
Growth and physiological changes in scaled carp and blue tilapia under behavioral stress in mono-and...
10
Growth and physiological changes in scaled carp and blue tilapia under behavioral stress in mono- and polyculture rearing using a recirculated water system S.E. PAPOUTSOGLOU 1, * , H. MILIOU 1 , N.P. KARAKATSOULI 1 , M. TZITZINAKIS 1 and S. CHADIO 1 1 Laboratory of Applied Hydrobiology, Faculty of Animal Production, Agricultural, University of Athens, Iera, Odos 75, 11855 Athens, Greece; 2 Laboratory of Anatomy and Physiology, Faculty of Animal Pro- duction, Agricultural University of Athens, Iera, Odos 75, 11855 Athens, Greece; *Author for corre- spondence (e-mail: [email protected]; phone: 01 5294401; fax: 01 5294401) Received ; accepted in revised form 14 February 2002 Key words: Biological stress, Blood parameters, Blue tilapia (Oreochomis aureus), Carcass composi- tion, Intensive culture, Scaled carp (Cyprinus carpio) Abstract. Growth performance, carcass composition, liver and blood parameters of scaled carp (C), Cyprinus carpio, and blue tilapia (T), Oreochromis aureus, reared for eight weeks in two monoculture (100%C and 100%T) and two polyculture (60%C–40%T and 40%C-60%T) conditions were investi- gated. In polyculture 40%C–60%T both species achieved the highest levels of specific growth rate and the lowest levels of food conversion ratio and carcass lipid content. In addition, they exhibited the high- est values of plasma pO 2 and pH and the lowest values of plasma pCO 2 , cholesterol and albumin, al- though the differences among treatments were not significant in the case of tilapia. Tilapia showed sig- nificantly lower plasma Cl - levels than under the other conditions. Carp in monoculture and tilapia in polyculture 60%C–40%T had the lowest levels of specific growth rate and significantly higher levels of liver lipids and plasma triglycerides than in the other groups. In addition, carp in monoculture exhibited a significantly higher haematocrit than in polyculture. No significant variations among treatments were observed concerning plasma cortisol, glucose, osmolality, Na + ,K + , HCO 3 - and HCO 3 - /H 2 CO 3 in either species. The combination of scaled carp and blue tilapia, in which blue tilapia were the main species, proved to be the best for both species. It was suggested that growth and physiological changes under mono- and polyculture rearing, in an intensive system, seem to be as a result of a different state of stress related to fish behavior. Introduction Growth performance of common carp and tilapia in polyculture has extensively been studied in freshwater ponds (Dadzie 1982; Zweig 1989; Milstein 1995). Al- though Cyprinus carpio and Oreochromis spp. are reared fish around the world, there is very limited knowledge about their mass production using an intensive closed or semi-closed recirculated water system (Macintosh and De Silva 1984; Papoutsoglou and Voutsinos 1988; Papoutsoglou et al. 1992; Van-Gorder 1998). To our knowledge, there is only one report (Papoutsoglou et al. 1992) investigating Aquaculture International 9: 509–518, 2001. © 2002 Kluwer Academic Publishers. Printed in the Netherlands.
-
Upload
agriculturalathensu -
Category
Documents
-
view
0 -
download
0
Transcript of Growth and physiological changes in scaled carp and blue tilapia under behavioral stress in mono-and...

Growth and physiological changes in scaled carp andblue tilapia under behavioral stress in mono- andpolyculture rearing using a recirculated water system
S.E. PAPOUTSOGLOU1,*, H. MILIOU1, N.P. KARAKATSOULI1, M.TZITZINAKIS1 and S. CHADIO1
1Laboratory of Applied Hydrobiology, Faculty of Animal Production, Agricultural, University of Athens,Iera, Odos 75, 11855 Athens, Greece; 2Laboratory of Anatomy and Physiology, Faculty of Animal Pro-duction, Agricultural University of Athens, Iera, Odos 75, 11855 Athens, Greece; *Author for corre-spondence (e-mail: [email protected]; phone: 01 5294401; fax: 01 5294401)
Received ; accepted in revised form 14 February 2002
Key words: Biological stress, Blood parameters, Blue tilapia (Oreochomis aureus), Carcass composi-tion, Intensive culture, Scaled carp (Cyprinus carpio)
Abstract. Growth performance, carcass composition, liver and blood parameters of scaled carp (C),Cyprinus carpio, and blue tilapia (T), Oreochromis aureus, reared for eight weeks in two monoculture(100%C and 100%T) and two polyculture (60%C–40%T and 40%C-60%T) conditions were investi-gated. In polyculture 40%C–60%T both species achieved the highest levels of specific growth rate andthe lowest levels of food conversion ratio and carcass lipid content. In addition, they exhibited the high-est values of plasma pO2 and pH and the lowest values of plasma pCO2, cholesterol and albumin, al-though the differences among treatments were not significant in the case of tilapia. Tilapia showed sig-nificantly lower plasma Cl− levels than under the other conditions. Carp in monoculture and tilapia inpolyculture 60%C–40%T had the lowest levels of specific growth rate and significantly higher levels ofliver lipids and plasma triglycerides than in the other groups. In addition, carp in monoculture exhibiteda significantly higher haematocrit than in polyculture. No significant variations among treatments wereobserved concerning plasma cortisol, glucose, osmolality, Na+, K+, HCO3
− and HCO3−/H2CO3 in either
species. The combination of scaled carp and blue tilapia, in which blue tilapia were the main species,proved to be the best for both species. It was suggested that growth and physiological changes undermono- and polyculture rearing, in an intensive system, seem to be as a result of a different state of stressrelated to fish behavior.
Introduction
Growth performance of common carp and tilapia in polyculture has extensivelybeen studied in freshwater ponds (Dadzie 1982; Zweig 1989; Milstein 1995). Al-though Cyprinus carpio and Oreochromis spp. are reared fish around the world,there is very limited knowledge about their mass production using an intensiveclosed or semi-closed recirculated water system (Macintosh and De Silva 1984;Papoutsoglou and Voutsinos 1988; Papoutsoglou et al. 1992; Van-Gorder 1998). Toour knowledge, there is only one report (Papoutsoglou et al. 1992) investigating
Aquaculture International 9: 509–518, 2001.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.

the growth of Cyprinus carpio and Oreochromis aureus in polyculture using aclosed recirculated water system.
Dadzie (1982) suggested that the polyculture of different tilapia species, or oftilapia in combination with other teleosts, remains one of the most promising pro-duction systems of raising tilapia in ponds. Furthermore, Papoutsoglou et al. (1992)in a recirculated system, as well as Milstein (1995) in ponds, found that the growthrate of carp and tilapia in polyculture depended upon the species percentage com-bination and the initial body size. In addition, Papoutsoglou et al. (1992) concludedthat the advantages of polyculture fish rearing, in the case of an intensive produc-tion system, are based on fish ethology and feeding behaviour. However, furtherexplanation is required on the mechanisms, which may affect the changes in growthrate of carp and tilapia when both are present, especially at different percentages.
The objective of the present study was to evaluate the implication of a behav-ioral–ethological stress under mono- and polyculture rearing of two different fishspecies, with almost the same environmental and food requirements, in an inten-sive production system. For this purpose, growth and physiological changes wereinvestigated in Cyprinus carpio and Oreochromis aureus reared in monoculture andtwo polyculture combinations for eight weeks using a closed recirculated system.
Materials and methods
Fish and experimental design
Scaled carp, Cyprinus carpio (L.), and blue tilapia, Oreochromis aureus (St.), wereobtained from stock populations at the Laboratory of Applied Hydrobiology of Ag-ricultural University of Athens, Greece. Fish were reared for eight weeks in tankswith 108-l capacity. Four treatments were examined: two monoculture (100% carpand 100% tilapia) and two polyculture (60% carp–40% tilapia and 40% carp–60%tilapia). Fifteen fish were placed in each tank and the number of individuals foreach species was: 15 carp (100%C), 15 tilapia (100%T), 9 carp and 6 tilapia(60%C-40%T), 6 carp and 9 tilapia (40%C–60%T). For each treatment two repli-cate groups were maintained. Mean (± S.E.M.) initial body weight of carp (126.2 ±2.94 g) and tilapia (121.0 ± 3.63 g) did not differ significantly between all groups.
The water used (exchange rate: 1.5 times per hour) was drawn from the 50-tonlaboratory recirculated system (dechlorinated, filtered and UV-sterilised) and pro-vided with a compressed air supply. This system’s physicochemical parameters(mean ± S.E.M.) were monitored daily (temperature = 23.9 ± 0.03 °C, DO = 5.7 ±0.03 ppm, pH = 7.26 ± 0.002, total ammonia = 2.55 ± 0.006 ppm, NH3 = 0.26 ±0.001 ppm, NO2
− = 0.024 ± 0.0004 ppm, Cl− = 60.3 ± 0.07 ppm, hardness = 202.9± 0.18 ppm CaCO3).
Constant photoperiod (8L:16D) was used during the experiment. Fish were fedby hand, three times per day, a pelleted diet of the following proximate composi-tion: moisture 9.5%, crude protein 47%, crude fat 11.5%, crude fibre 2%, ash 11.5%
510

and nitrogen free extractives 18.5%. The daily feeding level was 2 % of initial bodyweight. Feeding level was not adjusted during the experiment to prevent stresswhich might arise during weighing procedures. Fish were not fed for 48 h beforesampling, which took place in the morning for all groups.
Blood sampling and analytical methods
At the end of the experiment, fish were anaesthetized using 2-phenoxyethanol(0.00032 ml/g per l) and immobilized within 3 min. Blood samples were taken fromthe heart and ventral aorta by heparinized syringes. Blood from the heart was im-mediately used for haematocrit determination, electrolytes (Na+, Cl− and K+), pO2,pCO2 and pH measurements (288-Blood Gas System, Ciba-Corning). PlasmaHCO3
− and HCO3−/H2CO3 values were calculated using the Henderson-Hasselbach
equation and corrected according to Boutillier et al. (1984) and Heisler (1984).Blood from the ventral aorta was used for plasma glucose, cholesterol, triglycer-ides, (enzymatic colorimetric methods, Elitech) and osmolality (cryoscopic osmom-eter, Gonotec Osmomat 030) determinations. Plasma cortisol levels were measuredby radioimmunoassay (RIA) using a commercially available kit (Coat-A-CountCortisol, DPC) which has been validated for fish (Ainsworth et al. 1985). In thepresent study, the sensitivity of the assay was 0.2 �g/dl and intra- and interassaycoefficient of variation was 3.2 and 6.5%, respectively.
Conventional methods were used for specific growth rate (SGR), food conver-sion ratio (FCR) and hepatosomatic index determinations (Papoutsoglou and Tziha1996). For the evaluation of proximate carcass composition of each species, fivespecimens of each group were minced together without viscera, lyophilized, andanalyzed (four replicates for each group) according to Kjeldahl and Soxhlet meth-ods (A.O.A.C. 1984). Liver total lipids were determined individually (five repli-cates for each group) in both species according to Folch et al. (1957).
One-way analysis of variance was carried out for the data obtained from indi-vidual fish (liver and blood parameters) for each species studied. Comparison ofmeans was performed by Duncan’s multiple range test. Differences were consid-ered significant at P < 0.05. The power of ANOVA was higher than 0.8 for all pa-rameters tested (Searcy-Bernal 1994). Where appropriate, data were log trans-formed in order to obtain normal distribution and homogeneity of variance anduntransformed means (± S.E.M.) are presented. For blood parameters, the samplingtime (time from the adding of anaesthetic until the end of blood sample collectionfrom each fish) was used as a covariate, in order to eliminate the effects of thedifferences in the anaesthetization time of each individual, carp or tilapia, amongexperimental groups. For parameters not measured individually (growth perform-ance and carcass composition) duplicated means of the two replicate groups rearedunder each condition are given for both species.
511
Results
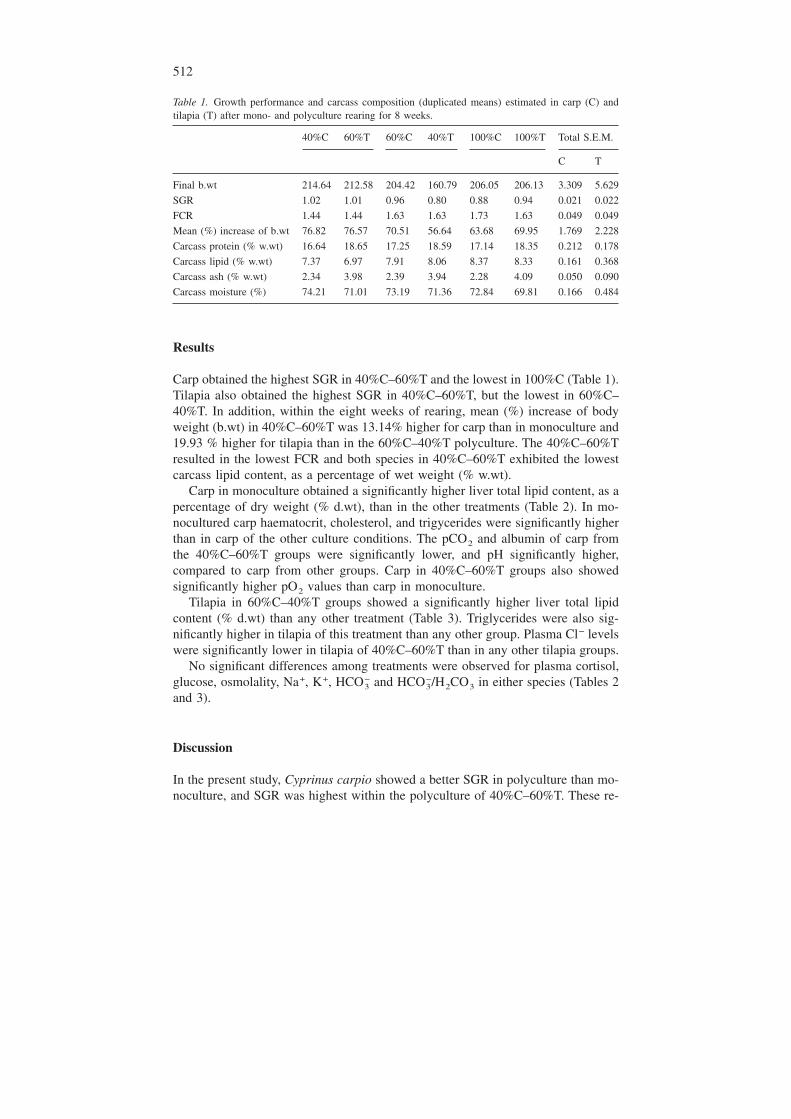
Carp obtained the highest SGR in 40%C–60%T and the lowest in 100%C (Table 1).Tilapia also obtained the highest SGR in 40%C–60%T, but the lowest in 60%C–40%T. In addition, within the eight weeks of rearing, mean (%) increase of bodyweight (b.wt) in 40%C–60%T was 13.14% higher for carp than in monoculture and19.93 % higher for tilapia than in the 60%C–40%T polyculture. The 40%C–60%Tresulted in the lowest FCR and both species in 40%C–60%T exhibited the lowestcarcass lipid content, as a percentage of wet weight (% w.wt).
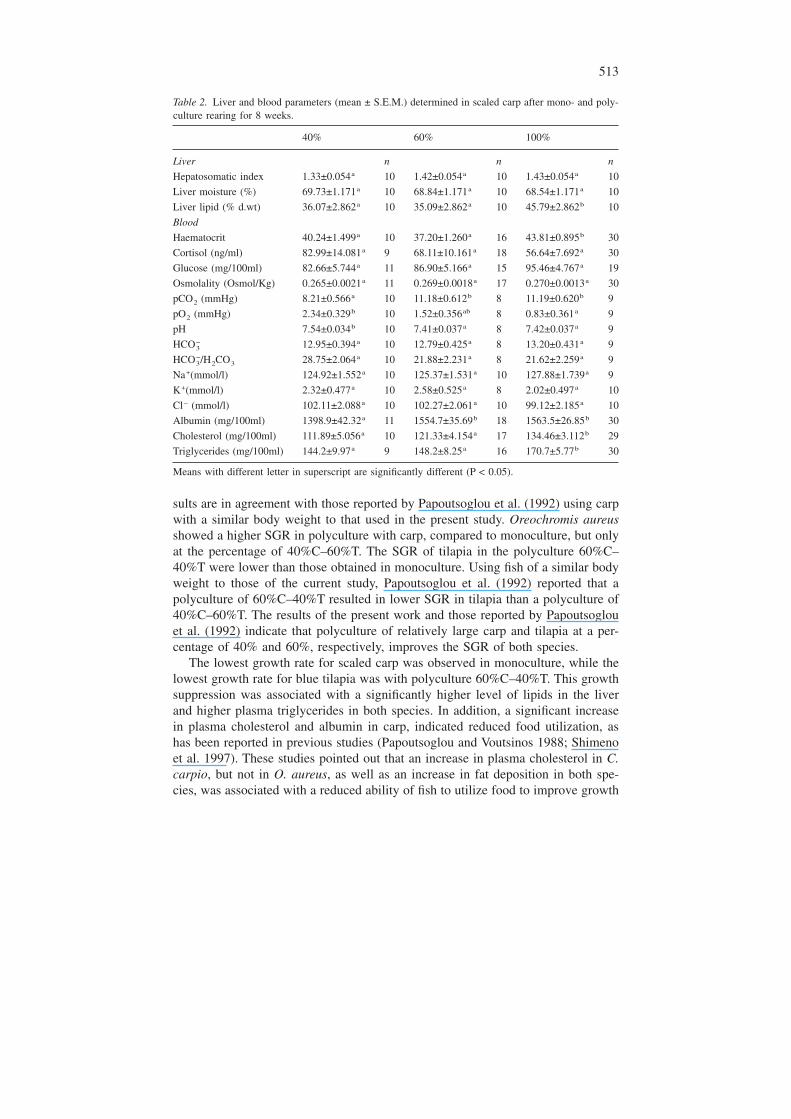
Carp in monoculture obtained a significantly higher liver total lipid content, as apercentage of dry weight (% d.wt), than in the other treatments (Table 2). In mo-nocultured carp haematocrit, cholesterol, and trigycerides were significantly higherthan in carp of the other culture conditions. The pCO2 and albumin of carp fromthe 40%C–60%T groups were significantly lower, and pH significantly higher,compared to carp from other groups. Carp in 40%C–60%T groups also showedsignificantly higher pO2 values than carp in monoculture.
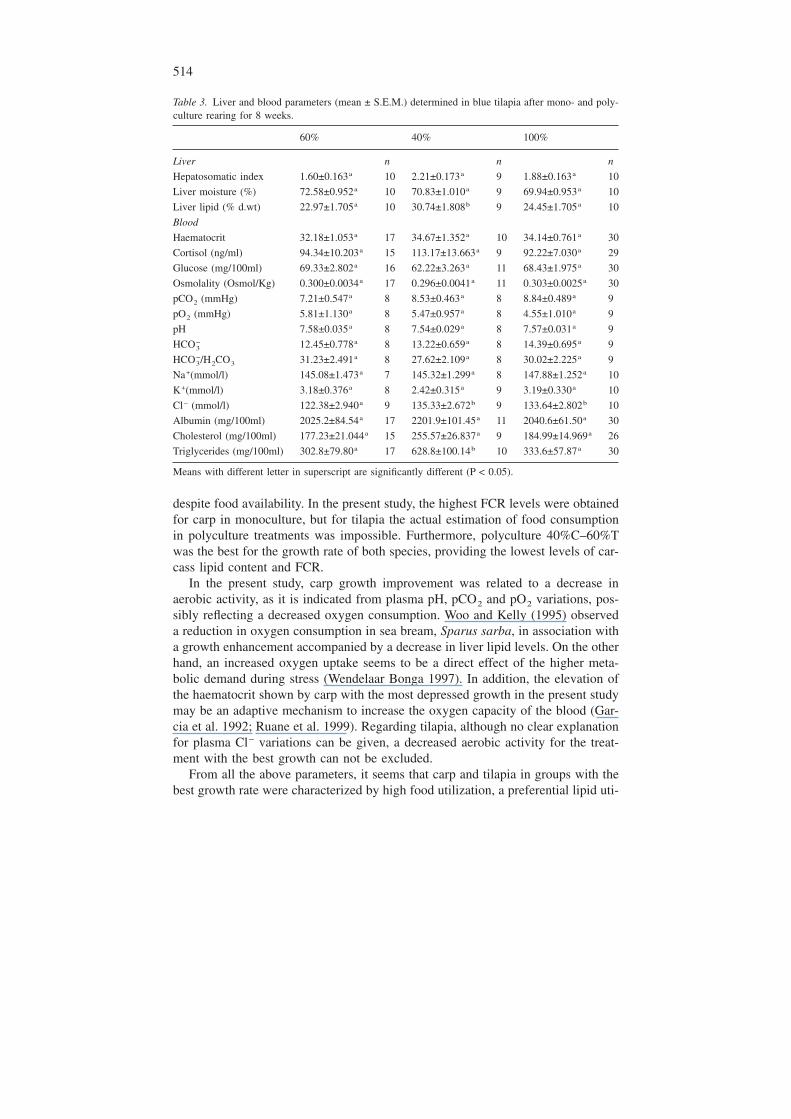
Tilapia in 60%C–40%T groups showed a significantly higher liver total lipidcontent (% d.wt) than any other treatment (Table 3). Triglycerides were also sig-nificantly higher in tilapia of this treatment than any other group. Plasma Cl− levelswere significantly lower in tilapia of 40%C–60%T than in any other tilapia groups.
No significant differences among treatments were observed for plasma cortisol,glucose, osmolality, Na+, K+, HCO3
− and HCO3−/H2CO3 in either species (Tables 2
and 3).
Discussion
In the present study, Cyprinus carpio showed a better SGR in polyculture than mo-noculture, and SGR was highest within the polyculture of 40%C–60%T. These re-
Table 1. Growth performance and carcass composition (duplicated means) estimated in carp (C) andtilapia (T) after mono- and polyculture rearing for 8 weeks.
40%C 60%T 60%C 40%T 100%C 100%T Total S.E.M.
C T
Final b.wt 214.64 212.58 204.42 160.79 206.05 206.13 3.309 5.629
SGR 1.02 1.01 0.96 0.80 0.88 0.94 0.021 0.022
FCR 1.44 1.44 1.63 1.63 1.73 1.63 0.049 0.049
Mean (%) increase of b.wt 76.82 76.57 70.51 56.64 63.68 69.95 1.769 2.228
Carcass protein (% w.wt) 16.64 18.65 17.25 18.59 17.14 18.35 0.212 0.178
Carcass lipid (% w.wt) 7.37 6.97 7.91 8.06 8.37 8.33 0.161 0.368
Carcass ash (% w.wt) 2.34 3.98 2.39 3.94 2.28 4.09 0.050 0.090
Carcass moisture (%) 74.21 71.01 73.19 71.36 72.84 69.81 0.166 0.484
512
sults are in agreement with those reported by Papoutsoglou et al. (1992) using carpwith a similar body weight to that used in the present study. Oreochromis aureusshowed a higher SGR in polyculture with carp, compared to monoculture, but onlyat the percentage of 40%C–60%T. The SGR of tilapia in the polyculture 60%C–40%T were lower than those obtained in monoculture. Using fish of a similar bodyweight to those of the current study, Papoutsoglou et al. (1992) reported that apolyculture of 60%C–40%T resulted in lower SGR in tilapia than a polyculture of40%C–60%T. The results of the present work and those reported by Papoutsoglouet al. (1992) indicate that polyculture of relatively large carp and tilapia at a per-centage of 40% and 60%, respectively, improves the SGR of both species.
The lowest growth rate for scaled carp was observed in monoculture, while thelowest growth rate for blue tilapia was with polyculture 60%C–40%T. This growthsuppression was associated with a significantly higher level of lipids in the liverand higher plasma triglycerides in both species. In addition, a significant increasein plasma cholesterol and albumin in carp, indicated reduced food utilization, ashas been reported in previous studies (Papoutsoglou and Voutsinos 1988; Shimenoet al. 1997). These studies pointed out that an increase in plasma cholesterol in C.carpio, but not in O. aureus, as well as an increase in fat deposition in both spe-cies, was associated with a reduced ability of fish to utilize food to improve growth
Table 2. Liver and blood parameters (mean ± S.E.M.) determined in scaled carp after mono- and poly-culture rearing for 8 weeks.
40% 60% 100%
Liver n n n
Hepatosomatic index 1.33±0.054a 10 1.42±0.054a 10 1.43±0.054a 10
Liver moisture (%) 69.73±1.171a 10 68.84±1.171a 10 68.54±1.171a 10
Liver lipid (% d.wt) 36.07±2.862a 10 35.09±2.862a 10 45.79±2.862b 10
Blood
Haematocrit 40.24±1.499a 10 37.20±1.260a 16 43.81±0.895b 30
Cortisol (ng/ml) 82.99±14.081a 9 68.11±10.161a 18 56.64±7.692a 30
Glucose (mg/100ml) 82.66±5.744a 11 86.90±5.166a 15 95.46±4.767a 19
Osmolality (Osmol/Kg) 0.265±0.0021a 11 0.269±0.0018a 17 0.270±0.0013a 30
pCO2 (mmHg) 8.21±0.566a 10 11.18±0.612b 8 11.19±0.620b 9
pO2 (mmHg) 2.34±0.329b 10 1.52±0.356ab 8 0.83±0.361a 9
pH 7.54±0.034b 10 7.41±0.037a 8 7.42±0.037a 9
HCO3− 12.95±0.394a 10 12.79±0.425a 8 13.20±0.431a 9
HCO3−/H2CO3 28.75±2.064a 10 21.88±2.231a 8 21.62±2.259a 9
Na+(mmol/l) 124.92±1.552a 10 125.37±1.531a 10 127.88±1.739a 9
K+(mmol/l) 2.32±0.477a 10 2.58±0.525a 8 2.02±0.497a 10
Cl− (mmol/l) 102.11±2.088a 10 102.27±2.061a 10 99.12±2.185a 10
Albumin (mg/100ml) 1398.9±42.32a 11 1554.7±35.69b 18 1563.5±26.85b 30
Cholesterol (mg/100ml) 111.89±5.056a 10 121.33±4.154a 17 134.46±3.112b 29
Triglycerides (mg/100ml) 144.2±9.97a 9 148.2±8.25a 16 170.7±5.77b 30
Means with different letter in superscript are significantly different (P < 0.05).
513
despite food availability. In the present study, the highest FCR levels were obtainedfor carp in monoculture, but for tilapia the actual estimation of food consumptionin polyculture treatments was impossible. Furthermore, polyculture 40%C–60%Twas the best for the growth rate of both species, providing the lowest levels of car-cass lipid content and FCR.
In the present study, carp growth improvement was related to a decrease inaerobic activity, as it is indicated from plasma pH, pCO2 and pO2 variations, pos-sibly reflecting a decreased oxygen consumption. Woo and Kelly (1995) observeda reduction in oxygen consumption in sea bream, Sparus sarba, in association witha growth enhancement accompanied by a decrease in liver lipid levels. On the otherhand, an increased oxygen uptake seems to be a direct effect of the higher meta-bolic demand during stress (Wendelaar Bonga 1997). In addition, the elevation ofthe haematocrit shown by carp with the most depressed growth in the present studymay be an adaptive mechanism to increase the oxygen capacity of the blood (Gar-cia et al. 1992; Ruane et al. 1999). Regarding tilapia, although no clear explanationfor plasma Cl− variations can be given, a decreased aerobic activity for the treat-ment with the best growth can not be excluded.
From all the above parameters, it seems that carp and tilapia in groups with thebest growth rate were characterized by high food utilization, a preferential lipid uti-
Table 3. Liver and blood parameters (mean ± S.E.M.) determined in blue tilapia after mono- and poly-culture rearing for 8 weeks.
60% 40% 100%
Liver n n n
Hepatosomatic index 1.60±0.163a 10 2.21±0.173a 9 1.88±0.163a 10
Liver moisture (%) 72.58±0.952a 10 70.83±1.010a 9 69.94±0.953a 10
Liver lipid (% d.wt) 22.97±1.705a 10 30.74±1.808b 9 24.45±1.705a 10
Blood
Haematocrit 32.18±1.053a 17 34.67±1.352a 10 34.14±0.761a 30
Cortisol (ng/ml) 94.34±10.203a 15 113.17±13.663a 9 92.22±7.030a 29
Glucose (mg/100ml) 69.33±2.802a 16 62.22±3.263a 11 68.43±1.975a 30
Osmolality (Osmol/Kg) 0.300±0.0034a 17 0.296±0.0041a 11 0.303±0.0025a 30
pCO2 (mmHg) 7.21±0.547a 8 8.53±0.463a 8 8.84±0.489a 9
pO2 (mmHg) 5.81±1.130a 8 5.47±0.957a 8 4.55±1.010a 9
pH 7.58±0.035a 8 7.54±0.029a 8 7.57±0.031a 9
HCO3− 12.45±0.778a 8 13.22±0.659a 8 14.39±0.695a 9
HCO3−/H2CO3 31.23±2.491a 8 27.62±2.109a 8 30.02±2.225a 9
Na+(mmol/l) 145.08±1.473a 7 145.32±1.299a 8 147.88±1.252a 10
K+(mmol/l) 3.18±0.376a 8 2.42±0.315a 9 3.19±0.330a 10
Cl− (mmol/l) 122.38±2.940a 9 135.33±2.672b 9 133.64±2.802b 10
Albumin (mg/100ml) 2025.2±84.54a 17 2201.9±101.45a 11 2040.6±61.50a 30
Cholesterol (mg/100ml) 177.23±21.044a 15 255.57±26.837a 9 184.99±14.969a 26
Triglycerides (mg/100ml) 302.8±79.80a 17 628.8±100.14b 10 333.6±57.87a 30
Means with different letter in superscript are significantly different (P < 0.05).
514

lization, and a decreased aerobic activity, whereas in the other treatments they werefound to be in a state of decreased fitness. An increased food conversion efficiencyin O. niloticus (Likongwe et al. 1996) and C. carpio (Qiu and Qin 1995), as well asan increased lipid utilization in O. niloticus (Alsop et al. 1999), S. sarba (Woo andKelly 1995) and Oncorhynchus mykiss (Kieffer et al. 1998), have also been ob-served under optimum environmental conditions for these species independently oftheir food habits. The opposite metabolic responses, combined to a depressedgrowth, might be explained by previous results (Woo and Kelly 1995; Rotllant etal. 2000). These demonstrated that certain chronic stress conditions elicited a pro-longed reduction in the levels of some hormones, such as thyroxine and/or growthhormone, which are known to enhance growth and lipid utilization (Farbridge et al.1992; Woo and Kelly 1995; Kelly et al. 1996; Papoutsoglou 1998).
Behavioral observations during the present experiment showed that tilapia be-came less aggressive in polyculture 40%C–60%T. Tilapia species are hierarchical(de Oliveira Fernades and Volpato 1993) and the formation of dominant fish andsubordinates seems to be dependent upon the number of individuals per tank. Onthe other hand, carp in monoculture showed a schooling behavior, which was dis-rupted in the presence of tilapia, especially when tilapia population density permit-ted, at the same time, a reduction in its aggressive behavior. Fish species display avariety of behavioral patterns in relation to their population density (Falter and De-backer 1988; Papoutsoglou et al. 1998). In most fish species, including commoncarp (Sehgal and Toor 1995), growth rate is inversely related to stocking density(Papoutsoglou et al. 1987). On the contrary, in O. aureus (Papoutsoglou et al. 1990),Salvenius alpinus (Jorgensen et al. 1993), and Dicentrarchus labrax (Papoutsoglouet al. 1998), an increase in fish density up to a certain level improves growth, prob-ably by reducing aggressiveness or due to a schooling behavior.
In the present study, after mono- and polyculture rearing for eight weeks, plasmacortisol levels did not significantly differ among the groups of each species studied.An increase in plasma cortisol levels has been accepted as an indicator of acutestress in fish (Barton and Iwama 1991; Pickering and Pottinger 1995; Papoutsoglouet al. 1999). On the other hand, during chronic stress, plasma cortisol levels maybe elevated (Papoutsoglou et al. 2000) or return to prestress values despite the con-tinued presence of stressors (Pickering and Pottinger 1987), suggesting that plasmacortisol may not be an appropriate indicator of chronic stress (Vijayan and Leath-erland 1988; Papoutsoglou 1998). In addition, the lack of significance in plasmaglucose, osmolality and blood electrolyte levels may be the result of recovery, as ithas been previously reported for scaled carp and other fish species under chronicstress (Ruane et al. 1999; Papoutsoglou et al. 2000; Rotllant et al. 2000). However,growth and physiological changes detected in the present study, showed that bothspecies were found in a different state of stress under monoculure than for the twopolyculture conditions which was in agreement with the observations concerningthe changes in their behavior.
Balm et al. (1994) detected a marked dichotomy in cortisol levels in tilapia, O.mossambicus, exposed to confinement in pairs and explained this difference interms of hierarchy, the low responders being dominant. Differences in the hypotha-
515

lamus-pituitary-adrenal axis activity associated with social rank have also been re-ported for other fish species (Peters et al. 1981; Pottinger and Pickering 1992). Inthe present study, differences in the hierarchical status of tilapia were more obviousin polyculture 60%C–40%T, the majority of tilapia individuals probably being sub-ordinates and high cortisol responders. Growth and physiological changes amongtreatments may reflect a previous more prolonged activation of interrenal cortisoloutput in tilapia of 60%C–40%T and in carp of monoculture than in their conspe-cific of other groups. Ruane et al. (1999) observed in rainbow and brown trout thatthe recovery of secondary stress responses is associated with the magnitude of thecortisol response. However, to date, schooling behavior of carp as a social statusassociated with increased interrenal activity has not, to our knowledge, been re-ported. Moreover, the variations in blood and liver parameters among treatments,observed in the present study, were more pronounced in carp than in tilapia. Thismay be explained by the ability of tilapia to down regulate the activity of the pi-tuitary interrenal axis more quickly than other fish species studied (Balm et al.1994). It should be noticed that no signs of illness (injuries, massive mucous se-cretion, fin erosion, etc.) were observed in any specimen, although a sensitivity ofhigh cortisol responders to infectious diseases has been suggested for other fishspecies (Maule et al. 1987).
It is concluded that an intensive polyculture rearing of scaled carp and blue ti-lapia at a percentage of 40% and 60%, respectively, contributes to a better physi-ological status of both species, indicating a decreased stress of behavioral–etho-logical origin.
Acknowledgements
We are grateful to Mr X. Vrettos for his technical assistance.
References
Ainsworth A.J., Bowser P.R. and Beleau M.H. 1985. Serum cortisol levels in channel catfish from pro-duction ponds. Progressive Fish-Culturist 47: 176–181.
Alsop D.H., Kieffer J.D. and Wood C.M. 1999. The effects of temperature and swimming speed oninstantaneous fuel use and nitrogenous waste excretion of the Nile tilapia. Physiological and Bio-chemical Zoology 72: 474–483.
A.O.A.C. 1984. Official Methods of Analysis. 14th edn. AOAC, Arlington, Virginia, USA.Balm P.H.M., Pepels P., Helfrich S., Hovens M.L.M. and Wendelaar Bonga S.E. 1994. Adrenocortico-
tropic hormone in relation to interrenal function during stress in Tilapia (Oreochromis mossambicus).General and Comparative Endocrinology 96: 347–360.
Barton B.A. and Iwama G.K. 1991. Physiological changes in fish from stress in aquaculture with em-phasis on the response and effects of corticosteroids. Annual Review of Fish Diseases, 3–26.
516

Boutillier R.G., Iwama G.K. and Heming T.A. 1984. Physicochemical parameters for use in fish respi-ratory physiology. In: Hoar W.S. and Randall D.J. (eds), Fish Physiology. Vol. 5. Academic Press,New York, pp. 403–430.
Dadzie S. 1982. Species combination in tilapia culture. Aquaculture 27: 295–299.de Oliveira Fernades M. and Volpato G.L. 1993. Heterogeneous growth in the Nile tilapia: Social stress
and carbohydrate metabolism. Physiology and Behavior 54: 319–323.Falter U. and Debacker L. 1988. Density-dependent behavioral shift in Oreochromis niloticus. In: Pullin
R.S.V., Bhukaswan T., Tonguthai K. and Maclean J.L. (eds), The second international symposiumon tilapia in aquaculture, Bangkok, Thailand, p. 599.
Farbridge K.J., Flett P.A. and Leatherland J.F. 1992. Temporal effects of restricted diet and compensa-tory increased dietary intake on thyroid function, plasma growth hormone levels and tissue lipidreserves of rainbow trout Oncorhynchus mykiss. Aquaculture 104: 157–174.
Folch J., Lees M. and Sloane-Stanley G.A. 1957. A simple method for the isolation and purification oftotal lipids from animal tissues. Journal of Biological Chemistry 226: 497–509.
Garcia M.P., Echevarria G., Martinez F.J. and Zamora S. 1992. Influence of blood sample collection onthe haematocrit value of two teleosts: rainbow trout (Oncorhynchus mykiss) and European sea bass(Dicentrarchus labrax L.). Comparative Biochemistry and Physiology 101: 733–736.
Heisler N. 1984. Acid-base regulation in fishes. In: Hoar W.S. and Randall D.J. (eds), Fish Physiology.Vol. 5. Academic Press, New York, pp. 315–401.
Jorgensen E.H., Christiansen J.S. and Jobling M. 1993. Effects of stocking density on food intake,growth performance and oxygen consumption in Arctic charr (Salvelinus alpinus). Aquaculture 110:191–204.
Kelly A.M., Kohler C.C. and Grau E.G. 1996. A mammalian growth hormone-releasing hormone in-creases serum growth hormone levels and somatic growth at suboptimal temperatures in tilapia.Journal of the World Aquaculture Society 27: 384–401.
Kieffer J.D., Alsop D. and Wood C.M. 1998. A respirometric analysis of fuel use during aerobic swim-ming at different temperatures in rainbow trout (Oncorhynchus mykiss). Journal of Experimental Bi-ology 201: 3123–3133.
Likongwe J.S., Stecko T.D., Stauffer J.R. Jr. and Carline R.F. 1996. Combined effects of water tempera-ture and salinity on growth and feed utilization of juvenile Nile tilapia Oreochromis niloticus (Lin-neaus). Aquaculture 146: 37–46.
Macintosh D.J. and De Silva S.S. 1984. The influence of stocking density and food ration on fry sur-vival and growth in Oreochromis mossambicus female x O. aureus male hybrids reared in a closedcirculated system. Aquaculture 41: 345–358.
Maule A.G., Schreck C.B. and Kaattari S.L. 1987. Changes in the immune system of coho salmon (On-corhynchus kisutch) during parr-to-smolt transformation and after implantation of cortisol. CanadianJournal of Fisheries and Aquatic Science 44: 161–166.
Milstein A. 1995. Fish-management relationships in Israeli commercial fish farming. Aquaculture Inter-national 3: 292–314.
Papoutsoglou S.E. 1998. Endocrinology of Fishes. Stamoulis Press, Athens, Greece, 599 pp.Papoutsoglou S.E. and Voutsinos G.A. 1988. Influence of feeding level on growth rate of Tilapia aureus
(Steindachner) reared in a closed circulated system. Aquaculture and Fisheries Management 19:291–298.
Papoutsoglou S.E. and Tziha G. 1996. Blue tilapia (Oreochromis aureus) growth rate in relation to dis-solved oxygen concentration under recirculated water conditions. Aquacultural Engineering 15: 181–192.
Papoutsoglou S.E., Papaparaskeva-Papoutsoglou E. and Alexis M.N. 1987. Effects of density on growthrate and production of rainbow trout (Salmo gairdneri Rich.) over a full rearing period. Aquaculture66: 9–17.
Papoutsoglou S.E., Voutsinos G.A. and Panetsos F. 1990. The effect of photoperiod and density ongrowth rate of Oreochromis aureus (Steindachner) reared in a closed water system. Animal ScienceReview 11: 73–87.
517

Papoutsoglou S.E., Petropoulos G. and Barbieri R. 1992. Polyculture rearing of Cyprinus carpio (L.)and Oreochromis aureus (St.) using a closed circulated system. Aquaculture 103: 311–320.
Papoutsoglou S.E., Tziha G., Vrettos X. and Athanasiou A. 1998. Effects of stocking density on behav-ior and growth rate of European sea bass (Dicentrarchus labrax) juveniles reared in a closed circu-lated system. Aquacultural Engineering 18: 135–144.
Papoutsoglou S.E., Miliou H., Chadio S., Karakatsouli N. and Zarkada A. 1999. Studies on stress re-sponses and recovery from removal in gilthead sea bream Sparus aurata (L.) using recirculated sea-water system. Aquacultural Engineering 21: 19–32.
Papoutsoglou S.E., Mylonakis G., Miliou H., Karakatsouli N.P. and Chadio S. 2000. Effects of back-ground color on growth performances and physiological responses of scaled carp (Cyprinus carpioL.) reared in a closed circulated system. Aquacultural Engineering 22: 309–318.
Peters G., Klinger H. and Delventhal H. 1981. Stress diagnosis of eels. In: Pickering A.D. (ed.), Stressand fish. Academic Press, London, pp. 333–334.
Pickering A.D. and Pottinger T.G. 1987. Crowding causes prolonged leucopenia in salmonid fish, de-spite interrenal acclimation. Journal of Fish Biology 30: 701–712.
Pickering A.D. and Pottinger T.G. 1995. Biochemical effects of stress. In: Hochashka P.W. and Mom-msem T.P. (eds), Biochemistry and molecular biology of fishes. Vol. 5. Elsevier, Amsterdam, pp.349–379.
Pottinger T.G. and Pickering A.D. 1992. The influence of social interaction on the acclimation of rain-bow trout, Oncorhynchus mykiss (Walbaum) to chronic stress. Journal of Fish Biology 41: 435–447.
Qiu D. and Qin K. 1995. Influence of salinity on energy budgets of juvenile common carp (Cyprinuscarpio L.). Journal of Fisheries, China Shuichan Xuebao 19: 35–42.
Rotllant J., Balm P.H.M., Ruane N.M., Pérez-Sánchez S.E., Wendelaar-Bonga S.E. and Tort L. 2000.Pituitary proopiomelanocortin-derived peptides and hypothalamus-pituitary-interrenal activity ingilthead sea bream (Sparus aurata) during prolonged crowding stress: differential regulation ofadrenocorticotropin hormone and a-melanocyte-stimulating hormone release by corticitropin-releas-ing hormone and thyrotropin-releasing hormone. General and Comparative Endocrinology 119: 152–163.
Ruane N.M., Wendelaar Bonga S.E. and Balm P.H.M. 1999. Differences between rainbow trout andbrown trout in the regulation of the pituitary-interrenal axis and physiological performance duringconfinement. General and Comparative Endocrinology 115: 210–219.
Searcy-Bernal R. 1994. Statistical power and aquacultural research. Aquaculture 127: 371–388.Sehgal H.S. and Toor H.S. 1995. The carp. Aquaculture 129: 113–117.Shimeno S., Shikata T., Hosokawa H., Masumoto T. and Kheyyali T. 1997. Metabolic response of feed-
ing rates in common carp, Cyprinus carpio. Aquaculture 151: 371–377.Van-Gorder S. 1998. Tilapia in recirculating aquaculture systems. Aquaculture Magazine 24: 21–23.Vijayan M.M. and Leatherland J.F. 1988. Effects of stocking density on the growth and stress-response
in brook charr, Salvenius fontinalis. Aquaculture 75: 159–170.Wendelaar Bonga S.E. 1997. The stress response in fish. Physiological Reviews 77: 591–625.Woo N.Y.S. and Kelly S.P.K. 1995. Effects of salinity and nutritional status on growth and metabolism
of Sparus sarba in a closed seawater system. Aquaculture 135: 229–238.Zweig R.D. 1989. Evolving water quality in a common carp and blue tilapia high production pond.
Hydrobiologia 171: 11–21.
518








![Rearing Healthy Calves Manual 2nd ed (1)[2] copy](https://static.fdokumen.com/doc/165x107/6326a762051fac18490ddddd/rearing-healthy-calves-manual-2nd-ed-12-copy.jpg)











