
How specifically are action verbs represented in the neural motor system: An fMRI study
8
How specifically are action verbs represented in the neural motor system: An fMRI study Wessel O. van Dam ⁎, Shirley-Ann Rueschemeyer, Harold Bekkering Donders Institute for Brain, Cognition and Behavior, Radboud University Nijmegen, The Netherlands abstract article info Article history: Received 14 April 2010 Revised 21 June 2010 Accepted 28 June 2010 Available online 7 July 2010 Keywords: Embodiment Semantics Action Specificity Embodied accounts of language processing suggest that sensorimotor areas, generally dedicated to perception and action, are also involved in the processing and representation of word meaning. Support for such accounts comes from studies showing that language about actions selectively modulates the execution of congruent and incongruent motor responses (e.g., Glenberg & Kaschak, 2002), and from functional neuroimaging studies showing that understanding action-related language recruits sensorimotor brain areas (e.g. Hauk, Johnsrude, & Pulvermueller, 2004). In the current experiment we explored the basis of the neural motor system's involvement in representing words denoting actions. Specifically, we investigated whether the motor system's involvement is modulated by the specificity of the kinematics associated with a word. Previous research in the visual domain indicates that words denoting basic level category members lacking a specific form (e.g., bird) are less richly encoded within visual areas than words denoting subordinate level members (e.g., pelican), for which the visual form is better specified (Gauthier, Anderson, Tarr, Skudlarski, & Gore, 1997). In the present study we extend these findings to the motor domain. Modulation of the BOLD response elicited by verbs denoting a general motor program (e.g., to clean) was compared to modulation elicited by verbs denoting a more specific motor program (e.g., to wipe). Conform with our hypothesis, a region within the bilateral inferior parietal lobule, typically serving the representation of action plans and goals, was sensitive to the specificity of motor programs associated with the action verbs. These findings contribute to the growing body of research on embodied language representations by showing that the concreteness of an action-semantic feature is reflected in the neural response to action verbs. © 2010 Elsevier Inc. All rights reserved. Introduction Embodied theories of language maintain that brain areas generally dedicated to perception and action are also involved in the representation and comprehension of language stimuli. Embodied language theories differ from one another with respect to the putative role of these brain areas in processing semantic information. Some theories suggest that sensorimotor areas become an integral part of lexical-semantic representations over time as words and their referents continuously coincide (Pulvermueller, 2005). Others sug- gest that lexical-semantic meaning is derived from internal simula- tions of past experiences with words' referents (Barsalou, 2008; Simmons et al., 2008). Common to all embodied theories, however, is the idea that lexical-semantic meaning is grounded in real-world sensorimotor experiences. Embodied accounts differ fundamentally from symbolic accounts, which contend that meaning can be derived from syntactic relations between abstract language symbols (Kintsch, 2008) or statistical co-occurrences of linguistic forms (Landauer and Dumas, 1997). A middle ground between strictly embodied and disembodied accounts, which argue for grounding when necessary and statistical associations when sufficient is also represented in the literature (Mahon and Caramazza, 2008; Vigliocco et al., 2004). Converging evidence in favour of embodiment comes from behavioral, electrophysiological and neuroimaging studies. In a reaction time study, Glenberg and Kaschak (2002) showed that participants are faster to respond to sentences like “Open the drawer” if their response is compatible with the action implied in the sentence on a motor level (i.e., moving the hand towards the body). The authors argue that responses are facilitated because comprehending language about actions recruits the same neural resources required for action execution; thus, comprehending the sentence primes a congruent motor act. Rueschemeyer et al. (2010) report a similar effect on the single word level. They showed that responses to words denoting functional manipulable objects typically brought either towards or away from the body (e.g., cup or key, respectively), interacted with the required response directionality. That is, responses to words were facilitated if the required response action was congruent to the action typically performed on the word's referent (i.e., responses to cup were NeuroImage 53 (2010) 1318–1325 ⁎ Corresponding author. P.O. Box 9104, 6500 HE Nijmegen, The Netherlands. E-mail address: [email protected] (W.O. van Dam). 1053-8119/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2010.06.071 Contents lists available at ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of How specifically are action verbs represented in the neural motor system: An fMRI study

NeuroImage 53 (2010) 1318–1325
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r.com/ locate /yn img
How specifically are action verbs represented in the neural motorsystem: An fMRI study
Wessel O. van Dam ⁎, Shirley-Ann Rueschemeyer, Harold BekkeringDonders Institute for Brain, Cognition and Behavior, Radboud University Nijmegen, The Netherlands
⁎ Corresponding author. P.O. Box 9104, 6500 HE NijmE-mail address: [email protected] (W.O. van
1053-8119/$ – see front matter © 2010 Elsevier Inc. Adoi:10.1016/j.neuroimage.2010.06.071
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 April 2010Revised 21 June 2010Accepted 28 June 2010Available online 7 July 2010
Keywords:EmbodimentSemanticsActionSpecificity
Embodied accounts of language processing suggest that sensorimotor areas, generally dedicated toperception and action, are also involved in the processing and representation of word meaning. Supportfor such accounts comes from studies showing that language about actions selectively modulates theexecution of congruent and incongruent motor responses (e.g., Glenberg & Kaschak, 2002), and fromfunctional neuroimaging studies showing that understanding action-related language recruits sensorimotorbrain areas (e.g. Hauk, Johnsrude, & Pulvermueller, 2004). In the current experiment we explored the basis ofthe neural motor system's involvement in representing words denoting actions. Specifically, we investigatedwhether the motor system's involvement is modulated by the specificity of the kinematics associated with aword. Previous research in the visual domain indicates that words denoting basic level category memberslacking a specific form (e.g., bird) are less richly encoded within visual areas than words denotingsubordinate level members (e.g., pelican), for which the visual form is better specified (Gauthier, Anderson,Tarr, Skudlarski, & Gore, 1997). In the present study we extend these findings to the motor domain.Modulation of the BOLD response elicited by verbs denoting a general motor program (e.g., to clean) wascompared to modulation elicited by verbs denoting a more specific motor program (e.g., to wipe). Conformwith our hypothesis, a region within the bilateral inferior parietal lobule, typically serving the representationof action plans and goals, was sensitive to the specificity of motor programs associated with the action verbs.These findings contribute to the growing body of research on embodied language representations byshowing that the concreteness of an action-semantic feature is reflected in the neural response to actionverbs.
egen, The Netherlands.Dam).
ll rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Introduction
Embodied theories of language maintain that brain areas generallydedicated to perception and action are also involved in therepresentation and comprehension of language stimuli. Embodiedlanguage theories differ from one another with respect to the putativerole of these brain areas in processing semantic information. Sometheories suggest that sensorimotor areas become an integral part oflexical-semantic representations over time as words and theirreferents continuously coincide (Pulvermueller, 2005). Others sug-gest that lexical-semantic meaning is derived from internal simula-tions of past experiences with words' referents (Barsalou, 2008;Simmons et al., 2008). Common to all embodied theories, however, isthe idea that lexical-semantic meaning is grounded in real-worldsensorimotor experiences. Embodied accounts differ fundamentallyfrom symbolic accounts, which contend that meaning can be derivedfrom syntactic relations between abstract language symbols (Kintsch,
2008) or statistical co-occurrences of linguistic forms (Landauer andDumas, 1997). A middle ground between strictly embodied anddisembodied accounts, which argue for grounding when necessaryand statistical associations when sufficient is also represented in theliterature (Mahon and Caramazza, 2008; Vigliocco et al., 2004).
Converging evidence in favour of embodiment comes frombehavioral, electrophysiological and neuroimaging studies. In areaction time study, Glenberg and Kaschak (2002) showed thatparticipants are faster to respond to sentences like “Open the drawer”if their response is compatible with the action implied in the sentenceon amotor level (i.e., moving the hand towards the body). The authorsargue that responses are facilitated because comprehending languageabout actions recruits the same neural resources required for actionexecution; thus, comprehending the sentence primes a congruentmotor act. Rueschemeyer et al. (2010) report a similar effect on thesingle word level. They showed that responses to words denotingfunctional manipulable objects typically brought either towards oraway from the body (e.g., cup or key, respectively), interactedwith therequired response directionality. That is, responses to words werefacilitated if the required response action was congruent to the actiontypically performed on the word's referent (i.e., responses to cupwere

1319W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
faster if a movement was made towards the body rather than awayfrom the body). This indicates that very specific information abouthow an object is manipulated (i.e., movement trajectory) is activatedduring lexical retrieval. In a similar vein, Scorolli and Borghi (2007)demonstrated that compatibility effects are not only spatially but alsoeffector specific. In their study, participants were faster to respond toword pairs describing mouth actions than word pairs describing handactions when participants gave their response using their mouths.This congruency effect disappeared when participants respondedwith their feet. Thus, behavioral data to date indicate (1) that actionexecution can modulate language comprehension and (2) that thismodulation is fairly detailed (i.e., it reflects specific spatial andeffector-specific information).
Neuroimaging studies on action-language come to similar conclu-sions. For example, the comprehension of action verbs (Hauk et al.,2004; Kemmerer et al., 2008), action sentences (Tettamanti et al.,2005; Aziz-Zadeh et al., 2006) and words denoting manipulableobjects (Chao andMartin, 2000; Saccuman et al., 2006; Rueschemeyeret al., in press) reliably activate the cerebral motor system. In holdingwith what is reported in behavioral studies, patterns of brain activityare modulated in a spatially (Rueschemeyer et al., 2010) and effector-specific manner (Hauk et al., 2004).
It remains unclear whether motor activations in conjunction withaction-language reflect automatic, invariant effects or flexible,context-specific effects. In the latter case, action simulations maysimply serve to enrich lexical-semantic processing after a word hasbeen effectively accessed. This would mean that the neural motorsystem plays a role in language comprehensionwhen needed, but thatit does not support the mental representation of lexical-semanticmeaning. In support of this view, a number of studies have failed tofind motor activity for words with a putative action-semanticcomponent, when these words are presented in language contextsundermining their default motor meaning (e.g., Raposo et al., 2009;Kable et al., 2005, 2002; Rueschemeyer et al., 2007). For exampleRaposo and colleagues (2009) presented motor verbs withinidiomatic phrases (e.g., kick the bucket) and failed to find reliableeffects within the neural motor system (but see also Boulenger et al.,2009). Rueschemeyer and colleagues (2007) presented participantswith morphologically complex verbs built on a motor stem (e.g.,comprehend, which contains the motor verb stem prehend) andlikewise failed to see any effect within the neural motor system. Theseresults indicate that word forms that usually point towards a motormeaning do not activate resources in motor areas if the context inwhich the word is presented supports a non-motor interpretation.
Despite the fact that motor words in non-motor contexts fail toactivate cerebral motor areas, the finding that motor words inisolation and in contexts supporting amotor interpretation do activatethe motor system is quite robust. The question remains whetheractivation of the motor system is an all-or-nothing effect. In otherwords, do all words with a putative action-semantic componentactivate the neural motor system to the same degree? Or does thedegree of action specified by the word affect how involved the neuralmotor system is in processing the word?
Evidence from the visual domain suggests that the richness ofvisual information associated with a word modulates the level ofactivation in visual areas during word processing (Gauthier et al.,1997). Specifically, Gauthier and colleagues showed that wordsdenoting basic level members of a category lacking a specific form(e.g., bird) are less richly encoded within primary visual cortices thanmembers for which the visual form is better specified (e.g., pelican). Inthe current study, we investigate whether this finding can beextended to the motor domain. To this end we measured changes inthe hemodynamic response of participants while reading verbs thatdenoted very general (basic level) actions (e.g., to clean) andcontrasted this with the response to words denoting actionsassociated with very specific motor programs (e.g., to wipe). In
other words, while to clean refers to an action, it does not conveydetailed information about the exact motor parameters involved inperforming the act. In contrast, wipe, polish and hoover all describeactions at a level subordinate to the verb clean and are all highlyassociated with a very specific motoric act. We hypothesized that thisdifference in the richness of the action-semantic features of wordswould be reflected in a greater involvement of neural motor areas forsubordinate (e.g., wipe) vs. basic level (e.g., clean) verbs.
Experimental methods
Participants
Sixteen students of the Radboud University participated in thestudy, all of which were right-handed between 18 and 38 years of age(M=24, SD=4.63). All participants had normal or corrected-to-normal vision and no history of neurological disorders. Beforehand, allparticipants were informed about the experimental procedures, weregiven practice trials and signed informed consent. Afterward, allstudents were awarded 15 euros for participating. The data from onesubject were excluded from analysis because of an excessive amountof errors (accuracy level of less than 60% in one of the conditions) andthe data from another because of technical problems during therecording. The experiment was conducted in accordance with thenational legislation for the human protection and the HelsinkiDeclaration of 1975, revised in 2004.
Stimuli
A total of 108 Dutch words were used as experimental stimuli(stimuli with English translations can be seen in Table 1). Eighty-onewere Dutch verbs denoting either actions that you performmostly withyour arms/hands or verbs denoting abstract events and comprised thecritical experimental stimuli. The remaining 27 stimuli were Dutchverbs denoting actions that you performwith yourmouth and served ascatch trials (see procedures below). Critical stimuli belonged to one ofthree experimental conditions: (1) basic level action verb condition, i.e.,verbs denoting actions that are unspecific with regard to actionparameters (e.g., to greet, to clean), (2) subordinate level action verbcondition, i.e., verbs denoting actions that are highly specificwith regardto action parameters (e.g., to wave, to wipe), and (3) abstract verbcondition (e.g., to appreciate, to judge).
To test that stimuli were truly matched with regards topsycholinguistic variables, a questionnaire was administered to 20native Dutch speakers who did not participate in the fMRI experiment(see Table 2 for results). In this questionnaire, participants were askedto rate words on a 5-point scale with respect to (1) their familiaritywith the word (1=unfamiliar, 5=very familiar), (2) the imageabilityof the verb (1=very difficult to imagine the referent action, 5=veryeasy to imagine the referent action), (3) whether they thought theverb denoted an action performedwith the body (1=yes, 2=no) and(4) whether the verb specified a single action or multitude of actions(1=a single action, 2=more than one action).
The results of the questionnaire showed that verbs were matchedacross conditions with respect to familiarity (Basic: M=4.38,Subordinate: M=4.26; Abstract: M=4.35), F(2,18)=2,25, pN .1. Inorder to obtain an objective measure of word frequency, the meanlogarithmic lemma frequency per million for each condition wasdetermined using the CELEX lexical database (Baayen et al., 1993).This value was 1.21 for the Basic level action verbs, 0.91 for theSubordinate level action verbs and 1.28 for the Abstract verbs. Arepeated measures ANOVA indicated that there were no reliabledifferences in the mean logarithmic lemma frequency per millionbetween any of the experimental conditions, F(2,25)=1,38, pN .2.Words were also matched across conditions for length (Basic=8.0letters, Subordinate=7.3; Abstract=8.2) F(2,25)=1,61, pN .2. Thus,
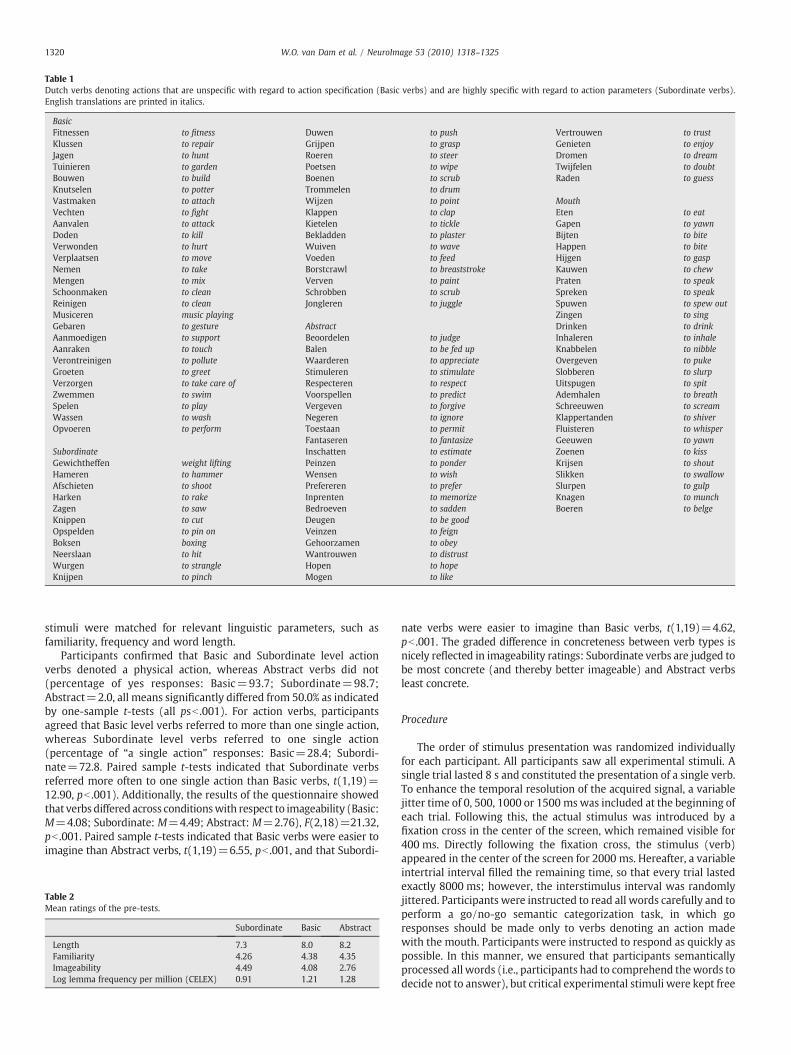
Table 1Dutch verbs denoting actions that are unspecific with regard to action specification (Basic verbs) and are highly specific with regard to action parameters (Subordinate verbs).English translations are printed in italics.
BasicFitnessen to fitness Duwen to push Vertrouwen to trustKlussen to repair Grijpen to grasp Genieten to enjoyJagen to hunt Roeren to steer Dromen to dreamTuinieren to garden Poetsen to wipe Twijfelen to doubtBouwen to build Boenen to scrub Raden to guessKnutselen to potter Trommelen to drumVastmaken to attach Wijzen to point MouthVechten to fight Klappen to clap Eten to eatAanvalen to attack Kietelen to tickle Gapen to yawnDoden to kill Bekladden to plaster Bijten to biteVerwonden to hurt Wuiven to wave Happen to biteVerplaatsen to move Voeden to feed Hijgen to gaspNemen to take Borstcrawl to breaststroke Kauwen to chewMengen to mix Verven to paint Praten to speakSchoonmaken to clean Schrobben to scrub Spreken to speakReinigen to clean Jongleren to juggle Spuwen to spew outMusiceren music playing Zingen to singGebaren to gesture Abstract Drinken to drinkAanmoedigen to support Beoordelen to judge Inhaleren to inhaleAanraken to touch Balen to be fed up Knabbelen to nibbleVerontreinigen to pollute Waarderen to appreciate Overgeven to pukeGroeten to greet Stimuleren to stimulate Slobberen to slurpVerzorgen to take care of Respecteren to respect Uitspugen to spitZwemmen to swim Voorspellen to predict Ademhalen to breathSpelen to play Vergeven to forgive Schreeuwen to screamWassen to wash Negeren to ignore Klappertanden to shiverOpvoeren to perform Toestaan to permit Fluisteren to whisper
Fantaseren to fantasize Geeuwen to yawnSubordinate Inschatten to estimate Zoenen to kissGewichtheffen weight lifting Peinzen to ponder Krijsen to shoutHameren to hammer Wensen to wish Slikken to swallowAfschieten to shoot Prefereren to prefer Slurpen to gulpHarken to rake Inprenten to memorize Knagen to munchZagen to saw Bedroeven to sadden Boeren to belgeKnippen to cut Deugen to be goodOpspelden to pin on Veinzen to feignBoksen boxing Gehoorzamen to obeyNeerslaan to hit Wantrouwen to distrustWurgen to strangle Hopen to hopeKnijpen to pinch Mogen to like
1320 W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
stimuli were matched for relevant linguistic parameters, such asfamiliarity, frequency and word length.
Participants confirmed that Basic and Subordinate level actionverbs denoted a physical action, whereas Abstract verbs did not(percentage of yes responses: Basic=93.7; Subordinate=98.7;Abstract=2.0, all means significantly differed from 50.0% as indicatedby one-sample t-tests (all psb .001). For action verbs, participantsagreed that Basic level verbs referred to more than one single action,whereas Subordinate level verbs referred to one single action(percentage of “a single action” responses: Basic=28.4; Subordi-nate=72.8. Paired sample t-tests indicated that Subordinate verbsreferred more often to one single action than Basic verbs, t(1,19)=12.90, pb .001). Additionally, the results of the questionnaire showedthat verbs differed across conditionswith respect to imageability (Basic:M=4.08; Subordinate: M=4.49; Abstract: M=2.76), F(2,18)=21.32,pb .001. Paired sample t-tests indicated that Basic verbs were easier toimagine than Abstract verbs, t(1,19)=6.55, pb .001, and that Subordi-
Table 2Mean ratings of the pre-tests.
Subordinate Basic Abstract
Length 7.3 8.0 8.2Familiarity 4.26 4.38 4.35Imageability 4.49 4.08 2.76Log lemma frequency per million (CELEX) 0.91 1.21 1.28
nate verbs were easier to imagine than Basic verbs, t(1,19)=4.62,pb .001. The graded difference in concreteness between verb types isnicely reflected in imageability ratings: Subordinate verbs are judged tobe most concrete (and thereby better imageable) and Abstract verbsleast concrete.
Procedure
The order of stimulus presentation was randomized individuallyfor each participant. All participants saw all experimental stimuli. Asingle trial lasted 8 s and constituted the presentation of a single verb.To enhance the temporal resolution of the acquired signal, a variablejitter time of 0, 500, 1000 or 1500 ms was included at the beginning ofeach trial. Following this, the actual stimulus was introduced by afixation cross in the center of the screen, which remained visible for400 ms. Directly following the fixation cross, the stimulus (verb)appeared in the center of the screen for 2000 ms. Hereafter, a variableintertrial interval filled the remaining time, so that every trial lastedexactly 8000 ms; however, the interstimulus interval was randomlyjittered. Participants were instructed to read all words carefully and toperform a go/no-go semantic categorization task, in which goresponses should be made only to verbs denoting an action madewith the mouth. Participants were instructed to respond as quickly aspossible. In this manner, we ensured that participants semanticallyprocessed all words (i.e., participants had to comprehend thewords todecide not to answer), but critical experimental stimuli were kept free
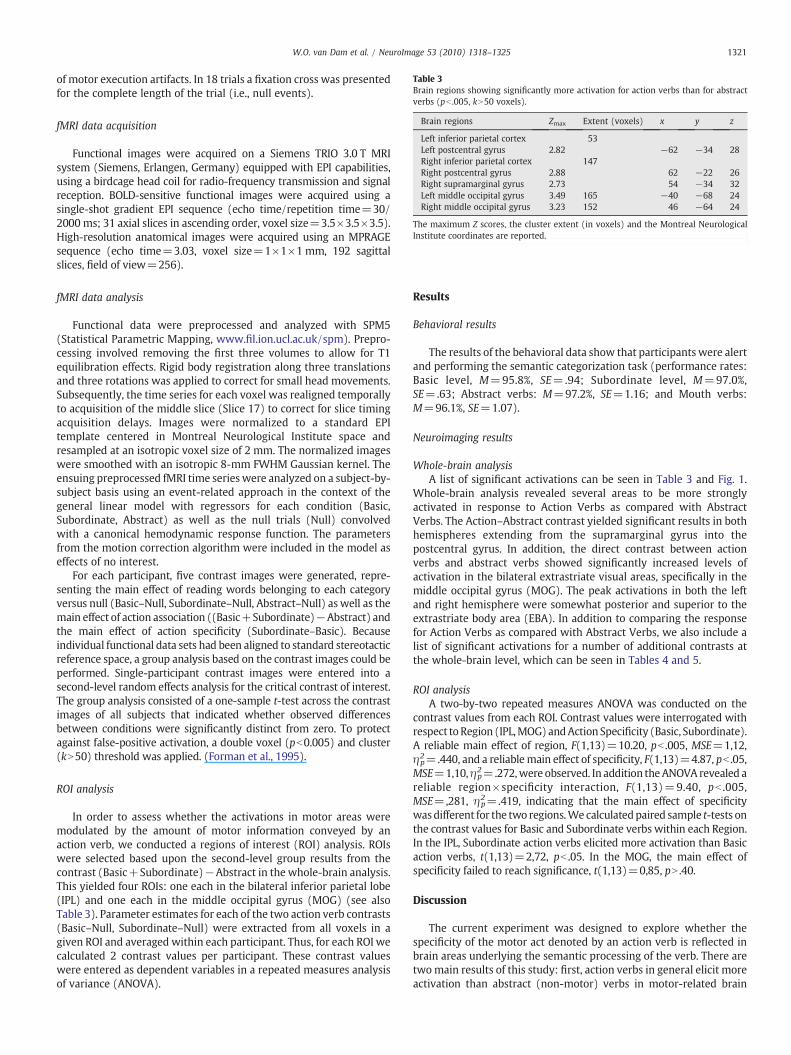
Table 3Brain regions showing significantly more activation for action verbs than for abstractverbs (pb .005, kN50 voxels).
Brain regions Zmax Extent (voxels) x y z
Left inferior parietal cortex 53Left postcentral gyrus 2.82 −62 −34 28Right inferior parietal cortex 147Right postcentral gyrus 2.88 62 −22 26Right supramarginal gyrus 2.73 54 −34 32Left middle occipital gyrus 3.49 165 −40 −68 24Right middle occipital gyrus 3.23 152 46 −64 24
The maximum Z scores, the cluster extent (in voxels) and the Montreal NeurologicalInstitute coordinates are reported.
1321W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
of motor execution artifacts. In 18 trials a fixation cross was presentedfor the complete length of the trial (i.e., null events).
fMRI data acquisition
Functional images were acquired on a Siemens TRIO 3.0 T MRIsystem (Siemens, Erlangen, Germany) equipped with EPI capabilities,using a birdcage head coil for radio-frequency transmission and signalreception. BOLD-sensitive functional images were acquired using asingle-shot gradient EPI sequence (echo time/repetition time=30/2000 ms; 31 axial slices in ascending order, voxel size=3.5×3.5×3.5).High-resolution anatomical images were acquired using an MPRAGEsequence (echo time=3.03, voxel size=1×1×1 mm, 192 sagittalslices, field of view=256).
fMRI data analysis
Functional data were preprocessed and analyzed with SPM5(Statistical Parametric Mapping, www.fil.ion.ucl.ac.uk/spm). Prepro-cessing involved removing the first three volumes to allow for T1equilibration effects. Rigid body registration along three translationsand three rotations was applied to correct for small head movements.Subsequently, the time series for each voxel was realigned temporallyto acquisition of the middle slice (Slice 17) to correct for slice timingacquisition delays. Images were normalized to a standard EPItemplate centered in Montreal Neurological Institute space andresampled at an isotropic voxel size of 2 mm. The normalized imageswere smoothed with an isotropic 8-mm FWHM Gaussian kernel. Theensuing preprocessed fMRI time series were analyzed on a subject-by-subject basis using an event-related approach in the context of thegeneral linear model with regressors for each condition (Basic,Subordinate, Abstract) as well as the null trials (Null) convolvedwith a canonical hemodynamic response function. The parametersfrom the motion correction algorithm were included in the model aseffects of no interest.
For each participant, five contrast images were generated, repre-senting the main effect of reading words belonging to each categoryversus null (Basic–Null, Subordinate–Null, Abstract–Null) as well as themain effect of action association ((Basic+Subordinate)−Abstract) andthe main effect of action specificity (Subordinate–Basic). Becauseindividual functional data sets had been aligned to standard stereotacticreference space, a group analysis based on the contrast images could beperformed. Single-participant contrast images were entered into asecond-level random effects analysis for the critical contrast of interest.The group analysis consisted of a one-sample t-test across the contrastimages of all subjects that indicated whether observed differencesbetween conditions were significantly distinct from zero. To protectagainst false-positive activation, a double voxel (pb0.005) and cluster(kN50) threshold was applied. (Forman et al., 1995).
ROI analysis
In order to assess whether the activations in motor areas weremodulated by the amount of motor information conveyed by anaction verb, we conducted a regions of interest (ROI) analysis. ROIswere selected based upon the second-level group results from thecontrast (Basic+Subordinate)−Abstract in the whole-brain analysis.This yielded four ROIs: one each in the bilateral inferior parietal lobe(IPL) and one each in the middle occipital gyrus (MOG) (see alsoTable 3). Parameter estimates for each of the two action verb contrasts(Basic–Null, Subordinate–Null) were extracted from all voxels in agiven ROI and averaged within each participant. Thus, for each ROI wecalculated 2 contrast values per participant. These contrast valueswere entered as dependent variables in a repeated measures analysisof variance (ANOVA).
Results
Behavioral results
The results of the behavioral data show that participants were alertand performing the semantic categorization task (performance rates:Basic level, M=95.8%, SE=.94; Subordinate level, M=97.0%,SE=.63; Abstract verbs: M=97.2%, SE=1.16; and Mouth verbs:M=96.1%, SE=1.07).
Neuroimaging results
Whole-brain analysisA list of significant activations can be seen in Table 3 and Fig. 1.
Whole-brain analysis revealed several areas to be more stronglyactivated in response to Action Verbs as compared with AbstractVerbs. The Action–Abstract contrast yielded significant results in bothhemispheres extending from the supramarginal gyrus into thepostcentral gyrus. In addition, the direct contrast between actionverbs and abstract verbs showed significantly increased levels ofactivation in the bilateral extrastriate visual areas, specifically in themiddle occipital gyrus (MOG). The peak activations in both the leftand right hemisphere were somewhat posterior and superior to theextrastriate body area (EBA). In addition to comparing the responsefor Action Verbs as compared with Abstract Verbs, we also include alist of significant activations for a number of additional contrasts atthe whole-brain level, which can be seen in Tables 4 and 5.
ROI analysisA two-by-two repeated measures ANOVA was conducted on the
contrast values from each ROI. Contrast values were interrogated withrespect toRegion (IPL,MOG) andAction Specificity (Basic, Subordinate).A reliable main effect of region, F(1,13)=10.20, pb .005, MSE=1,12,η2p=.440, and a reliablemain effect of specificity, F(1,13)=4.87, pb .05,
MSE=1,10, η2p=.272,were observed. In addition theANOVA revealed a
reliable region×specificity interaction, F(1,13)=9.40, pb .005,MSE=,281, η2
p=.419, indicating that the main effect of specificitywasdifferent for the two regions.We calculatedpaired sample t-tests onthe contrast values for Basic and Subordinate verbs within each Region.In the IPL, Subordinate action verbs elicited more activation than Basicaction verbs, t(1,13)=2,72, pb .05. In the MOG, the main effect ofspecificity failed to reach significance, t(1,13)=0,85, pN .40.
Discussion
The current experiment was designed to explore whether thespecificity of the motor act denoted by an action verb is reflected inbrain areas underlying the semantic processing of the verb. There aretwomain results of this study: first, action verbs in general elicit moreactivation than abstract (non-motor) verbs in motor-related brain
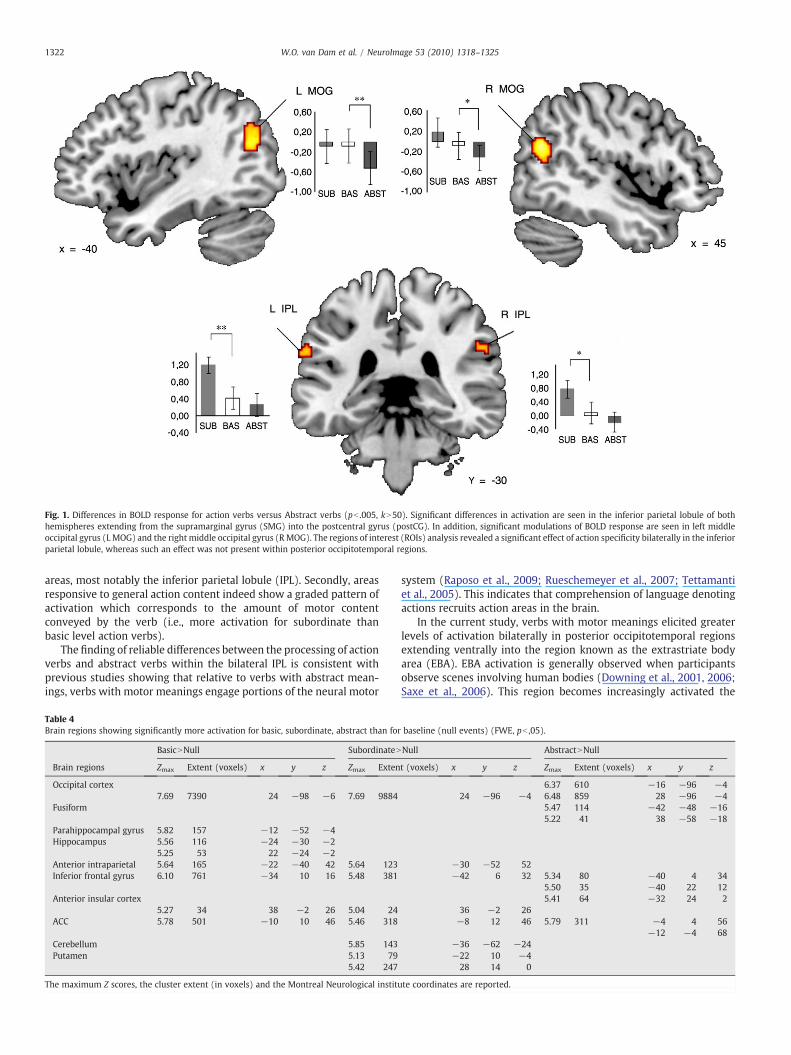
Fig. 1. Differences in BOLD response for action verbs versus Abstract verbs (pb .005, kN50). Significant differences in activation are seen in the inferior parietal lobule of bothhemispheres extending from the supramarginal gyrus (SMG) into the postcentral gyrus (postCG). In addition, significant modulations of BOLD response are seen in left middleoccipital gyrus (L MOG) and the right middle occipital gyrus (RMOG). The regions of interest (ROIs) analysis revealed a significant effect of action specificity bilaterally in the inferiorparietal lobule, whereas such an effect was not present within posterior occipitotemporal regions.
1322 W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
areas, most notably the inferior parietal lobule (IPL). Secondly, areasresponsive to general action content indeed show a graded pattern ofactivation which corresponds to the amount of motor contentconveyed by the verb (i.e., more activation for subordinate thanbasic level action verbs).
The finding of reliable differences between the processing of actionverbs and abstract verbs within the bilateral IPL is consistent withprevious studies showing that relative to verbs with abstract mean-ings, verbs with motor meanings engage portions of the neural motor
Table 4Brain regions showing significantly more activation for basic, subordinate, abstract than for
BasicNNull SubordinateN
Brain regions Zmax Extent (voxels) x y z Zmax Exten
Occipital cortex7.69 7390 24 −98 −6 7.69 9884
Fusiform
Parahippocampal gyrus 5.82 157 −12 −52 −4Hippocampus 5.56 116 −24 −30 −2
5.25 53 22 −24 −2Anterior intraparietal 5.64 165 −22 −40 42 5.64 123Inferior frontal gyrus 6.10 761 −34 10 16 5.48 381
Anterior insular cortex5.27 34 38 −2 26 5.04 24
ACC 5.78 501 −10 10 46 5.46 318
Cerebellum 5.85 143Putamen 5.13 79
5.42 247
The maximum Z scores, the cluster extent (in voxels) and the Montreal Neurological institu
system (Raposo et al., 2009; Rueschemeyer et al., 2007; Tettamantiet al., 2005). This indicates that comprehension of language denotingactions recruits action areas in the brain.
In the current study, verbs with motor meanings elicited greaterlevels of activation bilaterally in posterior occipitotemporal regionsextending ventrally into the region known as the extrastriate bodyarea (EBA). EBA activation is generally observed when participantsobserve scenes involving human bodies (Downing et al., 2001, 2006;Saxe et al., 2006). This region becomes increasingly activated the
baseline (null events) (FWE, pb ,05).
Null AbstractNNull
t (voxels) x y z Zmax Extent (voxels) x y z
6.37 610 −16 −96 −424 −96 −4 6.48 859 28 −96 −4
5.47 114 −42 −48 −165.22 41 38 −58 −18
−30 −52 52−42 6 32 5.34 80 −40 4 34
5.50 35 −40 22 125.41 64 −32 24 2
36 −2 26−8 12 46 5.79 311 −4 4 56
−12 −4 68−36 −62 −24−22 10 −428 14 0
te coordinates are reported.
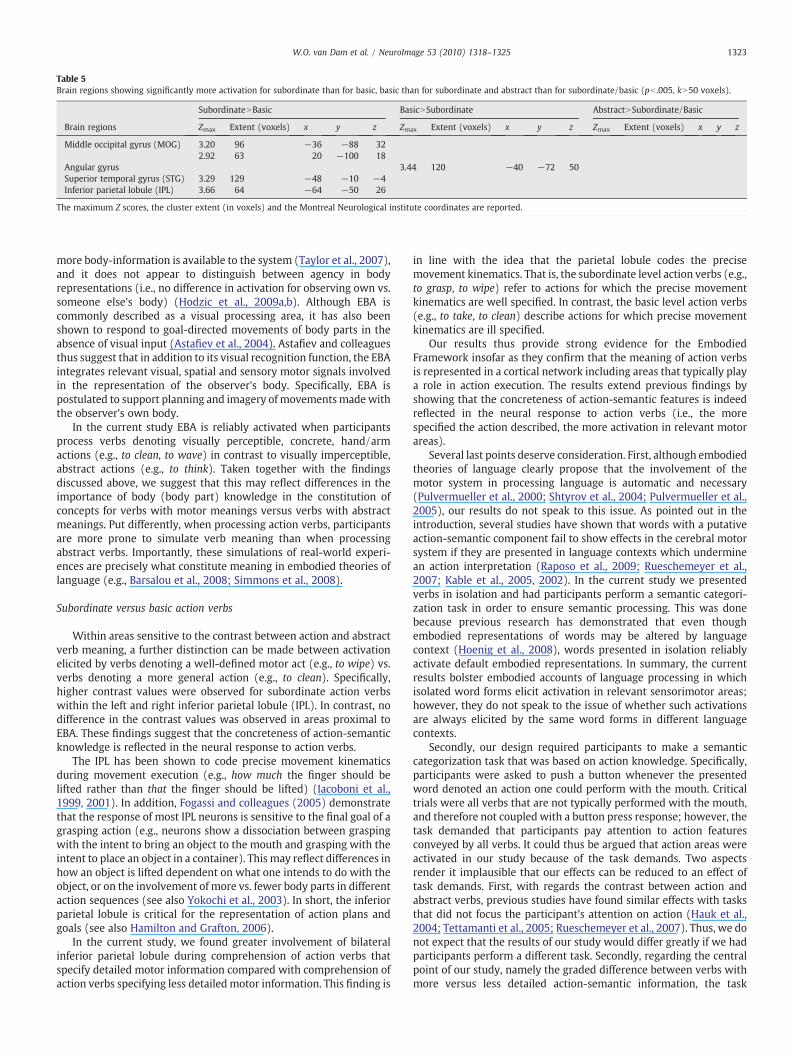
Table 5Brain regions showing significantly more activation for subordinate than for basic, basic than for subordinate and abstract than for subordinate/basic (pb .005, kN50 voxels).
SubordinateNBasic BasicNSubordinate AbstractNSubordinate/Basic
Brain regions Zmax Extent (voxels) x y z Zmax Extent (voxels) x y z Zmax Extent (voxels) x y z
Middle occipital gyrus (MOG) 3.20 96 −36 −88 322.92 63 20 −100 18
Angular gyrus 3.44 120 −40 −72 50Superior temporal gyrus (STG) 3.29 129 −48 −10 −4Inferior parietal lobule (IPL) 3.66 64 −64 −50 26
The maximum Z scores, the cluster extent (in voxels) and the Montreal Neurological institute coordinates are reported.
1323W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
more body-information is available to the system (Taylor et al., 2007),and it does not appear to distinguish between agency in bodyrepresentations (i.e., no difference in activation for observing own vs.someone else's body) (Hodzic et al., 2009a,b). Although EBA iscommonly described as a visual processing area, it has also beenshown to respond to goal-directed movements of body parts in theabsence of visual input (Astafiev et al., 2004). Astafiev and colleaguesthus suggest that in addition to its visual recognition function, the EBAintegrates relevant visual, spatial and sensory motor signals involvedin the representation of the observer's body. Specifically, EBA ispostulated to support planning and imagery of movementsmadewiththe observer's own body.
In the current study EBA is reliably activated when participantsprocess verbs denoting visually perceptible, concrete, hand/armactions (e.g., to clean, to wave) in contrast to visually imperceptible,abstract actions (e.g., to think). Taken together with the findingsdiscussed above, we suggest that this may reflect differences in theimportance of body (body part) knowledge in the constitution ofconcepts for verbs with motor meanings versus verbs with abstractmeanings. Put differently, when processing action verbs, participantsare more prone to simulate verb meaning than when processingabstract verbs. Importantly, these simulations of real-world experi-ences are precisely what constitute meaning in embodied theories oflanguage (e.g., Barsalou et al., 2008; Simmons et al., 2008).
Subordinate versus basic action verbs
Within areas sensitive to the contrast between action and abstractverb meaning, a further distinction can be made between activationelicited by verbs denoting a well-defined motor act (e.g., to wipe) vs.verbs denoting a more general action (e.g., to clean). Specifically,higher contrast values were observed for subordinate action verbswithin the left and right inferior parietal lobule (IPL). In contrast, nodifference in the contrast values was observed in areas proximal toEBA. These findings suggest that the concreteness of action-semanticknowledge is reflected in the neural response to action verbs.
The IPL has been shown to code precise movement kinematicsduring movement execution (e.g., how much the finger should belifted rather than that the finger should be lifted) (Iacoboni et al.,1999, 2001). In addition, Fogassi and colleagues (2005) demonstratethat the response of most IPL neurons is sensitive to the final goal of agrasping action (e.g., neurons show a dissociation between graspingwith the intent to bring an object to the mouth and grasping with theintent to place an object in a container). This may reflect differences inhow an object is lifted dependent on what one intends to do with theobject, or on the involvement of more vs. fewer body parts in differentaction sequences (see also Yokochi et al., 2003). In short, the inferiorparietal lobule is critical for the representation of action plans andgoals (see also Hamilton and Grafton, 2006).
In the current study, we found greater involvement of bilateralinferior parietal lobule during comprehension of action verbs thatspecify detailed motor information compared with comprehension ofaction verbs specifying less detailedmotor information. This finding is
in line with the idea that the parietal lobule codes the precisemovement kinematics. That is, the subordinate level action verbs (e.g.,to grasp, to wipe) refer to actions for which the precise movementkinematics are well specified. In contrast, the basic level action verbs(e.g., to take, to clean) describe actions for which precise movementkinematics are ill specified.
Our results thus provide strong evidence for the EmbodiedFramework insofar as they confirm that the meaning of action verbsis represented in a cortical network including areas that typically playa role in action execution. The results extend previous findings byshowing that the concreteness of action-semantic features is indeedreflected in the neural response to action verbs (i.e., the morespecified the action described, the more activation in relevant motorareas).
Several last points deserve consideration. First, although embodiedtheories of language clearly propose that the involvement of themotor system in processing language is automatic and necessary(Pulvermueller et al., 2000; Shtyrov et al., 2004; Pulvermueller et al.,2005), our results do not speak to this issue. As pointed out in theintroduction, several studies have shown that words with a putativeaction-semantic component fail to show effects in the cerebral motorsystem if they are presented in language contexts which underminean action interpretation (Raposo et al., 2009; Rueschemeyer et al.,2007; Kable et al., 2005, 2002). In the current study we presentedverbs in isolation and had participants perform a semantic categori-zation task in order to ensure semantic processing. This was donebecause previous research has demonstrated that even thoughembodied representations of words may be altered by languagecontext (Hoenig et al., 2008), words presented in isolation reliablyactivate default embodied representations. In summary, the currentresults bolster embodied accounts of language processing in whichisolated word forms elicit activation in relevant sensorimotor areas;however, they do not speak to the issue of whether such activationsare always elicited by the same word forms in different languagecontexts.
Secondly, our design required participants to make a semanticcategorization task that was based on action knowledge. Specifically,participants were asked to push a button whenever the presentedword denoted an action one could perform with the mouth. Criticaltrials were all verbs that are not typically performed with the mouth,and therefore not coupled with a button press response; however, thetask demanded that participants pay attention to action featuresconveyed by all verbs. It could thus be argued that action areas wereactivated in our study because of the task demands. Two aspectsrender it implausible that our effects can be reduced to an effect oftask demands. First, with regards the contrast between action andabstract verbs, previous studies have found similar effects with tasksthat did not focus the participant's attention on action (Hauk et al.,2004; Tettamanti et al., 2005; Rueschemeyer et al., 2007). Thus, we donot expect that the results of our study would differ greatly if we hadparticipants perform a different task. Secondly, regarding the centralpoint of our study, namely the graded difference between verbs withmore versus less detailed action-semantic information, the task

1324 W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
demands did not differ. In otherwords, in both conditions participantswere equally focussed on action in order to make a decision. Thus, forour principle contrast of interest, task requirements are the same andshould not contribute to observed differences.
While basic and subordinate level verbs in this study all refer toactions, it is the case that subordinate level verbs are more imageablethan basic level verbs. This is also supported by the results of ourquestionnaire. Thus, it could be argued the graded difference in IPLactivation reflects differences in the ability to image verbs with moreversus less detailed action information. In essence, however, this isprecisely what an embodied account of language would propose—namely that internal, intrinsic simulations underlie semantic mean-ing. Thus, we do not think this possibility is detrimental to our results.One recent study attempts to dissociate between mental imagery andaction verb understanding (Willems et al., in press). The results of thisstudy point to a dissociation between the two processes; however,language comprehension and motor imagery trials in this study weremodelled in completely different fashions. While the effects oflanguage comprehension were investigated for a period of 1.5 sstarting with the onset of the word presentation, effects of motorimagery initiated by a verb were investigated starting from themoment a participant chose to close his/her eyes after reading theverb and were then modelled for a duration of approximately 5 s.Thus, although this study presents a dissociation between languagecomprehension and motor imagery, the authors effectively look atearly language processes in their first contrast and late processes intheir second. Thus, it is entirely possible that one always images aftercomprehending a verb, but that the effects of imagery play a relativelylate role in word comprehension. This would be in line with theembodied language account proposed by Barsalou and colleagues(Barsalou et al., 2008; Simmons et al., 2008).
It should be noted that a differential neural response tosubordinate action verbs as compared with basic action verbs couldalso be accounted for in a distributed network account. That is, themeanings of basic verbs (that lack a unitary focal referent) are likely tobe more complex and distributed across different types of contentwhen compared with subordinate level verbs that have a relativelyfocal referent (Barsalou and Wiemer-Hastings, 2005). That is,differences in IPL activations might not reflect differences in therichness of action-semantic features associated with a concept butmerely be an artefact of the fact that subordinate action verbs lead toanatomically more consistent activations (due to a unitary focalreferent) as compared with basic verbs. Unfortunately we cannotdisentangle these two perspectives with the current data; however,future research looking at self-organizing clustering in fMRI data orrepetition suppression effects within single subjects could shed lighton this issue.
Conclusion
In the current study, we investigated the neural response to thecomprehension of basic action, subordinate action and abstract verbs.The results show increased activation in response to action verbs ingeneral in inferior parietal cortex, as well as posterior temporo-occipital cortex, two areas involved in processing of action and bodilyawareness, respectively. The relative contribution of the two actionverb types (i.e., basic and subordinate action verbs) differed in the twoactivated areas. Within inferior parietal cortex, a graded pattern ofactivation was observed, such that subordinate level verbs, whichconvey the most specific kinematic information, elicited the highestlevel of activation, while basic action verbs contributed less. In thetemporo-occipital areas, no significant difference was seen betweenthe action verb types. The results indicate (1) that language denotingactions elicits activation in motor areas of the brain and (2) that theamount of kinematic information conveyed by a verb is reflected inthe neural signature of language comprehension.
Acknowledgments
The authors would like to thank Paul Gaalman for technicalsupport.
Reprint requests should be sent to Wessel van Dam, DondersCentre for Brain, Cognition and Behaviour, Centre for Cognition,Montessorilaan 3, 6525 HR, Nijmegen, The Netherlands, or via e-mail:[email protected].
The study was supported by an NWO-Veni grant (No. 275-89-002)the the second author and an NWO-VICI grant (No. 453-05-001) tothe last author.
References
Astafiev, S.V., Stanley, C., Shulman, G., Corbetta, M., 2004. Extrastriate body area inhuman occipital cortex responds to the performance of motor actions. NatureNeuroscience 7, 542–548.
Aziz-Zadeh, L., Wilson, S.M., Rizzolatti, G., Iacoboni, M., 2006. Congruent embodiedrepresentations for visually presented actions and linguistic phrases describingactions. Current Biology 16, 1818–1823.
Baayen, R., Piepenbrock, R., Van Rijn, H., 1993. The CELEX Lexical Database (CD-ROM).Linguistic Data Consortium, University of Pennsylvania, Philadelphia, PA.
Barsalou, L.W., 2008. Grounded cognition. Annual Review of Psychology 59, 617–645.Barsalou, L.W., Wiemer-Hastings, K., 2005. Situating abstract concepts. In: Pecher, D.,
Zwaan, R. (Eds.), Grounding Cognition: The Role of Perception and Action inMemory,Language and Thought. Cambridge University Press, New York, pp. 129–163.
Barsalou, L.W., Santos, A., Simmons, W.K., Wilson, C.D., 2008. In: De Vega, M., Glenberg,A.M., Graesser, A.C. (Eds.), Language and simulation in conceptual processing.Symbols, embodiment, andmeaning. Oxford University Press, Oxford, pp. 245–283.
Boulenger, V., Hauk, O., Pulvermueller, F., 2009. Grasping ideas with your motor system:semantic somatotopy in idiom comprehension. Cerebral Cortex 19, 1905–1914.
Chao, L.L., Martin, A., 2000. Representation of manipulable man-made objects in thedorsal stream. Neuroimage 12, 478–484.
Downing, P.E., Jiang, Y., Shuman, M., Kanwisher, N., 2001. A cortical area selective forvisual processing of the human body. Science 293, 2470–2473.
Downing, P.E., Chan, A.W.Y., Peelen, M.V., Dodds, C.M., Kanwisher, N., 2006. Domainspecificity in visual cortex. Cerebral Cortex 16, 1453–1461.
Fogassi, L., Ferrari, P.F., Gesierich, B., Rozzi, S., Chersi, F., Rizzolatti, G., 2005. Parietallobe: from action organization to intention understanding. Science 308, 662–667.
Forman, S., Cohen, J., Fitzgerald, M., Eddy, W., Mintun, M., Noll, D., 1995. Improvedassessment of significant activation in functional magnetic resonance imaging(fMRI): Use of a cluster-size threshold. MRM 33, 637–647.
Gauthier, I., Anderson, A., Tarr, M., Skudlarski, P., Gore, J., 1997. Levels of categorizationin visual recognition studied using functional magnetic resonance imaging. CurrentBiology 7, 645–651.
Glenberg, A., Kaschak, M., 2002. Grounding language in action. Psychonomic Bulletin &Review 9, 558–565.
Hamilton, A.F.C., Grafton, S.T., 2006. Goal representation in human anterior intrapar-ietal sulcus. The Journal of Neuroscience 26 (4), 1133–1137.
Hauk, O., Johnsrude, I., Pulvermueller, F., 2004. Somatotopic representation of actionwords in human motor and premotor cortex. Neuron 41, 301–307.
Hodzic, A., Kaas, A., Muckli, L., Stirn, A., Singer, W., 2009a. Distinct cortical networks forthe detection and identification of human body. Neuroimage 45, 1264–1271.
Hodzic, A., Muckli, L., Singer, W., Stirn, A., 2009b. Cortical responses to self and others.Human Brain Mapping 30, 951–962.
Hoenig, K., Sim, E.-J., Bochev, V., Herrnberger, B., Kiefer, M., 2008. Conceptual flexibilityin the human brain: dynamic recruitment of semantic maps from visual, motor, andmotion-related areas. Journal of Cognitive Neuroscience 20 (10), 1799–1814.
Iacoboni, M., Woods, R.P., Brass, M., Bekkering, H., Mazziotta, J.C., Rizzolatti, G., 1999.Cortical mechanisms of human imitation. Science 286, 2526–2528.
Iacoboni, M., Koski, L.M., Brass,M., Bekkering, H.,Woods, R.P., Dubeau,M.C.,Mazziotta, J.C.,Rizzolatti, G., 2001. Reafferent copies of imitated actions in the right superior temporalcortex. Proceedings of the National Academy of Sciences, U.S.A. 98, 13995–13999.
Kable, J.W., Lease-Spellmeyer, J., Chatterjee, A., 2002. Neural substrates of action eventknowledge. Journal of Cognitive Neuroscience 14 (5), 795,805.
Kable, J.W., Kan, I.P., Wilson, A., Thompson-Schill, S.L., Chatterjee, A., 2005. Conceptualrepresentations of action in the lateral temporal cortex. Journal of Cognitive Neuro-science 17 (12), 1855–1870.
Kemmerer, D., Castillo, J.G., Talavage, T., Patterson, S., Wiley, C., 2008. Neuroanatomicaldistribution of five semantic components of verbs: evidence from fMRI. Brain &Language 107, 16–43.
Kintsch, W., 2008. Symbol systems and perceptual representations. In: De Vega, M.,Glenberg, A., Graesser, A. (Eds.), Symbols and Embodiment. Oxford Univ. Press,Oxford, pp. 145–164.
Landauer, T.K., Dumas, S.T., 1997. A solution to Plato's problem: the latent semanticanalysis theory of acquisition, induction, and representation of knowledge.Psychological Review 104, 211–240.
Mahon, B., Caramazza, A., 2008. A critical look at the embodied cognition hypothesis and anewproposal forgroundingconceptual content. Journalof Physiology–Paris 102, 59–70.
Pulvermueller, F., 2005. Brain mechanisms linking language and action. Nature ReviewsNeuroscience 6, 576–582.

1325W.O. van Dam et al. / NeuroImage 53 (2010) 1318–1325
Pulvermueller, F., Härle, M., Hummel, F., 2000. Neurophysiological distinction of verbcategories. Neuroreport 11, 2789–2793.
Pulvermueller, F., Shtyrov, Y., Ilmoniemi, R., 2005. Brain signatures of meaning access inaction word recognition. Journal of Cognitive Neuroscience 17 (6), 884–892.
Raposo,A.,Moss,H.E., Stamatakis, E.A., Tyler, L.K., 2009.Modulationofmotorandpremotorcorticesby actions, actionwordsandaction sentences.Neuropsychologia 47, 388–396.
Rueschemeyer, S.-A., Brass, M., Friederici, A.D., 2007. Comprehending prehending: neuralcorrelates of processing verbs with motor stems. Journal of Cognitive Neuroscience19, 855–865.
Rueschemeyer, S.-A., Pfeiffer, C., Bekkering, H., 2010. Body schematics: on the role of the bodyschema inembodied lexical-semantic representations.Neuropsychologia 48(3), 774–781.
Rueschemeyer, S.-A., Rooij, D., Lindemann, O., Willems, R., Bekkering, H., 2010. Thefunction of words: distinct neural correlates for words denoting differentlymanipulable objects. Journal of Cognitive Neuroscience 22 (8), 1844–1851.
Saccuman, M.C., Cappa, S.F., Bates, E.A., Arevalo, A., Della Rosa, P., Danna, M., Perani, D.,2006. The impact of semantic reference on word class: an fMRI study of action andobject naming. Neuroimage 32, 1865–1878.
Saxe, R., Jamal, N., Powell, L., 2006. My body or yours? The effect of visual perspective oncortical body representations. Cerebral Cortex 16, 178–182.
Scorolli, C., Borghi, A.M., 2007. Sentence comprehension and action: effector specificmodulation of the motor system. Brain Research 1130, 119–124.
Shtyrov, Y., Hauk, O., Pulvermueller, F., 2004. Distributed neuronal networks forencoding category-specific semantic information: the mismatch negativity toaction words. European Journal of. Neuroscience 19, 1083–1092.
Simmons, W.K., Hamann, S.B., Harenski, C.L., Hu, X.P., Barsalou, L.W., 2008. fMRIevidence for word association and situated simulation in conceptual processing.Journal of Physiology-Paris 102 (1–3), 106–119.
Taylor, J.C., Wiggett, A.J., Downing, P., 2007. Functional MRI analysis of body and bodypart representations in the extrastriate and fusiform body areas. Journal ofNeurophysiology 98, 1626–1633.
Tettamanti, M., Buccino, G., Saccuman, M.C., Gallese, V., Danna, M., Scifo, P., et al., 2005.Listening to action-related sentences activates fronto-parietal motor circuits.Journal of Cognitive Neuroscience 17, 273–281.
Vigliocco, G., Vinson, D.P., Lewis, W., Garrett, M., 2004. Representing the meanings ofobject and action words: the featural and unitary semantic space hypothesis.Cognitive Psychology 48, 422–488.
Willems, R.M., Toni, I., Hagoort, P., Cassasanto, D., 2010. Neural dissociations betweenaction verb understanding and motor imagery. Journal of Cognitive Neuroscience22 (10), 2387–2400.
Yokochi, H., Tanaka, M., Kumashiro, M., Iriki, A., 2003. Inferior parietal somatosensoryneurons coding face-hand coordination in Japanese macaques. Somatosensory &Motor Research 20, 115–125.




















