though the field rate (0.1 mg kg�1) of imazethapyr shifted thetwo soils microbial community structure (PCA analysis) in 1–30 d, the effects were minor on the GN/GP, fungi/bacteria and thestress level, which indicated that the field rate of imazethapyrwas safe to the two soils (soil HS and soil QL) microbial communitystructure.
The smaller microbial biomass C and total PLFA concentrationsindicated that soil HS, which had a lower organic matter and worsenutrient status, unfavored microbial functioning, increasing thetoxic effects of organic pollutant on the soil microbes (Labud

806 C. Zhang et al. / Chemosphere 81 (2010) 800–806
et al., 2007). However, the results showed that the effects of theherbicide imazethapyr on the microbial biomass (microbial bio-mass C and total PLFA concentrations), microbial community(GN/GP and fungi/bacteria), stress level, and PCA of PLFA in soilHS had patterns similar to that of soil QL. These findings may havebeen because soil HS has a stronger adsorption for imazethapyr,which reduced the effective amount of imazethapyr in soil.
Soil microorganisms can be studied in cultured communities orin whole communities. Generally, the former is performed on cul-turable microbes with the soil dilution plate method. The latter isused to assess changes in the entire microbial community struc-ture by determination of PLFA (Kong et al., 2008). In this study,PLFA analysis was used to supply not only the total biomass of soilmicrobes, but also the shifts of the entire microbial communitystructure (GN/GP, fungi/bacteria, the stress level, and PCA analy-sis). At the same time, the more accurate information on soilmicrobial biomass was supplied by PLFA analysis than that ofMBC. The shifts in the soil microbial community observed in thisstudy were confirmed by field trials (another experiment),although the PLFA profile data was not completely concordant.Further studies will use PLFA analysis combined with other meth-ods, such as community-level physiological profiles (CLPPs) andamplified ribosomal DNA restriction analysis (ARDRA), to providemore comprehensive, exact information regarding the effect ofthe herbicide on the structure and function of microbial communi-ties and the implications on ecosystem-level processes.
5. Conclusions
The application of imazethapyr led to a shift in the soil commu-nity structure during the initial period of incubation. Imazethapyrwas degraded slowly in the two soils, especially in the soil HS withthe lower pH. Soil microbial biomass C and total concentration ofPLFA were variable with incubation time, which were also reducedby the addition of imazethapyr. In the two soils, the ratio of GN/GP,the ratio of fungi/bacteria and the stress level of the microflorawere affected by not only the soil properties, treatments, incuba-tion time, but also two-way and three-way interactions. Imazeth-apyr addition also decreased the ratios of GN/GP and fungi/bacteria. A larger stress level, measured as the ratio of PLFA (cy-c17:0 + cyc19:0)/(16:1x7c + 18:1x7c), was found in the high con-centration (1 and 10 mg kg�1) herbicide treatment groups. Theeffects of imazethapyr at the field application on soil microbial bio-mass and microbial community were minor. PCA of the PLFAshowed that imazethapyr altered the soil microbial communitystructure. These findings stress that there is a high need for furtherinvestigations to increase our understanding of the interactions ofherbicide and the soil microbial community structure.
Acknowledgements
This work was supported by the National Key Technology R&DProgram (Grant No. 2006BAD08A03), Science and Technology Pro-ject for Food Production (2006BAD02A16) and National Key Projectof Fundamental Research (The 973 Program, Grant No.2009CB119006).
References
Baath, E., Anderson, T.H., 2003. Comparison of soil fungi/bacterial ratios in a pHgradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 35,955–963.
Bardgett, R.D., Hobbs, P.J., Frostegard, A., 1996. Changes in soil fungal: bacterialratios following reductions in the intensity of management of an uplandgrassland. Biol. Fertil. Soils 22, 261–264.
Battaglin, W.A., Furlong, E.T., Burkhardt, M.R., Peter, C.J., 2000. Occurrence ofsulfonylurea, sulphonamide, imidazolinone, and other herbicides in rivers,
reservoirs and ground water in the Midwestern United States, 1998. Sci. TotalEnviron. 248, 123–133.
Bohme, L., Langer, U., Bohme, F., 2005. Microbial biomass, enzyme activities andmicrobial community structure in two European long-term field experiments.Agr. Ecosyst. Environ. 109, 141–152.
Bossio, D.A., Scow, K.M., 1998. Impacts of carbon and flooding on soil microbialcommunities: phospholipid fatty acid profiles and substrate utilisationpatterns. Microb. Ecol. 35, 265–278.
Bossio, D.A., Scow, K.M., Gunapala, N., Graham, K.J., 1998. Determinants of soilmicrobial communities: effect of agricultural management, season, and soiltype on phospholipid fatty acid profiles. Microb. Ecol. 36, 1–12.
Chinalia, F.A., Killham, K.S., 2006. 2,4-Dichlorophenoxyacetic acid (2,4-D)biodegradation in river sediments of northeast-Scotland and its effect on themicrobial communities (PLFA and DGGE). Chemosphere 64, 1675–1683.
Donald, R.Z., George, W.K., 2006. Microbial community composition and functionacross an arctic tundra landscape. Ecology 87, 1659–1670.
Frostegard, A., Baath, E., Tulid, A., 1993. Shifts in the structure of soil microbialcommunities in limes forests as revealed by phospholipids fatty-acid analysis.Soil Biol. Biochem. 25, 723–730.
Goetz, A.J., Lavy, T.L., Gbur, E.E., 1990. Degradation and field persistence ofimazethapyr. Weed Sci. 38, 421–428.
Hammesfahr, U., Heuer, H., Manzke, B., Smalla, K., Bruhn, S.T., 2008. Impact of theantibiotic sulfadiazine and pig manure on the microbial community structure inagricultural soils. Soil Biol. Biochem. 40, 1583–1591.
Hollaway, K.L., Kookana, R.S., Noy, D.M., Smith, J.G., Wilhelm, N., 2006. Persistenceand leaching of imazethapyr and flumetsulam herbicides over a 4-year periodin the highly alkaline soils of south-eastern Australia. Aust. J. Exp. Agr. 46, 669–674.
Hund-Rinke, K., Simon, M., Lukow, T., 2004. Effects of tetracycline on the soilmicroflora: function, diversity, resistance. J. Soil Sediment. 4, 11–16.
Kong, C.H., Wang, P., Zhao, H., Xu, X.H., Zhu, Y.D., 2008. Impact of allelochemicalexuded from allelopathic rice on soil microbial community. Soil Biol. Biochem.40, 1862–1869.
Labud, V., Garcia, C., Hernandez, T., 2007. Effect of hydrocarbon pollution on themicrobial properties of a sandy and a clay soil. Chemosphere 66, 1863–1871.
Li, W.Z., Fan, D.S., Luan, Y.B., 2008. Current situation, problems and solutions ofethametsulfuron, chlorimuron-ethyl and imazethapyr. Agrochemicals 47, 781–789 (in Chinese).
Lin, Q.M., Wu, Y.G., Liu, H.L., 1999. Modification of fumigation extraction method formeasuring soil microbial biomass carbon. Chin. J. Ecol. 18, 63–66 (in Chinese).
Loux, M.M., Reese, K.D., 1993. Effect of soil type and pH on persistence andcarryover of imidazolinone herbicides. Weed Technol. 7, 452–458.
Lupwayi, N.Z., Harker, K.N., Clayton, G.W., Turkington, T.K., Rice, W.A., O’Donovan,J.T., 2004. Soil microbial biomass and diversity after herbicide application. Can.J. Plant Sci. 84, 677–685.
Ma, J.W., Chen, L., Wang, J.F., Jiang, G.P., Pan, C.P., 2007. Determination ofimazethapyr residues in soil by SPE–HPLC. Agrochemicals 46, 843–845 (inChinese).
Perucci, P., Scarponi, L., 1994. Effects of the herbicide imazethapyr on soil microbialbiomass and various soil enzyme activities. Biol. Fertil. Soils 17, 237–240.
Perucci, P., Dumontet, S., Bufo, S.A., Mazzatura, A., Casucci, C., 2000. Effects oforganic amendment and herbicide treatment on soil microbial biomass. Biol.Fertil. Soils 32, 17–23.
Piotrowska-Seget, Z., Engel, R., Nowak, E., Kozdroj, J., 2008. Successive soiltreatment with captan or oxytetracycline affects non-target microorganisms.World J. Microbiol. Biotechnol. 24, 2843–2848.
Ratcliff, A.W., Busse, M.D., Shestak, C.J., 2006. Changes in microbial communitystructure following herbicide (glyphosate) additions to forest soils. Appl. SoilEcol. 34, 114–124.
Renner, K.A., Meggitt, W.F., Penner, D., 1988. Effect of soil pH imazaquin andimazethapyr adsorption to soil and phototoxicity to corn (Zea mays). Weed Sci.36, 78–83.
Stougaard, R.N., Shwa, P.J., Martin, A.R., 1990. Effect of soil type and pH onadsorption, mobility, and efficacy of imazaquin and imazethapyr. Weed Sci. 38,67–73.
Wang, Y.P., Shi, J.Y., Wang, H., Qi, L., Chen, X.C., Chen, Y.X., 2007. The influence of soilheavy metals pollution on soil microbial biomass, enzyme activity, andcommunity composition near a copper smelter. Ecotoxicol. Environ. Safety 67,75–81.
Wang, M.C., Liu, Y.H., Wang, Q., Gong, M., Hua, X.M., Pang, Y.J., Hu, S.J., Yang, Y.H.,2008. Impacts of methamidophos on the biochemical, catabolic, and geneticcharacteristics of soil microbial communities. Soil Biol. Biochem. 40, 778–788.
Wu, J.S., Lin, Q.M., Huang, Q.Y., Xiao, H.A., 2006. Soil Microbial Biomass – Methodsand Application. Weather Press, Beijing, China.
Zabalza, A., Gaston, S., Sandalio, L.M., Rio, L.A., Royuela, M., 2007. Oxidative stress isnot related to the mode of action of herbicides that inhibit acetolactatesynthase. Environ. Exp. Bot. 59, 150–159.
Zhao, S., Ye, F., 2009. Application of imidazolinone herbicide and its degradation.Plant Prot. 35, 15–19 (in Chinese).
Zilli, J.E., Smiderle, O.J., Neves, M.C.P., Rumjanek, N.G., 2007. Microbial population insoil cultivated with soybean and treated with different herbicides in cerradoarea of Roraima. Acta Amaz. 37, 201–212.
Zilli, J.E., Botelho, G.R., Neves, M.C.P., Rumjanek, N.G., 2008. Effect of glyphosate andimazaquin on the soybean rhizoplane bacterial community and microbiologicalsoil characteristics. Rev. Bras. Cienc. Solo 32, 633–642.



















