
Helioseismic line shape estimation given stochastic excitation
Upload
khangminh22Category
view
2download
0
Growth and Development in Rats Given Recombinant HumanEpidermal Growth Factor1-48 as Neonates
J. W. Henck,1 J. F. Reindel, and J. A. Anderson
Department of Pathology and Experimental Toxicology, Parke-Davis Pharmaceutical Research Division,Warner-Lambert Company, Ann Arbor, Michigan 48105
Received June 23, 2000; accepted February 27, 2001
To assess effects of supraphysiologic doses of human recombi-nant epidermal growth factor1-48 (rhEGF1-48) on neonatal rats, 10litters of Wistar rats/treatment group were given 0 (formulatedvehicle), 10, 100, or 1000 mg/kg daily by subcutaneous injection onpostnatal days (PND) 1 through 6. Clinical signs, body weight,acquisition of developmental landmarks and reflexes, and behav-ior were monitored during treatment and for 5 weeks thereafter (toPND 42). A subset of animals was euthanized weekly from PND7–28 and necropsied. Selected tissues were examined microscopi-cally. Body weight gain at 1000 mg/kg during treatment wassignificantly less than control. Precocious incisor eruption, eyeopening, vaginal opening, and preputial separation occurred at100 and/or 1000 mg/kg. Acquisition of reflexes (negative geotaxis,wire maneuver, acoustic startle reflex, and visual placing) wasdelayed at 1000 mg/kg. Acquisition of adult locomotion was alsodelayed at 1000 mg/kg. These effects were transient, as locomotoractivity at PND 28 and 42 did not differ from control. Effects onacoustic-startle responding persisted in females to final assessment onPND 42. Habituation to repeated acoustic stimuli was impaired, aswell as response inhibition following a prepulse acoustic stimulus.rhEGF1-48 induced structural changes in the skin, retina, kidney, oraland nasal mucosa, lung, and liver. Many of these changes wereconsistent with the expected mitogenic activity of rhEGF1-48 and weretransient in nature, as severity and incidence diminished with time.An exception was changes observed in the retina at 1000 mg/kg(rosettes/folds and focal defects in the outer nuclear/photoreceptorlayers) that were still present 3 weeks after termination of treatment.Acceleration of developmental landmarks; suppression of reflexes,behavior, and somatic growth; and mitogenic responses in epidermaltissues have been reported in rodents treated with epidermal growthfactor (EGF) derived from various mammalian species. These resultsdemonstrate that a 48-amino acid fragment of human EGF producedby recombinant technology also induces such effects.
Key Words: rhEGF1-48 toxicity; human epidermal growth fac-tor1-48; neonates; rats; development; behavior.
Recombinant human epidermal growth factor1-48 (rhEGF1-48)is a 48-amino acid fragment of human epidermal growth factor
(EGF), produced inEscherichia coliusing recombinant DNAtechnology. rhEGF1-48 is a potent mitogenin vitro and retainsthe full spectrum of biological activity of the 53-amino acidendogenous form of human EGF in the absence of the final 5carboxy-terminus amino acids (unpublished data, Parke-DavisPharmaceutical Research). A variety of pharmacological ac-tions have been attributed to EGF, including stimulation of cellgrowth in vitro andin vivo, inhibition of gastric-acid secretion,stimulation of ulcer healing, and stimulation of prostaglan-din-E2 synthesis (Carpenter and Wahl, 1990). EGF has beenshown to promote gastrointestinal mucosa regeneration/heal-ing (Guglietta and Sullivan, 1995; Kontureket al., 1988;Musroeet al., 1991), leading to the proposal that EGF might beuseful in the treatment of gastrointestinal erosive/ulcerativedisorders, including necrotizing enterocolitis in infants. Re-combinant human EGF (100 ng/kg/h) was given to an8-month-old child with necrotizing enteritis, and resulted inhistological repair of intestinal mucosa and clinical recovery inapproximately 2 months (Sullivanet al., 1991).
The peptide fragment rhEGF1-48 induces intestinal cell pro-liferation (Haskinset al., 1995, 1997) and may have potentialuse in infants. In addition to the aforementioned pharmacolog-ical actions in adults, EGF also appears to play an importantrole in regulation of cell proliferation and differentiation duringdevelopment. Various forms of EGF administered to neonatalrodents produce both accelerating and retarding effects onsomatic and behavioral development (Calamandreiet al.,1993). Subcutaneous injection of mouse EGF in neonatal ratsand mice produces alterations in craniofacial developmentcharacterized by precocious eye opening and incisor eruption.However, retardation of somatic growth, inhibition of hairgrowth, and delayed development of some reflexes and behav-iors have also been observed in the same animals (Calamandreiand Alleva, 1989; Cohen, 1962; Hoath, 1986; Smartet al.,1989; Tam, 1985). Because of the potential for clinical use ofrhEGF1-48 in infants, it was considered necessary to determinein neonates if pharmacological responses during developmentwere similar to those of other forms of EGF, and to explore thepotential for unexpected toxicity. The present study was there-fore undertaken to evaluate potential effects on growth and
1 To whom correspondence should be addressed at: Eli Lilly and Company,P.O. Box 708, Drop Code GL43, Greenfield, IN 46140. Fax: (317) 277-7601.E-mail: [email protected].
TOXICOLOGICAL SCIENCES62, 80–91 (2001)Copyright © 2001 by the Society of Toxicology
80

development induced by administration of rhEGF1-48 to neona-tal rats.
MATERIALS AND METHODS
Animals. The rat was selected for this study because the biological activityof various forms of EGF, including human EGF, has been well characterizedin this species. Forty timed-pregnant female Wistar rats [Crl:(WI)BR VAF/Plust; Charles River Breeding Laboratories, Inc., Kingston, New York],obtained on gestation day (GD) 13, were allowed to deliver their offspring. Theday of delivery was considered postnatal day (PND) 0. Each dam was housedindividually (except during lactation) in a stainless steel, hanging wire-meshcage. Near the time of parturition, a solid stainless steel plate and bedding wereadded to the home cage. Following weaning on PND 21, surviving male andfemale offspring were segregated by sex into separate cages according to litter.Each dam and her offspring were uniquely identified. Food (Purina CertifiedLab Rodent Chowt No. 5002, Ralston Purina Co., St. Louis, MO) and waterwere availablead libitumthroughout the study. Environmental conditions werein accordance with theGuide for the Care and Use of Laboratory Animals(National Research Council, 1996).
On GD 20, dams with their respective litters were randomly assigned to 4experimental groups (10/group) and, although dams remained untreated, eachlitter was treated with vehicle or rhEGF1-48 at 10, 100, or 1000mg/kg. Alloffspring were evaluated for acquisition of developmental landmarks. Off-spring were assigned to subgroups (in general composed of 7–10 rats/sex/group) for reflex and behavior testing, or for pathologic evaluation. The firstmale and female of each litter were assessed for acquisition of reflexes andbehavioral parameters. No more than one rat/sex/litter was evaluated for theseparameters. The subsequent 5 males and females of each litter were designatedfor euthanasia at weekly intervals for evaluation of pathologic changes.
Test material. Recombinant human epidermal growth factor1-48
(rhEGF1-48), purified fromEscherichia coli,is a fragment of the 53-amino acidhuman EGF (hEGF1-53). The amino acid sequence is Asp(NH2)-Ser-Asp-Ser-Glu-Cys-Pro-Leu-Ser-His-Asp-Gly-Tyr-Cys-Leu-His-Asp-Gly-Val-Cys-Met-Tyr-Ile-Glu-Ala-Leu-Asp-Lys-Tyr-Ala-Cys-Asp(NH2)-Cys-Val-Val-Gly-Tyr-Ile-Gly-Glu-Arg-Cys-Glu(NH2)-Tyr-Arg-Asp-Leu-Lys-OH. It lacks the final 5carboxy terminus amino acids of hEGF1-53, Trp-Trp-Glu-Leu-Arg(COOH).rhEGF1-48 was reconstituted in a vehicle of 20 mM phosphate buffer containing0.01% Tween 80 (pH 6.0) and was dosed based on active moiety (49.1mg/ml).
Reconstituted rhEGF1-48 was further diluted with sterile 5% dextrose inwater (D5W), with a final concentration of 0.5-mg rat albumin/ml, to obtainsolutions of 0.2, 2, and 20mg/ml at 10, 100, and 1000mg/kg, respectively.Control animals received formulated vehicle diluted with D5W and rat albu-min in the same ratio as animals administered 1000mg/kg. Dosing solutionswere filtered through a 0.2-mm filter and were transferred to a sterile container.
Samples from each dosing solution were analyzed for drug concentration byan HPLC assay and were found to be within 10% of the intended value, withthe exception of the 10mg/kg dosing solution, which was below the limit ofdetection. Mitogenic activity of each dosing solution was assessed pre- andpost-dosing, and was within the acceptable range of biologic activity for theassay.
Treatment. Offspring received solutions of rhEGF1-48 or vehicle by sub-cutaneous injection in the intrascapular region, once daily, at a dose factor of50 ml/kg of body weight from PND 1 (approximately 24 h after completion ofdelivery) through 6. A neonatal treatment period was found to induce maximalbiological activity in previous studies with EGF in rats. Dose volumes werebased on the most recent individual body weights.
Doses were selected based on preliminary results of an exploratory studywith rhEGF1-48 in adult male Wistar rats, as well as on information from thescientific literature on various forms of EGF administered to rodents. In theexploratory study, 100 or 1000mg/kg administered as a single intravenousinjection or as a continuous infusion via a subcutaneously implanted mini-pump were well tolerated. Subcutaneous injection of mouse EGF has been
shown to produce precocious eye opening and incisor eruption, but alsoretardation of somatic growth, as well as some reflexes and behaviors, inneonatal mice at 2700 to 4000mg/kg/day (Calamandrei and Alleva, 1989;Tam, 1985), and in neonatal rats at 500mg/kg/day (Hoath, 1986). A compar-ative study of mouse EGF to human EGF also revealed accelerated eyeopening in neonatal mice at subcutaneous doses of 250 to 2000mg/kg/day forboth compounds (Smithet al., 1985). Based on these results, doses of 10, 100,and 1000mg/kg/day were selected.
Each dam was handled daily following arrival in the laboratory in an attemptto minimize maternal stress and any ensuing effects on offspring during thepostnatal period. Offspring were observed daily throughout the study forclinical signs and survival, and body weights were recorded. All offspring wereretained (i.e., litter sizes were not standardized) to provide a sufficient numberto fill all subgroups.
Developmental landmarks, reflexes, and behavior.The developmentallandmarks, reflexes, and behavioral parameters indicated in Table 1 wereevaluated from the PND listed until acquisition. For developmental landmarks,acquisition occurred when both pinnae were detached and both eyes wereopen, when both lower incisors erupted, and when the vaginal opening becamepatent. Preputial separation was determined according to the method of Ko-renbrotet al. (1977). Evaluation of the static righting reflex, wire maneuver,visual placing, and negative geotaxis was according to standard methodology(Irwin, 1968; U.S. EPA, 1985; Moseret al., 1988). Auditory startle wasevaluated by placing the animal in a sound-attenuating enclosure; an auditorystimulus was elicited by a hand clicker, and a positive response was recordedif flicking of the pinnae and/or a sudden jerk of the body was observed.
Offspring were assessed for acquisition of 4 unique stages of locomotion,according to the method of Vorhees and coworkers (Vorheeset al., 1979).Locomotion was monitored from the most primitive form, pivoting (Stage 1),to forward locomotion with the head low and body low (Stage 2), to forwardlocomotion with the body raised and head low (Stage 3), to adult locomotionwith the head and body raised (Stage 4). Acquisition of each of these 4 stageswas analyzed separately.
On PND 28 and 42, each animal was monitored for locomotor activity andhabituation to a novel environment during a 10-min test session in an auto-mated activity monitor (Digiscan Activity Monitor System with OASIS soft-ware, AccuScan Electronics, Inc., Columbus, OH). Prior to testing, diagnosticand calibration procedures were conducted on the activity monitor system, asdescribed previously (Hencket al., 1996). The duration of the test session wasconsidered sufficient to evaluate the initial response to the novel environmentof the activity monitor during test minute 1, as well as to observe the rate indecline of activity (considered a measure of habituation) over 10 consecutive1-min test periods. Five parameters (total distance, number of vertical move-ments, rest time, stereotypy time, and center time) were considered to beindicative of the spontaneous movement of a rat and were subjected togroup-mean comparison.
TABLE 1Schedule of Testing for Acquisition of Developmental Land-
marks, Reflexes, and Behavioral Parameters in Rats GivenrhEGF1-48 Neonatally
Parameter
PN daytesting
initiated Parameter
PN daytesting
initiated
Pinnae detachment 1 Auditory startle 5Incisor eruption 1 Wire maneuver 5Negative geotaxis 1 Preputial separation 21Locomotion 1 Vaginal opening 18, 19 or 20Static righting reflex 1 Activity monitor 28Eye opening 5 Activity monitor 42Visual placing 5 Acoustic startle 42
81NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48

On PND 42, following activity monitor testing, each animal was evaluatedfor the acoustic startle response, as well as prepulse inhibition and habituationto an acoustic stimulus. Prior to testing, calibration and sensitivity evaluationprocedures were conducted on the acoustic startle system, as described previ-ously (Hencket al., 1996). Each animal was given 70 trials, using the SR-LABStartle Response System (San Diego Instruments, San Diego, CA). The trialsalternated between a background noise of 70 dB (total of 30 trials), a noise-level (NL) tone of 120 dB (total of 20 trials), and a prepulse (PRE) tone of 90dB, followed in 100 ms by a noise level tone of 120 dB (total of 20 trials),separated by intertrial intervals ranging from 5 to 30 s. Peak response (max-imum input voltage) for the first NL and PRE trials, the rate of habituation over20 NL trials, and percent response inhibition resulting from the prepulse toneswere evaluated.
Pathology. On PND 7, 14, 21, and 28, 1 animal/sex/litter (if available)from each treatment group was anesthetized with ether and decapitated. Majortissues and organs were examined grossly. Formalin-fixed brain, kidneys,small and large intestines, and liver were weighed from animals in the first 5litters/group. To assess intestinal length, small and large intestine from theseanimals were extended to full length, and measured. Brain, liver, kidney,stomach, small and large intestine, and skin were fixed in 10% bufferedformalin. Eyes were fixed in 6% glutaraldehyde. Formalin-fixed tongue, nasalcavity, hard palate, trachea, mandibular salivary gland, esophagus, urinarybladder, lung, and thyroid were collected on PND 7 only, from 1 animal/sex/litter from the first 5 litters/group in the control and 1000mg/kg group. Alltissues were processed in paraffin, stained with hematoxylin and eosin, andexamined microscopically.
Statistical analysis. To control for the multiplicity of statistical compari-sons (i.e., reduce the number of false positive conclusions), all sets of devel-opmental and behavioral response measures (parameters) were divided intodistinct classes based on the relationship of the parameters. For example, alldevelopmental landmark data comprised one class, while all activity monitordata on a specific day comprised another. The level of significance for eachcomparison within the class was 0.05 divided by the square root of the numberof parameters in the class (Tukeyet al., 1985), to provide an approximateclasswise significance level of 5%. Organ weight data were not partitioned andwere analyzed at an approximate class-wise significance level of 1%.
The basic trend test employed was the sequential trend test, using therank-dose scale and rank-transformed data (Park, 1985; Tukeyet al., 1985).This test is equivalent to sequential application of the Kruskal-Wallis 1-wayanalysis of variance (ANOVA) by ranks (Kruskal and Wallis, 1952), with thetreatment effects being evaluated by dose-trend tests that have contrast coef-ficients for equally spaced (ranked) treatment groups. The sequential lineartrend test is designed to detect monotone changes in dose response; it does notdetect trend reversal or curvature. If the true dose response was not monotonic,the trend test was considered not sensitive enough to detect the treatmenteffect. A trend reversal test was then conducted, consisting of a quadratic trendtest performed at the 2-tailed 1% class-wise significance level. If the trendreversal test was significant and the high-dose trend test was not significant, thetreatment groups were compared to control by Dunnett’s test (Dunnett, 1955,1964), using rank-transformed data, and performed at the 5% class-wisesignificance level.
The monotonic dose-response relationship tested by the trend test was notconsidered realistic for the activity monitor parameters, because they havebeen demonstrated with several drugs to exhibit a U-shaped dose-responsecurve (Iversen and Iversen, 1981). Hence, Dunnett’s test on rank-transformeddata was performed in place of the trend test as the main analytical method.Acoustic startle data, as well as the activity monitor parameters total distanceand number of vertical movements, were subjected to profile analysis (Johnsonand Wichern, 1982) to evaluate the response by treatment group interaction(parallelism test) and, secondarily, to address the question of equal groupeffects; the raw data were used for this analysis. The methods of Greenhouseand Geisser (1959) were incorporated in the profile analysis to produceconservative tests for the parallelism hypotheses.
RESULTS
Clinical Observations and Growth
Generalized exfoliation occurred over the entire body inapproximately 70% of animals at 1000mg/kg during the weekfollowing treatment. Treatment-related ocular alterations oc-curred in approximately 20% of animals at 1000mg/kg duringthis period, and included corneal drying or opacity, enlargedeyelid, keratitis sicca, and unilateral eye enlargement; theseobservations appeared a few days after precocious eyeopening.
Eight of 103 animals at 1000mg/kg died or were euthanizedmoribund. No deaths or treatment-related clinical signs oc-curred at 10 or 100mg/kg.
Body-weight gain during the treatment period at 1000mg/kgwas less than control by 23% and 27% for males and females,respectively. No treatment-related body weight changes wereapparent at 10 or 100mg/kg throughout the study or at 1000mg/kg during the post-treatment period (PND 7–42).
Developmental Landmarks and Reflexes
Significantly (p , 0.025) accelerated acquisition of lowerincisor eruption (by 1–2 days), eye opening (by 3–5 days), andpreputial separation (by 4–7 days) occurred at 100 and 1000mg/kg, and of vaginal opening (by 12 days) occurred at 1000mg/kg (Table 2). Timing of pinnae detachment was not af-fected by treatment.
Development of all reflexes evaluated, with the exception ofstatic righting reflex, was significantly (p , 0.025) delayed inmales and/or females at 1000mg/kg (Table 3): negativegeotaxis (by 2 days), acoustic startle (by 1–2 days), wiremaneuver (by 2 days), and visual placement (by 1.5 days).
Behavior
No treatment-related effects were seen on initial stages oflocomotor development. Progression to the final stage of adultlocomotion was delayed by 1.5–2 days in males and females at1000mg/kg (Fig. 1), although this change was not statisticallysignificant. Evaluation of locomotor activity in an automatedactivity monitor on PND 28 and 42 revealed no treatment-related effect on horizontal or vertical activity, stereotypy, orhabituation to the novel environment of the open field (data notshown).
The acoustic startle response to 120 dB tones (evaluated asmaximum input voltage) for males at 10mg/kg, but not at 100or 1000mg/kg, was less than control during the majority of 20trials, by a maximum of 52% (Fig. 2). However, becausemaximum input voltage for many of these trials was within thehistorical control range for this laboratory, and because nodose-response relationship was evident, this apparent effectwas considered not biologically significant. Group mean max-imum input voltage for females at 1000mg/kg was comparableto control for the first 10–120 dB trials; however, for the next
82 HENCK, REINDEL, AND ANDERSON

10 trials, maximum input voltage was up to 2-fold greater thancontrol. Although comparison across the 20 trials revealed nostatistical significance, the mean and standard error for maxi-mum input voltage of females at 1000mg/kg during the ma-jority of the last 10 trials was greater than the mean andstandard error of control females, and observation of the dataindicated that responding did not diminish with repeated acous-tic stimuli (Fig. 2). Group mean percent response inhibitionfollowing a prepulse stimulus for 20–120-dB and 20 prepulsetrials is given in Table 4. Although no statistically significantdifferences were apparent for maximum input voltage for thefirst prepulse trial for males or females, group mean andstandard error percent response inhibition of females at 1000mg/kg was less than the mean and standard error of controls.
Pathology
Organ weight data and histopathologic findings are summa-rized in Tables 5 and 6, respectively. Alterations were noted inskin, eye, kidney, nasal and oral mucosa, lung, and liver ofmale and female rhEGF1-48-treated rats terminated at variousintervals.
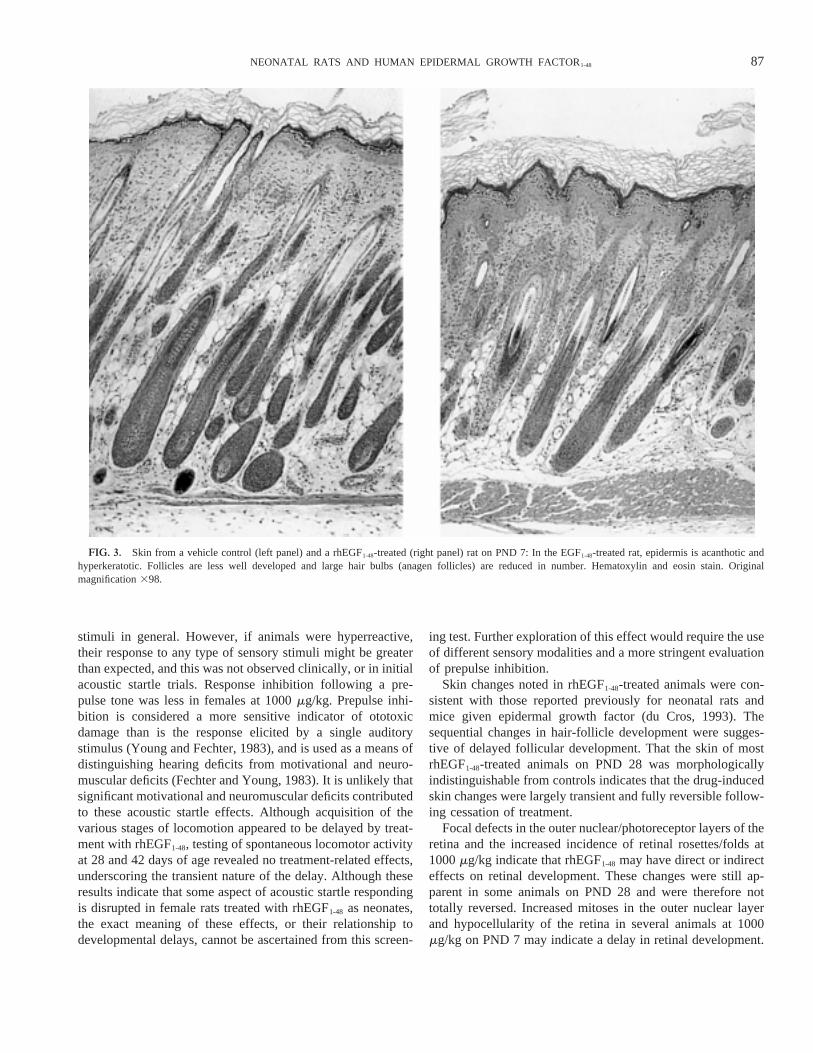
The skin of rhEGF1-48-treated animals had dose-related in-creases in the incidence and intensity of epidermal and follic-ular changes. Epidermal hyperkeratosis and acanthosis, and adecreased proportion of anagen hair follicles relative to cata-gen/telogen follicles occurred at all doses on PND 7 (Fig. 3).These changes were generalized, involving intrascapular (in-jection site) and flank skin at 1000mg/kg. Epidermal hyper-
TABLE 2Acquisition of Developmental Landmarks in Male and Female Rats Given rhEGF1-48
0 mg/kg dose 10mg/kg dose 100mg/kg dose 1000mg/kg dose
Pinnae detachmentMales 2.86 0.12 2.86 0.09 2.66 0.22 2.96 0.23Females 2.66 0.11 2.86 0.14 2.36 0.19 3.06 0.27
Incisor eruptionMales 12.26 0.20 12.76 0.19 10.96 0.37* 10.86 0.29*Females 12.06 0.30 12.26 0.25 10.86 0.48* 10.36 0.33*
Eye openingMales 14.96 0.22 14.66 0.14 11.56 0.12* 9.56 0.28*Females 14.96 0.16 14.66 0.10 11.66 0.14* 9.86 0.32*
Preputial separation 34.36 0.80 33.96 0.94 29.96 1.14* 27.36 0.19*Vaginal opening 32.96 0.89 31.26 0.44 31.16 0.52 20.66 0.63*
Note.Mean6 standard error postnatal day of acquisition. Litter was the unit of measure;n 5 9–10 litters/group.* p , 0.025 (0.05 divided by the square root of 4 individual parameters/sex), different from vehicle control for trend test.
TABLE 3Reflex Development in Male and Female Rats Given rhEGF1-48
0 mg/kg dose 10mg/kg dose 100mg/kg dose 1000mg/kg dose
Static righting reflexMales 3.06 0.21 3.36 0.26 3.16 0.23 3.96 0.26Females 4.26 0.36 3.16 0.28 3.16 0.23 4.06 0.37
Negative geotaxisMales 3.96 0.50 4.36 0.42 4.06 0.33 6.36 0.67*Females 4.26 0.49 4.46 0.50 4.06 0.60 5.26 0.70
Startle reflexMales 12.86 0.13 12.96 0.23 13.36 0.34 14.26 0.32*Females 12.26 0.20 12.36 0.15 12.96 0.35 14.76 0.41*
Wire maneuverMales 14.76 0.40 14.66 0.48 14.06 0.63 15.96 0.31Females 14.26 0.44 15.46 0.37 14.86 0.33 16.26 0.62*
Visual placementMales 18.46 0.43 18.46 0.54 18.96 0.59 20.06 0.44Females 18.96 0.35 17.86 0.59 19.86 0.62 20.56 0.42*
Note.Mean6 standard error postnatal day of acquisition. The individual animal was the unit of measure;n 5 8–10 animals/sex/group.* p , 0.025 (0.05 divided by the square root of 4 individual parameters/sex), different from vehicle control for trend test.
83NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48

keratosis accounted for skin scaling (exfoliation) noted clini-cally. On PND 14, the proportion of anagen to catagen/telogenhair follicles was reduced at 100 and 1000mg/kg, although theincidence of this finding was not as great as on PND 7. Thisfinding corresponded to the clinical observation of thin haircoats at 1000mg/kg. By Day 14, hyperkeratosis was stillevident at 1000mg/kg, and acanthosis was present in only oneanimal. Two 1000-mg/kg animals had increased numbers ofdistorted or deformed hair follicles. On PND 21, an increase inthe proportion of catagen and anagen hair follicles relative totelogen follicles was apparent in rhEGF1-48-treated rats at alldoses. Telogen follicles predominated in controls at this time.By PND 28, the skin of all but one rat at 1000mg/kg wasindistinguishable from control.
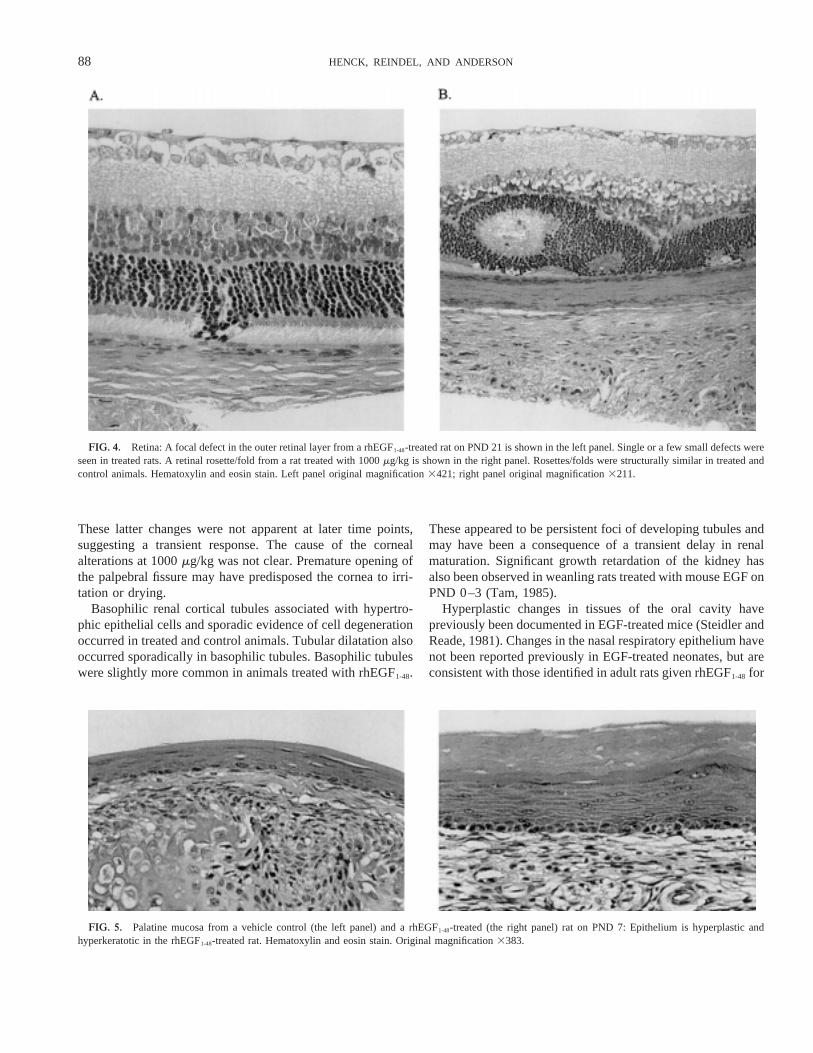
Retinal and corneal lesions were first evident on PND 7 and14, respectively. On PND 7, the retinal nuclear layers of rats at1000mg/kg were diffusely hypocellular and/or had increased
mitoses in the outer nuclear layers (delayed maturation). Theincidence of developmental malformations of the retina, clas-sified as rosettes/folds (peripheral and central), was also in-creased at 1000mg/kg on PND 7 through 28. Additional retinalmalformations, consisting of cell nuclei from the outer nuclearlayer in the photoreceptor layer, occurred in several animals at1000mg/kg on PND 14 and 21 (Fig. 4). Corneal neovascular-ization, neutrophilic inflammation, and reactive epithelial hy-perplasia were noted on PND 14 at 1000mg/kg. Cornealfindings corresponded to dry corneas noted grossly in severalrats. Neovascularization and neutrophilic inflammation werestill present in some animals at 1000mg/kg terminated on PND21 and/or 28.
Hypercellular epithelial cell foci, reminiscent of immaturerenal tubules, were observed in the renal cortex of rats at 1000mg/kg on PND 14–28. Multifocal basophilic renal tubules,indicative of slight tubular degeneration occurred with in-creased incidence and frequency at 1000mg/kg in rats termi-nated on PND 14–28.
Diffuse epithelial hyperplasia, hypertrophy, and hyperkera-tosis were apparent in the lingual and palatine mucosa of allanimals at 1000mg/kg (Fig. 5). Mucosal thickness was at least1.5- to 2-fold greater than control. Nasal respiratory epitheliumwas also hyperplastic at 1000mg/kg (Fig. 6). Respiratorymucosal height was increased and epithelial cells were hyper-trophic; goblet cells were increased in size and number withinthe pseudostratified epithelium. In lung, the cellularity of al-veolar septa of rhEGF1-48-treated animals was reduced com-pared to controls at 1000mg/kg (Fig. 7). Furthermore, alveoliwere larger in size and septal development appeared to be at amore advanced stage than in controls.
A slight increase in intensity and incidence of microvesicu-lar cytoplasmic vacuolation of hepatocytes occurred at 1000mg/kg on PND 7 and correlated with pale livers noted grossly.Liver weight relative to body weight was significantly in-creased in females at 1000mg/kg. Histological correlates forthe significant increase in small intestinal weight in males andfemales at 1000mg/kg were not apparent, and no treatment-related changes were apparent in the length of the small or
FIG. 2. Mean maximum input voltage across 20 noise level (120 dB) trialsin an acoustic startle paradigm. The individual animal was the unit of measure;n 5 9–10 animals/sex/group.
TABLE 4Percent Response Inhibition Resulting from a Prepulse Acoustic
Startle Stimulus in Male and Female Rats Given rhEGF1-48
0 mg/kg dose 10mg/kg dose 100mg/kg dose 1000mg/kg dose
Male 496 3.1 586 2.4 556 3.0 486 2.9Female 606 2.8 526 2.0 576 1.6 276 1.5
Note.Calculated as mean6 standard error percent response inhibition of themaximum input voltage over 20 alternating noise level (NL) and prepulse(PRE) trials, using the formula 100 – [(maximum input voltage PRE trials/maximum input voltage NL trial)3 100]; n 5 9–10 animals/sex/group.
FIG. 1. Mean6 standard error day of acquisition of locomotion Stages 1through 4. Stage 15 pivoting, Stage 25 forward locomotion with head andbody low, Stage 35 forward locomotion with head low and body raised, andStage 45 forward locomotion with head and body raised. Litter was the unitof measure;n 5 8–10 litters/group. Results from males-only are presentedbecause the response of males and females was similar.
84 HENCK, REINDEL, AND ANDERSON

large intestines. Drug-related microscopic alterations were notevident in stomach or small and large intestines.
DISCUSSION
rhEGF1-48-associated clinical observations occurred only at1000mg/kg and were either expected pharmacological effectsrelated to the skin (du Cros, 1993) or considered a consequenceof precocious eye opening. Body weight-gain suppression wasconsistent with previous reports in which rodents treated asneonates with various forms of EGF had growth retardation(Calamandrei and Alleva, 1989; Cohen, 1962; Hoath, 1986;Tam, 1985). Precocious incisor eruption and eye opening werealso expected, based on previous reports (Calamandrei andAlleva, 1989; Cohen, 1962; da Silvaet al., 1991; Hoath, 1986;Smart et al., 1989; Tam, 1985). Pinna detachment was notdelayed in the present study, as was observed in previousstudies, likely due to timing differences. In the present study,treatment was initiated on PND 1, rather than on PND 0, aswith previous studies.
Cleavage of the balanopreputial gland and vaginal opening,considered indicators of puberty in the rat, were accelerated inrhEGF1-48-treated animals. The influence of EGF on cleavage
of the balanopreputial gland has not been reported previously.Precocious vaginal opening has been reported in hamstersgiven subcutaneous injections of human EGF during the sec-ond postnatal week (advanced by 3 days) and in rats givensubcutaneous injections of human EGF during late gestation(advanced by 5–6 days) (da Silvaet al., 1991; Smartet al.,1989). Absence of effects on ovarian and uterine weights onthe day of vaginal opening in EGF-treated rats suggested thatadvancement of this developmental landmark was not due tosexual maturation, but rather to the altered development of theperineal epithelium (da Silvaet al., 1991). This idea wassupported by the work of Nelsonet al. (1991), who determinedthat 750 ng mouse EGF implanted in a slow-release capsule inovariectomized adult mice induced vaginal keratinization.Similar effects induced by estrogen were inhibited by an EGF-specific antibody, implicating EGF as a mediator of estrogenicaction in mice. Vaginal opening occurred earlier followingneonatal treatment in the present study than in previous rodentstudies, which treated subjects during late gestation or thesecond postnatal week. These differences suggest that thedegree of acceleration of vaginal opening depends on thedevelopmental stage of exposure.
Delayed acquisition of reflexes, as noted in the present
TABLE 5Postnatal Day 7 Relative Organ Weights and Lengths in Male and Female Rats Given rhEGF1-48
0 mg/kg dose 10mg/kg dose 100mg/kg dose 1000mg/kg dose
Body weight (g)Males 13.76 0.70 16.56 1.04 14.66 1.57 12.96 0.99Females 13.26 0.79 14.26 0.51 13.96 0.73 12.56 0.43
Organ weights (g/100 g bw)Brain
Males 6.046 0.248 5.256 0.216 5.836 0.508 5.696 0.195Females 5.906 0.160 5.556 0.122 5.856 0.148 5.806 0.128
KidneysMales 1.486 0.071 1.236 0.026 1.316 0.043 1.286 0.038Females 1.416 0.033 1.336 0.037 1.296 0.036 1.376 0.042
Small intestineMales 3.876 0.198 3.706 0.127 4.356 0.146 5.116 0.108*Females 4.086 0.168 3.936 0.050 4.356 0.113 5.326 0.060*
Large intestineMales 0.656 0.037 0.556 0.010 0.516 0.014 0.596 0.077Females 0.626 0.020 0.606 0.038 0.566 0.023 0.576 0.027
LiverMales 3.056 0.189 2.906 0.113 3.026 0.122 3.496 0.102Females 2.966 0.100 2.886 0.074 3.126 0.081 3.676 0.073*
Organ lengths (cm)Small intestine
Males 22.16 0.75 25.76 1.47 24.26 1.25 21.86 0.90Females 21.46 0.83 24.16 0.74 25.36 0.74 22.36 1.05
Large intestineMales 3.36 0.14 3.76 0.024 3.36 0.08 2.86 0.11*Females 3.46 0.18 3.66 0.19 3.36 0.15 3.06 0.03
Note.Mean6 standard error. The individual animal was the unit of measure;n 5 10 animals/sex/group; bw, body weight.* p , 0.001, different from vehicle control for trend test.
85NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48

study, has been reported in studies with several forms of EGF(Calamandreiet al., 1989; Smartet al., 1989; da Silvaet al.,1991). Delayed acquisition of the acoustic startle reflex is ofparticular interest because altered responses were still detectedin females at PND 42. These effects in young adults were
manifested as failures to reduce responding to repeated acous-tic stimuli, and to inhibit responding following a prepulse tone.Failure to decrease responding to repeated stimuli might be theresult of disruption of habituation (considered a measure ofsimple learning), or might result from enhanced reactivity to
TABLE 6Histopathologic Findings in Male and Female Rats Given rhEGF1-48
0 mg/kg dose 10mg/kg dose 100mg/kg dose 1000mg/kg dose
M F M F M F M F
Eye(s)Cornea: neovascularization — — — — — — 2/6 (14) 2/6 (14)
— — — — — — 2/5 (21) 3/8 (21)— — — — — — 2/4 (28) 3/5 (28)
Cornea: neutrophilic inflammation — — — — — — 1/6 (14) 2/6 (14)— — — — — — 1/5 (21) 1/8 (21)— — — — — — — —
Retina: hypocellular — — — — — — 2/5 (7) 1/5 (7)Cornea: reactive hyperplasia — — — — — — 1/6 (14) 1/6 (14)Retina: mitosis increased, outer nuclear
layer — — — — — — 4/5 (7) 3/5 (7)Retina: rosette 2/5 (7) 2/5 (7) 1/5 (7) 1/5 (7) 1/5 (7) 1/5 (7) 4/5 (7) 4/5 (7)
1/5 (14) 1/5 (14) — — 2/5 (21) 1/5 (14) 5/6 (14) 4/6 (14)2/5 (21) 2/5 (21) 1/5 (21) — 1/5 (21) — 4/5 (21) 4/8 (21)
— 2/4 (28) 1/5 (28) 2/6 (28) — 2/5 (28) 4/4 (28) 4/5 (28)Retina: cell nuclei in photoreceptor layer — — — — — — 3/6 (14) 4/6 (14)
— — — — — — 2/5 (21) 6/8 (21)— — — — 1/5 (28) — 3/4 (28) 2/5 (28)
Hard palate: epithelial hyperplasia — — NE NE NE NE 5/5 (7) 5/5 (7)Kidney(s)
Hypercellular focus, renal cortex — — — — — — — 3/5 (14)— — — — 1/5 (21) — 3/5 (21) 3/5 (21)— — — — — — — 1/5 (28)
Tubular basophilia — 2/5 (14) 1/5 (14) — 1/5 (14) — 2/5 (14) 3/5 (14)2/5 (21) 4/5 (21) 2/5 (21) 2/5 (21) 2/5 (21) 2/5 (21) 5/5 (21) 5/5 (21)3/5 (28) 2/5 (28) 4/5 (28) 3/6 (28) 1/5 (28) 3/5 (28) 3/4 (28) 5/5 (28)
Tubular dilatation 3/5 (14) 3/5 (14) 1/5 (14) 2/5 (14) 3/5 (14) 2/5 (14) 5/5 (14) 4/5 (14)1/5 (21) — 1/5 (21) 2/5 (21) — 1/6 (21) 4/5 (21) 2/8 (21)
— 1/4 (28) — 1/5 (28) — — — 2/5 (28)Liver: cytoplasmic vacuolation 3/5 (7) — 2/5 (7) 2/5 (7) 3/5 (7) 3/5 (7) 7/8 (7) 6/6 (7)Lung: hypocellular — — NE NE NE NE 5/5 (7) 5/5 (7)Nasal cavity: epithelial hyperplasia — — NE NE NE NE 5/5 (7) 5/5 (7)Skin
Epidermal acanthosis 1/5 (7) 2/5 (7) 2/6 (7) 1/7 (7) 6/6 (7) 7/8 (7) 10/10 (7) 8/8 (7)— — — — — — 1/8 (14) —
Hair follicle: Decreased anagen — — 2/6 (7) — 3/6 (7) 6/8 (7) 10/10 (7) 8/8 (7)— — — — 2/5 (14) 1/5 (14) 6/8 (14) 6/8 (14)
Hair follicle: Increased catagen — — 1/5 (21) 1/5 (21) — 3/5 (21) 5/5 (21) 5/5 (21)— — — — — — 1/4 (28) —
Epithelial hyperplasia, increased/irregulardebris in lumen — — — — — — 1/8 (14) 1/8 (14)
Hyperkeratosis — 1/5 (7) 4/6 (7) 4/7 (7) 6/6 (7) 8/8 (7) 10/10 (7) 8/8 (7)— — — — — — 6/8 (14) 1/8 (14)
Mononuclear infiltrate 1/5 (7) 5/5 (7) 6/6 (7) 6/7 (7) 6/6 (7) 8/8 (7) 6/10 (7) 8/10 (7)— — — — — — 1/8 (14) —— 1/5 (21) — — — — — —— 1/4 (28) — 1/5 (28) 1/5 (28) 2/5 (28) — 1/5 (28)
Tongue: epithelial hyperplasia — — NE NE NE NE 5/5 (7) 5/5 (7)
Note.Data expressed as number affected/number examined (postnatal day of necropsy); —, finding not observed; NE, not evaluated.
86 HENCK, REINDEL, AND ANDERSON
stimuli in general. However, if animals were hyperreactive,their response to any type of sensory stimuli might be greaterthan expected, and this was not observed clinically, or in initialacoustic startle trials. Response inhibition following a pre-pulse tone was less in females at 1000mg/kg. Prepulse inhi-bition is considered a more sensitive indicator of ototoxicdamage than is the response elicited by a single auditorystimulus (Young and Fechter, 1983), and is used as a means ofdistinguishing hearing deficits from motivational and neuro-muscular deficits (Fechter and Young, 1983). It is unlikely thatsignificant motivational and neuromuscular deficits contributedto these acoustic startle effects. Although acquisition of thevarious stages of locomotion appeared to be delayed by treat-ment with rhEGF1-48, testing of spontaneous locomotor activityat 28 and 42 days of age revealed no treatment-related effects,underscoring the transient nature of the delay. Although theseresults indicate that some aspect of acoustic startle respondingis disrupted in female rats treated with rhEGF1-48 as neonates,the exact meaning of these effects, or their relationship todevelopmental delays, cannot be ascertained from this screen-
ing test. Further exploration of this effect would require the useof different sensory modalities and a more stringent evaluationof prepulse inhibition.
Skin changes noted in rhEGF1-48-treated animals were con-sistent with those reported previously for neonatal rats andmice given epidermal growth factor (du Cros, 1993). Thesequential changes in hair-follicle development were sugges-tive of delayed follicular development. That the skin of mostrhEGF1-48-treated animals on PND 28 was morphologicallyindistinguishable from controls indicates that the drug-inducedskin changes were largely transient and fully reversible follow-ing cessation of treatment.
Focal defects in the outer nuclear/photoreceptor layers of theretina and the increased incidence of retinal rosettes/folds at1000mg/kg indicate that rhEGF1-48 may have direct or indirecteffects on retinal development. These changes were still ap-parent in some animals on PND 28 and were therefore nottotally reversed. Increased mitoses in the outer nuclear layerand hypocellularity of the retina in several animals at 1000mg/kg on PND 7 may indicate a delay in retinal development.
FIG. 3. Skin from a vehicle control (left panel) and a rhEGF1-48-treated (right panel) rat on PND 7: In the EGF1-48-treated rat, epidermis is acanthotic andhyperkeratotic. Follicles are less well developed and large hair bulbs (anagen follicles) are reduced in number. Hematoxylin and eosin stain. Originalmagnification398.
87NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48
These latter changes were not apparent at later time points,suggesting a transient response. The cause of the cornealalterations at 1000mg/kg was not clear. Premature opening ofthe palpebral fissure may have predisposed the cornea to irri-tation or drying.
Basophilic renal cortical tubules associated with hypertro-phic epithelial cells and sporadic evidence of cell degenerationoccurred in treated and control animals. Tubular dilatation alsooccurred sporadically in basophilic tubules. Basophilic tubuleswere slightly more common in animals treated with rhEGF1-48.
These appeared to be persistent foci of developing tubules andmay have been a consequence of a transient delay in renalmaturation. Significant growth retardation of the kidney hasalso been observed in weanling rats treated with mouse EGF onPND 0–3 (Tam, 1985).
Hyperplastic changes in tissues of the oral cavity havepreviously been documented in EGF-treated mice (Steidler andReade, 1981). Changes in the nasal respiratory epithelium havenot been reported previously in EGF-treated neonates, but areconsistent with those identified in adult rats given rhEGF1-48 for
FIG. 4. Retina: A focal defect in the outer retinal layer from a rhEGF1-48-treated rat on PND 21 is shown in the left panel. Single or a few small defects wereseen in treated rats. A retinal rosette/fold from a rat treated with 1000mg/kg is shown in the right panel. Rosettes/folds were structurally similar in treated andcontrol animals. Hematoxylin and eosin stain. Left panel original magnification3421; right panel original magnification3211.
FIG. 5. Palatine mucosa from a vehicle control (the left panel) and a rhEGF1-48-treated (the right panel) rat on PND 7: Epithelium is hyperplastic andhyperkeratotic in the rhEGF1-48-treated rat. Hematoxylin and eosin stain. Original magnification3383.
88 HENCK, REINDEL, AND ANDERSON

4 weeks by continuous infusion (Breideret al., 1996). Pulmo-nary changes in rhEGF1-48-treated neonates in this study weresuggestive of enhanced or accelerated maturation of peripherallung parenchyma, and are consistent with findings previouslyobserved in several animal species treated with EGF duringfetal stages of development (Cattertonet al., 1988; St. Georgeet al., 1991; Sundellet al., 1980).
The significance of the pale livers attributed to increasedmicrovesicular cytoplasmic vacuolation of hepatocytes intreated animals is uncertain. This change was only noted in rats
euthanized on PND 7, and was not associated with otherdegenerative or proliferative changes. Cytoplasmic vacuolationalso occurred to a lesser degree in several control animals. Thischange, reflecting fat accumulation in hepatocytes, may havebeen a consequence of the greater interval these animals wereaway from the dams on the day of euthanasia and consequentinanition resulting in immobilization of peripheral fat stores tothe liver (Hathcock, 1985), rather than a drug-related effect.Fatty vacuolation of hepatocytes has been previously reportedin EGF-treated neonatal mice (Heinberget al., 1965).
FIG. 6. Nasal respiratory epithelium from a vehicle control (left panel) and a rhEGF1-48-treated (right panel) rat on PND 7: Increased mucosal height andan increase in size and number of goblet cells are evident in the rhEGF1-48-treated animal. Original magnification3410.
FIG. 7. Lung alveoli epithelium from a vehicle control (left panel) and a rhEGF1-48-treated (right panel) rat on PND 7: Alveolar septal cellularity is reducedand alveolar spaces are larger in the rhEGF1-48-treated rat. Original magnification3402.
89NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48

It is difficult to distinguish whether many of the pathologicobservations were directly attributable to EGF effects on spe-cific tissues, or were due to indirect effects secondary togrowth retardation or EGF-related changes in cardiovascularparameters.
Results of this study indicate that treatment of neonatal ratswith the EGF fragment, rhEGF1-48 resulted in growth retarda-tion, accelerated developmental landmarks, and delayed acqui-sition of reflexes at 1000mg/kg, and mitogenic responses inepidermal tissues at$ 10 mg/kg. These changes correspond tothose resulting from neonatal administration of other forms ofEGF to rodents. With the exception of effects on acousticstartle responding and changes in the retina, treatment-relatedfindings were considered transient.
ACKNOWLEDGMENTS
The authors thank Denise Frahm and Michelle Cole for technical expertise,Walt Bobrowski for preparation of photomicrographs, Jamie Colgin and JoyceZandee for statistical analyses, and Dr. Robert Parker for peer review ofhistopathologic data.
REFERENCES
Breider, M. A., Bleavins, M. R., Reindel, J. F., Gough, A. W., and de la Iglesia,F. A. (1996). Cellular hyperplasia in rats following continuous intravenousinfusion of recombinant human epidermal growth factor.Vet. Pathol. 33,184–194.
Calamandrei, G., and Alleva, E. (1989). Epidermal growth factor has bothgrowth-promoting and growth-inhibiting effects on physical and neurobe-havioral development on neonatal mice.Brain Res. 477,1–6.
Calamandrei, G., Valanzano, A., and Alleva, E. (1993). Growth factors inbehavioral teratology.Ann. 1st Super. Sanita29, 185–196.
Carpenter, G., and Wahl, M. I. (1990). The epidermal growth factor family. InPeptide Growth Factors and Their Receptors(M.B. Sporn and A.D. Rob-erts, Eds.), pp. 69–171. Springer-Verlag, Berlin.
Catterton, W. Z., Escobedo, M. B., Sexson, W. R., Gray, M. E., Sundell,H. W., and Stahlman, M. T. (1988). Effects of epidermal growth factor onlung maturation in fetal rabbits.Am. Pediatr. Res. 13, 104–108.
Cohen, S. (1962). Isolation of a mouse submaxillary gland protein and accel-erating incisor eruption and eyelid opening in the newborn animal.J. Biol.Chem. 237,1555–1562.
da Silva, V. A., Smart, J. L., and McLean, A. E. (1991). Urogastrone-epidermal growth factor and aspects of sexual maturation in female rats asa function of age at treatment. J. Dev. Physiol. 15, 303–307.
du Cros, D. L. (1993). Fibroblast growth factor and epidermal growth factor inhair development.J. Invest. Dermatol. 101,106–113S.
Dunnett, C. W. (1955). A multiple comparison procedure for comparingseveral treatments with a control.J. Am. Stat. Assoc. 50, 1096–1121.
Dunnett, C. W. (1964). New tables for multiple comparisons with a control.Biometrics20, 482–491.
Fechter, L. D., and Young, J. S. (1983). Discrimination of auditory fromnon-auditory toxicity by reflex modulations audiometry: Effects of triethy-tin. Toxicol. Appl. Pharmacol.70, 216–227.
Greenhouse, S. W., and Geisser, S. (1959). On methods in the analysis ofprofile data.Psychometrika32, 95–112.
Guglietta, A., and Sullivan, P. B. (1995). Clinical applications of epidermalgrowth factor.Eur. J. Gastroenterol. Hepatol.7, 945–950.
Haskins, J. R., Breider, M. A., Bleavins, M. R., and de la Iglesia, F. A. (1997).Cellular changes in intestinal mucosa of Wistar rats after epidermal growthfactor1-48. Toxicologist36, 233.
Haskins, J. R., Lalwani, N. D., Bleavins, M. R., Breider, M. A., and de laIglesia, F. A. (1995). Cell proliferation in rat gastrointestinal tract uponcontinuous infusion of recombinant human EGF1–48.FASEB J. 9, 704.
Hathcock, J. N. (1985). Nutrient and non-nutrient effects on drug metabolism.Drug Nutr. Interact. 4, 217–234.
Heinberg, M., Weinstein, I., Leguire, V. S., and Cohen, S. (1965). Theinduction of fatty liver in neonatal animals by a paired protein (EGF) frommouse submaxillary gland.Life Sci. 4, 1625–1633.
Henck, J. W., Frahm, D., and Anderson, J. A. (1996). Validation of automatedbehavioral test systems.Neurotoxicol. Teratol. 18, 189–197.
Hoath, S. B. (1986). Treatment of the neonatal rat with epidermal growthfactor: Differences in time and organ response.Ped. Res. 20, 468–472.
Irwin, S. (1968). Comprehensive observational assessment: Ia. A systematicquantitative procedure for assessing the behavioral and physiologic state ofthe mouse.Psychopharmacologia13, 222–257.
Iversen, S. D., and Iversen, L. L. (1981).Behavioral Pharmacology. OxfordUniversity Press, New York.
Johnson, R. A., and Wichern, D. W. (1982).Applied Multivariate StatisticalAnalysis. Prentice-Hall, New York.
Konturek, S. J., Dembinski, A., Warzecha, Z., Brzozowski, T., and Gregory, H.(1988). Role of epidermal growth factor in healing of gastric duodenal ulcersin rats.Gastroenterology94, 1300–1307.
Korenbrot, C. C., Huhtaniemi, I. T., and Weiner, R. I. (1977). Preputialseparation as an external sign of pubertal development in the male rat.Biol.Reprod.17, 298–303.
Kruskal, W. H., and Wallis, W. A. (1952). Use of ranks in one-criterionvariance analysis. J. Am. Stat. Assoc.47, 583–621.
Moser, V. C., McCormick, J. P., Creason, J. P., and MacPhail, R. C. (1988).Comparison of chlordimeform and carbaryl using a functional observationalbattery.Fundam. Appl. Toxicol. 11, 189–206.
Musroe, T. A., Pierce, G. F., Morishima, C., and Deuel, T. F. (1991). Growthfactors-induced acceleration of tissue repair through direct and inductiveactivities in a rabbit dermal ulcer model.J. Clin. Invest.87, 694–703.
Nelson, K. G., Takahashi, T., Bossert, N. L., Walmer, D. K., and McLachlan,J. A. (1991). Epidermal growth factor replaces estrogen in the stimulation offemale genital-tract growth and differentiation. Proc. Natl. Acad. Sci. U.S.A.88, 21–25.
Park, Y. C. (1985). Nonparametric sequential trend test. Proceedings of theTenth Annual SAS Users Group International Conference, pp. 809–813.SAS, Cary, NC.
Smart, J. L., da Silva, V. A., Malheiros, L. R., Paumgartten, F. J., and Massey,R. F. (1989). Epidermal growth factor advances some aspects of develop-ment but retards others in both rats and hamsters.J. Dev. Physiol. 11,153–158.
Smith, J. M., Spron, M. B., Roberts, A. B., Derynck, R., Winkler, M. E., andGregory, H. (1985). Human transforming growth factor-a causes precociouseye opening in newborn mice.Nature315,515–516.
Steidler, N. E., and Reade, P. C. (1981). Histomorphological effects of epi-dermal growth factor on skin and oral mucosa in neonatal mice.Arch. OralBiol. 25, 37–43.
St. George, J. A., Read, L. C., Cranz, D. L., Tarantal, A. F., George-Nasci-mento, C., and Plopper, C. G. (1991). Effect of epidermal growth factor onthe fetal development of the tracheobronchial secretory apparatus in rhesusmonkeys.Am. J. Respir. Cell Mol. Biol. 4, 95–101.
Sullivan, P. B., Brueton, M. J., Tabara, Z. B., Goodland, R., Lee, C. Y., andWright, N. A. (1991). Epidermal growth factor in necrotizing enteritis.Lancet338,53–54.
90 HENCK, REINDEL, AND ANDERSON

Sundell, H. W., Gray, M. E., Sereniu, F. S., Escobedo, M. B., and Stahlman,M. T. (1980). Effects of epidermal growth factor on lung maturation in fetallambs.Am. J. Pathol. 100,707–725.
Tam, J. P. (1985). Physiological effects of transforming growth factor in thenewborn mouse.Science229,673–675.
Tukey, J. W., Ciminera, J. L., and Heyse, J. F. (1985). Testing the statisticalcertainty of a response to increasing doses of a drug. Biometrics 41,295–301.
U.S. Environmental Protection Agency, Office of Toxic Substances (1985).
Toxic Substances Control Act test guidelines: Final rule. Health effectstesting guidelines. Subpart G, Neurotoxicity.Fed. Reg. 50, 39458–39470.
Young, J. S., and Fechter, L. D. (1983). Reflex inhibition procedures foranimal audiometry: A technique for assessing ototoxicity.J. Acoust. Soc.Am.73, 1686–1693.
Vorhees, C. V., Butcher, R. E., Brunner, R. L., and Sobotka, T. J. (1979). Adevelopmental test battery for neurobehavioral toxicity in rats: A prelimi-nary analysis using monosodium glutamate, calcium carrageenan, and hy-droxyurea. Toxicol. Appl. Pharmacol.50, 267–282.
91NEONATAL RATS AND HUMAN EPIDERMAL GROWTH FACTOR1-48
Copyright © 2022 FDOKUMEN











![Effects of recombinant drug-specific single chain antibody Fv fragment on [ 3H]-desipramine distribution in rats](https://static.fdokumen.com/doc/165x107/631cd3f3d5372c006e04a0a0/effects-of-recombinant-drug-specific-single-chain-antibody-fv-fragment-on-3h-desipramine.jpg)








