
Identification and expansion of human osteosarcoma-cancer-stem cells by long-term 3-aminobenzamide...
13
Identification and Expansion of Human Osteosarcoma-Cancer-Stem Cells by Long-Term 3-Aminobenzamide Treatment RICCARDO DI FIORE, 1 ANDREA SANTULLI, 1,2 ROSA DRAGO FERRANTE, 1 MICHELA GIULIANO, 1 ANNA DE BLASIO, 1 CONCETTA MESSINA, 1,2 GIUSEPPE PIROZZI, 3 VIRGINIA TIRINO, 3 GIOVANNI TESORIERE, 1 AND RENZA VENTO 1 * 1 Dipartimento di Scienze Biochimiche, Universita`degli Studi di Palermo, Policlinico, Palermo, Italy 2 Istituto di Biologia Marina, Consorzio Universitario di Trapani, Erice Casa Santa, Trapani, Italy 3 Dipartimento di Oncologia Sperimentale, Istituto Nazionale dei Tumori, Fondazione G. Pascale, Napoli, Italy A novel cancer stem-like cell line (3AB-OS), expressing a number of pluripotent stem cell markers, was irreversibly selected from human osteosarcoma MG-63 cells by long-term treatment (100 days) with 3-aminobenzamide (3AB). 3AB-OS cells are a heterogeneous and stable cell population composed by three types of fibroblastoid cells, spindle-shaped, polygonal-shaped, and rounded-shaped. With respect to MG-63 cells, 3AB-OS cells are extremely smaller, possess a much greater capacity to form spheres, a stronger self-renewal ability and much higher levels of cell cycle markers which account for G1-S/G2-M phases progression. Differently from MG-63 cells, 3AB-OS cells can be reseeded unlimitedly without losing their proliferative potential. They show an ATP-binding cassette transporter ABCG2-dependent phenotype with high drug efflux capacity, and a strong positivity for CD133, marker for pluripotent stem cells, which are almost unmeasurable in MG-63 cells. 3AB-OS cells are much less committed to osteogenic and adipogenic differentiation than MG-63 cells and highly express genes required for maintaining stem cell state (Oct3/4, hTERT, nucleostemin, Nanog) and for inhibiting apoptosis (HIF-1a, FLIP-L, Bcl-2, XIAP, IAPs, and survivin). 3AB-OS may be a novel tumor cell line useful for investigating the mechanisms by which stem cells enrichment may be induced in a tumor cell line. The identification of a subpopulation of cancer stem cells that drives tumorigenesis and chemoresistance in osteosarcoma may lead to prognosis and optimal therapy determination. Expression patterns of stem cell markers, especially CD133 and ABCG2, may indicate the undifferentiated state of osteosarcoma tumors, and may correlate with unfavorable prognosis in the clinical setting. J. Cell. Physiol. 219: 301–313, 2009. ß 2009 Wiley-Liss, Inc. Stem cells are characterized by the ability to divide asymmetrically producing two daughter cells, one is a new stem cell with the capability for self-renewal without losing the proliferative capacity with each cell division, and the second is progenitor cell, which has the ability for differentiation and proliferation, but not the capability for self-renewal (Reya et al., 2001). Stem cells are immortal, and rather resistant to action of drugs (Dean et al., 2005). Normal stem cells in the adult organism are responsible for tissue renewal and repair of aged or damaged tissue (Molofsky et al., 2004). Recent studies performed in several types of human cancer, suggest that tumors are organized in a hierarchy of heterogeneous cell populations with the capability to maintain tumor formation/growth specifically residing in a small population of cells called cancer stem cells (CSCs) (Clarke et al., 2006; Dalerba et al., 2007). Whether a tumor derives from transformed organ stem cells or whether the CSC have acquired their self-renewal capacity during tumor development is still an open question (Immervoll et al., 2008). However, the concept of stem cells as a definite cell population in a niche which represent the source for tissue renewal is widely accepted (Schaffer, 2007), and a number of studies suggest that a small population of cells, the tumor initiating cells, with unique self-renewal properties, with malignant potential and therapy resistant exists in at least some, and perhaps all, solid tumors (Al-Hajj et al., 2003; Al-Hajj and Clarke, 2004; Singh et al., 2004; Gibbs et al., 2005; Wang and Dick, 2005; Li and Neaves, 2006; O’Brien et al., 2007; Maitland and Collins, 2008). Apart from the ability of self-renewal and proliferation, and the capability to be resistant to drugs, this functional subset of CSC have a number of characteristic initially established for human acute myeloid leukemia and subsequently extended to human solid tumors (Jordan et al., 2006; Ailles and Weissman, 2007; Graziano et al., 2008; Yang and Chang, 2008; Zou, 2008), which is the expression of typical markers of stem cells, including CD133, recognized as a stem cell marker for normal and cancerous tissues (Mizrak et al., 2008), and ATP-binding cassette transporters, by which the cells can pump out specific fluorescence dyes, such as Hoechst 33342 or rhodamine 123 (Bunting, 2002; Hadnagy et al., 2006). Although standard chemotherapy kills most cells in a tumor, CSC remain viable, thus, despite the small number of such cells, they might be the cause of tumor recurrence, sometimes many years after the ‘‘successful’’ treatment of primary tumor. Thus, further characterization of CSC is needed in order to find ways to destroy them, which might contribute significantly to the therapeutic management of malignant tumors. G. Tesoriere and R. Vento contributed equally to this work. Contract grant sponsor: Ministero dell’Istruzione, dell’Universita `e della Ricerca. *Correspondence to: Renza Vento, Dipartimento di Scienze Biochimiche, Universita ` degli Studi di Palermo, Policlinico, Via Del Vespro, 90127 Palermo, Italy. E-mail: [email protected] Received 19 August 2008; Accepted 14 November 2008 Published online in Wiley InterScience (www.interscience.wiley.com.), 21 January 2009. DOI: 10.1002/jcp.21667 ORIGINAL ARTICLE 301 Journal of Journal of Cellular Physiology Cellular Physiology ß 2009 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Identification and expansion of human osteosarcoma-cancer-stem cells by long-term 3-aminobenzamide...

ORIGINAL ARTICLE 301J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
Identification and Expansion ofHuman Osteosarcoma-Cancer-StemCells by Long-Term3-Aminobenzamide Treatment
RICCARDO DI FIORE,1 ANDREA SANTULLI,1,2 ROSA DRAGO FERRANTE,1MICHELA GIULIANO,1 ANNA DE BLASIO,1 CONCETTA MESSINA,1,2 GIUSEPPE PIROZZI,3
VIRGINIA TIRINO,3 GIOVANNI TESORIERE,1 AND RENZA VENTO1*1Dipartimento di Scienze Biochimiche, Universita degli Studi di Palermo, Policlinico, Palermo, Italy2Istituto di Biologia Marina, Consorzio Universitario di Trapani, Erice Casa Santa, Trapani, Italy3Dipartimento di Oncologia Sperimentale, Istituto Nazionale dei Tumori, Fondazione G. Pascale, Napoli, Italy
A novel cancer stem-like cell line (3AB-OS), expressing a number of pluripotent stem cell markers, was irreversibly selected from humanosteosarcoma MG-63 cells by long-term treatment (100 days) with 3-aminobenzamide (3AB). 3AB-OS cells are a heterogeneous andstable cell population composed by three types of fibroblastoid cells, spindle-shaped, polygonal-shaped, and rounded-shaped. Withrespect to MG-63 cells, 3AB-OS cells are extremely smaller, possess a much greater capacity to form spheres, a stronger self-renewalability and much higher levels of cell cycle markers which account for G1-S/G2-M phases progression. Differently from MG-63 cells,3AB-OS cells can be reseeded unlimitedly without losing their proliferative potential. They show an ATP-binding cassette transporterABCG2-dependent phenotype with high drug efflux capacity, and a strong positivity for CD133, marker for pluripotent stem cells, whichare almost unmeasurable in MG-63 cells. 3AB-OS cells are much less committed to osteogenic and adipogenic differentiation than MG-63cells and highly express genes required for maintaining stem cell state (Oct3/4, hTERT, nucleostemin, Nanog) and for inhibiting apoptosis(HIF-1a, FLIP-L, Bcl-2, XIAP, IAPs, and survivin). 3AB-OS may be a novel tumor cell line useful for investigating the mechanisms by whichstem cells enrichment may be induced in a tumor cell line. The identification of a subpopulation of cancer stem cells that drivestumorigenesis and chemoresistance in osteosarcoma may lead to prognosis and optimal therapy determination. Expression patterns ofstem cell markers, especially CD133 and ABCG2, may indicate the undifferentiated state of osteosarcoma tumors, and may correlatewith unfavorable prognosis in the clinical setting.
J. Cell. Physiol. 219: 301–313, 2009. � 2009 Wiley-Liss, Inc.
G. Tesoriere and R. Vento contributed equally to this work.
Contract grant sponsor: Ministero dell’Istruzione, dell’Universita edella Ricerca.
*Correspondence to: Renza Vento, Dipartimento di ScienzeBiochimiche, Universita degli Studi di Palermo, Policlinico, Via DelVespro, 90127 Palermo, Italy. E-mail: [email protected]
Received 19 August 2008; Accepted 14 November 2008
Published online in Wiley InterScience(www.interscience.wiley.com.), 21 January 2009.DOI: 10.1002/jcp.21667
Stem cells are characterized by the ability to divideasymmetrically producing two daughter cells, one is a new stemcell with the capability for self-renewal without losing theproliferative capacity with each cell division, and the second isprogenitor cell, which has the ability for differentiation andproliferation, but not the capability for self-renewal (Reya et al.,2001). Stem cells are immortal, and rather resistant to actionof drugs (Dean et al., 2005). Normal stem cells in the adultorganism are responsible for tissue renewal and repair ofaged or damaged tissue (Molofsky et al., 2004).
Recent studies performed in several types of human cancer,suggest that tumors are organized in a hierarchy ofheterogeneous cell populations with the capability to maintaintumor formation/growth specifically residing in a smallpopulation of cells called cancer stem cells (CSCs) (Clarke et al.,2006; Dalerba et al., 2007). Whether a tumor derives fromtransformed organ stem cells or whether the CSC haveacquired their self-renewal capacity during tumor developmentis still an open question (Immervoll et al., 2008). However, theconcept of stem cells as a definite cell population in a nichewhich represent the source for tissue renewal is widelyaccepted (Schaffer, 2007), and a number of studies suggest thata small population of cells, the tumor initiating cells, with uniqueself-renewal properties, with malignant potential and therapyresistant exists in at least some, and perhaps all, solid tumors(Al-Hajj et al., 2003; Al-Hajj and Clarke, 2004; Singh et al., 2004;Gibbs et al., 2005; Wang and Dick, 2005; Li and Neaves, 2006;O’Brien et al., 2007; Maitland and Collins, 2008). Apart from theability of self-renewal and proliferation, and the capability to beresistant to drugs, this functional subset of CSC have a numberof characteristic initially established for human acute myeloid
� 2 0 0 9 W I L E Y - L I S S , I N C .
leukemia and subsequently extended to human solid tumors(Jordan et al., 2006; Ailles and Weissman, 2007; Graziano et al.,2008; Yang and Chang, 2008; Zou, 2008), which is theexpression of typical markers of stem cells, including CD133,recognized as a stem cell marker for normal and canceroustissues (Mizrak et al., 2008), and ATP-binding cassettetransporters, by which the cells can pump out specificfluorescence dyes, such as Hoechst 33342 or rhodamine123 (Bunting, 2002; Hadnagy et al., 2006). Although standardchemotherapy kills most cells in a tumor, CSC remain viable,thus, despite the small number of such cells, they might be thecause of tumor recurrence, sometimes many years afterthe ‘‘successful’’ treatment of primary tumor. Thus, furthercharacterization of CSC is needed in order to find ways todestroy them, which might contribute significantly to thetherapeutic management of malignant tumors.

302 D I F I O R E E T A L .
Recently, using a sarcosphere culture system, has beenreported the identification of a subpopulation of stem-like cellsin osteosarcomas, and demonstrated that these cells expressthe key molecular machinery of embryonic stem cells(Gibbs et al., 2005). Osteosarcoma is the most commonnonhematologic malignancy of bone in children and adults,consisting of malignant cells that produce immature bone and ischaracterized by osteoid formation within the tumor (Tanget al., 2008). There are about 900 new cases of osteosarcomadiagnosed in the US each year with about 400 occurring inchildren and adolescents, younger than 20 years of age (Picci,2007). The most common treatment for osteosarcoma is acombination of chemotherapy, using drugs to kill the cancercells, and surgery to remove the cancer and some healthy tissue.Although chemotherapy protocols (Spangler, 2008) haveproduced survival rates in the 55–70% range for nonmetastaticextremity osteosarcoma, unfortunately, in 15–20% of casesmetastasis are present at the time of diagnosis and an additional20–25% of the patients develop metastasis during the course ofdisease, having a very poor prognosis despite achievements inmultimodal therapy (Marina et al., 2004). Recently (De Blasioet al., 2005), we have demonstrated that in humanosteosarcoma MG-63 cells, aberrant gene expression keeps Rbprotein constitutively inactivated throughhyperphosphorylation and this promotes uncontrolled cellproliferation. Brief-term treatment with 3-aminobenzamide(3AB), a potent inhibitor of poly(ADP-ribose) polymerase(PARP), induces morphological and biochemical features ofosteocyte differentiation, accompanied by an increase in thehypophosphorylated active form of Rb and in Rb/E2Fcomplexes, with downregulation of gene products required forproliferation (cyclin D1, b-catenin, c-Jun, c-Myc and Id2) andupregulation of those implicated in the osteoblastdifferentiation (p21/Waf1, osteopontin, osteocalcin, type Icollagen, N-cadherins and alkaline phosphatase). Our studysuggested that in MG-63 cells, 3AB treatment may induce aremodeling of chromatin with the reprogramming of geneexpression and the activation of differentiation.
Now, we demonstrate that long-term treatment of humanosteosarcoma MG-63 cells with 3AB, favors the enrichmentwith a cellular phenotype with stem cell characteristicsdemonstrated by morphological, flowcytometrical, andbiomolecular evidence.
Materials and MethodsCell culture
Human osteosarcoma MG-63 cells (Interlab Cell Line Collection,Genova, Italy) were routinely grown as previously described (DeBlasio et al., 2003). For production of 3AB-OS cells, MG-63 cellswere seeded on 10 cm tissue culture dishes at 0.5� 106 cells/dishand treated either with 5 mM 3AB or vehicle (control cells), for thetimes indicated in the results. The medium was changed every3 days and fresh dilutions of 3AB were added at each mediumchange. Stock solution of 3AB were prepared in dimethyl sulfoxide(DMSO) and diluted to final concentrations in the culture medium.The final concentration of DMSO never exceeded 0.25%, aconcentration which had no discernible effect on osteosarcomacells and which was employed as vehicle.
Assessment of cellular morphological changes
Cell morphology was evaluated using phase contrast microscopy,photographed and captured by a computer-imaging system (LeicaDC300F camera and Adobe Photoshop for image analysis).
Sphere formation assay
MG-63 and 3AB-OS cells were plated at a density of 6� 104 cells/well in 6-well ultra low attachment plates (Corning, Inc., Corning,
JOURNAL OF CELLULAR PHYSIOLOGY
NY) in N2 medium with 1% methylcellulose. N2 medium consistsof 2�DMEM/F12 with progesterone (20 nM), putrescine(100 mM), sodium selenite (30 nM), transferrin (25 mg/ml), insulin(20 mg/ml), human recombinant epidermal growth factor (EGF;10 ng/ml) and basic fibroblast growth factor (bFGF; 10 ng/ml) mixedwith an equal volume of 2% methylcellulose. Fresh aliquots of EGFand bFGF were added every other day. After 6 and 12 days ofculturing, colonies were counted by inverted phase contrastmicroscopy (Leica DC300F). Sphere assay was performedwith triplicate examination. All reagents were purchased fromSigma-Aldrich (Milan, Italy).
Rhodamine 123 exclusion assay
Suspension cells (1� 106) were incubated with complete mediumcontaining 0.5 mg/ml rhodamine 123 (Sigma-Aldrich), with orwithout 10 mM verapamil (Sigma) at 378C for 30 min. Cells werethen washed and allowed to incubate for 1 h in rhodamine-freemedium with or without 10 mM verapamil. Then Rhodaminefluorescence was measured on a flow cytometer equipped with a488 nm argon laser with 530 nm bandpass filter (FITC). At least10,000 events were analyzed for all samples and debris waseliminated by gating on forward versus side scatter; dead cells wereexcluded based on propidium iodide staining (Robey et al., 2008).All flow cytometric data were analyzed with Expo 32 software(Beckman-Coulter, Cassina De Pecchi, Milan, Italy).
Flow cytometry analysis
Cell were washed in 0.1% bovine serum albumin in phosphatebuffered saline (PBS) at 48C, and incubated with 10 ml antibody.Cells were then washed and assayed with a FACS Vantage(Becton & Dickinson, Cinasco, Milan, Italy) or Epics XL (Beckman-Coulter). Cells were incubated with antibodies against CD133,CD34, CD117 (Miltenyi Biotec, Calderara di Reno, Bologna, Italy);CD31, CD24, CD90, CD44, CD29 (BD Pharmingen, Cinasco,Milan, Italy); CD105, CD271 (Santa Cruz Biotechnology, Inc.,Segrate, Milan, Italy), on ice for 30 min. The antibodieswere FITC-, PE-, or Cy5-conjugated. For indirect labelingcells were incubated with antibodies against ABCG2, Ror2,FAK, PPARg followed by a compatible secondary antibodyFITC- or PE-conjugated (Santa Cruz Biotechnology, Inc.). Fordetection of intracellular antigens were used a fixation/permeabilization kit (CALTAG Laboratories, Torino, Italy).Expression of cell markers was determined by comparisonwith isotype control.
Hoechst 33342 exclusion assay and ABCB1/ABCG2 expression
MG-63 and 3AB-OS cells were seeded in 4-well chamber slides at1� 104 cells/well. After adhesion live cells were double labeledwith Hoechst 33342 dye (bis-benzimide trihydrochloride; Sigma)and with anti-ABCB1 or anti-ABCG2 antibody (Santa CruzBiotechnology, Inc.). Hoechst 33342 dye was added at 5mg/ml, andcells were incubated at 378C for 60 min. Then, cells were washed inPBS, incubated for 1 h with 10mg/ml of anti-ABCB1 or anti-ABCG2antibody at room temperature, washed again with PBS andincubated with 1:100 dilution of FITC-conjugated secondaryantibody (Santa Cruz Biotechnology, Inc.). Finally, cells werewashed two times with PBS and visualized and captured with a LeicaDC300F microscope with epifluorescent filters for Hoechstand FITC.
Immunofluorescence microscopy
Cells were fixed with methanol/acetic acid (3:1), washed with PBSand incubated with Hoechst 33342 for 10 min. After washing withPBS cells were incubated for 1 h with 10 mg/ml of anti-Oct3/4,anti-nucleostemin or anti-Nanog antibody (Santa CruzBiotechnology, Inc.), washed with PBS to remove excess antibodyand probed with FITC-conjugated secondary antibody (1:100)(Santa Cruz Biotechnology, Inc.) for 1 h at room temperature.

E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 303
The images were visualized with Leica DC300F microscope withepifluorescent filters for Hoechst and FITC.
Cell size analysis by flow cytometry
Cell suspension was centrifuged and resuspended in cold PBScontaining PI (2 mg/ml) on ice until flow cytometry was performed.A 488-nm argon laser was used as the probing beam. An indirectmeasurement of size was based on forward scatter which isproportional to cell size; the bigger the cell, the more light isscattered, the higher the detected signal. At least 10,000 eventswere analyzed for each sample and debris was eliminated by gatingon forward versus side scatter; dead cells were excluded based onpropidium iodide staining. All flow cytometric data were analyzedwith Expo 32 software (Beckman-Coulter).
Cell proliferation assay
MG-63 and 3AB-OS cells were plated at a density of 6� 104 cells/well in 6-well plates. Every 12 h cells were harvested andresuspended in PBS. An aliquot of cell suspension was diluted with0.4% trypan blue (Sigma–Aldrich), pipetted onto a hemocytometerand counted under a microscope at 200� magnification. Live cellsexcluded the dye, whereas dead cells admitted the dye andconsequently stained intensely with trypan blue. The number ofviable cells for each experimental condition was counted andrepresented on a linear graph. Doubling time (td) was estimated bythe following equation: td¼ (t2� t1) ln2/lnX2/X1; where X2 andX1 are the number of cells at t2 and t1.
Western blot analysis
Cells were washed in PBS and incubated in ice-cold lysis buffercontaining protease inhibitor cocktail for 20 min, as previouslyreported (Lauricella et al., 2006). Then, cells were sonicated threetimes for 10 sec. Equal amounts of protein samples (60 mg/lane)were subjected to SDS–polyacrylamide gel electrophoresis andthen transferred to a nitrocellulose membrane for detectionwith specific antibodies (Santa Cruz Biotechnology, Inc.). Thecorrect protein loading was verified by means of both redPonceau staining and immunoblotting for actin. Bands werevisualized with Chemidoc System and analyzed with QuantityOne software. The comparative CT method was used todetermine the fold of protein expression, normalized by internalcontrol actin.
Preparation of nuclear and cytosolic proteins
Nuclear and cytosolic proteins extracts were prepared accordingto the method described by Schreiber et al. (1989). Briefly, the cellswere collected, washed with PBS and resuspended in 400 ml coldbuffer containing 10 mM N-2-hydroxyethyl piperazine
TABLE 1. Primer pairs used in RT-PCR
Gene Forward primer 5(–3(
CD133 TCTTGACCGACTGAGACCCAACOct3/4 TGGAGAAGGAGAAGCTGGAGCAANanog CAAAGGCAAACAACCCACTTNucleostemin GGGAAGATAACCAAGCGTGTGRor2 CCTGGACACGACAGACACTGFAK GCGCTGGCTGGAAAAAGAGGAARunx2 CTCCCTGAACTCTGCACCAAGTCOsterix GCCAGAAGCTGTGAAACCTCALP CACTGCGGACCATTCCCACGTCTOsteocalcin CCCTCACACTCCTCGCCCTATTPPARg ACCCCTATTCCATGCTGTATGb-catenin CGTGGACAATGGCTACTCAAGCABCB1 CCCATCATTGCAATAGCAGGABCG2 AGTTCCATGGCACTGGCCATAGAPDH TGACATCAAGAAGGTGA
JOURNAL OF CELLULAR PHYSIOLOGY
ethanesulfonic acid (HEPES, pH 7.9), 10 mM KCl, 0.1 mMethylenediamine tetraacetic acid (EDTA), 3 mM MgCl2, 1 mMdithiothreitol (DTT), and 1 mM phenylmethylsulfonyl fluoride(PMSF), 0.7 mg/ml pepstatin, 1 mg/ml leupeptin, and 10 mg/mlaprotinin. After swelling on ice for 15 min, cells were lysed byadding 25 ml NP-40 10% and gently passed through a 27-gaugeneedle. The extract was centrifuged at 48C for 5 min at 400g,obtaining a nuclear pellet (fraction A) and a supernatant (fractionB). Fraction A was resuspended in 50ml of buffer containing 20 mMHEPES, pH 7.9, 400 mM KCl, 1 mM EDTA, 1 mM ethylene glycoltetraacetic acid, 1 mM DTT, 1 mM PMSF, 0.7 mg/ml pepstatin,1 mg/ml leupeptin and 10 mg/ml aprotinin. The tube was vigorouslyrocked on shaking platform at 48C for 30 min and centrifuged for5 min at 48C at 12,000g. The supernatant was used as nuclearextract. Fraction B was centrifuged for 20 min at 48C at 12,000gand the supernatant was employed as cytosolic fraction. Proteinextracts were stored at �808C until testing.
RT-PCR analysis
RNA was isolated using RNAsy mini kit (Qiagen, Milan, Italy).cDNA was amplified from 1 mg of RNA and PCR was performedas previously reported (De Blasio et al., 2003). The amplifiedproducts were resolved by agarose gel electrophoresis (1%agarose, 0.5 mg/ml ethidium bromide) then visualized withChemidoc System and data were analyzed with Quantity Onesoftware. The comparative CT method was used to determine thefold of gene expression in the samples, normalized by internalcontrol GAPDH. The primers (Proligo USA, Milan, Italy) areprovided in Table 1.
ResultsProduction of 3AB-OS cells from human osteosarcomaMG-63 cells by prolonged 3AB treatment
In Figure 1 human osteosarcoma MG-63 cells, seeded on 10 cmtissue culture dishes, were allowed to adhere overnight andthen were treated either with vehicle (control) or with 5 mM3AB as described in Materials and Methods Section. Cultureswere analyzed by microscopy daily and imagines were acquiredby a light microscope equipped with an analyzing chamber. Ascells reached confluence, cultures were harvested with trypsin-EDTA and passaged to a new dish for further expansion. Until5 days of incubation the cells grew, under the two experimentalconditions, as already previously described (De Blasio et al.,2003). More precisely, control MG-63 cells progressivelyformed a cell population that assumed a spindle-shapemorphology in confluent wave-like (A), while 48–72 h of 3ABtreatment, produced a cell population exhibiting a progressivelowering in cell number with the appearance of a cell population
Reverse primer 5(–3(
ACTTGATGGATGCACCAAGCACAA GGCAGATGGTCGTTTGGCTGAATA
ATTGTTCCAGGTCTGGTTGCCCTCCAAGAAGTTTCCAAAGGAAGTTATGATTTGGGATGTGCTCGGTGGGTGCTGGCTGGTAGG
CT GGGGTGGTAGAGTGGATGGACGGCTGCAAGCTCTCCATAACC
T GCGCCTGGTAGTTGTTGTGAGCATAAAGCCGATGTGGTCAGCCAACTCGTAAGGAATCGCTTTCTGGGTCTCTGAGCTCGAGTCATTGCAGAGGAAGTTCAAACTTCTGCTCCTGATCAGGTAGGCAATTGTGAGGTCCACCACCCTGTTGCTGTA

Fig. 1. Production of 3AB-OS cells from human osteosarcoma MG-63 cells by prolonged 3AB treatment. The figure describes the morphologyof MG-63 cells treated either with vehicle (control, A) or with 5 mM 3AB (B–H) analyzed by contrast phase microscopy as described in Materialsand Methods and in Results Sections. Original magnification 200T.
304 D I F I O R E E T A L .
exhibiting marked morphological changes of their shape, withthe formation of long dendritic-like protrusions and theappearance of osteocyte-like cells with stellate morphology (B).These cells appeared connected to each other via junctionsresembling small channels (C). Surprisingly, when 3ABtreatment was prolonged for longer time (more than 6 days),osteocyte-like cells progressively died (D) and a newheterogeneous cell population appeared which progressivelyinvaded the culture dish growing as cell cluster of rapidlyexpanding fibroblastoid cells compatible with apparentlyundifferentiated mesenchymal stem cell (MSC) morphology (E).Confluency was achieved by day 15 when the cell cluster wasdetached by trypsin-treatment and transferred to new 10 cmculture dishes in the presence of 3AB. After this first passage,the adherent cells divided more rapidly and became confluentwithin 7 days. The cells were passaged again and cultures weremaintained in the presence of 3AB for 15 passages (about100 days). Thereafter, cells were transferred in culture mediumwithout 3AB to evaluate whether the phenotypes obtained byprolonged 3AB treatment were reverted. As no reversion wasobserved, since that moment cells were grown in mediumwithout 3AB. Such a stable expanding cell populations, whichhad been passaged serially for more than 20 months (73passages) and which has a strong self-renewal capability,appeared to be composed of cells with three differentmorphologies, where two types of adherent polygonal-shaped(F) and-spindle-shaped (G) fibroblastoid cells prevailed, mixedwith single phase-bright cells of round shape, most of whichunattached or only weakly attached to the substrate (H).Conclusively, Figure 1 describes the progressive substitutionand enrichment of human osteosarcoma MG-63 cells with aheterogeneous and stable cell population composed byfibroblastoid cells, spindle-shaped, polygonal-shaped, androunded-shaped, selected by long-term 3AB treatment. Wehave indicated this stable heterogeneous cell population, whichgrows in our laboratory in the absence of 3AB since 1 year, as3AB-osteosarcoma cells (3AB-OS cells). In comparison, studieson long-term culture of MG-63 cells grown in the absence of3AB show that these cells probably undergo senescence. In fact,MG-63 cells can be reseeded for a limited number of times,as they reach confluency and maintain their spindle-shapemorphology until about 15 passages. Thereafter, by newreseeding the cells suddenly dye, apparently by necrosis(not shown).
JOURNAL OF CELLULAR PHYSIOLOGY
Analysis of multidrug resistance phenotype inMG-63 and 3AB-OS cells by rhodamine 123 andHoechst 33342 dyes exclusion assay,immunofluorescence microscopy, flow cytometry,and RT/PCR
The ability of cancer cells to exclude small lipophilic molecules,such as the fluorescent dyes Rhodamine 123 (Rho-123) andHoechst 33342, is exploited for the individuation and isolationof cells which are characterized by low-staining after beingincubated with the fluorescent dyes and which are associatedwith multidrug resistance (MDR) transporters expression.These criteria are widely accepted for defining stem cells(Hadnagy et al., 2006). Here, we have performed analysis toevaluate the ability of MG-63 and 3AB-OS cells to efflux thesedyes. The Figure 2A shows that when the cells were incubatedwith rho-123 alone (left part), in MG-63 cells the ability toexclude the dye is represented by two peaks, one at lowerfluorescence (a, with major ability to exclude rho-123) and oneat higher fluorescence (b, with minor ability to exclude rho-123). Moreover, simultaneous incubation of the cells with rho-123 and 10 mM verapamil, potent inhibitor of P-glycoprotein/ABCB1 (right part), induces the disappearing of the lowfluorescence peak, thus suggesting that this peak can beassociated to the ABCB1 activity. In 3AB-OS cells (Fig. 2B) theincubation with rho-123, either with or without verapamil,demonstrates the presence of only the higher fluorescencepeak (b) which was not influenced by verapamil treatment, thussuggesting that 3AB-OS cells does not express the ABCB1protein. Analysis by fluorescence microscopy performed bydouble labeling with Hoechst 33342 dye and anti-ABCB1 oranti-ABCG2 antibodies, show in MG-63 cells (Fig. 2C,E, leftparts) a higher blue fluorescence intensity than in 3AB-OS cells(Fig. 2D,F, left parts); the Figure 2 also shows that ABCB1transporter appears to be well expressed in MG-63 cells(C, middle part) but not in 3AB-OS cells (D, middle part),whereas ABCG2 transporter is much less expressed in MG-63cells (E, middle part) than in 3AB-OS cells (F, middle part).Finally, in Figure 2 merge analysis (C–F, right parts) showsthat the increased capacity to efflux Hoechst dye stronglycorrelated with ABCG2 transporter expression. In Figure 2cytofluorimetric analysis confirms that, with respect to MG-63cells (G), 3AB-OS cells (H) express a much higher percentage ofABCG2-positivity. Comparative analysis of ABCB1 and ABCG2
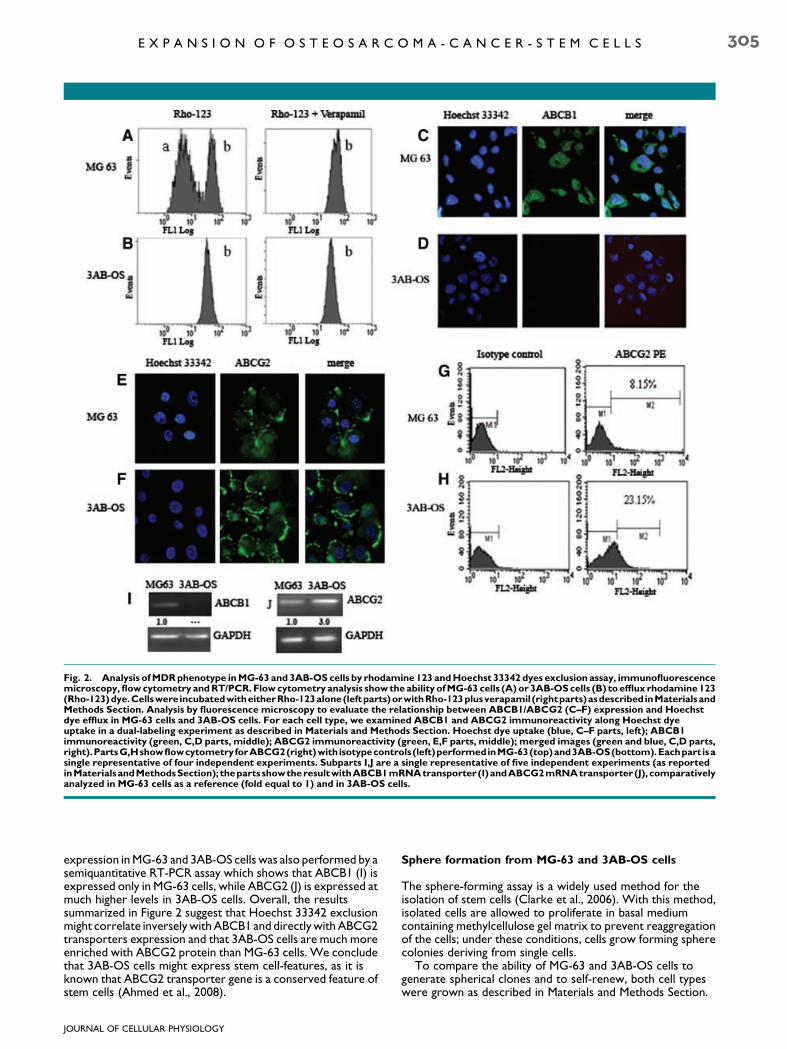
Fig. 2. Analysis of MDR phenotype in MG-63 and 3AB-OS cells by rhodamine 123 and Hoechst 33342 dyes exclusion assay, immunofluorescencemicroscopy, flow cytometry and RT/PCR. Flow cytometry analysis show the ability of MG-63 cells (A) or 3AB-OS cells (B) to efflux rhodamine 123(Rho-123)dye.Cellswere incubatedwitheitherRho-123alone(leftparts)orwithRho-123plusverapamil (rightparts)asdescribedinMaterialsandMethods Section. Analysis by fluorescence microscopy to evaluate the relationship between ABCB1/ABCG2 (C–F) expression and Hoechstdye efflux in MG-63 cells and 3AB-OS cells. For each cell type, we examined ABCB1 and ABCG2 immunoreactivity along Hoechst dyeuptake in a dual-labeling experiment as described in Materials and Methods Section. Hoechst dye uptake (blue, C–F parts, left); ABCB1immunoreactivity (green, C,D parts, middle); ABCG2 immunoreactivity (green, E,F parts, middle); merged images (green and blue, C,D parts,right).PartsG,HshowflowcytometryforABCG2(right)withisotypecontrols(left)performedinMG-63(top)and3AB-OS(bottom).Eachpart isasingle representative of four independent experiments. Subparts I,J are a single representative of five independent experiments (as reportedinMaterialsandMethodsSection); theparts showtheresultwithABCB1mRNAtransporter(I)andABCG2mRNAtransporter (J), comparativelyanalyzed in MG-63 cells as a reference (fold equal to 1) and in 3AB-OS cells.
E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 305
expression in MG-63 and 3AB-OS cells was also performed by asemiquantitative RT-PCR assay which shows that ABCB1 (I) isexpressed only in MG-63 cells, while ABCG2 (J) is expressed atmuch higher levels in 3AB-OS cells. Overall, the resultssummarized in Figure 2 suggest that Hoechst 33342 exclusionmight correlate inversely with ABCB1 and directly with ABCG2transporters expression and that 3AB-OS cells are much moreenriched with ABCG2 protein than MG-63 cells. We concludethat 3AB-OS cells might express stem cell-features, as it isknown that ABCG2 transporter gene is a conserved feature ofstem cells (Ahmed et al., 2008).
JOURNAL OF CELLULAR PHYSIOLOGY
Sphere formation from MG-63 and 3AB-OS cells
The sphere-forming assay is a widely used method for theisolation of stem cells (Clarke et al., 2006). With this method,isolated cells are allowed to proliferate in basal mediumcontaining methylcellulose gel matrix to prevent reaggregationof the cells; under these conditions, cells grow forming spherecolonies deriving from single cells.
To compare the ability of MG-63 and 3AB-OS cells togenerate spherical clones and to self-renew, both cell typeswere grown as described in Materials and Methods Section.

Fig. 3. Sphere formation from MG-63 and 3AB-OS cells. Phase contrast images of monoclonal sarcospheres formed from self-renewing cells.Sarcospheres were formed from MG-63 cells and 3AB-OS cells plated at a density of 6 T 104 cells/well in 6-well ultra low attachment plates in N2medium with 1% methylcellulose as described in Materials and Methods Section. After 6 and 12 days of culturing, the colonies were quantitated byinverted phase contrast microscopy.
306 D I F I O R E E T A L .
Figure 3 shows that MG-63 and 3AB-OS cells were bothcapable of forming spheres, but, after 6 days in culture,those formed by 3AB-OS cells were more numerous and largerthan those formed by MG-63 cells. In particular, MG-63 cellsformed spheres at a frequency of approximately 1/750(80.00� 11.2 colonies/6� 104 cells), while 3AB-OS cellsformed spheres at a frequency of approximately 1/350(171.42� 18.4 colonies/6� 104cells). After 12 days thedimension of the spheres was similar in MG-63 and 3AB-OScells, but the number of spheres formed by 3AB-OS cells wasmuch greater (>69%) than with MG-63 cells. To investigate theability of the cells for self-renewing in the sarcosphere system,cultured MG-63 and 3AB-OS spheres were dissociated tosingle cells and allowed to grow in monolayers to 70%confluence and then reseeded as single cells in 1%methylcellulose media. The results demonstrated the ability toself-renew via the formation of secondary spheres repeatedly(not shown).
Characterization of 3AB-OS cells by proliferativepotential, cell size and cell cycle regulators
In addition to self-renewal ability, CSCs are characterized byhigh proliferative potential after placement in culture (Baguley,2006), small cell size (De Paiva et al., 2006), and special cellcycle regulator expression (Herrera et al., 1996). Thus, weperformed experiments to compare MG-63 and 3AB-OS cellsfor these characters, aimed at evidencing in 3AB-OS cells somedistinctive properties of stem cells. The results are summarizedin Figure 4. As shown in part A, with respect to MG-63 cells,3AB-OS cells possess a higher proliferative output, exhibiting adoubling time of approximately 22 h, whereas MG-63 cellsshow a doubling time of 28 h; in addition, 3AB-OS cells areendowed with characteristics similar to those of progenitorcells as they can be reseeded unlimitedly without losing theirproliferative potential (not shown). In contrast, as abovedescribed, MG-63 cells reach confluence with a homogeneousmorphology and after confluency, can be reseeded a limitednumber of times (�15 times) after which irreversibly lost theirproliferative potential, undergoes senescence and dye (notshown). The unlimited replicative potential of 3AB-OS cells is
JOURNAL OF CELLULAR PHYSIOLOGY
also reported in Figure 4E where Western blotting analysisshows high levels of human telomerase reverse transcriptase(hTERT) expression, while the protein resulted unmeasurablein MG-63 cells. Small size seems to represent one of theimportant features of stem cells. Although the role andmechanism of cell size in stem cell regulation have not beencompletely defined, it has been reported that cell size is relatedto cell cycle, cell proliferation, and differentiation. To analyzethis parameter, we performed flow cytometry based onforward scatter, which is an indirect measurement of size. InFigure 4B comparative cytofluorimetric analysis indicates that3AB-OS cells (a) consisted of cells which are extremely smallerthan MG-63 cells (b). We then allowed to characterize the cellsby cell cycle markers, starting from the functional analysis ofthe retinoblastoma protein (pRb) which is known to influencethe timing of G1-S and that, when absent or altered, leads toan abbreviated G1 phase with reduced cell size. We havepreviously shown that in MG-63 cells, pRb is constitutivelyinactivated by hyperphosphorylation (De Blasio et al., 2005) andthat this is one of the reasons for the deregulation of cellproliferation. Here (Fig. 4C), we compare the status of Rbphosphorylation and that of some key regulators of cell cycleprogression in the two cell lines. The analysis of the functionalstatus of the G0/G1 restriction marker pRB evidences that,respect to MG-63 cells, 3AB-OS cells express much higherlevels of ppRb, the hyperphosphorylated/inactive Rb form (a,left part), accompanied by a potent increase in cyclin D1 (a, rightpart) responsible for mediating pRB phosphorylation with theconsequent G1-S phase progression. Since in a number ofcancer cells cyclin D1 is under the control of the aberrantnuclear translocation of b-catenin which also stronglycorrelated to cancer invasivity (Hoang et al., 2004; Shiratsuchiet al., 2007), we examined the levels ofb-catenin in 3AB-OS andin MG-63 cells and its localization. In Figure 4C RT/PCR (b, leftpart) and Western blot analysis of whole cell extracts (b, rightpart) show that, with respect to MG-63 cells, 3AB-OS cellsexpress much higher levels of b-catenin. Furthermore,localization analysis (c) demonstrated that in 3AB-OS cells thenuclear level of b-catenin was much higher than in MG-63 cells.Overall, these results suggest a much higher invasive potentialfor 3AB-OS than for MG-63 cells. Moreover, Figure 4D shows
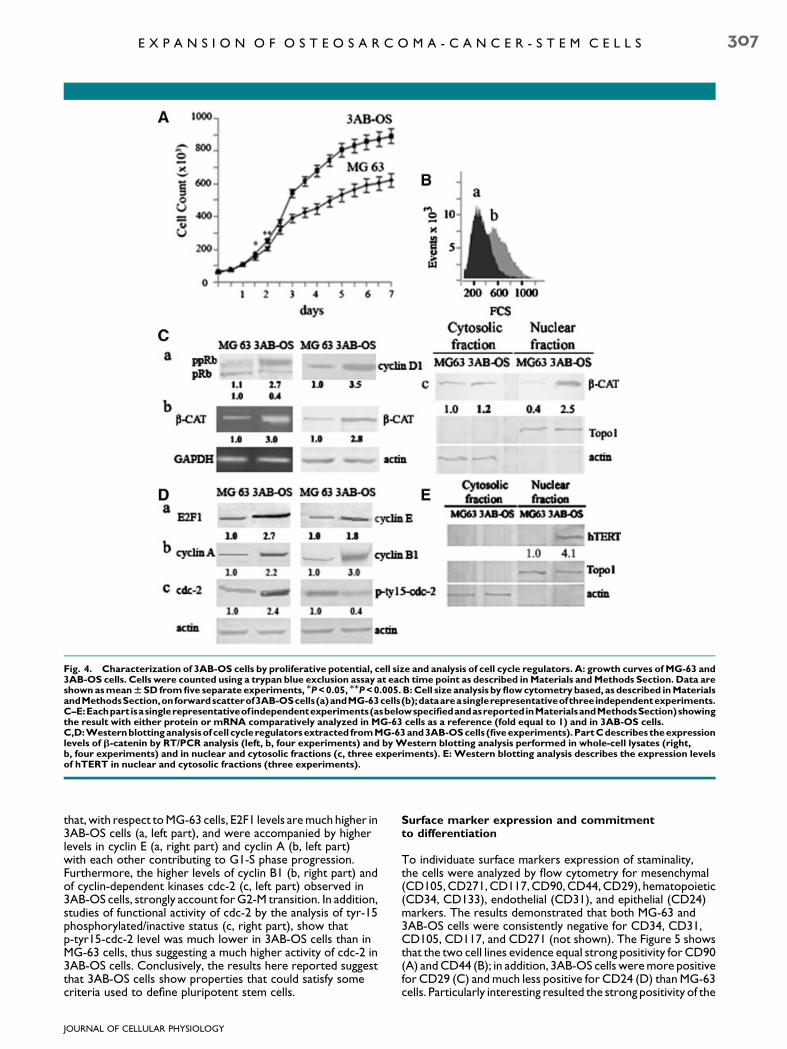
Fig. 4. Characterization of 3AB-OS cells by proliferative potential, cell size and analysis of cell cycle regulators. A: growth curves of MG-63 and3AB-OS cells. Cells were counted using a trypan blue exclusion assay at each time point as described in Materials and Methods Section. Data areshown as mean W SD from five separate experiments, MP < 0.05, MMP < 0.005. B: Cell size analysis by flow cytometry based, as described in MaterialsandMethodsSection,onforwardscatterof3AB-OScells(a)andMG-63cells(b);dataareasinglerepresentativeofthreeindependentexperiments.C–E:Eachpart isasinglerepresentativeof independentexperiments(asbelowspecifiedandasreportedinMaterialsandMethodsSection)showingthe result with either protein or mRNA comparatively analyzed in MG-63 cells as a reference (fold equal to 1) and in 3AB-OS cells.C,D:Westernblotting analysisof cell cycleregulators extracted fromMG-63 and3AB-OS cells (fiveexperiments).PartCdescribes theexpressionlevels of b-catenin by RT/PCR analysis (left, b, four experiments) and by Western blotting analysis performed in whole-cell lysates (right,b, four experiments) and in nuclear and cytosolic fractions (c, three experiments). E: Western blotting analysis describes the expression levelsof hTERT in nuclear and cytosolic fractions (three experiments).
E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 307
that, with respect to MG-63 cells, E2F1 levels are much higher in3AB-OS cells (a, left part), and were accompanied by higherlevels in cyclin E (a, right part) and cyclin A (b, left part)with each other contributing to G1-S phase progression.Furthermore, the higher levels of cyclin B1 (b, right part) andof cyclin-dependent kinases cdc-2 (c, left part) observed in3AB-OS cells, strongly account for G2-M transition. In addition,studies of functional activity of cdc-2 by the analysis of tyr-15phosphorylated/inactive status (c, right part), show thatp-tyr15-cdc-2 level was much lower in 3AB-OS cells than inMG-63 cells, thus suggesting a much higher activity of cdc-2 in3AB-OS cells. Conclusively, the results here reported suggestthat 3AB-OS cells show properties that could satisfy somecriteria used to define pluripotent stem cells.
JOURNAL OF CELLULAR PHYSIOLOGY
Surface marker expression and commitmentto differentiation
To individuate surface markers expression of staminality,the cells were analyzed by flow cytometry for mesenchymal(CD105, CD271, CD117, CD90, CD44, CD29), hematopoietic(CD34, CD133), endothelial (CD31), and epithelial (CD24)markers. The results demonstrated that both MG-63 and3AB-OS cells were consistently negative for CD34, CD31,CD105, CD117, and CD271 (not shown). The Figure 5 showsthat the two cell lines evidence equal strong positivity for CD90(A) and CD44 (B); in addition, 3AB-OS cells were more positivefor CD29 (C) and much less positive for CD24 (D) than MG-63cells. Particularly interesting resulted the strong positivity of the
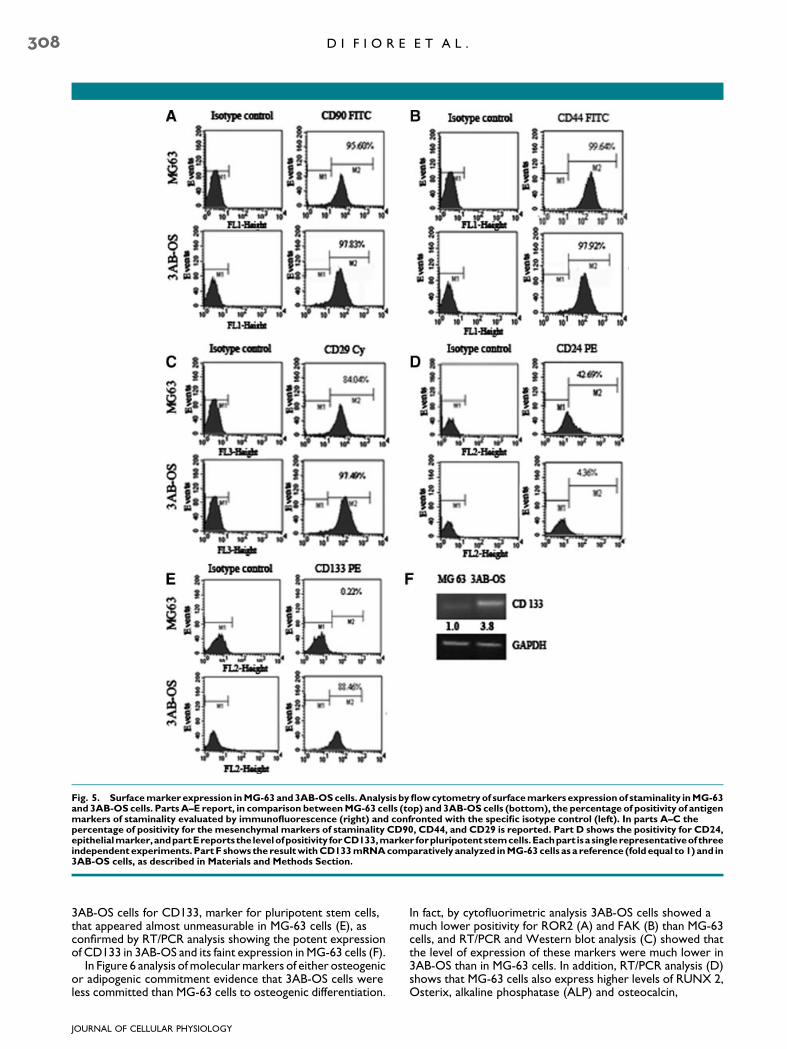
Fig. 5. Surface marker expression in MG-63 and 3AB-OS cells. Analysis by flow cytometry of surface markers expression of staminality in MG-63and 3AB-OS cells. Parts A–E report, in comparison between MG-63 cells (top) and 3AB-OS cells (bottom), the percentage of positivity of antigenmarkers of staminality evaluated by immunofluorescence (right) and confronted with the specific isotype control (left). In parts A–C thepercentage of positivity for the mesenchymal markers of staminality CD90, CD44, and CD29 is reported. Part D shows the positivity for CD24,epithelialmarker,andpartEreportsthe levelofpositivity forCD133,marker forpluripotentstemcells.Eachpart isasinglerepresentativeof threeindependent experiments. Part F shows the result with CD133 mRNA comparatively analyzed in MG-63 cells as a reference (fold equal to 1) and in3AB-OS cells, as described in Materials and Methods Section.
308 D I F I O R E E T A L .
3AB-OS cells for CD133, marker for pluripotent stem cells,that appeared almost unmeasurable in MG-63 cells (E), asconfirmed by RT/PCR analysis showing the potent expressionof CD133 in 3AB-OS and its faint expression in MG-63 cells (F).
In Figure 6 analysis of molecular markers of either osteogenicor adipogenic commitment evidence that 3AB-OS cells wereless committed than MG-63 cells to osteogenic differentiation.
JOURNAL OF CELLULAR PHYSIOLOGY
In fact, by cytofluorimetric analysis 3AB-OS cells showed amuch lower positivity for ROR2 (A) and FAK (B) than MG-63cells, and RT/PCR and Western blot analysis (C) showed thatthe level of expression of these markers were much lower in3AB-OS than in MG-63 cells. In addition, RT/PCR analysis (D)shows that MG-63 cells also express higher levels of RUNX 2,Osterix, alkaline phosphatase (ALP) and osteocalcin,

Fig. 6. Commitment of MG-63 and 3AB-OS cells to differentiation. Parts A–F report the analysis of molecular markers of osteogenic andadipogenic differentiation in MG-63 cells (left) and 3AB-OS cells (right). In A,B,E, flow cytometry shows the percentage of ROR2, FAK (markers ofosteogenic differentiation) andPPARg (marker ofadipogenicdifferentiation) positivity (b) inconfrontwith isotype control (a).Each part isa singlerepresentative of three independent experiments. C,D,F show the result with either protein or mRNA comparatively analyzed in MG-63 cells as areference (fold equal to 1) and in 3AB-OS cells, as reported in Materials and Methods Section. C,D show the expression levels of the osteogenicdifferentiation markers ROR2, FAK, Runx2, Osterix, alkaline phosphatase (ALP), and osteocalcin (OSTEOC). F shows the levels of PPARg.Each part is a single representative of four independent experiments.
E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 309
mesenchymal markers of osteogenic differentiation. Similarly,3AB-OS cells were less committed than MG-63 cells toadipogenic differentiation, as suggested by the analysis of theadipogenic marker PPARg performed by cytofluorimetry (E),which shows an higher positivity of MG-63 cells, and alsoconfirmed by RT/PCR and Western blot analysis (F) showingthe stronger levels of expression of this marker in MG-63 cells.
Expression of genes required for the maintaining of stemcell state and for the inhibition of apoptosis
Embryonic stem cells are immortal and pluripotent cellscharacterized by the expression of a group of genes (stemnessgenes) required to maintain the stem cell-like feature. Thesegenes include Oct3/4 and Nanog—transcription factors whichform the core regulatory network that ensures both thesuppression of genes that lead to differentiation and themaintenance of pluripotency (Pan and Thomson, 2007) andnucleostemin—a putative GTPase preferentially expressed inthe nucleoli of embryonic stem cells and several cancer celllines—which play a key role in controlling cell cycleprogression (Ye et al., 2008). In Figure 7 the comparative
JOURNAL OF CELLULAR PHYSIOLOGY
analysis of these genes performed by immunofluorescence inMG-63 cells and 3AB-OS cells, shows that although both the celllines expressed Oct3/4 (A), nucleostemin (B), and Nanog (C),nevertheless in 3AB-OS cells these proteins— in particularNanog—appeared more potently expressed than in MG-63cells, as also confirmed by RT/PCR analysis (D).
Among the signals that influence division, differentiation, andfunction of stem cells, oxygen availability plays a key role andhypoxia-inducible factor HIF-1a is the key molecule which canregulate stem cell function through activation of a number ofgenes which contribute to their survival. In a number ofconditions HIFs upregulated Oct3/4 expression, suggesting aspecific mechanism whereby HIFs can affect stem cell functionand promote tumor growth (Semenza, 2007). Here Westernblotting analysis (E) demonstrates that in MG-63 cells HIF-1alevel was almost unmeasurable, whereas it was potentlyexpressed in 3AB-OS cells. The results complexively suggestthat 3AB-OS cells expressed much higher levels of staminalitymarkers than MG-63 cells.
Malignant cells commonly have defects in cell death controland loss of apoptosis can lead to tumorigenesis. Among theknown apoptotic inhibitors, the anti-apoptotic proteins Bcl-2,
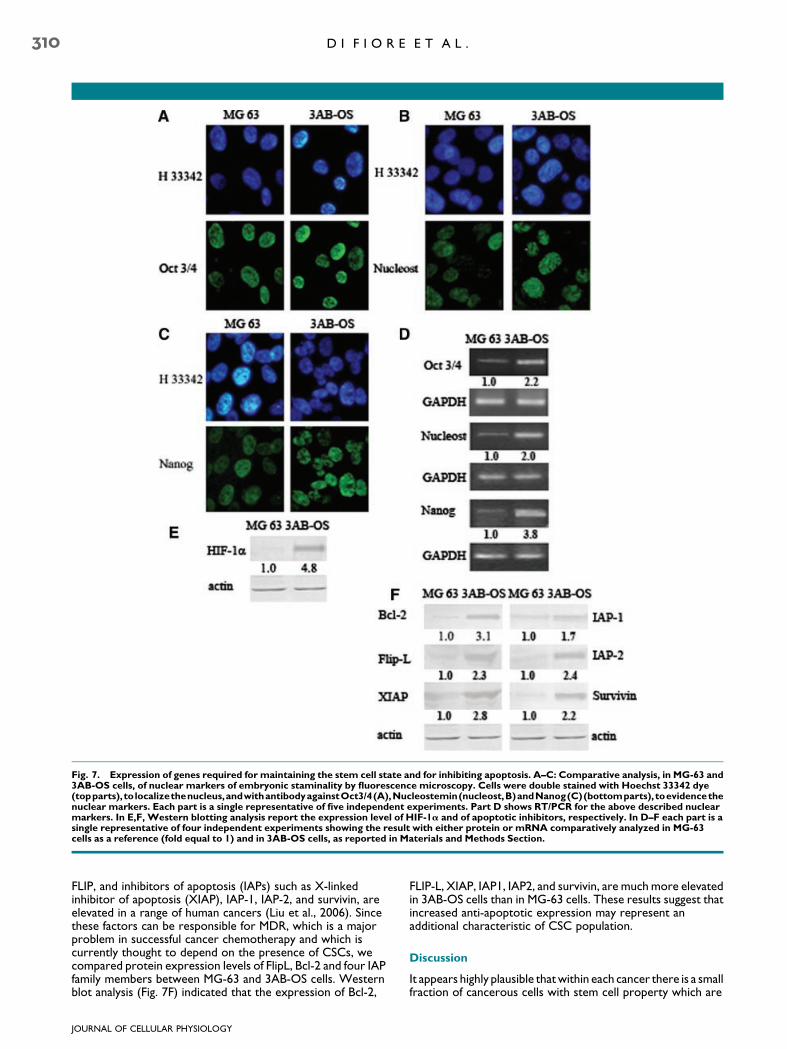
Fig. 7. Expression of genes required for maintaining the stem cell state and for inhibiting apoptosis. A–C: Comparative analysis, in MG-63 and3AB-OS cells, of nuclear markers of embryonic staminality by fluorescence microscopy. Cells were double stained with Hoechst 33342 dye(topparts), tolocalizethenucleus,andwithantibodyagainstOct3/4(A),Nucleostemin(nucleost,B)andNanog(C)(bottomparts), toevidencethenuclear markers. Each part is a single representative of five independent experiments. Part D shows RT/PCR for the above described nuclearmarkers. In E,F, Western blotting analysis report the expression level of HIF-1a and of apoptotic inhibitors, respectively. In D–F each part is asingle representative of four independent experiments showing the result with either protein or mRNA comparatively analyzed in MG-63cells as a reference (fold equal to 1) and in 3AB-OS cells, as reported in Materials and Methods Section.
310 D I F I O R E E T A L .
FLIP, and inhibitors of apoptosis (IAPs) such as X-linkedinhibitor of apoptosis (XIAP), IAP-1, IAP-2, and survivin, areelevated in a range of human cancers (Liu et al., 2006). Sincethese factors can be responsible for MDR, which is a majorproblem in successful cancer chemotherapy and which iscurrently thought to depend on the presence of CSCs, wecompared protein expression levels of FlipL, Bcl-2 and four IAPfamily members between MG-63 and 3AB-OS cells. Westernblot analysis (Fig. 7F) indicated that the expression of Bcl-2,
JOURNAL OF CELLULAR PHYSIOLOGY
FLIP-L, XIAP, IAP1, IAP2, and survivin, are much more elevatedin 3AB-OS cells than in MG-63 cells. These results suggest thatincreased anti-apoptotic expression may represent anadditional characteristic of CSC population.
Discussion
It appears highly plausible that within each cancer there is a smallfraction of cancerous cells with stem cell property which are

E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 311
responsible for initiating and maintaining the tumor (Bjerkviget al., 2005). Thus, now, a fundamental problem in cancerresearch is the identification within a tumor of the CSCs. Thecancer stem-cell hypothesis has important implications for earlydetection, prevention, and treatment of cancer, as theseaberrant stem cells may provide targets for the development oftherapeutic strategies and the studies in this field can helpelucidate the origin of the molecular complexity of humancancers. Here we report that long-term treatment of humanosteosarcoma MG-63 cells with 3AB irreversibly selected aphenotype stem-like cells which possess a number of markergenes of pluripotent embryonic and MSCs. MSCs are raremultipotent progenitor cells, self-renewable, with the capacityto differentiate into several distinct mesenchymal lineages,including bone, cartilage, adipose, tendon, and muscle tissue.These cells exhibit a unique in vitro expansion capacity and,interestingly, also have the potential to trans-differentiate intoother types of tissue, indicating pluripotency (Reger et al.,2008). Defining a MSC remains controversial since there is littleconsensus concerning their characteristics, which reportedlydiffer among species, laboratories and isolation techniques.Thus, these cells are currently defined by a combination ofphysical, morphologic, phenotypic, and functional properties,none of which have been demonstrated to be physiologic. Morerecent studies have identified MSC by positive selection basedon a number of surface antigens associated with MSCs, such asCD13, CD29, CD44, CD73 (SH3/SH4), CD90, CD105 (SH2),CD166, STRO-1 and others (Park et al., 2007; Nadri et al.,2008). To date, all MSC populations analyzed by clonal assaysare heterogeneous, with individual cells capable of varyingdifferentiation potential and expansion capacity.
Osteosarcoma is among a group of mesenchymalmalignancies that exhibits clinical, histologic, and molecularheterogeneity (Tang et al., 2008). The biology ofsarcomagenesis is understood poorly. In our laboratory wehave recently demonstrated that treatment with 3AB initiallyallows MG-63 human osteosarcoma cells to reduce the growthfor entering osteocyte differentiation. Thereafter, 3ABtreatment induced osteocytes death accompanied byprogressive enrichment with a new heterogeneous cellpopulation composed by fibroblastoid cells. The paper alsodemonstrates that the prolonged treatment with 3ABirreversibly selected and expanded this heterogeneous cellpopulation which express a number of marker genes of bothpluripotent embryonic stem cells and MSCs. These cellsrepresent a stable cell population which, after the cells had beenpassaged serially for more than 20 months (73 passages), stillretain its characteristics which were maintained even afterwithdrawn of 3AB. Thus, these cells, which we have indicated as3AB-OS cells, are growing in our laboratory in the absence of3AB since 1 year. The ability of cancer cells to exclude smalllipophilic molecules such as the fluorescent dyes Rhodamine123 and Hoechst 33342 depends on the presence of ABC(ATP-binding cassette) transporters actively pumping out thesemolecules and it is exploited for the identification of putativestem cells in a variety of human cancer (Challen and Little,2006). The emergence of drug-resistant tumor cells remains amajor clinical problem for cancer treatment. Numerous clinicaldata revealed that the MDR phenotype in tumors is associatedwith the overexpression of certain ABC transporters (Bunting,2002). The P-glycoprotein (Pgp/MDR1/ABCB1)-mediatedmultidrug resistance was the first discovered and probably stillis the most widely observed mechanism in clinical MDR (Ieiriet al., 2004). There are two other ABC transporters, whichhave been definitively demonstrated to participate in the MDRof tumors: the MDR protein 1 (MRP1/ABCC1), and themitoxantrone resistance protein (MXR/BCRP/ABCG2)(Bart et al., 2004; Choi, 2005). In our study we have shown that3AB-OS cells can efflux Hoechst 33342 dye more efficaciously
JOURNAL OF CELLULAR PHYSIOLOGY
than MG-63 cells, whereas were totally unable to effluxRhodamine 123 dye which efficiently was extruded fromMG-63 cells. In addition, 3AB-OS cells showed a strongexpression of ABCG2 and did not express ABCB1, whileMG-63 cells displayed a strong expression of ABCB1 (alsoconfirmed by its potent inhibition induced by verapamil) and aweak expression of ABCG2. The expression of ABCG2 wasrecently reported in a number of cancers (leukemia, germ cellcancers, as well as cancers of the breast, prostate and lung) andit was interpreted as a conserved feature of stem cells(Abbott, 2003; Doyle and Ross, 2003; Kawabata et al., 2003;Hoei-Hansen et al., 2006).
In addition to MDR expression, we have comparativelyevaluated in 3AB-OS and MG-63 cells, sphere formation, cellsize, cell cycle with correlated gene expression, growth curvewith proliferative potential, surface marker expression andcommitment to differentiation expression, together withexpression of genes required for the maintaining of stemcell state and for inhibiting apoptosis.
The capacity to form sarcospheres and to self-renew in aculture system is an important requirement to suggest thepresence of stem cells in a tumor and this has been recentlyshown for a number of tumors (Clarke et al., 2006). 3AB-OScells formed spheres at a higher frequency than MG-63 cells,possess a much greater self-renewing ability and a higherproliferative output; 3AB-OS cells are endowed withcharacteristics similar to those of stem cells since they can bereseeded unlimitedly without losing their proliferativepotential, while MG-63 cells after confluency can be reseededonly a limited number of times. This character is also supportedby the results showing that, in contrast to MG-63 cells, 3AB-OScells express high levels of hTERT, catalytical subunit oftelomerase, known to be correlated to the unlimitedproliferation capacity of malignant cells (Burns et al., 2005).
Respect to MG-63 cells, 3AB-OS cells show shortened cellcycle and smaller size, express an higher levels ofhyperphosphorylated, inactive form of Rb, accompanied byhigher levels in cyclin D1, cyclin E and E2F1 which may accountfor the accelerated G1/S transition; they also exhibit muchhigher levels of cyclin A and cyclin B1, with a much higher level ofCdc-2 activity accounting for mitosis activation (Deshpandeet al., 2005; Stark and Taylor, 2006); 3AB-OS cells also displayeda much higher level of nuclear b-catenin, which is known tocontrol cyclin D1 expression and that, together with CD29, isinvolved in the regulation of integrin-dependent mechanisms ofpluripotent stem cells proliferation (Hoang et al., 2004;Hartmann, 2006; Chiba et al., 2007); in addition, 3AB-OS cellswere much less committed than MG-63 cells to osteogenicdifferentiation, displaying a lower level of ROR2, FAK, RUNX2,Osterix and ALP, mesenchymal markers of osteogenicdifferentiation (Liu et al., 2007; Salasznyk et al., 2007) andalso displaying a much lower adipogenic commitment, assuggested by the lower level of PPARg, mesenchymal markerof adipocytic differentiation (Lin et al., 2007; Yanagisawa et al.,2007).
Concerning the expression of genes required for themaintaining of stem cell state, likely to MG-63 cells, 3AB-OScells were strongly positive for CD90 and CD44 and morepositive for CD29 (MSC markers). Importantly, 3AB-OS cellswere strongly positive for CD133 (>88%) marker forpluripotent stem cells (Shmelkov et al., 2005; Neuzil et al.,2007), and express high levels of the POU transcription factorOct3/4, Nanog and nucleostemin, markers characteristic forembryonic stem cells, epiblast stem cells and primordial germcells (Ye et al., 2008) which are critical for maintainingembryonic stem cell character and promoting pluripotency,allowing propagation over multiple passages (Darr et al., 2006).In MG-63 cells CD133 was almost unmeasurable and Oct3/4,Nanog and nucleostemin were only dimly positive.

312 D I F I O R E E T A L .
It is known that embryonic stem and progenitor cellsfrequently occupy hypoxic ‘‘niches’’ and respond totheir hypoxic microenvironment by stimulating severalhypoxia-inducible factors (Simon and Keith, 2008). Our resultsshow that 3AB-OS cells markedly express HIF-1a which isknown as a key molecule for stem cell survival and which wellcorrelated with the high level of Oct3/4 and Nanog, whilein MG-63 cells HIF-1a resulted almost unmeasurable.
Although the expression of ABC transporters could renderstem cells resistant to drugs, it is not the sole determinant ofresistance, as the reluctance to enter apoptosis could be equallyor more important. Here we report that, with respect toMG-63 cells, 3AB-OS cells are provided with a number ofanti-apoptotic activity (FlipL, Bcl-2, XIAP, IAP1, IAP2, andsurvivin) which may represent an additional characteristicof CSC population.
We conclude that human osteosarcoma MG-63 cellscontains a rare population of cancer stem-like cells able to self-renew. This population may be selected by 3AB treatmentwhich produces a stable, irreversible stem-like phenotype(3AB-OS cells) characterized by very high levels of CD133, highlevels of ABCG2, nucleostemin, Oct3/4 and Nanog, and anumber of other distinctive characters which render these cellsstaminal. We still do not know how 3AB treatment can inducesuch a stem cell selection and enrichment. It is possible thatlong-term treatment with 3AB kills MG-63 cells, whereas therare CD133þ population present escape from death.However, as our previous study in MG-63 cells has shown that24–72 h of 3AB treatment induces marked changes in geneexpression, it may also be possible that long-term treatmentwith 3AB induces epigenetic changes responsible for thegeneration of the 3AB-OS stem cell population. This is one ofthe questions that we are interested in exploring, in particularsince emerging evidence suggests that osteosarcoma should beregarded as a differentiation disease caused by genetic andepigenetic changes that interrupt osteoblast differentiationfrom MSCs (Tang et al., 2008). However, the findings suggestthat 3AB-OS may be a novel tumor cell line which may provideevidence for cancer development and may be useful forinvestigating the mechanisms by which CSCs may originate.We are interested in evaluating whether 3AB may be exploitedto enrich other cancer with CSCs. The identification of asubpopulation of CSCs that drives tumorigenesis andchemoresistance in osteosarcoma may lead to new approachesfor determining prognosis and optimal therapy. Initial tumorsize and response to neoadjuvant chemotherapy are today themost accepted prognostic factors used for postoperativestratification of chemotherapy. Expression patterns of stem cellmarkers, especially CD133 and ABCG2, may indicate thedifferentiated/undifferentiated state of osteosarcoma tumors,and may correlate with a favorable/unfavorable prognosis in theclinical setting. New drugs designed to efficiently kill these stemcells subpopulation of osteosarcoma may lead to more effectivetreatments for both primary tumors and metastases. We areaimed at determining which drugs are capable of killing 3AB-OScells overcoming MDR. We are performing analysis ofepigenetic changes which could indicate which gene would beresponsible for generating 3AB-OS. The evaluation of capabilityto trans-differentiate into both mesenchymal andnonmesenchymal lineages is one of our next goal. We areinterested in transplanting 3AB-OS cells and MG-63 cells intoimmunocompromised mice to evaluate whether transplantedcells form tumors.
Acknowledgments
This study was supported by grants from ‘‘Ministerodell’Istruzione, dell’Universita e della Ricerca.’’ We thankProf. G. Calvaruso for his help in cytofluorimetry analysis.
JOURNAL OF CELLULAR PHYSIOLOGY
Literature Cited
Abbott BL. 2003. ABCG2 (BCRP) expression in normal and malignant hematopoietic cells.Hematol Oncol 21:115–130.
Ahmed F, Arseni N, Glimm H, Hiddemann W, Buske C, Feuring-Buske M. 2008. Constitutiveexpression of the ATP-binding cassette transporter ABCG2 enhances the growthpotential of early human hematopoietic progenitors. Stem Cells 26:810–818.
Ailles LE, Weissman IL. 2007. Cancer stem cells in solid tumors. Curr Opin Biotechnol18:460–466.
Al-Hajj M, Clarke MF. 2004. Self-renewal and solid tumor stem cells. Oncogene 23:7274–7282.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. 2003. Prospective
identification of tumorigenic breast cancer cells. Proc Natl Acad Sci 100:3983–3988.Baguley BC. 2006. Tumor stem cell niches: A new functional framework for the action of
anticancer drugs. Recent Patents Anticancer Drug Discov 1:121–127.Bart J, Hollema H, Groen HJ, de Vries EG, Hendrikse NH, Sleijfer DT, Wegman TD, Vaalburg
W, van der Graaf WT. 2004. The distribution of drug-efflux pumps, P-gp, BCRP, MRP1 andMRP2, in the normal blood–testis barrier and in primary testicular tumours. Eur J Cancer40:2064–2070.
Bjerkvig R, Tysnes BB, Aboody KS, Najbauer J, Terzis AJA. 2005. The origin of the cancer stemcell: Current controversies and new insights. Nat Rev Cancer 5:899–904.
Bunting KD. 2002. ABC transporters as phenotypic markers and functional regulators ofstem cells. Stem Cells 20:11–20.
Burns JS, Abdallah BM, Guldberg P, Rygaard J, Schrøder HD, Kassem M. 2005. Tumorigenicheterogeneity in cancer stem cells evolved from long-term cultures of telomerase-immortalized human mesenchymal stem cells. Cancer Res 65:3126–3135.
Challen GA, Little MH. 2006. A side order of stem cells: The Sp phenotype. Stem Cells 24:3–12.Chiba T, Zheng YW, Kita K, Yokosuka O, Saisho H, OnoderaM, Miyoshi H, Nakano M, Zen Y,
Nakanuma Y, Nakauchi H, Iwama A, Taniguchi H. 2007. Enhanced self-renewal capability inhepatic stem/progenitor cells drives cancer initiation. Gastroenterology 133:937–950.
Choi CH. 2005. ABC transporters as multidrug resistance mechanisms and the developmentof chemosensitizers for their reversal. Cancer Cell Int 5:30.
Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, Visvader J, Weissman IL, WahlGM. 2006. Cancer stem cells-perspectives on current status and future directions: AACRworkshop on cancer stem cells. Cancer Res 66:9339–9344.
Dalerba P, Cho RW, Clarke MF. 2007. Cancer stem cells: Models and concepts. Annu RevMed 58:267–284.
Darr H, Mayshar Y, Benvenisty N. 2006. Overexpression of NANOG in human ES cellsenables feeder-free growth while inducing primitive ectoderm features. Development133:1193–1201.
De Blasio A, Musmeci MT, Giuliano M, Lauricella M, Emanuele S, D’Anneo A, Vassallo B,Tesoriere G, Vento R. 2003. The effect of 3-aminobenzamide, inhibitor of poly(ADP-ribose) polymerase, on human osteosarcoma cells. Int J Oncol 23:1521–1528.
De Blasio A, Messina C, Santulli A, Mangano V, Di Leonardo E, D’Anneo A, Tesoriere G,Vento R. 2005. Differentiative pathway activated by 3-aminobenzamide, an inhibitor ofPARP, in human osteosarcoma MG-63 cells. FEBS Lett 579:615–620.
De Paiva CS, Pflugfelder SC, Li DQ. 2006. Cell size correlates with phenotype andproliferative capacity in human corneal epithelial cells. Stem Cells 24:368–375.
Dean M, Fojo T, Bates S. 2005. Tumour stem cells and drug resistance. Nat Rev Cancer5:275–284.
Deshpande A, Sicinski P, Hinds PW. 2005. Cyclins and cdks in development and cancer: Aperspective. Oncogene 24:2909–2915.
Doyle LA, Ross DD. 2003. Multidrug resistance mediated by the breast cancer resistanceprotein BCRP (ABCG2). Oncogene 22:7340–7358.
Gibbs PC, Kukekov VG, Reith JD, Tchigrinova O, Suslov ON, Scott EW, Ghivizzani SC,Ignatova TN, Steindler DA. 2005. Stem-like cells in bone sarcomas: Implications fortumorigenesis. Neoplasia 7:967–976.
Graziano A, D’Aquino R, Tirino V, Desiderio V, Rossi A, Pirozzi G. 2008. The stem cellhypothesis in head and neck cancer. J Cell Biochem 103:408–412.
Hadnagy A, Gabouryb L, Beaulieua R, Balickia D. 2006. SP analysis may be used to identifycancer stem cell populations. Exp Cell Res 312:3701–3710.
Hartmann C. 2006. A Wnt canon orchestrating osteoblastogenesis. Trends Cell Biol 16:151–159.
Herrera RE, Sah VP, Williams BO, MaKela TP, Weinberg RA, Jacks T. 1996. Altered cell cyclekinetics, gene expression, and G1 restriction point regulation in Rb-deficient fibroblasts.Mol Cell Biol 16:2402–2407.
Hoang BH, Kubo T, Healey JH, Yang R, Nathan SS, Kolb EA, Mazza BA, Meyers PA, Gorlick R.2004. Dickkopf 3 inhibits invasion and motility of Saos-2 osteosarcoma cells by modulatingthe Wnt-b-catenin pathway. Cancer Res 64:2734–2739.
Hoei-Hansen CE, Sehested A, Juhler M, Lau YF, Skakkebaek NE, Laursen H, Rajpert-de MeytsE. 2006. New evidence for the origin of intracranial germ cell tumours from primordialgerm cells: Expression of pluripotency and cell differentiation markers. J Pathol 209:25–33.
Ieiri I, Takane H, Otsubo K. 2004. The MDR1 (ABCB1) gene polymorphism and its clinicalimplications. Clin Pharmacokinet 43:553–576.
Immervoll H, Hoem D, Sakariassen PØ, Steffensen OJ, Molven A. 2008. Expression of the‘‘stem cell marker’’ CD133 in pancreas and pancreatic ductal adenocarcinomas. BMCCancer 8:48.
Jordan CT, Guzman ML, Noble M. 2006. Cancer stem cells. N Engl J Med 355:1253–1261.Kawabata S, Oka M, Soda H, Shiozawa K, Nakatomi K, Tsurutani J, Nakamura Y, Doi S,
Kitazaki T, Sugahara K, Yamada Y, Kamihira S, Kohno S. 2003. Expression and functionalanalyses of breast cancer resistance protein in lung cancer. Clin Cancer Res 9:3052–3057.
Lauricella M, Emanuele S, D’Anneo A, Calvaruso G, Vassallo B, Carlisi D, Portanova P, VentoR, Tesoriere G. 2006. JNK and AP-1 mediate apoptosis induced by bortezomib in HepG2cells via FasL/caspase-8 and mitochondria-dependent pathways. Apoptosis 11:607–625.
Li L, Neaves WB. 2006. Normal stem cells and cancer stem cells: The niche matters. CancerRes 66:4553–4557.
Lin TH, Yang RS, Tang CH, Lin CP, Fu WM. 2007. PPARg inhibits osteogenesis via the down-regulation of the expression of COX-2 and iNOS in rats. Bone 41:562–574.
Liu G, Yuan X, Zeng Z, Tunici P, Ng H, Abdulkadir IR, Lu L, Irvin D, Black KL, Yu JS. 2006.Analysis of gene expression and chemoresistance of CD133þ cancer stem cells inglioblastoma. Mol Cancer 5:67.
Liu Y, Bhat RA, Seestaller-Wehr LM, Fukayama S, Mangine A, Moran RA, Komm BS, BodinePV, Billiard J. 2007. The orphan receptor tyrosine kinase Ror2 promotes osteoblastdifferentiation and enhances ex vivo bone formation. Mol Endocrinol 21:376–387.
Maitland NJ, Collins AT. 2008. Prostate cancer stem cells: A new target for therapy. J ClinOncol 26:2862–2870.
Marina N, Gerbhardt M, Teot L, Gorlick R. 2004. Biology and therapeutic advances forpediatric osteosarcoma. Oncologist 9:422–441.

E X P A N S I O N O F O S T E O S A R C O M A - C A N C E R - S T E M C E L L S 313
Mizrak D, Brittan M, Alison MR. 2008. CD133: Molecule of the moment. J Pathol 214:3–9.Molofsky AV, Pardal R, Morrison J. 2004. Diverse mechanisms regulate stem cell self-renewal.
Curr Opin Cell Biol 16:700–707.Nadri S, Soleimani M, Kiani J, Atashi A, Izadpanah R. 2008. Multipotent mesenchymal stem
cells from adult human eye conjunctiva stromal cells. Differentiation 76:223–231.Neuzil J, Stantic M, Zobalova R, Chladova J, Wang X, Prochazka L, Dong L, Andera L, Ralph SJ.
2007. Tumour-initiating cells vs. cancer ‘stem’ cells and CD133: What’s in the name?Biochem Biophys Res Commun 355:855–859.
O’Brien CA, Pollett A, Gallinger S, Dick JE. 2007. A human colon cancer cell capable ofinitiating tumour growth in immunodeficient mice. Nature 445:106–110.
Pan G, Thomson JA. 2007. Nanog and transcriptional networks in embryonic stem cellpluripotency. Cell Res 17:42–49.
Park BW, Hah YS, Kim DR, Kim JR, Byun JH. 2007. Osteogenic phenotypes and mineralizationof cultured human periosteal-derived cells. Arch Oral Biol 52:983–989.
Picci P. 2007. Osteosarcoma (osteogenic sarcoma). Orphanet J Rare Dis 23:2–6.Reger RL, Tucker AH, Wolfe MR. 2008. Differentiation and characterization of human MSCs.
Methods Mol Biol 449:93–107.Reya T, Morrison SJ, Clarke F, Weissman IL. 2001. Stem cells, cancer, and cancer stem cells.
Nature 414:105–111.Robey RW, Shukla S, Finley EM, Oldham RK, Barnett D, Ambudkarb SV, Fojo T, Bates SE.
2008. Inhibition of P-glycoprotein (ABCB1)- and multidrug resistance-associated protein 1(ABCC1)-mediated transport by the orally administered inhibitor, CBT-1W. BiochemPharmacol 75:1302–1312.
Salasznyk RM, Klees RF, Williams WA, Boskey A, Plopper GE. 2007. Focal adhesion kinasesignaling pathways regulate the osteogenic differentiation of human mesenchymal stemcells. Exp Cell Res 313:22–37.
Schaffer D. 2007. Exploring and engineering stem cells and their niches. Curr Opin Chem Biol11:355–356.
Schreiber E, Matthias P, Muller M, Schaffner W. 1989. Rapid detection of DNA bindingproteins with mini-extracts prepared from a small number of cells. Nucleic Acid Res17:6419–6424.
JOURNAL OF CELLULAR PHYSIOLOGY
Semenza GL. 2007. Evaluation of HIF-1 inhibitors as anticancer agents. Drug Discov Today12:853–859.
Shiratsuchi H, Nakashima T, Hirakawa N, Toh S, Nakagawa T, Saito T, Tsuneyoshi M,Komune S. 2007. beta-Catenin nuclear accumulation in head and neck mucoepidermoidcarcinoma: Its role in cyclin D1 overexpression and tumor progression. Head Neck29:577–584.
Shmelkov SV, St. Clair R, Lyden D, Rafii S. 2005. Molecules in focus: AC133/CD133/Prominin-1. Int J Biochem Cell Biol 37:715–719.
Simon MC, Keith B. 2008. The role of oxygen availability in embryonic development and stemcell function. Nat Rev Mol Cell Biol 9:285–296.
Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, CusimanoMD, Dirks PB. 2004. Identification of human brain tumour initiating cells. Nature 432:396–401.
Spangler JG. 2008. Bone biology and physiology: Implications for novel osteoblasticosteosarcoma treatments? Med Hypotheses 70:281–286.
Stark GR, Taylor WR. 2006. Control of the G2/M transition. Mol Biotechnol 32:227–248.
Tang N, Song WX, Luo J, Haydon RC, He TC. 2008. Osteosarcoma development and stemcell differentiation. Clin Orthop Relat Res 466:2114–2130.
Wang JC, Dick JE. 2005. Cancer stem cells: Lessons from leukemia. Trends Cell Biol 15:494–501.
Yanagisawa M, Suzuki N, Mitsui N, Koyama Y, Otsuka K, Shimizu N. 2007. Effects ofcompressive force on the differentiation of pluripotent mesenchymal cells. Life Sci 81:405–412.
Yang YM, Chang JW. 2008. Current status and issues in cancer stem cell study. Cancer Invest26:741–755.
Ye F, Zhou C, Cheng Q, Shen J, Chen H. 2008. Stem-cell-abundant proteins Nanog,Nucleostemin and Musashi1 are highly expressed in malignant cervical epithelial cells. BMCCancer 8:108–113.
Zou GM. 2008. Cancer initiating cells or cancer stem cells in the gastrointestinal tract andliver. J Cell Physiol 217:598–604. Review.




















