On the anionic homo‐ and copolymerization of ethyl‐ and butyl‐2‐cyanoacrylates

Homo tsaichangensis
and Gigantopithecus
Mark A. S. McMenamin
Meanma Press
2015
Published January 28, 2015

2
Meanma Press South Hadley, Massachusetts, USA ©2015 by Mark A. S. McMenamin All rights reserved. McMenamin, Mark A. S. Homo tsaichangensis and Gigantopithecus/Mark A. S. McMenamin ISBN 1-893882-19-5 ISBN13 978-1-893882-19-5 Printed in the United States of America
c 10 9 8 7 6 5
DOI: 10.13140/2.1.3463.7121

3
INTRODUCTION
EVER SINCE Ralph von Koenigswald (von Koenigswald 1931, 1935, 1952), Pierre
Teilhard de Chardin (Teilhard 1926) and other Western scientists entered Asia in the
early decades of the 20th Century to search for paleontological data bearing on human
origins, Asia has been seen as rich in promise for this compelling area of paleontological
research. Recent new discoveries in the region are beginning to greatly increase our
understanding of Pleistocene higher primate evolution in East Asia.
An archaic human mandible ("Penghu Man" or Penghu 1, here named Homo
tsaichangensis n. sp.) dredged from the Penghu Channel off the west coast of Taiwan
shows intriguing similarities to a newly reported jaw fragment from Tegal, central Java,
here assigned to Gigantopithecus cf. G. blacki. These Pleistocene creatures may have
inhabited similar environments as part of what is referred to here as the Tegal-Penghu
Biogeographic Province. In addition to the primates, this province hosted the
proboscidean Stegodon and the hyaenid Crocuta. Morphological similarity between the
mandibular dentition of H. tsaichangensis n. sp. and Gigantopithecus blacki von
Koenigswald, 1935 suggest shared dietary preferences; indeed, the increased size of teeth
in H. tsaichangensis n. sp. contrasts sharply with the typical evolutionary trend of
"dentognathic reduction" (Chang et al. 2015) in successive species within the genus
Homo. The similarities represent an unusual case of convergent evolution between the
genus Homo and an unrelated group of primates.

4
MAJOR RANGE EXTENSION FOR GIGANTOPITHECUS
On December 1, 2014, The Jakarta Post reported the discovery of a Gigantopithecus jaw
fragment from the Semedo excavation site, Semedo Village, Kedungbanteng District,
Tegal regency, central Java (Maryono 2014). The jaw fragment was found in association
with the fossil remains of a dwarf proboscidean belonging to the genus Stegodon. Fossils
of the pongid genus Gigantopithecus, the largest known primate of all time, were
previously known to occur only in China, India, and Vietnam. Gigantopithecus has
therefore experienced a major range extension with discovery of the Java specimen. This
new specimen is referred to here as Gigantopithecus cf. G. blacki.
ANCIENT HOMO FROM TAIWAN
On January 27, 2015, Chang et al. (2015) described the fossil of an archaic Homo
mandible that had been dredged from the Penghu Channel off the west coast of Taiwan.
The specimen was recovered from the seafloor by fishermen at a depth of approximately
90 meters, at a site about 25 kilometers from the coastline. The fossil was sold to a local
antique shop. A fossil collector named Kun-Yu Tsai later purchased the fossil. Mr. Tsai
donated the specimen to the National Museum of Natural Sciences in Taichung soon after
he recognized the importance of the discovery.
Although in their description of the fossil Chang et al. (2015) declined to give the
creature a new species name, it is true that important ancient higher primate species have
been established on far less extensive fossil material. For example, Gigantopithecus
blacki von Koenigswald, 1935 was named from isolated teeth (von Koenigswald 1935,
1952). It seems evident that the new Taiwan fossil, referred to as "Penghu Man" or
Penghu 1, represents a distinctive new hominoid species. It is here named Homo
tsaichangensis n. sp. to honor both the fossil collector who donated the fossil to the
museum in Taichung and the senior author of the initial scientific report on the find
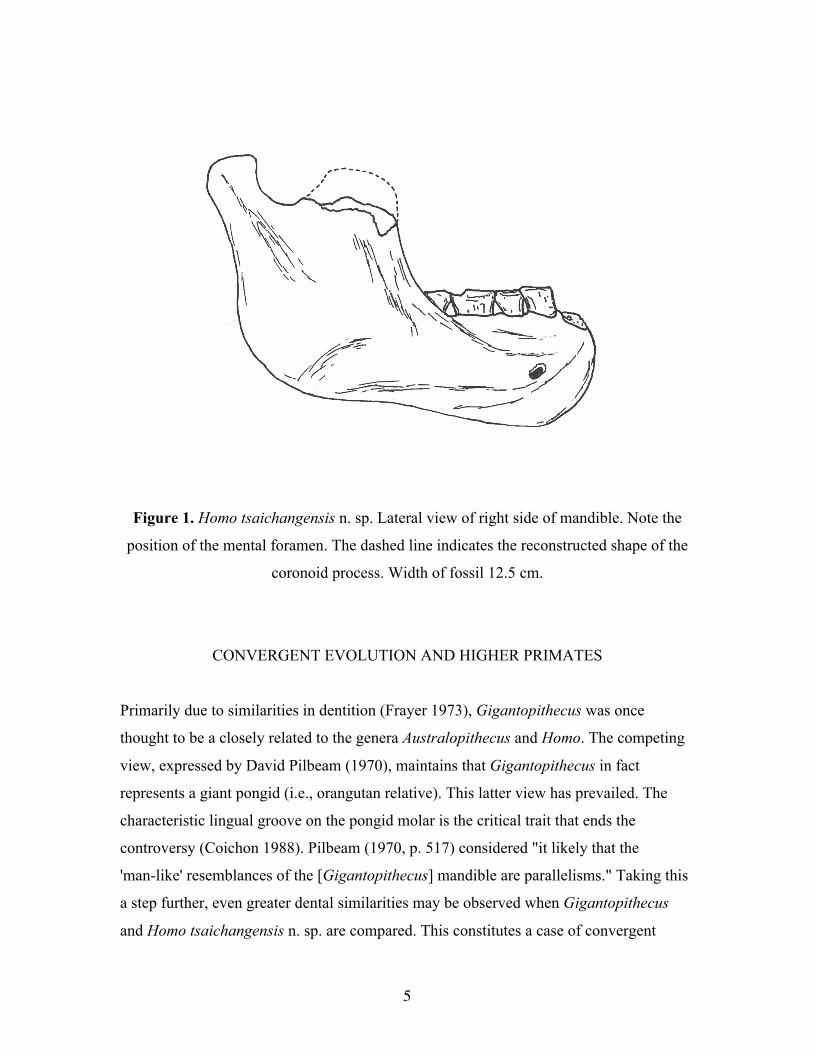
(Figure 1).
5
Figure 1. Homo tsaichangensis n. sp. Lateral view of right side of mandible. Note the
position of the mental foramen. The dashed line indicates the reconstructed shape of the
coronoid process. Width of fossil 12.5 cm.
CONVERGENT EVOLUTION AND HIGHER PRIMATES
Primarily due to similarities in dentition (Frayer 1973), Gigantopithecus was once
thought to be a closely related to the genera Australopithecus and Homo. The competing
view, expressed by David Pilbeam (1970), maintains that Gigantopithecus in fact
represents a giant pongid (i.e., orangutan relative). This latter view has prevailed. The
characteristic lingual groove on the pongid molar is the critical trait that ends the
controversy (Coichon 1988). Pilbeam (1970, p. 517) considered "it likely that the
'man-like' resemblances of the [Gigantopithecus] mandible are parallelisms." Taking this
a step further, even greater dental similarities may be observed when Gigantopithecus
and Homo tsaichangensis n. sp. are compared. This constitutes a case of convergent

6
evolution that reflects similarities in dietary preferences between the two species.
Consider that the ratios of the maximum value of C1/M1 (width measured along the
predominant zahnreihe for each tooth in question) are similar between female G. blacki
and H. tsaichangensis n. sp., with values of C1/M1 = 0.875 and C1/M1 = 0.789,
respectively.
The major range extension of Gigantopithecus across the Sunda Shelf into Java,
and the range extension of an archaic species of Homo westward into Taiwan, confirms
the observation that a huge paleogeographic province (Teilhard 1930) extended from
Java, across the Sunda Shelf and up the southeastern Asian coast into Taiwan during
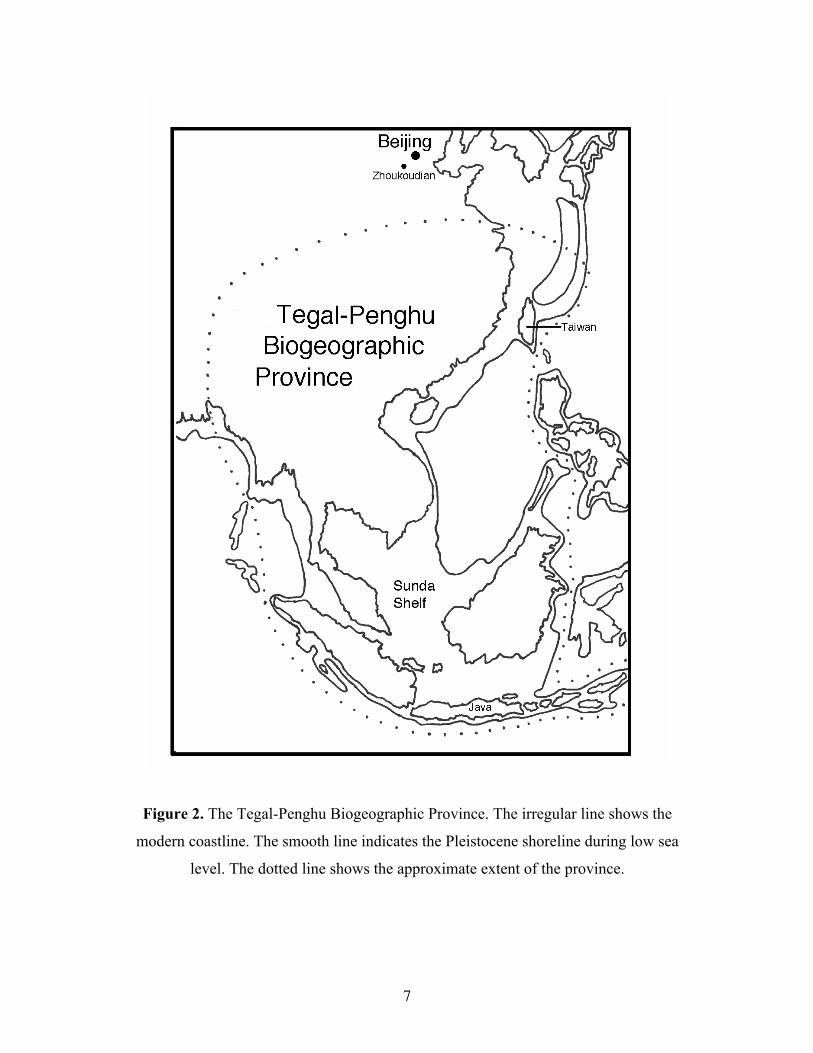
Pleistocene episodes of lowered sea level. This province, here called the Tegal-Penghu
Biogeographic Province (Figure 2), was populated by Stegodon and other distinctive
Pleistocene mammals. The province also hosted a diverse group of higher primates
including Gigantopithecus, Homo erectus and Homo tsaichangensis n. sp. The
Tegal-Penghu Biogeographic Province may be co-extensive with the biogeographic range
occupied by the Stegodon-Ailuropoda faunal assemblage (Zeitoun et al. 2005).
7
Figure 2. The Tegal-Penghu Biogeographic Province. The irregular line shows the
modern coastline. The smooth line indicates the Pleistocene shoreline during low sea
level. The dotted line shows the approximate extent of the province.

8
Based on its presumed co-occurrence with the hyaenid Crocuta crocuta ultima (an index
taxon known to have lived 500,000-250,000 years ago based on its occurrence at
Zhoukoudian locality 1 in northern China; Teilhard and Young 1929; Teilhard 1934; Pei
1934; Kahlke and Chow 1961; Chang et al. 2015) and the likelihood that the H.
tsaichangensis n. sp. mandible was deposited during a period of low sea level that
occurred 240,000-280,000 years ago, we may infer that H. tsaichangensis n. sp. lived
approximately 250,000 years ago and was thus contemporary with Gigantopithecus.
Gigantopithecus is known to have had a diet rich in bamboo, and this plant may also have
constituted part of the diet of H. tsaichangensis n. sp. In accordance with this inference,
Chang et al. (2015, p. 7-8) note that H. tsaichangensis n. sp. might have evolved "from a
gracile-jawed local H. erectus population, but this is contrary to the general trend of
dentognathic reduction in Pleistocene Homo." Thus, the dietary requirements of H.
tsaichangensis n. sp. probably explain the unexpected reversal in the evolutionary trend
of dentognathic reduction.

9
SYSTEMATIC PALEONTOLOGY
Class Mammalia
Order Primates
Family Hominidae
Tribe Hominini
Subtribe Hominina
Genus Homo
Homo tsaichangensis n. sp.
(Fig. 1)
Synonymy:
"Penghu 1 mandible", Chang et al., 2015
Type specimen. Catalog Number F051911 [Access Number NMNS006655], National
Museum of Natural Science, Taichung, Taiwan.
Diagnosis. Large teeth (anterior row) with long incisors and premolars. Ramus and
corpus mandibulae relatively low in height. Corpus mandibulae robustly thick and
relatively low. Incisal part of jaw is wide; condyle very small. Chin absent; only a single
mental foramen. Root morphology of M2 unique, developing an extra root between the
lingual projections of the distal and mesial roots. M3 absent.
Remarks. This species is known from a right mandible that preserves both the symphysis
and five teeth (C1, P3, P4, M1, M2). The lack of M3 appears to be a species characteristic
in H. tsaichangensis n. sp. The coronoid process is broken. The dashed line in Fig. 1
shows a reconstruction of the coronoid process. Even with the end of the coronoid
process broken off, it seems clear that the H. tsaichangensis n. sp. coronoid process was
upright, much more like the coronoid process of H. neanderthalensis (Gibraltar) than like
the coronoid process of H. erectus (Zhoukoudian) that curves forward at its tip.

10
The jaw ramus in Homo tsaichangensis n. sp. slants back posteriorly at a greater incline
(30°) than is typical for either Homo erectus (Zhoukoudian; 15°) or Homo
neanderthalensis (Gibraltar; 20°). Although Homo tsaichangensis n. sp. might be
compared to Hexian (H. erectus) from China's Anhui Province, I agree with Chang et al.
(2015) that it is unlikely that H. erectus was the sole species (albeit with clinal variation)
of the genus Homo in the Tegal-Penghu Biogeographic Province.
Homo tsaichangensis n. sp. has a considerably more prognathous aspect than
either Homo erectus (Zhoukoudian) or Homo neanderthalensis (Gibraltar), and is more
closely comparable to both male and female specimens of Gigantopithecus blacki in this
regard. H. tsaichangensis n. sp. shares with H. neanderthalensis (Gibraltar) a downward
expansion of the lower edge of the jaw directly underneath the molars and premolars. In
H. tsaichangensis n. sp., however, the point of greatest expansion (i.e., where the lower
jaw is locally greatest in height) is between P3 and M1, whereas in H. neanderthalensis
(Gibraltar) this point is closer to the M1-M2 junction.
Interestingly, the mental foramen in H. neanderthalensis (Gibraltar) is just
slightly posterior of the premolar-molar junction, whereas in H. tsaichangensis n. sp. the
mental foramen is located just beneath the junction of the two premolars, in accord with
the more prognathous character of the H. tsaichangensis n. sp. jaw. Significantly, H.
tsaichangensis n. sp. shares this feature (placement of the mental foramen) in common
with male Gigantopithecus blacki, where the mental foramen is directly beneath (or just
slightly anterior to) the junction between the two premolars. In contrast, female
Gigantopithecus blacki develops the mental foramen directly underneath P4. Thus we see
a progression as follows in the placement of the mental foramen in these chinless or
weak-chin mandibles going from the most anterior placement to the most posterior
placement:
G. blacki male anterior to P3-P4 junction
H. tsaichangensis n. sp. P3-P4 junction
G. blacki female P4
H. neanderthalensis (Gibraltar, Zafarraya) just posterior to P4-M1 junction

11
Therefore, Homo tsaichangensis n. sp. with regard to this character falls intermediate
between male and female Gigantopithecus blacki, a relationship that clearly indicates that
H. tsaichangensis n. sp. belongs to a new and previously undescribed species of Homo,
one that is homoplasic with respect to Gigantopithecus.
Distribution and age. Pleistocene, approximately 250,000 years old. Stratigraphic
position unknown, as a marine dredge recovered the only known specimen. The
depositional environment may have been an estuarine, fluvial or lacustrine environment.
Etymology. The new species is named for Kun-Yu Tsai and Dr. Chun-Hsiang Chang.
ACKNOWLEDGEMENTS
I wish to thank S. Wheatley for assistance with this research.
BIBLIOGRAPHY
Chang, C.-H., Y. Kaifu, M. Takai, R. T. Kona, R. Grün, S. Matsu'ura, L. Kinsley and L.-K. Lin.
2015. The first archaic Homo from Taiwan. Nature Communications
DOI:10.1038/ncomms7037.
Ciochon, R. L. 1988. Gigantopithecus: The king of all apes. Animal Kingdom 91(2):32-39.
Cranston, P. S., F.-T. Krell, K. Walker and D. Hewes. 2015. Wiley's Early View constitutes valid
publication for date-sensitive nomenclature. Systematic Entomology 40:2-4.
Frayer, D. W. 1973. Gigantopithecus and its relationship to Australopithecus. American Journal
of Physical Anthropology 39:413-426.
Kahlke, H. D. and B. Chow. 1961. A summary of stratigraphical and paleontological
observations in the lower layers of Choukoutien, Locality 1, and on the chronological
position of he site. Vertebr. Palasiat. 3:212-240.
Maryono, A. 2014. Fossils of rare, ancient animals found in Tegal. The Jakarta Post, December
1, 2014: http://www.thejakartapost.com/news/2014/12/01/fossils-rare-ancient-animals-
found-tegal.html.

12
Pei, W. 1934. On the Carnivora from Locality 1 of Choukoutien. Palaeontol. Sin. Series C 8:1-
216.
Pilbeam, D. 1970. Gigantopithecus and the origins of Hominidae. Nature 225:516-519.
Teilhard de Chardin, P. 1926. How and where to search [for] the oldest man in China. Bulletin of
the Geological Society of China 5(3-4):201-206.
Teilhard de Chardin, P. 1930. Quelques observations biogéographiques en Chine. Comptes Rend.
somm. Soc. de Biogéographie 7(60):94-96.
Teilhard de Chardin, P. 1934. Les fouilles préhistoriques de Péking. Rev. quest. scientif.
25(8):181-193.
Teilhard de Chardin, P. and C. C. Young. 1929. Preliminary report on the Chou-Kou-Tien
fossiliferous deposits. Bulletin of the Geological Society of China 8(3):173-202.
von Koenigswald, G. H. R. 1931. Fossielen uit Chineesche Apotheken in West-Java. De
Mijningenieur 11:189-193.
von Koenigswald, G. H. R. 1935. Eine fossile Säugetierfauna mit Simia aus Südchina. Proc.
Sect. Sci., K. Akad. Wetensch. Amsterdam 38(8):872-879.
von Koenigswald, G. H. R. 1952. Gigantopithecus blacki von Koenigswald, a giant fossil
hominoid from the Pleistocene of Southern China. Anthropological Papers of the
American Museum of Natural History 43:291-325.
Zeitoun, V., A. Seveau, H. Forestier, H. Thomas, A. Lenoble, F. Laudet, P.-O. Antoine, R.
Debruyne, L. Ginsburg, P. Mein, C. Winayalai, N. Chumdee, T. Doyasa, A. Kijngam and
S. Nakbunlung. 2005. Découverte d'un assemblage faunique à Stegodon-Ailuropoda dans
une grotte du Nord de la Thaïlande (Ban Fa Suai, Chiang Dao). C. R. Paleovol 4:255-
264.
Copyright © 2022 FDOKUMEN