
Genetic diversity and host range studies of turnip curly top virus
9
Genetic diversity and host range studies of turnip curly top virus Sara Razavinejad • Jahangir Heydarnejad • Mehdi Kamali • Hossain Massumi • Simona Kraberger • Arvind Varsani Received: 15 August 2012 / Accepted: 27 November 2012 / Published online: 9 December 2012 Ó Springer Science+Business Media New York 2012 Abstract Turnip curly top virus (TCTV) is a unique ge- minivirus that has recently been characterised as infecting turnips in Iran. The genome of TCTV shares \ 68 % pair- wise identity with other geminiviruses and has a genome organisation similar to that of curtoviruses and topocuvirus. The replication-associated protein (Rep) bears the highest similarity to curtovirus Reps (48.5–69.0 %); however, in the case of the capsid protein (CP), the extent of similarity is only 39.5–44.5 %. We constructed an agroinfectious clone of TCTV and undertook host range studies on ten plant species; in three species (turnip, sugar beet and cowpea), we detected infection which presents curly top symptoms in turnip and sugar beet. The efficiency of TCTV infection in agroinoculated turnip plants was 71.7 %, and the infection was successfully transmitted to 80 % of the healthy turnip plants used in the insect trans- mission studies by Circulifer haematoceps under green- house conditions. We also determined the genome sequence of 14 new TCTV isolates from southern Iran isolated from turnips. We observed *13 % diversity amongst all the TCTV isolates and found evidence of recombination in the CP- and Rep-coding regions of the genomes. Keywords Turnip curly top virus Á Curly top disease Á Geminivirus Á Iran Introduction The family Geminiviridae including geminate virions and circular ssDNA genomes is potentially one of the largest catalogued plant virus families. Based on the host range, genome organisation and insect vector, geminiviruses are classified into four genera namely Mastrevirus, Curtovirus, Topocuvirus and Begomovirus [6]. Over the last five years, a variety of novel viruses have been characterised, which potentially represent new genera: Eragrostis curvula streak virus (ECSV) [23], Beet curly top Iran virus (BCTIV) [2], Turnip curly top virus (TCTV) [5], Grapevine cabernet franc-associated virus (GCFaV) [14], and Citrus chlorotic dwarf-associated virus (CCDaV) [16]. Briddon et al. [5] recently reported on TCTV, a novel geminivirus from Iran, which has a genome consisting of four overlapping genes in the complementary sense, similar to those of topocurvirus and curtoviruses to some extent (Fig. 1a). However, the virion sense encodes two genes unlike curtoviruses which encode three, with capsid protein (CP) sharing 39.5–44.5 % identity to those of curtoviruses. Despite the homologous ORFs between TCTV and curt- oviruses in the complementary sense, there is low sequence identity except for C4 which shares *69.4 % amino acid identity with curtoviruses. The C4 protein in curtoviruses is a symptoms determinant protein, and it is not surprising S. Razavinejad Á J. Heydarnejad (&) Á M. Kamali Á H. Massumi Department of Plant Protection, College of Agriculture, Shahid Bahonar University of Kerman, Kerman, Iran e-mail: [email protected] S. Kraberger Á A. Varsani School of Biological Sciences, University of Canterbury, Private Bag 4800, Christchurch 8140, New Zealand A. Varsani Biomolecular Interaction Centre, University of Canterbury, Private Bag 4800, Christchurch 8140, New Zealand A. Varsani Electron Microscope Unit, University of Cape Town, Rondebosch, Cape Town 7701, South Africa 123 Virus Genes (2013) 46:345–353 DOI 10.1007/s11262-012-0858-y
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Genetic diversity and host range studies of turnip curly top virus

Genetic diversity and host range studies of turnip curly top virus
Sara Razavinejad • Jahangir Heydarnejad •
Mehdi Kamali • Hossain Massumi •
Simona Kraberger • Arvind Varsani
Received: 15 August 2012 / Accepted: 27 November 2012 / Published online: 9 December 2012
� Springer Science+Business Media New York 2012
Abstract Turnip curly top virus (TCTV) is a unique ge-
minivirus that has recently been characterised as infecting
turnips in Iran. The genome of TCTV shares \68 % pair-
wise identity with other geminiviruses and has a genome
organisation similar to that of curtoviruses and topocuvirus.
The replication-associated protein (Rep) bears the highest
similarity to curtovirus Reps (48.5–69.0 %); however, in
the case of the capsid protein (CP), the extent of similarity
is only 39.5–44.5 %. We constructed an agroinfectious
clone of TCTV and undertook host range studies on ten
plant species; in three species (turnip, sugar beet and
cowpea), we detected infection which presents curly top
symptoms in turnip and sugar beet. The efficiency of
TCTV infection in agroinoculated turnip plants was
71.7 %, and the infection was successfully transmitted to
80 % of the healthy turnip plants used in the insect trans-
mission studies by Circulifer haematoceps under green-
house conditions. We also determined the genome
sequence of 14 new TCTV isolates from southern Iran
isolated from turnips. We observed *13 % diversity
amongst all the TCTV isolates and found evidence of
recombination in the CP- and Rep-coding regions of the
genomes.
Keywords Turnip curly top virus � Curly top disease �Geminivirus � Iran
Introduction
The family Geminiviridae including geminate virions and
circular ssDNA genomes is potentially one of the largest
catalogued plant virus families. Based on the host range,
genome organisation and insect vector, geminiviruses are
classified into four genera namely Mastrevirus, Curtovirus,
Topocuvirus and Begomovirus [6]. Over the last five years,
a variety of novel viruses have been characterised, which
potentially represent new genera: Eragrostis curvula streak
virus (ECSV) [23], Beet curly top Iran virus (BCTIV) [2],
Turnip curly top virus (TCTV) [5], Grapevine cabernet
franc-associated virus (GCFaV) [14], and Citrus chlorotic
dwarf-associated virus (CCDaV) [16].
Briddon et al. [5] recently reported on TCTV, a novel
geminivirus from Iran, which has a genome consisting of
four overlapping genes in the complementary sense, similar
to those of topocurvirus and curtoviruses to some extent
(Fig. 1a). However, the virion sense encodes two genes
unlike curtoviruses which encode three, with capsid protein
(CP) sharing 39.5–44.5 % identity to those of curtoviruses.
Despite the homologous ORFs between TCTV and curt-
oviruses in the complementary sense, there is low sequence
identity except for C4 which shares *69.4 % amino acid
identity with curtoviruses. The C4 protein in curtoviruses is
a symptoms determinant protein, and it is not surprising
S. Razavinejad � J. Heydarnejad (&) � M. Kamali � H. Massumi
Department of Plant Protection, College of Agriculture, Shahid
Bahonar University of Kerman, Kerman, Iran
e-mail: [email protected]
S. Kraberger � A. Varsani
School of Biological Sciences, University of Canterbury, Private
Bag 4800, Christchurch 8140, New Zealand
A. Varsani
Biomolecular Interaction Centre, University of Canterbury,
Private Bag 4800, Christchurch 8140, New Zealand
A. Varsani
Electron Microscope Unit, University of Cape Town,
Rondebosch, Cape Town 7701, South Africa
123
Virus Genes (2013) 46:345–353
DOI 10.1007/s11262-012-0858-y

that the symptoms presented by TCTV infections are
indeed similar to those presented by curtoviruses. TCTV
induces curly top symptoms; in turnips, additional symp-
toms such as erection and curling inward of leaves, the
infected leaves becoming thick and brittle and swelling of
veins on the lower leaf surfaces with fluid being exuded
from the petioles or midribs on the lower leaf surfaces are
observed [5]. In addition to turnip, radish and four weed
species have been identified as natural hosts of TCTV, and
the virus has been shown to be transmitted by the leaf-
hopper, Circulifer haematoceps, under greenhouse condi-
tions [19]. In order to satisfy the Koch’s postulate, verify
that the symptoms observed in natural infection and
determine the host range of TCTV, we constructed an
infectious clone and undertook infection studies on ten
plant species. We also isolated 14 new TCTV isolates from
turnip samples from Fars province in southern Iran, which
we analysed for phylogenetic relationships, recombination
and basic strain classification.
Materials and methods
Construction of TCTV infectious clone
The TCTV-[IR:Zaf:B11:06] isolate (accession number
GU456685), sampled from turnip-growing farms in Zafar-
Abad (lat.: 29.4018, long.: 52.5838; 20 km south of Shiraz,
Fars Province, Iran) [5], showing inward leaf curling, thick
and brittle leaves, swelling of veins on the lower leaf
surfaces (Fig. 1b), was used as the virus source for con-
struction of an infectious clone.
TCTV cloned into pGEM�-3Zf(?) (Promega Biotech)
plasmid vector (2,981 nucleotides; designated as pGEM2.9)
was digested with KpnI and EcoRI restriction enzymes to
yield a 2,057-bp fragment of TCTV genome containing
replication origin. The 2,057-bp fragment was gel purified
by Qiaquick Gel Extraction Kit (Qiagen, Germany)
according to manufacturer’s protocol and cloned into the
binary vector pGreen0029 [11] restricted with KpnI and
EcoRI to generate a pGreen2.0 recombinant plasmid. A gel-
purified full-length TCTV genome obtained by digestion of
pGEM2.9 plasmid using KpnI restriction enzyme was
cloned into pGreen2.0 digested by the same enzyme. The
resulting plasmid, labelled pGreen5.038, containing partial
dimer of TCTV genome (full-length ? 2057 bp fragment;
Fig. 2a), was used to transform Escherichia coli cells (strain
JM107). Purified construct (pGreen5.038) and pSoup
(helper plasmid) were used simultaneously to transform
Agrobacterium tumefaciens strain C58 using the freeze/
thaw method according to Boulton [3].
Infectivity tests of TCTV construct
Transformed agrobacterium with pGreen5.038 and pSoup
plasmids were grown for 48–72 h at 28 �C on agar plates,
and the harvested bacterial cells were suspended in sterile
water. Inoculation of three leaves turnip (Brassica rapa L.
cv. White globe purple top), cabbage, sugar beet, spinach,
squash, cucumber, cowpea, datura, tobacco and tomato
seedlings (Table 1) were carried out by needle injection
with *10 ll of agrobacterium suspension at the crown,
near the surface of the soil, as described by Grimsley et al.
[10]. Second group of turnip seedlings were inoculated by
A. tumefaciens cultures bearing pGreen plasmid without a
TCTV partial dimer as negative control.
BCTIV
Rep (C1)
Rep (C1 + C2)
MP ? (V3)
Reg ? (V2)
CP (V1)
Rep (C1)
TCTV
Trap ? (C2)
CP (V1)
Ren (C3)
MP ? (V2)
(C4)?
Rep (C1)
(C4) ?
Trap ? (C2)
MP? (V2)
CP (V1)
Ren (C3)
Topocuvirus
Rep (C1)
SD (C4)reg(V2)
CP (V1)
Ren (C3)Trap/ss (C2)
Curtovirus
MP (V3)
A
Vector: Circulifer haematoceps Vector: Circulifer haematoceps
Vector: Circulifer tenellus Vector: Micrutalis malleifera
B
Fig. 1 a Genome organisation of curtoviruses, topocurvirus, BCTIV
and TCTV, b naturally infected turnip to Turnip curly top virus
showing leaf erection, inward rolling of leaf margin and swelling of
veins on the lower leaf surfaces
346 Virus Genes (2013) 46:345–353
123

Agroinoculated plants were kept in a greenhouse
(*12 h of light and *12 h of dark at 25 �C) and moni-
tored for curly top symptoms. Total DNA from inoculated
plants (either partial dimer construct or pGreen plasmid
without insert) was extracted 4 weeks post agroinoculation.
PCR assays were used to confirm TCTV infection in
agroinoculated plants coupled with visual curly top
symptoms presentation.
Total DNA from plant samples was extracted according
to the method described by Zhang et al. [24]. TCTV
infection of the samples was tested by PCR using two
primer pairs TCTV-840-F (GGCCAATGATGCTTTCTGG
KpnI(2430)
SacI(2563)
HindIII(185)
BglII(683)
NdeI(815)
EcoRI(1506)
KpnI(2430) HindIII(185)
BglII(683)
NdeI(815)
EcoRI(1506)
Full-length genome Partial-length genome
SacI(2563)
AFig. 2 a Schematic
representation of linear TCTV
partial dimer construct showing
potential stem loop structure
and cleavage sites of restriction
enzymes. Symptoms of
agroinoculated plants by TCTV
partial construct showing leaf
erection, inward leaf curling and
swelling of veins on the lower
leaf surfaces on turnip (b, c),
inward leaf curling and
production of swelling and
spine-like outgrowths of the
veins on the lower leaf surfaces
on sugar beet (d, e), pustule-like
(f), and necrotic spots (g) on
cowpea
Table 1 Characteristics of agroinoculated plant species by TCTV infectious construct, total number and number of TCTV infected plant
Common name Scientific name Family Number of infected
plant/number of
agroinoculated plants
Turnip Brassica rapa L. cv. White globe purple top Brassicaceae 43/60
Cabbage Brassica oleracea L. cv. capitata Brassicaceae 0/40
Sugar beet Beta vulgaris L. cv. Br1 Chenopodiaceae 2/30
Spinach Spinacia oleracea L. cv. Virofly Chenopodiaceae 0/30
Squash Cucurbita pepo L. cv. Maragheh Cucurbitaceae 0/40
Cucumber Cucumis sativus L. cv. Peto Seed Cucurbitaceae 0/30
Cowpea Vigna unguiculata L. cv. Mashhad Legominosae 15/35
Datura Datura maxima Sesse & Moc. Solanaceae 0/20
Tobacco Nicotiana Clevelendii Gray. Solanaceae 0/10
Tomato Lycopersicon esculentum Mill. cv. Queen Solanaceae 0/35
Virus Genes (2013) 46:345–353 347
123

GA)/TCTV-1594-R (AATATCATGGGACCACCAGGA)
and TCTV-97-F (CTTGGTCGTGTGGTCCCAAAGG)/
TCTV-1594-R which direct to amplify a 775-bp fragment
covering a part of V1, C2 and C3 ORFs and 1,517-bp
fragments covering complete V1, V2 and parts of C2 and
C3 ORFs of the TCTV genome. PCR conditions and
amplification conditions were used as described by
Heydarnejad et al. [12].
Leafhopper transmission of virus from agroinoculated
plants
Instar nymphs and/or adults of C. haematoceps collected
from sesame growing farms in Sirjan (180 km southwest of
Kerman province, Iran) were used for transmission study.
The method described by Heydarnejad et al. [12] was used
to obtain non-viruliferous leafhoppers from the established
colonies of these C. haematoceps. Different nymphal stages
and/or adults of non-viruliferous leafhoppers were used to
transmit TCTV from the infected agroinoculated turnip
plants to the healthy seedlings under greenhouse conditions
(*25 �C). The inoculated plants were examined for TCTV
infection 4 weeks later for curly top symptoms and by PCR
using two sets of primers described above.
Cloning and sequencing of new TCTV isolates
Fourteen turnip samples showing curly top symptoms were
collected from Zafar-Abad (n = 7), Homayejan (n = 4)
and Lapouei (n = 3) in the Fars province of southern Iran.
Total DNA was extracted from the leaves using CTAB
method as described by Zhang et al. [24]. The DNA was
then used as a template for rolling circle amplification
using TempliPhi Amplification Kit (GE Healthcare, USA)
as described by Shepherd et al. [20], and the resulting
concatemers were digested with BglII restriction endonu-
clease to yield unit length genomes. The unit length-
restricted fragments were cloned into a pUC19 plasmid
linearised with BamHI (BglII compatible cloning site). The
resulting clones were sequenced by primer walking at
Macrogen Inc. (South Korea).
Sequence analyses
The sequences of the TCTV isolates were assembled using
DNAMAN (v. 5.2.9). These were aligned with all available
sequences of curtoviruses, topocuvirus, other curto-like
sequences (BCTIV and Spinach curly top Arizona virus,
SCTAV) and ECSV (as an out-group for downstream
phylogenetic analysis) using MUSCLE [8]. Nucleotide
pairwise identities were calculated using SDTv 1.0 [18].
Full genome maximum likelihood phylogenetic trees were
constructed with the substitution model GTR?G4 (deter-
mined at the best-fit model by RDP 4 [17] using PHYML
3.0 [9] with 1,000 bootstrap replicates. Maximum likeli-
hood phylogenetic trees of the protein sequences were
constructed with PHYML 3.0 [9] using LG model of amino
acid substitutions with approximate likelihood-ratio test
(aLRT) for branch support [1]. Recombination analysis
was performed using RDP, BOOTSCAN, MAXCHI,
CHIMERA, SiScan and 3SEQ methods implemented in
RDP4 (v 4.13) [17].
Results
Agroinoculation of monopartite TCTV-cloned genome
Amongst agroinoculated plants, three plant species
including turnip, sugar beet and cowpea reacted to TCTV
inoculation of partial dimer construct, and symptoms
appeared. Typical TCTV symptoms including leaf erec-
tion, inward rolling of leaf margin and swelling of veins on
the lower leaf surfaces (Fig 2b, c) appeared in 71.7 %
(43/60) (Table 1) of the inoculated turnip plants. Symp-
toms were observed 10 days post agroinoculation on turnip
plants. The infection of symptomatic plants was confirmed
by PCR using TCTV-840-F/TCTV-1594-R and TCTV-97-
F/TCTV-1594-R primer pairs. While 775 and 1,517-bp
fragments were amplified in PCR using extracted DNA of
symptomatic plants, no symptoms were observed or TCTV
viral DNA detected in the negative control turnip plants,
inoculated with pGreen without insert.
Relatively lower infection rates of 6.7 % (2/30) were
observed in agroinoculated sugar beet plants with late
symptoms being observed at 6 weeks post agroinoculation.
These symptoms included inward leaf curling, swelling of
veins and vein enation on the lower leaf surfaces (Fig. 2d,
e). Agroinoculation of the cowpea plants resulted in
infection, 10 days post agroinoculation. Pustule-like and/or
necrotic spots were observed around the veins of cotyledon
leaves in 42.9 % (15/35) of the inoculated plants (Fig. 2f;
Table 1). The pustule-like spots gradually changed to
necrotic spots (Fig. 2g) but seldom extended to other
leaves.
Either TCTV viral DNA was not detected or symptoms
were not observed in the following: agroinoculated cab-
bage (Brassica oleracea L. cv. Capitata), spinach (Spinacia
oleracea L. cv. Virofly), squash (Cucurbita pepo L. cv.
Maragheh), cucumber (Cucumis sativus L. cv. Peto Seed),
Jimson weed (Datura maxima Sesse & Moc.), tobacco
(Nicotiana Clevelendii Gray.), and tomato (Lycopersicon
esculentum Mill. cv. Queen) plants.
348 Virus Genes (2013) 46:345–353
123
Vector transmission of TCTV-cloned genome
Circulifer haematoceps was demonstrated to transmit
TCTV (80 %; 16/20) from agroinoculated turnip plants at
to the healthy seedlings under greenhouse conditions
(*25 �C). Symptoms were observed 10 days after the start
of leafhopper feeding, and typical curly top symptoms
similar to those of agroinoculated plants were noted, and
the infection was confirmed by PCR.
Sequences analysis
The TCTV isolates are phylogenetically distinct from curt-
oviruses, topocuvirus, BCTIV and SCTAV. Genome-wide
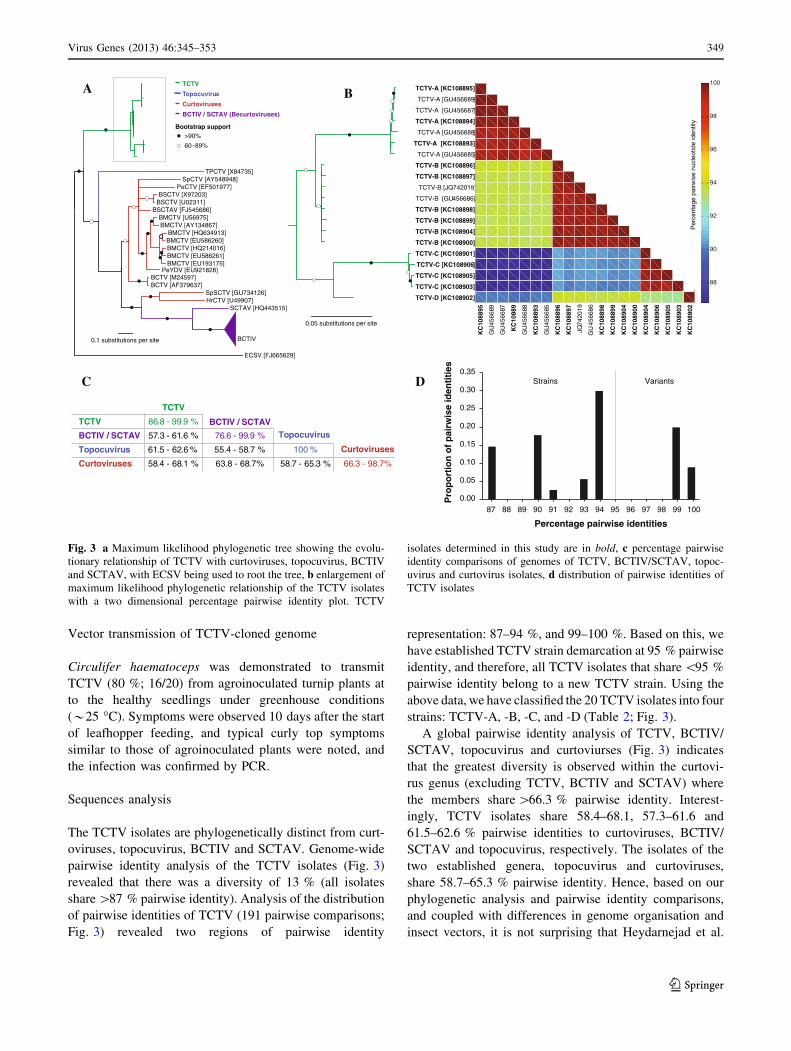
pairwise identity analysis of the TCTV isolates (Fig. 3)
revealed that there was a diversity of 13 % (all isolates
share [87 % pairwise identity). Analysis of the distribution
of pairwise identities of TCTV (191 pairwise comparisons;
Fig. 3) revealed two regions of pairwise identity
representation: 87–94 %, and 99–100 %. Based on this, we
have established TCTV strain demarcation at 95 % pairwise
identity, and therefore, all TCTV isolates that share \95 %
pairwise identity belong to a new TCTV strain. Using the
above data, we have classified the 20 TCTV isolates into four
strains: TCTV-A, -B, -C, and -D (Table 2; Fig. 3).
A global pairwise identity analysis of TCTV, BCTIV/
SCTAV, topocuvirus and curtoviurses (Fig. 3) indicates
that the greatest diversity is observed within the curtovi-
rus genus (excluding TCTV, BCTIV and SCTAV) where
the members share [66.3 % pairwise identity. Interest-
ingly, TCTV isolates share 58.4–68.1, 57.3–61.6 and
61.5–62.6 % pairwise identities to curtoviruses, BCTIV/
SCTAV and topocuvirus, respectively. The isolates of the
two established genera, topocuvirus and curtoviruses,
share 58.7–65.3 % pairwise identity. Hence, based on our
phylogenetic analysis and pairwise identity comparisons,
and coupled with differences in genome organisation and
insect vectors, it is not surprising that Heydarnejad et al.
TCTV-A [KC108895]
TCTV-A [GU456689]
TCTV-A [GU456687]
TCTV-A [KC108894]
TCTV-A [GU456688]
TCTV-A [KC108893]
TCTV-A [GU456685]
TCTV-B [KC108896]
TCTV-B [KC108897]
TCTV-B [JQ742019]
TCTV-B [GU456686]
TCTV-B [KC108898]
TCTV-B [KC108899]
TCTV-B [KC108904]
TCTV-B [KC108900]
TCTV-C [KC108901]
TCTV-C [KC108906]
TCTV-C [KC108905]
TCTV-C [KC108903]
TCTV-D [KC108902]
88
90
92
94
96
98
100
KC
1088
95
GU
4566
89
GU
4566
87
KC
1088
9
GU
4566
88
KC
1088
93
GU
4566
85
KC
1088
96
KC
1088
97
JQ
7420
19
GU
4566
86
KC
1088
98
KC
1088
99
KC
1089
04
KC
1089
00
KC
1089
04
KC
1089
06
KC
1089
05
KC
1089
03
KC
1089
02
>90%
60–89%
TCTV
Curtoviruses
Bootstrap support
0.05 substitutions per site
BCTIV / SCTAV (Becurtoviruses)
Topocuvirus
87 88 89 90 91 92 93 94 95 96 97 98 99 1000.00
0.05
0.10
0.20
0.30
0.35
0.15
0.25
Strains Variants
Percentage pairwise identities
Pro
po
rtio
n o
f p
airw
ise
iden
titi
e s
ECSV [FJ665629]
TPCTV [X84735] SpCTV [AY548948]
PeCTV [EF501977] BSCTV [X97203]
BSCTV [U02311] BSCTAV [FJ545686]
BMCTV [U56975] BMCTV [AY134867]
BMCTV [HQ634913] BMCTV [EU586260] BMCTV [HQ214016] BMCTV [EU586261] BMCTV [EU193175]
BCTV [M24597] BCTV [AF379637]
SpSCTV [GU734126] HrCTV [U49907]
SCTAV [HQ443515]
BCTIV0.1 substitutions per site
Per
cent
age
pairw
ise
nucl
eotid
e id
entit
y
A
D
PeYDV [EU921828]
TCTV
BCTIV / SCTAVTopocuvirus
Curtoviruses
TCTV 86.8 - 99.9 %
BCTIV / SCTAV 57.3 - 61.6 % 76.6 - 99.9 %
Topocuvirus 61.5 - 62.6 % 55.4 - 58.7 % 100 %
Curtoviruses 58.4 - 68.1 % 63.8 - 68.7% 58.7 - 65.3 % 66.3 - 98.7%
C
B
Fig. 3 a Maximum likelihood phylogenetic tree showing the evolu-
tionary relationship of TCTV with curtoviruses, topocuvirus, BCTIV
and SCTAV, with ECSV being used to root the tree, b enlargement of
maximum likelihood phylogenetic relationship of the TCTV isolates
with a two dimensional percentage pairwise identity plot. TCTV
isolates determined in this study are in bold, c percentage pairwise
identity comparisons of genomes of TCTV, BCTIV/SCTAV, topoc-
uvirus and curtovirus isolates, d distribution of pairwise identities of
TCTV isolates
Virus Genes (2013) 46:345–353 349
123

[12] and Briddon et al. [5] proposed that BCTIV and
TCTV be considered as members of new genera (Fig. 3a,
b, c, d).
We found evidence for two putative recombination
events amongst the TCTV isolates. TCTV-B and –D are
recombinants with a domain in the CP region which is most
similar to that of TCTV-C (Fig. 4), whereas TCTV-B is a
double recombinant with a domain spanning the Rep that is
most similar to that of TCTV-C and -D (Fig. 4). Maximum
likelihood phylogenetic analysis of the CP and Rep further
supports this evidence (Fig. 4). TCTV CPs share \44.6,
44.6 and 22.9 % amino acid identity to the CPs of curtovi-
ruses, BCTIV/SCTAV and topocuvirus, respectively
(Fig. 5). On the other hand, the TCTV Reps share \68.9,
30.9 and 62.1 % amino acid identity with the Reps of curt-
oviruses, BCTIV/SCTAV and topocuvirus, respectively.
Discussion
In Iran, three geminiviruses associated with curly top dis-
eases: BSCTV [4], BCTIV [2] and TCTV [5] have been
reported in sugar beet or turnip-growing farms. The bio-
logical properties of TCTV are most similar to those of
members of the genus Curtovirus; however, the genome
organisation is most similar to that of topocuvirus. Further,
phylogenetically, TCTV is distinct and monophyletic [5].
The typical curly top symptoms, leafhopper transmission
by C. haematoceps aligns these characteristics with those
of BCTIV [2, 5, 13, 19]; however, in contrast to BCTIV,
we found TCTV had almost no infectivity in sugar beet
(6.7 % efficiency). These results corroborate with obser-
vations made in surveys of TCTV infection in turnip farms
in Homayejan (50 km west of Shiraz, Fars province, Iran)
where sugar beet was infected with BCTIV [19], thereby
suggesting that sugar beet plants are not a TCTV suitable
host. Our results reveal the limited host range of TCTV
which is similar to that of Horseradish curly top curtovirus
(HrCTV) [15], which, similar to, Spinach severe curly top
virus (SpSCTV) shares only 67.1–70.6 % pairwise identity
with other curtoviruses. Amongst ten plant species agro-
inoculated with TCTV construct, only three plant species
(Table 1) were infected. Furthermore, attempts to find
possible additional hosts of the virus within or around the
severely TCTV-infected farms led to only the virus being
detected in symptomatic radish plants (genome accession #
JQ742019) [19] and four symptomless weed species.
Despite numerous attempts, we were unable to recover any
full TCTV genomes from the four symptomless weed
samples. Nevertheless, more studies are necessary to
identify natural hosts of the virus in cultivated crops and
possible reservoirs in uncultivated plants.
The genome of curtoviruses comprises 6–7 ORFs, three
encoded in the virion sense and 3–4 in the complementary
sense, which separated by one IR [6] and curtoviruses, are
transmitted by the leafhopper Circulifer tenellus (Baker)
Table 2 Details of TCTV isolates, hosts and accession numbers. Isolates determined in this study are in bold
GenBank accession Field id Sampling year Host Location
GU456685 TCTV-A [IR:Zaf:B11:06] 2006 Brassica rapa Zafar Abad, Fars province (southern Iran)
GU456686 TCTV-B [IR:Hom1:2K:09] 2009 Brassica rapa Homayejan, Fars province (southern Iran)
GU456687 TCTV-A [IR: Hom2:8K:09]: 2009 Brassica rapa Homayejan, Fars province (southern Iran)
GU456688 TCTV-A [IR: Hom3:4K:09] 2009 Brassica rapa Homayejan, Fars province (southern Iran)
GU456689 TCTV-A [IR: Hom3:7K:09] 2009 Brassica rapa Homayejan, Fars province (southern Iran)
JQ742019 TCTV-B [IR:Hom:T57K:Tur:10] 2010 Raphanus sativus Homayejan, Fars province (southern Iran)
KC108893 TCTV-A [IR:Lap:L13:Tur:12] 2012 Brassica rapa Lapoui, Fars province (southern Iran)
KC108894 TCTV-A [IR:Zaf:Z8:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108895 TCTV-A [IR:Lap:L2-P:Tur:12] 2012 Brassica rapa Lapoui, Fars province (southern Iran)
KC108896 TCTV-B [IR:Lap:L16:Tur:12] 2012 Brassica rapa Lapoui, Fars province (southern Iran)
KC108899 TCTV-B [IR:Hom:H6:Tur:12] 2012 Brassica rapa Homayejan, Fars province (southern Iran)
KC108898 TCTV-B [IR:Zaf:Z2-14:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108900 TCTV-B [IR:Zaf:Z2-18:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108897 TCTV-B [IR:Zaf:Z9:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108904 TCTV-B [IR:Hom:H8:Tur:12] 2012 Brassica rapa Homayejan, Fars province (southern Iran)
KC108905 TCTV-C [IR:Zaf:Z2-1:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108906 TCTV-C [IR:Zaf:Z5-2:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108901 TCTV-C [IR:Hom:H1:Tur:12] 2012 Brassica rapa Homayejan, Fars province (southern Iran)
KC108903 TCTV-C [IR:Zaf:Z10:Tur:12] 2012 Brassica rapa Zafar Abad, Fars province (southern Iran)
KC108902 TCTV-D [IR:Hom:H9:Tur:12] 2012 Brassica rapa Homayejan, Fars province (southern Iran)
350 Virus Genes (2013) 46:345–353
123

[7, 21, 22]. The TCTV genome has six ORFs, two in viral and
four in complementary strand. Despite having different
genome arrangements in the complementary sense ORFs,
products of all TCTV ORFs, other than V2, are most similar
to the counterpart ORFs of curtoviruses. However, the
complementary-sense ORFs of TCTV other than C4 show
little sequence similarity to their curtovirus homologs. Our
analysis of TCTV isolates revealed that there is *13 %
diversity, and, as with all other geminiviruses, recombina-
tion seems to play a key role in its evolution. The CP of all the
curtoviruses share [80.2 % amino acid identity; however,
they only bear an amino acid identity of 39.5–44.5 % to
those of TCTV. Nevertheless, the Reps of curtoviruses
share [53.1 % amino acid identity; whereas they share
48.5–69.0 % amino acid identity to TCTV Reps, clearly
suggesting that TCTV has a highly divergent CP. It is
interesting to note that in general there is high diversity in the
Rep of curtoviruses relative to TCTV; we presume this to be
as a result of recombination. It is highly likely, that when
coupled with new molecular tools and next generation
sequencing technologies, novel curto-like viruses will be
discovered from the Middle East which seems to be the
centre of curto-like virus diversity in contrast to the New
World, where most of the curtoviruses have been found.
Broad turnip survey in the Middle East will give us a better
indication of the incidence and diversity of TCTV.
PeYDV [EU921828]
BCTIV [EU273817]
BCTIV [EU273817]
PeYDV [EU921828]
0.5 amino acid substitutions per siter
>90%
60–89%
TCTV
Curtoviruses
Bootstrap support
BCTIV / SCTAV (Becurtoviruses)
Topocuvirus
ECSV [FJ665629] TPCTV [X84735]
SpCTV [AY548948]
PeCTV [EF501977]
BSCTV [X97203] BSCTV [U02311]
BSCTAV [FJ545686]
BMCTV [U56975] BMCTV [AY134867] BMCTV [HQ634913] BMCTV [EU586260] BMCTV [HQ214016]
BMCTV [EU586261] BMCTV [EU193175]
BCTV [M24597] BCTV [AF379637]
SpSCTV [GU734126]
HrCTV [U49907] SCTAV [HQ443515]
ECSV [FJ665629] TPCTV [X84735]
SpCTV [AY548948]
PeCTV [EF501977]
BSCTV [X97203] BSCTV [U02311]
BSCTAV [FJ545686]
BMCTV [U56975] BMCTV [AY134867]
BMCTV [HQ634913] BMCTV [EU586260]
BMCTV [HQ214016]
BMCTV [EU586261] BMCTV [EU193175]
BCTV [M24597] BCTV [AF379637]
SpSCTV [GU734126]
HrCTV [U49907]
SCTAV [HQ443515]
TCTV-A [KC108895] TCTV-A [GU456689] TCTV-A [GU456687]
TCTV-A [KC108894 ]
TCTV-A [GU456688]TCTV-A [KC108893] TCTV-A [GU456685]
TCTV-B [KC108896]
TCTV-B [KC108897 ]
TCTV-B [JQ742019]TCTV-B [GU456686]
TCTV-B [KC108898 ]
TCTV-B [KC108899 ] TCTV-B [KC108904 ]
TCTV-B [KC108900 ]
TCTV-C [KC108901] TCTV-C [KC108906] TCTV-C [KC108905 ]
TCTV-C [KC108903]
TCTV-D [KC108902 ]
TCTV-A [KC108895]
TCTV-A [GU456689]
TCTV-A [GU456687] TCTV-A [KC108894 ]
TCTV-A [GU456688]
TCTV-A [KC108893]
TCTV-A [GU456685]
TCTV-B [KC108896] TCTV-B [KC108899 ]
TCTV-B [JQ742019]
TCTV-B [GU456686]
TCTV-B [KC108898 ] TCTV-B [KC108899 ]
TCTV-B [KC108904 ]
TCTV-B [KC108900 ]
TCTV-C [KC108901] TCTV-C [KC108906] TCTV-C [KC108905 ]TCTV-C [KC108903]
TCTV-D [KC108902 ]
0.5 amino acid substitutions per siter
RepCP
BC
TCTV-BTCTV-D
Recombinant
TCTV-C
Potential major parents
Potential minor parent
TCTV-A
Detection methods
RGBMCT
P-value
1.8 x 10- 16
TCTV-B TCTV-CTCTV-D
TCTV-A RGBMCT 2.8 x 10 - 28
Event
II I
I
II
V2 (Mp)
Cp C4
Rep
C2 (trap)
Ren
A
Fig. 4 a Iillustration and details of the recombination events detected
by RDP 4. R, G, B, M, C, S and T indicate detection by the RDP
GENCONV, BOOTSCAN, MAXCHI, CHIMERA, SISCAN and
3SEQ methods, respectively (p value determined by the method
indicated in bold type), b maximum likelihood phylogenetic analysis
of the CP and Rep amino acid sequences of TCTV, curtoviruses,
topocurvirus, BCTIV, SCTAV (ECSV was used to root the CP
phylogenetic tree and BCTIV and SCTAV were used to root the Rep
phylogenetic tree). TCTV isolates determined in this study are in bold
Virus Genes (2013) 46:345–353 351
123

GenBank Accession #
KC108893: TCTV-A [IR: Lap13:Tur:12]
KC108894: TCTV-A [IR: Zaf8:Tur:12]
KC108895: TCTV-A [IR: Lap2-P:Tur:12]
KC108896: TCTV-B [IR: Lap16:Tur:12]
KC108897: TCTV-B [IR: Zaf9:Tur:12]
KC108898: TCTV-B [IR: Zaf2-14:Tur:12]
KC108899: TCTV-B [IR: Hom6:Tur:12]
KC108900: TCTV-B [IR: Zaf2-18:Tur:12]
KC108901: TCTV-C [IR: Hom1:Tur:12]
KC108902: TCTV-D [IR: Hom9:Tur:12]
KC108903: TCTV-C [IR: Zaf10:Tur:12]
KC108904: TCTV-B [IR: Hom8:Tur:12]
KC108905: TCTV-C [IR: Zaf2-1:Tur:12]
KC108906: TCTV-C [IR: Zaf5-2:Tur:12]
Acknowledgments This study was supported by a grant from
Shahid Bahonar University of Kerman, Kerman, Iran and a block
grant from the University of Cape Town, South Africa, awarded to
Arvind Varsani.
References
1. M. Anisimova, O. Gascuel, Syst. Biol. 55, 539–552 (2006)
2. H.R. Bolok Yazdi, J. Heydarnejad, H. Massumi, Virus Genes 36,
539–545 (2008)
3. M. Boulton, Plant Virology Protocols: From Viral Sequence to
Protein Function, in Methods in Molecular Biology, vol. 451, ed.
by G.D. Foster, I.E. Johansen, Y. Hong, P.D. Nagy (Humana,
Totowa, 2008), pp. 503–523
4. R.W. Briddon, D.C. Stenger, I.D. Bedford, J. Stanley, K. Iza-
dpanah, P.G. Markham, Eur. J. Plant Pathol. 104, 77–84 (1998)
5. R.W. Briddon, J. Heydarnejad, F. Khosrowfar, H. Massumi, D.P.
Martin, A. Varsani, Virus Res. 152, 169–175 (2010)
PeYDV [EU921828]
BCTIV [EU273817]
BSCTV [X97203] BSCTV [U02311]
BSCTAV [FJ545686]
BMCTV [U56975] BMCTV [AY134867] BMCTV [HQ634913] BMCTV [EU586260] BMCTV [HQ214016]
BMCTV [EU193175]
BCTV [M24597] BCTV [AF379637]
SpSCTV [GU734126] SCTAV [HQ443515]
ECSV [FJ665629] TPCTV [X84735]
SpCTV [AY548948]
PeCTV [EF501977]
BMCTV [EU586261]
HrCTV [U49907]
TCTV-A [KC108895]
TCTV-A [GU456689]
TCTV-A [GU456687] TCTV-A [KC108894]
TCTV-A [GU456688]
TCTV-A [KC108893]
TCTV-A [GU456685]
TCTV-B [KC108896] TCTV-B [KC108897]
TCTV-B [JQ742019]
TCTV-B [GU456686]
TCTV-B [KC108898] TCTV-B [KC108899]
TCTV-B [KC108904]
TCTV-B [KC108900]
TCTV-C [KC108901] TCTV-C [KC108906] TCTV-C [KC108905]TCTV-C [KC108903]
TCTV-D [KC108902]
PeY
DV
[EU
9218
28]
BC
TIV
[EU
2738
17]
BS
CT
V [X
9720
3]B
SC
TV
[U02
311]
BS
CTA
V [F
J545
686]
BM
CT
V [U
5697
5]B
MC
TV
[AY
134
867]
BM
CT
V [H
Q63
4913
]B
MC
TV
[EU
586
260]
BM
CT
V [H
Q21
4016
]
BM
CT
V [E
U19
3175
]
BC
TV
[M24
597]
BC
TV
[AF3
7963
7]
SpS
CT
V [G
U73
4126
]S
CTA
V [H
Q44
3515
]
EC
SV
[FJ6
6562
9]T
PC
TV
[X84
735]
SpC
TV
[AY
548
948]
PeC
TV
[EF5
0197
7]
BM
CT
V [E
U5
8626
1]
HrC
TV
[U49
907]
TC
TV-
A [
KC
1088
95]
TC
TV
-A [G
U45
6689
]
TC
TV
-A [
GU
4566
87]
TC
TV-
A [
KC
1088
94]
TC
TV
-A [G
U45
6688
]
TC
TV-
A [
KC
1088
93]
TC
TV
-A [G
U45
6685
]
TC
TV-
B [
KC
1088
96]
TC
TV-
B [
KC
1088
97]
TC
TV
-B [J
Q74
2019
]
TC
TV
-B [
GU
4566
86]
TC
TV-
B [
KC
1088
98]
TC
TV-
B [
KC
1088
99]
TC
TV-
B [
KC
1089
04]
TC
TV-
B [
KC
1089
00]
TC
TV-
C [
KC
1089
01]
TCT
V-C
[K
C10
8906
]T
CT
V-C
[K
C10
8905
]T
CT
V-C
[K
C10
8903
]
TC
TV-
D [
KC
1089
02]
20
30
40
50
60
70
80
90
100
Perc
enta
ge a
min
o ac
id id
entit
y
CP
Rep
TC
TV
Cur
tovi
ruse
s
BCTIV / SCTAV
Topocuvirus
Fig. 5 Two dimensional percentage amino acid identity plot of the CP and Rep of TCTV isolates with curtoviruses, BCTIV, SCTAV,
topocuvirus and ECSV. Isolates determined in this study are in bold
352 Virus Genes (2013) 46:345–353
123

6. J.K. Brown, C.M. Fauquet, R.W. Briddon, M. Zerbini, E. Mori-
ones, J. Navas-Castillo, in Virus Taxonomy: Classification andNomenclature of Viruses: Ninth Report of the InternationalCommittee on Taxonomy of Viruses, ed. by A.M.Q. King, M.J.
Adams, E.B. Carstens, E.J. Lefkowitz (Elsevier Academic Press,
San Diego, 2011), pp 251-374
7. L.F. Chen, K. Brannigan, R. Clark, R.L. Gilbertson, Plant Dis. 94,
99–108 (2010)
8. R.C. Edgar, Nucleic Acids Res. 32, 1792–1797 (2004)
9. S. Guindon, J.F. Dufayard, V. Lefort, M. Anisimova, W. Hordijk,
O. Gascuel, Syst. Biol. 59(3), 307–321 (2010)
10. N. Grimsley, B. Hohn, T. Hohn, R. Walden, Proc. Natl. Acad.
Sci. USA 83, 3282–3286 (1986)
11. R.P. Hellens, E.A. Edwards, N.R. Leyland, S. Bean, P.M. Mul-
lineaux, Plant Mol. Biol. 42, 819–832 (2000)
12. J. Heydarnejad, N. Keyvani, S. Razavinejad, H. Massumi, A.
Varsani, Arch. Virol. (2012). doi:10.1007/s00705-012-1485-6
13. E. Hosseini Abhari, J. Heydarnejad, H. Massumi, A. Hosseini
Pour, K. Izadpanah, In Proceedings of the Second Asian Con-ference on Plant Pathology (National University of Singapore,
Singapore, 2005), p. 62
14. B. Krenz, J.R. Thompson, M. Fuchs, K.L. Perry, J. Virol. 86,
771515 (2012)
15. K.A. Klute, S.A. Nadler, D.C. Stenger, J. Gen. Virol. 77,
1369–1378 (1996)
16. G. Loconsole, P. Saldarelli, H. Doddapaneni, V. Savino, G.P.
Martelli, M. Saponari, Virology 432, 162–172 (2012)
17. D.P. Martin, P. Lemey, M. Lott, V. Moulton, D. Posada, P. Le-
feuvre, Bioinformatics 26, 2462–2463 (2010)
18. B. Muhire, D.P. Martin, J.K. Brown, J. Navas-Castillo, E. Mor-
iones, F.M. Zerbini, R. Rivera-Bustamante, V.G. Malathi, R.W.
Briddon, A. Varsani, Arch. Virol. (Ref. AVIROL-D-12-00641R1)
(In press)
19. S. Razavinejad, J. Heydarnejad, Iran. Plant Pathol. J. (2012) (In
press)
20. D.N. Shepherd, D.P. Martin, P. Lefeuvre, A.L. Monjane, B.E.
Owor, E.P. Rybicki, A. Varsani, J. Virol. Methods 149, 97–102
(2008)
21. M.J. Soto, M.L. Gilbertson, Phytopathology 93, 479–484 (2003)
22. M.J. Soto, L.F. Chen, Y.S. Seo, R.L. Gilbertson, Virology 341,
257–270 (2005)
23. A. Varsani, D.M. Shepherd, K. Dent, A.L. Monjane, E.P. Ryb-
icki, D.P. Martin, Virol. J. 6, 36 (2009)
24. Y.P. Zhang, J.K. Uyemoto, B.C. Kirkpatrick, J. Virol. Methods
71, 45–50 (1998)
Virus Genes (2013) 46:345–353 353
123




















