
ICOVIR-5 Shows E2F1 Addiction and Potent Antiglioma Effect In vivo
10
2007;67:8255-8263. Cancer Res Marta M. Alonso, Manel Cascallo, Candelaria Gomez-Manzano, et al. In vivo Effect ICOVIR-5 Shows E2F1 Addiction and Potent Antiglioma Updated version http://cancerres.aacrjournals.org/content/67/17/8255 Access the most recent version of this article at: Cited Articles http://cancerres.aacrjournals.org/content/67/17/8255.full.html#ref-list-1 This article cites by 37 articles, 13 of which you can access for free at: Citing articles http://cancerres.aacrjournals.org/content/67/17/8255.full.html#related-urls This article has been cited by 1 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on February 23, 2014. © 2007 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on February 23, 2014. © 2007 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of ICOVIR-5 Shows E2F1 Addiction and Potent Antiglioma Effect In vivo

2007;67:8255-8263. Cancer Res Marta M. Alonso, Manel Cascallo, Candelaria Gomez-Manzano, et al.
In vivoEffect ICOVIR-5 Shows E2F1 Addiction and Potent Antiglioma
Updated version
http://cancerres.aacrjournals.org/content/67/17/8255
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/67/17/8255.full.html#ref-list-1
This article cites by 37 articles, 13 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/67/17/8255.full.html#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

ICOVIR-5 Shows E2F1 Addiction and Potent
Antiglioma Effect In vivo
Marta M. Alonso,1Manel Cascallo,
3Candelaria Gomez-Manzano,
1Hong Jiang,
1
B. Nebiyou Bekele,2Anna Perez-Gimenez,
3Frederick F. Lang,
1Yuji Piao,
1
Ramon Alemany,3and Juan Fueyo
1
1Brain Tumor Center and 2Department of Biostatistics, University of Texas M. D. Anderson Cancer Center, Houston, Texas and3Translational Research laboratory, Institut Catala d’Oncologia Barcelona, Barcelona, Spain
Abstract
During 2007, f200,000 people in the United States will bediagnosed with brain tumors. Gliomas account for 77% ofprimary malignant brain tumors, and the prognosis hashardly changed in the past 20 years, with only 30% of patientswith malignant glioma surviving 5 years after diagnosis.Oncolytic adenoviruses are promising therapies for thetreatment of gliomas. Here, report the antiglioma activity ofthe tumor-selective ICOVIR-5 adenovirus, which encompassesan early 1A adenoviral (E1A) deletion in the retinoblastoma(Rb) protein-binding region, substitution of the E1A promoterfor E2F-responsive elements, and an RGD-4C peptide motifinserted into the adenoviral fiber to enhance adenoviraltropism. Mechanistic studies showed a dramatic addiction ofICOVIR-5 to the E2F1 oncogene in vitro and in vivo . Thisaddiction was mediated by the occupancy of the ectopicadenoviral E2F1-responsive elements by the endogenous E2F1protein resulting in high level of E1A expression in cancer cellsand potent antiglioma effect. Importantly, we showed for thefirst time the ability of oncolytic adenoviruses to enhance E2Ftranscriptional activity in vivo , and we provided directevidence of the interaction of the E2F1 protein with nativeand ectopic adenovirus promoters. Restoration of Rb functionled to the association of Rb/E2F1 repressor complexes withICOVIR-5 ectopic E2F1 promoter and subsequent down-modulation of E1A, dramatically impairing adenoviral repli-cation. In xenografted mice, intratumoral injection of ICOVIR-5 resulted in a significant improvement of the median survival(P < 0.0001), and furthermore, led to 37% of long-termsurvivors free of disease. The antitumor activity of ICOVIR-5suggests that it has the potential to be an effective agent in thetreatment of gliomas. [Cancer Res 2007;67(17):8255–63]
Introduction
Glioblastoma multiforme, the most common primary braintumor, is clinically and pathologically malignant. Current treatmentstrategies involve surgery, chemotherapy, and radiotherapy, aloneand in combination, but the prognosis for patients with thesetumors has not changed dramatically in the past 20 years (1).Targeted therapies directed against the fundamental geneticabnormalities of cancer offer the promise of more rational and
effective treatments. However, with the exception of a few agentsbeing studied in early-stage clinical trials (2), current antigliomatherapies do not target specific genetic abnormalities in cancercells. One major target is the genetic alterations affecting thep16INK4a, cyclin-dependent kinase 4, and cyclin D1 proteins thatgovern the phosphorylation of the retinoblastoma (Rb) proteinleading to E2F activation (3). Indeed, the resultant inactivation ofthis pathway and consequential excess of ‘‘free’’ E2F may well be acondition for glioblastoma development (4).Oncolytic adenoviruses are promising therapies for the treat-
ment of gliomas. However, untargeted viral replication and thepaucity of coxsackie-adenovirus receptors (CAR) on tumor cells arethe major hurdles for adenovirus-based treatment. Currently, thereare three main strategies used to improve the potency andselectivity of oncolytic adenoviruses. The first involves themodification of the interaction between adenoviral proteins andtumor suppressor genes or oncogenes (5). Another strategy is basedon the modification of the adenoviral tropism to render cancercells susceptible to adenoviral infection. The third strategy involvesthe insertion of ectopic regulatory elements to restrict theexpression of early adenoviral genes to cancer cells. On the basisof studies of the transforming regions of the early 1A adenoviral(E1A) protein (6, 7), oncolytic adenoviruses targeting the Rbpathway were developed (8, 9). Because cancer cells are charac-terized by a paucity of the CAR (10), E1A-mutant adenoviruseswere modified so that they could infect tumor cells via CAR-independent mechanisms. In the best-examined strategy, increasedinfectivity was achieved through the addition of the RGD-4C motifwithin the viral fiber knob of the adenoviral construct (11, 12).Although RGD-modified E1A-mutant adenoviruses showed en-hanced infectivity and an augmented anticancer effect (12), theexcessive levels of E1A in normal cells would risk E1A-mediatedtoxicity (13, 14). Among the strategies that could be used to controlthe level of expression of E1A in normal cells is the insertion of cellcycle–dependent promoters, such as E2F1, as the main regulatorsof E1A expression (15–17). We hypothesized that an oncolyticadenovirus harboring all three modifications will display animproved selectivity while retaining a robust and effectiveantiglioma effect. However, currently, there are no reports of thepreclinical characterization of oncolytic adenoviruses that incor-porate all three modifications.In the present study, we analyzed the antiglioma effect of a new
retargeted oncolytic adenovirus (ICOVIR-5) in which both the leveland function of E1A are controlled at the transduction, transcrip-tion, and protein/protein interaction levels. ICOVIR-5 infectioninduced the up-regulation of E2F1 transcriptional activity andoccupation of the ectopic E2F1-responsive elements by theendogenous E2F1 protein in vivo . In stark contrast, adenoviralactivation was suppressed by Rb in normal cells. Intratumoral
Note: R. Alemany and J. Fueyo contributed equally to this work.Request for reprints: Juan Fueyo, Department of Neuro-Oncology, Box 1002,
University of Texas M. D. Anderson Cancer Center, 1515 Holcombe Boulevard,Houston, TX 77030. Phone: 713-834-6221; E-mail: [email protected].
I2007 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-06-4675
www.aacrjournals.org 8255 Cancer Res 2007; 67: (17). September 1, 2007
Research Article
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

administration of ICOVIR-5 resulted in a significant increase in themedian survival of tumor-bearing animals (P < 0.001) andprolonged long-term survival.
Materials and Methods
Cell lines and culture conditions. The glioma cell lines U251 MG andU87 MG were obtained from the American Type Culture Collection. Cell
lines were maintained in DMEM/F12 (1:1, v/v) supplemented with 10% fetal
bovine serum in a humidified atmosphere containing 5% CO2 at 37jC.Normal human astrocytes (NHA) were purchased from Clonetics/BioWhit-taker and maintained according to the manufacturer’s instructions.
Adenovirus construction and infection. Construction of wild-type
(WT) adenovirus (Ad300; ref. 18), WT-RGD (19), D-24 (8), and D-24-RGD(11, 12) has previously been described. Adenoviral constructs inacti-vated by UV light (12) and mock-infected cells were used as controls.
ICOVIR-2 was constructed as previously described (20). Here, we have
used a derivative of ICOVIR-2, named ICOVIR-5, which contains aKozak sequence preceding the E1A start codon. To insert the Kozak
sequence before E1A, a Kpn I fragment from pShuttle-DM-E2F-D-24containing the E2F promoter and E1A was subcloned into pGEM-3Z
(Promega), and this plasmid was used to replace the E1A translationstart site using oligonucleotides with the Kozak sequence. The Kpn I
fragment containing the E2F-E1A modified with the Kozak sequence
was returned to pShuttle-DM-E2F-D24 to obtain pShuttle-DM-E2F-
KD24. Finally, pShuttle-DM-E2F-D24 was recombined by homologousrecombination with pVK503 that contains complete Ad5 genome with
RGD-modified fiber.
Luciferase assays. Cells (3 � 104 per well in 24-well dishes) weretransfected with 250 ng of the E2F1 reporter construct (21) using FuGENE 6
transfection reagent (Roche Diagnostics Corp.). One hour after transfection,
cells were mock infected or treated with the indicated viral constructs or
Ad-h-galactosidase (Ad-h-Gal; an adenovirus that carries the cDNA of theh-galactoside used as a specificity control). Cells were harvested 24 h afterinfection, and the reporter activity was measured using the Dual Luciferase
assay (Promega). Transfections were normalized for efficiency using pRL-
CMV (Promega) and expressed as folds of induction relative to that ofmock-treated cells (assigned an arbitrary value of 1).
Chromatin immunoprecipitation assays. The chromatin immunopre-
cipitation assay was done by use of the chromatin immunoprecipitation assay
kit (Upstate Biotechnology) by following the manufacturer’s instructions. E2F1(KH-95), pRb (BD Biosciences), or mouse immunoglobulin G (IgG) antibodies
(Santa Cruz Biotechnology) were used to immunoprecipitate the cross-linked
chromatin. The following primers were used in the PCR (33 cycles) to amplifya 272-bp fragment in the E2F1 promoter and the adjacent viral genome:
5¶-TGTCTGTCCCCACCTAGGAC-3¶ and 5¶-GCGGTTCCTATTGGCTTTAAC-3¶.E2 primers were designed to amplify a 52-bp fragment in the E2 promoter
containing two binding sites for E2F1: 5¶-TCGAACAAAAGCGCGAATTTAA-3¶and 5¶-TTAAACTCTTTCCCGCGCTTTGATCAGT-3¶.
Cell cycle analysis. The cell cycle phase distribution was analyzed by
measuring the DNA content, as previously described (22).
Infection with exogenous wild-type Rb or p21. The AdCMV-pA (emptyadenovirus), Rb, or p21 adenoviruses used in this study and their infectivity
have previously been described (22, 23). Cell viability was monitored daily
and was quantified using the trypan blue exclusion test.Cell viability assay. Cristal violet and 3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide (MTT) experiments were done to quantify cell
viability, as previously described (12).
Viral replication assays. We seeded human glioma and NHA cells at5 � 104 per well in six-well plates, which, 20 h later, were infected with the
indicated adenovirus at a multiplicity of infection (MOI) of 1. Three days
after infection, we scraped the cells into culture medium and lysed them
with three cycles of freezing and thawing. We used the tissue cultureinfection dose replication assay (TCID50 method) to determine the final
viral titration. Briefly, the cell lysates were clarified by centrifugation and the
supernatants were serially diluted in medium for infecting A549 cells in96-well plates. We analyzed the cells for cytopathic effect 10 days after
infection. Final titers were determined as plaque-forming units (pfu) usingthe validation method developed by Quantum Biotechnology.
Immunoblotting assay. Samples containing identical amounts of
protein (20 Ag) were subjected to SDS-Tris-glycine gel electrophoresis.
Membranes were incubated with the following antibodies: E2F1, E1A (SantaCruz Biotechnology), fiber (NeoMarkers, Lab Vision Corp.), and a-tubulin(Sigma). The membranes were developed according to the enhanced
chemiluminescence protocol (Amersham Bioscience).
TaqMan analysis. Quantitative PCR analysis was done on a Chromo 4sequence detection system (Bio-Rad) as described elsewhere (15). For the
detection of E2F1 and E1A mRNA transcripts, we used primers cited
elsewhere (15, 24). The cycling conditions for PCR were as follows: 10 min at
95jC for 1 cycle, 15 s at 95jC, and 1 min at 60jC for 40 cycles. To determinerelative gene expression, the comparative threshold cycle (CT) method was
used (24).
Bioluminescence imaging. Cells were transfected with 250 ng of E2F1reporter plasmid (21) using FuGENE 6 transfection reagent (Roche). Where
indicated, cells were cotreated with AdpRb. Cells were harvested 48 h after
treatment and implanted in the brains of athymic mice. We anesthetized
the mice with isoflurane 48 h later and imaged for E2F-Luc–inducedluciferase expression after the i.p. injection of D-luciferin (4 mg/g of body
weight) using the IVIS imaging system (Xenogen). Acquisition parameters
were as follows: exposure time, 5 min; binning, 4; no filter; f/stop, 1; field of
view, 10 cm.Animal studies. Nude (nu/nu) mice were obtained from the breeding
facility at the University of Texas M.D. Anderson Cancer Center. U87 MG
human glioma cells (5 � 105) were engrafted into the caudate nucleus ofathymic mice using a previously described guide-screw system (25). On days
3, 5, and 7 after the implantation of tumor cells, animals were treated with
5-AL intratumoral injections of the indicated adenovirus (all 3 � 108 pfu/
mouse). All animal studies were done in the veterinary facilities of TheUniversity of Texas M.D. Anderson Cancer Center in accordance with
institutional, state, and federal laws and ethical guidelines for experimental
animal care.
Immunohistochemical analysis. Tumor xenografts and paraffin-embedded sections of the mouse tumors were treated with either goat
anti-hexon antibody (Chemicon) or goat anti-E1A (Santa Cruz Biotechnol-
ogy; ref. 26). For immunohistochemical staining, Vectastain ABC kits (VectorLaboratories) were used according to the manufacturer’s instructions.
Statistical analysis. For the in vitro experiments, statistical analyses
were done with the two-tailed Student t test. Data are expressed as mean
F SD or 95% confidence intervals (95% CI). The in vivo cytopathic effect ofICOVIR-5 on human glioma xenografts was assessed by plotting survival
curves according to the Kaplan-Meier method. Survival in different
treatment groups was compared using the log-rank test. Pairwise
comparisons of the difference in the proportion of surviving mice weredone with the Fisher exact test.
Results
E1A expression is mediated by E2F in ICOVIR-5–infectedcells. ICOVIR-5 is a third-generation retargeted oncolytic adenovi-rus in which both the level and function of a mutant E1A arecontrolled at the transduction (integrin-mediated adenoviralinfection), transcription (E2F1 regulated), and protein/proteininteraction (unable to bind Rb) levels. The structure of a similarconstruct (ICOVIR-2) has previously been described (20). Wehypothesized that ICOVIR-5 will interact with free E2F1 in cancercells and with Rb/E2F1 repressor complexes in normal quiescentcells. To assess the transcriptional activity of E2F1 in cancer andnormal cells, we transfected U87MG cells, U251MG cells, or arrestedNHAs with an E2F-Luc reporter construct (21). E2F1-mediatedluciferase activity was 12- and 14-fold higher in glioma cells,respectively, than in arrested NHAs, in which the E2F1 activity washardly detectable (P < 0.001; data not shown). Because adenoviralinfection results in increased E2F1 activity, we next examined the
Cancer Research
Cancer Res 2007; 67: (17). September 1, 2007 8256 www.aacrjournals.org
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

responsiveness of the E2F1 promoter to adenovirus infection.Glioma cells infected with any of the three adenoviruses showed asignificant (10-fold) increase in luciferase activity in comparisonwith mock-infected cells (P < 0.001). Similar results (9.5- and 6.8-foldincreases in cells infected with WT-RGD and D-24-RGD adenovirus,respectively) were obtained in growth-arrested NHAs (Fig. 1A).However, ICOVIR-5 infection in NHAs transfected with E2F-Luc didnot induce a significant increase in the activity of the E2F promoter(1.4-fold increase in comparison with adenovirus control; P > 0.05).Twenty-four hours after infection with ICOVIR-5, chromatin
immunoprecipitation assays revealed the physical interaction ofthe E2F1 protein and the ectopic E2F1-responsive promoterencompassed in the adenoviral genome in glioma cells (Fig. 1B).In addition, immunoprecipitation with the Rb protein did notreveal the association of Rb/E2F1 complexes with ICOVIR-5,strongly suggesting that ICOVIR-5 promoters are occupied by freeE2F1 protein in cancer cells. The opposite was seen in normal cells.Thus, chromatin immunoprecipitation assays showed that the Rbprotein was recruited to the recombinant E2F-responsive elementsof ICOVIR-5 in serum-starved NHAs, suggesting the association ofRb/E2F1 repressor complexes with the ectopic E2F1-responsiveelements in the adenovirus.
Cell cycle profiles of ICOVIR-5–infected human glioma andnormal cells. Our model predicted that ICOVIR-5 replicationdepends on the interaction of ICOVIR-5 with free E2F1. Becausethe free transcriptional activity of E2F1 is observed during thetransition from the late G1 to the S phase (27, 28), we analyzed thecell cycle profile of ICOVIR-5–infected glioma and normal cells.Infection with WT-RGD, D-24-RGD, and ICOVIR-5 showed a similarproportion of cells in the S phase (57%, 43%, and 35%, respectively,in U87 MG cells; 50%, 52%, and 45%, respectively, in U251 MG cells).Beyond 24 h, the cell cycle profile was so disrupted that the S-phasepopulation could not be accurately quantified. In contrast, thepercentage of ICOVIR-5–infected NHAs in the S phase wassignificantly lower (8%; P < 0.001) than the percentage of WT-RGD–infected cells (36%) or D-24-RGD–infected cells (15%;Fig. 1C). These data suggested that ICOVIR-5 infection renderscell cycle profiles potentially favorable to viral replication in cancerbut not in normal cells. To test this hypothesis, we examinedwhether the transcriptional activity of E2F1, the levels of E2F1mRNA, and the percentage of cells in the S phase were associatedwith differential expression levels of E1A in ICOVIR-5–infectedcancer cells and arrested NHAs (Fig. 1C and D). Whereas E2F1mRNA and E1A protein were highly expressed in normal cells and
Figure 1. Characterization of the E2F1promoter activity in the context ofICOVIR-5 infection. A, E2F-mediatedluciferase activity in glioma and normal cells.U87 MG, U251 MG, and arrested NHAswere plated and, 24 h later, infected(at 50 MOIs) with the indicated oncolyticadenoviruses. Cell lysates were harvested24 h after infection and luciferase activitywas determined. UV-inactivatedadenovirus (UVi), a replication-deficientadenovirus expressing the LacZ gene(b-Gal), or a simulated adenovirus infection(Mock) was used as control. Threeindependent assays were done in triplicate.*, significant differences in U87 MG andU251 MG glioma cells (P < 0.001). #,nonsignificant differences in NHAs(P > 0.05). B, examination of the occupancyof the E2F-responsive elements ofICOVIR-5 during viral infection. Chromatinimmunoprecipitation assay was done inU87 MG, U251 MG, or arrested NHAs.After infection with ICOVIR-5 or D-24-RGD,cell lysates were collected andimmunoprecipitated with anti-E2F1 or anti-Rb antibodies. Primers were designed torecognize a unique region encompassingthe E2F1 promoter and adenoviral E1Aregion (E2Fre/E1A) as well as the viralgenome or the E2 promoter of theadenovirus (E2). C, S-phase population inadenovirus-infected cells. U87 MG, U251MG, and arrested NHAs were plated andinfected with the indicated viruses at 50MOI. Cells were collected at the indicatedhours post infection (hpi) and the DNAcontent was analyzed by flow cytometry.Representative example of the results of twoindependent experiments. D, expressionlevels of E1A and E2F1 in ICOVIR-5–infected cells. U87 MG, U251 MG, orarrested NHAs were plated and infected withthe indicated viruses at 50 MOI. Cell lysateswere collected at the indicated hours postinfection. E1A expression was analyzed byWestern blot. a-Tubulin expression is shownas the loading control. E2F1 mRNAexpression was quantified at the same timepoints by quantitative RT-PCR.
New Oncolytic Strategies for Glioma Treatment
www.aacrjournals.org 8257 Cancer Res 2007; 67: (17). September 1, 2007
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

cancer cells infected with control adenoviruses, their expressionwas hardly detectable in ICOVIR-5–infected NHAs. Therefore,normal cells seem to be resistant to ICOVIR-5 by maintaining tightcontrol of cell cycle progression, thereby preventing E2F1-mediatedactivation of E1A transcription.
ICOVIR-5 displays a robust replication phenotype andantiglioma effect in vitro. Qualitative (crystal violet) andquantitative (MTT) dose-dependent assays showed that ICOVIR-5infection resulted in a cytopathic effect on U87 MG and U251 MGglioma cells. In particular, crystal violet staining showed thatICOVIR-5 infection resulted in noticeable cell death in both celllines at an MOI of 1 (Fig. 2A). MTT assays showed that the LC50 ofICOVIR-5 in both glioma cell lines ranged between MOIs of 1 and 5.Importantly, higher doses (5–10 MOIs) were required in cells
infected with D-24 (Fig. 2B), suggesting that the fiber modificationenhances ICOVIR-5 infectivity and, therefore, the capability of thevector to replicate in and kill cancer cells in comparison withadenovirus expressing wild-type fiber.To ascertain whether the cytopathic effect was due to effec-
tive replication, we carried out TCID50 assays in U87 MG andU251 MG glioma cells. ICOVIR-5 replicated efficiently in both celllines (8.3 � 107 and 9.1 � 108 pfu/mL, respectively), producingslightly inferior viral titers to those of D-24-RGD (8.7 � 108 and8.9 � 109 pfu/mL, respectively; Fig. 2C). Importantly, ICOVIR-5not only showed levels of replication similar to those of theadenovirus control in CAR-positive cells (U251 MG) but alsodisplayed a more efficient replication phenotype than the wild-type adenovirus in glioma cells expressing low levels of CAR
Figure 2. Antiglioma effect in vitro. Aand B, examination of the cytopathic effect.Cells were plated and, 24 h later, wereinfected with oncolytic adenoviruses atdoses ranging from 0.1 to 10 MOIs. Thedose-dependent cytopathic effect wasassessed by crystal violet (A ) or MTT (B)assays. Representative crystal violet assayresult. MTT analyses were done thrice intriplicate wells. UV-inactivated adenovirusand simulated adenovirus infections wereused as negative controls. C, analysis ofthe replication phenotype. Glioma cell lineswere plated and infected with oncolyticadenoviruses at 1 MOI and cell lysateswere used to infect A549 cells 72 hpostinfection. Viral titers were determinedby the TCID50 method and expressed asplaque-forming units. Three independentassays were done for each glioma cell line.D, expression of early and late adenoviralgenes. For the immunoblot analysis of E1Aand fiber expression, cells were plated and,24 h later, infected with the differentadenoviruses. Then, cells were harvested36 h after the infection and protein andmRNA were extracted. Representativeresult from immunoblot analyses for E1Aand fiber proteins. a-Tubulin is shown asthe loading control.
Cancer Research
Cancer Res 2007; 67: (17). September 1, 2007 8258 www.aacrjournals.org
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

(U87 MG). The results of the replication assays were consistentwith the levels of expression of early and late adenoviral genes, asassessed byWestern blot analyses of E1A and fiber proteins (Fig. 2D).
Restoration of the Rb pathway abrogates ICOVIR-5 antican-cer effect. We previously showed that the restoration of Rbfunction in glioma cells substantially decreased the cytopathiceffects of the D-24 and D-24-RGD adenoviruses (8, 12). Todetermine whether ICOVIR-5–mediated cell death also dependson the cell cycle regulatory function of the Rb protein, we treatedglioma cells with replication-deficient adenoviral vectors express-ing either Rb or p21 (or Ad5CMV-pA) and infected the cells withICOVIR-5 or D-24-RGD. U251 MG and U87 MG glioma cell culturespretreated with Rb followed by ICOVIR-5 infection showed less celldeath (5 F 2.3%, and 8 F 3.4%, respectively) than did culturesinfected with Ad5CMV-pA (91.4 F 1.9% and 90.2 F 2.3%; P < 0.001;Fig. 3A). We next examined the ability of the cyclin-dependentkinase inhibitor p21, a regulator of Rb function, to reduce the effectof ICOVIR-5 on the viability of wild-type Rb cells. p21 pretreatmentalmost completely protected cells from the effect of ICOVIR-5, asreflected by 90.1 F 2.1% and 87.4 F 3.8% increases in the viabilityof U87 MG and U251 MG cells, respectively (P < 0.001; Fig. 3A).
We carried out TCID50 assays to correlate cell death with viralreplication, which showed a dramatic decrease in the replicationcapability of ICOVIR-5 in cells overexpressing Rb or p21 (Fig. 3B).Interestingly, the rescue of glioma cells from the cytopathic effectof D-24-RGD in response to the exogenous expression of Rb andp21 was significantly lower than the rescue of cells from thecytopathic effect of ICOVIR-5 (P < 0.01). Accordingly, the ICOVIR-5titers were lower than the D-24-RGD titers in these cells, suggestingthat ICOVIR-5 is more sensitive to the status of the Rb pathwaythan adenoviruses encompassing exclusively CR2 mutations in E1Abut not the E2F-mediated regulation of E1A (15).
Recruitment of Rb/E2F1 repressor complexes to the E2F-ectopic promoter of ICOVIR results in the down-modulation ofE1A expression. To ascertain whether the activation of the Rbpathway results in low levels of E1A in cells infectedwith ICOVIR-5,weevaluated the expression levels of E1A in ICOVIR-5–infected cells. Incultures pretreated with Rb, we detected very low levels of E1AmRNAtranscripts (1.5 F 1-fold and 2.1 F 1.2-fold in U87 MG and U251 MGcells, respectively) than in cultures infected with Ad5CMV-pA. Similarresults were seen in cultures pretreated with p21 (2.5 F 1-fold and3.4 F 1.2-fold in U87 MG and U251 MG cells, respectively; Fig. 3C).
Figure 3. Restoration of Rb activityrestricts ICOVIR-5 replication. A ,examination of cell viability: Cells wereplated and, 24 h later, infected withadenoviral vectors expressing pRb, p21, oran empty expression cassette (CMV ).Forty-eight hours later, cells were infectedwith ICOVIR-5 or D-24-RGD adenovirus.Cytotoxicity was evaluated 7 d later by thetrypan blue exclusion method. Columns,mean of three independent experiments,each done in triplicate; bars, SD. * and #,P < 0.001. B, analysis of the replicationphenotype. Cells were pretreated as in theviability experiments and, 48 h later, wereinfected with oncolytic adenoviruses at10 MOIs. Day 3 after infection, viral titerswere determined by the TCID50 method.Columns, mean of three independentexperiments; bars, SD. * and #, P < 0.001.C, expression of E1A. Cells weresubjected to the same pretreatment as inthe replication experiments, after whichRNA was extracted and E1A mRNA levelswere assessed by quantitative RT-PCR. D,interaction of the endogenous E2F1 proteinand the ectopic E2F-responsive elements.U87 MG and U251 MG cells were platedand, 24 h later, treated with an adenoviralvector expressing pRb, p21, or CMV at100 MOI. Forty-eight hours later, cells wereinfected with ICOVIR-5 at an MOI of 10.Then, 2 d later, cell lysates were subjectedto chromatin immunoprecipitation (ChIP )assays. Samples were immunoprecipitatedwith E2F1, pRb, or IgG antibodies andectopic E2F1 promoter occupancy wasassessed by PCR. Representative resultsfrom one of three experiments.
New Oncolytic Strategies for Glioma Treatment
www.aacrjournals.org 8259 Cancer Res 2007; 67: (17). September 1, 2007
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

We next determined whether the Rb-mediated restriction ofE1A expression was dependent on the association of newlyformed Rb/E2F repressor complexes with ICOVIR-5 promoters.For this study, U87 MG and U251 MG glioma cells were infectedwith the AdpRb or the Ad5CMV-pA adenovirus at an MOI of 100and were infected 72 h later with ICOVIR-5 at an MOI of 10.We then assessed whether ectopic Rb might associate with E2F1-responsive promoters. This showed that the E2F promotersequences encompassed in the genome of ICOVIR-5 wereprecipitated with a Rb-specific antibody, but this did not occurin the absence of Rb pretreatment (Fig. 3D). These resultssupport the model whereby Rb expression directly counteractsthe E2F1-mediated augmentation of gene transcription in thecontext of ICOVIR-5 infection.We next examined whether the higher dependence of ICOVIR-5
than that of D-24-RGD on an E2F1 free status could also result inan increase in the therapeutic index of D-24-RGD in tumor overnormal tissue. In this experiment, 3 days after serum starvation, weinfected NHAs with ICOVIR-5 at MOIs of 0.1 to 10 and thencytotoxicity was evaluated 7 days later (Fig. 4A). This showed thatICOVIR-5 elicited a 20 F 5.2% decrease in cell viability at an MOI of10, whereas D-24-RGD elicited a 50% decrease in cell viability at anMOI of only 5 (Fig. 4A).To determine the therapeutic index (i.e., viral replication in tumor
cells/viral replication in normal cells), we compared adenovirusreplication in serum-starved glioma cells and NHAs. As expected,
D-24-RGD displayed an attenuated replication phenotype in NHAs,but more importantly, ICOVIR-5 was unable to replicate in thiscell line. In contrast, the Ad300 and WT-RGD adenovirusesreplicated efficiently in both gliomas and NHAs (Figs. 4B and 2C).In addition, the expression of E1A mRNA and protein was reduced(2F 0.3-fold) in arrested NHAs treatedwith ICOVIR-5 in comparisonwith NHAs treated with Ad300 (14.4 F 3.3-fold), WT-RGD (17.5 F5-fold), D-24 (5 F 1.4-fold), or D-24-RGD (8.5 F 2.9-fold; Fig. 4C).Confirming the severely impaired replication capability ofICOVIR-5, fiber expression was absent in ICOVIR-5–infectedsamples (Fig. 4D).
ICOVIR-5 infection enhances the E2F1 activity in vivo. First,we aimed to detect the transcriptional activity of E2F1 within aglioma xenograft in vivo . For this purpose, we implanted U87 MGcells transfected with the E2F-Luc construct in the brains of nudemice and then measured E2F1 activity with the use of biolumi-nescence imaging. We detected E2F1-mediated luciferase expres-sion in vivo in tumors (1.6 � 105 light units; Fig. 5A). Importantly,the intratumoral injection of ICOVIR-5 produced strong luciferaseexpression (Fig. 5A) throughout the tumor but no signal wasdetected outside the brain. In addition, the pretreatment of U87-E2F-Luc cells with an empty adenoviral vector followed by ICOVIR-5 injection resulted in luciferase levels in the tumors that were notsignificantly different from levels in tumors treated with ICOVIR-5(data not shown). In contrast, the pretreatment of U87-E2F-Luc cells with Rb protein resulted in less luciferase expression
Figure 4. Determination of the therapeutic index of ICOVIR-5. A, examination of cell viability in NHAs infected with oncolytic adenoviruses. NHAs were plated in 96-wellplates and arrested by serum starvation for 3 d followed by infection with oncolytic adenoviruses at the indicated doses. Seven days after infection, cell viabilitywas assessed by MTT assays. Columns, mean of three independent experiments, each run in triplicate; bars, SD. B, determination of the replication phenotype innormal cells infected with oncolytic adenoviruses. Arrested NHAs were plated and infected with oncolytic viruses at a dose of 1 MOI. On day 3 after infection,cells were harvested and viral titers were determined with the TCID50 method. Combined results of three independent experiments. C, examination of E1A mRNAlevels. Arrested NHAs were plated and infected with oncolytic viruses at 50 MOI. Thirty-six hours after infection, mRNA was analyzed by quantitative RT-PCR.D, examination of E1A and fiber proteins. Arrested NHAs were plated and infected with oncolytic viruses at 50 MOI. Thirty-six hours after infection, E1A and fiberexpression was analyzed by Western blot. a-Tubulin expression is shown as the loading control. Representative immunoblot.
Cancer Research
Cancer Res 2007; 67: (17). September 1, 2007 8260 www.aacrjournals.org
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
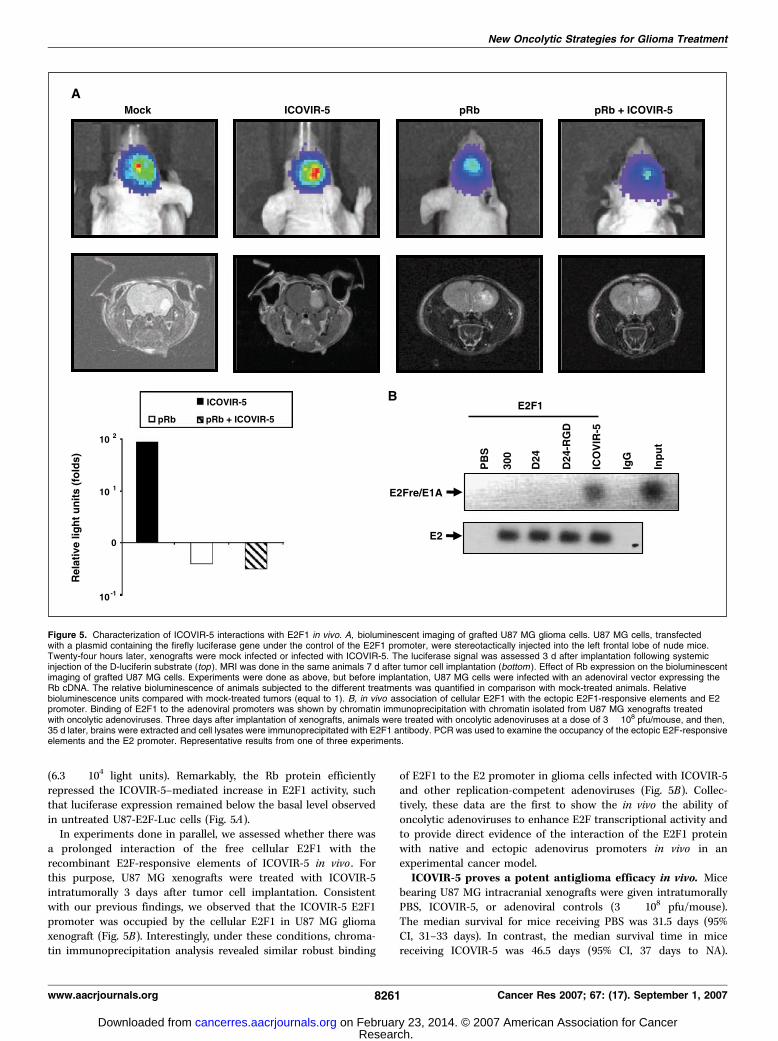
(6.3 � 104 light units). Remarkably, the Rb protein efficientlyrepressed the ICOVIR-5–mediated increase in E2F1 activity, suchthat luciferase expression remained below the basal level observedin untreated U87-E2F-Luc cells (Fig. 5A).In experiments done in parallel, we assessed whether there was
a prolonged interaction of the free cellular E2F1 with therecombinant E2F-responsive elements of ICOVIR-5 in vivo . Forthis purpose, U87 MG xenografts were treated with ICOVIR-5intratumorally 3 days after tumor cell implantation. Consistentwith our previous findings, we observed that the ICOVIR-5 E2F1promoter was occupied by the cellular E2F1 in U87 MG gliomaxenograft (Fig. 5B). Interestingly, under these conditions, chroma-tin immunoprecipitation analysis revealed similar robust binding
of E2F1 to the E2 promoter in glioma cells infected with ICOVIR-5and other replication-competent adenoviruses (Fig. 5B). Collec-tively, these data are the first to show the in vivo the ability ofoncolytic adenoviruses to enhance E2F transcriptional activity andto provide direct evidence of the interaction of the E2F1 proteinwith native and ectopic adenovirus promoters in vivo in anexperimental cancer model.
ICOVIR-5 proves a potent antiglioma efficacy in vivo. Micebearing U87 MG intracranial xenografts were given intratumorallyPBS, ICOVIR-5, or adenoviral controls (3 � 108 pfu/mouse).The median survival for mice receiving PBS was 31.5 days (95%CI, 31–33 days). In contrast, the median survival time in micereceiving ICOVIR-5 was 46.5 days (95% CI, 37 days to NA).
Figure 5. Characterization of ICOVIR-5 interactions with E2F1 in vivo. A, bioluminescent imaging of grafted U87 MG glioma cells. U87 MG cells, transfectedwith a plasmid containing the firefly luciferase gene under the control of the E2F1 promoter, were stereotactically injected into the left frontal lobe of nude mice.Twenty-four hours later, xenografts were mock infected or infected with ICOVIR-5. The luciferase signal was assessed 3 d after implantation following systemicinjection of the D-luciferin substrate (top ). MRI was done in the same animals 7 d after tumor cell implantation (bottom ). Effect of Rb expression on the bioluminescentimaging of grafted U87 MG cells. Experiments were done as above, but before implantation, U87 MG cells were infected with an adenoviral vector expressing theRb cDNA. The relative bioluminescence of animals subjected to the different treatments was quantified in comparison with mock-treated animals. Relativebioluminescence units compared with mock-treated tumors (equal to 1). B, in vivo association of cellular E2F1 with the ectopic E2F1-responsive elements and E2promoter. Binding of E2F1 to the adenoviral promoters was shown by chromatin immunoprecipitation with chromatin isolated from U87 MG xenografts treatedwith oncolytic adenoviruses. Three days after implantation of xenografts, animals were treated with oncolytic adenoviruses at a dose of 3 � 108 pfu/mouse, and then,35 d later, brains were extracted and cell lysates were immunoprecipitated with E2F1 antibody. PCR was used to examine the occupancy of the ectopic E2F-responsiveelements and the E2 promoter. Representative results from one of three experiments.
New Oncolytic Strategies for Glioma Treatment
www.aacrjournals.org 8261 Cancer Res 2007; 67: (17). September 1, 2007
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

Moreover, whereas all the mice treated with PBS died by day 37,37% of mice treated with ICOVIR-5 survived more than 90 days(Fig. 6A). Examination of the brains of asymptomatic long-termsurvivors showed complete tumor regression in all animals butone in which a small tumor remained (data not shown).
Examination of the brains of ICOVIR-5–treated mice that diedbetween 35 and 50 days after treatment showed that their deathsresulted from the mass effect of large tumors. Microscopicexamination of the brains of animals with tumors, however,revealed the presence of prominent viral inclusions, whichsuggested efficient infection and replication (Fig. 6B). Accordingly,the inclusion bodies showed immunohistochemical staining forE1A and hexon proteins. Intratumoral expression of E1A was alsoconfirmed by quantitative reverse transcription-PCR (RT-PCR;Fig. 6C). Immunohistochemical analyses of the brains of the long-term survivors using both anti-E1A and anti-hexon antibodies didnot reveal viral particles.
Discussion
Deregulation of E2F occurs in virtually all human cancers as adownstream result of mutations in several components of the Rbpathway (29). In this work, we showed that an adenovirusengineered to exploit aberrant E2F in cancer cells and tightregulation of E2F in normal cells allowed for enhanced selectivitywhile exerting a potent antiglioma effect in vitro and in vivo .It has been reported that E2F-responsive promoters are active
in glioma cells because of an excess of free E2F as a result ofloss of pRb/E2F repressor complexes (30–32). Interestingly, theE2F1 promoter encompasses E2F binding sites (21, 32). TheICOVIR model is based on the potential of E2F1 promoter forautoregulation by E2F1 protein. Importantly, our group haspreviously shown that E2F1 is overexpressed in malignant gliomasand that E2F overexpression is strongly associated with a poorprognosis in patients with high-grade gliomas (33). In light ofthese results, the correlation between E2F1 free activity andICOVIR-5–mediated oncolytic effect is significant. Our resultsproved that in the context of cancer cells, the viral E2F-responsivepromoter was occupied by free E2F (Figs. 1B and 5B), leading toincreased E1A expression and cell cycle progression, thusgenerating an auto-feedback loop that reinforced E1A activityand, subsequently, viral replication. Accordingly, arrested NHAastrocytes did not display E2F activity, and further infection withICOVIR-5 did not result in induction of S phase and led toattenuated E1A expression and an impaired replication phenotype(Figs. 1A–D and 4B). Analyses of the cytopathic effect of ICOVIR-5in normal and cancer cells showed an exceedingly broadtherapeutic index in comparison with D-24 and D-24-RGD. Inaddition, restoration of the Rb pathway in cancer cells resulted inthe formation of repressor Rb/E2F complexes with the adenoviralE2F-responsive elements leading to a drastic abrogation of celldeath (Fig. 3A). Moreover, restoration of the Rb pathway inglioma cells treated with D-24-RGD rescued viability in less than50% of infected cells. Interestingly, we were able to show the E2F1transcriptional activity and the in vivo binding of the tumoralE2F1 to the E2F-responsive elements in ICOVIR-5 infectedtumors. To our knowledge, this is the first time that the in vivoability of the virus to enhance E2F transcriptional activity hasbeen shown.The modification of the fiber HI loop of adenoviruses by the
insertion of the RGD-4C motif (34) greatly enhances the adeno-virus tropism and infectivity and thus oncolytic potency (11, 12).This seems to be true also in the context of ICOVIR-5. Whenwe compared the potency of ICOVIR-5 with the adenovirusD-24, ICOVIR-5 exerted a more robust anticancer effect thanD-24 (Figs. 2A–B and 6A). However, one could argue that this
Figure 6. Antiglioma efficacy of ICOVIR-5 in vivo. A, Kaplan-Meier survivalcurves for overall survival in ICOVIR-5–, D-24-RGD–, D-24–, and control-treatedathymic mice bearing U87 MG intracranial xenografts. Intracranial implantationof U87 MG cells (day 0) was followed by intratumoral injection (days 3, 5, and 7)of ICOVIR-5 (n = 14), D-24-RGD (D24-RGD ; n = 10), D-24 (D24 ; n = 15), orwild-type (300 ; n = 10) adenovirus or PBS (Mock ; n = 10). The P values weredetermined by log-rank test and represent a comparison of survival of ICOVIR-5–treated mice with that of mice treated with the other oncolytic agents. B, crosssection of U87 MG xenografts treated with PBS or ICOVIR-5 from mice that diedbetween 30 and 40 d after cell implantation. The tissue section was incubatedwith anti-E1A or anti-hexon antibodies. Arrowheads, viral inclusion bodies;T, tumor; N, necrosis. Magnification, �40. C, expression of E1A mRNA fromtumors of animals treated with the indicated agents as assessed by quantitativeRT-PCR. Representative findings from one of three independently doneexperiments.
Cancer Research
Cancer Res 2007; 67: (17). September 1, 2007 8262 www.aacrjournals.org
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

enhancement could come from the excess of E2F activity inglioma cells. Nevertheless, the fact that D-24-RGD is even morepotent than ICOVIR-5 seems to indicate that the E2F-responsiveelements provide more selectivity than potency. Interestingly,whereas normal astrocytes are consistently negative for avintegrins, neoplastic astrocytes in vivo and in vitro showedincreased expression of RGD-related integrins (35, 36), indicatingthat integrin retargeting could result in high levels of transductionwithin the tumor but not in the normal parenchyma. In fact, ourdata support this hypothesis because we were unable to detect anysign of viral particles in normal brain tissue. Nevertheless, theultimate test of selectivity can only be obtained in a human clinicaltrial due to the nonpermissive nature of the mouse model toadenoviral replication.To date, there is only one other construct (ONYX411) that
combines the addition of E2F promoter elements with the CR2mutation in E1A (15). However, ONYX411, in addition to remainingstrictly dependent on CAR expression for infection, differs fromICOVIR-5 in the addition of E2F responsible elements to regulatenot only E1A but also E4. These types of constructs may presentsome mechanistic and technical problems. It is normally difficult topredict the effect of every regulatory element separately. Moreworrisome is the evidence that addition of transcriptional elements
to regulate E4 could lead to genomic instability and defectiveadenovirus replication (37).The broad applicability of ICOVIR-5 based on the disruption of
the Rb pathway, a hallmark of cancer, indicates that E2F-dependentadenoviruses constitute an attractive therapeutic strategy for manytumors including gliomas. ICOVIR-5 represents an advance inoptimizing the potency and selectivity of oncolytic adenovirusesand as such deserves a further clinical testing in patients withmalignant gliomas.
Acknowledgments
Received 12/19/2006; revised 5/8/2007; accepted 6/11/2007.Grant support: NIH RO1, ‘‘Run for the Rose’’ Foundation, and National Brain
Tumor Foundation (J. Fueyo); SAF2002-04122-C03 and BIO2005-08682-C03-01 from the‘‘Ministerio de Ciencia y Tecnologia,’’ Spain (R. Alemany); and an American BrainTumor Association ‘‘Harper Rowland’’ fellowship (M.M. Alonso). R. Alemany belongs tothe Network of Cooperative Research on Cancer (C03/10), Instituto Carlos III of theMinisterio de Sanidad y Consumo, Spain.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Joy Gumin for assistance with the imaging assays; Blanca Luena, VerleneHenry, and Jennifer Edge for technical assistance with the animal experiments; andBetty P. Notzon (Department of Scientific Publications, M.D. Anderson Cancer Center)and Jan Esenwein (Department of Neuro-Oncology, M.D. Anderson Cancer Center) foreditorial assistance.
New Oncolytic Strategies for Glioma Treatment
www.aacrjournals.org 8263 Cancer Res 2007; 67: (17). September 1, 2007
References1. Stupp R, Mason WP, van den Bent MJ, et al.Radiotherapy plus concomitant and adjuvant temozo-lomide for glioblastoma. N Engl J Med 2005;352:987–96.
2. Reardon DA, Rich JN, Friedman HS, Bigner DD. Recentadvances in the treatment of malignant astrocytoma.J Clin Oncol 2006;24:1253–65.
3. Ueki K, Ono Y, Henson JW, et al. CDKN2/p16 or RBalterations occur in the majority of glioblastomas andare inversely correlated. Cancer Res 1996;56:150–3.
4. Hahn WC, Weinberg RA. Rules for making humantumor cells. N Engl J Med 2002;347:1593–603.
5. O’Shea CC. Viruses—seeking and destroying the tumorprogram. Oncogene 2005;24:7640–55.
6. Berk AJ. Functions of adenovirus E1A. Cancer Surv1986;5:367–87.
7. Whyte P, Williamson NM, Harlow E. Cellular targetsfor transformation by the adenovirus E1A proteins.Cell 1989;56:67–75.
8. Fueyo J, Gomez-Manzano C, Alemany R, et al. Amutant oncolytic adenovirus targeting the Rb pathwayproduces anti-glioma effect in vivo . Oncogene 2000;19:2–12.
9. Heise C, Hermiston T, Johnson L, et al. An adenovirusE1A mutant that demonstrates potent and selectivesystemic anti-tumoral efficacy. Nat Med 2000;6:1134–9.
10. Krasnykh V, Dmitriev I, Navarro JG, et al. Advancedgeneration adenoviral vectors possess augmented genetransfer efficiency based upon coxsackie adenovirusreceptor-independent cellular entry capacity. CancerRes 2000;60:6784–7.
11. Suzuki K, Fueyo J, Krasnykh V, et al. A conditionallyreplicative adenovirus with enhanced infectivity showsimproved oncolytic potency. Clin Cancer Res 2001;7:120–6.
12. Fueyo J, Alemany R, Gomez-Manzano C, et al.Preclinical characterization of the antiglioma activityof a tropism-enhanced adenovirus targeted to theretinoblastoma pathway. J Natl Cancer Inst 2003;95:652–60.
13. White E. Regulation of the cell cycle and apoptosisby the oncogenes of adenovirus. Oncogene 2001;20:7836–46.
14. Berk AJ. Recent lessons in gene expression, cell cyclecontrol, and cell biology from adenovirus. Oncogene2005;24:7673–85.
15. Johnson L, Shen A, Boyle L, et al. Selectivelyreplicating adenoviruses targeting deregulated E2Factivity are potent, systemic antitumor agents. CancerCell 2002;1:325–37.
16. Jakubczak JL, Ryan P, Gorziglia M, et al. An oncolyticadenovirus selective for retinoblastoma tumor suppres-sor protein pathway-defective tumors: dependence onE1A, the E2F-1 promoter, and viral replication forselectivity and efficacy. Cancer Res 2003;63:1490–9.
17. Ryan PC, Jakubczak JL, Stewart DA, et al. Antitumorefficacy and tumor-selective replication with a singleintravenous injection of OAS403, an oncolytic adeno-virus dependent on two prevalent alterations in humancancer. Cancer Gene Ther 2004;11:555–69.
18. Jones N, Shenk T. Isolation of deletion and substitu-tion mutants of adenovirus type 5. Cell 1978;13:181–8.
19. Hong SS, Gay B, Karayan L, Dabauvalle MC,Boulanger P. Cellular uptake and nuclear delivery ofrecombinant adenovirus penton base. Virology 1999;262:163–77.
20. Majem M, Cascallo M, Bayo-Puxan N, et al. Control ofE1A under an E2F-1 promoter insulated with themyotonic dystrophy locus insulator reduces the toxicityof oncolytic adenovirus Ad-D24RGD. Cancer Gene Ther2006;13:696–705.
21. Johnson DG, Ohtani K, Nevins JR. Autoregulatorycontrol of E2F1 expression in response to positive andnegative regulators of cell cycle progression. Genes Dev1994;8:1514–25.
22. Gomez-Manzano C, Fueyo J, Kyritsis AP, et al.Characterization of p53 and p21 functional interactionsin glioma cells en route to apoptosis. J Natl Cancer Inst1997;89:1036–44.
23. Fueyo J, Gomez-Manzano C, Yung WK, Kyritsis AP. Thefunctional role of tumor suppressor genes in gliomas:clues for future therapeutic strategies. Neurology 1998;51:1250–5.
24. Livak KJ, Schmittgen TD. Analysis of relative geneexpression data using real-time quantitative PCR andthe 2(�DDC(T)) method. Methods 2001;25:402–8.
25. Lal S, Lacroix M, Tofilon P, et al. An implantableguide-screw system for brain tumor studies in smallanimals. J Neurosurg 2000;92:326–33.
26. Falkeholm L, Grant CA, Magnusson A, Meoller E.Xylene-free method for histological preparation: amulticentre evaluation. Lab Invest 2001;81:1213–21.
27. Johnson DG, Schwarz JK, Cress WD, Nevins JR.Expression of transcription factor E2F1 induces quies-cent cells to enter S phase. Nature 1993;365:349–52.
28. Sherr CJ. The Pezcoller lecture: cancer cell cyclesrevisited. Cancer Res 2000;60:3689–95.
29. Sherr CJ, McCormick F. The RB and p53 pathways incancer. Cancer Cell 2002;2:103–12.
30. Adams PD, Kaelin WG, Jr. Transcriptional control byE2F. Semin Cancer Biol 1995;6:99–108.
31. Sellers WR, Rodgers JW, Kaelin WG, Jr. A potenttransrepression domain in the retinoblastoma proteininduces a cell cycle arrest when bound to E2F sites.Proc Natl Acad Sci U S A 1995;92:11544–8.
32. Parr MJ, Manome Y, Tanaka T, et al. Tumor-selectivetransgene expression in vivo mediated by an E2F-responsive adenoviral vector. Nat Med 1997;3:1145–9.
33. Alonso MM, Fueyo J, Shay JW, et al. Expression oftranscription factor E2F1 and telomerase in glioblasto-mas: mechanistic linkage and prognostic significance.J Natl Cancer Inst 2005;97:1589–600.
34. Dmitriev I, Krasnykh V, Miller CR, et al. An adenovirusvector with genetically modified fibers demonstratesexpanded tropism via utilization of a coxsackievirus andadenovirus receptor-independent cell entry mechanism.J Virol 1998;72:9706–13.
35. Paulus W, Baur I, Schuppan D, Roggendorf W.Characterization of integrin receptors in normaland neoplastic human brain. Am J Pathol 1993;143:154–63.
36. Riemenschneider MJ, Mueller W, Betensky RA,Mohapatra G, Louis DN. In situ analysis of integrinand growth factor receptor signaling pathways inhuman glioblastomas suggests overlapping relationshipswith focal adhesion kinase activation. Am J Pathol 2005;167:1379–87.
37. Working PK, Lin A, Borellini F. Meeting productdevelopment challenges in manufacturing clinical gradeoncolytic adenoviruses. Oncogene 2005;24:7792–801.
Research. on February 23, 2014. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from




















