
Heavy Metal and Arsenic Resistance of the Halophyte Atriplex halimus L. Along a Gradient of...
16
Heavy Metal and Arsenic Resistance of the Halophyte Atriplex halimus L. Along a Gradient of Contamination in a French Mediterranean Spray Zone Jacques Rabier & Isabelle Laffont-Schwob & Anca Pricop & Ahlem Ellili & Gabriel D’Enjoy-Weinkammerer & Marie-Dominique Salducci & Pascale Prudent & Brahim Lotmani & Alain Tonetto & Véronique Masotti Received: 8 February 2014 /Accepted: 12 May 2014 # Springer International Publishing Switzerland 2014 Abstract Elements uptake, histological distributions as well as mycorrhizal and physiological statuses of Atriplex halimus were determined on trace metal and metalloid polluted soils from the surrounding spray zones of a former lead smelter in the South-East coast of Marseille (France). Analyses of heavy metal and arsenic distribution in soil and plant organs showed that A. halimus tolerance is largely due to exclusion mechanisms. No specific heavy metal concentration in leaf or root tissues was observed. However, accu- mulation of salts (NaCl, KCl, Mg and Ca salts) on leaf bladders and peripheral tissues of roots was observed and may compete with metal element absorption. Oc- currence of endomycorrhizal structures was detected in roots and may contribute to lower element transfer from root into the aerial parts of plants. The non- destructive measurements of leaf epidermal chloro- phylls, flavonols and phenols showed a healthy state of the A. halimus population on the metal and metal- loid polluted sites. Considering the low metal bioac- cumulation and translocation factors along with a reduced metal stress diagnosis, A. halimus appeared as a good candidate for phytostabilization of trace metals and metalloids and notably arsenic in contam- inated soils of the Mediterranean spray zone. Howev- er, its invasive potential has to be determined before an intensive in situ use. Keywords Mediterranean saltbush . Root symbioses . Inorganic contamination . Phytostabilization . Salt- affected soils . Non-invasive sensors Water Air Soil Pollut (2014) 225:1993 DOI 10.1007/s11270-014-1993-y Jacques Rabier and Isabelle Laffont-Schwob contributed equally to this work. J. Rabier : I. Laffont-Schwob (*) : A. Pricop : A. Ellili : G. D’Enjoy-Weinkammerer : M.<D. Salducci : V. Masotti Aix Marseille Université, CNRS, IRD, Avignon Université, Institut Méditerranéen de Biodiversité et d’Ecologie marine et continentale (IMBE), Campus Saint Charles, Case 4, 3, place Victor Hugo, 13331 Marseille cedex 03, France e-mail: [email protected] A. Pricop : P. Prudent Aix Marseille Université, CNRS, Laboratoire de Chimie de l’Environnement, FRE 3416, Case 29, 3 place Victor Hugo, 13331 Marseille cedex 3, France A. Ellili Unité de Physiologie et Biochimie de la Tolérance au Sel des Plantes, Faculté des Sciences, Département de Biologie, Université de Tunis El Manar, 1060 Tunis, Tunisia B. Lotmani Laboratoire Protection des végétaux, Unité culture in vitro, Université A. Ibn Badis de Mostaganem, Mostaganem, Algeria A. Tonetto Aix Marseille Université, PRATIM, 3 place Victor Hugo, 13331 Marseille cedex 3, France
Transcript of Heavy Metal and Arsenic Resistance of the Halophyte Atriplex halimus L. Along a Gradient of...

Heavy Metal and Arsenic Resistance of the Halophyte Atriplexhalimus L. Along a Gradient of Contamination in a FrenchMediterranean Spray Zone
Jacques Rabier & Isabelle Laffont-Schwob & Anca Pricop & Ahlem Ellili &Gabriel D’Enjoy-Weinkammerer & Marie-Dominique Salducci & Pascale Prudent &Brahim Lotmani & Alain Tonetto & Véronique Masotti
Received: 8 February 2014 /Accepted: 12 May 2014# Springer International Publishing Switzerland 2014
Abstract Elements uptake, histological distributionsas well as mycorrhizal and physiological statuses ofAtriplex halimus were determined on trace metal andmetalloid polluted soils from the surrounding sprayzones of a former lead smelter in the South-East coastof Marseille (France). Analyses of heavy metal andarsenic distribution in soil and plant organs showedthat A. halimus tolerance is largely due to exclusionmechanisms. No specific heavy metal concentrationin leaf or root tissues was observed. However, accu-mulation of salts (NaCl, KCl, Mg and Ca salts) on leafbladders and peripheral tissues of roots was observedand may compete with metal element absorption. Oc-currence of endomycorrhizal structures was detectedin roots and may contribute to lower element transferfrom root into the aerial parts of plants. The non-
destructive measurements of leaf epidermal chloro-phylls, flavonols and phenols showed a healthy stateof the A. halimus population on the metal and metal-loid polluted sites. Considering the low metal bioac-cumulation and translocation factors along with areduced metal stress diagnosis, A. halimus appearedas a good candidate for phytostabilization of tracemetals and metalloids and notably arsenic in contam-inated soils of the Mediterranean spray zone. Howev-er, its invasive potential has to be determined beforean intensive in situ use.
Keywords Mediterranean saltbush . Root symbioses .
Inorganic contamination . Phytostabilization . Salt-affected soils . Non-invasive sensors
Water Air Soil Pollut (2014) 225:1993DOI 10.1007/s11270-014-1993-y
Jacques Rabier and Isabelle Laffont-Schwob contributed equallyto this work.
J. Rabier : I. Laffont-Schwob (*) :A. Pricop :A. Ellili :G. D’Enjoy-Weinkammerer :M.<D. Salducci :V. MasottiAix Marseille Université, CNRS, IRD, Avignon Université,Institut Méditerranéen de Biodiversité et d’Ecologie marine etcontinentale (IMBE),Campus Saint Charles, Case 4, 3, place Victor Hugo,13331 Marseille cedex 03, Francee-mail: [email protected]
A. Pricop : P. PrudentAix Marseille Université, CNRS, Laboratoire de Chimie del’Environnement, FRE 3416,Case 29, 3 place Victor Hugo, 13331 Marseille cedex 3,France
A. ElliliUnité de Physiologie et Biochimie de la Tolérance au Sel desPlantes, Faculté des Sciences, Département de Biologie,Université de Tunis El Manar,1060 Tunis, Tunisia
B. LotmaniLaboratoire Protection des végétaux, Unité culture in vitro,Université A. Ibn Badis de Mostaganem,Mostaganem, Algeria
A. TonettoAix Marseille Université, PRATIM,3 place Victor Hugo, 13331 Marseille cedex 3, France

1 Introduction
At the beginning of the Calanques hills (Marseille, S-EFrance), Mediterranean coastal ecosystems interfacepollution from abandoned industrial sites and pollutedsea sprays from urban effluents. Occurrence of heavymetals and metalloids from industrial activities, such aslead (Pb) and arsenic (As), was detected in sea sedi-ments and soils along the site. This pollution is consid-ered by the French Institute of Human Health (INVS) tobe unsafe for the population living near this site (Lasalle2007). There is a need of global understanding of thepollutant fluxes at the level of the different compart-ments (soil, water, plants) and the biocoenose relation-ships. Preliminary studies on this area were previouslydone: main physico-chemical characteristics, trace metaland metalloid (MM) contamination levels in soil and intwo species, i.e. Rosmarinus officinalis and Globulariaalypum growing within the former industrial site and notsubmitted to sea spray, were previously characterized(Testiati et al. 2013; Affholder et al. 2013).
Along the polluted coastline, an halophytic vegeta-tion developed amongst with the species Atriplexhalimus L. The latter is a widespread Mediterraneanshrub species (Osmond et al. 1980; McArthur andSanderson 1984; Ortíz-Dorda et al. 2005) with highresistance to various abiotic stresses such as drought(Le Houérou 1992), salinity (Bajji et al. 1998, 2002)and heavy metals (Lutts et al. 2004; Lefèvre et al. 2009;Manousaki and Kalogerakis 2009; Mateos-Naranjoet al. 2013). In addition to its ability to grow on degradedsoils and in very harsh conditions such as soil salinity(Pourrat and Dutuit 1994; Martinez et al. 2004),A. halimus has the property to produce an abundantfoliar biomass even during unfavourable periods of theyear (Kessler 1990). These characteristics enable thisplant to be used in the phytoremediation of contaminat-ed soils (see review byWalker et al. 2014), for example,for phytoextraction of cadmium and zinc in South-Eastern Spain (Lutts et al. 2004). However, since thisspecies shows a high genetic variability (Abbad et al.2004; Ortíz-Dorda et al. 2005), phytoextraction capaci-ties may greatly vary from one population to another.Since biotic and abiotic parameters may also impact thetranslocation abilities of this species, an alternative strat-egy is to use it for phytostabilization. However, thepresence as a key factor in horizontal transfer of nutri-ents, namely, root symbionts, remains under-documented. The arbuscular mycorrhizal (AM) status
of the genus Atriplex is controversial, and theChenopodiaceae family is normally defined as non-mycorrhizal. However, more than 10 species of theAtriplex genus were found to form endomycorrhizalassociations (Allen 1983; Johnson-Green et al. 1995;Aguilera et al. 1998; Barrow and Aaltonen 2001;Asghari et al. 2005; Sonjak et al. 2009). Furthermore,dark septate endophytes that can form mutualistic,mycorrhiza-like associations with their host plants(Jumpponen 2001) have been observed in roots of somespecies of the genus Atriplex (Sonjak et al. 2009). To thebest of our knowledge, there is no report about the AMstatus of A. halimus in France. This study, first, aimed todetermine if spontaneous A. halimus were naturallyendomycorrhized.
Moreover, heavy metals may alter the physiologicalstatus of plants growing on contaminated soils (Clijstersand Assche 1985). We also aimed to determine theheavy metal and metalloid contents of roots and leavesof A. halimus and of soils where the plants were spon-taneously growing, that are calcareous soils and subject-ed to marine aerosols deposits, and to establish if chlo-rophyll and some flavonoids contents of leaves werealtered. All these pieces of information were gathered todefine whether or not A. halimus is a good candidate forphytostabilization in the context of heavy metal contam-inated soils of the Mediterranean spray zone.
2 Material and Methods
2.1 The Study Area
Spontaneous populations of A. halimus from three sitesnamed Calanque de Saména, Calanque des Trous andCap Croisette in the National Park of Calanques werechosen to study the potential use of this species for aphytostabilization process under the mixed influence ofthe sea spray and metal and metalloid (MM) pollution.In 1925, the former silver-lead smelter factory ofl’Escalette (Fig. 1) ceased its activities, and approxi-mately 20,000 m3 of slag was deposited on the oldfactory site, but slag was also scattered along the coastas several main deposits and as road fill. The first twosites nearby visible slag deposits were chosen as poten-tially polluted and the latter as reference site since it isfar from slag deposits and from the former smelterfactory, so potentially the less contaminated site(Testiati et al. 2013; Affholder et al. 2013). Prior
1993, Page 2 of 16 Water Air Soil Pollut (2014) 225:1993

localization of the biggest slag deposit was checkedfrom aerials photography archives particularly for thereference site Cap Croisette (IGN 1926, 1961). In orderto check correlations between the Na and MM concen-trations in plant and soils to the sources of contamina-tion, the distance between each plant from the sea or theroad was evaluated from the multiplex GPS data andwith GPS visualizer (GPS visualizer 2013).
2.2 Sample Collection
Sample collection was done on five healthy-lookingA. halimus individuals in each population and on theirmycorhizospheric soil.
Five soil samples were collected per site from therooting zone of each selected plant on the top 15 cm ofsoils (after removal of the litter) and sieved at 2 mm for
trace and major elements analyses (Na, K, As, Cu, Fe,Mn, Pb, Zn).
Roots and leaves were collected for MM analyses. Theroot samples (ca. 5 g) were collected in the top 15 cm ofsoils of the five-plant individuals on each site then pooledin three samples for MM analysis. More roots wererandomly taken on each individual for assessment ofarbuscular mycorrhizae (AM) and/or dark septate endo-phyte (DSE) colonization and kept separately. A total of10 g of leaves were collected at different levels (high-medium-low) on the five plants of each site and thenpooled forMManalyses.More leaves per individual weretaken for the ex situ plant physiological index calibration.
2.3 Soil Salinity Estimation
Measurements of pH and salinity (calculated from con-ductivity) were determined by potentiometry in a 1:5
Fig. 1 Location of the three A. halimus stations nearby the former industrial site of l’Escalette, South-East of Marseille, France (filled circleindicates A. halimus sampling sites on the main picture and individual localization of the five individuals per site on the small pictures)
Water Air Soil Pollut (2014) 225:1993 Page 3 of 16, 1993

soil:water suspension using a Multi 3420 SET B-WTWpHmeter and ECmeter (Baize 1988) on the five soilsamples per site.
2.4 Trace and Major Element Analysis
The five soils samples per site were dried at roomtemperature and ground to 0.2 mm (tungstengrinder with titanium sieve). Total metal and majorelement concentrations were determined after soildigestion with aqua regia (1HNO3/2HCl) in a mi-crowave oven (milestone Start D) as described byAffholder et al. (2014).
Roots (three pooled samples per site) and leaves (onepooled sample per site) were washed apart with Milli-Qwater then dried at 40 °C during 1 week. Afterwards,dry weights were determined, and the samples wereground separately to 0.2 mm. Total metal concentrationsin roots and leaves were determined by acid digestion(HNO3:H2O2:ultra-pure H2O, volume proportion ratio2:1:1) with a microwave oven (milestone Start D).
Solutions obtained for soils and plant organ mineral-ization were filtrated at 0.45 μm. Cu, Fe, Pb, Mn and Znwere analyzed by ICP-AES (Jobin Yvon, Spectra 2000),As was analyzed by graphite furnace AAS (ThermoScientific ICE 3000 series AA spectrometer), whileflame AES (Thermo Scientific ICE 3000) was used forNa and K measurements.
Quality assurance-quality controls and accuracy werechecked using standard plant reference materials (DC73349, NCS-China) and standard soil reference mate-rials (CRM 049-050, RTC-USA) with accuracies within100±10 %.
Two factors were calculated as an indicator of thephytoremediation potential of A. halimus according toYoon et al. (2006): (1) bioconcentration factor (BC=metal concentration in root/metal concentration in soil)and (2) translocation factor (TF=metal concentration inleaf/metal concentration in root).
2.5 Root Symbiont Observations
The root samples were rinsed first under tap waterthen deionized water and stored in alcohol (60 %,v/v) at room temperature until proceeding. The per-centage of symbiont colonization was estimated byvisual observation of fungal colonization after clear-ing roots in 10 % KOH and staining withlactophenol blue solution, according to Phillips and
Hayman (1970). A minimum of 50 root segments(1-cm long) per plant was counted. AM frequencywas expressed as a percentage of colonization perroot sample. Occurrence of DSE was also recordedat the same time, and DSE frequency was deter-mined as previously described for AM frequency.
2.6 In Situ Non-destructive Plant Physiological IndexMeasurements
Plant physiological indices were optically estimatedusing a non-destructive measurement of constitutiveand induced epidermal phenols, flavonols, anthocyaninschlorophylls and the chlorophyll-to-flavonoids rationamed nitrogen balance index (NBI) of in situ plants,using a Multiplex® 3 (FORCE-A, Orsay, France). Thisportable fluorimetric device uses fluorescence technol-ogy with multiple excitations. Different combinations ofthe blue-green, red and far-red fluorescence signals atthe various excitation bands could be used as indices ofdifferent compounds (Cerovic et al. 2008; Agati et al.2011). For each individual, 25 measurements were doneon five plants for each site. The surface measured bymultiplex on each plant is hence equivalent to about10 % of the surface of an average A. halimus bush of1 m in diameter.
2.7 Ex Situ Plant Physiological Index Calibration
To elaborate scale values of multiplex index in arange from healthy to chlorotic or necrotic tissues,25 excised samples were chosen from apical buds,healthy leaves, yellowed leaves and dead shoots.They were laid on the 10-cm black surface accesso-ry of the Multiplex® 3 on their adaxial and abaxialsides and measured with the multiplex sensor forfive random layouts. Moreover, to estimate refer-ence values of multiplex indices for senescing pro-cess in leaf, five sets of eight excised healthy leaveswere measured as described above at time zero thenafter a period during which they were floated onwater above three filter paper layers in 9-cm diam-eter Petri dishes. The Petri dishes covered withaluminium foils were incubated in the dark at24 °C for 15 days to induce leaf senescence. Themeasurements done at the beginning of the treatmentwere repeated at the end of the incubation period.
1993, Page 4 of 16 Water Air Soil Pollut (2014) 225:1993

2.8 Scanning Electron Microscopy (SEM) andElemental Analysis
Root and leaf samples were prepared as previously de-scribed by Rabier et al. (2008). Transversal sections30-μm thick were cut at −25 °C using a cryomicrotome(Cryo-cut II microtome Reichert-Jung), then immediatelyplaced on SEM specimen holders and carbon metalized(10–15 nm) for observation under an ESEMPhilips XL 30microscope with detector EDAX sdd apollo 10. The ener-gy dispersive X-ray spectra (EDXS) for the compositionalanalyses were performed by point analysis with 3,000–4,000 cps (count per second) and a collection time of2 min. X-ray mapping was performed for 20 min to givethe elemental distribution for each selected element (Ca, K,Mg, Cl, Mn, S, P, Ni, Al, Mg, Fe and Si). In all cases, thevoltage was 20 kV. SEM images were obtained withbackscattered electron or secondary electron imaging.
2.9 Statistical Analysis
Statistical analyses and control charts were performedfor all data using JMP 10 statistical software (SASInstitute, Cary, NC, USA). Differences between MMconcentrations in plant parts and soils, plant stress indi-ces and fungal colonization percentages in the threepopulations of A. halimus were compared using theparametric Tukey’s test after log transformation of thevalues for data following a log normal distribution. Thenon-parametric Wilcoxon rank sum test (Kruskal-Wallistest) andWilcoxon each pair test were used for data withnon-normal distributions.
3 Results and Discussion
3.1 Soil Main Features
Soils were alkaline with typical pH (ca 8.6) from cal-careous zone (Table 1) as already reported by Testiatiet al. (2013) around the former industrial site ofl’Escalette. Soil salinity measurements showed that soilswere from non-saline to slightly saline with no signifi-cant difference between the three sites. No significantvariations of K were observed; however, Na averagesoil concentration was higher in Calanque de Saménathan that in Cap Croisette. This may be explained by thedifference of sea spray distribution. The plant popula-tion of Cap Croisette is 100–140 m away from the
seashore, while those of Calanque des Trous andCalanque de Saména are both 10–40 m away (Fig. 1and Table 1).
3.2 MM Concentrations in Soil and A. halimus Samples
Various metals and metalloids were detected in the soilsamples in link with the historical anthropic activities(Table 2). Cd data were not shown since theywere underthe detection level (0.1 mg kg−1). Sb which was presentat concentrations under 4 mg kg−1 in roots and far lowertoxic than As (Dietl et al. 1997; Foster et al. 2005) wasnot shown. The main element detected was Fe. Thepresence of As and Pb on the sites is in accordance withcontamination from the past activity of the former smelt-er of l’Escalette but less concentrated than the contam-ination measured in the slag deposit (Laffont-Schwobet al. 2011a; Testiati et al. 2013). Concentrations of As,Fe and Mn were significantly higher at Calanque deSaména than that in the two other sites. This high levelof As in Calanque de Saména is probably related toremains originating from the former creeping chimneys(Affholder et al. 2013; Testiati et al. 2013). Concentra-tions of Pb and Zn were significantly lower in CapCroisette than that in the other sites. Obviously, the soilconcentration variations of As between 43 and670 mg kg−1 for Calanque de Saména and Pb between37 and 1,293 mg kg−1 for Calanque des Trous indicate apoint source pollution deposit. Differently, the Pb soilconcentrations variations from 17 to 129mg kg−1 for theless polluted Cap Croisette could be mistaken withtraffic-related pollution (Triboit et al. 2010). Most ofthe variability of concentrations within one site can beexplained by the distance from the contaminated fillmaterial located under the road (Figs. 1 and 2). This isespecially the case for the main contaminants, i.e. As, Pband Zn on the three sites, but the differences may bemasked at lower concentrations as for Cu because thevalues are close to another non-contaminated referencesite (Affholder et al. 2013). In Fig. 2, the highest elementconcentrations in soils were detected nearby the road inCalanques de Saména, and these concentrations propor-tionally decreased when more distant from the road. InCalanque des Trous, all samples were along the road,and no pattern of element distribution could be ob-served. In Cap Croisette, element concentrations werelow along the transect from 25 to 140 m from the road.This shows clearly that the dispersion of pollutioncomes from the solid waste deposits used as building
Water Air Soil Pollut (2014) 225:1993 Page 5 of 16, 1993

materials of the road. The mechanism of dispersion onthe sea spray area may be different than the ridges zoneuphill submitted to the plume of smoke from the formerchimney of the smelter (Affholder et al. 2013; Testiatiet al. 2013). Moreover, the sea sprays might havewashed down most of the atmospheric pollution. Nev-ertheless, an observable contamination was highlightedspecifically for As in soils of Saména and for Pb and Znin soils of Calanque des Trous. Indeed, contaminationfactors (concentration ratio between soil and back-ground values) calculated from an estimation of thelocal contamination background levels in soils (5, 7.5,43 and 66 mg kg−1 for As, Cu, Pb and Zn, respectivelyas reported by Affholder et al. 2013), presented valuearound 60 for As in Calanque of Saména and values of 8and 14 for Zn and Pb, respectively, in Calanque desTrous, while values of contamination factors were onlyof 1 and 2 for soils of Cap Croisette whatever theelement.
However, only low concentrations of MM werefound in roots and leaves compared with the levels ofpollutants in the soils (Table 3). Similar results of lowMM transfer from soil to root in the surroundings of thethree sites were previously reported in the rare speciesAstragalus tragacantha (Laffont-Schwob et al. 2011a)and in Tamarix gallica L. (Abou Jaoudé et al. 2012),respectively. Further investigations on stem MM con-tents may be of interest to conclude on the potential MMtranslocation in A. halimus.
From a point of view of potentially using this plantspecies as a phytoextraction candidate, no individualexhibited concentrations in leaves higher than1.0 g kg−1 for Cu, Pb or As, and higher than10.0 g kg−1 for Zn. Therefore, A. halimus cannot beconsidered as a hyperaccumulator of any of theseelements according to the criteria set by Baker andBrooks (1989). No accumulation of MM was shownas BC and TF were inferior to one except for Zn-
Table 1 Soil characteristics of the three selected sites
Determined parameters Selected sites
Calanque de Saména Calanque des Trous Cap Croisette
pH 8.6±0.2 a 8.6±0.2 a 8.5±0.2 a
Salinity (mg kg−1) 0.5±0.4 a 0.9±0.8 a 0.2±0.1 a
Na (mg kg−1) 2,486±1,798 a 2,084±1,265 ab 867±242 b
K (mg kg−1) 5,440±1,573 a 4,917±2,969 a 4,978±1,314 a
Road distance from the sample sites (m) 17±13 b 8±3 b 88±47 a
Sea distance (m) 18±7 b 26±15 b 110±30 a
Means (±standard deviation) followed by different letters in the same line are significantly different (Wilcoxon test, p≤0.05, N=5)
Table 2 Average metal and Asconcentrations (mg kg−1) in soilsamples from each site
Means (±standard deviation (sd))followed by different letters in thesame line are significantly differ-ent (Wilcoxon test, p≤0.05, N=5)aFor major pollutants, when sdwas too high, value of each repli-cate is given in brackets to de-scribe pollution heterogeneity
Element Concentration (mg kg−1)
Calanque de Saména Calanque des Trous Cap Croisette
As 300±310 a
(60, 125, 606, 668, 43)a13±10 b
(13, 29, 14, 6, 4)a8±7 b
(5, 6, 7, 4, 20)a
Cu 27±6 a 28±17 a 18±11 a
Fe 33,860±14,612 a 11,566±3,998 b 10,600±3,699 b
Mn 540±240 a 191±99 b 271±78 b
Pb 263±141 a
(164, 211, 432, 393, 116)a628±633 a
(405, 1,293,1,317, 90, 37)a45±47 b
(16, 31, 19, 30, 129)a
Zn 355±299 a
(258, 213, 884, 262, 156)a528±446 a
(348, 1,016, 997, 195, 86)a88±58 b
(56, 50, 67, 77, 190)a
1993, Page 6 of 16 Water Air Soil Pollut (2014) 225:1993

translocation factors on all sites (Table 4). These on fieldresults are in agreement with previous studies undercontrolled conditions on A. halimus seedlings for Pb,Zn and Cu (Lotmani et al. 2011) and A. halimus cuttingsfor Cd and Pb (Manousaki and Kalogerakis 2009).
Moreover, MM accumulation was below the domes-tic animal toxicity (Mendez and Maier 2008). Whateverthe detected element, all bioconcentration factors werelow. For Na and K, apparent BC were two- to threefoldhigher, and especially, TF were high in accordance withthe excretion capacity of salt bladders of A. halimus(Grigore and Toma 2010). However, contribution fromsea sprays is not negligible, and TF may beoverestimated.
As hypothesized by Manousaki and Kalogerakis(2011), salt excretion organs of halophytes such as saltbladders of A. halimus are not always specific to sodiumand chloride ions, and other potentially toxic ions suchas zinc, copper or lead may be excreted through the saltbladders from leaf tissues onto the leaf upper layer. Asshown in Tables 3 and 4, the MM transfer is negligiblein comparison of Na and K.
Moreover, EDXS cartography was performed for allthe detected peaks of the X-ray spectra on root and leafsamples of A. halimus from the three sites to preciseelement localization in plant tissues. However, MMwere under the threshold of detection for EDXS. EDXS
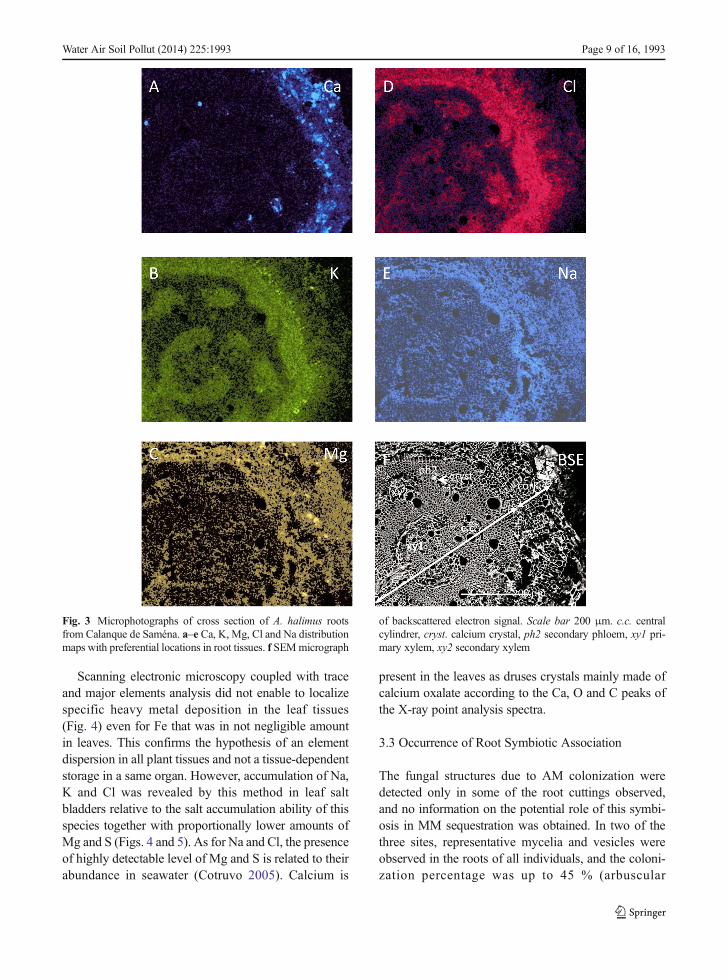
maps of root distribution for Si and Al are not shownbecause there are probably soil elements located on theouter layer of the root even after washing root samples,and no detection in the root tissues was obtained by thismethod. We hypothesized that there is no preferentialaccumulation of these elements in a specific tissue dif-ferently from our previous results for nickel in rootphloem ofGrevillea exul (Rabier et al. 2008). However,in root of A. halimus, Ca, Mg and K were detectedmostly in the cork both as concretions and diffuse layer(Fig. 3). The level of detected elements was by increas-ing order Ca<Mg<K. K was also present in the primaryand secondary xylem. Na and Cl were diffused in thecork and sparingly in the xylem. Moreover, Na wasmore concentrated near the inner boundary of the cork.Input from directly deposited marine aerosols is proba-bly the cause for a supplement of Ca, K and Mg(Whipkey et al. 2000). Because the macronutrients Ca,Mg and K were all found as diffuse layers and concre-tions in the roots, two kinds of interpretation can be putforward. On the one hand, a difference of soluble mac-ronutrients and the saline ion penetrations results fromtheir competition at the level of membrane transporters(Hu and Schmidhalter 2005; Kronzucker and Britto2010). This imbalance might also affect the absorptionof MM. On the other hand, the presence of concretionsof Ca, Mg and K might also incorporate traces of MM
Fig. 2 As, Fe, Pb and Znconcentrations (mg kg−1) in eachsoil samples versus the distance tothe road. Filled circle indicatesmain element concentrations insoil (mg kg−1).Grey line indicatessmooth line (JMP 10 statisticalsoftware). Cu and Mn valueswhere omitted for the sake ofclarity
Water Air Soil Pollut (2014) 225:1993 Page 7 of 16, 1993

via solid solution in the crystal lattice but under thedetection level for EDXS. Thus, the mineral may actas a storehouse for toxic ions (Skinner and Jahren 2003).
The results shown were obtained with root parts fromCalanque de Saména (Fig. 3), but similar concretions ofCa and Mg were observed for the two others sites.
Table 3 Major and trace elementaverage concentrations (mg kg−1)in root and leaf parts fromA. halimus samples on each site
Means for element concentrationsin leaves are from only onepooled sample for each site. Forcomparison of means (±standarddeviation, N=3) for element con-centrations in roots, the data in asame row followed by differentletters were significantly differentusing
ud l under de tec t ion leve l(<1.4 mg kg−1 for As)
*p≤0.05, Tukey’s parametricmultiple comparison test
**p≤0.05, nonparametric com-parisons for all pairs using Dunnmethod for joint ranking
Plant part Element Concentrations (mg kg−1)
Calanque de Saména Calanque des Trous Cap Croisette
Leaf As udl udl udl
Cu 5.8 10.2 9.8
Fe 126 137 161
Mn 34 56 55
Pb 1.9 1.3 1.3
Zn 122.8 161 150
Na 40.5×103 50.6×103 46.1×103
K 51.8×103 43.3×103 44.2×103
Root As 7.1±0.5 26±1 7±1
Cu* 12.0±0.2 c 15.9±0.5 a 13.7±0.3 b
Fe* 1,144±9 c 3,988±142 a 2,344±7 b
Mn* 94±2 c 467±48 a 134±6 b
Pb* 58±7 a 200±24 b 25±5 c
Zn** 72.8±0.5 ab 145±4 a 68±1b
Na** (5.16±0.57)×103 ab (7.11±0.58)×103 a (2.45±0.55)×103 b
K** (12.48±0.45)×103 a (9.24±0.13)×103 b (10.87±0.24)×103 ab
Table 4 Average transfer factorsof elements in A. halimus from thethree sites
udl under detection level
Elements Selected sites
Calanque de Saména Calanque des Trous Cap CroisetteBioconcentration factor (element concentration in root/element concentration in soil)
As 0.02 2.00 0.87
Cu 0.44 0.57 0.76
Fe 0.03 0.34 0.22
Mn 0.17 2.44 0.49
Pb 0.22 0.32 0.55
Zn 0.20 0.27 0.77
Na 2.07 3.41 2.82
K 2.28 1.88 2.18
Translocation factor (element concentration in leaf/element concentration in root)
As udl udl udl
Cu 0.48 0.64 0.71
Fe 0.11 0.03 0.07
Mn 0.36 0.12 0.41
Pb 0.03 0.006 0.05
Zn 1.68 1.11 2.20
Na 7.85 7.11 18.8
K 4.19 4.68 4.06
1993, Page 8 of 16 Water Air Soil Pollut (2014) 225:1993
Scanning electronic microscopy coupled with traceand major elements analysis did not enable to localizespecific heavy metal deposition in the leaf tissues(Fig. 4) even for Fe that was in not negligible amountin leaves. This confirms the hypothesis of an elementdispersion in all plant tissues and not a tissue-dependentstorage in a same organ. However, accumulation of Na,K and Cl was revealed by this method in leaf saltbladders relative to the salt accumulation ability of thisspecies together with proportionally lower amounts ofMg and S (Figs. 4 and 5). As for Na and Cl, the presenceof highly detectable level of Mg and S is related to theirabundance in seawater (Cotruvo 2005). Calcium is
present in the leaves as druses crystals mainly made ofcalcium oxalate according to the Ca, O and C peaks ofthe X-ray point analysis spectra.
3.3 Occurrence of Root Symbiotic Association
The fungal structures due to AM colonization weredetected only in some of the root cuttings observed,and no information on the potential role of this symbi-osis in MM sequestration was obtained. In two of thethree sites, representative mycelia and vesicles wereobserved in the roots of all individuals, and the coloni-zation percentage was up to 45 % (arbuscular
Fig. 3 Microphotographs of cross section of A. halimus rootsfrom Calanque de Saména. a–e Ca, K, Mg, Cl and Na distributionmaps with preferential locations in root tissues. f SEMmicrograph
of backscattered electron signal. Scale bar 200 μm. c.c. centralcylindrer, cryst. calcium crystal, ph2 secondary phloem, xy1 pri-mary xylem, xy2 secondary xylem
Water Air Soil Pollut (2014) 225:1993 Page 9 of 16, 1993

mycorrhizal colonization percentage was 41 %, 45 %and not detected, for Calanque de Saména, Calanquedes Trous and Cap Croisette, respectively). However,low occurrence of arbuscules was observed and mayindicate that nutrient exchange between the plant rootsand mycorrhizal fungi was low. These results confirmthe previous observation by He et al. (2002) on theoccurrence of endomycorrhizae in this plant speciesand confirm our previous results on polluted soils(Laffont-Schwob et al. 2011b). Furthermore, AMsymbiotic associations may be of interest inrhizoremediation process as reviewed by Kamaludeen
and Ramasamy (2008) and may contribute to lowerelement transfer into the aerial parts of plants (Christieet al. 2004). Otherwise, noDSEwas observed in the rootfragments contrariwise to the observations of Barrowand Aaltonen (2001) in Atriplex canescens.
3.4 Plant Stress Biomarkers
3.4.1 Setting-Up of Reference Values
Twenty percent of variation of chlorophyll indices wasobserved between abaxial and adaxial healthy leaves
Fig. 4 Microphotographs of cross section of A. halimus leavesfrom Calanque des Trous. a–e Ca, K, Mg, Cl and Na distributionmaps with preferential locations in leaf tissues, i.e. salt bladdersand external layers of mesophyll. f SEM micrograph of
backscattered electron signal. Scale bar 200 μm. col. collenchy-ma, cryst. calcium crystal, ep. epidermis, sb. salt bladder, kr. Kranzcell, ph1 primary phoem, ph2 secondary phloem, xy1 primaryxylem, xy2 secondary xylem
1993, Page 10 of 16 Water Air Soil Pollut (2014) 225:1993

under controlled conditions (Table 5). Moreover, refer-ence levels of chlorophylls under controlled conditionswere maximal in the adaxial side of healthy leaves anddecreased of 80 and 20 % in the yellowed and thesenescent leaves, respectively (Table 5). A. halimusleaves showed slight and slow senescence process whenincubating detached leaves in the dark, i.e. no visualdifference in chlorophyll loss was detected betweennon-senescent and senescent leaves after 15 days. An-thocyans are detected only on young buds or onyellowed leaves but are under the detection level formature plants (Table 5). Reference levels of flavonolswere maximal on the adaxial side of healthy andyellowed leaves, and lower in the abaxial side of healthyand yellowed leaves (Table 5). Leaf epidermal phenolindexes were maximal for dead and yellowed leaves.Nitrogen balance indices were significantly lower foryellowed leaves towards all the other conditions.
3.4.2 Field Measurements
A plant stress status of the individuals from the threepopulations was given by non-destructive measure-ments of chlorophyll and flavonoid indices on the field.Twenty percent of variation of chlorophyll indices inleaves was observed between the collection sites (Ta-ble 5) but were in the same order of variation as betweenchlorophyll indices of abaxial and adaxial healthy leavesunder controlled conditions (Table 5). Anthocyan indi-ces of plants from the three sites were under the detec-tion level. Neither the flavonol indices nor the leafepidermal phenol indices or the nitrogen balance indiceswere significantly different between the sites, althoughthe mean values were the lowest at Cap Croisette.
Thus, the main conclusion that can be drawn is thatthe mean values of in situ chlorophyll indices for thethree sites correspond to the reference indices for
Fig. 5 SEMmicrographs and elemental analysis of leaf cross sections of A. halimus. a Cross section of a leaf. b and c Shape and elementalanalysis of a calcium oxalate crystal. d and e Shape of salt bladders. f Elemental analysis of salt bladders
Water Air Soil Pollut (2014) 225:1993 Page 11 of 16, 1993

healthy leaves and let suppose a MM tolerance of thisspecies. This is in accordance with the visual observa-tions that no significant percentage of yellowed ornecrotized leaves of A. halimus was observed on thethree sites. Contrariwise to most plant species (Beckerand Apel 1993; Thimann 1980), A. halimus leavesshowed slight and slow senescence process when incu-bating detached leaves in the dark, i.e. no visual differ-ence in chlorophyll loss was detected between non-senescent and senescent leaves after 15 days. Theanthocyan index is not a good marker for A. halimusstress status because it was detected only on youngbuds, fruits or on yellowed leaves but was under thedetection level for mature plants. Globally, significantdifferences for all plant biomarkers were observed forthe different physiological stages and the leaf side. Thedifference of UV absorbing pigments between the leafadaxial and the abaxial sides is in accordance with theresults of several authors on different plant species(Cerovic et al. 2008; Bidel et al. 2007). Flavonols werenot detected into the dead shoots. A linear regressionexpression was calculated to describe the change offlavonol indices of individual plants as a function of
Pb concentration of their mycorhizospheric soil: flavo-nol index=0.65378+0.0002479×Pb mg kg−1 soil. Theprobability was p=0.163, and a weak correlation of R2=0.37 was obtained between flavonol index and Pb soilconcentrations. Therefore, the flavonol index is weaklylinked to the soil pollution levels. Previous results on theprotective mechanism of the antioxidant activities onbarley (Hordeum vulgare) leaf senescence test (Arnaoand Hernández-Ruiz 2009) and on Quercus ilexresprouts after fire (El Omari et al. 2003) suggest thatthe presence of antioxidant activities in Mediterraneanplant species protects them against a variety of reactiveoxygen species (ROS)-producing stresses. Globally,these non-destructive values for the three sites wereclose to those of the adaxial healthy leaves. This leadto conclude to a good health status of the threeA. halimus populations. However, since the differentphenological stages of the references are simultaneouslypresent on a same individual and because the plantsfrom each site were submitted to an heterogeneousheavy metal and As distribution, this contributes to thedistribution of the values and the lack of clear significantstatistical difference between the sites. Some
Table 5 Biomarkers, i.e. chlorophyll, flavonol, anthocyan and leaf epidermal phenol indices in A. halimus leaves from the three sites andreference values of multiplex indices for healthy and senescing leaves and dead shoots of A. halimus
Plant material Chlorophyllindex
Anthocyanindex
Flavonolindex
Leaf epidermalphenol index
Nitrogenbalance index
Field measurements
Shoots from Calanque de Saména 2.20±0.13 c udl 0.69±0.03 a 96.6±121.9 a 6.17±12.8 a
Shoots from Calanque des Trous 2.73±0.12 a udl 0.90±0.03 a 134±147 a 0.42±0.06 a
Shoots from Cap Croisette 2.28±0.12 c udl 0.64±0.03 a 65.1±41.5a 0.60±0.12a
Reference levels of phenological stages and sides of leaves
ap. buds 1.8±0.1 e 0.050±0.003 c 0.44±0.02 a 24.4±0.8 b 0.65±0.05a
ab. h. leaves 1.8±0.1 e udl 0.29±0.02 b 12.1±0.7 c 0.90±0.12a
ad. h. leaves 2.3±0.1 c udl 0.74±0.03 a 15.4±0.9 c 0.42±0.04a
ab. y. leaves 0.37±0.02 f 0.19±0.02 b 0.50±0.04 a 184±10 a 0.12±0.005b
ad. y. leaves 0.43±0.01 f 0.25±0.01 a 0.7±0.1 a 147±12 a 0.08±0.02b
d. shoots 0.61±0.02 f udl udl 290±12 a 1.14±0.04a
Reference levels of leaf senescence process
t 0 ad. h. leaves 2.5±0.2 a udl 0.52±0.07 a 37±9 a 0.75±0.12a
t 15 ad. d. s. leaves 2.01±0.15 a udl 0.4±0.2 b 24±9 b 0.91±0.34a
Plant health biomarker values are expressed as an average value for multiplex analyses (25 flashes per plant) and as the mean ± SE (N=5) foreach population. Means followed by different letter in a same column are significantly different at p≤0.05ND not detected, udl under detection level, ap. buds apical buds, ab. h. leaves abaxial side of healthy leaves, ad. h. leaves adaxial side ofhealthy leaves, ab. y. leaves abaxial side of yellowed leaves, ad. y. leaves adaxial side of yellowed leaves, d. shoots dead shoots, t 0 ad. h.leaves time zero adaxial side of healthy leaves, t 15 ad. d. s. leaves adaxial side of excised healthy leaves after 15 days of dark-inducedsenescence
1993, Page 12 of 16 Water Air Soil Pollut (2014) 225:1993

explanation may also be obtained from the literature, i.e.the scavenging of reactive oxygen under stressed envi-ronments is one of the suggested roles of flavonoids inplants (Türkan and Demiral 2009). Adaptation of plantcells to high NaCl salinity involves osmotic adjustmentand the compartmentalization of toxic ions, whereas anincreasing body of evidence suggests that high salinityalso includes oxidative stress (Khavari-Nejad et al.2006; Türkan and Demiral 2009). The threeA. halimus populations are submitted to sea sprays;however, the coupled effect of salt and trace metalsand metalloids may differ in Cap Croisette from bothother sites. A next step would be to characterize thegenetical variability of these three populations with theaim to distinguish in- and extrinsic factors affectingA. halimus resistance to MM and particularly to As.
These results confirmed that some flavonoids areimplicated in stress tolerance of A. halimus such as saltand drought and may be extended to other stresses suchas metal contamination on the studied sites.
Most of our results are congruent with a lower effecton A. halimus ecophysiological status in the less pollut-ed site, i.e. Cap Croisette. However, no drastic effects ofMM pollution on spontaneous populations of this spe-cies were noticed on the field.
In conclusion, considering the low trace elementtranslocation in the aerial parts certainly involving ex-clusion mechanisms along with the plant health diagno-sis and a deep root-system, A. halimus appears as a goodcandidate for phytostabilization of trace metals and met-alloids and notably As in salt-affected contaminatedsoils of the Mediterranean littoral. Under controlledconditions, similar levels of Zn, Cu and Pb in shootparts of various Algerian populations of A. halimus havebeen previously detected (Lotmani et al. 2011). Theseauthors confirmed that these elements were preferential-ly stored in the root parts rather than transferred in theaerial parts of the plants. Moreover, further study on salteffect on heavy metal absorption by A. halimus,endomycorrhized or not, may be of interest. Previousstudies of antagonism of metal versus salt on Atriplexgenus concentrated on root absorption (Lefèvre et al.2009) or adsorption on dead tissues (Sawalha et al.2009). Our observations lead to suppose that leaf ab-sorption of elements present in sea spray might also playa role in metal tolerance of A. halimus submitted tometal-polluted soils on the seashore. The apparent highMM resistance of A. halimus could be linked to toler-ance mechanisms both at the level of the root by actions
of salinity levels and symbiotic associations and at thelevel of leaf surface and bladders.
Observing the wide spontaneous occurrence of thisplant species on polluted salt-affected soils, its use forphytostabilization maybe of interest. However, the bio-geographical status of A. halimus in Southern France isstill discussed. If some coastal and insular populationsare probably native, as suggested by the genetic resultsof Ortíz-Dorda et al. (2005), this plant species is alsofrequently planted and naturalized along the FrenchMediterranean coast. Therefore, its invasive potentialhas to be determined before an in situ use even if thisspecies is currently suggested as a substitute of thehighly invasive Baccharis halimifolia L. for cultivationand ornamental purposes (Filippi and Aronson 2010;Heywood and Brunel 2008). Our field study was on aprotected area included in the National Park ofCalanques, South-East of France, and our results sug-gest to combine scientific approaches in ecology andplant physiology when dealing with pollution inprotected areas.
Acknowledgments The authors thank Thomas Devenoges,Soumia Djilalli and Cécile Evrard for their technical assistanceof root preparation and endomycorrhizal colonization countingand Laurent Vassalo and Carine Demelas for their analyticalassistance for trace and major element measurements. Manythanks to Alma Heckenroth, who conceived the map of the col-lection sites, and to Frédéric Médail for helpful discussion on thebiogeographic distribution of A. halimus. The authors are gratefulto Lidwine Le Mire Pécheux from the National Park of Calanquesfor helpful discussion on environment management. This studywas funded by the French Research National Agency (ANRMarséco 2008 CESA 018) and supported by the National Innova-tive Cluster on Risks Management. This research has been madepossible by participation in the EU COST Action FA 0901favouring links between scientists of various countries on halo-phytes. Isabelle Laffont-Schwob is grateful to Tim Flowers andJames Aronson that gave her the opportunity to participate to thisCOSTAction.
References
Abbad, A., Cherkaoui, M., Wahid, N., El Hadrami, A., &Benchaabane, A. (2004). Variabilités phénotypique etgénétique de trois populations naturelles d’Atriplex halimus.Comptes Rendus Biologies, 327, 371–380.
Abou Jaoudé, R., Pricop, A., Laffont-Schwob, I., Prudent, P.,Rabier, J., Masotti, V., De Dato, G., & De Angelis, P.(2012). Evaluating the potential use of Tamarix gallica L.for phytoremediation practices in heavy-metal polluted soils.Geophysical Research Abstracts, 14, EGU2012–EGU12887.
Water Air Soil Pollut (2014) 225:1993 Page 13 of 16, 1993

Affholder, M. C., Prudent, P., Masotti, V., Coulomb, B., Rabier, J.,Nguyen-The, B., & Laffont-Schwob, I. (2013). Transfer ofmetals and metalloids from soil to shoots in wild rosemary(Rosmarinus officinalis L.) growing on a former lead smeltersite: human exposure risk. Science of the Total Environment,454–455, 219–229.
Affholder, M.C., Pricop, A.-D., Laffont-Schwob, I. Coulomb, B.,Rabier, J., Borla, A., Demelas, C., & Prudent, P. (2014). As,Pb, Sb and Zn transfers from soil to root of wild rosemary: donative symbionts matter? Plant and Soil. doi:10.1007/s11104-014-2135-4.
Agati, G., Cerovic, Z. G., Pinelli, P., & Tattini, M. (2011). Light-induced accumulation of ortho-dihydroxylated flavonoids asnon-destructively monitored by chlorophyll fluorescence ex-citation techniques. Environmental and ExperimentalBotany, 73, 3–9.
Aguilera, L. E., Gutierrez, J. R., &Moreno, R. J. (1998). Vesiculararbuscular mycorrhizae associated with saltbushes Atriplexspp. (Chenopodiaceae) in the Chilean arid zone. RevistaChilena de Historia Natural, 71, 291–302.
Allen, M. F. (1983). Formation of arbuscular mycorrhizae inAtriplex gardneri, Chenopodiaceae: seasonal response in acold desert. Mycologia, 75, 773–776.
Arnao, M. B., & Hernández-Ruiz, J. (2009). Protective effect ofmelatonin against chlorophyll degradation during the senescenceof barley leaves. Journal of Pineal Research, 46(1), 58–63.
Asghari, H. R., Marschner, P., Smith, S. E., & Smith, F. A. (2005).Growth response of Atriplex nummularia to inoculation witharbuscular mycorrhizal fungi at different salinity levels. Plantand Soil, 273, 245–256.
Baize, D. (1988). Guide des analyses courantes en pédologie.Paris: Institut de la Recherche Agronomique.
Bajji, M., Kinet, J. M., & Lutts, S. (1998). Salt stress effectson roots and leaves of Atriplex halimus L. and theircorresponding callus cultures. Plant Science, 137, 131–142.
Bajji, M., Kinet, J. M., & Lutts, S. (2002). Osmotic and ioniceffects of NaCl on germination, early seedling growth, andion content of Atriplex halimus (Chenopodiaceae).CanadianJournal of Botany, 80, 297–304.
Baker, A. J. M., & Brooks, R. R. (1989). Terrestrial higher plantswhich hyperaccumulate metallic elements—a review of theirdistribution, ecology and phytochemistry. Biorecovery, 1,81–126.
Barrow, J. R., & Aaltonen, R. E. (2001). Evaluation of the internalcolonization of Atriplex canescens (Pursh) Nutt. roots bydark septate fungi and the influence of host physiologicalactivity. Mycorrhiza, 11, 199–205.
Becker, W., & Apel, K. (1993). Differences in gene expressionbetween natural and artificially induced leaf senescence.Planta, 189, 74–79.
Bidel, L. P. R., Meyer, S., Goulas, Y., Cadot, Y., & Cerovic, Z. G.(2007). Responses of epidermal phenolic compounds to lightacclimation: in vivo qualitative and quantitative assessmentusing chlorophyll fluorescence. Journal of ExperimentalBotany, 58, 1753–1760.
Cerovic, Z. G., Moise, N., Agati, G., Latouche, G., Ben Ghozlen,N., & Meyer, S. (2008). New portable optical sensors for theassessment of winegrape phenolic maturity based on berryfluorescence. Journal of FoodComposition and Analysis, 21,650–654.
Christie, P., Li, X., & Chen, B. (2004). Arbuscular mycorrhiza candepress translocation of zinc to shoots of host plants in soilsmoderately polluted with zinc. Plant and Soil, 261, 209–217.
Clijsters, H., & Assche, F. (1985). Inhibition of photosynthesis byheavy metals. Photosynthesis Research, 7, 31–40.
Cotruvo, J. A. (2005). Water desalinization processes and associ-ated health and environmental issues. Water Conditioningand Purification, 1, 13–17.
Dietl, C., Reifenhäuser, W., & Peichl, L. (1997). Association ofantimony with traffic occurrence in airborne dust, depositionand accumulation in standardized grass cultures. Science ofthe Total Environment, 205(2–3), 235–244.
El Omari, B., Fleck, I., Aranda, X., Abadía, A., Cano, A., Marino,B., & Arnao, C. (2003). Total antioxidant activity inQuercusilex resprouts after fire. Plant Physiology and Biochemistry,41, 41–47.
Filippi, O., & Aronson, J. (2010). Plantes invasives en régionméditerranéenne: quelles restrictions d'utilisation préconiserpour les jardins et les espaces verts? Ecologia Mediterranea,36, 31–54.
Foster, S., Maher, W., Krikowa, F., Telford, K., & Ellwood, M.(2005). Observations on the measurement of total antimonyand antimony species in algae, plant and animal tissues.Journal of Environmental Monitoring, 7, 1214–1219.
Institut national de l’information géographique et forestière (IGN).(1926, 1961) France & IGNF_PVA_1-0__1926-08-06__C3145-0351_1926_NP8_2050, scale 1: 9.916,I G N F _ P VA _ 1 - 0 _ _ 1 9 6 1 - 0 3 - 2 5 _ _ C 3 2 4 5 -0371_CDP1700_9055, scale 1: 8.975, 13008, Marseille 8eArrondissement retrieved from http://geoportail.gouv.fr.Accessed 6 May 2013.
GPS visualizer. (2013). http://www.gpsvisualizer.com/. Accessed10 May 2013.
Grigore, M.-N., & Toma, C. (2010). Salt secreting structures ofhalophytes. An integrative approach, Romanian AcademicPress, Bucharest, 290 pp (in Romanian, extended summaryin English). ISBN 978-973-27-1911-4.
He, X., Mouratov, S., & Steinberger, Y. (2002). Spatial distributionand colonization of arbuscular mycorrhizal fungi under thecanopies of desert halophytes. Arid Land Research andManagement, 16, 149–160.
Heywood, V., & Brunel, S. (2008). Code of conduct on horticul-ture and invasive alien plants. Council of Europe Conventionon the Conservation of European Wildlife and NaturalHabitats Standing Committee 28th meeting Strasbourg, 24–27 November 2008.
Hu, Y., & Schmidhalter, U. (2005). Drought and salinity: a com-parison of their effects on mineral nutrition of plants. Journalof Plant Nutrition and Soil Science, 168, 541–549.
Johnson-Green, P. C., Kenkel, N. C., & Booth, T. (1995).Distribution and phenology of arbuscular-mycorrhizae alonga salinity gradient at an inland salt pan. Canadian Journal ofBotany, 73, 1318–1327.
Jumpponen, A. (2001). Dark septate endophytes—are they my-corrhizal? Mycorrhiza, 11, 207–211.
Kamaludeen, S. P. B., & Ramasamy, K. (2008). Rhizoremediationof metals: harnessing microbial communities. Indian Journalof Microbiology, 48, 80–88.
Kessler, J. J. (1990). Atriplex forage as a dry season supplementa-tion feed for sheep in the Montane Plains of the Yemen ArabRepublic. Journal of Arid Environments, 19, 225–234.
1993, Page 14 of 16 Water Air Soil Pollut (2014) 225:1993

Khavari-Nejad, R. A., Bujar, M., & Attaran, E. (2006). Evaluationof anthocyanin contents under salinity (NaCl) stress in Bellisperennis L. In M. A. Khan & D. J. Weber (Eds.),Ecophysiology of High Salinity Tolerant Plants, Series:Tasks for Vegetation Science (pp. 127–134). New York:Springer.
Kronzucker, H. J., & Britto, D. T. (2010). Sodium transport inplants: a critical review. New Phytologist, 189, 54–81.
Laffont-Schwob, I., Dumas, P. J., Pricop, A., Rabier, J., Miché, L.,Affre, L., Masotti, V., Prudent, P., & Tatoni, T. (2011a).Insights on metal-tolerance and symbionts of the rare speciesAstragalus tragacantha aiming at phytostabilization of pol-luted soils and plant conservation. Ecologia Mediterranea,37, 57–62.
Laffont-Schwob, I., D’Enjoy-Weinkammerer, G., Pricop, A.,Prudent, P., Masotti, V., & Rabier, J. (2011b). Evaluation ofa potential candidate for heavy metal phytostabilization inpolluted sites of the Mediterranean littoral (SE Marseille):endomycorrhizal status, fitness biomarkers and metal contentof Atriplex halimus spontaneous populations. EcologicalQuestions, 14, 89–90.
Lasalle, J.L. (2007). Présence de plomb et d’arsenic sur le littoralsud de Marseille: une etude de santé (juillet 2005). INVSreport.
Le Houérou, H. N. (1992). The role of saltbushes (Atriplex spp.) inarid lands rehabilitation in the Mediterranean basin: a review.Agroforestry Systems, 18, 107–148.
Lefèvre, I., Marchal, G., Meerts, P., Corréal, E., & Lutts, S. (2009).Chloride salinity reduces cadmium accumulation by theMediterranean halophyte species Atriplex halimus L.Environmental and Experimental Botany, 65, 142–152.
Lotmani, B., Fatarna, L., Berkani, A., Rabier, J., Prudent, P., &Laffont-Schwob, I. (2011). Selection of Algerian populationsof theMediterranean saltbush,Atriplex halimus, tolerant to highconcentrations of lead, zinc, and copper for phytostabilizationof heavy metal-contaminated soils. The European Journal ofPlant Science and Biotechnology, 5, 20–26.
Lutts, S., Lefèvre, I., Delpérée, C., Kivits, S., Dechamps, C.,Robledo, A., & Correal, E. (2004). Heavy metal accumula-tion by the halophyte species Mediterranean saltbush.Journal of Environmental Quality, 33, 1271–1279.
Manousaki, E., & Kalogerakis, N. (2009). Phytoextraction of Pband Cd by the Mediterranean saltbush (Atriplex halimus L.):metal uptake in relation to salinity. Environmental Scienceand Pollution Research, 16, 844–854.
Manousaki, E., & Kalogerakis, N. (2011). Halophytes present newopportunities in phytoremediation of heavy metals and salinesoils. Industrial and Engineering Chemical Research, 50,656–660.
Martinez, J. P., Lutts, S., Schanck, A., Bajji, M., & Kinet, J. M.(2004). Is osmotic adjustment required for water stress resis-tance in the Mediterranean shrub Atriplex halimus L.?Journal of Plant Physiology, 161, 1041–1051.
Mateos-Naranjo, E., Andrades-Moreno, L., Cambrollé, J., & Perez-Martin, A. (2013). Assessing the effect of copper on growth,copper accumulation and physiological responses of grazingspecies Atriplex halimus: ecotoxicological implications.Ecotoxicological and Environmental Safety, 90, 136–142.
McArthur, E.D., & Sanderson, C. (1984). Distribution, systemat-ics and evolution of Chenopodiaceae: an overview. In A.RTiedemann., E.D. McArthur, H.C. Stutz, R. Stevens, K.L.
Johnson (Eds.), Proceedings of the symposium on the biologyof Atriplex and related Chenopods (pp. 14-24). USDepartment of Agriculture, Forest Service, USA.
Mendez, M. O., &Maier, R.M. (2008). Phytoremediation of minetailings in temperate and arid environments. Reviews inEnvironmental Science and Biotechnology, 7, 47–59.
Ortíz-Dorda, J., Martínez-Mora, C., Correal, E., Simón, B., &Cenis, J. L. (2005). Genetic structure of Atriplex halimuspopulations in the Mediterranean Basin. Annals of Botany,95, 827–834.
Osmond, C. B., Björkman, O., & Anderson, D. J. (1980).Physiological processes in plant ecology: towards a synthe-sis with Atriplex. Berlin: Springer-Verlag.
Phillips, J. M., & Hayman, D. S. (1970). Improved procedures forclearing roots and staining parasitic and vesicular arbuscularmycorrhizal fungi for rapid assessment of infection.Transactions of the British Mycological Society, 55, 159–161.
Pourrat, Y., & Dutuit, P. (1994). Etude précoce des effetsmorphologiques et physiologiques du rapport sodium/calcium in vitro sur une population d’Atriplex halimus.In AUPLEF—UREF (Ed. ) , Quel Avenir pourl’Amélioration des Plantes? (pp. 283–295). Paris: JohnLibbey Eurotext.
Rabier, J., Laffont-Schwob, I., Notonier, R., Fogliani, B., &Bouraïma-Madjebi, S. (2008). Anatomical element localiza-tion by EDXS inGrevillea exul var. exul under Nickel stress.Environmental Pollution, 156, 1156–1163.
Sawalha, M. F., Peralta-Videa, J. R., Sanchez-Salcido, B., &Gardea-Torresdey, J. L. (2009). Sorption of hazardous metalsfrom single and multi-element solutions by saltbush biomassin batch and continuous mode: interference of calcium andmagnesium in batch mode. Journal of EnvironmentalManagement, 90, 1213–1218.
Skinner, H. C. W., & Jahren, A. H. (2003). Biomineralization. InH. F. Holland & K. K. Turekian (Eds.), Treatise on geochem-istry 8 (pp. 117–184). New York: Elsevier.
Sonjak, S., Udovic, M., Wraber, T., Likar, M., & Regvar, M.(2009). Diversity of halophytes and identification ofarbuscular mycorrhizal fungi colonising their roots in anabandoned and sustained part of Secovlje saltern. SoilBiology & Biochemistry, 41, 1847–1856.
Testiati, E., Parinet, J., Massiani, C., Laffont-Schwob, I., Rabier, J.,Pfeifer, H. R., Lenoble, V., Masotti, V., & Prudent, P. (2013).Tracemetal andmetalloid contamination levels in soils and intwo native plant species of a former industrial site: evaluationof the phytostabilization potential. Journal of HazardousMaterials, 248–249, 131–141.
Thimann, K. V. (1980). Senescence in plants. Boca Raton: CRCPress.
Triboit, F., Laffont-Schwob, I., Demory, F., Soulié-Märsche,I., Rabier, J., Despréaux, M., & Thiéry, A. (2010).Heavy metal lability in porewater of highway detentionpond sediments in South-Eastern France in relation tosubmerged vegetation. Water, Air, & Soil Pollution,209(1-4), 229–240.
Türkan, I., & Demiral, T. (2009). Recent developments in under-standing salinity tolerance. Environmental and ExperimentalBotany, 67(1), 2–9.
Walker, D. J., Lutts, S., Sánchez-García, M., & Correal, E. (2014).Atriplex halimus L.: its biology and uses. Journal of AridEnvironments, 100-101, 111–121.
Water Air Soil Pollut (2014) 225:1993 Page 15 of 16, 1993

Whipkey, C. E., Capo, R. C., Chadwick, O. A., & Stewart, B. W.(2000). The importance of sea spray to the cation budget of acoastal Hawaiian soil: a strontium isotope approach.Chemical Geology, 168, 37–48.
Yoon, J., Cao, X., Zhou, Q., & Ma, L. Q. (2006).Accumulation of Pb, Cu and Zn in native plants growingon a contaminated Florida site. Science of the TotalEnvironment, 368, 456–464.
1993, Page 16 of 16 Water Air Soil Pollut (2014) 225:1993












