
Hitchhiker’ polynoid polychaetes in cold deep waters and their potential influence on benthic soft...
9
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234 Antarctic Science 22(4), 399–407 (2010) & Antarctic Science Ltd 2010 doi:10.1017/S0954102010000210 ‘Hitchhiker’ polynoid polychaetes in cold deep waters and their potential influence on benthic soft bottom food webs STEFANO SCHIAPARELLI 1 , MARIA CHIARA ALVARO 2 , JEHNS BOHN 3 and GIANCARLO ALBERTELLI 1 1 Dipartimento per lo Studio del Territorio e delle sue Risorse (Dip.Te.Ris.), Universita ` di Genova, C.so Europa 26, Genova I-16132, Italy 2 Museo Nazionale dell’Antartide (MNA), Universita ` di Genova, Viale Benedetto XV n85, Genova I-16132, Italy 3 Zoologische Staatssammlung Mu ¨nchen, Mu ¨nchhausenstrasse 21, Mu ¨nchen D-81247, Germany [email protected] Abstract: We describe a new association for Antarctica, involving an holothuroid host, Bathyplotes bongraini Vaney, 1914, and a parasitic polynoid polychaete, Eunoe opalina McIntosh, 1885, which lives on the host body. Both species have never been recorded in the study area, the Ross Sea. The ecological definition of this partnership is difficult to assess, being a mix of phoresis, protective association, parasitism and, possibly, kleptocommensalism. Eunoe opalina emerges also as a true predator, ingesting several food items that do not belong to the diet of Bathyplotes. We compare this association with analogous examples known from shallow tropical environments as well as bathyal and abyssal depths. Given the conspicuous similarities between the deep water and high latitude examples of this kind of association, a possible common origin is hypothesized. Although the role of such a kind of parasitic relationships in Antarctic communities remains to be fully evaluated, it seems evident that, at high latitudes, where trophic levels are simplified and food webs do not have much redundancy, the impact of such a ‘multitasking’ predator-parasite as E. opalina might be of a greater magnitude than its shallow water tropical counterpart. Received 5 December 2009, accepted 4 March 2010 Key words: Antarctica, Holothuroidea, parasitism, Polychaeta, Ross Sea, ‘symbiotic’ association Introduction Marine ‘partnerships’ or ‘symbiotic’ associations (sensu De Bary 1878) have only recently been recognized to play a very important role in shaping marine communities, since they may affect their structure to an extent as important as predation or physical disturbance (Hay et al . 2004). In particular, parasitism may increase connectance (links/species; Dunne et al . 2002) and affect nestedness, chain length and linkage density of food webs dramatically (Lafferty et al . 2006), and may play a major role, with consequences that have still to be quantified, in most marine environments. Historically, the study of ‘symbioses’ has been focused on reef communities, where these kind of partnerships are widespread and abundant, whilst examples occurring in temperate and cold areas, especially polar waters, have been little studied. Nevertheless, in recent years, partnerships involving Antarctic marine invertebrates have received increasing interest with the result that a provisional list of different associations occurring in this area now numbers 23–25 different examples, mainly shifted towards parasitism, with polychaetes and molluscs being the commonest symbionts (Schiaparelli et al. 2007 and unpublished data). Among the polychetous annelids, the setting-up of close associations with other marine invertebrates is a rather common phenomenon at all latitudes (Martin & Britayev 1998), especially in the case of Polynoidae. This family of 1666 species (Fauchald & Barnich 2009a), mainly of generalist carnivores (Fauchald & Jumars 1979), also numbers several examples of species living as ‘symbionts’ on different invertebrate hosts (e.g. Pettibone 1982, 1993, Britayev et al . 2003). Among polychaetes, 55% of commensal polychaete species belong to the Polynoidae (Martin & Britayev 1998). In the Southern Ocean, Polynoidae are well represented with about 82 species known (Fauchald & Barnich 2009b) out of a total of 677 species of polychaetes described so far for this area (RAMS 2009). For most of these polynoids, however, no ecological data are available and to date only Polynoe thouarellicola Hartmann-Schro ¨der, 1989, has been reported as a ‘commensal’, having been found in association with gorgonians of the genus Thouarella (Hartmann-Schro ¨der 1989). In the present contribution, we describe an Antarctic association from the Ross Sea involving the polychaete Eunoe opalina McIntosh, 1885 (Phyllodocida: Polynoidae) and the holothurian Bathyplotes bongraini Vaney, 1914 (Holothuroidea: Synallactidae), which have never been reported in association before, probably due to the well known bias of destructive sampling, as trawling or dredging, which may separate symbionts and hosts. The partnership between polynoids and holothuroids has been previously reported from shallow tropical waters, where it has been extensively studied from an ecological point of view (Britayev & Zamishliak 1996, Britayev et al. 399
Transcript of Hitchhiker’ polynoid polychaetes in cold deep waters and their potential influence on benthic soft...

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
Antarctic Science 22(4), 399–407 (2010) & Antarctic Science Ltd 2010 doi:10.1017/S0954102010000210
‘Hitchhiker’ polynoid polychaetes in cold deep waters and theirpotential influence on benthic soft bottom food webs
STEFANO SCHIAPARELLI1, MARIA CHIARA ALVARO2, JEHNS BOHN3 and GIANCARLO ALBERTELLI1
1Dipartimento per lo Studio del Territorio e delle sue Risorse (Dip.Te.Ris.), Universita di Genova, C.so Europa 26,
Genova I-16132, Italy2Museo Nazionale dell’Antartide (MNA), Universita di Genova, Viale Benedetto XV n85, Genova I-16132, Italy
3Zoologische Staatssammlung Munchen, Munchhausenstrasse 21, Munchen D-81247, Germany
Abstract: We describe a new association for Antarctica, involving an holothuroid host, Bathyplotes
bongraini Vaney, 1914, and a parasitic polynoid polychaete, Eunoe opalina McIntosh, 1885, which lives on
the host body. Both species have never been recorded in the study area, the Ross Sea. The ecological
definition of this partnership is difficult to assess, being a mix of phoresis, protective association, parasitism
and, possibly, kleptocommensalism. Eunoe opalina emerges also as a true predator, ingesting several food
items that do not belong to the diet of Bathyplotes. We compare this association with analogous examples
known from shallow tropical environments as well as bathyal and abyssal depths. Given the conspicuous
similarities between the deep water and high latitude examples of this kind of association, a possible
common origin is hypothesized. Although the role of such a kind of parasitic relationships in Antarctic
communities remains to be fully evaluated, it seems evident that, at high latitudes, where trophic levels are
simplified and food webs do not have much redundancy, the impact of such a ‘multitasking’ predator-parasite as
E. opalina might be of a greater magnitude than its shallow water tropical counterpart.
Received 5 December 2009, accepted 4 March 2010
Key words: Antarctica, Holothuroidea, parasitism, Polychaeta, Ross Sea, ‘symbiotic’ association
Introduction
Marine ‘partnerships’ or ‘symbiotic’ associations (sensu De
Bary 1878) have only recently been recognized to play a very
important role in shaping marine communities, since they may
affect their structure to an extent as important as predation or
physical disturbance (Hay et al. 2004). In particular, parasitism
may increase connectance (links/species; Dunne et al. 2002)
and affect nestedness, chain length and linkage density of food
webs dramatically (Lafferty et al. 2006), and may play a major
role, with consequences that have still to be quantified, in most
marine environments.
Historically, the study of ‘symbioses’ has been focused on
reef communities, where these kind of partnerships are
widespread and abundant, whilst examples occurring in
temperate and cold areas, especially polar waters, have been
little studied. Nevertheless, in recent years, partnerships
involving Antarctic marine invertebrates have received
increasing interest with the result that a provisional list of
different associations occurring in this area now numbers
23–25 different examples, mainly shifted towards parasitism,
with polychaetes and molluscs being the commonest
symbionts (Schiaparelli et al. 2007 and unpublished data).
Among the polychetous annelids, the setting-up of close
associations with other marine invertebrates is a rather common
phenomenon at all latitudes (Martin & Britayev 1998),
especially in the case of Polynoidae. This family of 1666
species (Fauchald & Barnich 2009a), mainly of generalist
carnivores (Fauchald & Jumars 1979), also numbers several
examples of species living as ‘symbionts’ on different
invertebrate hosts (e.g. Pettibone 1982, 1993, Britayev et al.
2003). Among polychaetes, 55% of commensal polychaete
species belong to the Polynoidae (Martin & Britayev 1998).
In the Southern Ocean, Polynoidae are well represented
with about 82 species known (Fauchald & Barnich 2009b)
out of a total of 677 species of polychaetes described so far
for this area (RAMS 2009). For most of these polynoids,
however, no ecological data are available and to date only
Polynoe thouarellicola Hartmann-Schroder, 1989, has been
reported as a ‘commensal’, having been found in association
with gorgonians of the genus Thouarella (Hartmann-Schroder
1989).
In the present contribution, we describe an Antarctic
association from the Ross Sea involving the polychaete
Eunoe opalina McIntosh, 1885 (Phyllodocida: Polynoidae)
and the holothurian Bathyplotes bongraini Vaney, 1914
(Holothuroidea: Synallactidae), which have never been
reported in association before, probably due to the well
known bias of destructive sampling, as trawling or
dredging, which may separate symbionts and hosts.
The partnership between polynoids and holothuroids has
been previously reported from shallow tropical waters,
where it has been extensively studied from an ecological
point of view (Britayev & Zamishliak 1996, Britayev et al.
399

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
1999, Britayev & Lyskin 2002, Lyskin & Britayev 2005),
and for Californian temperate shallow water communities
(e.g. Dimock & Davenport 1971). A few more scattered
examples involving these partners have been described for
the deep waters of the Atlantic Ocean (e.g. Wesenberg-
Lund 1941, Kirkegaard & Billet 1980). In each of these
examples different species of polynoids and holothuroids
are involved, establishing relationships which only partially
overlap, if their ecological traits are taken into account.
Here we describe the southernmost case of such an
association, the first one for Antarctica, and review the
available literature comparing this example with the tropical
shallow water and deep water, non-Antarctic, counterparts.
The possible ecological role of these ‘symbiotic’ associations,
generally overlooked in deep waters environments, is
discussed in the light of the potential influence they may
have on benthic soft bottom food webs.
Methods
Polychaetes and holothurians were collected by two
different cruises in the Antarctic waters by the RV
Tangaroa. The first one took place in the Ross Sea, from
January–March 2004, sampling between 658S and 758S and
from 65–1570 m. The second one occurred between
February and March 2008, sampling from 668S and 768S
and from 150–3553 m. Both cruises used Orange Roughy
Trawls, Beam Trawls and dredges.
Immediately after collection, samples were carefully
observed and polychaetes photographed in situ on the
Table I. List of sampling stations in the Balleny Islands and in the Ross Sea areas.
Cruise St. Site Holothuroid species Latitude S Longitude E Depth (m) Date
BioRoss Cruise 218 Balleny Islands Bathyplotes bongraini 67815'13'' 164852'30'' 348 03/03/04
(TAN0402) (Sturge)
BioRoss Cruise 249 Balleny Islands Bathyplotes bongraini 66833'30'' 163801'04'' 555 05/03/04
(TAN0402) (Young)
IPY-CAML Cruise 56 Ross Sea shelf Bathyplotes bongraini 75837'58'' 169850'59'' 531 14/02/08
(TAN0802)
IPY-CAML Cruise 169 Ross Sea shelf Laetmogone sp. 71823'03'' 174844'07'' 2202 25/02/08
(TAN0802)
Fig. 1. Different views of the association
between Bathyplotes bongraini and
Eunoe opalina. a. B. bongraini
specimen photographed in situ
(Tangaroa 2008 Cruise, station 56).
b. Ventral view of a freshly collected
B. bongraini specimen carrying
E. opalina next to its mouth, between
the fields of tube feet. c. Particular
showing the cephalic end of
B. bongraini with E. opalina lying into
its ‘niche’ in the host tegument. The
polychaete ventral side is in direct
contact with the holothurian’s one.
d. E. opalina specimen: ventral view
(left) and dorsal view (right).
400 STEFANO SCHIAPARELLI et al.
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
host, in order to document their exact position on the
holothuroid. After fixation in 70% ethanol, every specimen
of E. opalina was dissected and its stomach isolated.
Stomachs were then dipped for half a day in a solution of
1% Rose Bengal, rinsed three times in water and their
contents extracted, dehydrated in increasing alcohol
concentrations (80%–90%–100%) and placed on a glass
slide for microscope examination. The dominance method
(Holden & Raitt 1974) was adopted to quantify the
abundance of food items.
As regards the host, B. bongraini, skin fragments from
three different parts (oral zone, central zone, anal zone) of
the holothurian body were cut both from dorsal and ventral
sides. Ossicles of the body wall were then obtained by
digesting the tissue with the proteolitic enzyme Tripsina
(Tiago et al. 2005). The ossicles were then rinsed in distilled
water, dehydrated and mounted on a glass slide with Eukitt.
SEM observations of chaetae, polychaete stomach content
and holothuroid skin and ossicles were performed with a
Leica Stereoscan 440, after dehydratation of samples with
HMDS (hexamethyldisilazane) (Nation 1983).
Results
Both cruises together provided a total of 86 specimens of
B. bongraini which carried 26 specimen of E. opalina.
Since we found only one polychaete per holothurian, the
estimated infestation rate is about 30% of the holothuroid
population. Due to logistic restrictions, the evaluation of
the infestation rate has been possible only in selected
stations (see Table I) and, therefore, the distribution of this
association may be much wider than herein considered.
Both species are here reported for the first time in the
Ross Sea. Bathyplotes bongraini (Fig. 1a) was previously
known from western Antarctica (Weddell Sea, between
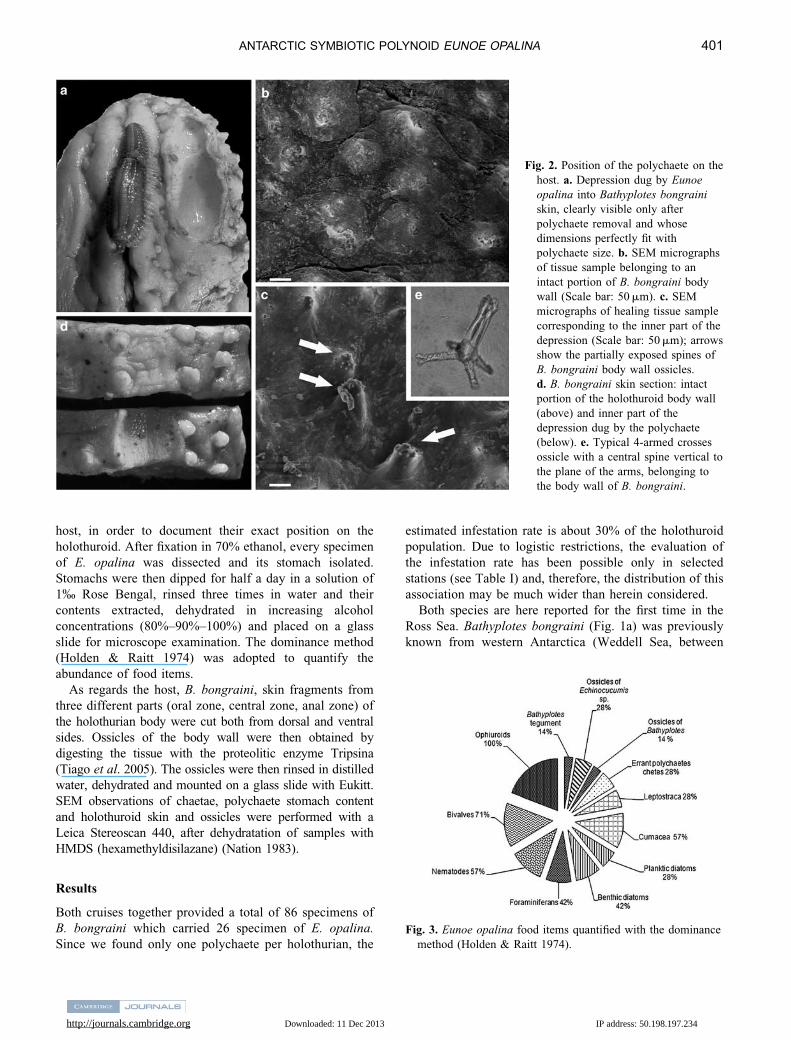
Fig. 2. Position of the polychaete on the
host. a. Depression dug by Eunoe
opalina into Bathyplotes bongraini
skin, clearly visible only after
polychaete removal and whose
dimensions perfectly fit with
polychaete size. b. SEM micrographs
of tissue sample belonging to an
intact portion of B. bongraini body
wall (Scale bar: 50 mm). c. SEM
micrographs of healing tissue sample
corresponding to the inner part of the
depression (Scale bar: 50 mm); arrows
show the partially exposed spines of
B. bongraini body wall ossicles.
d. B. bongraini skin section: intact
portion of the holothuroid body wall
(above) and inner part of the
depression dug by the polychaete
(below). e. Typical 4-armed crosses
ossicle with a central spine vertical to
the plane of the arms, belonging to
the body wall of B. bongraini.
Fig. 3. Eunoe opalina food items quantified with the dominance
method (Holden & Raitt 1974).
ANTARCTIC SYMBIOTIC POLYNOID EUNOE OPALINA 401

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
245–465 m; Antarctic Peninsula, 250 m) and eastern
Antarctica (Prydz Bay, 120–768 m; off Kemp Land,
603 m) (O’Loughlin 2002, O’Loughlin et al. 2009), while
E. opalina was known for the Antarctic Peninsula and the
Scotia Ridge (Fauchald 2009).
In all the cases we have examined, E. opalina (Fig. 1d)
was exclusively found on the host’s ventral side, mainly
next to the mouth of the holothuroid, between tube feet
fields (Fig. 1b & c) (in one case only it was on the opposite
side), where it was lying in a depression excavated into the
holothurian tegument. This cavity remained clearly visible
even after polychaete removal, perfectly fitting the worm’s
shape and size (Fig. 2a).
SEM observation of the host’s tissue samples, taken from
an intact portion of the holothurian body (Fig. 2b) and from
the inner part of the depression dug by the polychaete
(Fig. 2c), respectively, showed the presence of a different skin
texture in the two areas. It is clear that the polychaete activity
has somehow removed part of the tegument, almost exposing
(Fig. 2c, arrows) the 4-armed crosses ossicles (Fig. 2e), typical
of B. bongraini tegument, that emerged beneath the thin,
perhaps healing, tissue. Where the tegument is intact, the
ossicles do not protrude and are completely embedded.
The stomach content analysis of seven polychaetes
(dominance method, according to Holden & Raitt 1974)
(Fig. 3) allowed the identification of a variety of prey items
(Fig. 4a), as: ossicles of Bathyplotes (14%) and other
holothuroid species (Echinocucumis sp.) (28%) (Fig. 4d),
bivalves (71%), foraminiferans (42%), benthic (42%) and
planktic (28%) diatoms, ophiuroids (100%), chaetae of
errant polychaetes (28%), microcrustaceans fragments
(Fig. 4b & c) belonging to Cumacea (57%) and to
Leptostraca (28%) and nematodes (57%).
Discussion
Representatives of most marine phyla are known to live in
close association with echinoderms, establishing different
typologies of partnerships, ranging from phoresis to parasitism
(e.g. Barel & Kramers 1977, Jangoux 1987a, 1987b).
Among echinoderms, Holothuroidea harbours the highest
number of symbionts, reaching a total of 145 species
belonging to ten different Phyla (Jangoux 1987b, table I,
p. 223). These associated organisms may live inside the
holothuroid body, as in fishes (Carapidae) and crabs
(Pinnotheriodae), or outside and/or transdermically, as in
the case of bivalves (Galeommatoidea) and gastropods
(Eulimidae) (Waren 1983, Jangoux, 1987a, Ng & Manning
2003, Middelfart & Craig 2004, Parmentier & Vanderwalle
2005). On the whole, it has been estimated that, in temperate
and tropical waters, about 40 different species of holothuroids
are the hosts in 50 recognized cases of ‘symbioses’ involving
polychaetes (Martin & Britayev 1998).
To date, the best known association between a
polychaete and its holothuroid host is that of the polynoid
Fig. 4. SEM micrographs of Eunoe
opalina stomach content. a. Overview
of the main food items (Scale bar:
100 mm). b,c. Particular of
microcrustacean fragments.
d. Echinocucumis sp. ossicle
(Scale bar: 20 mm).
402 STEFANO SCHIAPARELLI et al.
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
Gastrolepidia clavigera Schmarda, 1861, a shallow water
‘polyxenous’ species from the Indo-West Pacific region,
which is known to live on 13 different holothuroids of the
families Stichopodidae and Holothuriidae (Martin &
Britayev 1998). Gastrolepidia clavigera has been studied
in detail from an ecological point of view and, therefore,
represents a good reference example to be compared with
E. opalina, the most ‘extreme’ case of such a kind of
relationship, occurring in the cold Antarctic waters.
The first macroscopic difference between these two
polychaetes is that E. opalina was found in only one
specimen per host, always placed ventrally next to the
mouth of the holothurian’s body, where it lies in a deep
depression produced by the polychaete itself. Gastrolepidia
clavigera may be present with up to four specimens per host,
usually living on the holothurian dorsal side (Britayev &
Zamishliak 1994, 1996, Britayev et al. 1999) where they may
form temporary depressions that disappear after polychaete
removal (Britayev & Zamishliak, 1996). Gastrolepidia
clavigera is also known to crawl into the mouth or cloaca of
the holothurian host in case of threats (Britayev & Zamishliak
1994, 1996), a behaviour not observed in E. opalina, although
it has to be taken into account the fact that no in vivo long-term
observations are available for this species.
A comparison between the stomach contents of the two
polychaetes reveals that both species have a wide variety of
food items, which almost perfectly overlaps from a
qualitative point of view, including: holothuroid ossicles,
crustacean fragments, multicellular and unicellular algae,
chaetae and jaws of polychaetes (other than G. clavigera
or E. opalina), detritus, foraminifera and fragments of
the shells of bivalves (Britayev & Lyskin 2002 for
G. clavigera; present data for E. opalina, Fig. 3).
Unfortunately, it is not possible to make a direct,
quantitative comparison of food items due to the different
methodologies used but some very important differences
can be highlighted about the origin of the holothuroid
ossicles found in the stomachs. In G. clavigera, the stomach
content analysis clearly indicates that the polychaete
feeds on the skin of the host from which it has been
collected (Britayev & Lyskin 2002). Eunoe opalina
contains B. bongraini ossicles and another distinctive kind
of ossicles, clearly not belonging to the host species. These
ossicles fall within the morphological range of the genus
Echinocucumis and may belong to a not yet described species
(O’Loughlin, personal communication 2009).
This finding opens several scenarios, the most obvious one
being that of a host switch of E. opalina during its lifetime.
However, considering the size of the known Echinocucumis
species, which are really small holothurians, almost comparable
to E. opalina in average size, a permanent association with this
holothuroid genus can be excluded. The possibility that these
ossicles could already be present in the sediment as loose items
and that these are passively ingested by the polychaete during
its feeding activity seems highly improbable, as these have
been found in quantities in the stomachs and always in very
good condition (not damaged, etched or worn). Most
interestingly, the same ossicle type was also found in
stomachs of E. opalina specimens from the Weddell Sea
(unpublished data). It is therefore possible that E. opalina
undertakes opportunistic predation on other holothuroid species
(i.e. Echinocucumis), if these are somehow detected by the
polychaete during the host’s grazing activity on the bottom.
The fact that Echinocucumis ossicles have been found in
E. opalina stomachs both from the Ross Sea and the Weddell
Sea, possibly indicates that the environmental settings in which
B. bongraini specimens is found are very similar, despite the
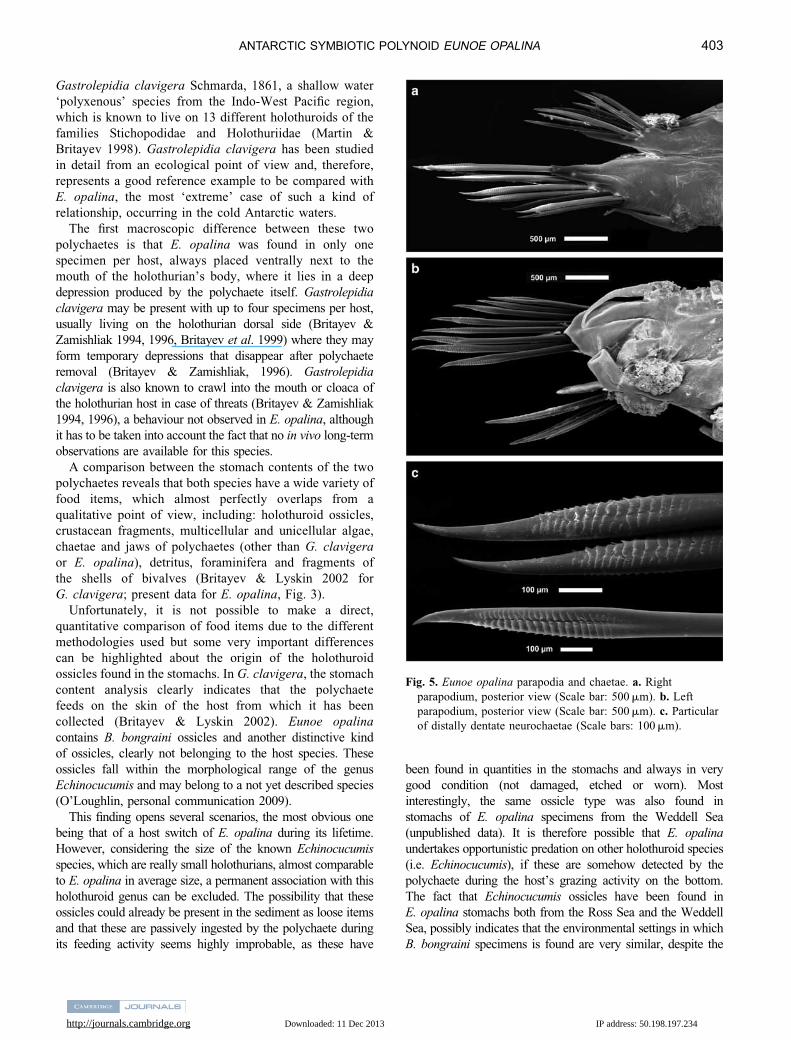
Fig. 5. Eunoe opalina parapodia and chaetae. a. Right
parapodium, posterior view (Scale bar: 500 mm). b. Left
parapodium, posterior view (Scale bar: 500 mm). c. Particular
of distally dentate neurochaetae (Scale bars: 100 mm).
ANTARCTIC SYMBIOTIC POLYNOID EUNOE OPALINA 403
http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
distance between the two areas, and have almost the same
species composition.
Although belonging to different genera, it is clearly evident
that G. clavigera and E. opalina are adapted to live in
association with holothuroids, both showing morphological
peculiarities linked to their symbiotic mode of life. As
documented by Britayev & Zamishlyak (1994), G. clavigera
has specially designed attaching structures, called ‘ventral
sucker-like lobes’. These features have probably evolved to
enable the adherence of the polychaete to several hosts,
accounting for the potentially different ‘tegument properties’
encountered from time to time. In contrast, E. opalina does
not have these structures, but shows distally dentate spines
(Fig. 5a–c), known to occur in other symbiotic polynoids
(Martin & Britayev 1998).
In summary, G. clavigera is a ‘polyxenous’ polychaete
with several host species, while E. opalina is a ‘monoxenous’
polychaete, always associated with B. bongraini. On the other
hand, it is clear that E. opalina has a wider prey range
including other holothurian species with which no ‘symbiotic’
partnerships are established.
From an ecological point of view, these differences are
rather intriguing and might be possibly related to the
ecosystems in which these associations have been established,
and not only to the taxonomic and morphological differences
between the two symbionts.
The latitudinal and bathymetric distribution of the
documented associations involving polynoids and holothuroids
show that most of them occur in the Northern Hemisphere,
at intermediate latitudes and on the continental shelf
(shallower than 200 m, Fig. 6a).
The relationship between E. opalina and B. bongraini,
even if it occurs between 348–555 m depth (Fig. 6b),
should be considered in the category of shelf examples, as
it is known that the Antarctic continental shelf is much
deeper than other continental shelves, due to the weight of
the ice cap (Anderson 1999). On the other hand, it is also
known that most Antarctic species are extremely eurybathic
and recolonized the shelf only when it again became
available after ice retreat during interglacials (Brey et al.
1996).
Several similarities between Antarctic and deep water
representatives of ‘symbiotic’ associations involving
polynoids and holothurians species can also be highlighted.
For example, the presence of a ‘scar’ on the host’s ventral
body wall clearly resembles what has already been described
for the polychaetes Harmothoe bathydomus Ditlevsen, 1917
and Eunoe laetmogonensis Kirkegaard & Billet, 1980,
respectively associated with the holothuroids Bathyplotes
natans (Sars, 1868) and Laetmogone violacea Theel, 1879
(Wesenberg-Lund 1941, Kirkegaard & Billet 1980), both
occurring in deep Atlantic waters (800–3200 m). The first
Fig. 6. Latitudinal and bathymetric distribution of the
documented associations involving polynoid polychaetes and
holothuroids worldwide distributed. Northern and southern
latitude values have been combined. a. Shelf water records
(0–200 m). b. Deepwater records (200–3700 m).
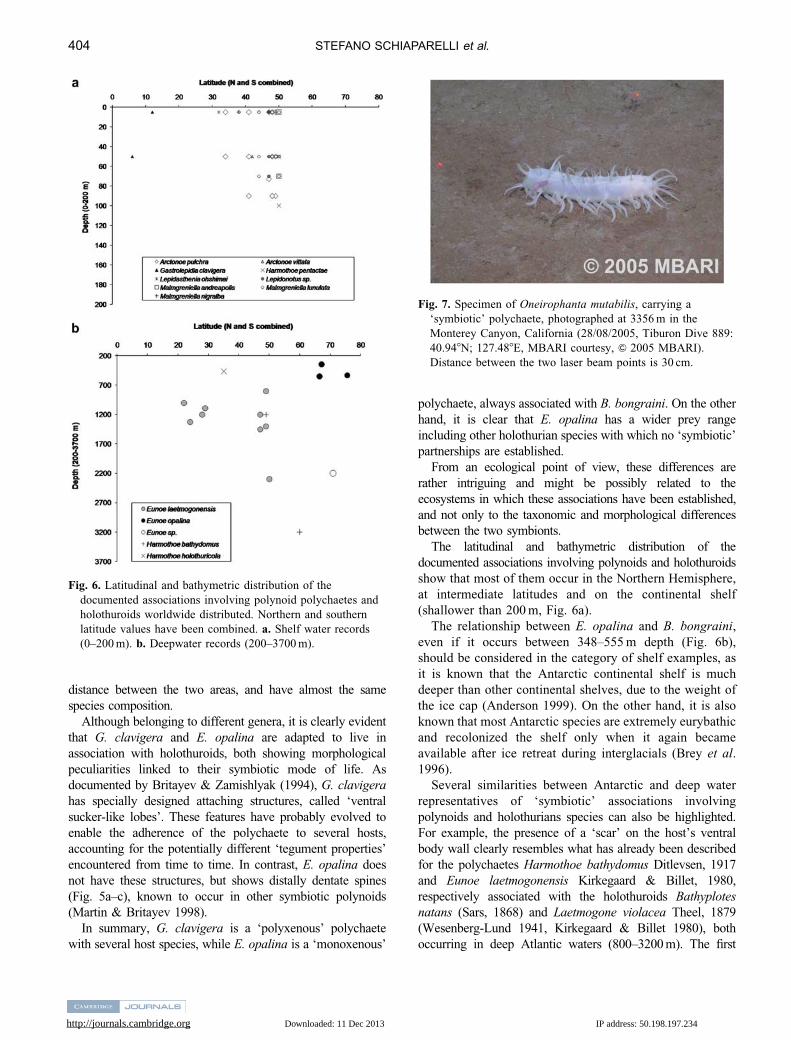
Fig. 7. Specimen of Oneirophanta mutabilis, carrying a
‘symbiotic’ polychaete, photographed at 3356 m in the
Monterey Canyon, California (28/08/2005, Tiburon Dive 889:
40.948N; 127.488E, MBARI courtesy, & 2005 MBARI).
Distance between the two laser beam points is 30 cm.
404 STEFANO SCHIAPARELLI et al.

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
example, in particular, is remarkable not only for the
similarities with the Antarctic case, but also because the
partners involved in the association are another Bathyplotes
species and a polychaete that, although now included in
Harmothoe, a genus closely allied to Eunoe, has been
considered indistinguishable from E. opalina by Ditlevsen
(1917, p. 26).
It is therefore possible that these two deep water polynoid-
holothuroid associations share a common origin with the
Antarctic one. The disjointed, bipolar distribution could be
explained in the light of the ‘thermohaline expressway’
hypothesis, according to which some Antarctic species could
have spread into deep water basins outside Antarctica,
by following the increased production of cold, hyperhaline
water masses during glacial cycles (Strugnell et al. 2008).
Independent information on phylogenetic relationships of both
polynoids and holothuroids would be required to test this
hypothesis but, unfortunately, no such studies are yet available.
Deep water polynoid-holothuroid partnerships might be
much more common and widespread than believed. Using
images collected during deepwater ROV surveys, we were
able to document two further examples. The first one refers
to a specimen of Oneirophanta mutabilis Theel, 1879,
photographed at 3356 m, off Monterey (California), with a
polynoid moving on its body (Fig. 7). This record increases
the maximum known depth limit for a polynoid associated
with an holothuroid (the previous deepest record was of
3136 m for H. bathydomus, Ditlevsen 1917), and adds a new
host record in this kind of association. A further example is
that of Laetmogone sp. (probably an undescribed species,
O’Loughlin personal communication 2009), photographed
at 2202 m in the Ross Sea. This latter species carried an
undetermined polynoid whose cephalic end was clearly
inserted within the holothuroid tentacles crown (Fig. 8).
A precise, strict definition of the relationship between
E. opalina and B. bongraini is not possible, as it appears to
be a mix of phoresis (the polychaete is carried by the host),
protective association (the polychaete is also protected from
predators by its host), parasitism (the polychaete removes and
ingests host skin and ossicles) and kleptocommensalism (due
to the possibility that some of the preys are removed from the
host mouth as documented in Fig. 8). Nevertheless, it is clear
from the variety of food items collected, and from the removal
of host tegument, that the polychaete may have both a
detrimental effect on its host and a negative impact on other
bottom dwelling invertebrates living in the same environment.
Considering the formula to calculate Connectance,
corrected to include parasites (C 5 L0/F(F 1 P), where
C 5 connectance, L0 5 number of links, F 5 free living
species and P 5 parasite species) (see Lafferty et al. 2006
for details), it emerges that the presence of E. opalina in the
loop determines an increase of connectance from 0.25
(holothuroid-detritus chain with a single link, where
detritus is contemplated as a ‘non-phylogenetic category’,
Dunne et al. 2002) to 0.33 (detritus-B. bongraini-E. opalina
chain, with two links), in accordance to what is expected
when parasites are included in food webs (Lafferty et al.
2006). Interestingly, if we take into account the double role
of E. opalina, that can be considered simoultaneously a free
living species (F) and a parasite (P), by including in the
calculation also the food items (i.e. bivalves, ophiuroids
and the other holothuroid species) which are not part of the
B. bongraini diet, the number of total links rises to five and
the connectance decreases to 0.16.
Given the major role that parasites have been demonstrated
to play on complex food webs in general (Lafferty et al.
2006), and in the light of the above calculations, it is
reasonable to suppose that in Antarctica, where trophic levels
are simplified, no ‘modern’ predators exist (Aronson & Blake
2001) and food webs do not have much redundancy, the
impact of such a ‘multitasking’ predator-parasite as E. opalina
might be even of a greater magnitude, deeply affecting
Connectance and other food web features.
Further studies dedicated to the quantification of the
effects that the ‘symbiotic’ interactions may have in
Antarctica, will enable some of the hypotheses proposed
here to be tested and will contribute to our understanding of
the ecological constraints which affect invertebrates living
at high latitudes.
Acknowledgements
We would like to acknowledge the crew of the Tangaroa
expeditions ‘BioRoss TAN0402’ and ‘IPY-CAML
TAN0802’ for logistic support during both cruises. A
particular acknowledgement is due to Anne Nina Loerz
Fig. 8. Undetermined specimen of Laetmogone sp. carrying a
symbiotic polychaete, photographed in situ at 2202 m in the
Ross Sea (IPY-CAML Cruise, TAN0802, st. 169). The head
of the polychaete (arrow) is clearly inserted within the
holothuroid oral tentacle crown. This behaviour could be
interpreted as kleptocommensalism, with the polychaete possibly
stealing food items before the holothuroid swallow them.
ANTARCTIC SYMBIOTIC POLYNOID EUNOE OPALINA 405

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
(NIWA) and Sadie Mills (NIWA) for collaboration during the
cruises. We are indebted to the New Zealand Ministry of
Fisheries (MFish) and NIWA (Wellington) for the financial
support of the cruises and related study activities. We are
extremely grateful to MBARI (www.mbari.org) for the
permission to use the picture from the Tiburon Dive 889,
and to Mark O’Loughlin (Marine Biology Section, Museum
Victoria) for the useful commentaries and suggestions. Geoff
Read (NIWA, Wellington) kindly provided the E. opalina
classification. Gaia Magioncalda and Sylvie de Sousa Pereira
contributed to laboratory analyses. Two anonymous reviewers
are acknowledged for useful comments on the manuscript.
This paper is a contribution to the multinational Latitudinal
Gradient Project and contribution #28 to the Census of
Antarctic Marine Life (CAML).
References
ANDERSON, J.B. 1999. Antarctic marine geology. Cambridge: Cambridge
University Press, 289 pp.
ARONSON, R.B. & BLAKE, D.B. 2001. Global climate change and the origin
of modern benthic communities in antarctica. American Zoologist, 41,
27–39.
BAREL, C.D. & KRAMERS, P.G. 1977. A survey of the echinoderm associates
of the north-east Atlantic area. Zoologische Verhandelingen, 156,
1–159.
BREY, T., DAHM, C., GORNY, M., KALGES, M., STILLER, M. & ARNTZ, W.E.
1996. Do Antarctic benthic invertebrates show an extended level of
eurybathy? Antarctic Science, 8, 3–6.
BRITAYEV, T.A. & ZAMISHLYAK, E.A. 1994. The biology of a commensal
polychaete Gastrolepidia clavigera (Polychaeta, Polynoidae) with
remarks on the biology of host holothurians (Holothuriidae,
Stichopodidae). Zoologicheskii Zhurnal, 73, 39–53.
BRITAYEV, T.A. & ZAMYSHLYAK, E.A. 1996. Association of the commensal
scaleworm Gastrolepidia clavigera (Polychaeta: Polynoidae) with
holothurians near the coast of South Vietnam. Ophelia, 45, 175–190.
BRITAYEV, T.A., DOIGNON, G. & EECKHAUT, I. 1999. Symbiotic
polychaetes from Papua New Guinea associated with echinoderms,
with descriptions of three new species. Cahiers de Biologie Marine, 40,
359–374.
BRITAYEV, T.A. & LYSKIN, S.A. 2002. Feeding of the symbiotic polychaete
Gastrolepidia clavigera (Polynoidae) and its interactions with its hosts.
Doklady Biological Sciences, 385, 352–356.
BRITAYEV, T.A., KRYLOVA, E.M., AKSYUK, T.S. & COSEL, R. 2003.
Association of Atlantic hydrothermal mytilids of the genus
Bathymodiolus spp. (Mollusca: Mytilidae) with the polychaeta
Branchipolynoe aff. seepensis (Polychaeta: Polynoidae): commensalism
or parasitism? Doklady Biological Sciences, 391, 371–374.
DE BARY, A.H. 1878. Uber Symbiose. Tagblatt der 51. Versammlung
Deutscher Naturforscher und Aerzte in Cassel. Kassel: Baier &
Lewalter, 121–126.
DIMOCK, R.V. & DAVENPORT, D. 1971. Behavioural specificity and the
induction of host recognition in a symbiotic polychate. Biological
Bulletin, 141, 472–484.
DITLEVSEN, H. 1917. Annelids. I. The Danish Ingolf Expedition, 4(4), 1–71.
DUNNE, J.A., WILLIAMS, R.J. & MARTINEZ, N.D. 2002. Network structure
and biodiversity loss in food webs: robustness increases with
connectance. Ecology Letters, 5, 558–567.
FAUCHALD, K. 2009. Eunoe opalina McIntosh, 1885. In FAUCHALD, K., ed.
World Polychaeta database. Accessed through: World Register of
Marine Species at http://www.marinespecies.org/aphia.php?p5
taxdetails&id5327847 on 22 September 2009.
FAUCHALD, K. & JUMARS, P. 1979. The diet of worms: a study of polychaete
feeding ecology. Oceanography and Marine Biology Annual Review,
17, 193–284.
FAUCHALD, K. & BARNICH, R. 2009a. Polynoidae. In FAUCHALD, K., ed.
World Polychaeta database. Accessed through: World Register of
Marine Species at http://www.marinespecies.org/aphia.php?p5
taxdetails&id5939 on 24 September 2009.
FAUCHALD, K. & BARNICH, R. 2009b. Polynoidae. In FAUCHALD, K., ed.
World Polychaeta database. Accessed through: DE BROYER, C., CLARKE,
A., KOUBBI, P., PAKHOMOV, E., SCOTT, F., VANDEN BERGHE, E. & DANIS,
B., eds. The SCAR-MarBIN Register of Antarctic Marine Species
(RAMS) at http://www.scarmarbin.be/rams.php?p5taxdetails&id5939
on 24 September 2009.
HARTMANN-SCHRODER, G. 1989. Polynoe thouarellicola n. sp. aus der
Antarktis, assoziiert mit Hornkorallen, und Wiederbeschreibung von
Polynoe antarctica Kinberg, 1858 (Polychaeta, Polynoidae).
Zoologische Anzaiger, 3/4, 205–221.
HAY, M.K., PARKER, J.D., BURKEPILE, D.E., CAUDILL, C.C., WILSON, A.E.,
HALLINAN, Z.P. & CHEQUER, A.D. 2004. Mutualism and aquatic
community structure: the enemy of my enemy is my friend. Annual
Review of Ecology, Evolution and Systematics, 35, 175–197.
HOLDEN, M.J. & RAITT, D.F.S. 1974. Manual of fisheries science. 2.
Methods of resource investigation and their application. FAO Fisheries
Technical Paper, No. 115, 1–211.
JANGOUX, M. 1987a. Diseases of Echinodermata. III. Agents metazoans
(Annelida to Pisces). Disease of Aquatic Organisms, 3, 59–83.
JANGOUX, M. 1987b. Diseases of Echinodermata. IV. Structural
abnormalities and general considerations on biotic diseases. Disease
of Aquatic Organisms, 3, 221–229.
KIRKEGAARD, J.B. & BILLET, D. 1980. Eunoe laetmogonensis, a new species
of polynoid worm, commensal with the bathyal holothurian Laetmogone
violacea, in the north-east Atlantic. Steenstrupia, 6, 101–109.
LAFFERTY, K.D., DOBSON, A.P. & KURIS, A.M. 2006. Parasites dominate food web
links. Proceedings of the National Academy of Sciences, 103, 11211–11216.
LYSKIN, S.A. & BRITAYEV, T.A. 2005. Symbionts of holothurians from
South Vietnam: intra- and interspecific interactions. Doklady Biological
Sciences, 401, 116–119.
MARTIN, D. & BRITAYEV, T.A. 1998. Symbiotic Polychaetes: review of
known species. Oceanography and Marine Biology Annual Review, 36,
217–340.
MIDDELFART, P. & CRAIG, M. 2004. Description of Austrodevonia sharnae n.
gen. n. sp. (Galeommatidae: Bivalvia), en ectocommensal of Taeniogyrus
australianus (Stimpson, 1855) (Synaptidae: Holothuroidea). Molluscan
Research, 24, 211–219.
NATION, J.L. 1983. A new method using hexamethildisilazane for
preparation of soft insect tissues for scanning electron microscopy.
Stain Technology, 58, 347–351.
NG, P.K.L. & MANNING, R.B. 2003. On two new genera of pea crabs parasitic
in holothurians (Crustacea: Decapoda: Brachyura: Pinnotheridae) from the
Indo-West Pacific, with notes on allied genera. Proceedings of the
Biological Society of Washington, 116, 901–919.
O’LOUGHLIN, P.M. 2002. Report on selected species of BANZARE and ANARE
Holothuroidea, with reviews of Meseres Ludwig and Heterocucumis Panning
(Echinodermata). Memoirs of Museum Victoria, 59, 297–325.
O’LOUGHLIN, P.M., MANJON-CABEZA, M.E. & RUIZ, F.M. 2009. Antarctic
holothuroids from the Bellingshausen Sea, with descriptions of new
species (Echinodermata: Holothuroidea). Zootaxa, 2016, 1–16.
PARMENTIER, E. & VANDEWALLE, P. 2005. Further insight on carapid-
holothuroid relationships. Marine Biology, 146, 455–465.
PETTIBONE, M.H. 1982. Annelida. In PARKER, S.B., ed. Synopsis and
classification of living organisms. New York: McGraw Hill, 1–43.
PETTIBONE, M.H. 1993. Scaled Polychaetes (Polynoidae) associated with
ophiuroids and other invertebrates and review of species referred to
Malmgrenia Mclntosh and replaced by Malmgreniella Hartman, with
descriptions of new taxa. Smithsonian Contributions to Zoology, 538, 1–92.
406 STEFANO SCHIAPARELLI et al.

http://journals.cambridge.org Downloaded: 11 Dec 2013 IP address: 50.198.197.234
REGISTER OF ANTARCTIC MARINE SPECIES (RAMS). 2009. Polychaeta.
Accessed through: DE BROYER, C., CLARKE, A., KOUBBI, P., PAKHOMOV,
E., SCOTT, F., VANDEN BERGHE, E. & DANIS, B., eds. The SCAR-MarBIN
Register of Antarctic Marine Species (RAMS) at http://www.
scarmarbin.be/rams.php?p5taxdetails&id5883 on 23 October 2009.
SCHIAPARELLI, S., GHIRARDO, C., BOHN, J., CHIANTORE, M., ALBERTELLI, G. &
CATTANEO-VIETTI, R. 2007. Antarctic associations: the parasitic
relationship between the gastropod Bathycrinicola tumidula (Thiele,
1912) (Ptenoglossa: Eulimidae) and the comatulid Notocrinus virilis
Mortensen, 1917 (Crinoidea: Notocrinidae) in the Ross Sea. Polar
Biology, 30, 1545–1555.
STRUGNELL, J.M., ROGERS, A.D., PRODOHL, P.A., COLLINS, M.A. & ALLCOCK,
A.L. 2008. The thermohaline expressway: the Southern Ocean as a
centre of origin for deep-sea octopuses. Cladistics, 24, 853–860.
TIAGO, C.G., BRITES, A.D. & KAWAUCHI, G.Y. 2005. A simple enzymatic
method for examining calcite ossicles of Echinodermata. Journal of
Microscopy, 218, 240–246.
WAREN, A. 1983. A generic revision of the family Eulimidae (Gastropoda,
Prosobranchia). The Journal of the Molluscan Studies, S13, 1–96.
WESENBERG-LUND, E. 1941. Notes on Polychaeta I. 1. Harmothoe
bathydomus H. Ditlevsen refound. Videnskabelige Meddelelser fra
Dansk naturhistorisk Forening i Kobenhaven, 105, 31–32.
ANTARCTIC SYMBIOTIC POLYNOID EUNOE OPALINA 407











![JNTU ONLINE EXAMINATIONS [Mid 2 - lica] - Webs](https://static.fdokumen.com/doc/165x107/633727b2d63e7c790105aa60/jntu-online-examinations-mid-2-lica-webs.jpg)








