
IL-2 signals through Sgk1 and inhibits proliferation and apoptosis in kidney cancer cells
15
ORIGINAL ARTICLE IL-2 signals through Sgk1 and inhibits proliferation and apoptosis in kidney cancer cells Rosario Amato & Miranda Menniti & Valter Agosti & Rosalia Boito & Nicola Costa & Heather M. Bond & Vito Barbieri & Pierosandro Tagliaferri & Salvatore Venuta & Nicola Perrotti Received: 29 January 2007 / Revised: 14 March 2007 / Accepted: 16 April 2007 / Published online: 15 June 2007 # Springer-Verlag 2007 Abstract The interleukin-2 is a cytokine that is essential for lymphocytic survival and function. Ectopic expression of the IL-2 receptor in epithelial tissues has been reported previously, although the functional significance of this expression is still being investigated. We provided novel structural and functional information on the expression of the IL-2 receptor in kidney cancer cells and in other normal and neoplastic human epithelial tissues. In A-498 kidney cancer cells, we showed that IL-2 binding to its own receptor triggers a signal transduction pathway leading to the inhibition of proliferation and apoptosis. We found that the inhibition of proliferation is associated with Erk1/2 dephosphorylation, whereas the survival signals appear to be mediated by Sgk1 activation. This investigation focuses on the IL-2 induced regulation of Sgk1 and describes a role of the IL-2 receptor and Sgk1 in the regulation of epithelial tumor cell death and survival. Keywords Interleukin-2 . Sgk1 . Kidney cancer cells . Cell proliferation . Apoptosis Abbreviations IL-2 Interleukin-2 IL-2R Interleukin-2 Receptor Sgk1 Serum and glucocorticoid regulated kinase Erk1/2 Extracellular regulated kinase 1/2 Shc-1 Src homology 2 domain containing J Mol Med (2007) 85:707–721 DOI 10.1007/s00109-007-0205-2 Electronic supplementary material The online version of this article (doi:10.1007/s00109-007-0205-2) contains supplementary material, which is available to authorized users. In loving memory of Salvatore Venuta, 1944–2007. R. Amato : M. Menniti : V. Agosti : R. Boito : H. M. Bond : V. Barbieri : P. Tagliaferri : S. Venuta : N. Perrotti (*) Department of Experimental and Clinical Medicine “G. Salvatore”, University “Magna Graecia”, Campus Biomedico, Località Germaneto, Viale Europa, Catanzaro 88100, Italy e-mail: [email protected] N. Costa Department of Pharmacobiology, University “Magna Graecia”, Catanzaro, Italy ROSARIO AMATO he received his medical degree from the University of Catanzaro (UMG) in Italy. He is presently a Ph.D. Student in molecular on- cology and experimental immu- nology at the University of Catanzaro. His research interests include cell signaling and the molecular mechanisms of drug resistance in neoplastic cells. NICOLA PERROTTI he studied medicine at the Uni- versity of Napoli (Italy), trained in Internal Medicine, Diabetes and Metabolic Diseases, worked as a visiting scientist at the diabetes branch, NIDDK, NIH, Bethesda, USA. He is now pro- fessor of General Pathology at the University of Catanzaro. Re- search interests include the mo- lecular basis of insulin signal transduction in the regulation of cell metabolism, survival and proliferation.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of IL-2 signals through Sgk1 and inhibits proliferation and apoptosis in kidney cancer cells

ORIGINAL ARTICLE
IL-2 signals through Sgk1 and inhibits proliferationand apoptosis in kidney cancer cells
Rosario Amato & Miranda Menniti & Valter Agosti &Rosalia Boito & Nicola Costa & Heather M. Bond &
Vito Barbieri & Pierosandro Tagliaferri &Salvatore Venuta & Nicola Perrotti
Received: 29 January 2007 /Revised: 14 March 2007 /Accepted: 16 April 2007 / Published online: 15 June 2007# Springer-Verlag 2007
Abstract The interleukin-2 is a cytokine that is essentialfor lymphocytic survival and function. Ectopic expressionof the IL-2 receptor in epithelial tissues has been reportedpreviously, although the functional significance of thisexpression is still being investigated. We provided novelstructural and functional information on the expression ofthe IL-2 receptor in kidney cancer cells and in other normaland neoplastic human epithelial tissues. In A-498 kidneycancer cells, we showed that IL-2 binding to its ownreceptor triggers a signal transduction pathway leading tothe inhibition of proliferation and apoptosis. We found thatthe inhibition of proliferation is associated with Erk1/2dephosphorylation, whereas the survival signals appear tobe mediated by Sgk1 activation. This investigation focuseson the IL-2 induced regulation of Sgk1 and describes a roleof the IL-2 receptor and Sgk1 in the regulation of epithelialtumor cell death and survival.
Keywords Interleukin-2 . Sgk1 . Kidney cancer cells .
Cell proliferation . Apoptosis
AbbreviationsIL-2 Interleukin-2IL-2R Interleukin-2 ReceptorSgk1 Serum and glucocorticoid regulated kinaseErk1/2 Extracellular regulated kinase 1/2Shc-1 Src homology 2 domain containing
J Mol Med (2007) 85:707–721DOI 10.1007/s00109-007-0205-2
Electronic supplementary material The online version of this article(doi:10.1007/s00109-007-0205-2) contains supplementary material,which is available to authorized users.
In loving memory of Salvatore Venuta, 1944–2007.
R. Amato :M. Menniti :V. Agosti :R. Boito :H. M. Bond :V. Barbieri : P. Tagliaferri : S. Venuta :N. Perrotti (*)Department of Experimental and Clinical Medicine“G. Salvatore”, University “Magna Graecia”,Campus Biomedico, Località Germaneto, Viale Europa,Catanzaro 88100, Italye-mail: [email protected]
N. CostaDepartment of Pharmacobiology, University “Magna Graecia”,Catanzaro, Italy
ROSARIO AMATO
he received his medical degreefrom the University of Catanzaro(UMG) in Italy. He is presently aPh.D. Student in molecular on-cology and experimental immu-nology at the University ofCatanzaro. His research interestsinclude cell signaling and themolecular mechanisms of drugresistance in neoplastic cells.
NICOLA PERROTTIhe studied medicine at the Uni-versity of Napoli (Italy), trainedin Internal Medicine, Diabetesand Metabolic Diseases, workedas a visiting scientist at thediabetes branch, NIDDK, NIH,Bethesda, USA. He is now pro-fessor of General Pathology atthe University of Catanzaro. Re-search interests include the mo-lecular basis of insulin signaltransduction in the regulation ofcell metabolism, survival andproliferation.

Introduction
Interleukin-2 (IL-2) regulates lymphocyte proliferation anddifferentiation and plays a critical role in the immuneresponse, especially in the cellular-mediated responseagainst viruses and neoplastic cells [1, 2]. The binding ofIL-2 to its own receptor is the first step in the action of IL-2.The receptor is composed of at least three subunits: IL-2receptor alpha (IL-2RA), beta (IL-2RB), and gamma (IL-2RG) chains. The alpha chain (CD25) is a unique andspecific marker for the IL-2 receptor, whereas the beta chain(CD122) is common to both the IL-2 and IL-15 receptors,and the gamma chain (p64-CD132) is common to allcytokine receptors. Homodimeric alpha chains form a low-affinity receptor, while homodimeric beta chains produce amedium-affinity receptor and heterodimeric alpha and betachains constitute a high-affinity receptor [3].
The usefulness of IL-2 in the immunotherapy of kidneycarcinomas, melanomas, and metastatic cancer is consoli-dated in clinical oncology, although it is still unclearwhether IL-2 acts against specific neoplastic antigensthrough lymphocyte activation only and/or with a directsynergistic action on epithelial neoplastic cells, interactingwith a specific or related receptor. In this study, wedemonstrate for the first time the presence of the IL-2receptor in multiple epithelial tumor cells and normalhuman tissue. It is then possible that binding of IL-2 to itsown receptor might activate cell proliferation and survivalsignals, thus affecting the results of IL-2 immunotherapy ofcarcinomas. We demonstrate that IL-2 inhibits proliferationin kidney cancer cells downregulating the Erk 1-2 path-ways. IL-2 has been demonstrated to promote cell survivalby activating Akt in lymhoid cells [4].
Focusing on a renal cancer cell model, we asked whetherthe expression of the IL-2 receptor on epithelial cells mighthave consequences on cell survival. PI3-K-dependentsurvival signals can be mediated by either Akt or Sgk(serum and glucocorticoid regulated kinase), important anti-apoptotic effectors that inactivates the pro-apoptotic pro-teins Bad and Forkhead [5]. We show that the IL-2/IL-2receptor complex signals through PI3-K and that IL-2activates Sgk1, but not Akt at the posttranslational level andactivates Sgk1 at the transcriptional level.
Sgk1 is a serine/threonine protein kinase, approximately50% homologous to protein kinase B (Akt/PKB). It isregulated at a posttranslational level by insulin [6] serum,and cyclic-adenosine monophosphate (cAMP) [7] throughthe phosphoinositide-dependent kinases 1 and 2 (PDK1/PDK2) that phosphorylate Thr 256 and Ser 422, respec-tively [8]. Sgk1 is also regulated by steroids at thetranscriptional level and regulates enzymes in cell metab-olism [9]. In fact, Sgk1 is a hyperosmotic stress-inducedgene [10] and more, the promoter of Sgk1 is under tight
control of the onco-supperpressor protein p53 [11]. Morerecently, Sgk1 has been shown to mediate cell survival anddrug resistance to platinoid and taxani compounds in womenwith breast cancer pretreated with glucocorticoids [12].
We demonstrate that IL-2 inhibits proliferation andapoptosis in A-498 cells kidney cancer cells. In addition,we provide evidence that the activation of Sgk1 isnecessary for the IL-2-dependent inhibition of caspase8 activation observed on A-498 cells. These data, combinedwith IL-2’s action on lymphocytic cells, underscores thenecessity for a review of the role of IL-2 in rationaleclinical use and a differentiation marker.
Materials and methods
The information related to the reagents employed, theconstructs and the cell culture conditions, including thebinding competition experiments, are described inthe supplementary files, together with the methods andthe legends of the experiments presented in the supplemen-tary files.
Cell stimulation and inhibitors treatment
Cell cultures were serum-starved 24 h before stimulationwith any effector, unless otherwise specified in the figurelegends. LY294002 (50 μM, Sigma-Aldrich, Milan Italy)was added to A-498 cells 30 min before every stimulation.Il-2 and/or insulin were used at doses and times, asindicated in the figure legends.
The transfections of Sgk1-specific siRNA (Darmachon,Upstate) and dominant negative Sgk1 (D222A) wereperformed by Lipofectamine 2000™ (Invitrogen), accord-ing to the instructions of the manufacturer. For the caspase8 assay, doxorubicin (0.1 μg/ml) was added for 12 h to cellspretreated or not, for 2 h, with IL-2 (1.0×10−8 M), in thepresence or absence of the dominant negative Sgk1(D222A). To evaluate the Erk1/2 phosphorylation, the cellswere exposed for 30 min to 1×10−8 M of IL-2 in thepresence or absence of dominant negative for Sgk1.
Western blot, immunoprecipitation, and in vitrokinase assay
Western Blot Cell extracts of A-498 cells were preparedand immunoblotted as described [4]. Rabbit anti-phosphoSer422 Sgk1 (Santa Cruz) immunoglobulins were diluted in3% bovine serum albumin (BSA) in T-PBS, whereas anti-total-Sgk1 immunoglobulins were diluted in 5% non-fatdried milk in T-PBS. When phospho-specific anti-Sgk1antibodies were used, cell extracts were pre-cleared byincubation at 4°C for 120 min with normal rabbit
708 J Mol Med (2007) 85:707–721

immunoglobulins pre-bound to protein G-UltraLink (15 μl;Pierce Rockford, IL).
Immunoprecipitation and in vitro kinase assay Cells werestimulated by insulin or IL-2 as indicated in the figurelegends. Cell monolayers were then solubilized and analiquot (450 μl) was immunoprecipitated with rabbit anti-Sgk1 antibody (7 μl) for the immune complex kinase assayusing the artificial substrate peptide Arg-Pro-Arg-Thr-Ser-Thr-Phe (1 mM), as previously reported [7]. Sgk1 activitywas calculated by subtracting the basal level incorporationof 32P catalyzed by immunoprecipitates of non-stimulatedA-498. To average data from multiple experiments, the datawere expressed as a percentage of the mean Sgk1 activitymeasured in cells incubated in the presence of IL-2. Dataare means ± SE of duplicate measurements obtained in sixindependent experiments.
Semiquantitative reverse transcriptase polymerase chainreaction (RT-PCR) cDNA was synthesized from 1 μg totalcellular RNA using SuppererScript III Rnase H-reversetranscriptase (Invitrogen) and 2.5 μM random hexamers(invitrogen). cDNA aliquots were amplified by PCR (ABapplied biosystem) for 25 cycles using ReadyMix™ TaqPCR reaction Mix with MgCl2 (Sigma), in the presence ofprimers specific for exons 1 and 4 of Sgk1 (Fwd ATGACGGTGAAAACTGAGGCT,RevTGCCACAGAAGGTGGATGTT).
The relative amounts of the amplified products in theabsence and presence of IL-2 were compared with theamounts of the amplified products obtained by PCR (25cycles) in the presence of primers specific for glyceraldehydesphosphate dehydrogenase (GAPDH). (FwdCACCATCTTCCAGGAGCGAG, Rev TCACGCCACAGTTTCCCGGA).The amount of amplified product was evaluated by scanningdensitometry (NIH Image).
Immunofluorescence A-498 cells grown on coverslips werefixed with 3.7% formaldehyde for 20 min. Fixed cells werethen permeabilized with 0.5% Triton ×-100 for 1 min andwashed with PBS (pH 7.4). The prepared samples wereincubated with primary antibody in PBS (pH 7.4) contain-ing BSA (1 mg/ml) and Tween-20 (0.2%) for 2 h at roomtemperature. Rabbit anti-CD25-FITC conjugated (Serotec,UK), rabbit anti-Sgk1 (Upstate), and rabbit anti-caspase8 (Santa Cruz) antibodies were diluted 200 times in PBS/BSA. Samples were washed with PBS and incubated, whenneeded, with goat anti-rabbit immunoglobulins coupperledto FITC (Alexa fluor, Molecular probe-Invitrogen). Sam-ples were visualized using a Leica TC SP2 microscope witha ×63 objective and processed with Leica confocalsoftware.
D222A (Sgk1 dominant negative) continous cell line HeLacells were transfected with either pcDNA4TO D222ASgk1or pcDNA4TO empty vector by Lipofectamine 2000(Invitrogen) according to the manufacturer’s instructions.Forty-eight hours after transfection cells were selectedbased on their ability to grow in the presence of Zeocin(400 μg/ml). A total of 102 independent clones wereisolated. Fifty clones were pooled and the transgeneexpression was verified by Western blotting with mycimmunoglobulins, RT-PCR, and immunofluorescence (datanot shown).
Proliferative assay Propidium iodide staining was used toevaluate the effect of IL-2 on cell cycle progression. Serumstarved A-498 cells were treated for 24 h with IL-2, asindicated. Cells, fixed in ethanol 70% and kept overnight at−20°C, were incubated for 1 h at room temperature in PBScontaining propidium iodide (20 γ/ml), NP40 0.1% andribonuclease 40 γ/ml, washed and subjected to fluorescenceactivated cell sorter (FACS) analysis by FACScan©
cytometer (Becton Dickinson) using CellQuest software.The cell doublets were excluded by the FL2A-FL2W gatingmethod and the fluorescence intensity was recorded on anFL2A histogram.
Cell proliferation was evaluated by measuring the totalnumber of cells with a Burker chamber. Cellular viabilitywas monitored by staining with trypan blue. A-498 cellswere plated at a density of 5×104 cells/ml in six-wellplates. After 24 h of serum starvation, cells were incubatedin the absence and in the presence of increasing doses ofIL-2, as indicated. Forty-eight hours later the cells werewashed in cold PBS and trypsinized; 0.1 ml from each ofthe three treatments were removed, 0.1 ml of trypan bluewas added and stained, and unstained cells were counted. Asimilar procedure was followed when proliferation wasassayed in A-498 cells transfected with Sgk1 specific andscrambled siRNAs (Darmachon–Upstate). Each determina-tion of total cell number was performed on samples pooledfrom two wells in a six-well plate. The mean of fourcounting of the cells in the presence of either IL-2 or 10%serum was expressed as a percentage of the number of cellsstained in basal condition.
Apoptosis
Hypodiploid DNA analysis Apoptosis was detected ashypodiploid DNA peak and measured as the percentageof hypodiploid cells over the total number of A-498 cell.The hypodiploid peak was measured in 72-h serum-starvedcells, in cells grown in the presence of either fetal bovineserum (FBS) 10% or IL-2 (1.0×10−8 M). Cells werewashed in PBS 1× (without MgCl2 and CaCl2) andincubated 1 h at 4°C in the presence of propidium iodide
J Mol Med (2007) 85:707–721 709

staining (PI). Cells were washed and analyzed by FACScan©
cytometer (Becton Dickinson) using CellQuest software. Thecell doublets were excluded by the FL2A-FL2W gatingmethod and the fluorescence intensity was recorded on anFL2A histogram. Hypodiploid cells were counted asapoptotic cells.
Oligonucleosomal DNA fragmentation
Detection of DNA laddering by gel electrophoresis. DNAwasextracted from cultures of control and doxorubicin treatedA-498 cells in the absence and in the presence of IL-2 (1.5×10−8 M). Cells were lysed in 400 μl of Apo lysis buffer(10 mMTris–HCl [pH 7.4], 10 mM EDTA, 0.2% Triton×-100), and then processed as previously reported [13].
Animal procedure
Male (C57BL/6) mice at 9 weeks of age were treated witheither 10,000 units (3.4×10−8 M) of IL-2 or diluent (PBS),by subcutaneous injection, and killed after 3 days. Kidneyand liver of IL-2 stimulated and control groups weresolubilized in lysis buffer. Samples were separated bySDS-PAGE, transferred to polyvinylidene difluoride(PVDF) (Amersham) and immunoblotted with rabbit anti-phospho Ser422 Sgk1 immunoglobulins (Santa Cruz), andreprobed with rabbit anti-Sgk1 immunoglobulins (Upstate).
Results
IL-2 receptor beta chain is expressed in neoplasticand normal human tissues
We performed a profile expression for the IL-2 receptorbeta chain on a pre-made, normalized array containingnormal and neoplastic human tissues. The beta chainmessage was detected in several neoplastic tissue analyzed,as expected [14], with highest signals in rectum, ovary,stomach, kidney, colon, and breast tumors. The beta chaintranscript was also detected in the majority of normaltissues analyzed, including liver, salivary gland, fetalkidney, heart, skeletal muscle, stomach, duodenum andjejunum, nucleus accumbens, parietal lobe, and cerebrumareas (see supplementary data 1 for the images).
The IL-2 receptor is expressed in a renal carcinoma cell line
To define the receptor and post-receptor events involved inIL-2 signaling, we elected to study A-498 human kidneycarcinoma cells. Binding competition experiments to intactcells in culture suggested that the A-498 cells possess an
IL-2 receptor that contains two binding sites—one withhigh affinity and the other with medium-low affinity.Binding competition curves revealed allosteric propertiesfor the interleukin 2 receptor, suggesting intersubunitnegative cooperativity of ligand binding, at low interleukin2 concentrations [15]. A scatchard plot analysis of thebinding assay for the medium-low affinity componentrevealed a specific binding for the IL-2 receptor, compara-ble to what has been described for a lymphocyte cell line.In addition, we calculated an approximate number of 6.0×10−9 receptor sites per cell (Fig. 1a, b).
The presence of alpha, beta, and gamma chains of theIL-2 receptor was demonstrated in A-498, HEK293T, andJurkat cells by RT-PCR. The message coding for the betachain, the main transductive subunit of this receptorialcomplex, was found in the three cell types (Fig. 2a); themessage coding for the gamma chain was also found in the
Fig. 1 a Binding competition of 125I-IL-2 in A-498 cells. 2.0×106
A-498 cells were incubated at 4°C for 12 h in HEPES binding bufferwith radiolabeled IL-2 (0.1 ng/ml) in the absence or in the presence ofincreasing concentrations of unlabeled ligand (6.6×10−9, 1.0×10−8,1.5×10−8, 2.3×10−8, 3.6×10−8, 5.0×10−8, 6.3×10−8, 8.3×10−8, and8.3×10−7 M). Specific binding was expressed as a percent of thebinding capacity in the absence of the cold ligand. b Scatchard plot of125I-IL-2 binding. Scatchard analysis of the IL-2 binding for thesecond component of the binding curve (concentration unlabeled IL-2ranging from 3.6×10−8 to 8.3×10−7 M)
710 J Mol Med (2007) 85:707–721

three cell types, whereas the message coding for the alphachain was found only in A-498 and Jurkat cells (Fig. 2a).The message coding for the beta chain of the IL-2 receptorwas also detected, in A-498 and HEK293T cells, byNorthern blot analysis (data not shown). The sequence ofthe amplified products corresponding to the beta subunit ofthe IL-2 receptor was confirmed by automatic sequencing(data not shown). As the A-498 cells have been extensivelyused in the present paper, the expression of the alpha andgamma subunits have been further verified by immuno-fluorescence and Western blotting. Interestingly, the alphachain of the IL-2 receptor was detected by immunofluores-cence, only in the presence of IL-2 (Fig. 2b). Theexpression of the gamma chain of the IL-2 receptor inA-498 also increased as time progressed under IL-2stimulation, as assessed by Western blotting of gammachain immunoprecipitates (Fig. 2c). The gamma chain ofthe IL-2 receptor in A-498 cells migrates with an apparent
molecular weight of 64 kDa, as described in the literaturefor one of the isoforms of the IL-2 receptor gamma chain.
IL-2 has an anti-proliferative effect on A-498 cells
To understand the downstream effects IL-2 binding to itsown receptor, we investigated the involvement of IL-2 incell proliferation.
A growth rate analysis showed that IL-2, in A-498carcinoma cells, caused a dose-dependent anti-proliferativeeffect (Fig. 3a). Forty-eight after plating 10×104 cells, wecounted 70×104±80,000 cells, in serum-treated samples;52×104±90,000 cells, in serum-starved samples; 42×104±100,000 and 22.5×104±92,000 cells in samples treatedwith low dose and high dose of interleukin, respectively.FACS analysis of propidium-iodide-stained cells showedthat treatment with IL-2 decreased the percentage of cells in
Fig. 2 a Lower gel: IL-2Rβ chain mRNA expression in variouscancer cells. IL-2R beta chain mRNA was detected by RT-PCR indifferent cancer cell lines: lymphoid Jurkat cells (lane 1), HEK293T(lane 2), A-498 (lane 3). The fragment size is around 490 bp, asexpected. Upper gel: IL-2R alpha and gamma chain mRNAexpression detected by RT-PCR in various cancer cells: A-498 (lane1), lymphoid Jurkat cells (lane 2), HEK293T (lane 3). b Localizationof IL-2Rα chain (CD25) in A-498 cells. Low confluent monolayers ofserum-starved A-498 cells, grown on glass coverslips, were stimulatedwith IL-2 (1.0×10−8 M) for 4 h. Fields A, B, C, D, and E show cells
positive for CD25 detected with rabbit anti-CDC25 flouresceinisothiocyanate (FITC) immunoglobulins. Field F shows a controlimmunofluorescence detected with goat anti-rabbit FITC immuno-globulins. Field G shows the immunofluorescence detected with rabbitanti-CDC25 FITC immunoglobulins, in the absence of IL-2 stimula-tion. c Effect of IL-2 on IL-2R gamma chain expression. Serum-starved A-498 cells were stimulated with human IL-2 (1.0×10−8 M)for 0 min (lane 1), 60 min (lane 2), and 90 min (lane 3). The arrowindicates immunoprecipitated IL-2Rγ chain probed by Westernblotting with specific antibodies
J Mol Med (2007) 85:707–721 711
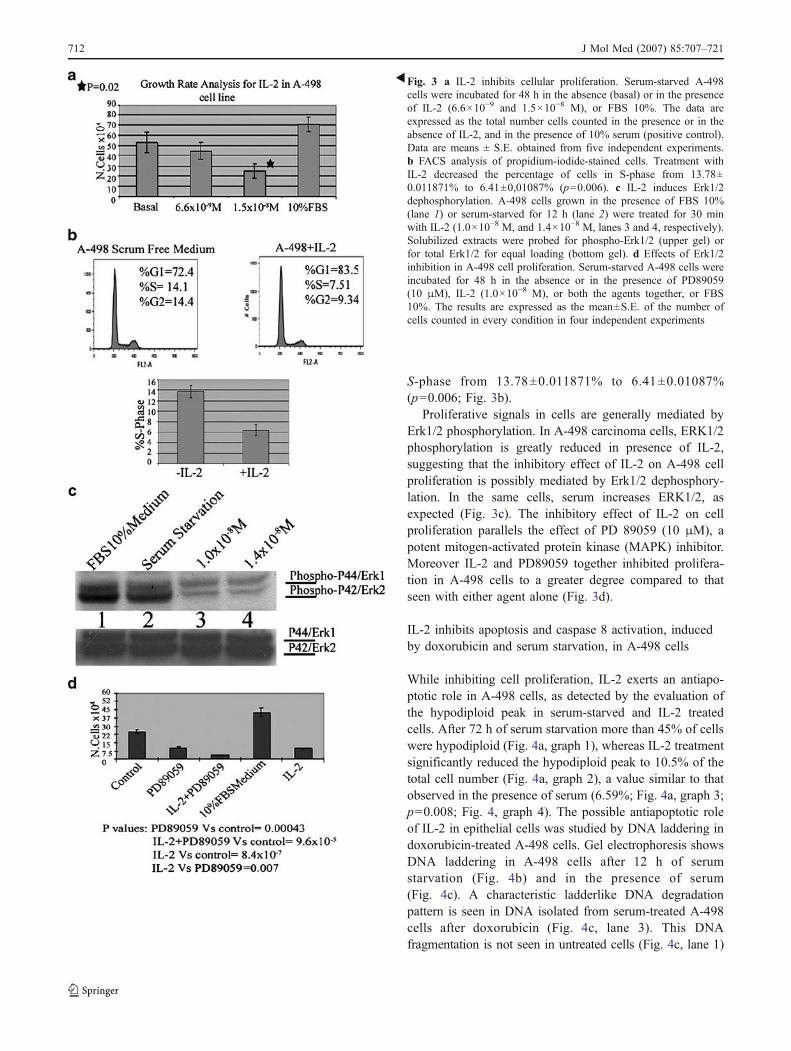
S-phase from 13.78±0.011871% to 6.41±0.01087%(p=0.006; Fig. 3b).
Proliferative signals in cells are generally mediated byErk1/2 phosphorylation. In A-498 carcinoma cells, ERK1/2phosphorylation is greatly reduced in presence of IL-2,suggesting that the inhibitory effect of IL-2 on A-498 cellproliferation is possibly mediated by Erk1/2 dephosphory-lation. In the same cells, serum increases ERK1/2, asexpected (Fig. 3c). The inhibitory effect of IL-2 on cellproliferation parallels the effect of PD 89059 (10 μM), apotent mitogen-activated protein kinase (MAPK) inhibitor.Moreover IL-2 and PD89059 together inhibited prolifera-tion in A-498 cells to a greater degree compared to thatseen with either agent alone (Fig. 3d).
IL-2 inhibits apoptosis and caspase 8 activation, inducedby doxorubicin and serum starvation, in A-498 cells
While inhibiting cell proliferation, IL-2 exerts an antiapo-ptotic role in A-498 cells, as detected by the evaluation ofthe hypodiploid peak in serum-starved and IL-2 treatedcells. After 72 h of serum starvation more than 45% of cellswere hypodiploid (Fig. 4a, graph 1), whereas IL-2 treatmentsignificantly reduced the hypodiploid peak to 10.5% of thetotal cell number (Fig. 4a, graph 2), a value similar to thatobserved in the presence of serum (6.59%; Fig. 4a, graph 3;p=0.008; Fig. 4, graph 4). The possible antiapoptotic roleof IL-2 in epithelial cells was studied by DNA laddering indoxorubicin-treated A-498 cells. Gel electrophoresis showsDNA laddering in A-498 cells after 12 h of serumstarvation (Fig. 4b) and in the presence of serum(Fig. 4c). A characteristic ladderlike DNA degradationpattern is seen in DNA isolated from serum-treated A-498cells after doxorubicin (Fig. 4c, lane 3). This DNAfragmentation is not seen in untreated cells (Fig. 4c, lane 1)
�Fig. 3 a IL-2 inhibits cellular proliferation. Serum-starved A-498cells were incubated for 48 h in the absence (basal) or in the presenceof IL-2 (6.6×10−9 and 1.5×10−8 M), or FBS 10%. The data areexpressed as the total number cells counted in the presence or in theabsence of IL-2, and in the presence of 10% serum (positive control).Data are means ± S.E. obtained from five independent experiments.b FACS analysis of propidium-iodide-stained cells. Treatment withIL-2 decreased the percentage of cells in S-phase from 13.78±0.011871% to 6.41±0,01087% (p=0.006). c IL-2 induces Erk1/2dephosphorylation. A-498 cells grown in the presence of FBS 10%(lane 1) or serum-starved for 12 h (lane 2) were treated for 30 minwith IL-2 (1.0×10−8 M, and 1.4×10−8 M, lanes 3 and 4, respectively).Solubilized extracts were probed for phospho-Erk1/2 (upper gel) orfor total Erk1/2 for equal loading (bottom gel). d Effects of Erk1/2inhibition in A-498 cell proliferation. Serum-starved A-498 cells wereincubated for 48 h in the absence or in the presence of PD89059(10 μM), IL-2 (1.0×10−8 M), or both the agents together, or FBS10%. The results are expressed as the mean±S.E. of the number ofcells counted in every condition in four independent experiments
712 J Mol Med (2007) 85:707–721

or in cells treated with doxorubicin after IL-2 preincubation(Fig. 4c, lane 2), proving that IL-2 is able to preventdoxorubicin-induced apoptosis. In serum-starved cells, DNAladdering was detected in the control, untreated cells,suggesting that serum starvation, in A-498 cells, can, byitself, induce DNA degradation (Fig. 4b, lane 1). Doxorubi-cin treatment greatly increased DNA degradation, with asignificant increment in the lower molecular weight compo-nents (Fig. 4b, lane 2). DNA degradation was greatlyreduced, compared to the control and doxorubicin treatedcells (Fig. 4b, lanes 1 and 2), when cells were incubated withIL-2 before doxorubicin treatment (Fig. 4b, lane 3), thusproving that IL-2 can indeed prevent apoptosis induced byboth serum starvation and doxorubicin treatment.
To better define the mechanisms involved in the IL-2mediated protection from apoptosis, we studied theactivation of caspase 8 in doxorubicin and IL-2 treatedA-498 cells. When cells were grown in the presence ofserum, virtually no caspase activation was observed inabsence of doxorubicin (Fig. 5a, lane 1). Again, doxoru-bicin treatment induced the activation of caspase8 (Fig. 5a, lane 2) and this effect was inhibited by IL-2(Fig. 5a, lane 3). Similar experiments were carried out inserum-starved cells (see supplementary data 2).The results,in the presence and in the absence of serum, are veryconsistent with what is described above in the DNAladdering experiments.
Active caspase 8 is predominantly found in the cyto-plasm [16]. We hypothesize that IL-2 may exert the anti-apoptotic effect by favoring the nuclear localization ofcaspase 8. Confocal imaging of doxorubicin-treated A-498cells, in the absence of IL-2 (Fig. 5b), showed that the redfluorescence of doxorubicin localized in the nucleus,whereas the green fluorescence of caspase 8 localizedpredominantly in the cytoplasm. IL-2 induced the nuclearlocalization of caspase 8 and colocalization with the nuclearred fluorescence of doxorubicin in most of the cells, thusrevealing a possible mechanism by which IL-2 exerts itsanti-apoptotic effect in this model. Figure 5b showsrepresentative fields by confocal microscopy. The IL-2effect on nuclear localization of caspase 8 was significant(p=0.0005, χ2 test).
IL-2 regulates Sgk1 at the transcriptional level
We then investigated the possible regulation of IL-2 ondownstream effectors such as Sgk1. Exposure of cells toIL-2, for 5 and 24 h, increased the amount of the Sgk1cDNA, as detected by reverse PCR (Fig. 6a). The increasein the expression of Sgk1 was specific, as IL-2 was unableto affect the expression of GAPDH, in the same conditions(Fig. 6a). At the protein level Sgk1 was detected in whole-celllysates by Western blotting with a specific rabbit anti-Sgk1antibody. The antibody specifically detected endogenous
Fig 4 a FACS analysis of the hypodiploid peak in A-498 cells. IL-2(graph 2) protects cells from apoptosis induced by serum starvation(graph 1) resulting in a percentage of apoptosis similar to that observedin the presence of serum (graph 4). The difference in the percentage ofapoptosis in cells treated with IL-2 and serum-starved cells is significant(graph 3). b and c DNA laddering in doxorubicin-treated A-498 cells. Inserum-starved cells (b), DNA laddering was detected in the control,untreated cells (b, lane 1). Doxorubicin treatment greatly increased
DNA degradation, with a significant increment in the lower molecularweight components (b, lane 2). DNA degradation was greatly reduced,when cells were incubated with IL-2 before doxorubicin treatment (b,lane 3). In serum-treated cells (c), a characteristic ladderlike DNAdegradation pattern is seen in DNA isolated from A-498 cells aftertreatment with doxorubicin (c, lane 3). This DNA fragmentation is notseen in untreated cells (c, lane 1) or in cells treated with doxorubicinafter IL-2 preincubation (c, lane 2)
J Mol Med (2007) 85:707–721 713

Sgk1 (see supplementary data 3). The amount of the Sgk1protein, increased 4 h after exposure of the cells to IL-2(1.0×10−8 M) and further increased after 12 h (Fig. 6b showsa representative gel). Both the panels include a normaliza-tion with anti β-tubulin immunoglobulins. The increase inthe expression of Sgk1 protein was quantified by scanningdensitometry of three independent experiments (Fig. 6d).The amount of the Sgk1 protein, as compared to basallevels, increased twofold within 4 h, and 2.5-fold within12 h of IL-2 treatment (1.0×10−8 M). To assess thesubcellular localization of Sgk1 after IL-2 stimulation, weperformed an immunofluorescence analysis of Sgk1 in A-498 cells. Upon stimulation with IL-2 (1.0×10−8 and 1.4×10−8 M), we discovered that there was more fluorescencedetected in the treated vs untreated cells, suggesting thatIL-2 caused an increase in total Sgk1 present in thecytoplasm (Fig. 6c). Interestingly, the IL-2 treatmentinduced a striking cell swelling, when compared to the
serum-starved cells (untreated) or to the serum- treatedcells (FBS 10%) [17].
IL-2 regulates Sgk1 in a posttranslational manner in vitroand in vivo
We detected an increase in the phosphorylation state ofSgk1, specifically on serine residue 422 (S422), uponstimulation with IL-2 (Fig. 7a). Phosphorylated Sgk1 wasspecifically detected by rabbit immunoglobulins raisedagainst Phospho-S422 Sgk1. Total Sgk1 was detected withthe rabbit anti-Sgk1 antibody (bottom). The phosphoryla-tion was detected within 30 min of IL-2 treatment and wasdose-dependent (Fig. 7a and c) and time-dependent(Fig. 7b). The IL-2 dependent phosphorylation of Serine422 on Sgk1 was measured by densitometry and expressedas the mean of five different experiments (Fig. 7c). It hasbeen previously described that Sgk1 is a downstream
Fig. 5 a IL-2 prevents caspase 8 activation induced by doxorubicin, inthe presence of FBS 10%. A-498 cells were incubated for 24 h, without(lane 1) or with doxorubicin (lane 2), or with doxorubicin after a2-h pretreatment of IL-2 (lane 3). Solubilized cell extracts were probedfor caspase 8. The upper band represents pro-caspase 8 and the lowerband represents the cleaved form (activated) of caspase 8. Equal proteinloading was confirmed by β-tubulin blotting. b Caspase 8 localizationby confocal microscopy. Caspase 8 (green) in A-498 cells, 24 h after
the addition of doxorubicin (red) in the absence (upper field) and in thepresence (lower field) of a 2-h pre-incubation with IL-2. Caspase8 (green) is predominantly cytoplasmic after treatment with doxoru-bicin alone, whereas it is predominantly nuclear and colocalizes withdoxorubicin when the drug is added after pretreatment with IL-2.Thesmall field on the top shows a control immunofluorescence with goatanti-rabbit FITC immunoglobulins. The IL-2 effect on caspase 8 locali-zation is significant at 0.0005 level (lateral table)
714 J Mol Med (2007) 85:707–721

target of phosphatidylinositol-3-kinase (PI3-kinase). Thephosphorylation of S422 on Sgk1 by IL-2 is mediated byPI3-kinase, as it is abolished by the specific inhibitor ofthis enzyme, LY294002 (Fig. 7b). The active, phosphor-ylated kinase was immuno-precipitated from IL-2 stim-ulted A-498 cells (see supplementary data 4). A kinaseassay (Fig. 7d) confirmed that the IL-2 dependentphosphorylation of Sgk1 led to an increase in the activityof the immuno-precipitated kinase. As expected, pretreat-ment with LY294002 completely abolished Sgk1 activa-tion under IL-2 treatment. The results are presented as theaverage of five independent experiments. To substantiatethis IL-2 dependent posttranslational modification ofSgk1, we further investigated whether this effect couldbe extended to an in vivo situation. In mice, IL-2 (3.3×10−8 M) induced a specific phosphorylation of S422 onSgk1 (Fig. 7e) in kidney, but not in liver. PhosphorylatedSgk1 was specifically detected by rabbit immunoglobulinsraised against Phospho-S422 Sgk1, but not by normalrabbit immunoglobulins (Fig. 7e).
Sgk is essential for IL2-dependent protectionfrom apoptosis
To better define the mechanism of IL-2 dependentprotection from apoptosis and the possible role of Sgk1,we performed experiments using Sgk1-specific siRNA anda dominant negative mutant (D222A) of Sgk1 in transientand in stably transfected cell lines.
We studied the effect of IL-2 on doxorubicin-inducedcaspase 8 activation in A-498 cells by means of scrambledor Sgk1-specific siRNA. The almost complete inhibition ofSgk1 expression by Sgk1-specific siRNA was verified byimmunodetection of the Sgk1 protein (Fig. 8a). Lanes 1 to3 show the results obtained in cells expressing normallevels of Sgk1. In basal conditions (Fig. 8a, lane 1), most ofthe caspase 8 were detected in the uncleaved, high-molecular-weight form. The addition of doxorubicin greatlydecreases the uncleaved form leading to the appearance ofhighly degraded, low-molecular-weight peptides (Fig. 8a,lane 2). IL-2 (Fig. 8a, lane 3) completely reverted the effect
Fig. 6 a Reverse PCR. IL-2 increased the amount of Sgk1 specificcDNA, amplified semiquantitative RT-PCR (top panel). Reverse PCRwas normalized by the amplification of GAPDH cDNA, in identicalconditions. b Induction of Sgk1 in response to IL-2 stimulation. IL-2increased the amount of Sgk1 protein in A498 cells, as detected byWestern blotting with Sgk1-specific immunoglobulins. Equal loading,was verified by anti-β-tubulin immunoglobulins. c Immunofluores-cence of endogenous Sgk1. IL-2 increases the amount of Sgk1detected in the cytoplasm of A-498 cells. Serum-starved A-498 cells
were incubated in the absence (untreated) or in the presence of variousconcentrations of IL-2 (1.0×10−8 and 1.4×10−8 M), as indicated, or inthe presence of serum (FBS 10%) for 4 h (colums 1, 2, and 3 representtriplicate experiments). The field on the extreme right shows a controlimmunofluorescence with goat anti-rabbit FITC immunoglobulins.d Densitometric scanning. The increase in Sgk1 expression wasmeasured by densitometric scanning and expressed as a ratio betweenthe OD at times 4 and 12 h and the OD at time 0. The results are theaverage±S.E. of three independent experiments
J Mol Med (2007) 85:707–721 715

of doxorubicin to the pattern observed in the absence of thedrug, thus confirming the role of IL-2 in protecting cellsfrom doxorubicin-dependent apoptosis. In the absence ofSgk1 (Fig. 8a, lanes 4 to 6), most of the caspase 8 weredetected in the uncleaved, high-molecular-weight form inthe absence of either doxorubicin and IL-2 (Fig. 8a, lane 4).The addition of doxorubicin greatly decreases theuncleaved form, leading to the appearance of highlydegraded, low molecular weight peptides (Fig. 8a, lane 5).IL-2 (Fig. 8a, lane 6) was unable to revert the effect ofdoxorubicin to the pattern observed in the absence of the
drug, thus confirming that Sgk1 is indeed necessary tomediate the effect of IL-2 on doxorubicin-dependentcaspase 8 activation. A graph showing quantitation bydensitometry from three independent experiments is pre-sented on the bottom of the panel.
In cells transiently transfected with a vector coding foram Myc-tagged dominant negative mutant of Sgk1(D222A), intermediate apoptotic figures appear, suggestingthat the inhibition of the endogenous Sgk1 activates anapoptotic pathway. Interestingly, also under these circum-stances and in the presence of fetal calf serum (FCS) 10%,
Fig. 7 a IL-2 increases Sgk1 phosphorylation in A-498 cells. A-498cells were incubated for 30 min without (lane 1) or with either twoconcentrations of IL-2 (lanes 2 and 3) or serum 10% FBS (lane 4), asindicated. Sgk1 was detected in solubilized cell extracts with rabbitimmunoglobulins specific for phopsho Ser 422Sgk (top) or for totalSgk1 (bottom). b IL-2 induced phosphorylation of Sgk1 is dependenton PI3-Kinase. Sgk1 phosphorylation was detected by blotting withanti-phospho-Ser 422 Sgk immunoglobulins, in A-498 cells treated byIL-2 (1.0×10−8 M) in the absence (lanes 1 to 4) or in the presence(lanes 5 to 7) of LY294002 (50 μM). Equal protein loading wasconfirmed by reprobing the filter with anti-Sgk1 immunoglobulins(bottom). c The phosphorylation of Sgk1 on Serine 422 was quantifiedby scanning densitometry. The results are expressed as the mean±S.E.of five independent experiments. d IL-2 dependent activation of Sgk1.
A-498 cells were stimulated for 30 min with increasing concentrationsof IL-2 (3.0×10
−9M and 4.6×10
−8M), with IL-2 (4.6×10
−8M) after
a 30-min incubation with LY294002 (50 μm), or with insulin (1 μM).The kinase activity associated to Sgk1 immunoprecipitates wasexpressed as the percent of the increase of the radioactivity (cpm)incorporated when the kinase was immunoprecipitated from unstimu-lated cells. Data are means ± S.E. of six independent experiments. e Invivo Sgk1 activation under IL-2 stimulation. Sgk1 phosphorylationwas detected in solubilized extracts prepared from kidneys and liversof four mice treated with vehicle alone (lanes 5, 6, 7, 8) or with IL-2(10,000 I.U. or 1.0×10 mol), subcutaneously, 3 days before thesacrifice (lanes 1, 2, 3, 4). Equal protein loading was confirmed byreprobing the filter with rabbit immunoglobulins specific for Sgk1(bottom)
716 J Mol Med (2007) 85:707–721

IL-2 was unable to inhibit completely doxorubicin-dependentcaspase 8 activation, thus confirming that IL-2 dependentprotection from apoptosis requires a functional endogenousSgk1 (see supplementary data 5).
The use of dominant negative mutant is sometimesbiased by the ability of the mutant to interphere withmultiple signal transduction pathways. We also producedstably transfected HELA cell lines expressing an myc-tagged dominant negative (DN) mutant of Sgk1 (D222A,DN pool; Fig. 8b). The alpha, beta, and gamma subunits ofthe IL-2 receptor were detected in HELA cells by RT-PCR(data not shown). In control HELA cells stably transfectedwith an empty vector, IL-2 inhibits the activation of caspase8 induced by doxorubicin (Fig. 8b, lanes 1, 2, 3).Interestingly, in cells stably transfected with the dominantnegative mutant, a certain degree of caspase activation is
detected, even in the absence of doxorubicin (Fig. 8b, lane 4),suggesting that this particular mutant is able by itself toincrease the percentage of apoptotic cells (R. Amato et al.manuscript in preparation). Moreover, in this cell line, IL-2is unable to inhibit the activation of caspase 8 induced bydoxorubicin (Fig. 8b, lanes 5 and 6).
Caspase-8 is the prototypical apoptosis initiator caspasedownstream of TNF superfamily death receptors. Wedetected the protein expression of FasL (a member of theTNF family) and Fas (CD 95) in our Hela cell lines stablytransfected by vectors coding for either wild-type Sgk1 orthe dominant negative Sgk1 mutant. A dramatic increase inthe expression of Fas (CD95) was observed in the cellsexpressing the dominant negative mutant (D222A) of Sgk1(Fig. 8c, lane 3) compared to the empty vector transfectedcells (Fig. 8c, lane 1) and the wild-type Sgk1 transfected
Fig. 8 a Sgk1 gene silencing and caspase 8 activation. Sgk1-specificsiRNA technology showed that Sgk1 is essential for the IL-2-dependentprotection from doxorubicin-induced apoptosis. A quantification of threeindependent experiments by scanning densitometry is shown. b Inter-lukin-2 protection from doxorucin-induced caspase 8 activation in stablytransfected HeLa cells expressing a dominant negative mutant of Sgk1.The inhibition of endogenous Sgk1 by means of a dominant negativetechnology showed that a functional Sgk1 is essential for the IL-2-
dependent protection from doxorubicin-induced apoptosis. c Expressionof CD95/Fas and CD/95 Fas L in stably transfected HeLa cellsexpressing a dominant negative mutant of Sgk1. The expression ofCD95/Fas is greatly increased in Hela cells lines expressing thedominant negative mutant (D222A) of Sgk 1 (lane 3), whereas theexpression of CD95/FasL is greatly reduced in Hela cells stablytransfected with a vector coding for wild type Sgk 1 (lane 2)
J Mol Med (2007) 85:707–721 717

cells (Fig. 8c, lane 2). On the other hand, the expression ofFasL was decreased in wild-type Sgk1 transfected cells(Fig. 8c, lane 2) compared to the other two cell lines.
(See supplementary data 5 for additional independentexperiments using siRNA silencing and transiently trans-fected A-498 cells expressing the D222A-dominant nega-tive mutant of Sgk1.)
Discussion
The role of IL-2 in proliferation, survival, and cytotoxicityhas been well investigated in hematopoetic cell lines [18].However, the function of this cytokine remains to be well-defined in epithelial tissues [14].
The role of the IL-2 receptor in epithelial cell proliferationis controversial [19]. This work defines a new role of the IL-2receptor activation in a human kidney carcinoma cell line.
The complete IL-2 receptor is composed of three subunits—alpha, beta, and gamma—and differences in the efficiency ofreceptor binding and transduction have been described depend-ing on its dimerization [20–23]. We confirmed the expressionof the beta chain of the IL-2 receptor, as previously described,in neoplastic tissues (lung, colon, stomach carcinoma)[24–27]. Moreover, we described for the first time theexpression of the beta chain in other neoplastic tissues(ovary, uterus, breast, and kidney), in embryonic tissues, inHEK293T cells, and in several normal tissues. As it is alwayspossible that human tumor extracts are contaminated byinfiltrating lymphocytes, we elected to study IL-2 signaling ina renal cancer cell model (A-498 cells), to verify whether theexpression and the activation of the IL-2 receptor pathway inepithelial cells might have consequences on cell survival. IL-2-binding competition experiments demonstrate that the receptorcontains two binding sites—the first component being of highaffinity and the other with middle to low affinity.
The nature of the binding curve suggests that allostericproperties exist in the heterodimeric binding domain of theIL-2 receptor complex [28, 29]. It is well established thatthe IL-2 receptor alpha chain is the only one that is specificfor the IL-2 receptor [22]. We therefore consider the alphachain as a selective marker for IL-2 receptor expression andimmunofluorescence was used to detect the alpha chain inA-498 cells. In addition, receptor internalization pits wereclearly visible after 4 h of IL-2 treatment, suggestingnormal receptor recycling [30].
We found that IL-2 treatment of A-498 cells induces theexpression of the IL-2 receptor gamma chain, in a time-dependent manner. Two differentially migrating forms ofthe gamma chain have been documented [31], a p64isoform restricted to lymphoid tissues and a p74 isoformrestricted to non-lymphoid tissue. In lymphoid tissue, IL-2acts as a growth factor by exerting a pro-proliferative
phenotype. We detected the p64 isoform in A-498 cells;however, IL-2 did not act as a growth factor in this model.Instead of inducing proliferation, IL-2 inhibited prolifera-tion. Therefore, we can deduce that the anti-proliferativephenotype induced by IL-2 in A-498 cells is independentfrom the isoform of the gamma chain that is present. In fact,it has been shown in some tissues that the IL-2 receptoralpha/beta chains are more limiting for the signal transduc-tion cascades initiated by receptor–ligand binding [30].
IL-2 has previously been described as a pro-proliferativein lymphoid tissue. Based on these assumptions, wewondered whether a similar effect could be documentedin cancer epithelial cells as well. Our growth rate analysisshowed that IL-2 plays an anti-proliferative role in thismodel. Analysis of the cell cycle by propidium iodidestaining and FACS demonstrated that IL-2 is able tosignificantly reduce the percentage of cells in S-phase, thusconfirming a cell growth inhibitory role for IL-2. A similarinhibition of cell proliferation was obtained when A-498cells were treated with PD89059, an inhibitor of MEK. IL-2plus PD89059, together, caused an even greater block ofproliferation as compared to that induced by either effectoralone. The significance of Erk-1/2 phosphorylation withinthe context of IL-2 regulation of cell proliferation [32] wasconfirmed by the finding that the phosphorylation of Erk1/2was clearly decreased in cells treated with IL-2.
Another important effect of IL-2 treatment of A-498cells is the inhibition of doxorubicin-dependent apoptosisand caspase 8 activation.
Sgk1 is well-known to be a key element in metabolicevents, including PKA and insulin-mediated sodium absorp-tion in the kidney [33]. It is well accepted that Sgk1 worksas a potent anti-apoptotic effector. However, the role of thiskinase remains to be elucidated [34, 35]. For example, inbreast cancer, Sgk1 has been shown to be upregulated,whereas it seems to be downregulated in prostatic cancer[36]. Therefore, we focused on the possible involvement ofSgk1 in IL-2-dependent regulation of apoptosis.
Fig. 9 Sgk1 in IL-2 signal transduction and apoptosis
718 J Mol Med (2007) 85:707–721

After binding to its own specific receptor, IL-2 signalsthrough Sgk1, a typical downstream effector of PI3-kinase[37–40]. IL-2 induced a strong increase in the level of Sgk1protein expression, which can be attributed to the activationof the transcription of the gene coding for Sgk1, asevidenced by the reverse PCR experiments. We also foundthat IL-2 can activate Sgk1 at the posttranslational levelthrough a PI3-kinase dependent phosphorylation of serine422 of Sgk1, a critical residue required for the activation ofthis kinase. In addition, we showed that the phosphoryla-tion of Sgk1 by IL-2 resulted in the activation of the kinase.This IL-2-dependent posttranslational activation of Sgk1may have physiological relevance as it occurs “in vivo” asit can be detected in kidney upon intra-peritoneal adminis-tration in mice.
The activation of caspase 8 in A-498 cells has been usedto monitor the IL-2 effects on apoptosis [41]. It has beenwell documented that doxorubicin-induced cell death ismediated by caspase 8 activation [42–44]. We have shownthat IL-2 completely inhibits caspase 8 activation inducedby doxorubicin treatment. Sgk1 has been demonstrated tobe involved in the transduction of survival signals frominsulin, IGF1 and steroids; moreover, it has been shown tobe upregulated in several epithelial human cancers [45–47].The observation that IL-2 may activate Sgk1 at thetranscriptional and posttranslational level, in kidney cancercells, raises important concerns for the effectiveness andsafety of the immunotherapy in these tumors. The activa-tion of anti-apoptotic pathways may in fact trigger theescape from effective treatment and favor the survival ofcancer cells. As a matter a fact, we demonstrated, by FACSanalysis and gel laddering experiments, that IL-2 canprevent apoptosis induced by doxorubicin treatment andserum starvation.
The importance of Sgk1 in mediating the IL-2 effect oncaspase 8 activation has been further studied in cellstransiently and stably transfected with a vector coding forthe dominant negative mutant of Sgk1 (D222A) and bymeans of Sgk1 specific siRNA. The functional inhibition ofthe SGK1 kinase by the dominant negative mutant and theinhibition of the expression of the kinase by means ofSgk1-specific siRNA gave similar results. The appearanceof intermediate apoptotic figures in cells transfected withthe vector coding for a dominant negative mutant of Sgk1(D222A), suggests that the inhibition of the endogenousSgk1 activates an apoptotic pathway.
Although we are unable at present to clearly describe themechanism by which Sgk1 acts on caspase 8 activation, wenoticed that the expression of the dominant negative mutantof Sgk1 increases the expression of Fas (CD95), whereasthe hyper-expression of wild-type Sgk1 decreases theexpression of FasL. These observations strongly suggestthat the susceptibility to caspase 8 activation in cells
expressing the dominant negative mutant of Sgk1 mayinvolve the regulation of Fas/FasL expression as previouslydescribed in different cancer cell lines [48]. We noticed thatthe IL-2-dependent inhibition of caspase 8 activation is notobserved in cells transfected with a dominant negativemutant of Sgk1, as well as in cells treated with the Sgk-1-specific siRNA, suggesting that a functional endogenousSgk1 is necessary for mediating the IL-2-dependentprotection from apoptosis induced by doxorubicin.
Caspase 8 can have a nuclear or a cytoplasmiclocalization. As documented by others [49, 50], caspase8 is incapable of activation and may be sumoylated [51]when localized within the nucleus, whereas it is fully activewhen localized within the cytoplasm. Our experiments byconfocal imaging show that caspase 8 is predominantlycytoplasmic when cells are treated with doxorubicin alone,whereas it shifts to the nucleus when cells are treated withIL-2 before the addition of doxorubicin.
Taken together our data prove that interleukin 2inhibits proliferation in A-498 cells and inhibits apoptosissignaling through Sgk1. The observations are summarizedin Fig. 9. This double effect is only seemingly controver-sial. Promitogenic and anti-apoptotic functions of IL-2 canbe uncoupled in T lymphocytes by specific signals [52].On the other hand, a role for Sgk1 in survival andhypertrophic response has been recently described incardiomyocytes [53]. Within this model the hypertrophycould be the final result of the inhibition of bothproliferation and apoptosis. On a more clinical side, ourfindings may suggest to review the knowledge about thefunction and the therapeutic usefulness of IL-2 in humancancer, as cell-mediated cytotoxicity might be inhibitied inrenal cell carcinoma cells exposed to IL-2.
Acknowledgments We thank Charity Nofziger and ProfessorBonnie Blazer-Yost for critical reading and helpful discussion of themanuscript, and Nicola Amodio for the caspase 8 confocal experi-ment’s acquisition.
This work was supported by Fondazione Carical, Cluster 04work package 6A, Cofin 2003/20032003063143_003, Cofin 2005 /068017_004, Interlink II04C0G4EM.
References
1. David D, Bani L, Moreau JL, TreilhouMP, Nakarai T, JoussemetM,Ritz J, Dupont B, Pialoux G, Theze J (1998) Regulatory dysfunctionof the interleukin-2 receptor during HIV infection and theimpact of triple combination therapy. Proc Natl Acad Sci USA95:11348–11353
2. Upton MP, Parker RA, Youmans A, McDermott DF, Atkins MB(2005) Histologic predictors of renal cell carcinoma response tointerleukin-2-based therapy. J Immunother 28:488–495
3. Bazan JF (1992) Unraveling the structure of IL-2. Science257:410–412
J Mol Med (2007) 85:707–721 719

4. Kelly E, Won A, Refaeli Y, Van Parijs L (2002) IL-2 and relatedcytokines can promote T cell survival by activating AKT1.J Immunol 168:597–603
5. Brunet A, Park J, Tran H, Hu LS, Hemmings BA, Greenberg ME(2001) Protein kinase SGK mediates survival signals by phos-phorylating the forkhead transcription factor FKHRL1 (FOXO3a).Mol Cell Biol 21:952–965
6. Kobayashi T, Cohen P (1999) Activation of serum- and glucocor-ticoid-regulated protein kinase by agonists that activate phosphati-dylinositide 3-kinase is mediated by 3-phosphoinositide-dependentprotein kinase-1 (PDK1) and PDK2. Biochem J 339:319–328
7. Perrotti N, He RA, Phillips SA, Haft CR, Taylor SI (2001)Activation of serum-and glucocorticoid-induced protein kinase(Sgk) by cyclic AMP and insulin. J Biol Chem 276:9406–9412
8. Frodin M, Antal TL, Dummler BA, Jensen CJ, Deak M,Gammeltoft S, Biondi RM (2002) A phosphoserine/threonine-binding pocket in AGC kinases and PDK1 mediates activation byhydrophobic motif phosphorylation. EMBO J 21:5396–5407
9. Menniti M, Iuliano R, Amato R, Boito R, Corea M, Le Pera I,Gulletta E, Fuiano G, Perrotti N (2005) Serum and glucocorticoid-regulated kinase Sgk1 inhibits insulin-dependent activation ofphosphomannomutase 2 in transfected COS-7 cells. Am J PhysiolCell Physiol 288:C148–C155
10. Leong ML, Maiyar AC, Kim B, O’Keeffe BA, Firestone GL(2003) Expression of the serum- and glucocorticoid-inducibleprotein kinase, Sgk, is a cell survival response to multiple types ofenvironmental stress stimuli in mammary epithelial cells. J BiolChem 278:5871–5882
11. Maiyar AC, Huang AJ, Phu PT, Cha HH, Firestone GL (1996)p53 stimulates promoter activity of the sgk. serum/glucocorticoid-inducible serine/threonine protein kinase gene in rodent mammaryepithelial cells. J Biol Chem 271:12414–12422
12. Wu W, Chaudhuri S, Brickley DR, Pang D, Karrison T, ConzenSD (2004) Microarray analysis reveals glucocorticoid-regulatedsurvival genes that are associated with inhibition of apoptosis inbreast epithelial cells. Cancer Res 64:1757–1764
13. Zheng, Li D-C, Zhang Z-D, Zhao J, Ge J-F (2005) Anti-gastriccancer active immunity induced by FasL/B7-1gene-modifiedtumor cells. World J Gastroenterol 11:3204–3211:1757–1764
14. Suminami Y, Kashii Y, Law JC, Lin WC, Stanson J, Reichert TE,Rabinowich H, Whiteside TL (1988) Molecular analysis of theIL-2 receptor beta chain gene expressed in human tumor cells.Oncogene 16:1309–1317
15. Christoffersen CT, Bornefeldt KE, Rotella CM, Gonzales NVissing H, Shymko RM, Hoeve JT, Groffen J, Heisterkamp N,De Meyts P (1994) Negative cooperativity in the insulin-likegrowth factor-I receptor and a chimeric IGF-I/insulin receptor.Endocrinology 1:472–475
16. Shikama Y, U M, Miyashita T, Yamada M (2001) Comprehensivestudies on subcellular localization and cell death inducingactivities of eight GFP-tagged apoptosis related caspase. Exp CellRes 264:315–325
17. Waldegger S, Barth P, Raber G, Lang F (1997) Cloning andcharacterization of a putative human serine/threonine proteinkinase transcriptionally modified during anisotonic and isotonicalterations of cell volume. Proc Natl Acad Sci U SA 94:4440–4445
18. Schimpl A, Berberich I, Kneitz B, Kramer S, Santner-Nanan B,Wagner S, Wolf M, Hunig T (2002) IL-2 and autoimmune disease.Cytokine Growth Factor Rev 13:369–378
19. Lesur O, Brisebois M, Thibodeau A, Chagnon F, Lane D, Fullop T(2004) Role of IFN-gamma and IL-2 in rat lung epithelial cellmigration and apoptosis after oxidant injury. Am J Physiol LungCell Mol Physiol 286:L4–L14
20. Fukushima K, Hara-Kuge S, Ideo H, Yamashita K (2001)Carbohydrate recognition site of interleukin-2 in relation to cellproliferation. J Biol Chem 276:31202–31208
21. Sharon M, Siegel JP, Tosato G, Yodoi J, Gerrard TL, Leonard WJ(1988) The human interleukin 2 receptor beta chain (p70). Directidentification, partial purification, and patterns of expression onperipheral blood mononuclear cells. J Exp Med 167:1265–1270
22. Weissman AM, Harford JB, Svetlik PB, Leonard WL, Depper JM,Waldmann TA, Greene WC, Klausner RD (1986) Only high-affinity receptors for interleukin 2 mediate internalization ofligand. Proc Natl Acad Sci U SA 83:1463–1466
23. Wang Xi, Rickert M, Garcia KC (2005) Structure of thequaternary complex of interleukin-2 with its, ß, and χc receptors.Science 310:1159–1163
24. Kradin RL, Xia W, Pike M, Byers HR, Pinto C (1996) Interleukin-2 promotes the motility of dentric cells and their accumulation inlung and skin. Pathobiology 64:180–186
25. Thèze J, Alzari PM, Bortoglio J (1996) Intereleukin 2 and itsreceptor recent advances and new immunological functions.Immunol Today 17:481–484
26. Reinecker HC, Podolsky D (1995) Human intestinal epithelialcells express functional cytokine receptor sharing the common γcchain oh the IL-2 receptor. Proc Natl Acad Sci U SA 92:8353–8357
27. McMillian DN, Kernohan NM, Flett ME, Heys SD, Deehan DJ,Sewell HF, Walker F, Eremin O (1995) Interleukin-2 receptorexpresion in human solid tumor cells in situ and in vitro: evidencefor a direct role in regulation of tumor cell proliferation. Int JCancer 60:766–772
28. Vamosi G, Bodnar A, Vereb G, Jenei A, Goldman CK, Langowski J,Toth K, Matyus L, Szollosi J, Waldmann TA, Damjanovich S (2004)IL-2 and IL-15 receptor alpha-subunits are coexpressed in asupramolecular receptor cluster in lipid rafts of T cells. Proc NatlAcad Sci USA 101:11082–11087
29. Robb RJ, Greene WC, Rusk CM (1984) Low and high affinitycellular receptors for interleukin 2. Implications for the level ofTac antigen. J Exp Med 160:1126–1146
30. Gesbert F, Sauvonnet N, Dautry-Varsat A (2004) Clathrin-independent endocytosis and signalling of interleukin 2 receptorsIL-2R endocytosis and signalling. Curr Top Microbiol Immunol286:119–148
31. Morrone G, Bond HM, Cuomo C, Agosti V, Petrella A, PagnanoAM, Della Corte A, Marasco O, Venuta S (1995) Differentialregulation of the expression of interleukin-2 receptor gamma-chain during the in vitro differentiation of human myeloid cells.Biochem J 308:909–914
32. Romerio F, Zella D (2002) MEK and ERK inhibitors enhance theanti-proliferative effect of interferon-alpha2b. FASEB J 16:1680–1682
33. Faletti CJ, Perrotti N, Taylor SI, Blazer-Yost BL (2002) sgk: anessential convergence point for peptide and steroid hormoneregulation of ENaC-mediated Na+ transport. Am J Physiol CellPhysiol 282:C494–C500
34. Sakoda H, Gotoh Y, Katagiri H, Kurokawa M, Ono H, Onishi Y,Anai M, Ogihara T, Fujishiro M, Fukushima Y, Abe M, Shojima N,Kikuchi M, Oka Y, Hirai H, Asano T (2003) Differing roles of Aktand serum and glucocorticoid-regulated kinase in glucose metabo-lism, DNA synthesis, and oncogenic activity. J Biol Chem278:25802–25807
35. Vallon V, Lang F (2005) New insights into the role of serum- andglucocorticoid-inducible kinase SGK1 in the regulation of renalfunction and blood pressure. Curr Opin Nephrol Hypertens 14:59–66
36. Rauhala HE, Porkka KP, Tolonen TT, Martikainen PM, TammelaTL, Visakorpi T (2005) Dual-specificity phosphatase 1 and serum/glucocorticoid-regulated kinase are downregulated in prostatecancer. Int J Cancer 117:738–745
37. Biondi RM, Kieloch A, Currie RA, Deak M, Alessi DR (2001)The PIF-binding pocket in PDK1 is essential for activation of S6Kand SGK, but not PKB. EMBO J 20:4380–4390
720 J Mol Med (2007) 85:707–721

38. Lang F, Cohen P (2001) Regulation and physiological roles ofserum- and glucocorticoid-induced protein kinase isoforms.Science’s Signal Transduct Knowledge Environment 108:RE17
39. Park J, Leong ML, Buse P, Maiyar AC, Firestone GL, HemmingsBA (1999) Serum and glucocorticoid-inducible kinase (SGK) is atarget of the PI 3-kinase-stimulated signaling pathway. EMBO J18:3024–3033
40. Alessi DR, James SR, Downes CP, Holmes AB, Gaffney PR,Reese CB, Cohen P (1997) Characterization of a 3-phosphoinosi-tide-dependent protein kinase which phosphorylates and activatesprotein kinase B alpha. Curr Biol 7:261–269
41. Lens SM, Kataoka T, Fortner KA, Tinel A, Ferrero I, MacDonaldRH, Hahne M, Beermann F, Attinger A, Orbea HA, Budd RC,Tschopp J (2002) The caspase 8 inhibitor c-FLIP(L) modulates T-cell receptor-induced proliferation but not activation-induced celldeath of lymphocytes. Mol Cell Biol 22:5419–5433
42. Boehrer S, Kukoc-Zivojnov N, Nowak D, Bergmann M, Baum C,Puccetti E, Weidmann E, Hoelzer D, Mitrou PS, Chow KU (2004)Upon drug-induced apoptosis expression of prostate-apoptosis-response-gene-4 promotes cleavage of caspase-8, bid and mito-chondrial release of cytochrome c. Hematology 9:425–431
43. Chen JS, Konopleva M, Andreeff M, Multani AS, Pathak S, MehtaK (2004) Drug-resistant breast carcinoma (MCF-7) cells areparadoxically sensitive to apoptosis. J Cell Physiol 200:223–234
44. Wu XX, Kakehi Y, Mizutani Y, Kamoto T, Kinoshita H, IsogawaY, Terachi T, Ogawa O (2002) Doxorubicin enhances TRAIL-induced apoptosis in prostate cancer. Int J Oncol 20:949–954
45. Tangir J, Bonafe N, Gilmore-Hebert M, Henegariu O, ChambersSK (2004) SGK1, a potential regulator of c-fms related breastcancer aggressiveness. Clin Exp Metastasis 21:477–483
46. Buse P, Tran SH, Luther E, Phu PT, Aponte GW, Firestone GL(1999) Cell cycle and hormonal control of nuclear-cytoplasmiclocalization of the serum- and glucocorticoid-inducible proteinkinase, Sgk, in mammary tumor cells. A novel convergence pointof anti-proliferative and proliferative cell signaling pathways.J Biol Chem 274:7253–7263
47. Waldegger S, Klingel K, Barth P, Sauter M, Rfer ML, Kandolf R,Lang F (1999) h-sgk serine-threonine protein kinase gene astranscriptional target of transforming growth factor beta in humanintestine. Gastroenterology 116:1081–1088
48. Li W, Wang S, Chen C, Zhuang G (2006) Induction of tumor cellapoptosis via Fas/DR5. Cell Mol Immunol 3:467–471
49. Schickling O, Stegh AH, Byrd J, Peter ME (2001) Nuclearlocalization of DEDD leads to caspase-6 activation through itsdeath effector domain and inhibition of RNA polymerase Idependent transcription. Cell Death Differ 8:1157–1168
50. Ferrando-May E (2005) Nucleocytoplasmic transport in apoptosis.Cell Death Differ 12:1236–1276
51. Besnault-Mascard L, Leprince C, Auffredou MT, Meunier B,Bourgeade MF, Camonis J, Lorenzo HK, Vazquez A (2005)Caspase-8 sumoylation is associated with nuclear localization.Oncogene 24:3268–3273
52. Nelson BH, Martyak TP, Thompson LJ, Moon JJ, Wang T (2003)Uncoupling of promitogenic and antiapoptotic functions of IL-2by Smad-dependent TGF-beta signaling. J Immunol 170:5563–5570
53. Aoyama T, Matsui T, Novikov M, Park J, Hemmings B,Rosenzweig A (2005) Serum and glucocorticoid-responsivekinase-1 regulates cardiomyocyte survival and hypertrophicresponse. Circulation 111:1652–1659
J Mol Med (2007) 85:707–721 721




















