
Honduran Folk Entomology
29
285 Reports Honduran Folk Entomology 1 jeffery w. bentley and gonzalo rodrı ´guez Casilla 2695, Cochabamba, Bolivia (bentley@ albatros.cnb.net). 17 x 00 Do people discover their world or create it? If people discover the categories of nature, then folk taxonomy of living things should have formal similarities cross-cul- turally because of the biological integrity of our planet. If people create the categories of nature, each culture should order living things uniquely (see Berlin 1992, Brown 1984, Hunn 1990, Ellen 1993). The universalist or intellectualist school claims that some living things are so perceptually salient, so biologically real, that they are “crying out to be named” (Berlin 1992:53). American college students faced with a pile of bird skins from the Peruvian Amazon classify them the same way as Jivaro and ornithologists on the basis of the birds’ morphology (Boster 1987, Boster, Berlin, and O’Neill 1986). The cul- tural relativist or utilitarian school observes that plants and animals are named because people use them; “sur- vival placed a premium on knowledge of utilitarian value” (Hunn 1990:117; see also Conklin 1962:129). The Sahaptin of the Columbia River have an extensive folk taxonomy of edible plants, while hundreds of species of culturally insignificant flowering plants are perempto- rily dismissed as “just a flower” (Hunn 1990:198–99). We try to reconcile the utilitarian and universalist perspec- tives by showing how cultural importance and the ease of observation of animal morphology interact in influ- encing which species are named, which are lumped into residual categories (Hays 1983), which are confused, and which are ignored in ethnobiological systems. Many re- searchers have noticed that cultural importance and morphological attributes are key to folk classifications (Hays 1982; Brown 1984; Posey 1984; Arioti 1985; Atran 2001 by The Wenner-Gren Foundation for Anthropological Re- search. All rights reserved 0011-3204/2001/4202-0005$1.00. 1. RENARM supported Bentley’s work and GTZ funded Rodrı ´- guez’s. The Departamento de Proteccio ´n Vegetal, Zamorano, pro- vided office space and materials. Ana Gonza ´ lez, Catrin Meir, Harold C. Conklin, Mauricio Bellon, Phil Young, Stephen Sherwood, and eight anonymous referees made valuable comments. Ana Gonza ´lez helped design and teach the insect ecology seminar, offered to farm- ers on various occasions, where we learned some of the folk names recorded in this paper. Ronald Cave identified insects since 1987 and revised an earlier draft. The literature review was made possible by the help of Ricardo Godoy and the staff of the Tozzer Library at Harvard University. Keith Andrews created the institutional and intellectual environment. Our biggest debt is to the rural people of Honduras. 1985; Ellen 1986, 1993; Berlin 1992) without distinguish- ing their roles. Paul Sillitoe’s (1996) ethnography of the folk ecology of the Wola people of Highland New Guinea is scientif- ically sophisticated and reveals a comprehensive, sym- pathetic knowledge of the Wola’s view of their land. Both universalist and utilitarian explanations apply here. Wola knowledge of some topics is highly detailed; for example, there are 64 named varieties of sweet potato. At the same time, Wola are completely unaware of, for example, microorganisms, including the ones that cause disease, and have only a “hazy” idea of the relationship of some grubs to their adult forms and only partial knowledge of insect metamorphosis. They label butter- flies, spiders, and some other major morphotypes of ar- thropods but do not name various beetles and larvae at all. They do not give separate names to the many kinds of grubs, even the ones that they realize are different species, because these grubs are not important to them. Yet for topics that are important to them, such as rat damage to their gardens, local knowledge is not so much incomplete as highly contradictory to modern science. For example, Wola believe that if someone who has re- cently eaten meat sees a garden, that crop may be dev- astated by a rat attack. Farmers build screens of cane grass to shield their sweet potato plants from the sight of possible meat eaters who may be passing by. In the modest hypothetical scheme we propose in this paper, folk knowledge has an uneven texture which can be explained by comparing the cultural importance (util- ity) of the domain with its ease of observation (conspic- uousness, perceptual salience). We say that a species is easy to observe if it is large, social, colorful, abundant, noisy, and diurnal (Berlin 1990:23–24; 1992:81; Atran 1987:150; Bentley 1992a, b, 1993, 1994). Many species go unobserved because they are very small, solitary, cryp- tic, rare, silent, or nocturnal. Hunn (1977) emphasizes perceptual salience: the more distinctive a species is, the more likely it is to be named. The boundaries of biolog- ical categories are formed along the lines of disconti- nuities in nature. While this notion is thoughtful and logical, we propose that perceptual salience is less rel- evant than ease of observation; species that look very much alike are commonly named if there is a cultural reason to do so. Of the common grain crops, maize is by far the most salient, followed by rice, while rye, oats, barley, and wheat are nearly identical to a city person, but farmers who grow these crops distinguish them all. Their ease of observation and the motivation of cultural importance allow farmers to name them in spite of their lack of perceptual salience. By “cultural importance” we mean perceived impor- tance within a specific culture, whether useful or harm- ful. The utilitarian school has emphasized economic use.
-
Upload
uuniversidaddepanam -
Category
Documents
-
view
1 -
download
0
Transcript of Honduran Folk Entomology

285
Reports
Honduran Folk Entomology1
j effery w. bentley and gonzalo rodr ıguezCasilla 2695, Cochabamba, Bolivia ([email protected]). 17 x 00
Do people discover their world or create it? If peoplediscover the categories of nature, then folk taxonomy ofliving things should have formal similarities cross-cul-turally because of the biological integrity of our planet.If people create the categories of nature, each cultureshould order living things uniquely (see Berlin 1992,Brown 1984, Hunn 1990, Ellen 1993). The universalistor intellectualist school claims that some living thingsare so perceptually salient, so biologically real, that theyare “crying out to be named” (Berlin 1992:53). Americancollege students faced with a pile of bird skins from thePeruvian Amazon classify them the same way as Jivaroand ornithologists on the basis of the birds’ morphology(Boster 1987, Boster, Berlin, and O’Neill 1986). The cul-tural relativist or utilitarian school observes that plantsand animals are named because people use them; “sur-vival placed a premium on knowledge of utilitarianvalue” (Hunn 1990:117; see also Conklin 1962:129). TheSahaptin of the Columbia River have an extensive folktaxonomy of edible plants, while hundreds of species ofculturally insignificant flowering plants are perempto-rily dismissed as “just a flower” (Hunn 1990:198–99). Wetry to reconcile the utilitarian and universalist perspec-tives by showing how cultural importance and the easeof observation of animal morphology interact in influ-encing which species are named, which are lumped intoresidual categories (Hays 1983), which are confused, andwhich are ignored in ethnobiological systems. Many re-searchers have noticed that cultural importance andmorphological attributes are key to folk classifications(Hays 1982; Brown 1984; Posey 1984; Arioti 1985; Atran
� 2001 by The Wenner-Gren Foundation for Anthropological Re-search. All rights reserved 0011-3204/2001/4202-0005$1.00.
1. RENARM supported Bentley’s work and GTZ funded Rodrı-guez’s. The Departamento de Proteccion Vegetal, Zamorano, pro-vided office space and materials. Ana Gonzalez, Catrin Meir, HaroldC. Conklin, Mauricio Bellon, Phil Young, Stephen Sherwood, andeight anonymous referees made valuable comments. Ana Gonzalezhelped design and teach the insect ecology seminar, offered to farm-ers on various occasions, where we learned some of the folk namesrecorded in this paper. Ronald Cave identified insects since 1987and revised an earlier draft. The literature review was made possibleby the help of Ricardo Godoy and the staff of the Tozzer Library atHarvard University. Keith Andrews created the institutional andintellectual environment. Our biggest debt is to the rural people ofHonduras.
1985; Ellen 1986, 1993; Berlin 1992) without distinguish-ing their roles.
Paul Sillitoe’s (1996) ethnography of the folk ecologyof the Wola people of Highland New Guinea is scientif-ically sophisticated and reveals a comprehensive, sym-pathetic knowledge of the Wola’s view of their land. Bothuniversalist and utilitarian explanations apply here.Wola knowledge of some topics is highly detailed; forexample, there are 64 named varieties of sweet potato.At the same time, Wola are completely unaware of, forexample, microorganisms, including the ones that causedisease, and have only a “hazy” idea of the relationshipof some grubs to their adult forms and only partialknowledge of insect metamorphosis. They label butter-flies, spiders, and some other major morphotypes of ar-thropods but do not name various beetles and larvae atall. They do not give separate names to the many kindsof grubs, even the ones that they realize are differentspecies, because these grubs are not important to them.Yet for topics that are important to them, such as ratdamage to their gardens, local knowledge is not so muchincomplete as highly contradictory to modern science.For example, Wola believe that if someone who has re-cently eaten meat sees a garden, that crop may be dev-astated by a rat attack. Farmers build screens of canegrass to shield their sweet potato plants from the sightof possible meat eaters who may be passing by.
In the modest hypothetical scheme we propose in thispaper, folk knowledge has an uneven texture which canbe explained by comparing the cultural importance (util-ity) of the domain with its ease of observation (conspic-uousness, perceptual salience). We say that a species iseasy to observe if it is large, social, colorful, abundant,noisy, and diurnal (Berlin 1990:23–24; 1992:81; Atran1987:150; Bentley 1992a, b, 1993, 1994). Many speciesgo unobserved because they are very small, solitary, cryp-tic, rare, silent, or nocturnal. Hunn (1977) emphasizesperceptual salience: the more distinctive a species is, themore likely it is to be named. The boundaries of biolog-ical categories are formed along the lines of disconti-nuities in nature. While this notion is thoughtful andlogical, we propose that perceptual salience is less rel-evant than ease of observation; species that look verymuch alike are commonly named if there is a culturalreason to do so. Of the common grain crops, maize is byfar the most salient, followed by rice, while rye, oats,barley, and wheat are nearly identical to a city person,but farmers who grow these crops distinguish them all.Their ease of observation and the motivation of culturalimportance allow farmers to name them in spite of theirlack of perceptual salience.
By “cultural importance” we mean perceived impor-tance within a specific culture, whether useful or harm-ful. The utilitarian school has emphasized economic use.

286 F current anthropology
table 1Categories of Honduran Folk Entomology andExamples
CulturallyUnimportant
CulturallyImportant
Easy toobserve
Mud dauber waspsEarwigsSpiders
Social beesSocial wasps
Difficult toobserve
Parasitic waspsNematodes
Pest caterpillars(especially earlyinstars andLepidopterareproduction)
The universalist school has countered that many ani-mals are named that are not strictly useful (Hays 1982:93; Berlin 1992:89; see also Brown 1992). Defining “cul-turally important” to include harmful species and notjust useful ones gives the utilitarian argument widerrange.
Cultural importance and ease of observation influenceeach other. People take the time to observe useful insectssuch as bees and harmful ones such as crop pests. Hon-duran peasants ignore harmless and useless species suchas mud dauber wasps (Hymenoptera: Sphecidae), whichdo not sting and make nothing people use, despite thefact that some of these wasps make mud nests shapedlike pan pipes and others nests shaped like footballs, allcommon along with nests of other shapes aroundfarmhouses.
Arguing against the role of cultural relativism in folkclassification, Berlin (1992:80) writes, “To the extent thatone is able to predict which plants and animals in somesociety will be named without prior knowledge of thecultural significance of those organisms, the Utilitarianargument loses much of its force.” Several cross-culturaldomains of plants and animals are, however, importantenough to be classified in detail in most languages. Theseinclude domesticated and game animals; edible plants,seasonings, and medicines; firewood; cordage and tex-tiles; weeds and other crop pests; pests of the humanbody and of our animal intimates; dangerous or painfulorganisms; and anything used for games, toys, ornament,ritual, or art.
Many large animals and vascular plants are either use-ful, harmful, or impossible to ignore and thereforenamed. However, folk classification of entomofauna of-ten lumps thousands of species into a single categorybecause the creatures are small and hard to see (Atran1987; see also Berlin 1992:81). Insects are a challenge toclassify, even for entomological taxonomists, because oftheir sheer number—30 to 50 million species (Erwin1988; Wilson 1988; 1992:143). Because insects evolvedbefore Pangea assembled and broke up again, individualinsect families tend to range farther than vertebrate fam-ilies (DiMichele et al. 1992). Insects persist in interactingwith people, and therefore ethnoentomology can be usedto test cross-cultural hypotheses with any human group.A traditional community is able to name most of thebirds, mammals, and trees in the local environment butmust cram several million insect species into (at most)a few hundred categories. Insects are perfect for illus-trating the biological and cultural criteria a communityuses to name, lump, confuse, or ignore living things. Wepresent a case study of Honduran ethnoentomology.
To reject the universalist hypothesis, we would needto find folk taxonomies ordered along the lines of theutility of the organisms; folk names for taxa would bebased on cultural criteria (e.g., use, harm), and folkknowledge would be deeper for the culturally importantcreatures than for the perceptually salient ones. To rejectthe utilitarian hypothesis, we would need to find folktaxonomies ordered along the lines of the creatures’ mor-phology; plants and animals would be named for their
physical characteristics, and folk knowledge would bedeeper for the easy-to-observe than for the culturally im-portant. According to the utilitarian school, folk tax-onomies should be based on taxa that the people in aspecific culture use (e.g., Hunn 1982, 1990). Supportersof the universalist hypothesis argue that all languageshave words for the major morphotypes of insects andhave other similarities (Berlin 1992). We show that thesetwo perspectives are complementary, since (1) folk tax-onomies show both tendencies, (2) some folk categoriesare named for their roles in local culture and others fortheir biological properties, and (3) folk knowledge isdeepest for creatures that are both culturally importantand easily observed.
We divide rural Honduran folk entomology into fourcategories according to cultural importance and ease ofobservation: (1) the culturally important and easily ob-served (e.g., bees, social wasps), (2) the easily observedbut culturally unimportant (e.g., mud dauber wasps, ear-wigs, spiders), (3) the culturally important but difficultto observe (e.g., pest caterpillars), and (4) the culturallyunimportant and difficult to observe (e.g., parasiticwasps, nematodes). We discuss (nonstandard, rural) Hon-duran (Spanish) folk knowledge, taxonomy, and seman-tics of insects and other terrestrial invertebrates for eachcategory (table 1).
Each category has its own epistemology, taxonomicstructure, and semantics. Easily observed and culturallyimportant taxa have deep folk knowledge2 and hierar-chical taxonomies. They tend to be named for their phys-ical characteristics more than for their cultural impor-tance, and some have semantically opaque names. Foreasily observed but culturally unimportant taxa folkknowledge is thinner and taxonomies are broad and shal-low. Their names tend to reflect their appearance (na-ture). For the culturally important but difficult-to-ob-serve taxa folk knowledge is complex, and much of it
2. We originally used the term “thick knowledge.” We are gratefulto Peter Baker for pointing out that in British English one of themeanings of “thick” is “stupid.”

Volume 42, Number 2, April 2001 F 287
clashes with modern science.3 Taxonomies include bi-nomial folk names that reflect the insects’ interactionwith humans. Difficult-to-observe and culturally un-important organisms are not known, named, orclassified.
We identified each creature as culturally important ornot and as easy or difficult to observe. We ranked a crea-ture as culturally important if we knew that Honduranfarmers considered it a pest, a danger or a nuisance, aplaything, or of any utility at all. It was harder to identifyorganisms as easy or difficult to observe. We classed so-cial insects, larger ones, and brightly colored flying onesas easily observed. We considered cryptic and nocturnalanimals (unless they were social) as difficult to observe.Although some caterpillars are quite conspicuous, weclassified most of them as difficult to observe; many arecryptically colored, and most are too small to be verynoticeable until their later instars. Few of the creatureswere difficult to classify by cultural importance. We did,however, end up classifying all bees as “important” be-cause campesinos distinguish the otherwise unimpor-tant ones from the troublesome or useful ones. A fewtaxa were hard to classify as easy versus difficult to ob-serve (notably the pest Lepidoptera larvae), but our find-ings would have been little altered by reclassifying them.In future work we may want to make “ease of obser-vation” a longer scale, with a category for insects thatare themselves conspicuous but for which key aspectsof their lives are difficult to observe.
folk knowledge
The folk knowledge of culturally important and easilyobserved groups is deep. Almost all descriptions of folkknowledge have dealt with culturally important and eas-ily observed domains, and this has produced the im-pression that all folk knowledge is deep. Scholars of tra-ditional people tend to discuss topics of importance tothe people themselves, topics on which they are experts.Traditional agriculture in general is dependent on elab-orate systems of folk knowledge (Netting 1993:321; seealso Wilken 1987 and Wilk 1991).
We agree that folk knowledge can be quite sophisti-cated. For example, Honduran campesinos can describethe brood chambers, worker and queen morphology, andhoney quality of the bees whose hives they harvest (seePosey and Carmago 1985). They understand that beesand wasps lay eggs and that the workers tend the brood.They distinguish species of bees for their utility: somegive honey and others do not. Some of the honeys aremedicinal; the honey of the jimerito (Trigona angustala,Hymenoptera: Apidae) is used as an ointment for injuredeyes (see Chittampalli and Mulcahy 1990). Some honeysare merely edible, and others are considered poisonous.
3. We avoid the phrase “Western science.” Many of the Easterncountries, such as Japan, now have more than a passing familiaritywith “Western” science, while much of the culture of Latin Amer-ican countries such as Honduras is of Western European (Spanish)origin. One could write of “Northern” science, but it makes moresense to omit the geographic stereotyping.
The bees must also be distinguished because one stings,some bite, one secretes a blister-causing liquid, and oth-ers are passive.
Folk knowledge about these insects is sometimesahead of current entomological thought. For example,campesinos told Bentley that leaf-cutter ants (Hymen-optera: Formicidae: Attini) have a nahual (an animal soulcompanion), a snake or a lizard. They said that digginginto a nest until one found the lizard would cause thenest to die (see also Hunn 1977:262). While digging upleaf-cutter ant nests with campesinos we have seen acoral snake emerge from one of the ant tunnels and un-earthed a nest of reptile eggs on a bed of spent leaf tissuein a chamber. It is apparent that leaf-cutter ants do havereptilian commensals (see Holldobler and Wilson 1990:471). Again, dozens of campesinos told us that socialwasps eat flower nectar, but entomologist colleagues in-sisted that social wasps preyed on insects. Formal re-search in vespid diet confirms both ideas. Adult socialwasps drink nectar but forage for caterpillars and otherinsects to feed to their brood (Reeve 1991, Gadagkar1991, Jeanne 1991, Hunt 1991).
Most of the folk knowledge of the easily observed butnot culturally important taxa tends to agree with modernscience: dragonflies live around water; spiders weavewebs; mud dauber wasps have spiders in their nests; Junebeetles emerge with the first rains; dung beetles roll ballsof manure; and earwigs live in maize plants. These ex-amples may seem trivial; they are facts that entomolo-gists and Honduran farmers know but few find remark-able. Campesinos know of some predatory insects, suchas army ants, but few know that social wasps hunt forinsects or that many other insects are beneficial preda-tors of insect pests (Gonzalez 1993). Honduran farmersknow that spiders and fire ants prey on insects but notthat there are many other serious predators of other in-sects. Once we had shown farmers wasps and ants prey-ing on pests, they continued to notice it on their own.This new information did not clash with local knowl-edge; folk knowledge is thinner than modern science fortopics that are not culturally important.
For the culturally important but difficult-to-observetaxa, local knowledge and modern science part company.Beliefs in caterpillars that rain from the sky, pests createdby spontaneous generation, and wasps that lay papaya-eating worms are some examples. Farmers watchingtheir maize fields being eaten by caterpillars that seemto have come from nowhere may be forced to adopt ex-planations that are consistent with local observationsbut not with modern science. Without the benefit ofdevices such as microscopes and without conceptualtools such as germ theory and metamorphosis, peoplemay conclude that disease is caused by spirits (Last 1981)and caterpillars are produced by spontaneous generation(Bentley, Rodrıguez, and Gonzalez 1994).
Anthropologists have been reluctant to discuss gapsand misunderstandings in folk knowledge. Chambers(1983:84) has criticized ethnoscience for focusing oncompetent informants and large, well-known domains.Vayda and Setyawati (1995) write that cognitive anthro-

288 F current anthropology
table 2Characteristics of Folk Knowledge in EachCategory
CulturallyUnimportant
CulturallyImportant
Easy toobserve
Thin butconsistentwith for-mal (so-calledWestern)science
Deep, muchof it un-known toformalscience
Difficult toobserve
Absent Complexbut ofteninconsis-tent withformalscience
pological accounts of traditional knowledge discuss lin-guistic distinctions of little practical relevance and aredeficient in describing knowledge and ignorance aboutinsects which could be useful for informing pest-man-agement practices.
Of all the insects, Honduran campesinos generally rec-ognize only bees and wasps as reproducing sexually.They say that caterpillars reproduce by spontaneous gen-eration. The reproduction of pest Lepidoptera is econom-ically important but difficult to observe. The cogollero(fall armyworm, Spodoptera frugiperda [Lepidoptera:Noctuidae]), a maize pest, for example, is a dull graymoth as an adult. Few farmers name the moth; evenfewer notice its egg masses, cream-colored blobs on fenceposts and maize leaves. The tiny larvae hatch and glidethrough the air on silk threads that they spin. They landand search for a maize whorl to live in and feed on.Honduran farmers notice them when the caterpillarshave molted two or three times and grown big enoughto be easily seen and to cause enough damage to worryabout. Cogolleros pupate in the soil or in the maize ear.The brown pupae escape rural people’s attention. Othertraditional peoples have misunderstood insect reproduc-tion by generally failing to grasp the notion of meta-morphosis (see Winarto 1996).
There is little or no folk knowledge about the taxa thatare culturally unimportant and difficult to observe. Hon-duran farmers do not recognize the causal agents of dis-ease (Bentley 1990, 1991), and most of them (along withanthropologists and most other nonentomologists) areunaware of the existence of insect parasitoids (of otherinsects), especially of the abundant but almost micro-scopic parasitic wasps.
In summary (see table 2), for insects that are culturallyimportant and easily observed, folk knowledge is deep:local people often know more about them than scientistsdo. This local knowledge can be empirically verified bythe scientific method. For insects that are not culturallyimportant but are easily observed, folk knowledge isthin: local people know them in a way that scientistscan understand, although local knowledge may be lesscomplete than that of specialized natural scientists. Lo-cal knowledge of the culturally important but difficultto observe is gritty: local people may have beliefs andperceptions which are at odds with scientific notions andcannot always be tested with the scientific method.About insects that are difficult to observe and of no cul-tural importance, local people know very little.
taxonomy
Berlin (1992) divides folk taxonomies into hierarchicallevels: kingdom, life-form, intermediate, generic, spe-cific, and varietal. There is an obvious similarity withformal biology: kingdom, phylum, class, order, family,genus, and species. The key Berlinian level is the generic,which includes the most basic primary meaningful cat-egories; their labels are simple (Berlin 1992:27; Conklin1969:106). Folk species usually have binomial labels (Ber-
lin 1992). Intermediate categories are rare (Brown 1984:4; Berlin 1992:27).
Our study deals with a single life-form, insecto. (TheStandard Spanish word bicho is rarely used in Honduras.)Like Brown’s (1984:16) WUG, insecto includes not justinsects but other terrestrial invertebrates. Spiders andcentipedes are insectos and so are slugs, which are mol-lusks. Each of the four categories of taxa has its owntaxonomic properties. Culturally important and easilyobserved taxa are ordered in hierarchies: they are the onlytaxa with intermediate categories. Some folk genera arepolytypic (divided into folk species). Culturally unim-portant but easily observed taxa are lists of genericnames, without categories of intermediate or specificrank. Culturally important but difficult-to-observe taxamay be taxonomically quite different from those of mod-ern science. There are some polytypic folk genera, withspecies labeled with productive binomials. Culturallyunimportant and difficult-to-observe species escapeclassification.
Outlining a test of the universalist hypothesis, Berlin(1992:267) predicts that the following morphotypes (iffound locally) are likely to be named in any ethnobio-logical system of classification: ants, wasps, bees, flies,butterflies and moths, grasshoppers, dragonflies, cicadas,ticks, roaches, beetles/bugs, weevils, spiders, scorpions,fleas/lice/chiggers, caterpillars, and millipedes. Ourwork fails to disprove his hypothesis. Honduran Spanishlabels bees, wasps, flies, caterpillars, and most of theothers on Berlin’s list (table 3). Unlike Standard Spanish,Rural Honduran Spanish has no single term for ant orbeetle (escarabajo refers only to some of the larger spe-cies). Weevils, fleas, lice, and chiggers are probably fa-miliar to people more for their cultural importance aspests than for their morphology. To Berlin’s list of majormorphotypes we would add earwigs (order Dermaptera)and grubs (larval Coleoptera).
Hymenoptera (bees, wasps, and ants) are some of thelarger, more colorful insects. Many are diurnal. Some ofthe nests of the social ones are larger than a person’s

Volume 42, Number 2, April 2001 F 289
table 3Honduran Folk Categories for Terrestrial Invertebrates (Unique Beginner Animales, Life Form Insectos) Classi-fied by Cultural Importance and Ease of Observation
Intermediate Generic Specific English Common Name Translation of Spanish Category
Babosa(Gastropoda: Veronicellidae)(Sarasinula plebeia and others)
Slug Slobberer Important, difficult
LipıMoclija(Gastropoda: Limacidae)
Slug Unanalyzable Important, difficult
RealilloReal(Diplopoda)
Millipede Little coina
Coina
Unimportant, easy
Ciempies(Chilopoda)
Centipede Hundred legs Unimportant, easy
Lombriz(Annellida)
Earthworm Earthworm Unimportant, easy
Arana(Araneae)
Spider Spider Unimportant, easy
Arana meacaballosPicacaballos(Theraphosidae)
Tarantula Horse pisserb
Horse stingerImportant, easy
Pendejo(Opilionnes)
Daddy longlegs Pubic hairc Unimportant, easy
Cazampulga(unidentified small spiders)
Flea hunter Unimportant, easy
Alacran(Scorpiones)
Scorpion Scorpion Important, easy
Coloradilla(Acari: Trombiculidae)
Chigger Little red one Important, difficult
Garrapata(Acari: Ixodidae)
Tick Leg grabber Important, easy
Garrapata chataDermacentor imitans
Thick (blood-filled) tick Important, easy
Garrapata menuditaColoraditaDermacentor imitans
Small tickSmall red tick
Important, easy
Mosca(Diptera, especially Muscidae)
Mosca Fly Fly Unimportant, easy
Queresa (eggs and larvae)(Calliphoridae)
Screwworm Unanalyzable Important, easy
Mosca de la queresa (adult)(Calliphoridae)
Screwworm Queresa fly Important, easy
Mosca tabano(Tabanidae)
Horse fly Horse fly Important, easy
Mosca lambesudorChupasudor(Syrphidae)
Syrphid fly Sweat lickerSweat sucker
Unimportant, easy
Mosca de la frutaespecially Ceratitis spp.
and Anastrepha spp.(Tephritidae)
Fruit fly Fruit fly Important, difficult
Zancudo(Culicidae)
Mosquito Long legs Important, easy
Mosquito(various small Diptera)
Gnat Little fly Important, difficult
Mosquito(various small Diptera)
Gnat Little fly Important, difficult
Avispa(Vespidae)
Wasp Wasp Important, easy
TurmaCampanillasCaucsirilCaushogoPolybia spp., usually P.
occidentalis
Scrotumc
Little bellc
UnanalyzableUnanalyzable
Important, easy
Turma de las largasPolybia diguetana
Scrotum, the long kindc Important, easy

290 F current anthropology
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Turma de las redondasPolybia occidentalis
Scrotum, the round kindc Important, easy
Turma de toroPolybia rejecta
Bull’s scrotumc Important, easy
CatalaChileraChilizatausually Polistes spp.
UnanalyzableChile-liked
Important, easy
Catala de las rojasPolistes major, P. instabilis,
and P. erythrocephalus
Red catala Important, easy
Catala de las negrasMischocyttarus spp.
Black catala Important, easy
AhorcadoraPolistes sp.
Stranglere Important, easy
JarritoPolybia emaciata
Little jarc Important, easy
Pico de chancoAlas blancasParachartergus apicalis
Pig snoutc
White wingsImportant, easy
ChirechanchoEpipona sp.
Pig snoutc Important, easy
CarniceroComecarneAgelaia cajennensis
Butcherf
Meat eaterf
Important, easy
QuitacalzonPapelilloProtopolybia acutiscutis
Underwear removerLittle paperc
Important, easy
Media lunaApoica thoracica
Half-moonc Important, easy
PupusaMetapolybia azteca
Stuffed tortilla Important, easy
PanalBrachygastra mellifica
Honeycombg Important, easy
GuitarronCorroncha de cuzucoPanza de burroSynoeca septentrionalis
Bass guitarh
Armadillo’s shellc
Donkey’s bellyc
Important, easy
CaseritaCasitas de tierra(Sphecidae)
Mud dauber Little housec Unimportant, easy
AvisponRey de aranasCazaranas(Pompilidae)
Big waspKing of spidersSpider hunter
Unimportant, easy
Avispa de la papayaToxotripana curvicauda(Diptera: Tephritidae)
Papaya fly Papaya waspi Important, difficult
Abeja(especially Apidae)
Bee Bee Important, easy
BlancoColmena grande
Bee HiveBig hive
Important, easy
Abeja blanco de castillaApis mellifera
European honeybee Castilian beej Important, easy
Abeja aluvaBlanco aluvaMelipona beecheii
Stingless bee Aluva (unanalyzable) beeAluva hive
Important, easy
Abeja moraBlanco moroMelipona fasciata
Moro beek
Moro hivek
Important, easy
Morroco Stingless bee Unanalyzable Important, easyMorrocoTrigona amaltheaTrigona nigerrima
Unanalyzable Important, easy
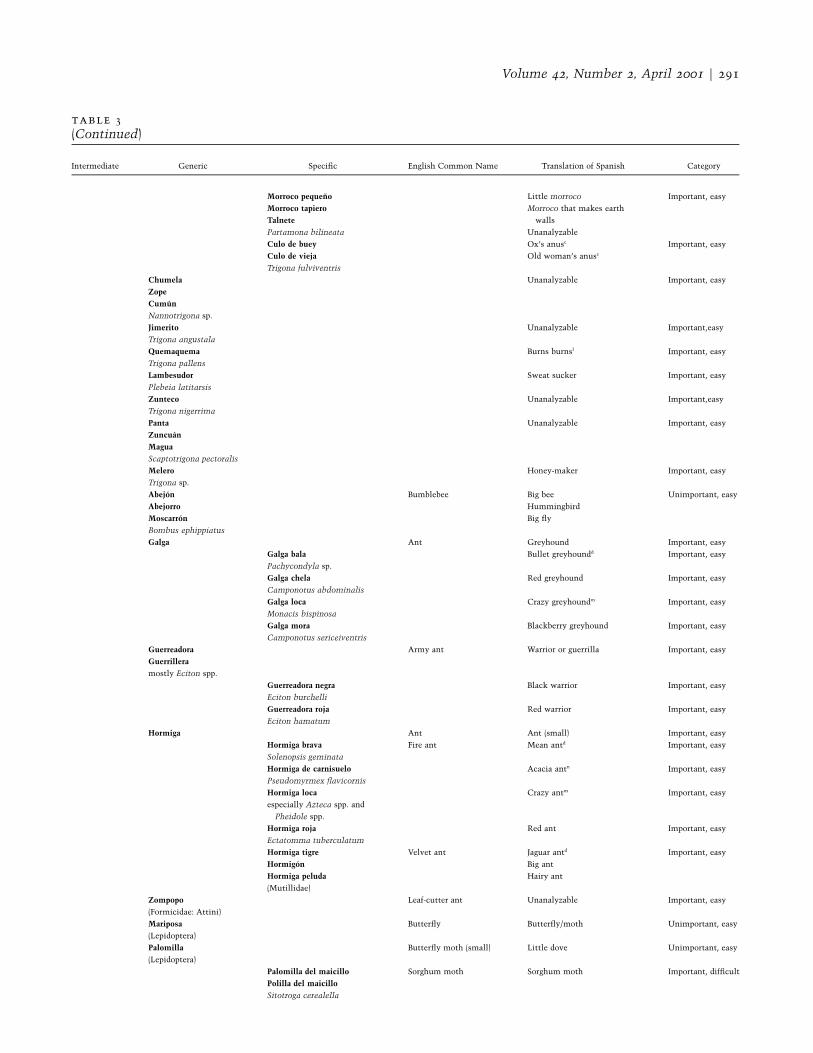
Volume 42, Number 2, April 2001 F 291
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Morroco pequenoMorroco tapieroTalnetePartamona bilineata
Little morrocoMorroco that makes earth
wallsUnanalyzable
Important, easy
Culo de bueyCulo de viejaTrigona fulviventris
Ox’s anusc
Old woman’s anusc
Important, easy
ChumelaZopeCumunNannotrigona sp.
Unanalyzable Important, easy
JimeritoTrigona angustala
Unanalyzable Important,easy
QuemaquemaTrigona pallens
Burns burnsl Important, easy
LambesudorPlebeia latitarsis
Sweat sucker Important, easy
ZuntecoTrigona nigerrima
Unanalyzable Important,easy
PantaZuncuanMaguaScaptotrigona pectoralis
Unanalyzable Important, easy
MeleroTrigona sp.
Honey-maker Important, easy
AbejonAbejorroMoscarronBombus ephippiatus
Bumblebee Big beeHummingbirdBig fly
Unimportant, easy
Galga Ant Greyhound Important, easyGalga balaPachycondyla sp.
Bullet greyhoundd Important, easy
Galga chelaCamponotus abdominalis
Red greyhound Important, easy
Galga locaMonacis bispinosa
Crazy greyhoundm Important, easy
Galga moraCamponotus sericeiventris
Blackberry greyhound Important, easy
GuerreadoraGuerrilleramostly Eciton spp.
Army ant Warrior or guerrilla Important, easy
Guerreadora negraEciton burchelli
Black warrior Important, easy
Guerreadora rojaEciton hamatum
Red warrior Important, easy
Hormiga Ant Ant (small) Important, easyHormiga bravaSolenopsis geminata
Fire ant Mean antd Important, easy
Hormiga de carnisueloPseudomyrmex flavicornis
Acacia antn Important, easy
Hormiga locaespecially Azteca spp. and
Pheidole spp.
Crazy antm Important, easy
Hormiga rojaEctatomma tuberculatum
Red ant Important, easy
Hormiga tigreHormigonHormiga peluda(Mutillidae)
Velvet ant Jaguar antd
Big antHairy ant
Important, easy
Zompopo(Formicidae: Attini)
Leaf-cutter ant Unanalyzable Important, easy
Mariposa(Lepidoptera)
Butterfly Butterfly/moth Unimportant, easy
Palomilla(Lepidoptera)
Butterfly moth (small) Little dove Unimportant, easy
Palomilla del maicilloPolilla del maicilloSitotroga cerealella
Sorghum moth Sorghum moth Important, difficult

292 F current anthropology
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Gusano(larvae of several insects, es-
pecially Lepidoptera)
Worm Wormo Unimportant, easy
Gusano peludo(several hairy larvae)(Arctiidae)
Hairy worm Important, easy
Gusano doradoEstigmene acrea(Arctiidae)
Golden worm Important, easy
Gusano quemador(Arctiidae)
Burning wormd Important, easy
Gusano cogollerolarvae of Spodoptera frugiperda(Noctuidae)
Fall armyworm Whorl wormp Important, difficult
Gusano medidorlarvae of Mocis latipes(Noctuidae)
Grasslooper Measurer Important, difficult
Falso medidorlarvae of Trichoplusia ni and
Pseudoplusia includens(Noctuidae)
False grasslooper False measurer Important, difficult
Gusano eloterolarvae that eat corn, e.g., Heli-
coverpa zea(Noctuidae)
Corn seed worm Corn ear worm Important, difficult
Gusano cortadorlarvae that cut the cornstalk,
e.g., Agrotis spp.
Cutworm Cutter Important, difficult
Gusano cuerudocutworms, e.g., Spodoptera sunia
Armyworm Leathery worm Important, difficult
Gusano cachudoGusano corronchudoespecially larvae of Manduca
sexta(Sphingidae)
Horned worm Horned wormThick, leathery worm
Important, difficult
Gusano barrenadorlarvae of Diatraea lineolata(Pyralidae)
Drillerq Important, difficult
Gusano barrenador de canalarvae of Diatraea saccharalis(Pyralidae)
Cane driller Important, difficult
Gusano de . . .(larvae of various Lepidoptera)
Worm of . . .
Gusano del pepinolarvae of Diaphania nitidalis(Pyralidae)
Cucumber worm Important, difficult
Gusano del melonlarvae of Diaphania hyalinata(Pyralidae)
Cantaloupe worm Important, difficult
Gusano del repollolarvae of Ascia monuste and
Leptophobia aripa(Pieridae)
Cabbage worm Important, difficult
RasquinaGusano del repolloPlutella xylostella
Scratcherr
Cabbage wormImportant, difficult
Langosta(larvae of Noctuidae, e.g.,
Mocis latipes)
Locust Important, difficult
Coralillo(larvae of Elasmopalpus lig-
nosellus)(Pyralidae)
Little coral snakes Important, easy
Coyota Female coyote Important, easy

Volume 42, Number 2, April 2001 F 293
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Torsalo(larvae of Dermatobia hominis)(Cuterebridae)
Botfly Unanalyzable Important, difficult
Clavito(larvae of Culicidae)
Mosquito larvae Little nails Unimportant, easy
Gallina ciega(larvae of Scarabaeidae, espe-
cially Phyllophaga spp.)
White grub Blind chicken Important, difficult
Gusano alambre(larvae of Coleoptera, Elateridae)
Wire wormt Important, difficult
Cucaracha de agua(Hydrophilidae)
Water cockroachs Unimportant, easy
RonronEspecially Phyllophaga spp.(Scarabaeidae Subf:
Melolonthinae)
(Onomatopoeic)h Unimportant, easy
Rueda mojonMierdero(Scarabaeidae Subf:
Scarabaeinae)
Dung beetle Turd rollerShitter
Unimportant, easy
Escarabajo(larger beetles of several
families)
Beetle Unimportant, easy
TronadorTrastras(Elateridae)
Click beetle Crackerh
(Onomatopoeic)hUnimportant, easy
CarapachoCarapachitoMegascelis spp. (Chrysomelidae)
and Eutheola spp.(Scarabaeidae)
Carapaces
Little carapaces
Unimportant, easy
BurroCacheton(Meloidae, Cantharidae,
Cerambycidae)
DonkeyBig cheeks
Unimportant, easy
Trozapalo(Passalidae)
Log breakeru Unimportant, easy
CandelillaLuciernaga(Lampyridae)
Firefly Little candleLight maker
Unimportant, easy
CamaleonTaladro(Buprestidae)
ChameleonDrillv
Unimportant, easy
Gorgojo(especially Curculionidae and
Bostrichidae)
Weevil Weevil Important, difficult
Picudo(especially Curculionidae)
Weevil Big snouts Important, difficult
TortuguillaMallaPulgon(Chrysomelidae, especially Dia-
brotica spp.)
Leaf beetle Little turtles
Meshw
Big fleas
Important, easy
Pulga(Siphonaptera)
Flea Flea Important, difficult
NiguaTunga penetrans(Siphonaptera: Tungidae)
Chigoe flea Chigoe flea Important, difficult
Ladilla(Pthirus pubis)(Phthiraptera: Pthiridae)
Crab louse Unanalyzable Important, difficult
Piojo(Pediculus humanus)(Phthiraptera: Pediculidae)
Louse Louse Important, difficult
Cusuquito(larvae of Myrmeleontidae)(Neuroptera)
Ant lion Little armadillos Important, easy
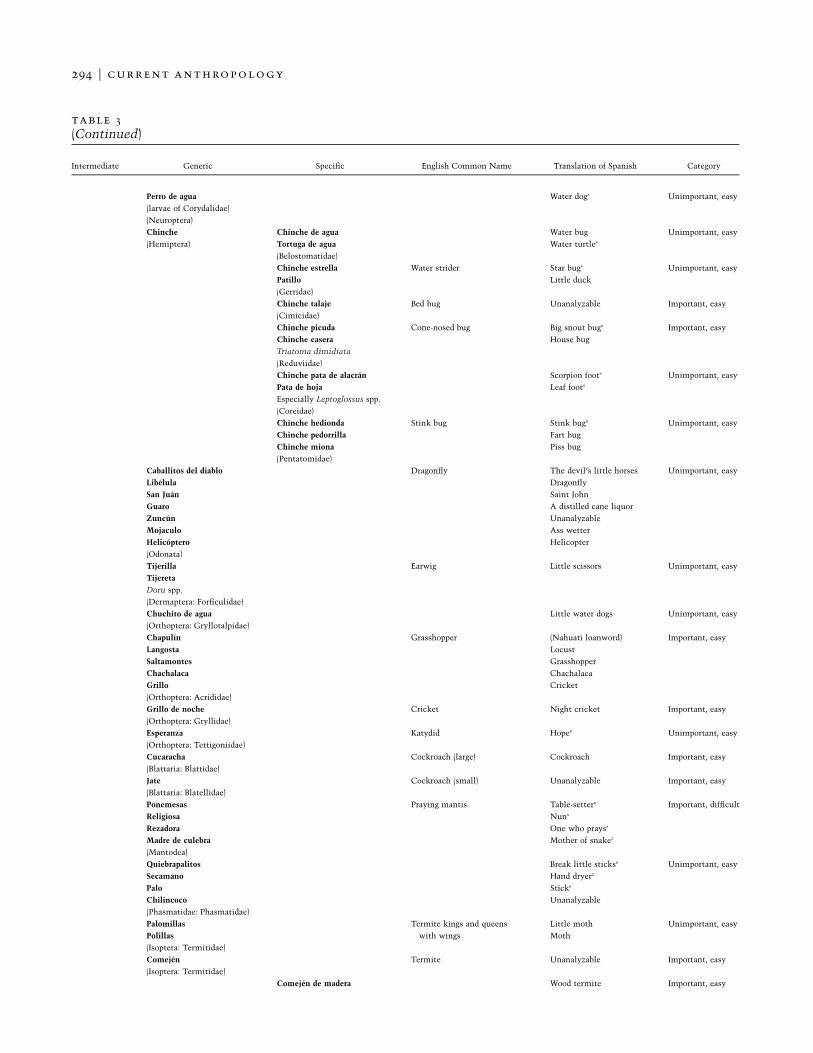
294 F current anthropology
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Perro de agua(larvae of Corydalidae)(Neuroptera)
Water dogs Unimportant, easy
Chinche(Hemiptera)
Chinche de aguaTortuga de agua(Belostomatidae)
Water bugWater turtles
Unimportant, easy
Chinche estrellaPatillo(Gerridae)
Water strider Star bugs
Little duckUnimportant, easy
Chinche talaje(Cimicidae)
Bed bug Unanalyzable Important, easy
Chinche picudaChinche caseraTriatoma dimidiata(Reduviidae)
Cone-nosed bug Big snout bugs
House bugImportant, easy
Chinche pata de alacranPata de hojaEspecially Leptoglossus spp.(Coreidae)
Scorpion foots
Leaf foots
Unimportant, easy
Chinche hediondaChinche pedorrillaChinche miona(Pentatomidae)
Stink bug Stink bugx
Fart bugPiss bug
Unimportant, easy
Caballitos del diabloLibelulaSan JuanGuaroZuncunMojaculoHelicoptero(Odonata)
Dragonfly The devil’s little horsesDragonflySaint JohnA distilled cane liquorUnanalyzableAss wetterHelicopter
Unimportant, easy
TijerillaTijeretaDoru spp.(Dermaptera: Forficulidae)
Earwig Little scissors Unimportant, easy
Chuchito de agua(Orthoptera: Gryllotalpidae)
Little water dogs Unimportant, easy
ChapulınLangostaSaltamontesChachalacaGrillo(Orthoptera: Acrididae)
Grasshopper (Nahuati loanword)LocustGrasshopperChachalacaCricket
Important, easy
Grillo de noche(Orthoptera: Gryllidae)
Cricket Night cricket Important, easy
Esperanza(Orthoptera: Tettigoniidae)
Katydid Hopey Unimportant, easy
Cucaracha(Blattaria: Blattidae)
Cockroach (large) Cockroach Important, easy
Jate(Blattaria: Blatellidae)
Cockroach (small) Unanalyzable Important, easy
PonemesasReligiosaRezadoraMadre de culebra(Mantodea)
Praying mantis Table-setters
Nuns
One who prayss
Mother of snakez
Important, difficult
QuiebrapalitosSecamanoPaloChilincoco(Phasmatidae: Phasmatidae)
Break little stickss
Hand dryerz
Sticks
Unanalyzable
Unimportant, easy
PalomillasPolillas(Isoptera: Termitidae)
Termite kings and queenswith wings
Little mothMoth
Unimportant, easy
Comejen(Isoptera: Termitidae)
Termite Unanalyzable Important, easy
Comejen de madera Wood termite Important, easy

Volume 42, Number 2, April 2001 F 295
table 3(Continued)
Intermediate Generic Specific English Common Name Translation of Spanish Category
Comejen de tierra Earth termite Important, easyComejen de pelota(Nasutitermes spp.)
Ball termite Important, easy
Piojillo(Thysanoptera)
Thrip Little louses Important, difficult
ChicharraChiquirın(Homoptera: Cicadidae)
Cicada Cicada(Onomatopoetic)h
Unimportant, easy
Lomo de camelloTorito(Homoptera: Membracidae)
Camel’s humpLittle bull
Unimportant, easy
Espuma de sapoSapilloSalivazoEspumonPonchito(Homoptera: Cercopidae
immature)
Spittlebug Toad foamLittle toadGlob of salivaBig foamLittle punch
Unimportant, easy
Lorito verdeEspecially Empoasca krae-
meri(Homoptera: Cicadellidae)
Empoasca Little green parrots Important, difficult
PulgonPiojillo(Homoptera: Aphidiidae)
Aphid Big fleas
Little louses
Important, easy
Mosca blanca(Bemisia tabaci)(Homoptera: Aleyrodidae)
Whitefly White fly Important, difficult
aRolled up, looks like coin.bBelieved to urinate while plucking hair for nest from horse’s leg, causing horse to lose its hoof.cAllusion to shape of nest.dStings.eSting produces choking sensation.fEats carrion.gMakes honey.hAllusion to sound it makes.iMimics a wasp.jOriginally brought to Latin America from Spain.kMoro means “Moor” and mora means “blackberry,” but in this case moro is probably unanalyzable.lSecretes a burning liquid on attacker’s skin.mRuns around.nLives in symbiosis inside the thorns of the bullhorn acacia.oIn Spain oruga is the word for “caterpillar,” but in Latin America gusano is generally used for both “worm” and “caterpillar.”pLives in and eats maize whorls.qDrills into cornstalk.rScratches into flesh of cabbage.sAllusion to appearance.tCalqued from English by agronomists?uLives in fallen timber.vBores into trees.wMakes the leaves it eats look like mesh.xDefends itself with foul-smelling urine.yBrings good luck.zIs believed to deform hand of person who picks it up.
head and have unique shapes and colors. Hymenopteraare culturally important for their honey, their ediblebrood, and their defense strategy (stinging, biting, andblistering). Fifty-one (38%) of the folk names we recordedfor insectos are for Hymenoptera. The only two inter-mediate taxa in Honduran folk entomology are for Hy-menoptera: bee (abeja) and wasp (avispa).
We would expect a detailed folk taxonomy for insects
that people eat (see Conconi 1982, Dufour 1987, Posey1987, Moran 1991, Setz 1991). Honduran peasants eatsome social wasp brood and honey, and they have a com-plex classification for wasps, with many folk genera anda few specifics. Some of the social wasp folk genera arepolytypic, among them, catala (Polistes spp. and someMischocyttarus spp.) and turma (some Polybia spp.). TheJicaque of Honduras, formerly hunter-gatherers, eat wasp

296 F current anthropology
brood and classify wasps in nearly the same way as His-panic Hondurans (Oltrogge 1975). In contrast, the Bribri,forest horticulturists of Costa Rica, classify a fairly sim-ilar wasp fauna with binomial labels (Starr and Bozzoli1990). Most Honduran bee names are generics, but thereare three folk species in the genus abejas de blanco,“hive bees” (Apis mellifera and two Melipona spp.). Allthree species live in the forest and are also tended in thevillages. In the woods, the bees nest in hollow treebranches, which campesinos cut off and bring home tohang from their front porches, harvesting the honey reg-ularly, somewhat as described by Posey (1983) for theKayapo. The three “hive” (blanco) species are muchlarger than other bees. This folk genus is classified bysize, not by use, since at least two species of smaller beesare also brought home and protected but are not classi-fied as abejas de blanco. The folk genus morroco (severalsmaller Meliponinae bees) is also polytypic.
Campesinos say, “Wasps sting and don’t make honey.Bees don’t sting and do give honey.” However, the (avis-pa) panal, “honeycomb (wasp)” (Brachygastra mellifica),is a honey-making vespid wasp, and the European hon-eybee, abeja de castilla (Apis mellifera) stings, unlikeother local bees. In spite of this ambiguity, Honduranfarmers classify the panal as a wasp and the honeybeeas a bee, as do entomologists.
Hondurans classify ants (Hymenoptera: Formicidae) infour folk genera—zompopos, guerreadoras, hormigas,and galgas—but have no word for “ant.” Zompopos (leaf-cutter ants—Formicidae: Attini, especially Atta spp. andAcromyrmex spp.) are not classified as ants. Few insectsare more conspicuous or perceptually salient. Large andred or black, they travel in long columns carrying cres-cent-shaped pieces of leaves like sails. Some of theirtrails through the tropical vegetation are as bare and wideas human paths. They are common, and some speciesare diurnal. The mounded entrances to mature coloniescover several square meters. They occasionally attackmaize or other crops and can strip an orange tree bareovernight.
Army ants (guerreadoras), “warriors” (especially Eci-ton burchelli and other Eciton spp.), are the next-most-salient ants; the colonies move constantly and can fieldseveral million workers each, fanning out in long col-umns and eating every small animal they catch (see Holl-dobler and Wilson 1990: chap. 16). The Honduran folkname is grammatically feminine, suggesting that it mayhave evolved from ∗ hormiga guerreadora. All other antsfall into two residual categories. Large ones are galgas(literally “greyhounds”). Small ones are hormigas (“ants”in Standard Spanish). There are several folk species ofgalga and hormiga. Stinging and nonstinging folk speciesare distinguished, for example, hormiga brava (Solen-opsis geminata) and galga bala (Pachycondyla sp.) areknown mainly for their bite and sting.
Several species of orange hairy caterpillars, gusano pe-ludo (family Arctiidae—especially Estigmene acrea—and Megalopygidae), are distinguished. The megalopygidspecies have urticating hairs that burn to the touch,while arctiid caterpillars are harmless, fuzzy Batesian
mimics. Many campesinos fail to distinguish the imi-tators from the burning caterpillars. This contrasts withlocal knowledge of bees, but in contrast to bees, none ofthe hairy caterpillars are useful and therefore they canall receive the same behavioral response (Hays 1982:92):avoidance.
Campesinos consider few other insects as importantas bees or wasps and classify them at the biological orderor family level. However, they may single out a few fam-ilies because of their harmfulness. Tabanidae (horse flies)are distinguished from other flies because of their bite(see Posey 1984:133).
Important, easily observed species may be named inorders which are otherwise not highly classified. Thereare few Honduran terms for the various true bugs (orderHemiptera). One of these is chinche picuda (especiallyTriatoma dimidiata [Hemiptera: Reduviidae]), a largered-and-black bug that lives in people’s houses and sucksblood from humans and other warm-blooded animals.Campesinos are becoming aware through public healthprograms that it transmits Chagas’ disease. The Spanishterm used in these programs, chinche (true bug), unfor-tunately leads to some confusion.
Honduran campesinos know some insects because oftheir role in children’s play. Dry sand patches are oftendimpled with the conical traps of ant lions (Neuroptera:Myrmeleontidae). The late Arnulfo Flores, a Honduranfarmer, showed us how to blow the sand out of the trapsand collect the fat, squirming insects and said, “We usedto play with them when we were kids.”
Culturally important, easily observed taxa are finelycategorized. Many of the terminal taxa are folk speciesthat coincide with Linnaean species. The only two in-termediate categories (bee, wasp) in Honduran folk en-tomology are culturally important and easily observed.This category could be used as an illustration for a par-adigmatic description of biological folk categories, withhierarchical taxonomic levels and some poly-typic folk genera divided into binomial folk species. Theother three categories could not.
Hondurans label the following invertebrates eventhough they have little if any local cultural significance:millipede, centipede, earthworm, spider, harvestman,hover fly, mosquito larvae (not recognized as the youngof mosquitoes), butterfly, small butterfly (palomilla), wa-ter scavenger beetle (Hydrophilidae), June beetle, largebeetle, click beetle, timber beetle, lightning bug, metallicwoodboring beetle (Buprestidae), larval dobsonfly, truebug, dragonfly, earwig, mole cricket, katydid, termite re-productives, cicada, treehopper, spittle bug, mud dauberwasp, tarantula wasp, and bumblebee. These categoriesare folk genera, but their organization is not very hier-archical. They are not subordinate to intermediate ranks,and they rarely have subordinate specific ranks. Thislong, flat taxonomy of generic categories lumps inver-tebrates at the biological order or family level, with theresult that each folk genus includes hundreds orthousands of biological species. The taxonomy of theeasily observed but culturally unimportant has little

Volume 42, Number 2, April 2001 F 297
table 4Taxonomic Properties of Each Category
CulturallyUnimportant
CulturallyImportant
Easy toobserve
Shallow, oftena long stringof genericterms. Organ-isms namedto the levelof Linnaeanorders orfamilies.
Deeper, hierar-chical (oftenincludingintermediateand specificlevels). Organ-isms frequentlynamed to thelevel of Linn-aean species.
Difficult toobserve
None Deep (e.g., withfolk species),but adult andjuvenile formsare not neces-sarily classifiedtogether andadults may evenbe lumped inlarge, residualcategories.Some stages ofsome organismsare labeled tothe level of theLinnaeanspecies.
structure. It could be represented as an index or a findinglist (Conklin 1969:107).
Campesinos classify some insects at the order level orlump several families together while singling out otherinsect families for names of their own. They notice can-delillas (lightning bugs—Coleoptera: Lampyridae) be-cause of their light and find the insects difficult to rec-ognize in the daytime. Click beetles (tronadores)(Coleoptera: Elateridae) have little cultural importanceas adults, but they are noticeable; they can snap a jointbetween two thoracic segments with enough force tohurl themselves into the air.
Small, cryptic arthropods that would otherwise escapeattention demand a name if they are pests of the humanbody such as ticks, chiggers, and lice.
Culturally important mimics are named but may bemisclassified from a modern scientific perspective. Cam-pesinos label the papaya “wasp” at the biological specieslevel; they know that the avispa de papaya causes wormsto appear in papaya fruit. Entomologists, in contrast,know it as a fly (Toxotrypana curvicauda [Diptera: Te-phritidae]) that, except for its long ovipositor (egg-layingorgan) and two wings (wasps have four), is an uncannymimic of a tiger-colored social wasp, Agelaia cajennensis(Hymenoptera: Vespidae).
Because they are difficult to observe, all grain-dwellingHonduran beetles (at least 25 species) are classified asgorgojos, “weevils” (Hoppe 1986), including true weevils(Curculionidae) and members of at least three other fam-ilies (Bostrichidae, Tenebrionidae, Cucujidae). Theyspend their first three life stages buried in stored food.Many farmers confuse the parasitic wasps of the weevilswith the weevils themselves. However, they classifyweevils as picudos if they feed on beans, chiles, and othercrops in the field (rather than in storage). Gorgojos andpicudos are contrasted ecologically (field versus storage),not by morphotype, because of their cultural importanceas pests.
Most Lepidoptera (butterflies and moths) are special-ized plant eaters in their larval (caterpillar) stage. Someare pests, and these are important and labeled at thebiological species level. Caterpillars that feed on wildplants are labeled by the residual term gusano. The in-sects themselves are often difficult to observe—small,colored to blend with the host plant, and buried in planttissue—but are noticed because of the attention thatfarmers pay to their crops. As Hunn (1982) observed inChiapas, Honduran campesinos label pest caterpillarsbut classify the adults as separate species. There aremany names for pest caterpillars, while the adults arelumped into larger, almost residual categories such asmariposa, “moth/butterfly.” Crop varieties are often bi-nomial folk species (Hunn and French 1984; Berlin 1992:24), and so are many crop pests.
Few anthropologists have discussed what local peopledo not label. It is easier to notice what is present thanto notice what is missing (Hearst 1991). However, cross-culturally, there are consistent gaps in local classifica-tions. Many organisms are too difficult to observe andtoo unimportant to be included at all in Honduran folk
entomology. Most parasitic wasps are solitary and toosmall to be seen easily, in spite of being among the mostnumerous insects on Earth (LaSalle and Gauld 1991). Ter-restrial nematodes tend to be microscopic and soil-dwell-ing. Insects of the order Collembola are small, flightless,and soil-dwelling; even some relatively large insects likegreen lacewings (Neuroptera: Chrysopidae) are notnamed even though they are occasionally seen. Greenlacewings are difficult to see because they are solitary,pale green, and nocturnal. Millions of species of mitesgo unobserved and unlabeled. The exception that provesthe rule is the coloradilla (chigger—Acari: Trombiculi-dae), a larval mite that digs into human skin and causesan agonizing itch.
Some highly salient species are unnamed because theyare so scarce that they are rarely observed. We noticeda colony of wasps (Brachygastra smithi [Hymenoptera:Vespidae]) with an asymmetrical, lumpy nest envelope.The wasps stung us when we touched their tree. Weasked several campesinos about the species. They hadnever seen it before and recognized it as a new speciesbut had no name for it. The colony moved on within afew days, and in four years we never saw another.
In summary (see table 4), intermediate categories arefound only in the culturally important and easily ob-served group. Folk genera may be divided into species ifthey are culturally important, whether easy or difficultto observe. The easily observed but culturally unimpor-tant taxa have little hierarchical organization and cor-respond roughly to scientific orders and families. Folk

298 F current anthropology
table 5Semantics of Each Category
CulturallyUnimportant
CulturallyImportant
Easy toobserve
Most named fora natural char-acteristic
Named for anatural char-acteristic;fewer namedfor the rolethey playwhen inter-acting withhumans
Difficult toobserve
Not named Many namedfor their rolein humanculture; somenamed for anatural char-acteristic
classification of culturally important but difficult-to-ob-serve organisms may be inconsistent with modern sci-entific taxonomy, especially where a species mimics adistantly related one, where creatures are small, or wherepeople fail to associate larvae with their adults. The cul-turally unimportant and difficult to observe areunclassified.
semantics
Some plant and animal names are semantically opaque.Most other names are coined from either appearance orutility (or damage). As Berlin (1992:27) has observed, en-coding salient morphological and behavioral features inethnobiological names makes a large vocabulary easy tolearn and remember. In Honduras, most of the culturallyunimportant and easily observed invertebrates arenamed for their appearance and behavior, and a pluralityof the culturally important and difficult-to-observe crea-tures are named for their importance (e.g., the crops theyattack). Some culturally important and easily observedinvertebrates are named for their natural attributes andothers for their cultural roles, but some names in allthree of these categories are unanalyzable.
The culturally important and easily observed insectstend to be named for natural rather than cultural traits.Of 66 categories which we judged to belong to this group,34 (52%) were named for natural attributes. For example,the leaf beetle is tortuguilla (“little turtle”) because ithas a hard round shell. Most wasp and bee species arenamed after an object that the nest resembles: a pig’ssnout, an ox’s anus, or a bass guitar.
Seventeen (26%) of the culturally important and easilyobserved insects are named for their cultural roles. Forexample, the “underwear remover” (the social wasp Pro-topolybia acutiscutis) is named for the way it attackshumans, and so is a bee called quema quema (“burnyburny”) (Trigona pallens), which burns its victims witha toxic secretion. As we have seen, a wasp that makesedible honey is called panal, “honeycomb.” The taran-tula (Theraphosidae) is called (arana) meacaballo(“horse-pissing spider”) because campesinos insist thattarantulas urinate on a horse’s foot and make it lose itshoof. (Most agronomists deny the validity of this belief.)The comecarne (“meat eater”) wasp (also called carni-cero, “butcher”) (Agelaia cajennensis) feeds on carrionand sometimes annoys farmers butchering an animal.
Fifteen (23%) of the culturally important and easilyobserved insects have semantically opaque names, someof which are old Spanish words and a few of which areloans from Native American languages. The 29 catego-ries with semantically opaque names in Honduran folkentomology are spread fairly evenly and do not correlatewith either cultural importance or ease of observation.This result was unexpected. Balee (1989) notes that cul-tivated plants (which are culturally important and easilyobserved) are labeled by single-word, unanalyzablelexemes.
We expected that the easily observed but culturallyunimportant creatures would be named for natural at-
tributes. Of 41 insects in this category, 31 (76%) are sonamed. For example, daddy longlegs (harvestmen) (orderOpiliones) are called pendejos (“pubic hairs”) becausewhen they are huddled in a gregarious mass they looklike a clump of body hair. Dung beetles (Coleoptera: Scar-abaeidae: Scarabaeinae) are ruedamojon (“turd roller”);the lightning bug is candelilla (“little candle”). Eight(20%) have unanalyzable, semantically opaque names.Only two categories (5%) are named for the way in whichthe insects interact with humans. Hover flies are namedchupasudor, “sweat-sucker,” for their habit of lappingsweat from the arms of people at work.
We expected that culturally important but difficult-to-observe insects would be named for their interactionwith humans. Only 13 (38%) of the 34 categories in thisgroup are named for their cultural importance. Most ofthe caterpillar pest species are named for the fruit theyattack (e.g., gusano del melon [Diaphania hyalinata(Lepidoptera: Pyralidae)]) or the kind of damage they do(e.g., gusano cogollero, “whorl worm”). Another 13 ofthese insects are named for natural characteristics, againreflecting the overall Honduran bias toward naturalrather than cultural insect names. The babosa (slug) (sev-eral gastropod families, especially Veronicellidae) isnamed for the trail of slime it leaves. Picudos (field wee-vils) are named for their snouts. Gusano cachudo (severalhorned caterpillars of the family Sphingidae, especiallyManduca sexta) is named for its horn. It seems paradox-ical that any difficult-to-observe insects could be namedfor their physical characteristics, but while difficult toobserve they are not invisible. Six (18%) of the culturallyimportant and difficult-to-observe insects have seman-tically opaque names.
In summary (see table 5), 80 (57%) of all categories arenamed for natural attributes. This lends modest supportto the universalist hypothesis, and, as would be expected,the tendency is especially strong for the easily observedbut culturally unimportant. Some names, especially forthe culturally important insects, reflect the creatures’

Volume 42, Number 2, April 2001 F 299
interaction with humans. Thirty-two (23%) of all cate-gories are named for their interaction with humans,which lends a little support to the utilitarian hypothesis.Almost all of those names (30, 94%) are for culturallyimportant taxa. The 29 (21%) categories labeled withunanalyzable names are evenly spread through the wholelexical set. Binomial folk species generally label cultur-ally important insects, whether easy or difficult to ob-serve. The names for pests are highly productive (tomatoworm, melon worm, etc.).
discussion
We began by asking whether people discover their worldor create it—that is, whether they know and name livingthings for their natural (universal) qualities or for theirculturally specific roles in human life. On semantic ev-idence, the rural people of Central Honduras pay moreattention to natural attributes but name a substantialminority of creatures for their roles in human cul-ture—suggesting that the people both discover and createtheir world. Honduran campesinos discover (and label)nature’s major morphotypes, the biologists’ orders andfamilies—the ants and the butterflies “crying out fornames.” This supports the universalist argument. Theydiscriminate finer categories according to local culturalpriorities of avoiding pain, playing, getting food and shel-ter, and managing pests. This supports the cultural rel-ativist argument, misnamed “utilitarian” in that culturedeals with creatures as much for their nuisance value asfor their utility. When a culture classifies the creaturesthat nature camouflages, some species are confused withunrelated ones; some relationships between adults andoffspring are misunderstood. Culture ignores the micro-scopic species and others that nature hides.
We propose that people first discover their world. Asthe universalist argument suggests, they notice andname the great categories of natural things that cry outfor labels. At least with insects, they name major mor-photypes (dragonflies, for example) just because thoseorganisms are so perceptually salient, even when theyare perceived to have no utility or harm value for hu-mans. However, as people make a living, they create orat least modify their world. They notice more subtledetails of coloring, habitat, locomotion, etc., to distin-guish pests from nonpests, food from the inedible, thesafe from the dangerous (consistent with the utilitarianargument). Traditional rural people label the insect worldalong universalist lines about to the level of the Linnaeanorder or family but generally label entomofauna at the(formal, biological) genus or species level only when nec-essary for utilitarian reasons. In other words, as the univ-ersalist perspective suggests, nature provides people withthe basic framework for biological taxonomies, thenames for living things and the folk knowledge of them.However, people elaborate on that basic system in cul-turally specific ways to make a living, to play, to avoidpain, and occasionally to meet spiritual and other cul-turally mediated needs.
Given the limits of unaided human observation, the
millions of Earth’s species, and other demands on peo-ple’s attention, traditional peoples cannot label all in-vertebrates. However, ethnoentomology has ample cat-egories for discussing work and play and for wonderingabout living things. Folk classification of terrestrial in-vertebrates is reasonably comprehensive and usuallyconsistent with formal, scientific entomology. Tradi-tional rural people know insects more intimately thananyone except entomologists, but few entomologistsknow how to harvest wasp honey or are aware that leaf-cutter ants host lizard lodgers.
We hypothesize that cross-culturally, fish, mammals,birds, trees, weeds, and crops will also be associated withdeep knowledge and deep taxonomies and will be namedfor a mix of natural and cultural properties (because thesetaxa are culturally important, usually, and easily ob-served). Disease organisms and most pests of crops, live-stock, and the human body will be associated with grittyknowledge and taxonomies that are somewhat stratifiedand will often be named for their utility (harm) value(because they are culturally important but difficult toobserve). Larger insects, large but inedible fungi, and use-less but harmless herbaceous plants will be associatedwith thin knowledge and flat taxonomies in categoriesformed at high Linnaean levels, and their names willdescribe their appearance (because they are culturallyunimportant but easy to observe). The very small, rare,cryptic beings, such as most nematodes, bacteria, andmicroscopic fungi, will be associated with no localknowledge, no taxonomies, and no names (because theyare difficult to observe and culturally unimportant).
References Citeda r i o t i , m a r i a . 1985. Edible animals of the Ituri Forest, Af-
rica, in the ethnozoology of the Efe Bambuti. Journal of Ethno-biology 5:21–28.
a t r a n , s c o t t . 1985. The nature of folk-botanical life forms.American Anthropologist 87:298–315.
———. 1987. The essence of folkbiology: A reply to Randall andHunn. American Anthropologist 89:149–51.
b a l e e , w. 1989. Nomenclatural patterns in Ka’apor ethnobo-tany. Journal of Ethnobiology 9:1–24.
b e n t l e y, j e f f e ry w. 1990. Conocimiento y experimentos es-pontaneos de campesinos hondurenos sobre el maız muerto.Manejo Integrado de Plagas 17(September):16–26.
———. 1991. ¿Que es hielo? Percepciones de los campesinoshondurenos sobre enfermedades del frijol y otros cultivos. In-terciencia 16:131–37.
———. 1992a. Learning about biological pest control. ILEIANewsletter 8:16–17.
———. 1992b. “The epistemology of plant protection: Hondurancampesino knowledge of pests and natural enemies,” in Pro-ceedings of a seminar on crop protection for resource-poorfarmers. Edited by R. W. Gibson and A. Sweetmore. Chatham,U.K.: Technical Centre for Agricultural and Rural Co-operationand Natural Resources Institute.
———. 1993. What farmers don’t know. Ceres 26(141):42–45.———. 1994. “Stimulating farmer experiments in non-chemical
pest control in Central America,” in Beyond Farmer First: Ru-ral people’s knowledge, agricultural research, and extensionpractice. Edited by I. Scoones and J. Thompson, pp. 147–50.London: Intermediate Technology Publications.
b e n t l e y, j e f f e ry w. , g o n z a l o ro d r ı g u e z , a n d a n a

300 F current anthropology
g o n z a l e z . 1994. Science and people: Honduran campesinosand natural pest control in Honduras. Agriculture and HumanValues 11:178–82.
b e r l i n , b r e n t . 1990. “The chicken and the egg-head revis-ited: Further evidence for the intellectualist bases of ethnobiol-ogical classification,” in Ethnobiology: Implications and appli-cations (Proceedings of the First International Congress ofEthnobiology [Belem, 1988]), vol. 1. Edited by D. A. Posey, pp.19–33. Belem: Museu Paraense Emılio Goeldi.
———. 1992. Ethnobiological classification: Principles of catego-rization of plants and animals in traditional societies. Prince-ton: Princeton University Press.
b o s t e r , j a m e s . 1987. Agreement between biological classifi-cation systems is not dependent on cultural transmission.American Anthropologist 89:914–20.
b o s t e r , j a m e s , b r e n t b e r l i n , a n d j o h n o ’ n e i l l .1986. The correspondence of Jivaroan to scientific ornithology.American Anthropologist 88:569–83.
b ro w n , c e c i l h . 1984. Language and living things: Uniform-ities in folk classification and naming. New Brunswick: Rut-gers University Press.
———. 1992. British names for American birds. Journal of Lin-guistic Anthropology 2:30–50.
c h a m b e r s , ro b e r t . 1983. Rural development: Putting thelast first. New York: Wiley.
c h i t t a m p a l l i , p u d m a , a n d f . d a v i d m u l c a h y. 1990.“Honey and sugar in the treatment of wounds and ulcers inbiomedicine and in the Ayurveda,” in Ethnobiology: Implica-tions and applications (Proceedings of the First InternationalCongress of Ethnobiology [Belem, 1988]), vol. 2. Edited by D.A. Posey, pp. 155–64. Belem: Museu Paraense Emılio Goeldi.
c o n c o n i , j u l i e t a r a m o s e l o r d u r d e . 1982. Los insec-tos como fuente de proteınas en el futuro. Mexico, D.F.: Edito-rial Limusa.
c o n k l i n , h a ro l d c . 1962. Lexicographical treatment of folktaxonomies. International Journal of American Linguistics 28,pt. 2(4):119–41.
———. 1969. “Ethnogenealogical method,” in Cognitive anthro-pology. Edited by Stephen A. Tyler, pp. 93–122. New York:Holt, Rinehart and Winston.
d i m i c h e l e , w i l l i a m a . , a n d ro b e r t w. h o o k , w i t hr i c h a r d b e e r b o w e r , j u r g e n a . b o y, ro b e r t a .g a s t a l d o , n i c h o l a s h o t t o n i i i , t o m l . p h i l l i p s ,s t e p h e n e . s c h e c k l e r , w i l l i a m a . s h e a r , a n dh a n s - d i e t e r s u e s . 1992. “Paleozoic terrestrial ecosys-tems,” in Terrestrial ecosystems through time. Edited by A. K.Behrensmeyer, J. D. Damuth, W. A. Di Michele, R. Potts, H.D. Sues, and S. L. Wing, pp. 204–325. Chicago: University ofChicago Press.
d u f o u r , d a r n a l . 1987. Insects as food: A case study fromthe Northwest Amazon. American Anthropologist 89:383–97.
e l l e n , ro y f . 1986. Ethnobiology, cognition, and the structureof prehension: Some general theoretical notes. Journal of Eth-nobiology 6:83–98.
———. 1993. The cultural relations of classification: An analysisof Nuaulu animal categories from Central Seram. Cambridge:Cambridge University Press.
e r w i n , t e r ry l . 1988. “The tropical forest canopy: The heartof biotic diversity,” in Biodiversity. Edited by E. O. Wilson, pp.123–29. Washington, D.C.: National Academy Press.
g a d a g k a r , r a g h a v e n d r a . 1991. “Belongaster, Mischocyt-tarus, Parapolybia, and independent-founding Ropalidia,” inThe social biology of wasps. Edited by K. G. Ross and R. W.Matthews, pp. 149–91. Ithaca: Cornell University Press.
g o n z a l e z , a n a . 1993. Elaboracion y evaluacion de cursos decontrol biologico para agricultores y extensionistas. Thesis, Es-cuela Agrıcola Panamericana, Zamorano, Honduras.
h a y s , t e r e n c e e . 1982. Utilitarian/adaptationist explanationsof folk biological classification: Some cautionary notes. Journalof Ethnobiology 2:89–94.
———. 1983. Ndumba folk biology and general principles of eth-nobiological classification and nomenclature. American An-thropologist 85:592–611.
h e a r s t , e l i o t . 1991. Psychology and nothing. American Sci-entist 79:432–43.
h o l l d o b l e r , b e r t , a n d e d w a r d o . w i l s o n . 1990. Theants. Cambridge: Belknap Press.
h o p p e , t . 1986. Storage insects of basic food grains in Hondu-ras. Tropical Science 26:25–38.
h u n n , e u g e n e s . 1977. Tzeltal folk zoology: The classifica-tion of discontinuities in nature. New York: Academic Press.
———. 1982. The utilitarian factor in folk biological classifica-tion. American Anthropologist 84:830–47.
———. 1990. Nch’i-Wana, “The Big River”: Mid-Columbia Indi-ans and their land. Seattle: University of Washington Press.
h u n n , e u g e n e , a n d d a v i d h . f r e n c h . 1984. Alterna-tives to taxonomic hierarchy: The Sahaptin case. Journal ofEthnobiology 4:73–92.
h u n t , j a m e s h . 1991. “Nourishment and the evolution of thesocial Vespidae,” in The social biology of wasps. Edited by K.G. Ross and R. W. Matthews, pp. 426–50. Ithaca: Cornell Uni-versity Press.
j e a n n e , ro b e r t l . 1991. “The swarm-founding Polistinae,”in The social biology of wasps. Edited by K. G. Ross and R. W.Matthews, pp. 191–231. Ithaca: Cornell University Press.
l a s a l l e , j . , a n d i . d . g a u l d . 1991. “Parasitic Hymenop-tera and the biodiversity crisis,” in Insect parasitoids: 4th Eu-ropean workshop—Perugia 3–5 April, 1991, Appendice, pp.315–34. REDIA 74.
l a s t , m u r r a y. 1981. The importance of knowing about notknowing. Social Science and Medicine 15B:387–92.
m o r a n , e m i l i o f . 1991. Human adaptive strategies in Ama-zonian blackwater ecosystems. American Anthropologist 93:361–82.
n e t t i n g , ro b e r t m c c . 1993. Smallholders, householders:Farm families and the ecology of intensive, sustainable agri-culture. Stanford: Stanford University Press.
o l t ro g g e , d . f . 1975. “La etnoentomologıa de algunas cate-gorıas de la orden Himenoptera entre los jicaques.” Ponenciasde los miembros del Instituto Linguıstico de Verano, A.C. enla XIV Reunion de Mesa Redonda de la Sociedad Mexicana deAntropologıa y la I Reunion de Antropologos e Historiadoresde Centroamerica y Mexico, 22 al 28 de junio. Tegucigalpa:Summer Institute of Linguistics.
p o s e y, d a r r e l l a . 1983. Keeping of stingless bees by theKayapo Indians of Brazil. Journal of Ethnobiology 3:63–73.
———. 1984. Hierarchy and utility in a folk biological taxonomicsystem: Patterns in classification of arthropods by the KayapoIndians of Brazil. Journal of Ethnobiology 4:123–39.
———. 1987. Ethnoentomological survey of Brazilian Indians. En-tomologia Generalis 12:191–202.
p o s e y, d a r r e l l a . , a n d j o a o m a r i a f r a n c o d e c a r -m a g o . 1985. Additional notes on the classification andknowledge of stingless bees (Meliponinae, Apidae, Hymenop-tera) by the Kayapo Indians of Gorotire, Para, Brazil. Annals ofCarnegie Museum 54:247–74.
r e e v e , h u d s o n k . 1991. “Polistes,” in The social biology ofwasps. Edited by K. G. Ross and R. W. Matthews, pp. 99–148.Ithaca: Cornell University Press.
s e t z , e l e o n o r e z u l n a r a f r e i r e . 1991. Animals in theNambiquara diet: Methods of collection and processing. Jour-nal of Ethnobiology 11:1–22.
s i l l i t o e , p a u l . 1996. A place against time: Land and envi-ronment in the Papua New Guinea Highlands. London: Har-wood Academic Publishers.
s t a r r , c h r i s t o p h e r k . , a n d m a r i a e u g e n i a b o z -z o l i d e w i l l e . 1990. “Social wasps among the Bribri ofCosta Rica,” in Ethnobiology: Implications and applications(Proceedings of the First International Congress of Ethnobiol-ogy [Belem, 1988]), vol. 1. Edited by D. A. Posey, pp. 187–94.Belem: Museu Paraense Emılio Goeldi.
v a y d a , a n d r e w p . , a n d i n d a h s e t y a w a t i . 1995.“Questions about culture-related considerations in research oncognition and agro-ecological change: Illustrations from studiesof agricultural pest management in Java,” in Cultural dynam-

Volume 42, Number 2, April 2001 F 301
ics in development processes. Edited by A. de Ruijter and L.van Vucht Tijssen, pp. 259–78. Amsterdam: UNESCO.
w i l k , r i c h a r d r . 1991. Household ecology: Economicchange and domestic life among the Kekchi Maya in Belize.Tucson: University of Arizona Press.
w i l k e n , g e n e c . 1987. Good farmers: Traditional agricul-tural resource management in Mexico and Central America.Berkeley: University of California Press.
w i l s o n , e d w a r d o . 1988. “The current state of biologicaldiversity,” in Biodiversity. Edited by E. O. Wilson, pp. 3–18.Washington, D.C.: National Academy Press.
———. 1992. The diversity of life. Cambridge: Belknap Press.w i n a r t o , y u n i t a t r i w a r d a n i . 1996. Seeds of knowl-
edge: The consequences of integrated pest management school-ing on a rice farming community in West Java. Ph.D. diss.,Australian National University, Canberra, Australia.
Assessing the Biological Status ofHuman Populations1
napoleon wolansk i and anna siniarskaDepartment of Human Ecology, Center for ScientificResearch and Postgraduate Studies CINVESTAV,Merida, Yucatan, AP 73 Cordemex, CP 97310, Mexico/Department of Human Ecology, Institute of Ecology ofthe Polish Academy of Sciences, ul. Konopnickiej 1,Lomianki, Poland ([email protected]). 9 x 00
The biological status of human populations is an im-portant object of research on human evolution and ad-aptation to contemporary living conditions and on prac-tical applications related to its role as a mirror ofsocioeconomic transformations. Every human biologicaltrait reflects living conditions, but the phenotype is theproduct of all the traits taken together. Moreover, cul-tural adaptation modifies and replaces biological adap-tations and must therefore also be taken into consider-ation. The biological status of a population describes itspotential for health in terms of both negative and posi-tive indices. Body height and weight are usually takenas positive indices of health, especially in childhood. Thebiological status of an adult is the result of growth anddevelopment. From this the health of the environmentis also assessed. Biological status, health status, nutri-tional status, and reproductive fitness are interrelatedbut have independent meanings and values. The lastthree of these are also related to certain psychologicaland social problems. In our discussion the term “biolog-
� 2001 by The Wenner-Gren Foundation for Anthropological Re-search. All rights reserved 0011-3204/2001/4202-0006 $1.00.
1. This work was supported by Mexican CONACyT grants nos.1324-1325-S9206 and 26469H. We are grateful for the editorial as-sistance of Francisco Gurri and the important remarks of an anon-ymous referee.
ical status” is in general broader than the other threeterms.
“Adaptation” is here understood as the structural andfunctional characteristics of individuals that enhancetheir survival and reproduction and enable them to copewith their environment. Adaptation may be genetic orcultural. “Adaptive changes” are understood as a patternof adaptation and/or adjustment to the environment, bi-ological and/or cultural. Any change in environmentalconditions causes adaptive changes in human popula-tions. In contemporary populations these changes areusually assessed in terms of a synthetic biologicalmarker, stature. Although such comparisons are basedon several traits, each trait is considered separately (Wo-lanski 1966, 1990). Indices relating one trait to anotherare sometimes used to eliminate differences resultingfrom variation in stature or weight, but there is a lackof such relations between traits involved in the samephysiological process. Some progress in this direction hasbeen provided by factor analysis, which creates a smallernumber of noncorrelated factors or components. Unfor-tunately, these nonmeasurable factors or components aredifficult to identify and interpret.
Studies of populations living under different social andeconomic conditions and at various cultural levels haverevealed that it is impossible to distinguish them interms of any single trait. For example, if individuals rep-resenting a certain population are tall, their nutritionaland health conditions can be expected to be good; how-ever, the same population may show low endurance fit-ness, considered a negative trait. People from one pop-ulation may display high muscular strength but lowpersistence fitness (stamina), whereas others have highpersistence fitness but low muscular strength. Many bi-ological traits—for example, the various respiratory, car-diovascular, and blood traits involved in transporting ox-ygen to the tissues—interact. If vital capacity alone wereexamined, the process of respiration would not be fullyunderstood. In the case of some environmental influ-ences, vital capacity, blood pressure, or hematocrit indexmay not show any changes while changes are apparentin ventilation and/or hemoglobin concentration and/orheart output. The question is which population is bio-logically better off.
The main aim of this work is to show that the kindof evaluation just described is insufficient. Thus, theproblem becomes how to assess the biological status ofhuman populations as an indicator of health and how tointerpret variations in individual biological traits. Weshall present an attempt to elucidate this problem usinginvestigations conducted in different geographicalregions and including populations living under differentsocioeconomic conditions. We want to show how con-temporary human populations in a country that is eth-nically rather homogeneous adapt to their living con-ditions by presenting their phenotypic differences. Wesuggest that the criteria used for the assessment of bi-ological traits need to be revised; instead of attemptingto assign positive or negative values to traits we should

302 F current anthropology
Fig. 1. Greatest differences between Polish populations for certain somatic, physiological, and psychomotortraits. Arrows point to population in which the value of the traits in question is greatest or best. Abbreviations:accur., accuracy of movements; adip. tis., thickness of subcutaneous fat tissue; agil., agility; apnea, duration ofapnea; balance stat. (turn.), static (or turning) sense of equilibrium; chest circ., chest circumference; ch. mov.,chest movement range; dyn. str., dynamic strength; eryth., erythrocyte count; eu-eu, head breadth; expl. str.legs (h.), explosive power of lower (upper) extremities; FEV, forced expiratory volume per second; flexib., spineflexibility; g-op, head length; Hg, hemoglobin concentration; Ht, hematocrit index; IRV (ERV), inspiratory (expi-ratory) reserve volume; linear meas., various linear measures; leuc., leucocyte count; n-gn, total facial height;orien., spatial orientation; persist., persistence fitness (stamina); Pu rest, pulse rate at rest; Pu work, pulse rateat work; react. h. (leg), reaction time of hands (legs); respir. freq., frequency of respiration; speed, speed of move-ments; stat., stature; thl-thl, chest breadth; VC, vital capacity; Vmax p maximal minute ventilation; weight,body weight; xi-ths, chest depth; zy-zy, face breadth (Siniarska 1984).
consider the broad range of living conditions to whichthey are adapted.
materials and methods
The synthesis presented below is based on data we col-lected between 1971 and 1993 in Poland and Mexico andcomparisons with the data of other researchers. In 1959and 1979, 3,235 subjects (1,618 males) and 2,856 subjects(1,420 males), respectively, between 2 and 20 years of agewere studied in Warsaw (Wolanski 1962, Wolanski andLasota 1964, Kozioł and Wolanski 1982). In 1960 a sim-ilar study of 2,600 children and youths (1,305 males) ofthe same age was conducted in Polish villages in the areaof Kurpie and Suwałki (Wolanski and Lasota 1964, Wo-lanski 1973). Between 1963 and 1968, 3,119 subjects(1,570 males) between 5 and 17 years of age were studied
in various parts of Poland: rural areas (Suwałki and Kur-pie regions), coastal areas (the Hel Peninsula), an area oflow mountains (Pieniny Mountains), cities (Warsaw andKatowice), and a heavily industrialized region (Silesia)(Pyzuk and Wolanski 1972). In 1971, 297 subjects (156males) between 21 and 70 years of age were studied intwo villages in the Kurpie region, Jeglijowiec and Brzo-zowka (Szemik 1986). Between 1975 and 1978, 5,692 sub-jects (2,951 males) between 2 and 80 years of age werestudied in a rural area (the Suwałki region), in regionsundergoing initial industrialization (the Lublin coal ba-sin and the Bełchatow industrial center), and in a heavilyindustrialized region (Silesia) (Siniarska 1984, Siniarskaand Wolanski 1986). The last of these studies took placein 1993 in Mexico, where 642 subjects (321 males) be-tween 2 and 18 years of age were studied in Merida and

Volume 42, Number 2, April 2001 F 303
Fig. 2. Differences in respiratory-circulatory functions under various climatic and altitude conditions and con-ditions created by industrialization and urbanization (especially air pollution). Abbreviations: VC, vital capac-ity of lungs; V, lung ventilation at rest per minute; Ap, apnea, time without breathing; Hb, hemoglobin concen-tration; Hct, hematocrit index; BP, arterial blood pressure; HR, heart rate at rest; Q, minute heart volume atrest (cardiac output) (Pyzuk and Wolanski 1972, Kozioł and Wolanski 1982).
Progreso, Yucatan (Wolanski 1994). Descriptions of thestudy areas and their demographic characteristics and ofthe methods of measurement have appeared elsewhere(Pyzuk 1973; Wolanski, Siniarska, and Szemik 1982; Sin-iarska 1984, 1996; Wolanski 1994). Various morpholog-ical and physiological variables and some psychomotorcharacteristics were used in the summary paper (Siniar-ska 1996).
The illustrative material in this paper is based mostlyon males for two reasons. First, boys often demonstrategreater variability of response to environmental and cul-tural variables than girls, and therefore the influence ofmany environmental factors is more clearly seen. Sec-ond, the changes in females are of the same character,and therefore it seemed appropriate to save space by pre-senting our results using only one gender.
results
Trait values in various populations. Siniarska’s (1984)data on morphological, physiological, and psychomotortraits show that different populations have differentmaximal (or optimal) values of various traits. Of the pop-ulations under study (fig. 1) the tallest are those fromthe heavily industrialized area of Silesia. Possible reasonsfor this are higher incomes, greater consumption of foodsrich in animal protein, and a tendency toward greater
body size in polluted environments. People in this en-vironment show the lowest values for physical fitness,for which the highest values are found in one of the areasjust beginning industrialization, Bełchatow. This latterpopulation shows the lowest values for respiratory traits,for which the highest values are found in the rural ag-ricultural villages of Suwałki. The Suwałki populationalso shows the lowest blood pressure and resting respi-ration frequency but a high hematocrit index and max-imal ventilation. Both maximal ventilation and staturehave the lowest values in the rural population from an-other area in the initial phase of industrialization (theLublin coal basin), which also has a low working heartrate and a low hematocrit index.
Summing up the positive trait values in the popula-tions studied, (1) the rural agricultural population hasthe best respiratory traits and hematocrit index; (2) therural population under initial industrialization shows alow working heart rate and the greatest stamina; (3) thetown population under initial industrialization showslong apnea duration, high erythrocyte count, high speedof movement, great explosive power, short reaction time,and good sense of balance; and (4) the population fromthe heavily industrialized area shows great stature andbody weight, high hemoglobin concentration, and highspine flexibility. From these results it is impossible to

304 F current anthropology
table 1Results of Motor Fitness Tests in Polish Villages,Towns, and Industrializing Areas
Trait Villages Towns Industrializing Areas
Agility andcoordinationSpine flexibility 1 2 3Turning balance 2 1 3Static balance 1 2 3Movement memory 1 3 2Spatial orientation 1 – 3Throwing accuracy 2 3 1Running agility 3 1 2
Strength and powerGrip strength 3 1 2Back lifting
strength3 – 1
Explosive power ofarms
2 1 3
Explosive power oflegs
1 2 3
SpeedReaction time 2 1 3Arm movement 2 1 3Leg movement 2 1 3
StaminaBurpee test 1 2 3Kraus-Weber test 1 2 3Hanging with arms 1 2 3
Endurance fitness (ox-ygen power)
2 3 1
note: 3, high; 2, moderate; 1, low.
Fig. 3. Stature of adult males in various social strataof Polish towns, small towns, and villages and in twoKurpie villages, Jeglijowiec and Brzozowka (data fromBielicki and Welon 1982, Bielicki and Waliszko 1991,Szemik 1986). Abbreviations: CE, parents have collegeeducation; C, parents are clerks; SW, parents areskilled workers; USW, parents are unskilled workers;ST, from small towns; VSW, skilled workers from vil-lages; VP, peasants; Jeg, from Jeglijowiec; Brz, fromBrzozowka.
categorize the populations in terms of biological well-being. It cannot be said that the rural agricultural pop-ulation is better off because of its respiratory traits orthat being tall makes the heavily industrialized societyhealthier or that better motor fitness shows the meritsof living in relatively small industrial towns. It can onlybe said that these populations differ.
Differences in physiological functions. In the popu-lation from the coastal area, the Hel Peninsula, individ-uals have low vital capacity, high minute lung volume,low hemoglobin concentration, high blood pressure, andlow heart output. In the population from the PieninyMountains, ca. 700 m above sea level, the values forthese traits are reciprocal: high vital capacity, low min-ute volume, high hemoglobin concentration and hemat-ocrit index, low blood pressure, and high heart output(fig. 2). Which is better? If the hemoglobin concentrationsof 12% typical of the coastal areas are considered normal,then the mountain dwellers can be judged anemic. How-ever, we know that populations with low hemoglobinalso have high vital capacity and minute heart volume,which means that oxygen transport is efficient in a dif-ferent way. Again, people working in agriculture needefficiency in different motor traits from those requiredfor work in industry or in offices. All these differencesshould reflect the conditions under which each popu-lation lives.
The differences between mountain and coastal popu-
lations are also seen in other physiological traits. How-ever, if we assume that all these traits play an importantrole in oxygen transport it is normal for some traits tohave elevated values and others lower values under dif-ferent climatic and probably nutritional conditions be-cause the final result is expected to be the same: thedelivery, under the same workload, of sufficient oxygento the tissues. Therefore it cannot be said that one pop-ulation shows better biological status than others be-cause of the value of a single trait. In populations livingin heavily industrialized areas, the values of almost allthe traits studied are elevated. It may be that a trait thatnormally shows a low value increases under critical con-ditions to help in oxygen delivery and this phenomenonplays a special role in the biological reserve system.Where all physiological traits show elevated values, thismay indicate a lack of reserves that may be destructiveto the organism. The organism adjusts to particular cli-mates and nutritional regimes in different ways, and thefact that physiological traits show high values in somepopulations does not mean that their biological status issatisfactory.
Differences in motor properties. Studies of psycho-motor traits measured in rural, urban, and industrializ-ing-area populations show that high running agility andstatic muscular strength (table 1) characterize the ruralpopulation. The urban population shows a high move-ment memory (proprioceptive sense), throwing accuracy,

Volume 42, Number 2, April 2001 F 305
Fig. 4. Minimal and maximal mean stature in adultmales from various ethnic groups (Williams 1931), set-tlements (villages and towns; Wolanski 1962, 1973,1987; Szemik 1986), social strata (Bielicki and Wal-iszko 1991), and continents, including individuals ofAfrican and European origin (Comas 1971; Evelethand Tanner 1976, 1990; Das 1993). Abbreviations: Mc,Maya and ladinos; vv, two villages; vt, village andtown; vc, village and city; Eu, European; Ea, Euro-pean-ancestry; Aa, African-ancestry; Au, Australian;As, Asian; Af, African; Im, Indo-Mediterranean; Ai,American Indian; Po, Polynesian.
Fig. 5. Standard deviations of stature in adult malesfrom various ethnic groups (Williams 1931), settle-ments (villages, towns; Wolanski 1962, 1973, 1987;Szemik 1986), social strata (Bielicki and Waliszko1991), and continents, including individuals of Afri-can and European origin (Eveleth and Tanner 1976,1990). Abbreviations as for figure 4.
and endurance fitness. The populations of the areas un-der initial industrialization have the highest values forspine flexibility, balance, spatial orientation, explosivepower of arms and legs, reaction time, arm and leg move-ment speed, and stamina. Again, on the basis of motorfunctions and physical fitness we cannot say which pop-ulation is fittest in general terms. Each population showstraits that express its adjustments to a particular work-load, nutritional regime, and set of living conditions.
Differences in body size. Martorell (Martorell and Ha-bicht 1986) believes that greater differences in staturebetween various social strata than between the uppersocial strata of various ethnic groups show that “the var-iation that can be attributed to the environment is sev-eral times greater than that which can be attributed togenetics.” This can be understood as a suggestion thatdifferences between ethnic groups are not a matter ofgenetic differences alone. In our opinion differences inbiological traits among ethnic groups may be related todifferences in gene pools, but they may also be dependenton the environmental conditions under which biologicaldevelopment takes place (nutritional habits, traditionalphysical activity, cultural practices). Martorell is prob-ably right about the developmental adjustment of stat-ure, a phenotypic effect of typical polygenic traits. How-ever, averaging on the population level as an effect ofinbreeding determines the phenotypic picture of a pop-ulation. The dispersion of a trait value in the populationis probably also important.
The differences between different populations and so-cial strata within a population are compared in figs. 3–5.The differences in stature reported by Williams (1931)between Maya and ladinos in the same village in Yucatanare greater than those between two neighboring Polishvillages with different kinds of rural economy (Szemik1986) and between various social strata in Poland (Bie-licki and Waliszko 1991)—the reverse of Martorell’s re-sult—but less than those between urban and rural pop-ulations in Poland (Wolanski 1962, 1973).
Differences between populations should increase withgeographic distance because distance increases environ-mental differences. Face and head breadths are, however,more different between close (see fig. 2) than betweendistant populations (Siniarska 1984), while the differencein mean stature between two neighboring populations isgreater than that between social strata in towns, and thedifference in stature between towns is greater than thedifference between average villages and average towns.Small island populations in Polynesia show greater dif-ferences in stature than the populations of other conti-nents. Rather large standard deviations are found in thesmall populations of Polynesia and Africa, and the dif-ferences between standard deviations in these popula-tions are also large (fig. 5). The large Asian populationsshow the smallest standard deviations and differencesbetween them. Finally, increasingly negative (poor) liv-ing conditions affect the organism in one direction onlyto some extent. For example, poorer living conditionsretard maturation, but excessive emotional stress (dis-tress) can accelerate sexual maturation (Kowalska, Val-sik, and Wolanski 1964, Hulanicka 1986, Prokopec, Dut-kova, and Vignerova 1987).
These considerations indicate that Martorell’s obser-

306 F current anthropology
Fig. 6. Annual increase in length of lower extremitiesin boys from Warsaw in 1959 and 1979 (Wolanski1962, 1987) and in Maya from Yucatan in 1993 (Wo-lanski 1994).
vation tells us only that differences in living conditionsaffecting stature in different human groups (as mean val-ues) are greater between social strata than between theethnic groups he studied. The stabilized local popula-tions are “well” adapted to their regional and local en-vironmental conditions, but social strata are not as“well” adapted because of mobility between strata andrapid changes in living conditions. It is not uncommonfor adults to live under different conditions from thosethey experienced as children.
Differences in body proportions. Whether body pro-portions are related to genetic differences or express nu-tritional status is the subject of active debate. Body pro-portions depend on nutrition, hormonal activity,workload in the period of growth, and other things (La-sota, Tomaszewska, and Wolanski 1966, Carter 1984).Maya from Yucatan have short lower and relatively longupper extremities, and Polish populations have longlower and relatively short upper extremities (Wolanskiand Siniarska 1997). The analysis of annual growth in-crements shows that the increase in lower extremitylength is seen at about the fourth or sixth year of lifeand between 10 and 13 years in various populations (Wo-lanski and Siniarska 2000). There are also differences inrate of growth between ethnic groups. However, studiesof the population of Warsaw between 1959 and 1979 re-vealed changes in the magnitude of annual incrementsand a small shift in the age of their occurrence (fig. 6)showing that this phenomenon depends strongly on en-vironmental factors. (Figure 6 presents data for boys, butthe same phenomenon is observed in girls [Wolanski andSiniarska 2000].) Thus, shorter legs probably representpoorer nutritional status, especially in the period of theirintensive growth. If this is true, different body propor-tions may be related to environmental factors and rep-
resent developmental adjustments, but genetic compo-nents cannot be excluded.
Compensatory mechanisms. In addition to the above-mentioned differences between populations, there areseveral compensatory (self-control) mechanisms at thepopulation level. For example, these populations arecharacterized by positive secular changes in stature anda tendency toward the arithmetic mean. As a result, thechanges in stature between generations of the populationlevel are different from those expected from averagingthe values for parents and their offspring.
A model for some more complex changes in the caseof lung volume is as follows: Vital capacity is generallyvery closely related to body size. With the increase inbody size, both an increase in vital capacity and a ten-dency toward the arithmetic mean should be expected.Secular changes toward a slimmer body will probablyaffect vital capacity negatively. However, pollution pro-motes an increase in vital capacity and ventilation, whilea more sedentary lifestyle or less physical activity re-duces vital capacity.
discussion
Human organisms have become genetically differenti-ated in the course of evolution, but this differentiationdoes not reflect the ecosystems and niches they occupytoday. Contemporary humans live under more uniformconditions than those in which their ancestors evolvedhundreds of thousands of years ago in other parts of theworld. At the same time, current living conditions (re-lations between people, between different social groupsand groups of different occupations, etc.) create new de-mands and give rise to new characteristics (traits).
The biological status of a population cannot be as-sessed on the basis of a single trait. Each trait has itsown adaptive value and shows specific adaptive changesto particular living conditions. Some groups of traits arelinked with particular functions of the organisms suchas oxygen transport, metabolism, and/or adaptation toworkload. Some traits promote better results in running,others in throwing, gymnastics, swimming, etc. Thereis no one trait that is objectively good. Each trait has adifferent value for a different activity (performance). Itcannot be argued, for example, that tallness representsbetter adaptation. It is only the effect of adjustment togood nutrition, optimal movement activity, absence ofdisease, etc. In other words, it is a consequence of par-ticular living conditions and lifestyles against the back-ground of genetic predisposition. It has been suggestedthat each population has its own genetically determinedsensitivity to environmental factors and that under thesame conditions stature change in various populationsmay vary (Lauw and Henneberg 1997).
Surprisingly, given that we would expect geographicaldistance to increase genetic distance and to be linked tovariation in living conditions, differences in head andface breadths have proved greater between close neigh-bors than between distant populations, and there are sim-ilar findings with regard to stature. Contemporary Polish

Volume 42, Number 2, April 2001 F 307
villages are not stratified internally but differ amongthemselves with regard to localization and type of econ-omy. These differences between villages seem to corre-spond to the differences between social strata in cities.
Biological traits must be analyzed as a system, takinginto account not only their mean values but their dis-persion as well. This is also true of the gene pool. Thedifferent gene frequencies and the degree of heteroge-neity of individuals in a population probably influencethe mean and median values, skewness, and dispersionof the distribution. Because of this, mean values do notalways express the real genetic difference (distance). Inthis connection, two well-known regularities should bementioned: the secular trend toward greater stature un-der favorable living conditions and the tendency towardthe arithmetic mean. These two tendencies counter eachother, and the final effect approaches more average val-ues and limited dispersion. There are probably other suchmechanisms.
There is no satisfactory way to answer all the ques-tions or to resolve all the issues raised by these studies.Instead, they may serve as the beginning of a very im-portant discussion of the criteria used to assess the bi-ological status of a population as a mirror of socioeco-nomic and cultural changes.
conclusions
1. The different biological traits of the organism are notindependent; each trait represents the organism as awhole but plays its own role.
2. No one population is better adapted in biologicalterms than others; the assessment of individual traitsdoes not allow such a conclusion.
3. Differences between populations have adaptivemeaning. They correspond to the conditions (environ-mental and lifestyle) under which individuals developedand genetic selection (differential fertility and mortality).
4. Differences between populations, even those thatare easy to identify, are very hard to interpret. Theircauses are complex and their mechanisms latent. Oneof them is a replacement of functions (a compensatorymechanism) in the realization of a certain purpose, andanother is a self-control mechanism related to trait var-iation. Both mechanisms are very complicated, and theyare integrated into the internal environment of the or-ganism, which is under the control of the neurohormonalsystem.
5. Differences in stature between social strata may begreater than those between the upper strata of variousethnic groups for some samples, probably because of dif-ferences in living conditions. The results analyzed do notexclude the possibility of genetic differences among eth-nic groups in stature and other morphological and phys-iological traits (skin, hair, and eye color). The only con-clusion to be drawn is that stature has adaptive meaningin both evolutionary and environmental terms.
6. The biological status of a population cannot be iden-tified by a single trait. Indicators of biological statusshould be selected in terms of changes in environmental
conditions, mode of life, and the expected responses oforganisms and populations. Stature, which is a very sen-sitive indicator of socioeconomic changes, must be sup-ported by other indicators (for example, respiratory andpsychomotor traits).
References Cited
b i e l i c k i , t . , a n d h . w a l i s z k o . 1991. Urbanization-depen-dent gradients in stature among Polish conscripts in 1976 and1986. American Journal of Human Biology 3:419–24.
b i e l i c k i , t . , a n d z . w e l o n . 1982. Growth data as indica-tor of social inequalities: The case of Poland. Yearbook ofPhysical Anthropology 25:153–67.
c a r t e r , j . e . l . Editor. 1984. Physical structure of Olympicathletes. Pt. 2. Kinanthropometry of Olympic athletes. Basel:Karger.
c o m a s , j . 1971. “Anthropometric studies in Latin AmericanIndian populations,” in The ongoing evolution of Latin Ameri-can populations. Edited by F. M. Salzano, pp. 333-404. Spring-field: Thomas.
d a s , b . m . 1993. “Physical anthropology of the tribes of North-east India,” in New perspective in anthropology. Edited by P.K. Seth and S. Seth, pp. 159-92. New Delhi: MD Publications.
e v e l e t h , p . b . , a n d j . m . t a n n e r . 1976. World-wide vari-ation in human growth. Cambridge: Cambridge UniversityPress.
———. 1990. 2d edition. World-wide variation in human growth.Cambridge: Cambridge University Press.
h u l a n i c k a , b . 1986. Effect of psychological and emotionalfactors on age at menarche. Materiały Prace Antropologii 107:45–80.
k o w a l s k a , i . , j . a . v a l s i k , a n d n . w o l a n s k i . 1963.Season of the year in which menstruation begins in relation toage and social and geographical environment. Prace MateriałyNauk IMD 1:81–108.
k o z i o ł , r . , a n d n . w o l a n s k i . 1982. “Respiratory and car-diovascular system in populations from areas of different de-grees of industrialization,” in Ekologia populacji ludzkich. Ed-ited by N. Wolanski and A. Siniarska, pp. 397–438. Wrocław:Ossolineum.
l a s o t a , a . , h . t o m a s z e w s k a , a n d n . w o l a n s k i .1966. Differentiation of body proportions and certain body frag-ments in children with disturbed growth. Polish Endocrinology17(1–2):10–20.
l a u w, g . j . , a n d m . h e n n e b e r g . 1997. Lack of seculartrend in adult stature in white South African males born be-tween 1954 and 1975. Homo 48(1):54–61.
m a r t o r e l l , r . , a n d j . p . h a b i c h t . 1986. “Growth inearly childhood in developing countries,” in Human growth.Edited by F. Falkner and J. M. Tanner, vol. 3, pp. 241–62. NewYork: Plenum Press.
p ro k o p e c , m . , l . d u t k o v a , a n d j . v i g n e ro v a . 1987.End of secular trend in the age at menarche in Czech girls.Proceedings of the 8th International Anthropological PosterConference, Zagreb, pp. 28–31.
p y z u k , m . 1973. Regions and populations under study. Studiesin Human Ecology. 1:34–57.
p y z u k , m . , a n d n . w o l a n s k i . 1972. Respiratory and cardi-ovascular systems of children under various environmentalconditions. Warszawa: PWN.
s i n i a r s k a , a . 1984. Interpopulational and interfamilial differ-ences in some somatic, physiological, and psychomotor traits.Studies in Human Ecology 5:195–221.
———. 1996. “Family environment, parents’ constitutional char-acteristics, and biological development of children,” in Familyas an environment for human development. Edited by N. Wo-lanski and B. Bogin, pp. 145–83. Delhi: Kamla Raj Enterprises.
s i n i a r s k a , a . , a n d n . w o l a n s k i . 1986. Agility and

308 F current anthropology
movement co-ordination in Polish populations, from 2 to morethan 80 years of age. Studies in Human Ecology 7:189–203.
s z e m i k , m . 1986. Changes in somatic development of inhabi-tants of two neighboring villages in Kurpie region (Poland) be-tween 1961–1981. Studies in Human Ecology 7:65–93.
w i l l i a m s , g . r . 1931. Maya-Spanish crosses in Yucatan. Pa-pers of the Peabody Museum of American Archaeology andEthnology 13(1).
w o l a n s k i , n . 1962. Kinetics and dynamics of growth and dif-ferentiation of body proportions in children and youth. War-szawa: PZWL.
———. 1966. Environmental modification of human form andfunction. Annals of the New York Academy of Sciences 134:826–40.
———. 1973. Tables of basic data. Studies of Human Ecology 1:197–226.
———. 1987 “Data for control of child growth (biological refer-ence systems—Warszawa 1981),” in Ocena rozwoju dziecka wzdrowiu i chorobie. Edited by N. Wolanski and R. Kozioł, pp.359–413. Wrocław: Ossolineum.
———. 1990. Human population as bioindicator of environmen-tal conditions (Environmental factors in biological status of thepopulation of Poland). Studies in Human Ecology 9:295–321.
———. 1994. La familia como microambiente para el desarrollode los ninos: Estudios en la poblacion de Merida y Progreso,1993 (Resultados preliminares). Final technical report,CONACyT ref. no. 1324-S9206, Merida, Mexico, CINVESTAV.
w o l a n s k i , n . , a n d a . l a s o t a . 1964. Physical develop-ment of countryside children and youth aged 2 to 20 years ascompared with the development of town youth of the sameage. Zeitschrift fur Morphologie und Anthropologie 54:272–92.
w o l a n s k i , n . , a n d a . s i n i a r s k a . 1997. Body proportionsin populations of Yucatan (Abstract). American Journal of Hu-man Biology 9(1):144.
———. 2000. “Differences in body proportions between Yucate-can and Polish populations: Genes or living conditions?” inLatvijas Vesturnieki, pp. 166–92. Riga: Latvijas Vestures Insti-tuta Apgads.
w o l a n s k i , n . , a . s i n i a r s k a , a n d m . s z e m i k . 1982.“Polskie badania populacyjne,” in Ekologia populacji ludzkich.Edited by N. Wolanski and A. Siniarska, pp. 721–38. Wrocław:Ossolineum.
Ceramic Age Seafaring andInteraction Potential in theAntilles: A Computer Simulation1
richard t. callaghanDepartment of Archaeology, University of Calgary,Calgary, Alberta, Canada T2N 1N4. 5 x 00
� 2001 by The Wenner-Gren Foundation for Anthropological Re-search. All rights reserved 0011-3204/2001/4202-0007$1.00.
1. Funding for this work was provided by a University of CalgaryResearch Services Pilot Grant and a Social Sciences and HumanitiesResearch Council of Canada Research Grant. Donna Fremont ofthe University of Calgary, Computer Science Department, carriedout the computer programming. I thank the Belizeans and Vene-zuelans who helped to construct rafts and test canoes, especiallyMorris Lesley of Sand Hill, Belize, and the Ye’Kwana people of thevillage of Esmeralda, Venezuela.
Computer simulations of ancient maritime voyaging pat-terns have shed light on a number of issues regardingthe movements of prehistoric and historic peoples. Thefirst simulation of this kind was designed by Levison,Ward, and Webb (1973) to investigate Polynesian disper-sal. The technique was later used to determine which ofthe Bahamas Islands is likely to have been the first onwhich Columbus landed (Fuson 1987). Probably the mostcomplex of these simulations are those of Geoffrey Irwinand his colleagues (Irwin 1989, 1990, 1992; Irwin, Bick-ler, and Quirke 1990), focused on exploration, coloni-zation, and settlement patterns in Polynesia. A handfulof other such works have been conducted elsewhere.This study uses computer simulations of the maritimeenvironment and performance characteristics of aborig-inal watercraft to investigate whether technology or theenvironment would have influenced patterns of humanmovement among the islands of the Antilles or betweenthe islands and the South American mainland in theCeramic Age, beginning in the first few centuries b.c.(fig. 1). Two models are tested: chance discovery throughunintentional drift voyages (people lost at sea) and di-rected voyages (intentional exploration).
The evidence for a northern South American origin ofthe earliest ceramic-producing horticulturists in the An-tilles is very clear (Rouse 1992:71–104; cf. Haviser 1997).These peoples, termed Saladoid, appear to have movedrapidly through the Antilles as far as eastern Hispaniolain the second half of the 1st millennium b.c. (Rouse1986:39; 1992:79–80). In passing through the Lesser An-tilles they initially occupied the northeast coasts of thehigh islands, presumably preferring those locations be-cause they were the most heavily forested parts of theislands and resembled their original home on the main-land (Rouse 1992:9). As Rouse points out, these are thewindward sides of the islands, and their settlementwould indicate that the Saladoid peoples had a good com-mand of seamanship. This group or some of its variantsgave rise around a.d. 600 (p. 92) to the Ostionoid peoples,who then moved west into eastern Cuba. Eventually, theOstionoid peoples gave rise to the Taıno groups encoun-tered by Columbus (pp. 72–73).
Two dugout canoe designs were evaluated in thisstudy. One is based on the Stargate canoe (fig. 2), recov-ered in the Bahamas by Stephanie Schwabe and the lateRob Palmer. This canoe is remarkably similar to canoesstill being used by the Ye’Kwana and other native groupson the Orinoco River of Venezuela today (Callaghan andSchwabe n.d.). The other (fig. 3) is a platform-style canoefound widely today around the Caribbean mainland andsimilar to those depicted in the early Spanish chroniclesof the islands (Callaghan 1993). Canoes of the two styleswere analyzed in Venezuela and Belize respectively todetermine their performance characteristics, includingspeed, leeway, carrying capacity, and stability. Thesecharacteristics were also analyzed using naval architec-ture programs. The data were then used in a simulationmodel of the Caribbean environment. The environmen-tal factors considered were winds, currents, gale and hur-

Volume 42, Number 2, April 2001 F 309
Fig. 1. The Caribbean region, showing staging area for voyages.
ricane frequencies, and sea-swell conditions, and the dataon these factors were taken from pilot charts and sailingdirections compiled by the U.S. Navy and other agencies.The computer program employed for the analysis hasbeen described elsewhere (Callaghan 1999).
One of the most important questions to be asked inapplying the model is whether the data presented in thepilot charts for the study area, compiled since the early19th century, are representative of the time period ofinterest here. The most important climatic factor af-fecting the model is surface wind circulation during theperiod from approximately 2,500 b.p. and 500 b.p. Surfacewinds not only affect vessels directly but also are theprimary determinants of surface current direction.Therefore the question is whether surface wind circu-lation for the period differed significantly from presentconditions.
According to Clarke’s (1989:44) summary of weatherpatterns in the area today, the Caribbean lies within thewind belt known as the Northeast Trades. With the ex-ception of disturbances from tropical cyclones, theweather is quite stable. The prevailing winds are easterlyand usually steadiest in the south of the region duringthe period between December and May. Summer and fallare warmer and more humid than winter and spring.Cloud cover and rainfall increase, as does thunderstormactivity, and winds are often lighter and more variable.
Tropical cyclones are most likely in summer and fall.The northern limit of the Northeast Trades is 28� N lat.and is reached between July and September. At this time,the strongest and steadiest winds pass through the mid-dle of the region; near the northern limit they tend tobe more variable. The limit shifts south to about 24� Nbetween February and April. On average the winds blow11–15 knots from the east-northeast. The northern Les-ser Antilles experience the steadiest winds in the sum-mer months; for the more southern islands and the coastof South America winds are steadiest in winter becauseof the southern shift of the central portion of the tradewind belt. The Bahamas are geographically outside of theCaribbean region but during the Ceramic Age were cul-turally within it. The northern Bahamas, north of 24� N,are beyond the trade winds in winter, and at this timethey experience lighter winds that are more variable indirection and occasional strong winds from the north.The wind shifts east to southeast in the summer withthe return of the trades.
Hodell et al. (1991) present a high-resolution recon-struction of the Caribbean climate for the past 10,500years based on 18O/16O ratios in ostracod shells fromHaiti’s Lake Miragoane. Variation in the ratios reflectschanges in precipitation for the period. From about 2,400b.p. to 1,500 b.p. the 18O/16O values and variation are verysimilar to those for the past 900 years (1991:fig. 2). The

310 F current anthropology
Fig. 2. The Stargate canoe, South Andros Island, Bahamas.
values indicate a drying trend for both periods (p. 792).For the intervening years from 1,500 to 900 b.p. the val-ues indicate a brief wetter period but one not as wet asthe early to mid-Holocene. While there is variation inrainfall for the period of interest here, 500 to 2,500 b.p.,it does not approach the overall variation for the past10,500 years. Hodell et al. note the correlation betweenprecipitation anomalies and variation in the annual cli-matic cycle in the Caribbean region discussed above:“Enhancement of the annual cycle led to years of an-omalously high precipitation, whereas a reduction led toa deficient rainy season” (p. 792). Thus reconstructionof variation in precipitation should be an accurate in-dicator of variation in the annual cycle.
The annual cycle is controlled by the summer dis-placement of the North Atlantic subtropical high by thenorthward movement of the Intertropical ConvergenceZone and the reverse movement in winter. Hodell et al.compared their data with the changes in annual cycleintensity estimated from the seasonal insolation differ-ence at the top of the atmosphere at 10� N between Au-gust and February and found the changes in the two re-cords for their 10,500-year period to be similar. Thisreinforces the conclusion that while variations from pre-sent climatic conditions including surface wind patternsexisted during the Caribbean Ceramic Age, they werenot substantial. It also suggests a major mechanism forvariation in both precipitation and the annual cycle inthe form of “orbitally forced (Milankovitch) variationsin solar insolation” (p. 792).
The field on which the simulation program operatesis the Caribbean Sea, the Gulf of Mexico, and the sur-rounding mainland (U.S. Navy 1995). The area is dividedinto two-degree Marsden squares (two degrees of latitudeby two degrees of longitude), with each square containingwind and current vectors as they have been recorded tooccur for a particular month of the year. A separate fieldis used for each month. Any starting position can bechosen on the field. Positions 20 or 30 nautical miles offthe coast were chosen in order to prevent all vessels fromsimply returning to the nearest coast.
The operator chooses a watercraft type and indicatesits initial position. For each vessel type there are data onthe speed that it can be paddled and the effect on it ofvarious wind speeds. Vessels can be allowed either todrift before the wind or to be paddled in a specific di-rection. Each two-degree Marsden square contains eightwind vectors (cardinal and intercardinal points) and thepercentage of the total number of observations in whichthe wind has blown from each direction. The percentageof observations in which calms are recorded is also given.The wind direction is chosen randomly by the programbut is weighted to reflect actual observations. The effectof the chosen wind on a particular vessel is then addedto the current vector for each two-degree square and thenew position is calculated. If vessels are paddled in aspecific direction, this needs to be added to the othervectors in order to obtain the new position. The durationover which the vectors affect the vessel between posi-tions is 24 hours, following Levison, Ward, and Webb(1973:24–25). The procedure is repeated until the vesseleither reaches an island (or any area designated as a suc-cess) or is forced back to the mainland (or off the field).Up to 1,000 runs from the same start can be simulatedat a time.
Two questions were asked of the simulation. The firstquestion was how likely it was that the Saladoid peoplesfrom South America would have discovered the Antillesby chance. Each of the two canoes was placed at differentpoints along a 600-nautical-mile staging area off thecoast of northern South America, and the simulation wasrun on the assumption that the vessels were allowed todrift. Although this may not seem a likely response tobeing lost at sea, it has the advantage of allowing thevessel to cover the maximum distance without expen-diture of energy. In a storm situation there is often noother rational option. The simulation was run underweather conditions for each of four months—January,April, July, and October. The percentages of successfulchance discoveries of the islands ranged from 0.3% underApril conditions to 0.1% under October conditions. Thehigher success rates for April conditions have some sig-

Volume 42, Number 2, April 2001 F 311
Fig. 3. A platform-style canoe found widely around the Caribbean mainland and historically recorded in theislands.
nificance in that April is the only month in which nohurricanes have ever been recorded (Clarke 1989:50).
Work in Polynesia has provided a means of estimatingrisk to the crew. Risk is determined by length of timeat sea. Lengthy drift voyages in open boats due to ship-wreck or other misfortune are well known for the PacificOcean under conditions similar to those of the Carib-bean. The maximum recorded drift seems to be on theorder of seven to eight months. Several recorded voyagescovered distances of ca. 3,000 miles over a period of sixto ten weeks, and a great number covered shorter dis-tances (Howay 1944). Levison, Ward, and Webb (1973:21) used the survival probabilities presented by McCanceet al. (1956), based on 27,000 persons lost at sea, to rep-resent the cumulative percentage of crew losses. Fromthis I estimate that a successful drift voyage from theSouth American coast taking four to five weeks wouldhave involved a probable crew loss of 10% to12%—meaning that one or two crew members out of atotal of eight to ten can be expected to have died en route.
A slight difference in the success rates of the two ves-sels is attributable to the difference in the effects of thewind on their different shapes. There was little differencein the success rates from various points along the coast.Although success rates were low, given the long stretchof coast from which success is possible and the longhistory of coastal adaptations in the region it seems prob-able that such drift voyages occurred.
Chance discovery of Grenada from a position off thenorth coast of Trinidad is not indicated by the simula-tion. The water gap between the continental islands ofTrinidad/Tobago and Grenada is the only gap along theLesser Antilles that cannot be seen across. It is possible,however, for an observer to see Trinidad and Grenada,Tobago and Grenada, or even all three islands from po-sitions at sea. For Trinidad and Grenada there is a 25-nautical-mile overlap in their sighting distances, and forTobago and Grenada there is a 15-nautical-mile overlap.There is also a 15-by-25-nautical-mile triangular area atsea from which all three islands can be observed given
reasonable visibility. It at first appears that someone in-tentionally exploring for new land would not have hadto leave sight of home (Trinidad or Tobago) in order tosight Grenada. However, for intentional discovery ofGrenada from Trinidad explorers in a canoe would havehad to steer a bearing considerably to the east of thetarget, and this would have virtually prevented theirpassing through the areas of mutual interisland visibility.Chance discovery still does not appear likely, as the areasof mutual visibility are relatively small and not on thedrift voyage paths.
Another possibility for detecting Grenada from Trin-idad or Tobago is the use of clouds as land indicators.Stationary cumulus clouds can form over high islandssuch as Grenada and indicate their location. Burch (1986:197) states that stationary cumulus clouds form at alllatitudes but may be obscured by lower clouds. Suchclouds when developed into cumulonimbus clouds canreach heights of more than 50,000 feet in the tropics(Admiralty Hydrographic Department 1941:50). Even astationary cloud with an upper height of 7,300 feet wouldmake Grenada detectable from the coast of Trinidad. Thequestion remains whether the first Ceramic Age explor-ers would have had the opportunity to learn about thiseffect in their initial forays north, particularly as suchclouds are most useful for navigation before midday(Burch 1986:197). Further, these explorers would stillhave had to become familiar with the wind and currentpatterns to steer the correct bearing.
The majority of the earliest Early Ceramic Age datesare north of the Guadeloupe Passage, the Fond Brule siteon Martinique being the only exception (Haviser 1997).Although the pattern may be a sampling bias, it does fitwith a chance discovery of the Greater Antilles andNorthern Lesser Antilles before the islands of the South-ern Lesser Antilles. This raises the possibility that thesettlement of the islands by Saladoid peoples was not asimple south-to-north progression.
The second question was whether movement alongthe island chain was so constrained by technology and

312 F current anthropology
the environment that it had to be conducted in “step-ping-stone” fashion or whether travel between the SouthAmerican mainland and the northern islands was pos-sible. To investigate this question the two canoes wereagain placed along the staging area off the north coast ofSouth America with weather conditions for January,April, July, and October. Now, rather than being allowedto drift, the canoes were paddled by their occupants(eight per canoe) in shifts of four, eight hours at a time.The speeds used were calculated from tests in the field,naval architecture programs (Callaghan 1999), and thehuman-endurance data provided by Horvath and Finney(1976). With regard to navigation skill the only assump-tion was that the occupants directed the canoe north-ward. Success in this series of experiments was definedby simply reaching the islands of the Greater Antillesfrom the same mainland area as in the previous seriesof experiments. A range of paddled speeds from 3.4 knotsto 2.0 knots was employed. All voyages were successfulunder these stipulations, with only slight variations oflanding sites despite variation of speed. Vessels ended upvariously in an area from eastern Puerto Rico to westernHispaniola within five to six days. For intentional voy-ages from all areas the probability of crew loss was lessthan 1%, which does not translate into a fatality. Returnvoyages from north to south did not indicate any sig-nificant differences; in both directions winds and cur-rents are moving across the path of the vessels.
Applying the results presented here to the Saladoidperiod, it appears likely that direct crossings of the Car-ibbean Sea were undertaken either between the main-land and islands like Puerto Rico or between any islandsof the Lesser Antilles not adjacent to one another. Oncethe locations of the Greater Antilles were known, directcontact was possible between the Venezuelan mainlandand islands such as Puerto Rico as some researchers havesuggested (Chanlatte Baik and Narganes Stordes 1989,Rodrıguez and Rivera 1991, Zucchi 1984). Movementwithin the Lesser Antilles need not have followed a step-ping-stone-like pattern. Only modest navigation skillwould have been required, and even moderate-sized ca-noes could have made a direct crossing in five to six days.
A direct route between Venezuela and Puerto Rico maynot at first seem advantageous. It would, however, havebeen only about two-thirds the distance involved in fol-lowing the curve of the Lesser Antilles island chain.Moreover, the channels between the Lesser Antilles of-ten have currents of 3 knots, and therefore voyagers fol-lowing the Lesser Antilles would have encounteredcrosscurrents three to four times the strength of thoseencountered in a direct crossing. The bottleneck effecton waves that had built up while crossing the Atlanticwould also have made crossing the channels less desir-able for small vessels than passing to the west (Stoneand Hays 1991:224). Finally, the deep valleys and highland of some of the Lesser Antilles, for example, Dom-inica, produce heavy squalls that are a danger to vessels(Defense Mapping Agency 1985:152).
Another factor in direct crossing between the main-land and Puerto Rico or Hispaniola that should be noted
is the formation of altocumulus lenticularis or mountainwave clouds (Burch 1986:197) on the south coast ofPuerto Rico. These are highly distinctive, sharply definedlenticular clouds that form when moist air passes overmountains (Admiralty Hydrographic Department 1941:230). Cumulus clouds form on the mountaintops, andlenticular clouds break off and drift out to sea. As thenortheast winds pass over islands like Puerto Rico theseclouds move southward into the Caribbean. They canmaintain their very distinctive shape for long distances,and I have observed them 240 nautical miles from themountain ranges that formed them. Under these con-ditions voyagers would have had a clear indication ofland for half of the distance from Venezuela to PuertoRico.
Overall, a direct route would have been shorter, safer,and easier than a route along the Lesser Antilles. If voy-ages were made in April, there would have been littledanger of encountering tropical storms in the open sea.The expected loss of crew over a five-to-six-day periodwould have been less than 1%. Finally, for the Taınotoward the end of the prehistoric period a direct routemay have had the advantage of avoiding islands of theLesser Antilles occupied by the reputedly hostile IslandCaribs. Neither the environment nor the available sea-faring technology would have forced people to travelalong any particular route. Any pattern that may emergefrom the analysis of lapidary materials (see, e.g., Ball1941; Chanlatte Baik 1983; Cody 1991a, b, c; Rodrıguez1991; Watters and Scaglion 1994; Watters 1997; Murphyet al. 2000) will be due to social, political, and economicfactors.
References Citeda d m i r a l t y h y d ro g r a p h i c d e p a r t m e n t . 1941. Admi-
ralty weather manual. London: H.M. Stationery Office.b a l l , s . h . 1941. The mining of gems and ornamental stones
by American Indians. Smithsonian Institution, Bureau ofAmerican Ethnology Bulletin 128, Anthropological Papers 13:1–77.
b u r c h , d a v i d . 1986. Emergency navigation. Camden: Inter-national Marine.
c a l l a g h a n , r . t . 1993. “Passages to the Greater Antilles:An analysis of watercraft and the marine environment,” inProceedings of the Fourteenth Congress of the InternationalAssociation for Caribbean Archaeology, Christ Church, Barba-dos, 1991. Edited by A. Cummins and P. King, pp. 33–39.Martinique.
———. 1999. Computer simulations of ancient voyaging. TheNorthern Mariner/Le Marin du nord 9(2):11–22.
c a l l a g h a n , r . t . , a n d s t e p h a n i e j . s c h w a b e . n.d.“Watercraft of the islands,” in Proceedings of the EighteenthInternational Congress for Caribbean Archaeology, Grenada,1999. In press.
c h a n l a t t e b a i k , l . a . 1983. Catalogo arqueologica de Vie-ques. Rıo Piedras: Museo de Antropologia, Historia y Arte,Universidad de Puerto Rico.
c h a n l a t t e b a i k , l . a . , a n d y. m . n a r g a n e s s t o r d e .1989. La nueva arqueologıa de Puerto Rico (Su proyeccion enLas Antilles). Museo del Hombre Dominicano Boletın 16 (22):9–63.
c l a r k e , j a m e s . 1989. Atlantic pilot atlas. Camden: Interna-tional Marine.

Volume 42, Number 2, April 2001 F 313
c o d y, a . k . 1991a. Prehistoric patterns of exchange in the Les-ser Antilles: Materials, models, and preliminary observations.Ann Arbor: UMI.
———. 1991b. “Distribution of exotic stone artifacts through theLesser Antilles: Their implications for prehistoric interactionand exchange,” in Proceedings of the Fourteenth Congress ofthe International Association for Caribbean Archaeology, Bar-bados, 1991. Edited by A. Cummins and P. King, pp. 204–26.
———. 1991c. “From the site of Pearls, Grenada: Exotic lithicsand radiocarbon dates,” in Proceedings of the Thirteenth Con-gress of the International Association for Caribbean Archae-ology, Curacao, 1990. Edited by E. N. Ayubi and J. B. Haviser,pp. 589–604. Reports of the Archaeological-Anthropological In-stitute of the Netherlands Antilles 9.
d e f e n s e m a p p i n g a g e n c y h y d ro g r a p h i c / t o p o -g r a p h i c c e n t e r . 1985. Sailing directions for the Carib-bean Sea. Vol. 1. Washington, D.C.
f u s o n , ro b e r t h . 1987. The log of Christopher Columbus.Camden: International Marine.
h a v i s e r , j . b . 1997. “Settlement strategies in the Early Ce-ramic Age,” in The indigenous people of the Caribbean. Editedby Samuel M. Wilson, pp. 57–69. Gainseville: University ofFlorida Press.
h o d e l l , d a v i d a . , j a s o n h . c u r t i s , g l e n a . j o n e s ,a n t o n i a h i g u e r a - g u n d y, m a r k b r e n n e r , m i -c h a e l w. b i n f o r d , a n d k a t h l e e n t . d o r s e y. 1991.Reconstruction of Caribbean climate change over the past10,500 years. Nature 352:790–93.
h o rv a t h , s . m . , a n d b . r . fi n n e y. 1976. “Paddling experi-ments and the question of Polynesian voyaging,” in Polynesiannavigation and voyaging. Edited by Ben R. Finney, pp. 47–54.Polynesian Society Memoir 39.
h o w a y, f . w. 1944. Some lengthy open-boat voyages in thePacific Ocean. American Neptune 4:53–57.
i r w i n , g e o f f r e y. 1989. Against, across, and down the wind.Journal of the Polynesian Society 98:167–206.
———. 1990. Human colonization and change in the remote Pa-cific. current anthropology 31:90–94.
———. 1992. The prehistoric exploration and colonization of thePacific. Cambridge: Cambridge University Press.
i r w i n , g e o f f r e y, s . b i c k l e r , a n d p . q u i r k e . 1990.Voyaging by canoe and computer. Antiquity 64:35–50.
l e v i s o n , m . , r . g . w a r d , a n d j . w. w e b b . 1973. Thesettlement of Polynesia: A computer simulation. Minneapolis:University of Minnesota Press.
m c c a n c e , r . a . , c . c . u n g l e y, j . w. l . c ro s fi l l ,a n d e . m . w i d d o w s o n . 1956. The hazards to men lost atsea, 1940–44. Medical Research Council [London] Special Re-port 291.
m u r p h y, a . r e g , d a v i d j . h o z j a n , c h r i s t y n . d em i l l e , a n d a l f r e d a . l e v i n s o n . 2000. Pre-Columbiangems and ornamental materials from Antigua, West Indies.Gems and Gemology 36:234–45.
ro d r ı g u e z , m . 1991. “Early trade networks in the Carib-bean,” in Proceedings of the Fourteenth Congress of the Inter-national Association for Caribbean Archaeology. Edited by Al-issandra Cummins and Philippa King, pp. 306–14. St. Ann’sGarrison: Barbados Museum and Historical Society.
ro d r ı g u e z , m . , a n d v. r i v e r a . 1991. “Puerto Rico andthe Caribbean Pre-Saladoid ‘crosshatch connection,’” in Pro-ceedings of the Twelfth Congress of the International Associa-tion for Caribbean Archaeology. Edited by Linda Sickler Rob-inson, pp. 45–51. Martinique.
ro u s e , i . 1986. Migrations in prehistory: Inferring populationmovement from cultural remains. New Haven: Yale Univer-sity Press.
———. 1992. The Tainos. New Haven: Yale University Press.s t o n e , w i l l i a m t . , a n d a n n e m . h a y s . 1991. A cruising
guide to the Caribbean. New York: Putnam.u . s . n a v y. 1995. Marine climatic atlas of the world. Asheville:
National Climatic Data Center.w a t t e r s , d . r . 1997. “Maritime trade in the prehistoric East-
ern Caribbean,” in The indigenous people of the Caribbean.Edited by Samuel M. Wilson, pp. 88–99. Gainseville: Univer-sity Press of Florida.
w a t t e r s , d . r . , a n d r . s c a g l i o n . 1994. Beads and pen-dants from Trants, Montserrat: Implications for the prehistoriclapidary industry of the Caribbean. Annals of Carnegie Mu-seum 63:215–37.
z u c c h i , a . 1984. “Nueva evidencia sobre la penetracıon degroupos Ceramicos a las Antillas Mayores,” in Relaciones pre-hispanicas de Venezuela. Edited by Erika Wagner, pp. 35–50.Caracas: Acta Cientifıca Venozolana.




















