
How do children coordinate simultaneous upper and lower extremity tasks? The development of dual...
21
How do children coordinate simultaneous upper and lower extremity tasks? The development of dual motor task coordination Nancy Getchell a, * and Jill Whitall b a Department of Health and Exercise Sciences, University of Delaware, 141 Human Performance Laboratory, Newark, DE 19716, USA b Department of Physical Therapy, University of Maryland, Baltimore, MD, USA Received 18 February 2003; revised 31 March 2003 Abstract When performing simultaneous clapping with walking or galloping, adults adopt coupled, consistent and stable dual motor task coordination (Whitall & Getchell, 1996); do develop- mental trends in this coordination exist? In this study, we measured and compared coupling characteristics, consistency across trials and variability of phasing in 4-, 6-, 8-, and 10-year- olds (n ¼ 44) as they also performed the same dual motor task. For walk/clap, children adopted specific coupling patterns like adults by 8 years and with the same consistency by 10 years. Across age, children became less variable in clap and step movements separately and as coupled together. In the gallop/clap, children did not resemble adults in coupling pat- terns by 10 years but all measures were becoming more consistent across age. We discuss dual motor task coordination as a function of age and task complexity using a ‘‘dynamic’’ perspec- tive within a developmental context. Ó 2003 Elsevier Science (USA). All rights reserved. Keywords: Dual motor task; Coordination; Motor development; Time-sharing J. Experimental Child Psychology 85 (2003) 120–140 www.elsevier.com/locate/jecp Journal of Experimental Child Psychology * Corresponding author. Fax: 1-302-831-3693. E-mail address: [email protected] (N. Getchell). 0022-0965/03/$ - see front matter Ó 2003 Elsevier Science (USA). All rights reserved. doi:10.1016/S0022-0965(03)00059-6
Transcript of How do children coordinate simultaneous upper and lower extremity tasks? The development of dual...

How do children coordinate simultaneousupper and lower extremity tasks?The development of dual motor
task coordination
Nancy Getchella,* and Jill Whitallb
a Department of Health and Exercise Sciences, University of Delaware, 141 Human Performance
Laboratory, Newark, DE 19716, USAb Department of Physical Therapy, University of Maryland, Baltimore, MD, USA
Received 18 February 2003; revised 31 March 2003
Abstract
When performing simultaneous clapping with walking or galloping, adults adopt coupled,
consistent and stable dual motor task coordination (Whitall & Getchell, 1996); do develop-
mental trends in this coordination exist? In this study, we measured and compared coupling
characteristics, consistency across trials and variability of phasing in 4-, 6-, 8-, and 10-year-
olds (n ¼ 44) as they also performed the same dual motor task. For walk/clap, childrenadopted specific coupling patterns like adults by 8 years and with the same consistency by
10 years. Across age, children became less variable in clap and step movements separately
and as coupled together. In the gallop/clap, children did not resemble adults in coupling pat-
terns by 10 years but all measures were becoming more consistent across age. We discuss dual
motor task coordination as a function of age and task complexity using a ‘‘dynamic’’ perspec-
tive within a developmental context.
� 2003 Elsevier Science (USA). All rights reserved.
Keywords: Dual motor task; Coordination; Motor development; Time-sharing
J. Experimental Child Psychology 85 (2003) 120–140
www.elsevier.com/locate/jecp
Journal of
Experimental
Child
Psychology
* Corresponding author. Fax: 1-302-831-3693.
E-mail address: [email protected] (N. Getchell).
0022-0965/03/$ - see front matter � 2003 Elsevier Science (USA). All rights reserved.
doi:10.1016/S0022-0965(03)00059-6

Introduction
Long after children achieve the complex task of independent walking, their move-
ments are still less coordinated than those of adults. In fact, the developmental pro-
cess of acquiring motor dexterity takes many years to complete. One aspect of thismotor skillfulness is the ability to produce steady state coordinated movements.
For example, studies of fine-motor skills such as bilateral tapping suggest that
around 11 years, children tap with similar stability or consistency as adults (Wolff,
Kotwica, & Obregon, 1998). Do other forms of limb coordination show similar
age-related trends? In particular, how do multilimb coordination patterns develop?
Humans often simultaneously perform distinct actions with separate limb girdles.
Despite the ubiquity of such multilimb activities, few studies on the development
of this ability exist. To this end, this study examines the age-related changes in hu-man coordination in a multilimb timesharing task involving the concurrent actions
of clapping and locomotion.
Researchers have addressed issues of coordination in adults using a dynamic ap-
proach. Accordingly, human rhythmic coordination of limbs has been successfully
modeled as coupled non-linear limit cycle oscillators (Amazeen, Amazeen, & Turvey,
1998; Donker, Beek, Wagenaar, & Mulder, 2001; Fitzpatrick, Schmidt, & Lockman,
1996; Kay &Warren, 1998). The coupled oscillator model predicts an attraction to en-
trainment between and among the oscillating objects. That is, although objects can os-cillate individually according to biomechanical properties, when coupled, they
oscillate together in predictable patterns. Two specific and related characteristics of
entrainment are of interest in rhythmic multilimb tasks. First, we can establish if
the limbs cycle or move at the same frequency, which represents frequency locking.
Second, because limit-cycle oscillators maintain fixed relations among each other,
we can establish if the activity of one limb girdle occurs at a consistent ‘‘attractive’’
point across trials or people within the cycle of the other girdle, which represents
phase-locking.1 Note that phase-locking cannot occur without frequency lockingbut that its value can vary; together these measures determine a specific coordination
mode. Across many actions and limbs, research in adults has consistently demon-
strated that for rhythmic tasks, limbs are attracted to frequency-locked, in phase co-
ordination (limbs move at the same time) or, to a slightly lesser degree, frequency-
locked, anti-phase coordination (limbs alternate) (e.g., Kay, Kelso, Saltzman, &
Sch€ooner, 1987; Kelso & Scholz, 1985; Turvey, Rosenblum, Schmidt, & Kugler,
1986). The consistency of these measures across trials is one method of assessing the
attractiveness of a particular movement pattern at a given time period. In addition,
1 Phase-locking refers to the co-incidence of two specific points on an oscillator or limb�s cycle in afrequency-locking situation. It is possible that one limb might speed up and slow down relative to the other
limb during a cycle but always arrive at the specified point at the same time. In the case here these points
defined a priori as the clap and whatever point in the step period that phase-locking actually takes place. In
practice this second point is usually near the footstrike (data not shown). Phase locking is in contrast to
phase entrainment where the two limbs would be perfectly in phase with each other throughout the entire
cycle.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 121

the variability of the phase-locking within a trial can be interpreted as one indication
of stability2 or the strength of coupling between the limbs (Haken, Kelso, & Bunz,
1985; Von Holst, 1973).
Humans can produce both symmetric (walk, run) and asymmetric (gallop, hop)
gait patterns; asymmetrical gaits tend to be more complex, variable and to appearlater during childhood (Whitall, 1989). The walk and run are described as out of
phase, because the temporal relation of one leg to the other is 50/50% (e.g., when
one leg starts its cycle, the other leg is half way through its cycle). When galloping,
individuals must rearrange their lower limb temporal coordination to produce an
approximate 66/33 or 75/25% relation between the legs (Whitall, 1989). In the gal-
lop, one leg is oriented as a lead in front of the body while the other is oriented as
the rear leg. It has been described as a step action with the leading foot followed by
a leap-step with the other, following foot (Clark & Whitall, 1989). Clapping, incontrast, is symmetric but in-phase since both arms do the same action at the same
time, namely, moving together to contact and apart in the transverse (horizontal)
plane.
Although these three interlimb movements described occur in rudimentary form
in the rhythmic stereotypies of infants, their existence in the context of upright vol-
untary actions occurs in the following approximate but invariant order: running at
around 1.5 years, galloping between 2 and 2.5 years and continuous clapping at
around 3 years. Of course, many actions require the simultaneous production ofboth upper and lower limb rhythmical movement; little is known regarding the de-
velopmental course of dual motor tasks. Of developmental interest, then, is when
and, more importantly, how these individual skills can become combined together.
Previously, we investigated the same dual motor task coordination in adults. Ex-
tending an earlier study (Muzii, Lamm, & Gentile, 1984), we asked participants to
walk or gallop and clap at the same time in self-paced trials (Whitall & Getchell,
1996). We predicted frequency-locked in-phase coordination between the arms and
legs in these tasks. Our results partially supported this prediction. Muzii et al.(1984) as well as Whitall and Getchell (1996) identified an attractive in-phase coor-
dination mode where hand contact and heel strike temporally coincided with one
clap for each step (see Fig. 1A). This type of 1:1 entrainment between the four limbs
has been termed absolute coordination by Von Holst (1973) and is very common in
biology (Collins & Stewart, 1992).
Although the 1:1 coordination mode occurred most commonly, participants also
adopted other modes of coordination. Muzii et al. (1984) termed a second mode
‘‘harmonic,’’ a variation of absolute coordination. In harmonic coordination, the ra-
2 We recognize that low variability around a specific phase-locking point is but one indicator of a
system�s stability. Response to a perturbation, for example, is another which is actually a strongerindicator because variability by itself might consist of sources other than an inability to stabilize on one
relationship. This is illustrated nicely by the frequency-unlocking stable coordination mode where the
increased variability of between-girdle phasing is a result of frequency-uncoupling and not the instability
of one of the limbs actions. For this reason, we only consider variability of between girdle coupling in
those trials where frequency-locking is established.
122 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140

tio of frequencies of the clap and heel strike consistently coincide at harmonic multi-
ple ratios such as 1:2 or 2:1 rather than 1:1. Participants almost exclusively adopted
this form in the gallop/clap, but rarely adopted it in the walk/clap combination. This
is a less common, but occurring coordination form in biology (Collins & Stewart,
1992). Some participants also adopted a third mode of coordination, when each gir-
dle appeared to consistently cycle at its own frequency (frequency unlocked) (see Fig.1B). This form of relative coordination (Von Holst, 1973), with consistent but inde-
pendent claps and steps, appeared quite commonly in the walk/clap trials. Finally, a
few adults demonstrated occasional trials of high variability where no regularity ex-
isted between the incidence of foot strike and the occurrence of the clap (see Fig. 1C).
In this case, no frequency locking existed between the limbs, owing to large cycle var-
iability, particularly of the arms. Looking across these results, adults almost exclu-
sively adopt stable coordination patterns, with the easier combination of clapping
and walking resulting in multiple, stable forms of coupling.
Fig. 1. Diagram of classification system for coordination mode with (numbered) circles representing con-
secutive claps across stride cycles. (A) Frequency-locking with absolute coordination represents the situ-
ation where arms and legs have the same frequency and the clap occurs at the same place in each step cycle
as denoted by circles piling on top of one another. (B) Frequency-unlocking with stable coordination rep-
resents the situation where arms and legs are moving at different frequencies but in a stable relationship
with each other as denoted by circles being in an orderly progressive fashion. (C) Frequency-unlocking
with variable coordination represents the situation where arms and legs are moving at different frequencies
that are themselves variable as denoted by circles being randomly scattered.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 123

Inherent in these multilimb tasks is the notion of time-sharing between two ac-
tivities, both of which may require attention. That is, participants were asked to
perform a task with the legs (walk or gallop) while concurrently performing a task
with the arms (clap). In previous research, children simultaneously performed a
cognitive/memory and cognitive/oral task while running or galloping without dis-ruption to the coordination structure of the locomotor task (Whitall, 1991). How-
ever, if the secondary task involved a temporal structure, as with singing, one
might expect a disruption in performance of one or both tasks; in fact, partici-
pants demonstrated this with signs of entrainment between the singing and foot-
falls or finger tapping (Whitall, 1991, 1996). Within a dynamic perspective, the
development of time-sharing between two motor tasks can be interpreted using
the concept of developing entrainment between oscillating systems. The developing
system initially appears to have weak entrainment or coupling across limbs (The-len, 1986). For example, initial walking coordination is variable before becoming
adult-like in stability after 3–6 months of experience (Clark, Whitall, & Phillips,
1988). After attaining a stable walking coordination pattern, infants subsequently
acquire other locomotor skills such as running and galloping, each of which is
also initially less stable than in adults (Whitall, 1989; Whitall & Getchell, 1996).
Entrainment between the legs, then, cycles between weak and stronger coupling
as the tasks become more complex and, at least initially, require increased atten-
tion (Whitall, 1997). Thus, even though independent walking, galloping, and clap-ping have all acquired various degrees of stability by the time the child is 4 years
of age (Fitzpatrick et al., 1996; Sutherland, Olshen, Cooper, & Woo, 1980; Whi-
tall, 1989, respectively), performing these upper and lower limb tasks concurrently
will increase attentional demands. The consequence of this dual motor task time-
sharing, then, might be to demonstrate early forms of entrainment as the dual mo-
tor task coordination ability develops across age. To our knowledge, however, the
actual developmental course of time-sharing (and entrainment) in two gross motor
tasks has not been investigated.In the present study, we wanted to determine when children of ascending ages
demonstrate similar coordination modes to adults when walking or galloping at
the same time as clapping. We expected a developmental trend in the type of coor-
dination mode adopted by children; that is, children of the younger ages might show
frequency-unlocked, variable phase-locking in both walk/clap and gallop/clap condi-
tions owing to the increased attentional requirements of dual motor task coordina-
tion early in development. Based on the concept of entrainment, we expected a
developmental trend from this uncoupled state towards adult-like coupling as indi-cated by frequency locking, specific phase-locking and decreased consistency across
and variability of phase-locking within trials. In addition, we hypothesized that chil-
dren in the walk/clap condition would perform in a more advanced way (earlier fre-
quency locking and more consistency across trials) than in the gallop/clap condition
because the latter is a more complex skill that has only relatively recently been ac-
quired and therefore is relatively less stable and more attention demanding. Given
the adult data, we assumed that children would develop towards at least two possible
coordination states in clap/walk but towards one in clap/gallop.
124 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140

Method
Participants
Participants were 44 children recruited from the Baltimore-Washington metropol-itan area with 11 in each of the following age groups: 4 years (M ¼ 4:3, SD ¼ :16), 6years (M ¼ 6:4, SD ¼ :31), 8 years (M ¼ 8:1, SD ¼ :07), and 10 years (M ¼ 10:2,SD ¼ :22). All children were right-handed, lacked significant musical experienceand were free of parent-reported learning disabilities, neurological, or behavioral im-
pairments. All children except the 10-year-olds were assessed to exclude ‘‘unidenti-
fied’’ learning disabilities using the Screen (Hresko, Reid, Hammill, Ginsburg, &
Baroody, 1988). Parents signed a consent form approved by the University of Mary-
land, School of Medicine, Institutional Human Subjects Assurances Committee.Children were paid and received prizes for their participation.
Apparatus
The participants performed locomotion trials on a 10.9 m conductive rubber gait
mat with a 10-V potential from one end to the other. Participants had copper tape
electrodes placed on the heels and balls of each foot; these were connected by cables
to a Labmaster A/D board via an amplifier, and then acquired by a computer usingAsyst data acquisition software. By contacting the gait mat with these copper elec-
trodes, the participants completed a circuit that provided a measure of voltage at
each foot strike. These differential voltages provided the spatial and temporal mea-
sures of the participant�s gait.Participants used small hand-held cymbals to facilitate clapping. A lapel style re-
mote microphone (Realistic Model 32-1221A) placed on the participant captured the
sound signal of the cymbals. This signal was amplified and collected using the Asyst
software program, which synchronized the sound data with the gait mat data.
Procedure
When children entered the laboratory, we provided them with a description of the
study and then fitted them with the copper electrodes and cymbals. Through visual
inspection, all participants demonstrated the ability to produce asymmetrical gait
during single task trials of galloping. All participants performed at a self-paced speed
with the instructions to ‘‘use an even, comfortable speed until I tell you to stop.’’Comprehension and clarification of the terms ‘‘even’’ and ‘‘comfortable’’ were
sought with practice trials of the single tasks. The participants were asked to perform
two simultaneous motor tasks (either walk or gallop while clapping at the same time)
along the length of the gait mat. No temporal or coupling instructions were provided
by the investigators. All participants then demonstrated three trials under each of the
following conditions in the order given: walk, clap, walk/clap, gallop, and gallop/
clap. Data collection lasted for a maximum duration of 15 s or the amount of time
the participants moved along the 10.9 m gait mat.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 125

Data reduction
Data collected with the Asyst software were converted into ASCII files and pro-
cessed using customized software. Phasing relations between the legs confirmed that
a trial was in fact a walk (50/50%) or gallop (P64=36%). For the clap-step couplinganalysis, each step cycle was normalized to a value of 100%. Heel strike represented
the beginning and end of the cycle. Hand contact was calculated as a proportion of
the normalized step cycle. These values were calculated for each cycle across the en-
tire trial and used to provide a mean and standard deviation value for each trial. In-
dividual values per step were then represented in a frequency distribution for each
trial so that the four classifications based on frequency-locking and coordination
could be assigned for each trial. Two criteria were used to distinguish between fre-
quency locking and unlocking. A trial was considered frequency-locked if the clapalways fell at a specific time within the step (or stride) and within �20% around
the modal value (see Figs. 1A and B). For example, in Fig. 1A, claps occur at or close
to half way through the stride cycle, indicating a modal phasing value of 50%. In our
previous study, these criteria allowed unambiguous categorization of all trials into
frequency-locked or frequency-unlocked categories.
Within the frequency-locked category, trials were defined as locked-absolute for
1:1 step frequency-locking while harmonically related trials were classified as
locked-harmonic where 1 clap occurred with every other step (1:2 or stride frequen-cy-locking). Finally, the trials categorized as frequency-unlocked were classified into
the two groups: unlocked-stable and unlocked-variable as shown in Figs. 1B and C.
In unlocked-stable trials, claps occurred at a stable frequency that was different from
that of the step cycle, indicating a non-linear relation between the two. Because of
the different frequencies, a large phasing variability exists between the two girdles.
In unlocked-variable trials, the claps and/or steps had a highly variable frequency
that, again, contributed to a large phasing variability across girdles. Within this clas-
sification scheme, only the unlocked-variable trials are an indication of a completelyuncoupled (no entrainment apparent) neuromuscular system.
Based on these classifications, the trials were analyzed at the following levels of
interest. To observe global attractive coordination modes, we classified all trials
available into the four categories described above. Next, we analyzed each partici-
pant to determine individual consistency in categorization across trials. Within the
frequency-locked categories only, we looked at the variability of the clap-step fre-
quency-locked coordination modes within each trial using the standard deviation
around the mean. This measure is one method of quantifying the stability of the co-ordination between clapping and stepping. Several of the relationships were statisti-
cally compared using v2 goodness of fit tests with adult data as the populationdistribution. Finally, the clap and steps were assessed individually for stability using
the coefficient of variation of the clap or step cycles. These were compared using a
4� 2� 2 (age� coordination mode� limb girdle) repeated measures ANOVA, witha Newman–Keuls post hoc test performed on significant results. Statistical analyses
were undertaken with a ¼ :05. Adult data from Whitall and Getchell (1996) are pre-sented on graphs for comparison.
126 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140
Results
Clap/walk at preferred speed
In the walk/clap condition, each participant produced three trials for a total of 33analyzable trials per age group. The number of stride cycles per trials varied from
approximately 10.6 for the 4-year-olds (average stride period¼ 943ms), 9.8 for the6-year-olds (average stride period¼ 1020ms), 8.02 for the 8-year-olds (average strideperiod¼ 1246ms) and 8.46 for the 10-year-olds (average stride period¼ 1181ms).Using the classification system as previously described, all of the walk/clap trials
could be classified into one of the global coordination patterns.
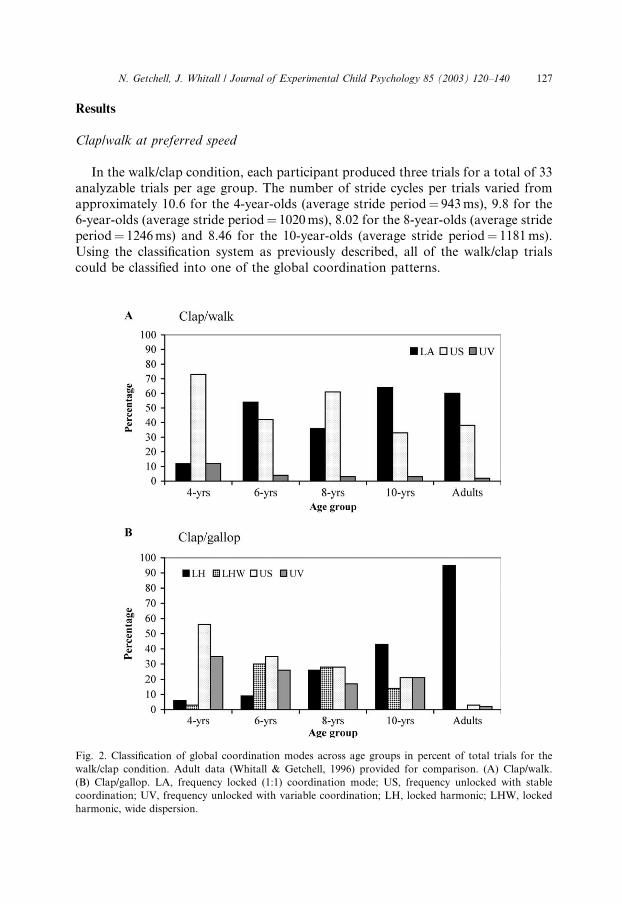
Fig. 2. Classification of global coordination modes across age groups in percent of total trials for the
walk/clap condition. Adult data (Whitall & Getchell, 1996) provided for comparison. (A) Clap/walk.
(B) Clap/gallop. LA, frequency locked (1:1) coordination mode; US, frequency unlocked with stable
coordination; UV, frequency unlocked with variable coordination; LH, locked harmonic; LHW, locked
harmonic, wide dispersion.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 127

Classification of global coordination patterns
The classification into the four coordination patterns is presented in Fig. 2A. Our
prediction that there would be a developmental trend from unlocked-variable to
locked coordination as a function of age was partially upheld. We had predicted that
the youngest groups would have a greater number of frequency unlocked, variableclassifications. In fact, the youngest groups did tend to be unlocked, but were un-
locked-stable rather than unlocked-variable. At the same time, there appeared to
be a developmental trend towards coupling the arms and legs together in a frequen-
cy-locked pattern. For the 4-year-old group, only 12% of the trials were classified as
either locked-absolute or unlocked-variable. The majority (75%) was classified as un-
locked-stable. Across the age groups, the percentage of trials classified as locked-ab-
solute increased as the percentage of trials classified as unlocked-stable and
unlocked-variable decreased.
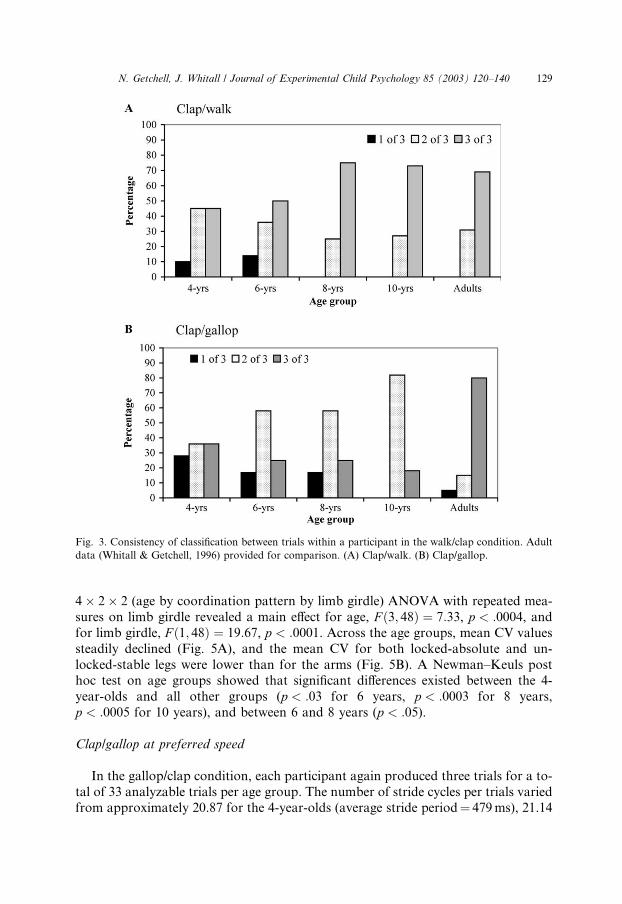
Consistency of classification across trials within a participant
Fig. 3A displays the number of trials for each participant that were classified as
the same coordination pattern in each age group. We had predicted that participants
would more consistently adopt a particular coordination mode over three trials as a
function of age group. In fact, the two youngest groups were less consistent in pat-
tern adoption. In contrast, the 8- and 10-year-old groups consistently adopted the
same coordination pattern as frequently as adults. No one in or after the 8-year-old group performed all three trials differently. Using v2 goodness of fit tests to com-pare age groups with frequency of trial classification within participants, we found a
significant difference, v2ð3;N ¼ 28Þ ¼ 17:6, p < :0005, in the frequency of all threetrials being classified the same. There was no significance when looking at age groups
and frequency of two out of three trials classified the same, v2ð3;N ¼ 15Þ ¼ 4:7,p < :20. Taken together, these results suggest that even the younger groups tendto couple limbs relatively consistently across trials, but not as consistently as adults
until 8 years old.
Variability of coordination within frequency-locked trials
This measure quantifies the strength of coupling between the arms and legs within
a trial in trials classified as locked-absolute. A lower dispersion value indicates a tigh-
ter coupling between the claps and steps across a given trial (e.g., claps consistently
occurred within 5% of the same place in the step cycle). Values indicate the percent-
age of trials that were within a particular range of phase dispersion (see Fig. 4A).
Even the youngest group has fairly stable frequency locking (100%6 15). As wehad hypothesized, across the age groups, there is a tendency to decrease variability.
Of the frequency-locked trials, 5% for the 6-year-olds, and 15% for the 8- and 10-
year-olds were dispersed less than 5%; this can be compared to 45% in the previously
reported adult group.
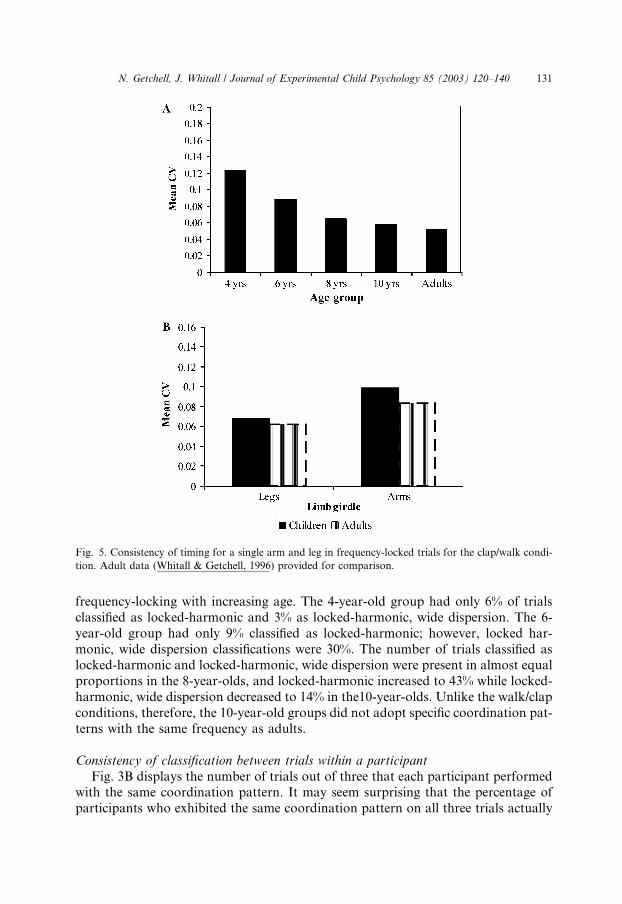
Variability of independent clap and step
The average values of the coefficients of variation (CV) for the participants with
locked-absolute and unlocked-stable classifications are presented in Fig. 5. A
128 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140
4� 2� 2 (age by coordination pattern by limb girdle) ANOVA with repeated mea-sures on limb girdle revealed a main effect for age, F ð3; 48Þ ¼ 7:33, p < :0004, andfor limb girdle, F ð1; 48Þ ¼ 19:67, p < :0001. Across the age groups, mean CV valuessteadily declined (Fig. 5A), and the mean CV for both locked-absolute and un-
locked-stable legs were lower than for the arms (Fig. 5B). A Newman–Keuls posthoc test on age groups showed that significant differences existed between the 4-
year-olds and all other groups (p < :03 for 6 years, p < :0003 for 8 years,
p < :0005 for 10 years), and between 6 and 8 years (p < :05).
Clap/gallop at preferred speed
In the gallop/clap condition, each participant again produced three trials for a to-
tal of 33 analyzable trials per age group. The number of stride cycles per trials variedfrom approximately 20.87 for the 4-year-olds (average stride period¼ 479ms), 21.14
Fig. 3. Consistency of classification between trials within a participant in the walk/clap condition. Adult
data (Whitall & Getchell, 1996) provided for comparison. (A) Clap/walk. (B) Clap/gallop.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 129

for the 6-year-olds (average stride period¼ 473ms), 16.13 for the 8-year-olds (aver-age stride period¼ 619ms) and 15.16 for the 10-year-olds (average stride pe-riod¼ 659ms). We added an additional classification of coordination pattern forthe gallop/clap. Some children had a clear modal phasing relationship (90% of claps
appeared at a specific point within the step cycle) but had a wider dispersion or
spread around that modal value (10% of the cycles were just over 20, but less than
25%) than the adults. According to Von Holst (1973), this might be considered a
form of relative coordination in the sense that the modal phasing relationship is al-
lowed to wander (phase wandering). We defined this coordination pattern as locked-
harmonic, wide dispersion.
Classification of global coordination patterns
The classification into the four coordination modes is presented in Fig. 2B. Just as
with the walk/clap condition, there is a developmental trend toward an increase in
Fig. 4. Stability of phasing in frequency-locked trials for the walk/clap condition. Adult data (Whitall &
Getchell, 1996) provided for comparison. (A) Clap/walk. (B) Clap/gallop. LH, frequency locked (1:2) co-
ordination mode; LHW, frequency locked (1:2) with wide dispersion around mean; US, frequency un-
locked with stable coordination; UV, frequency unlocked with variable coordination.
130 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140
frequency-locking with increasing age. The 4-year-old group had only 6% of trials
classified as locked-harmonic and 3% as locked-harmonic, wide dispersion. The 6-
year-old group had only 9% classified as locked-harmonic; however, locked har-
monic, wide dispersion classifications were 30%. The number of trials classified aslocked-harmonic and locked-harmonic, wide dispersion were present in almost equal
proportions in the 8-year-olds, and locked-harmonic increased to 43% while locked-
harmonic, wide dispersion decreased to 14% in the10-year-olds. Unlike the walk/clap
conditions, therefore, the 10-year-old groups did not adopt specific coordination pat-
terns with the same frequency as adults.
Consistency of classification between trials within a participant
Fig. 3B displays the number of trials out of three that each participant performedwith the same coordination pattern. It may seem surprising that the percentage of
participants who exhibited the same coordination pattern on all three trials actually
Fig. 5. Consistency of timing for a single arm and leg in frequency-locked trials for the clap/walk condi-
tion. Adult data (Whitall & Getchell, 1996) provided for comparison.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 131

declined from the 4- to the 10-year-old group. However, early age groups consisted
of many children who adopted unlocked-variable patterns across all trials whereas
more participants in the older age groups coupled their limbs in a frequency-locked
pattern for at least two trials. For the relationship between age group and frequency
of classification across trials, we found significant v2, v21ð3;N ¼ 5Þ ¼ 96:4, p < :0001;v22ð3;N ¼ 27Þ ¼ 624:1, p < :000; v23ð3;N ¼ 12Þ ¼ 152:3, p < :000. Again, in contrastto the walk/clap, children were not as consistent as adults across all trials in the type
of coordination pattern they exhibited, even by 10 years.
Variability of coordination within frequency-locked trials
As with the walk/clap, this indicates the tightness of coupling between the arms
and legs, with lower dispersion percentages indicating tighter coupling. Values indi-
cate the percentage of trials that were within a particular range of phase dispersion.There were no clear developmental trends in the tightness of frequency locking (Fig.
4B). Those few 4-year-olds who produced a frequency-locked trial displayed fairly
tight coupling, which did not persist in the older age groups.
Variability of independent clap and step
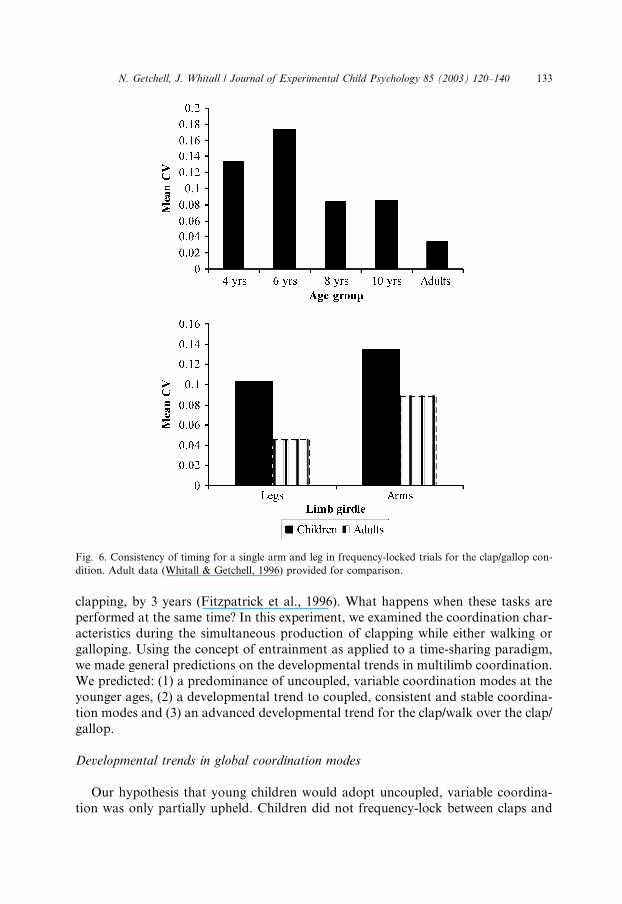
The average values of the coefficients of variation for the participants with locked-
harmonic and unlocked stable trials are presented in Fig. 6. A 4� 2� 2 (age by co-ordination pattern by limb girdle) ANOVA with repeated measures on limb girdleresulted in a main effect for age, F ð3; 44Þ ¼ 7:84, p < :0003, and for limb girdle,F ð1; 44Þ ¼ 8:25, p < :006. A Newman–Keuls post hoc analysis on age groups
showed significant differences between 4 and 8 years (p < :004), 4 and 10 years(p < :004), 6 and 8 years (p < :002) and 6 and 10 years (p < :002) groups. There wereno significant differences between the different coordination patterns, suggesting that
within-limb period stability does not affect between girdle stability.
Differences in walk/clap vs. gallop/clap
We hypothesized that the combination of walking and clapping would show
greater consistency over trials as well as greater frequency locking over time than gal-
loping and clapping. To test consistency across trials, we compared the proportion of
participants who coupled using the same coordination pattern for all three trials be-
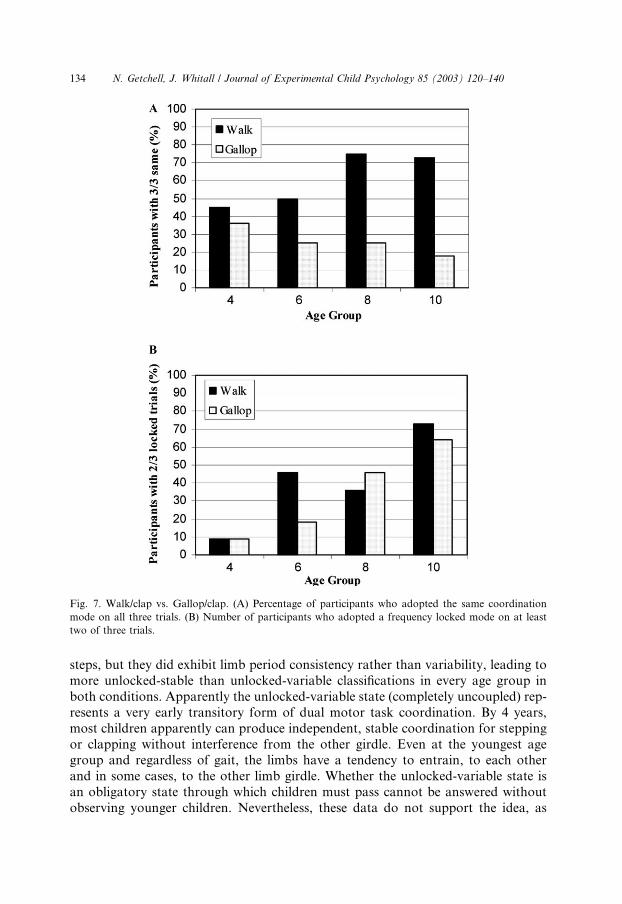
tween the walk/clap and gallop/clap conditions using a v2 goodness of fit test andfound the proportions to differ significantly, v2ð3;N ¼ 40Þ ¼ 295:3, p < :000 (seeFig. 7A). To test for greater frequency locking, we compared the proportion of par-
ticipants who coupled in a locked pattern for at least two trials between each,v2ð3;N ¼ 39Þ ¼ 17:07, p < :0007 (Fig. 7B). Taken together, these results suggest thatchildren couple the walk/clap more easily than the gallop/clap.
Discussion
By about 18 months, toddlers can produce relatively stable walking patterns
(Clark et al., 1988); galloping can occur as early as 2 years (Whitall, 1989), and stable
132 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140
clapping, by 3 years (Fitzpatrick et al., 1996). What happens when these tasks are
performed at the same time? In this experiment, we examined the coordination char-
acteristics during the simultaneous production of clapping while either walking or
galloping. Using the concept of entrainment as applied to a time-sharing paradigm,
we made general predictions on the developmental trends in multilimb coordination.We predicted: (1) a predominance of uncoupled, variable coordination modes at the
younger ages, (2) a developmental trend to coupled, consistent and stable coordina-
tion modes and (3) an advanced developmental trend for the clap/walk over the clap/
gallop.
Developmental trends in global coordination modes
Our hypothesis that young children would adopt uncoupled, variable coordina-tion was only partially upheld. Children did not frequency-lock between claps and
Fig. 6. Consistency of timing for a single arm and leg in frequency-locked trials for the clap/gallop con-
dition. Adult data (Whitall & Getchell, 1996) provided for comparison.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 133
steps, but they did exhibit limb period consistency rather than variability, leading to
more unlocked-stable than unlocked-variable classifications in every age group in
both conditions. Apparently the unlocked-variable state (completely uncoupled) rep-
resents a very early transitory form of dual motor task coordination. By 4 years,
most children apparently can produce independent, stable coordination for stepping
or clapping without interference from the other girdle. Even at the youngest agegroup and regardless of gait, the limbs have a tendency to entrain, to each other
and in some cases, to the other limb girdle. Whether the unlocked-variable state is
an obligatory state through which children must pass cannot be answered without
observing younger children. Nevertheless, these data do not support the idea, as
Fig. 7. Walk/clap vs. Gallop/clap. (A) Percentage of participants who adopted the same coordination
mode on all three trials. (B) Number of participants who adopted a frequency locked mode on at least
two of three trials.
134 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140

some would argue, that frequency-locked, in-phase coordination (absolute coordina-
tion) is intrinsic to the neuromuscular system in that it arises spontaneously with no
apparent external influence or particular intention on the part of the individual
(Sch€ooner & Kelso, 1988; Swinnen, Massion, & Heuer, 1994). Clearly, there is a de-velopmental aspect to acquiring absolute coordination in dual motor task coordina-tion.
Developmental time course of entrainment in dual motor task coordination
We hypothesized and found an age-related trend towards frequency coupling,
consistency across trials and decreased variability, with frequency locking leading to-
wards characteristics of previously analyzed adult performance. As indicated above,
however, a key unexpected finding was the high incidence of frequency unlocked butstable coordination modes that gave way over age to frequency-locked stable coor-
dination modes. These data provide insight into the development of dual motor task
coordination. Specifically, this indicates that children have or need to have stable fre-
quency-locking within girdle coordination prior to and not simultaneous to acquiring
stable frequency-locking between girdles in these types of multilimb tasks.
There is an important distinction to be made between dual task situations and
multilimb coordination. In multilimb coordination tasks, all four limbs are required
to do the same task at once, be it a specified coordination pattern (e.g., simultaneousflexion/extension patterns while sitting in a chair; Jeka, Kelso, & Kiemal, 1993a,
1993b) or an unspecified pattern (e.g., crawling on hands and feet; Whitall, For-
rester, & Getchell, 1997). Plausibly, the developmental time course of unitask multi-
limb coordination may emerge in a different way from dual task multilimb
coordination, with entrainment being simultaneous within and across limb girdles.
For example, prior to crawling, infants rock (simultaneous, in-phase coupling);
crawling begins when infants break this four limb symmetry by reaching forward
with an arm and creating a new, asymmetrical four limb coupling pattern (Goldfield,1989). We already know, however, that locomotor tasks that involve the arms in a
secondary role (upright locomotion) display the development of leg coordination
stability before arm coordination and before between girdle stability although no
one, to our knowledge has examined whether the development of arm coordination
stability is simultaneous to the between-girdle coupling. Given that these locomotor
tasks require no additional attention to the arms (unlike the dual task situation
here), it is plausible that the entrainment profile would not be the same as was found
in this experiment.In the task studied here, when frequency-locking between the arms and legs oc-
curred, the simplest and first available phase-locking necessary to complete the task
was the one arrived at by children from all age groups. When walking while clapping,
participants of all ages adopted a locked-absolute coupling pattern of one clap to one
step. However, locking in this manner would become more complex when the legs
shift to the temporal structure of galloping. In this case, the simplest and first avail-
able rhythm is to lock one clap to every stride, which was the most commonly adopted
pattern in the gallop/clap condition referred to as the locked-harmonic pattern.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 135

An expected age-related trend was the decreased variability of the between-girdle
phase-locking when frequency-locking between the girdles did occur over either task
condition. This becomes notable when considering that age-related increases in loco-
motion velocity would mathematically increase variability as defined by standard de-
viation.3 This finding is consistent with previous studies of bilateral coordinationtasks over this age range, for example, clapping (Fitzpatrick et al., 1996), tapping
(Wolff et al., 1998), and circle drawing (Robertson, 2001). According to coupled os-
cillator models, this indicates that children have increased coupling strength (greater
entrainment) as they get older. One exception to this developmental trend, however,
was displayed by 4-year-olds in the gallop/clap condition, who, surprisingly, showed
the greatest overall stability of any of the age groups studied. These data are repre-
sentative of only three participants who adopted locked frequency-coordination pat-
terns in this task condition. One possible explanation may be that when youngchildren first are able to adopt a very complex locked coordination mode, it must
be tightly coupled. In other words, for complex tasks the degrees of freedom are lim-
ited and some flexibility around a coupling point is actually gained over time. Cer-
tainly there is evidence in this experiment that the gallop/clap dual task was much
more challenging than the walk/clap.
The influence of dual motor task complexity
As hypothesized, the development towards 4-limb entrainment was both slower
and more limited in solutions in the gallop/clap compared to the walk/clap. When
comparing age groups across tasks, the participants performed the walk/clap using
the same proportions of coordination patterns as adults; in the gallop/clap, these
adult-like proportions were never attained. By 10 years, only 65% of the present par-
ticipants adopted a locked-harmonic coordination mode, compared to adulthood,
where 95% of the participants adopted the locked-harmonic coupling pattern (Whi-
tall & Getchell, 1996). Although 4-limb entrainment in the gallop/clap took longerover the course of childhood, by adulthood, it was almost exclusively one behavioral
form. Compare this to the walk/clap. In that condition, the youngest group had
nearly identical proportions of locked and unlocked classifications as in the gal-
lop/clap, but over time, a less clear developmental pattern of increasing four-limb en-
trainment to frequency-locked states existed. In fact, 6-year-olds adopted locked
patterns more frequently than 8-year-olds did, and both 10-year-olds and adults
adopted locked patterns only about 60% of the time. The remainder of the trials were
primarily uncoupled-stable. This suggests a multistability of two limb and four limbattractive coordination patterns for clapping and walking as found in the adults.
Why does the gallop/clap have a stronger developmental trend towards four-limb
entrainment than the walk/clap? One explanation may relate to task complexity and
attentional demands. One must specify and maintain an approximate 66/33 relation
3 As the absolute time of a step cycle decreases, the amount of time representing 15% or less of phase
dispersion decreases as well. Therefore, faster trials could yield greater percentages of phase dispersion
even if the absolute time of phase locking remains the same.
136 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140

between the legs while galloping—an out of phase rhythm that presumably requires
attentional input. By entraining the upper limbs to the temporal pattern of the lower
limbs, one can reduce complexity, as only one rhythm needs to be matched by both
limb girdles. The four-limb entrainment for the gallop/clap condition appears to be a
strongly attractive state, as it is adopted more frequently and more consistentlyacross time. Walking while clapping is less complex and requires less attention; this
might result in a weaker attraction towards four-limb entrainment because both
tasks can be completed successfully without interference between them.
A second and perhaps complementary explanation comes from the nature of non-
linear oscillators. A number of researchers (Jeka et al., 1993a, 1993b; Kelso & Jeka,
1992; Serrien & Swinnen, 1997; Treffner & Turvey, 1996) have proposed that symme-
try plays an important role in coupling patterns seen in non-linear oscillators. When
four limbs oscillate, within-girdle limb pairs couple more strongly, and tend to showright/left symmetry and either in-phase or anti-phase coupling (Jeka et al., 1993b).
Galloping forces the legs to adopt an asymmetric relation, which creates a rhythmi-
cal bias into the four-limb system. In essence, the coordination pattern of the gallop
acts as a driving force, creating a strong attraction towards four-limb entrainment.
From an evolutionary perspective, there is a biological purpose for flexible en-
trainment in the walk/clap condition. During bipedal locomotion, humans are far
more likely to walk than gallop from place to place. In such situations, strong cou-
pling among all limbs may be detrimental in functional tasks that require the handsto act independently of the legs. Thus it would be desirable to adopt uncoupled but
stable patterns even at the earliest ages. The continued presence of uncoupled-stable
classifications across age groups in the walk/clap condition may represent the range
of coordination modes available to successfully complete the task. Furthermore,
from a developmental perspective, it is clearly desirable to change from a ‘‘weakly
coupled, unstable state’’ to a ‘‘strongly coupled, stable state’’ and finally to a ‘‘flex-
ibly coupled state where both stability and adaptability to voluntary and involuntary
perturbations can co-exist’’ (Whitall, 1997). The flexibility of these particular dualmotor task conditions was explored in our previous adult study by asking partici-
pants to intentionally change their coordination states (Whitall & Getchell, 1996).
However, pilot testing indicated that children would not be able to accomplish these
kinds of tasks at all and therefore, in the current study, we asked the children to per-
form more practice trials (eight additional trials for a total of 11) of the two dual-
task conditions in order to look for short-term adaptations. As seen in Table 1, chil-
dren of all ages and in both conditions rarely became less coupled over the practice
trials. In both conditions, children tended to either maintain their coupling mode orbecome more coupled with practice. This effect is most obvious in the clap/gallop
condition, where children of all ages initially displayed looser or uncoupled coordi-
nation modes.
In general, we can say that the children are not highly skilled in these dual tasks
but that they demonstrate adult-like coordinative behavior under self-paced condi-
tions in walk/clap and approach it in gallop/walk by 10 years. Unlike other studies
of children�s developing bilateral coordination (e.g., Wolff et al., 1998) we will notdraw any inferences between these data and the growth of neural mechanisms such
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 137

as the myelination of the corpus callosum simply because our children were not
adult-like in one of the two tasks and were far from skillful in the other. Because ev-
idence suggests that myelination of the corpus callosum may be complete by 10 years
(Jeeves, Silver, & Milner, 1988), this mechanism is not sufficient (and at best onlynecessary) to the skilled performance of our task. We suggest, in fact, that the devel-
opment of these specific dual motor tasks (and their variations) might usefully be
compared in the future to the development of attentional processes as well as stand-
ing on their own as a method of studying dual motor task entrainment. Future re-
search could explore earlier ages to determine whether an uncoupled-variable state
of coordination is an obligatory initial state before entrainment of limbs. Alterna-
tively, one might focus on the role of intention and attention in dual motor task co-
ordination since the present paradigm, while giving specific instructions for the armsand legs (within girdle task level), did not specify a particular dual motor task coor-
dination (between girdle task level). The results might change if a specific intention to
adopt a coordination pattern and attention to a particular pattern are invoked rather
than allowed to vary.
Conclusions
Several important developmental trends in the adoption of dual motor tasks co-
ordination appear to exist. First, children—even younger ones—tend to stably coor-
dinate their limbs at some level. Even at early ages and despite complexity of
conditions, all but a few children were able to accomplish these dual motor tasks
with fairly stable coordination either between all limbs (mostly older children) or,
at best, within the arms and legs (younger and older children). The manner in which
the limbs coupled changed developmentally, in that the amount of frequency locking
increased with age and variability of adopted coordination modes across trials andtiming of limb coupling within trials decreased with age. The development of en-
trainment between two independent tasks is characterized by within-limb girdle sta-
bility before between-limb girdle stability. Differences in task complexity between the
walk and gallop lead to a slower development time-scale for the latter. Although
Table 1
Change in adopted coordination mode over 11 trials of practice
Change Coordination pattern
Walk/clap Gallop/clap
Age groups Age groups
4 6 8 10 4 6 8 10
Decrease 0 2 0 1 1 3 2 1
Same 7 6 8 6 3 3 5 2
Increase 3 3 3 2 6 5 2 5
Note. ‘‘Decrease’’ indicates a change to a less coupled mode, ‘‘increase’’, a change to a more coupled
mode, and ‘‘Same’’, no change in coupling.
138 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140

children can clap and gallop individually by (and generally well before) 4 years, the
oscillatory limb behaviors are not adult-like by 10 years in the more complex task. It
is not clear what leads to the change in this specific dual motor task coordination
between children and adults except that it is unlikely to be related solely to motoric
task specific practice or experience since adults do not often gallop let alone clap andgallop at the same time.
Acknowledgments
The work was supported by a National Institutes of Health Award (R29 HD
28157) to the second author. We would like to thank the participants and also Larry
Forrester and Carolyn Kelsey for their work on data collection and analysis, and Dr.Susan Hall as well as two anonymous reviewers for helpful comments on earlier ver-
sions of the manuscript.
References
Amazeen, P. G., Amazeen, E. L., & Turvey, M. T. (1998). Dynamics of human intersegmental
coordination: Theory and research. In D. Rosenbaum, & C. Collyer (Eds.), Timing of behavior (pp.
237–260). Cambridge, MA: MIT Press.
Clark, J., & Whitall, J. (1989). Changing patterns of locomotion: From walking to skipping. In M.
Woollacott, & A. Shumway-Cook (Eds.), Development of posture and gait across the lifespan (pp. 128–
151). Columbia: Univeristy of South Carolina Press.
Clark, J. E., Whitall, J., & Phillips, S. J. (1988). Human interlimb coordination and control: The first 6
months of independent walking. Developmental Psychobiology, 21, 445–456.
Collins, J. J., & Stewart, I. N. (1992). Symmetry-breaking bifurcation: A possible mechanism for 2:1
frequency-locking in animal locomotion. Journal of Mathematical Biology, 30, 827–838.
Donker, S. F., Beek, P. J., Wagenaar, R. C., & Mulder, T. (2001). Coordination between arm and leg
movements during locomotion. Journal of Motor Behavior, 33, 86–102.
Fitzpatrick, P., Schmidt, R. C., & Lockman, J. J. (1996). Dynamical patterns in the development of
clapping. Child Development, 67, 2691–2708.
Goldfield, E. (1989). Transition from rocking to crawling: Postural constraints on infant movement.
Developmental Psychology, 25, 913–919.
Haken, H., Kelso, J. A. S., & Bunz, H. (1985). A theoretical model of phase transitions in human hand
movements. Biological Cybernetics, 51, 347–356.
Hresko, W. P., Reid, D. K., Hammill, D. D., Ginsburg, H. P., & Baroody, A. J. (1988).Manual for screen.
Screening children for related early educational needs. Austin, TX: Pro-Ed.
Jeeves, M. A., Silver, P. H., & Milner, A. D. (1988). Role of the corpus callosum in the development of a
bimanual motor skill. Developmental Neuropsychology, 4, 305–323.
Jeka, J. J., Kelso, J. A. S., & Kiemal, T. (1993a). Spontaneous transitions and symmetry: Pattern dynamics
in human four-limb coordination. Human Movement Science, 12, 627–651.
Jeka, J. J., Kelso, J. A. S., & Kiemal, T. (1993b). Pattern switching in human multilimb coordination
dynamics. Bulletin of Mathematical Biology, 55, 829–845.
Kay, B. A., Kelso, J. A. S., Saltzman, E. L., & Sch€ooner, G. (1987). The space–time behavior of single andrhythmical movements: Data and model. Journal of Experimental Psychology: Human Perception and
Performance, 13, 178–192.
Kay, B. A., & Warren, W. (1998). A dynamic model of coupling between posture and gait. In D.
Rosenbaum, & C. Collyer (Eds.), Timing of behavior (pp. 293–322). Cambridge, MA: MIT Press.
N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140 139

Kelso, J. A. S., & Jeka, J. J. (1992). Symmetry breaking dynamics of human multilimb coordination.
Journal of Experimental Psychology: Human Perception and Performance, 18, 645–668.
Kelso, J. A. S., & Scholz, J. P. (1985). Cooperative phenomena in biological motion. In H. Haken (Ed.),
Complex systems: Operational approaches in neurobiology, physical systems, and computers (pp. 124–
149). Berlin: Springer.
Muzii, R. A., Lamm, C., & Gentile, A. M. (1984). Coordination of the upper and lower extremities.
Human Movement Science, 3, 337–354.
Robertson, S. D. (2001). Development of bimanual skill: The search for stable patterns of coordination.
Journal of Motor Behavior, 33, 114–126.
Sch€ooner, G., & Kelso, J. A. S. (1988). A dynamic pattern theory of behavioral change. Journal of
Theoretical Biology, 135, 501–524.
Serrien, D. J., & Swinnen, S. P. (1997). Coordination constraints induced by effector combination under
isofrequency and multifrequency conditions. Journal of Experimental Psychology: Human Performance
and Perception, 23, 1493–1510.
Sutherland, D. H., Olshen, R., Cooper, L., & Woo, S. L. Y. (1980). The development of mature gait.
Journal of Bone and Joint Surgery, 62A, 336–353.
Swinnen, S. P., Massion, J., & Heuer, H. (1994). Topics on interlimb coordination. In S. Swinnen, J.
Massion, H. Heuer, & P. Casaer (Eds.), Interlimb coordination: Neural, dynamical, and cognitive
constraints (pp. 2–25). San Diego: Academic Press.
Thelen, E. (1986). Development of coordinated movement: Implications for early human development. In
M. G. Wade, & H. T. A. Whiting (Eds.), Motor development in children: Aspects of coordination and
control (pp. 107–124). Dordrecht, Germany: Martinus Nijhoff.
Treffner, P. J., & Turvey, M. T. (1996). Symmetry, broken symmetry, and handedness in bimanual
coordination dynamics. Experimental Brain Research, 107, 463–478.
Turvey, M. T., Rosenblum, L. D., Schmidt, R. C., & Kugler, P. N. (1986). Fluctuations and phase
symmetry in coordinated rhythmic movements. Journal of Experimental Psychology: Human
Perception and Performance, 12, 564–583.
Von Holst, E. (1973). The behavioral physiology of animals and man: The collected papers of Erich Von
Holst (R. Martin, Trans.) (Vol. 1,. London: Methuen (Original work published, 1937).
Whitall, J. (1989). A developmental study of the interlimb coordination in running and galloping. Journal
of Motor Behavior, 21, 409–428.
Whitall, J. (1991). The developmental effect of concurrent cognitive and locomotor skills: Time-sharing
from a dynamical perspective. Journal of Experimental Child Psychology, 51, 245–266.
Whitall, J. (1997). The development of time-sharing: An old paradigm from a new perspective. Motor
Development: Research & Reviews, 1, 171–199.
Whitall, J., & Getchell, N. (1996). Multilimb coordination patterns in simultaneous dissimilar upper and
lower limb tasks. Human Movement Science, 15, 129–155.
Whitall, J., Forrester, L., & Getchell, N. (1997). The effect of non-specific task constraints on quadrupedal
locomotion: I Interlimb coordination. Motor Control, 1, 138–160.
Wolff, P. H., Kotwica, K., & Obregon, M. (1998). The development of interlimb coordination during
bimanual finger tapping. International Journal of Neuroscience, 93, 7–27.
140 N. Getchell, J. Whitall / Journal of Experimental Child Psychology 85 (2003) 120–140




















