
Discriminated Induction of SOS Functions in Escherichia coli by Alkylating Agents
Upload
independentCategory
view
3download
0
Genes That Are Uniquely Stress Regulated in Salt OverlySensitive (sos) Mutants1
Zhizhong Gong2, Hisashi Koiwa, Mary Ann Cushman, Anamika Ray, Davi Bufford, Shin Kore-eda,Tracie K. Matsumoto, Jianhua Zhu, John C. Cushman, Ray A. Bressan, and Paul M. Hasegawa*
Center for Plant Environmental Stress Physiology, 1165 Horticulture Building, Purdue University, WestLafayette, Indiana 47907–1165 (Z.G., H.K., T.K.M., J.Z., R.A.B., P.M.H.); Department of Biochemistry MS200,University of Nevada, Reno, Nevada 89557–0014 (M.A.C., S.K., J.C.C.); and Department of Biochemistry andMolecular Biology, Oklahoma State University, Stillwater, Oklahoma 74078 (A.R., D.B.)
Repetitive rounds of differential subtraction screening, followed by nucleotide sequence determination and northern-blotanalysis, identified 84 salt-regulated (160 mm NaCl for 4 h) genes in Arabidopsis wild-type (Col-0 gl1) seedlings. Probescorresponding to these 84 genes and ACP1, RD22BP1, MYB2, STZ, and PAL were included in an analysis of salt responsivegene expression profiles in gl1 and the salt-hypersensitive mutant sos3. Six of 89 genes were expressed differentially inwild-type and sos3 seedlings; steady-state mRNA abundance of five genes (AD06C08/unknown, AD05E05/vegetative storageprotein 2 [VSP2], AD05B11/S-adenosyl-l-Met:salicylic acid carboxyl methyltransferase [SAMT], AD03D05/cold regulated6.6/inducible2 [COR6.6/KIN2], and salt tolerance zinc finger [STZ]) was induced and the abundance of one gene (AD05C10/circadian rhythm-RNA binding1 [CCR1]) was reduced in wild-type plants after salt treatment. The expression of CCR1,SAMT, COR6.6/KIN2, and STZ was higher in sos3 than in wild type, and VSP2 and AD06C08/unknown was lower in themutant. Salt-induced expression of VSP2 in sos1 was similar to wild type, and AD06C08/unknown, CCR1, SAMT, COR6.6/KIN2, and STZ were similar to sos3. VSP2 is regulated presumably by SOS2/3 independent of SOS1, whereas the expressionof the others is SOS1 dependent. AD06C08/unknown and VSP2 are postulated to be effectors of salt tolerance whereas CCR1,SAMT, COR6.6/KIN2, and STZ are determinants that must be negatively regulated during salt adaptation. The pivotalfunction of the SOS signal pathway to mediate ion homeostasis and salt tolerance implicates AD06C08/unknown, VSP2,SAMT, 6.6/KIN2, STZ, and CCR1 as determinates that are involved in salt adaptation.
High soil salinity, which is caused typically byNaCl, results in ion toxicity and hyperosmotic stressleading to numerous pathologies including genera-tion of reactive oxygen species (ROS) and pro-grammed cell death (Niu et al., 1995; Zhu et al., 1997;Hasegawa et al., 2000b). Salt tolerance determinantsare categorized either as effectors that directly mod-ulate stress etiology or attenuate stress effects, or asregulatory molecules that are involved in stress per-ception, signal transduction, or modulation of effec-tor function. Enzymes that catalyze rate-limitingsteps in the biosynthesis of compatible solutes orosmoprotectants, e.g. sugar alcohol, quaternary am-monium, and tertiary sulfonium compounds, are cat-egorical examples of osmotic stress tolerance effec-tors (Hanson et al., 1994; Ishitani et al., 1997; Yoshibaet al., 1997; Nelson et al., 1998; Hasegawa et al.,2000b). Other effectors include proteins that protectmembrane integrity, control water or ion homeosta-sis, and ROS scavenging (Bray, 1994; Ingram and
Bartels, 1996; Hasegawa et al., 2000b). Determinantfunction of some effectors has been confirmed be-cause expression in transgenic plants enhances salttolerance sufficiency (Hasegawa et al., 2000b).
Regulatory determinants include transcription fac-tors and signal pathway intermediates that posttran-scriptionally activate effectors (Hasegawa et al.,2000b). Basic Leu zipper motif, MYB and MYC, andzinc finger transcription factors, including rd22BP1(MYC), AtMYB2 (MYB), DREB1A, and DREB2A (AP2domain), and ALFIN1 (zinc finger), interact with pro-moters of osmotic-regulated genes (Abe et al., 1997;Liu et al., 1998; Hasegawa et al., 2000b). The osmoticstress tolerance function of DREB1A in Arabidopsis(Kasuga et al., 1999) and ALFIN1 in alfalfa (Medicagosativa; Winicov, 2000) has been confirmed by ectopicexpression in transgenic plants. Regulatory interme-diates that modulate plant salt stress responses in-clude SOS3 (Ca21-binding protein), SOS2 (Suc non-fermenting-like) kinase, Ca21-dependent proteinkinases, and mitogen-activated protein kinases(Sheen, 1996; Halfter et al., 2000; Kovtun et al., 2000).Additional signal intermediates have been impli-cated in the plant response to salt (Hwang and Good-man, 1995; Mizoguchi et al., 1996; Mikami et al., 1998;Piao et al., 1999; Hasegawa et al., 2000b).
The Zhu laboratory recently has pioneered identi-fication of salt tolerance determinants using forward
1 This work was supported by a National Science FoundationPlant Genome award (no. DBI–9813360). This is Purdue UniversityAgricultural Experiment Station paper no. 16427.
2 Present address: Department of Plant Sciences, University ofArizona, Tucson, AZ 85721.
* Corresponding author; e-mail [email protected];fax 765–494–0391.
Plant Physiology, May 2001, Vol. 126, pp. 363–375, www.plantphysiol.org © 2001 American Society of Plant Physiologists 363

genetics in the plant model Arabidopsis (Hasegawaet al., 2000a; Sanders, 2000; Zhu, 2000). This effort hasidentified three complementation groups of ion hy-persensitive (salt overly sensitive [sos1-sos3]) mu-tants. Genetic and physiological data indicate thatSOS3, SOS2, and SOS1 are components of a signalpathway that regulates ion homeostasis and salt tol-erance and their functions are Ca21 dependent. Po-sitional cloning revealed that SOS1 encodes a puta-tive plasma membrane Na1/H1 antiporter, SOS2encodes a Suc non-fermenting-like (SNF) kinase, andSOS3 encodes a Ca21-binding protein with sequencesimilarity to the regulatory subunit of calcineurinand neuronal Ca21 sensors (Liu and Zhu, 1998; Liu etal., 2000; Shi et al., 2000). Molecular interaction andcomplementation analyses indicate that SOS3 is re-quired for activation of SOS2 that regulates SOS1transcription (Halfter et al., 2000; Shi et al., 2000),further confirming that the order of the signal path-way is SOS3 ¡ SOS2 ¡ SOS1 (Hasegawa et al.,2000a; Sanders, 2000; Zhu, 2000).
Herein is described a functional dissection of theplant salt stress response by analysis of gene expres-sion controlled by the SOS signal pathway. Differen-tial subtraction and northern analysis identified 84salt-regulated genes, the majority of which have notbeen annotated to be salt responsive in Arabidopsis.Comparison of salt regulation in wild type and sos3identified six genes that are controlled by the SOSsignal pathway. Transcription of one gene (VSP2) iscontrolled as an output from the SOS3/SOS2 path-way, similar to SOS1 (Shi et al., 2000), whereas reg-ulation of five other genes (AD06C08, CCR1, SAMT,COR6.6/KIN2, and STZ) is dependent on both SOS3and SOS2 as well. Because SAMT, COR6.6/KIN2, andSTZ are induced by NaCl, negative control by SOS3indicates that the SOS pathway functions to reducethe numerous signals induced by salt to those thatfunction more specifically, to mediate processes likeion homeostasis.
RESULTS
Identification of Expressed Sequence Tags (ESTs)Corresponding to Salt-Regulated Genes
A combination of approaches was used to identifystress-regulated transcriptional outputs from the SOSpathway on the premise that these are salt tolerance
determinants. A population of ESTs representingsalt-regulated transcripts was identified by screen-ing, through three rounds, a subtracted cDNA libraryprepared from wild-type (Col-0 gl-1) seedlingstreated for 4 h without or with 160 mm NaCl. Differ-ential dot-blot hybridization, and sequence andnorthern-blot analyses identified unique ESTs (salt-regulated ESTs [SREs]) that detected salt-regulatedtranscripts (Table I). The second and third rounds ofscreening eliminated highly repetitive clones fromthe initial subtracted library, including those ESTsidentified during the first round of screening. Thedifferential subtractive chain selection protocol wasused in the third round (Luo et al., 1999) to furtherenrich for less abundant cDNAs in the library. Thedifferent rounds of screening led to identification of84 nonredundant ESTs that hybridize to salt-regulated transcripts based on northern-blot analysis(Table II). The SREs represent the greatest numberthat has been used in one study to define the tran-scriptional response of plants to salt stress. Theseresults identify genes whose expression is most likelycontrolled by transcriptional activation, althoughother factors such as salt stress-dependent mRNAstability might contribute also to steady-state tran-script abundance.
Database comparisons of SREs using Blast pro-grams determined that the corresponding encodedproteins included those involved in primary metab-olism, cell wall synthesis or degradation, other cellu-lar functions, transport or nutrient assimilation, sig-naling, and defensive responses (Table II). The SREswere compared with Arabidopsis genome data usingBlastn. Blastp analysis was performed on any SREORF, without predicted function, that was identifiedin an Arabidopsis database. Blastx/Blastp analysiswas performed on SRE sequences that were unanno-tated as an ORF.
Several of the salt-responsive genes identified inthis evaluation encode components of octadecanoidsignaling through jasmonic acid (Table II, lipid sig-naling responses). The plant hormone appears to bederived from hydrolysis of membrane phospholipids(Koiwa et al., 1997). Triacyl glycerol lipase (AD03B08)can release free linolenic acid from phospholipids thatis then oxidized by lipoxygenase (AD04G12) and cy-clized by allene oxide synthase and allene oxide cy-clase (AD04D07) to 12-oxo-phytodienoic acid. Two
Table I. Arabidopsis (Col-1 gl1) SREs identified after successive rounds of differential subtractionSalt regulation was based on detection of altered steady-state transcript abundance after NaCl
treatment.
Round Dot Blot Sequenced Unique Clones Salt Regulated
First 1,000 137 34 25Second 6,000 157 91 42Third 3,000 320 80 17
Total 10,000 614 205 84
Gong et al.
364 Plant Physiol. Vol. 126, 2001

Tabl
eII
.A
rabi
dops
is(C
ol-0
and
gl1)
ESTs
corr
espo
ndin
gto
salt-
resp
onsi
vege
nes
dete
rmin
edby
nort
hern
-blo
tan
alys
isTo
tal
RN
Aw
asis
olat
edfr
omse
edlin
gstr
ansf
erre
dto
med
ium
with
out
orw
ith16
0m
MN
aCl
for
4h
SREs
wer
esu
bjec
ted
toB
last
nan
alys
isan
dill
ustr
ated
are
anno
tatio
nsba
sed
onth
ein
form
atio
nfr
omth
eA
rabi
dops
isge
nom
ese
quen
cepr
ojec
t:ch
rom
osom
eno
.,se
ctio
non
chro
mos
omes
2an
d4,
orB
AC
clon
eid
entif
icat
ion
no.(
for
chro
mos
omes
1,3,
and
5),a
ndge
ne/p
rote
inac
cess
ion
nos.
Hig
hlig
hted
inbo
ldar
eSR
Esth
atde
tect
edtr
ansc
ript
sdi
ffere
ntia
llyre
gula
ted
ingl
1an
dso
s3.
Tran
scri
ptab
unda
nce
isra
ted
base
don
the
scal
eill
ustr
ated
.
Gen
Ban
kA
cces
sion
No.
EST
No.
Chr
omo-
som
eB
AC
Clo
neN
o.ge
neID
prot
einI
DA
nnot
atio
non
Ara
bido
psis
Gen
e/H
omol
oga
Scor
eE
Val
ueC
omm
ents
gl-1
Con
trol
gl-1
Salt
bits
Prim
ary
met
abol
ism
Phot
osyn
thes
isB
E844
959
AD
04H
113
F5N
5A
TU05
218
AA
A21
570.
1A
TPsu
lfury
lase
492
e-13
81
11
11
11
11
BE8
4517
9A
D08
A06
225
At2
g051
00A
AD
3135
8.1
Puta
tive
chlo
roph
yll
a/b-
bind
ing
prot
ein
789
0.0
–1
1
BE8
4535
1A
D10
C12
469
AT4
g287
50C
AB
8146
3.1
Phot
osys
tem
Isu
buni
tPS
I-E-
like
prot
ein
513
e-14
41
11
11
11
11
BE8
4535
6A
D10
D06
1F1
2A21
F12A
21.1
1A
AF2
6934
.1Pu
tativ
ePS
BY
embl
CA
A11
248
728
0.0
11
11
1C
arbo
hydr
ate
BE8
4484
9A
D03
C01
47
dl45
75c
CA
B46
051.
1Pu
tativ
eb
-am
ylas
e10
780.
0G
ene
dupl
icat
ion?
–1
BE8
4499
8A
D05
E02
345
T22E
16A
T4g1
7090
T22E
16.7
0C
AB
8098
0.1
CA
B75
899.
12-
Oxo
glut
arat
ede
hydr
ogen
ase,
E1su
buni
t-lik
epr
otei
n24
22e
-63
11
1
Am
ino
acid
BE8
4483
4A
D03
A02
459
AT4
g236
00C
AB
7931
5.1
Tyro
sine
tran
sam
inas
e-lik
epr
otei
n60
5e-
172
11
11
11
.B
E844
976
AD
05B
091
F15I
1F1
5I1.
19A
AD
2578
3.1
Stro
ngsi
mila
rity
togb
lS77
096
alde
-hy
dede
hydr
ogen
ase
hom
olog
from
Bra
ssic
ana
pus
and
isa
mem
ber
ofPF
I001
71al
dehy
dede
hydr
ogen
ase
fam
ily
137
5e-3
1Pr
olin
ede
grad
atio
n?1
11
1
BE8
4506
3A
D06
C01
3T2
1L8
T21L
8.90
CA
B51
206.
1G
luta
min
e-de
pend
ent
aspa
ragi
nesy
nthe
tase
210
4e-5
31
11
1
BE8
4511
1A
D06
G04
482
AT4
g347
10C
AB
8018
8.1
Arg
inin
ede
carb
oxyl
ase
SPE2
809
0.0
Poly
amin
esy
nthe
sis
–1
11
1
Oth
ers
BE8
4498
9A
D05
D03
5K
24M
7–
–N
otan
nota
ted
55.4
6e-0
8C
ytoc
hrom
ep4
50–
11
pirl
lA29
368
Pros
tagl
andi
nom
ega-
hydr
oxyl
ase
(EC
1.14
.15.
-)cy
to-
chro
me
P450
4A4,
rabb
ita
48e-
05
Cel
lula
rfu
nctio
nC
ell
wal
lB
E844
904
AD
04B
125
MM
I9A
B01
9235
.1B
AA
9719
9.1
Rip
enin
gre
late
d,pr
otei
nlik
e;co
ntai
nssi
mila
rity
tope
ctin
este
rase
712
0.0
11
11
BE8
4508
7A
D06
E01
5F7
K24
––
Not
anno
tate
d33
93e
-92
11
1pi
rllT
1161
0Pr
obab
leci
nnam
yl-
alco
hol
dehy
drog
enas
e(E
C1.
1.1.
195)
CPR
D14
(Vig
naun
guic
ulat
a)a
191
2e-4
8
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 365
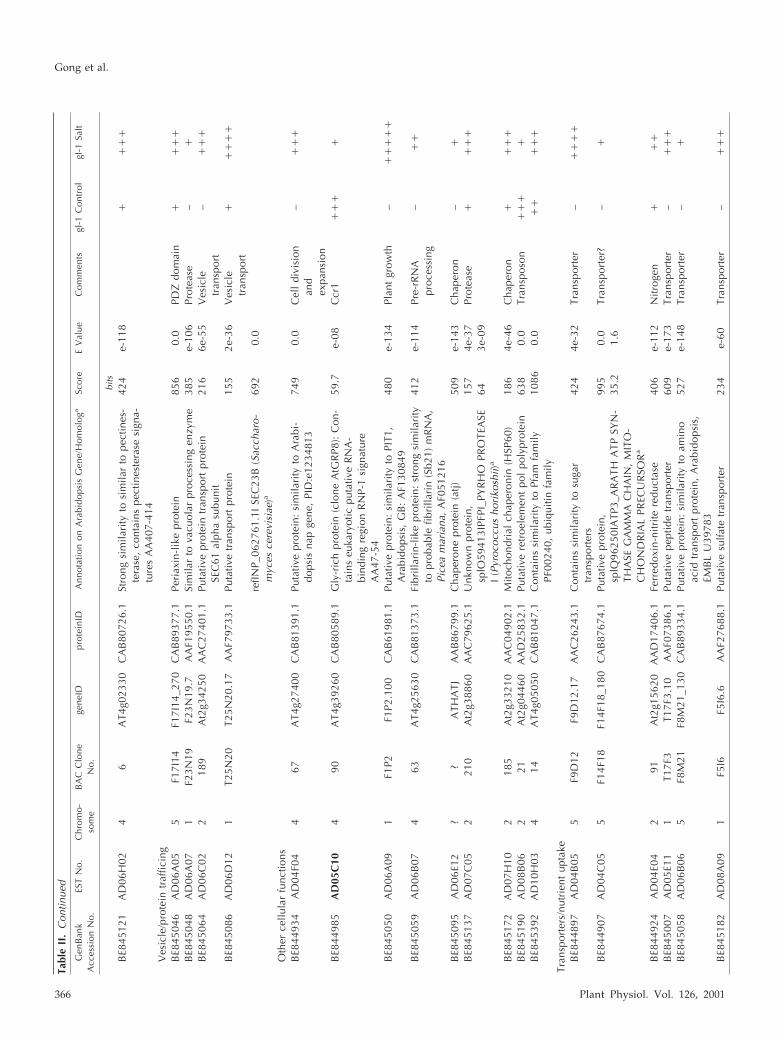
Tabl
eII
.C
ontin
ued
Gen
Ban
kA
cces
sion
No.
EST
No.
Chr
omo-
som
eB
AC
Clo
neN
o.ge
neID
prot
einI
DA
nnot
atio
non
Ara
bido
psis
Gen
e/H
omol
oga
Scor
eE
Val
ueC
omm
ents
gl-1
Con
trol
gl-1
Salt
bits
BE8
4512
1A
D06
H02
46
AT4
g023
30C
AB
8072
6.1
Stro
ngsi
mila
rity
tosi
mila
rto
pect
ines
-te
rase
,co
ntai
nspe
ctin
este
rase
sign
a-tu
res
AA
407-
414
424
e-11
81
11
1
Ves
icle
/pro
tein
traf
ficin
gB
E845
046
AD
06A
055
F17I
14F1
7I14
_270
CA
B89
377.
1Pe
riax
in-l
ike
prot
ein
856
0.0
PDZ
dom
ain
11
11
BE8
4504
8A
D06
A07
1F2
3N19
F23N
19.7
AA
F195
50.1
Sim
ilar
tova
cuol
arpr
oces
sing
enzy
me
385
e-10
6Pr
otea
se–
1B
E845
064
AD
06C
022
189
At2
g342
50A
AC
2740
1.1
Puta
tive
prot
ein
tran
spor
tpr
otei
nSE
C61
alph
asu
buni
t21
66e
-55
Ves
icle
tran
spor
t–
11
1
BE8
4508
6A
D06
D12
1T2
5N20
T25N
20.1
7A
AF7
9733
.1Pu
tativ
etr
ansp
ort
prot
ein
155
2e-3
6V
esic
letr
ansp
ort
11
11
1
refIN
P_06
2761
.1I
SEC
23B
(Sac
char
o-m
yces
cere
visi
ae)a
692
0.0
Oth
erce
llula
rfu
nctio
nsB
E844
934
AD
04F0
44
67A
T4g2
7400
CA
B81
391.
1Pu
tativ
epr
otei
n:si
mila
rity
toA
rabi
-do
psis
nap
gene
,PI
D:e
1234
813
749
0.0
Cel
ldi
visi
onan
dex
pans
ion
–1
11
BE8
4498
5A
D05
C10
490
AT4
g392
60C
AB
8058
9.1
Gly
-ric
hpr
otei
n(c
lone
AtG
RP8
):C
on-
tain
seu
kary
otic
puta
tive
RN
A-
bind
ing
regi
onR
NP-
1si
gnat
ure
AA
47-5
4
59.7
e-08
Ccr
11
11
1
BE8
4505
0A
D06
A09
1F1
P2F1
P2.1
00C
AB
6198
1.1
Puta
tive
prot
ein:
sim
ilari
tyto
PIT1
,A
rabi
dops
is,
GB
:A
F130
849
480
e-13
4Pl
ant
grow
th–
11
11
1
BE8
4505
9A
D06
B07
463
AT4
g256
30C
AB
8137
3.1
Fibr
illar
in-l
ike
prot
ein:
stro
ngsi
mila
rity
topr
obab
lefib
rilla
rin
(Sb2
1)m
RN
A,
Pice
am
aria
na,
AF0
5121
6
412
e-11
4Pr
e-rR
NA
proc
essi
ng–
11
BE8
4509
5A
D06
E12
??
ATH
ATJ
AA
B86
799.
1C
hape
rone
prot
ein
(atj)
509
e-14
3C
hape
ron
–1
BE8
4513
7A
D07
C05
221
0A
t2g3
8860
AA
C79
625.
1U
nkno
wn
prot
ein,
splO
5941
3IPF
PI_P
YR
HO
PRO
TEA
SEI
(Pyr
ococ
cus
hori
kosh
ii)a
157
644e
-37
3e-0
9Pr
otea
se1
11
1
BE8
4517
2A
D07
H10
218
5A
t2g3
3210
AA
C04
902.
1M
itoch
ondr
ial
chap
eron
in(H
SP60
)18
64e
-46
Cha
pero
n1
11
1B
E845
190
AD
08B
062
21A
t2g0
4460
AA
D25
832.
1Pu
tativ
ere
troe
lem
ent
pol
poly
prot
ein
638
0.0
Tran
spos
on1
11
1B
E845
392
AD
10H
034
14A
T4g0
5050
CA
B81
047.
1C
onta
ins
sim
ilari
tyto
Pfam
fam
ilyPF
0024
0,ub
iqui
tinfa
mily
1086
0.0
11
11
1
Tran
spor
ters
/nut
rien
tup
take
BE8
4489
7A
D04
B05
5F9
D12
F9D
12.1
7A
AC
2624
3.1
Con
tain
ssi
mila
rity
tosu
gar
tran
spor
ters
424
4e-3
2Tr
ansp
orte
r–
11
11
BE8
4490
7A
D04
C05
5F1
4F18
F14F
18_1
80C
AB
8767
4.1
Puta
tive
prot
ein,
splQ
9625
0IA
TP3_
AR
ATH
ATP
SYN
-TH
ASE
GA
MM
AC
HA
IN,
MIT
O-
CH
ON
DR
IAL
PREC
UR
SOR
a
995
35.2
0.0
1.6
Tran
spor
ter?
–1
BE8
4492
4A
D04
E04
291
At2
g156
20A
AD
1740
6.1
Ferr
edox
in-n
itrite
redu
ctas
e40
6e-
112
Nitr
ogen
11
1B
E845
007
AD
05E1
11
T17F
3T1
7F3.
10A
AF0
7386
.1Pu
tativ
epe
ptid
etr
ansp
orte
r60
9e-
173
Tran
spor
ter
–1
11
BE8
4505
8A
D06
B06
5F8
M21
F8M
21_1
30C
AB
8933
4.1
Puta
tive
prot
ein:
sim
ilari
tyto
amin
oac
idtr
ansp
ort
prot
ein,
Ara
bido
psis
,EM
BL
U39
783
527
e-14
8Tr
ansp
orte
r–
1
BE8
4518
2A
D08
A09
1F5
I6F5
I6.6
AA
F276
88.1
Puta
tive
sulfa
tetr
ansp
orte
r23
4e-
60Tr
ansp
orte
r–
11
1
Gong et al.
366 Plant Physiol. Vol. 126, 2001
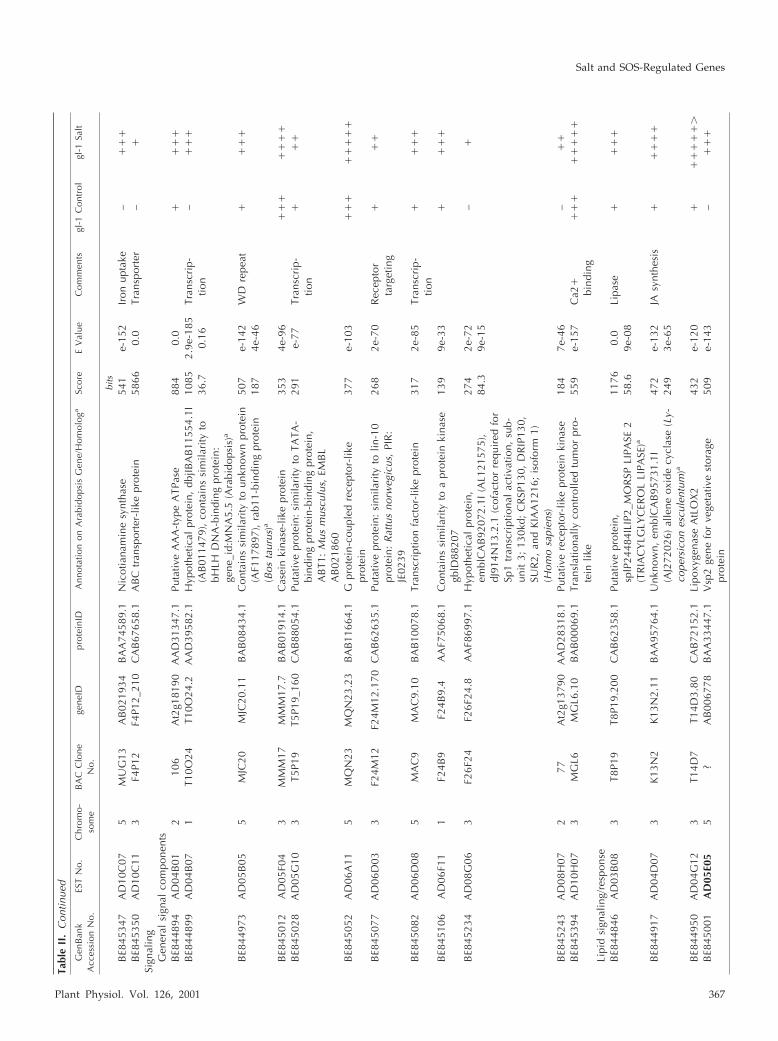
Tabl
eII
.C
ontin
ued
Gen
Ban
kA
cces
sion
No.
EST
No.
Chr
omo-
som
eB
AC
Clo
neN
o.ge
neID
prot
einI
DA
nnot
atio
non
Ara
bido
psis
Gen
e/H
omol
oga
Scor
eE
Val
ueC
omm
ents
gl-1
Con
trol
gl-1
Salt
bits
BE8
4534
7A
D10
C07
5M
UG
13A
B02
1934
BA
A74
589.
1N
icot
iana
min
esy
ntha
se54
1e-
152
Iron
upta
ke–
11
1B
E845
350
AD
10C
113
F4P1
2F4
P12_
210
CA
B67
658.
1A
BC
tran
spor
ter-
like
prot
ein
5866
0.0
Tran
spor
ter
–1
Sign
alin
gG
ener
alsi
gnal
com
pone
nts
BE8
4489
4A
D04
B01
210
6A
t2g1
8190
AA
D31
347.
1Pu
tativ
eA
AA
-typ
eA
TPas
e88
40.
01
11
1B
E844
899
AD
04B
071
T10O
24T1
0O24
.2A
AD
3958
2.1
Hyp
othe
tical
prot
ein,
dbjIB
AB
1155
4.1I
(AB
0114
79),
cont
ains
sim
ilari
tyto
bHLH
DN
A-b
indi
ngpr
otei
n:ge
ne_i
d:M
NA
5.5
(Ara
bido
psis
)a
1085
36.7
2.9e
-185
0.16
Tran
scri
p-tio
n–
11
1
BE8
4497
3A
D05
B05
5M
JC20
MJC
20.1
1B
AB
0843
4.1
Con
tain
ssi
mila
rity
toun
know
npr
otei
n(A
F117
897)
,ra
b11-
bind
ing
prot
ein
(Bos
taur
us)a
507
187
e-14
24e
-46
WD
repe
at1
11
1
BE8
4501
2A
D05
F04
3M
MM
17M
MM
17.7
BA
B01
914.
1C
asei
nki
nase
-lik
epr
otei
n35
34e
-96
11
11
11
1B
E845
028
AD
05G
103
T5P1
9T5
P19_
160
CA
B88
054.
1Pu
tativ
epr
otei
n:si
mila
rity
toTA
TA-
bind
ing
prot
ein-
bind
ing
prot
ein,
AB
T1:
Mus
mus
culu
s,EM
BL
AB
0218
60
291
e-77
Tran
scri
p-tio
n1
11
BE8
4505
2A
D06
A11
5M
QN
23M
QN
23.2
3B
AB
1166
4.1
Gpr
otei
n-co
uple
dre
cept
or-l
ike
prot
ein
377
e-10
31
11
11
11
1
BE8
4507
7A
D06
D03
3F2
4M12
F24M
12.1
70C
AB
6263
5.1
Puta
tive
prot
ein:
sim
ilari
tyto
lin-1
0pr
otei
n:R
attu
sno
rweg
icus
,PI
R:
JE02
39
268
2e-7
0R
ecep
tor
targ
etin
g1
11
BE8
4508
2A
D06
D08
5M
AC
9M
AC
9.10
BA
B10
078.
1Tr
ansc
ript
ion
fact
or-l
ike
prot
ein
317
2e-8
5Tr
ansc
rip-
tion
11
11
BE8
4510
6A
D06
F11
1F2
4B9
F24B
9.4
AA
F750
68.1
Con
tain
ssi
mila
rity
toa
prot
ein
kina
segb
lD88
207
139
9e-3
31
11
1
BE8
4523
4A
D08
G06
3F2
6F24
F26F
24.8
AA
F869
97.1
Hyp
othe
tical
prot
ein,
embl
CA
B92
072.
1I(A
L121
575)
,dJ
914N
13.2
.1(c
ofac
tor
requ
ired
for
Sp1
tran
scri
ptio
nal
activ
atio
n,su
b-un
it3;
130k
d;C
RSP
130,
DR
IP13
0,SU
R2,
and
KIA
A12
16;
isof
orm
1)(H
omo
sapi
ens)
274
84.3
2e-7
29e
-15
–1
BE8
4524
3A
D08
H07
277
At2
g137
90A
AD
2831
8.1
Puta
tive
rece
ptor
-lik
epr
otei
nki
nase
184
7e-4
6–
11
BE8
4539
4A
D10
H07
3M
GL6
MG
L6.1
0B
AB
0006
9.1
Tran
slat
iona
llyco
ntro
lled
tum
orpr
o-te
inlik
e55
9e-
157
Ca2
1bi
ndin
g1
11
11
11
1
Lipi
dsi
gnal
ing/
resp
onse
BE8
4484
6A
D03
B08
3T8
P19
T8P1
9.20
0C
AB
6235
8.1
Puta
tive
prot
ein,
splP
2448
4ILI
P2_M
OR
SPLI
PASE
2(T
RIA
CY
LGLY
CER
OL
LIPA
SE)a
1176
58.6
0.0
9e-0
8Li
pase
11
11
BE8
4491
7A
D04
D07
3K
13N
2K
13N
2.11
BA
A95
764.
1U
nkno
wn,
embl
CA
B95
731.
1I(A
J272
026)
alle
neox
ide
cycl
ase
(Ly-
cope
rsic
ones
cule
ntum
)a
472
249
e-13
23e
-65
JAsy
nthe
sis
11
11
1
BE8
4495
0A
D04
G12
3T1
4D7
T14D
3.80
CA
B72
152.
1Li
poxy
gena
seA
tLO
X2
432
e-12
01
11
11
1.
BE8
4500
1A
D05
E05
5?
AB
0067
78B
AA
3344
7.1
Vsp
2ge
nefo
rve
geta
tive
stor
age
prot
ein
509
e-14
3–
11
1
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 367

Tabl
eII
.C
ontin
ued
Gen
Ban
kA
cces
sion
No.
EST
No.
Chr
omo-
som
eB
AC
Clo
neN
o.ge
neID
prot
einI
DA
nnot
atio
non
Ara
bido
psis
Gen
e/H
omol
oga
Scor
eE
Val
ueC
omm
ents
gl-1
Con
trol
gl-1
Salt
bits
BE8
4509
1A
D06
E08
5M
TI20
MTI
20.3
BA
B08
850.
1Li
pid
tran
sfer
prot
ein;
glos
sy1
hom
olog
414
e-11
4–
11
1B
E845
100
AD
06F0
53
T02O
04T0
2O04
.11
AA
B63
638.
1Ja
smon
ate-
indu
cibl
epr
otei
nis
olog
,m
yros
inas
ebi
ndin
gpr
otei
nlik
e46
6e-
130
11
11
11
Hor
mon
ere
late
dB
E844
978
AD
05B
113
F28D
10F2
8D10
_50
CA
C03
536.
1A
tPP-
like
prot
ein:
prok
aryo
ticm
em-
bran
elip
opro
tein
lipid
atta
chm
ent
site
AA
199-
209
(AF1
3305
3),
S-ad
-en
osyl
-L-m
ethi
onin
e:sa
licyl
icac
idca
rbox
ylm
ethy
ltran
sfer
ase
(Cla
rkia
brew
eri)a
706
129
0.0
6e-2
9SA
–1
1
BE8
4540
1A
D05
C05
??
AF1
8382
7A
AF2
2295
.1B
eta-
gluc
osid
ase
hom
olog
(BG
1)35
150.
0–
11
11
BE8
4508
4A
D06
D10
3T1
0D17
T10D
17_9
0C
AB
8899
8.1
Nitr
ilase
220
2e-
501
11
1C
ell
deat
hB
E844
851
AD
03C
065
MQ
L5M
QL5
.19
BA
A97
167.
1Pa
lmito
yl-p
rote
inth
ioes
tera
sepr
ecur
-so
rlik
e42
2e-
117
Ant
i-ap
opto
sis?
11
1
BE8
4495
8A
D04
H10
3F2
8D10
F28D
10_7
0C
AC
0353
8.1
Leth
alle
af-s
pot
1ho
mol
ogLl
s1:
con-
tain
spr
okar
yotic
mem
bran
elip
opro
-te
inlip
idat
tach
men
tsi
teA
A15
0-16
0
472
e-13
2D
ioxy
gena
se1
11
Def
ense
resp
onse
Def
ense
prot
eins
BE8
4487
5A
D03
G09
1F2
2K20
F22K
20.1
9A
AC
0062
5.1
Alc
ohol
dehy
drog
enas
e48
6e-
136
11
11
BE8
4489
6A
D04
B03
5M
SG15
MSG
15.3
BA
B11
043.
1M
ande
loni
trile
lyas
e-lik
epr
otei
n51
9e-
146
Cya
noge
ne-
sis
–1
1
BE8
4492
6A
D04
E06
3M
DB
19M
DB
19.5
BA
B02
775.
1C
onta
ins
sim
ilari
tyto
endo
-1,3
-1,4
-be
ta- D
-glu
cana
sege
ne_i
d:M
DB
19.5
414
e-11
51
11
1
BE8
4497
2A
D05
B03
1F5
F19
F5F1
9.6
AA
D12
691.
1Si
mila
rto
gblY
0943
7m
yros
inas
ebi
nd-
ing
prot
ein
from
B.
napu
s19
48e
-49
–1
BE8
4499
0A
D05
D04
1F2
4B9
F24B
9.34
AA
F750
98.1
Met
allo
thio
nein
605
e-17
21
11
11
11
11
BE8
4540
3A
D06
E03
1F1
5K9
F15K
9.17
AA
C72
119.
1St
rong
sim
ilari
tyto
gblD
1455
0ex
tra-
cellu
lar
derm
algl
ycop
rote
in(E
DG
P)pr
ecur
sor
from
Dau
cus
caro
ta
745
0.0
11
1
BE8
4511
7A
D06
G10
1F1
4N23
F14N
23.2
5A
AD
3288
7.1
Sim
ilar
togl
utat
hion
eS-
tran
sfer
ase
TSI-
1(g
il219
0992
)76
30.
0C
yano
gene
-si
s1
11
11
1
BE8
4515
2A
D07
F07
223
6A
t2g4
3620
AA
B64
044.
1Pu
tativ
een
doch
itina
se54
9e-
154
11
11
1B
E845
153
AD
07F0
85
MO
J9M
OJ9
.4B
AB
1114
5.1
Poly
gala
ctur
onas
e-in
hibi
ting
prot
ein
509
e-14
2–
11
11
Cor
/RD
prot
eins
BE8
4485
0A
D03
C02
473
AT4
g306
50C
AB
7978
3.1
Stro
ngsi
mila
rity
tolo
wte
mpe
ratu
rean
dsa
lt-re
spon
sive
prot
ein
LTI6
A,
Ara
bido
psis
383
e-10
51
11
1
BE8
4485
4A
D03
D05
5F1
N13
F1N
13_1
10C
AC
0179
6.1
Col
d-re
gula
ted
prot
ein
CO
R6.
6(K
IN2)
341
2e-9
2–
11
11
1.
BE8
4485
5A
D03
D11
2F1
4N22
At2
g425
40A
AD
2299
9.1
Col
d-re
gula
ted
prot
ein
cor1
5apr
ecur
sor
519
e-14
–1
11
11
.
BE8
4486
5A
D03
F02
5K
24M
7D
1304
4B
AA
0237
6.1
Des
icca
tion-
resp
onsi
verd
29A
371
e-10
1–
11
11
1.
BE8
4502
2A
D05
G04
5T1
4C9
ATH
RD
22B
AA
0154
6.1
rd22
Gen
e44
8e-
124
–1
11
11
BE8
4504
9A
D06
A08
1F1
5M15
F5M
15.2
1A
AF7
9613
.1Si
mila
rto
cold
-reg
ulat
edpr
otei
nco
r47
127
4e-2
8?
?
Gong et al.
368 Plant Physiol. Vol. 126, 2001
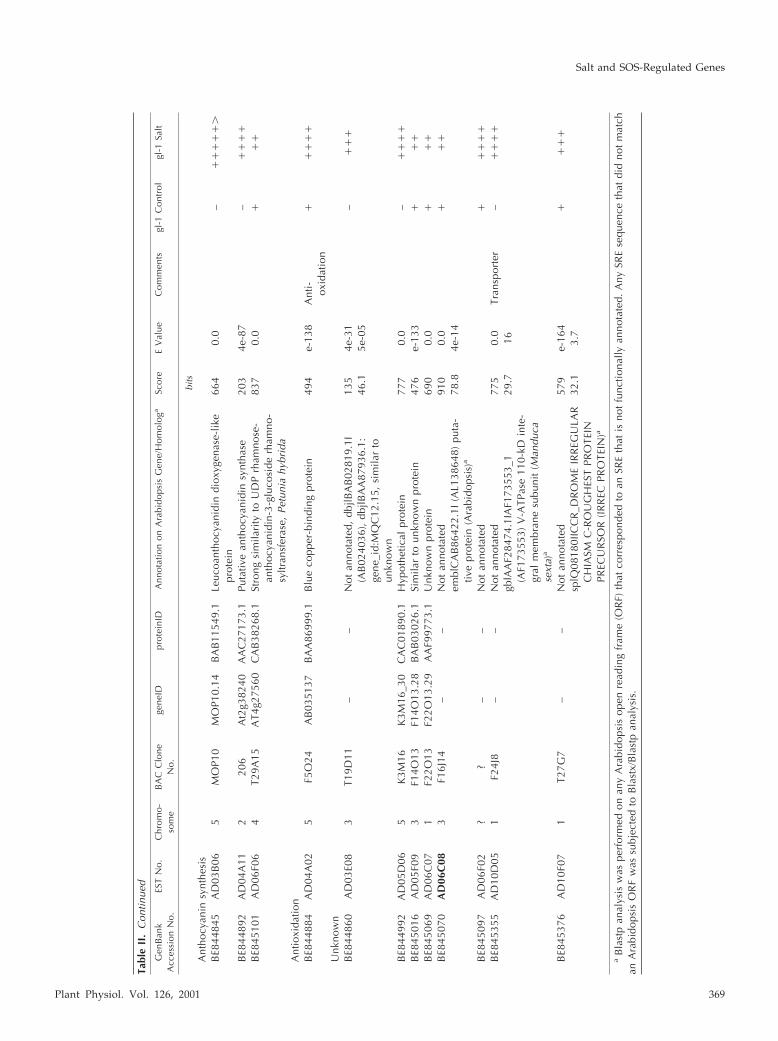
Tabl
eII
.C
ontin
ued
Gen
Ban
kA
cces
sion
No.
EST
No.
Chr
omo-
som
eB
AC
Clo
neN
o.ge
neID
prot
einI
DA
nnot
atio
non
Ara
bido
psis
Gen
e/H
omol
oga
Scor
eE
Val
ueC
omm
ents
gl-1
Con
trol
gl-1
Salt
bits
Ant
hocy
anin
synt
hesi
sB
E844
845
AD
03B
065
MO
P10
MO
P10.
14B
AB
1154
9.1
Leuc
oant
hocy
anid
indi
oxyg
enas
e-lik
epr
otei
n66
40.
0–
11
11
1.
BE8
4489
2A
D04
A11
220
6A
t2g3
8240
AA
C27
173.
1Pu
tativ
ean
thoc
yani
din
synt
hase
203
4e-8
7–
11
11
BE8
4510
1A
D06
F06
4T2
9A15
AT4
g275
60C
AB
3826
8.1
Stro
ngsi
mila
rity
toU
DP
rham
nose
-an
thoc
yani
din-
3-gl
ucos
ide
rham
no-
syltr
ansf
eras
e,Pe
tuni
ahy
brid
a
837
0.0
11
1
Ant
ioxi
datio
nB
E844
884
AD
04A
025
F5O
24A
B03
5137
BA
A86
999.
1B
lue
copp
er-b
indi
ngpr
otei
n49
4e-
138
Ant
i-ox
idat
ion
11
11
1
Unk
now
nB
E844
860
AD
03E0
83
T19D
11–
–N
otan
nota
ted,
dbjlB
AB
0281
9.1I
(AB
0240
36),
dbjlB
AA
8793
6.1:
gene
_id:
MQ
C12
.15,
sim
ilar
toun
know
n
135
46.1
4e-3
15e
-05
–1
11
BE8
4499
2A
D05
D06
5K
3M16
K3M
16_3
0C
AC
0189
0.1
Hyp
othe
tical
prot
ein
777
0.0
–1
11
1B
E845
016
AD
05F0
93
F14O
13F1
4O13
.28
BA
B03
026.
1Si
mila
rto
unkn
own
prot
ein
476
e-13
31
11
BE8
4506
9A
D06
C07
1F2
2O13
F22O
13.2
9A
AF9
9773
.1U
nkno
wn
prot
ein
690
0.0
11
1B
E845
070
AD
06C
083
F16J
14–
–N
otan
nota
ted
910
0.0
11
1em
blC
AB
8642
2.1I
(AL1
3864
8)pu
ta-
tive
prot
ein
(Ara
bido
psis
)a78
.84e
-14
BE8
4509
7A
D06
F02
??
––
Not
anno
tate
d1
11
11
BE8
4535
5A
D10
D05
1F2
4J8
––
Not
anno
tate
d77
50.
0Tr
ansp
orte
r–
11
11
gblA
AF2
8474
.1IA
F173
553_
1(A
F173
553)
V-A
TPas
e11
0-kD
inte
-gr
alm
embr
ane
subu
nit
(Man
duca
sext
a)a
29.7
16
BE8
4537
6A
D10
F07
1T2
7G7
––
Not
anno
tate
d57
9e-
164
11
11
splQ
0818
0IIC
CR
_DR
OM
EIR
REG
ULA
RC
HIA
SMC
-RO
UG
HES
TPR
OTE
INPR
ECU
RSO
R(IR
REC
PRO
TEIN
)a
32.1
3.7
aB
last
pan
alys
isw
aspe
rfor
med
onan
yA
rabi
dops
isop
enre
adin
gfr
ame
(OR
F)th
atco
rres
pond
edto
anSR
Eth
atis
not
func
tiona
llyan
nota
ted.
Any
SRE
sequ
ence
that
did
not
mat
chan
Ara
bido
psis
OR
Fw
assu
bjec
ted
toB
last
x/B
last
pan
alys
is.
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 369

SREs were annotated previously as jasmonic acid reg-ulated, VSP2 (AD05E05) and AD06F05. Furthermore,the genes of a number of SREs are involved in plantdefense and may be regulated by the octadecanoidsignal pathway. Some of these genes have been shownto express upon dehydration in tomato (Lycopersiconesculentum; Reymond et al., 2000). Several abscisic acid(ABA)-responsive SREs are included in Table II andthe plant hormone is a potentiator of octadecanoidsignaling.
Genes Differentially Regulated by Salt in WildType and sos3
The salt regulated expression profile of SRE tran-scripts, as well as that of previously characterizedstress-regulated genes (ACP1, RD22BP1, MYB2, STZ,and PAL), revealed that most are controlled similarlyin wild type and sos3. Six of the salt-responsivegenes, including STZ (Lippuner et al., 1996), weredifferentially regulated in wild type and sos3 (Fig. 1).Transcript abundance of two was lower (AD06C08/unknown and AD05E05/vegetative storage protein2[VSP2]) and of four was higher (encoding AD05C10/cold-circadian rhythm-RNA binding1 [CCR1], STZ/salt tolerance zinc finger [not shown], AD05B11/S-adenosyl-l-Met: salicyclic acid carboxyl methyl-transferase [SAMT], AD03D05/cold regulated/coldinducible [COR6.6/KIN2]) in the salt-sensitive mutant.SAMT and COR6.6/KIN2 transcript abundance wasslightly elevated in sos3 but the steady-state mRNAlevels were hyper-induced by salt treatment. CCR1was the only gene for which transcript abundance isreduced by salt treatment.
Salt regulation of VSP2 is similar in wild type andsos1 indicating that transcriptional activation is not
dependent on SOS1 (Fig. 2). Methyl jasmonate(MeJA) induces VSP2 transcript abundance in wildtype and sos3 (not shown). The SOS3/2 pathway andthe hormone seem to regulate VSP2 independently.Signal pathways often converge to regulate transcrip-tion of key effectors involved in cellular adaptation toenvironmental perturbation (Rep et al., 2000). VSP2 isa member of a two-gene family (87% nucleotide se-quence identity over the coding region) that encodesa protein with similarity to soybean VSPs (Berger etal., 1995; Utsugi et al., 1998), which are vacuolar-localized glycoproteins with acid phosphatase activ-ity (Mason and Mullet, 1990). These proteins arepresumed to be amino acid sinks during water deficitbut are important reduced nitrogen sources afterstress relief (Mason and Mullet, 1990).
CCR1, STZ, SAMT, COR6.6/KIN2, and AD06C08/unknown have similar expression profiles in sos1,sos2, and sos3, implicating these as transcriptionaloutputs requiring all components of the SOS path-way. CCR1 encodes a Gly-rich RNA-binding proteinimplicated in posttranscriptional regulation. CCR1and CCR2 comprise a two-gene family, and theirexpression is regulated by a diurnal circadian clock(Carpenter et al., 1994; Heintzen et al., 1997; Krepsand Simon, 1997). CCR1 or CCR2 expression nega-tively regulates either gene and this feedback looppresumably facilitates diurnal oscillation controlledby the master circadian clock (Heintzen et al., 1997).CCR1 and CCR2 steady-state mRNA levels are in-duced by cold but CCR1 transcript is negatively reg-ulated by ABA and dehydration, whereas CCR2 ex-pression is induced by dehydration (Carpenter et al.,1994). CCR1 transcript abundance was down-regulated in sos3 indicating that the SOS pathway isat least another negative regulator that controls CCR1expression downstream of the circadian rhythmclock (Heintzen et al., 1997; Kreps and Simon, 1997).These results indicate that control of circadian oscil-lations may be required during the salt stressresponse.
COR6.6//KIN2 and SAMT are both implicated inplant stress responses. COR6.6/KIN2 is linked in tan-dem to its homolog KIN1 (95% nucleotide sequenceidentity in the coding region) and both encode hy-drophilic peptides that are boiling soluble but ofunknown function (Thomashow, 1999). KIN1 and 2transcript abundance is cold and ABA induced (Kur-kela and Franck, 1990; Kurkela and Borg-Franck,1992; Thomashow, 1999) but KIN2 and not KIN1 ex-pression is modulated positively by drought and salt(Kurkela and Borg-Franck, 1992). SAMT catalyzes theformation of methylsalicylate from salicylic acid us-ing S-adenosyl-l-Met as the methyl donor (Ross etal., 1999). The volatile ester is implicated as a polli-nator attractant and a signal in plant defense medi-ated by salicylic acid (Ross et al., 1999; Dudareva etal., 2000).
Figure 1. Salt-responsive gene expression that is dependent on theSOS pathway in Arabidopsis. Genes that are differentially regulatedin wild-type (Col-0 gl1) and sos3. Total RNA (40 mg) from seedlingsgrown in liquid culture (14 d) and treated without or with 160 mM
NaCl for 4 h. The northern blot was hybridized with 32P-labeledprobe corresponding to: COR6.6/KIN2, SAMT, CCR1, VSP2, andAD06C08 (unknown). AtGAPDH (glyceraldehyde-3-phosphate-dehydrogenase) is the control.
Gong et al.
370 Plant Physiol. Vol. 126, 2001
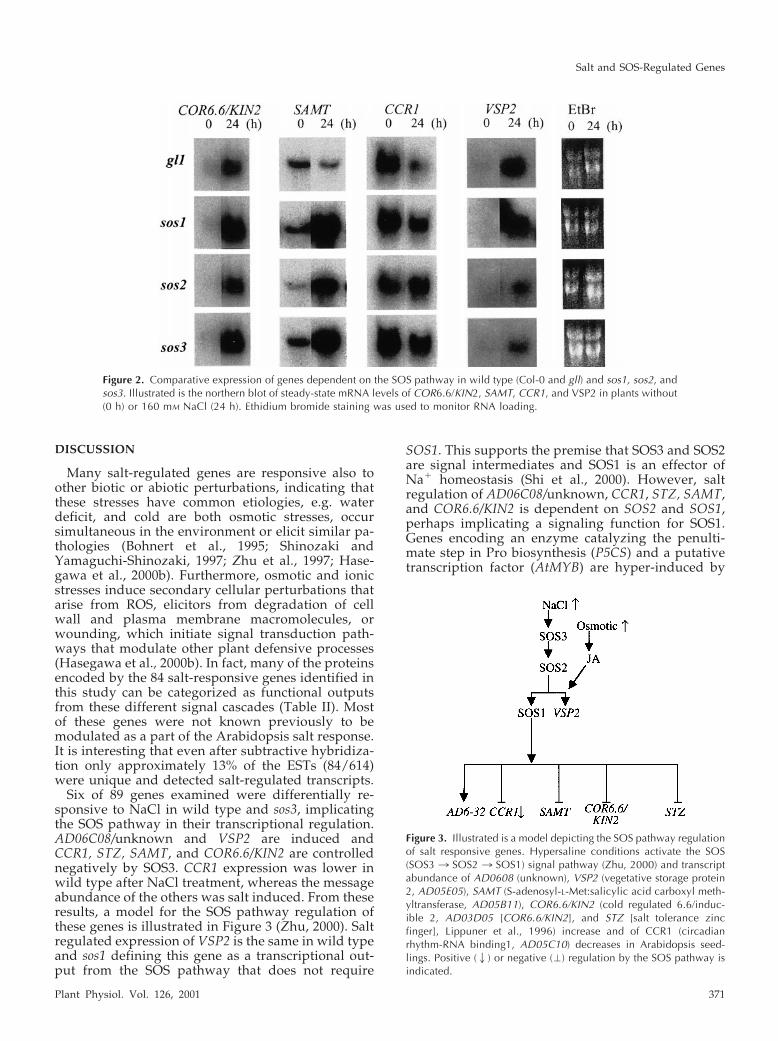
DISCUSSION
Many salt-regulated genes are responsive also toother biotic or abiotic perturbations, indicating thatthese stresses have common etiologies, e.g. waterdeficit, and cold are both osmotic stresses, occursimultaneous in the environment or elicit similar pa-thologies (Bohnert et al., 1995; Shinozaki andYamaguchi-Shinozaki, 1997; Zhu et al., 1997; Hase-gawa et al., 2000b). Furthermore, osmotic and ionicstresses induce secondary cellular perturbations thatarise from ROS, elicitors from degradation of cellwall and plasma membrane macromolecules, orwounding, which initiate signal transduction path-ways that modulate other plant defensive processes(Hasegawa et al., 2000b). In fact, many of the proteinsencoded by the 84 salt-responsive genes identified inthis study can be categorized as functional outputsfrom these different signal cascades (Table II). Mostof these genes were not known previously to bemodulated as a part of the Arabidopsis salt response.It is interesting that even after subtractive hybridiza-tion only approximately 13% of the ESTs (84/614)were unique and detected salt-regulated transcripts.
Six of 89 genes examined were differentially re-sponsive to NaCl in wild type and sos3, implicatingthe SOS pathway in their transcriptional regulation.AD06C08/unknown and VSP2 are induced andCCR1, STZ, SAMT, and COR6.6/KIN2 are controllednegatively by SOS3. CCR1 expression was lower inwild type after NaCl treatment, whereas the messageabundance of the others was salt induced. From theseresults, a model for the SOS pathway regulation ofthese genes is illustrated in Figure 3 (Zhu, 2000). Saltregulated expression of VSP2 is the same in wild typeand sos1 defining this gene as a transcriptional out-put from the SOS pathway that does not require
SOS1. This supports the premise that SOS3 and SOS2are signal intermediates and SOS1 is an effector ofNa1 homeostasis (Shi et al., 2000). However, saltregulation of AD06C08/unknown, CCR1, STZ, SAMT,and COR6.6/KIN2 is dependent on SOS2 and SOS1,perhaps implicating a signaling function for SOS1.Genes encoding an enzyme catalyzing the penulti-mate step in Pro biosynthesis (P5CS) and a putativetranscription factor (AtMYB) are hyper-induced by
Figure 2. Comparative expression of genes dependent on the SOS pathway in wild type (Col-0 and gll) and sos1, sos2, andsos3. Illustrated is the northern blot of steady-state mRNA levels of COR6.6/KIN2, SAMT, CCR1, and VSP2 in plants without(0 h) or 160 mM NaCl (24 h). Ethidium bromide staining was used to monitor RNA loading.
Figure 3. Illustrated is a model depicting the SOS pathway regulationof salt responsive genes. Hypersaline conditions activate the SOS(SOS3 ¡ SOS2 ¡ SOS1) signal pathway (Zhu, 2000) and transcriptabundance of AD0608 (unknown), VSP2 (vegetative storage protein2, AD05E05), SAMT (S-adenosyl-L-Met:salicylic acid carboxyl meth-yltransferase, AD05B11), COR6.6/KIN2 (cold regulated 6.6/induc-ible 2, AD03D05 [COR6.6/KIN2], and STZ [salt tolerance zincfinger], Lippuner et al., 1996) increase and of CCR1 (circadianrhythm-RNA binding1, AD05C10) decreases in Arabidopsis seed-lings. Positive (2) or negative (') regulation by the SOS pathway isindicated.
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 371

salt in sos1 compared with wild type (Liu and Zhu,1997). Furthermore, some transport proteins includesensor domains or function in association with sen-sors (Ozcan et al., 1998; Heinisch et al., 1999; Sabirovet al., 1999) as could the putative Na1/H1 antiporterSOS1 (Shi et al., 2000).
The experimental evidence presented here indi-cates that the SOS pathway controls expression ofonly a few salt stress-specific tolerance determinantgenes among the numerous genes (six of 89 in thisstudy) that are regulated in the plant response toNaCl treatment (Zhu et al., 1997). This is similar tothe paradigm that has been established recently forthe salt stress response of the unicellular eukaryoteyeast (Saccharomyces cerevisiae). Genome-wide arrayanalysis determined that osmotic upshock causes arapid and multi-fold increase in mRNA of between186 and 1,359 genes and reduced transcript abun-dance of more than 100 genes depending on theseverity of osmotic shock, the osmotic agent (NaCl orsorbitol), and time after treatment (Posas et al., 2000;Rep et al., 2000; J. Yale and H.J. Bohnert, unpublisheddata). Salt-induced expression of most is either par-tially or completely controlled by the high osmolarityglycerol and mitogen-activated protein kinase path-way. The yeast calcineurin pathway is analogous tothe Arabidopsis SOS pathway, controls ion ho-meostasis and is essential for salt tolerance in yeast(Mendoza et al., 1994, 1996), and affects expression ofmany fewer osmotic responsive genes (T.K. Matsu-moto, unpublished data). Like the SOS pathway, cal-cineurin regulates expression of genes that encodesalt tolerance effectors such as Na1 efflux transport-ers (Mendoza et al., 1994; Shi et al., 2000; Matusmotoet al., unpublished data).
It is conceivable that signal transduction throughthe SOS pathway that mediates salt tolerance mayhave a substantial component that involves posttran-scriptional activation of salt tolerance effectors, par-ticularly over the time span (minimum of 4 h) of thesalt treatment used in experiments reported here.Plant survival in severe stress likely requires veryimmediate cellular responses, whereas transcrip-tional regulation may be sufficient for stress recoveryand adaptation. Notwithstanding, salt induces tran-scriptional activation of genes in yeast within min-utes (Posas et al., 2000; Rep et al., 2000). Genes thatare transiently induced or weakly expressed furthercomplicate inference of function from expressionprofile analysis. The majority of yeast genes inducedby mild salt shock exhibit transient expression (Posaset al., 2000). Determinant gene transcript abundancedifferences in wild type and sos3 may be insufficientfor the resolution limits of the subtraction protocols,yet are biologically meaningful to salt stressadaptation.
It is interesting that the SOS pathway negativelycontrols the expression of four salt-regulated genes(SAMT, C0R 6.6/KIN2, STZ, and CCR1), three (SAMT,
C0R6.6/KIN2, and STZ) of which are induced byNaCl treatment. So, salt tolerance determinants in-clude genes that must be repressed, at least tempo-rally, during the plant stress response. Negativelyregulated genes may include those that contribute togrowth arrest necessary during the period of adjust-ment or ameliorate other etiologies that occur coin-cidentally with salt in the native environment of theorganism. CCR1 and COR6.6/KIN2 are cold induced,whereas SAMT is implicated in plant defense againstpathogens indicating their principal function is not insalt adaptation. Together, this suggests that a func-tion of the SOS pathway is to discriminate against themyriad of stress signals that are elicited by salt and tofocus the capacity of the plant to cope with the prin-cipal etiology; in this instance, ion dis-equilibrium.Furthermore, the SOS pathway may coordinate tem-poral gene expression to focus the availability ofeffectors, as required, during stress perception, ame-lioration, or adaptation. Confirmation that the genesidentified in this study encode tolerance determi-nants awaits molecular genetic confirmation by loss-or gain-of-function experimentation.
MATERIALS AND METHODS
Plant Material
Arabidopsis (ecotype Columbia-0 gl1) and sos1, sos2, andsos3 were wild-type and salt-hypersensitive genotypes, re-spectively (provided by Dr. Jian-Kang Zhu, University ofArizona). Seeds were surface disinfected and stratified for2 d at 4°C. Seeds were germinated in liquid medium (Mu-rashige and Skoog salts [Murashige and Skoog, 1962] and3% [w/v] Suc [pH 5.8]) in 250-mL flasks on a gyratoryshaker (80–100 rpm) under low-intensity Cool White fluo-rescent illumination (light/dark:16/8 h daily) at 22°C to24°C. After 14 d, seedlings were transferred to fresh me-dium without or with 160 mm NaCl for the time intervalindicated. Seedlings were harvested, frozen in liquid nitro-gen, and stored at 280°C.
Construction of Subtraction Libraries
Total RNA was isolated as described by Gong et al.(1997). mRNA was isolated using the Poly(A)1 RNA puri-fication kit (CLONTECH, Palo Alto, CA). The PCR-SelectcDNA subtraction kit (K1804-1, CLONTECH) was used toobtain subtracted cDNA libraries.
Subtraction of cDNAs Obtained from mRNA of Salt-Treated sos3 and gl1
Subtractive hybridization was used to identify cDNAscorresponding to salt regulated genes differentially ex-pressed in sos3. Both forward and reverse subtractive hy-bridizations were performed with salt-treated (160 mmNaCl, 4 h) seedlings. The forward subtraction used testercDNA obtained from mRNA of gl1 and driver cDNA fromsos3. In the reverse subtraction, the tester cDNA was ob-
Gong et al.
372 Plant Physiol. Vol. 126, 2001

tained from sos3 and driver cDNA from gl1. Driver cDNAsare the reference and are targets for elimination duringsubtraction leaving unique tester cDNAs. Forward andreverse subtractive hybridization was meant to identifysalt-responsive genes that are specifically regulated in sos3.
Subtraction of cDNAs Obtained from gl1 afterTreatment without or with NaCl
This subtraction was intended for identification of genesdifferentially regulated by salt in gl1. The forward subtrac-tion tester cDNA was obtained from mRNA of gl1 seed-lings 4 h after transfer to medium with 160 mm NaCl anddriver cDNA from gl1 plants grown in medium withoutsalt. In the reverse subtraction, tester and driver cDNA wasobtained from gl1 grown without and with 160 mm NaCl,respectively.
The subtracted libraries were subjected to two rounds ofPCR amplification, the second using nested primers foradaptors 1 and 2R (CLONTECH). The PCR products wereligated into pT-Adv (CLONTECH), and transformed intoEscherichia coli. The white colonies were isolated and insertsamplified by PCR.
Enrichment of Unique Salt-Regulated cDNAs in theSubtraction Library
A modification of the differential subtraction chainmethod (Luo et al., 1999) was used to enrich the subtractedlibrary for unique salt-regulated cDNAs. The driver for thissubtraction (driver 2) is a mixture of second PCR productfrom reverse subtraction (100 mL; tester DNA:gl1 withoutsalt, driver DNA:gl1 with salt) and PCR products amplifiedfrom highly repetitive salt-induced cDNAs isolated fromprevious rounds of screening (0.5 mL of each). To removeadaptor sequences from driver 2, the mixture was digestedat the restriction sites of adaptors 1 (SmaI and RsaI) and 2R(EagI and RsaI) by consecutive restriction digestion usingSmaI, and then EagI 1 RsaI. The digested DNA was ex-tracted with phenol and phenol/chloroform, and then pre-cipitated with ethanol. The precipitate was redissolved in100 mL of water and passed through the Microcon YM-30column (Amicon, Beverly, MA) to separate the DNA fromAdaptor fragments. Driver 2 DNA was recovered in 20 mLof water and used as driver in the following PCR subtrac-tion. About 10 times excess amount of driver 2 DNA (2 mL)was mixed with 1mL of forward subtraction library (driverand tester 5 gl1 grown without and with 160 mm NaCl,respectively) in hybridization buffer {5 mm HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid]-HCl (pH8.3), 12 mm NaCl, and 0.05 mm EDTA} in a total volume of12 mL. The hybridization solution was denatured at 94°Cfor 5 min, and incubated at 72°C for 12 h.
After hybridization, the mixture was precipitated withethanol, and digested with mung bean nuclease (Promega,Madison, WI) to remove adaptor sequence from productsof the driver-tester hybridization. Adaptor sequences intester-tester hybridization products are aligned withmatching ends and these are not digested by mung bean
nuclease. The mung bean nuclease digested mixture waspassed through the Microcon column and purified DNAwas rehybridized at 72°C. The mung bean nuclease diges-tion and DNA purification procedures were repeated. TheDNA was amplified by PCR using nested primers for 1 and2R, 11 cycles. The cloning of PCR products was as de-scribed as above.
Dot-Blot Analysis
For dot-blot analysis, cDNA inserts of forward subtrac-tion library clones were individually amplified by PCR and2 mL of PCR product was mixed with 2 mL of 0.6 m NaOH.Two microliters of the mixture was blotted onto each oftwo duplicate nylon membrane filters. Probes for dot-blotanalysis were either forward or reverse subtraction prod-ucts or cDNA of RNA from plants, as indicated. The prod-ucts of forward and reverse subtraction were digested withSmaI, RsaI, and EagI to remove the adapter sequences andlabeled with 32P using the Ready-To-Go kit (AmershamPharmacia Biotech, Piscataway, NJ). The labeled forwardprobes were hybridized to one membrane and the reverseprobes to the duplicate.
Template Preparation, DNA Sequencing, andData Analysis
Plasmid templates were prepared from selected bacterialcolonies by 96-well alkaline lysis minipreps according tothe manufacturer’s instructions (Edge BioSystems, Inc.,Gaithersburg, MD).
DNA sequencing reactions were conducted usingDyeDeoxy “Terminator PRISM” mix (Perkin-Elmer-ABI,Foster City, CA) according to the manufacturer’s instruc-tions in a multiplate thin-wall 96-well microplate on an MJResearch PTC-100-96 (MJ Research, Watertown, MA) pro-grammable thermal controller using the following profile:96°C for 30 s, 45°C for 15 s, and 60°C for 4 min for 49 cycles.Unincorporated dye terminators were removed by passingreactions over a 96-well gel filtration block (Edge BioSys-tems). Recovered sequencing reaction products were ana-lyzed on either an ABI 373A-XL Stretch or an ABI 3700capillary array automated DNA sequencing system(Perkin-Elmer Applied BioSystems). Raw sequence datawas analyzed using PHRED (Ewing and Green, 1998; Ew-ing et al., 1998) and Cross match to removal vector se-quences. Additional vector sequence removal and editingwas done manually using FACTURA software (Perkin-Elmer Applied BioSystems). Polished EST sequence fileswere assembled into singleton and contig files usingPHRAP (P. Green, unpublished data). EST identities weredetermined by sequence comparison to the nonredundantGenBank database using BLASTN (BLAST 2.0) using de-fault parameters (Altschul et al., 1997). In instances wherean unannotated match was obtained, BLASTX searcherswere conducted and sequence homology information wasused to assign putative identities. All EST sequencesreported here have been deposited in dbEST and can bebrowsed and retrieved from the NCBI website(http://www.ncbi.nlm.nih.gov) under accession numbersBE844684 through BE845405.
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 373

Northern-Blot Analysis of Putative Clones
Total RNA was isolated from seedlings and 40 mg fromeach sample was separated on 1.2% (w/v) agarose formal-dehyde gels and transferred to Hybond-N nylon mem-branes (Amersham) as previously described (Gong et al.,1997). The cDNA insert of each clone was amplified by PCRusing nested primers that hybridize to adaptors 1 and 2R,and purified from agarose gels using the Qiagen Gel Puri-fication kit. The probes for RD22BP1, MYB2, PAL, STZ, andACP1 were obtained by PCR amplification using cDNAobtained from mRNA of salt-treated gl1 as template;RD22BP1 (AB000875), forward primer: 59-ATGACGCTGT-TGATGAGGAG-39 and reverse primer: 59-TTTCGGATT-CTGGGTCTGAG-39 (0.56 kb); MYB2 (D14712), forwardprimer: 59-GAAATGGAAGATTACGAGCG-39 and reverseprimer: 59-TTAATTATACGAATACGATGTC-39 (1.0 kb);PAL (L33677), forward primer: 59-ATGGAGATT-AACG-GGGCACAC-39 and reverse primer: 59-ACGT-TCACCG-TTGGGACCAG-39 (1.1 kb); STZ (X95573), ORF; and ACP1(AF009228), forward primer: 59-CAA-AAGCCATTTTT-CAAATTTCAAACTCAG and reverse primer 59-GTTTT-CAATGATAGTGAAGAAAGATG-TAC-AAC (0.83 kb).The purified PCR products were labeled using 32P dCTPusing the Ready-To-Go kit. Blot-blot hybridization andwashes were as described (Gong et al., 1997). The blotswere stripped by boiling in 0.5% (w/v) SDS solution for 3min and were rehybridized with another probe.
ACKNOWLEDGMENTS
The authors would like to thank Sue Ann Hudiburg andJanet Rogers of the Oklahoma State University Recombi-nant DNA/Protein Resource Facility for providing oligo-nucleotide synthesis and automated DNA sequencing anal-ysis services.
Received September 29, 2000; returned for revision Decem-ber 15, 2000; accepted February 9, 2001.
LITERATURE CITED
Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T,Hosokawa D, Shinozaki K (1997) Role of ArabidopsisMYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 9: 1859–1868
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z,Miller W, Lipma DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database searchprograms. Nucleic Acids Res 25: 3389–3402
Berger S, Bell E, Sadka A, Mullet JE (1995) Arabidopsisthaliana Atvsp is homologous to soybean VspA and VspB,genes encoding vegetative storage protein acid phospha-tases, and is regulated similarly by methyl jasmonate,wounding, sugars, light and phosphate. Plant Mol Biol27: 933–942
Bohnert HJ, Nelson DE, Jensen RG (1995) Adaptation toenvironmental stresses. Plant Cell 7: 1099–1111
Bray EA (1994) Molecular responses to water deficit. PlantPhysiol 103: 1035–1040
Carpenter CD, Kreps JA, Simon AE (1994) Genes encodingglycine-rich Arabidopsis thaliana proteins with RNA-
binding motifs are influenced by cold treatment and anendogenous circadian rhythm. Plant Physiol 104: 1015–25
Dudareva N, Murfitt LM, Mann CJ, Gorenstein N, Ko-losova N, Kish CM, Bonham C, Wood K (2000) Develop-mental regulation of methyl benzoate biosynthesis andemission in snapdragon flowers. Plant Cell 12: 949–961
Ewing B, Green P (1998) Base-calling of automated se-quencer traces using Phred: II. Error probabilities. Ge-nome Res. 8: 186–194
Ewing B, Hillier L, Wendl MC, Green P (1998) Base-calling of automated sequencer traces using Phred: I.Accuracy assessment. Genome Res. 8: 175–185
Gong Z, Yamazaki M, Sugiyama M, Tanaka Y, Saito K(1997) Cloning and molecular analysis of structuralgenes involved in anthocyanin biosynthesis and ex-pressed in a forma-specific manner in Perilla frutescens.Plant Mol Biol 35: 915–927
Halfter U, Ishitani M, Zhu J-K (2000) The Arabidopsis SOS2protein kinase physically interacts with and is activatedby the calcium-binding protein SOS3. Proc Natl Acad SciUSA 97: 3735–3740
Hanson AD, Rathinasabapathi B, Rivoal J, Burnet M,Dillon MO, Gage DA (1994) Osmoprotective com-pounds in the Plumbaginaceae: a natural experimentinmetabolic engineering of stress tolerance. Proc NatlAcad Sci USA 91: 306–310
Hasegawa PM, Bressan RA, Pardo JM (2000a) The dawn ofplant salt tolerance genetics. Trends Plant Sci 5: 317–319
Hasegawa PM, Bressan RA, Zhu J-K, Bohnert HJ (2000b)Plant cellular and molecular responses to high salinity.Annu Rev Plant Physiol Plant Mol Biol 51: 463–499
Heinisch JJ, Lorberg A, Schmitz H-P, Jacoby JJ (1999) Theprotein kinase C-mediated MAP kinase pathway in-volved in the maintenance of cellular integrity in Saccha-romyces cerevisiae. Mol Microbiol 32: 671–680
Heintzen C, Nater M, Apel K, Staiger D (1997) AtGRP7, anuclear RNA-binding protein as a component of acircadian-regulated negative feedback loop in Arabidopsisthaliana. Proc Natl Acad Sci USA 94: 8515–8520
Hwang I, Goodman H (1995) An Arabidopsis thaliana root-specific kinase homolog is induced by dehydration, ABAand NaCl. Plant J 8: 37–43
Ingram J, Bartels D (1996) The molecular basis of dehy-dration tolerance in plants. Annu Rev Plant Physiol PlantMol Biol 47: 377–403
Ishitani M, Xiong L, Stevenson B, Zhu J-K (1997) Geneticanalysis of osmotic and cold stress signal transduction inArabidopsis: interactions and convergence of abscisicacid-dependent and abscisic acid-independent path-ways. Plant Cell 9: 1935–1949
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shi-nozaki K (1999) Improving plant drought, salt and freez-ing tolerance by gene transfer of a single stress-inducibletranscriptional factor. Nature Biotech 17: 287–291
Koiwa H, Bressan RA, Hasegawa PM (1997) Regulation ofprotease inhibitors and plant defense. Trends Plant Sci 2:379–384
Kovtun Y, Chiu W-L, Tena G, Sheen J (2000) Functionalanalysis of oxidative stress-activated mitogen-activated
Gong et al.
374 Plant Physiol. Vol. 126, 2001

protein kinase cascade in plants. Proc Natl Acad Sci USA97: 2940–2945
Kreps JA, Simon AE (1997) Environmental and geneticeffects on circadian clock-reggulated gene expression inArabidopsis. Plant Cell 9: 297–304
Kurkela S, Borg-Franck M (1992) Structure and expressionof KIN2, one of two cold- and ABA-induced genes ofArabidopsis thaliana. Plant Mol Biol 19: 689–692
Kurkela S, Franck M (1990) Cloning and characterizationof a cold- and ABA-inducible Arabidopsis gene. Plant MolBiol 15: 137–144
Lippuner V, Cyert MS, Gasser CS (1996) Two classes ofplant cDNAs differentially complement yeast calcineurinmutants and increase salt tolerance of wild-type yeast.J Biol Chem 271: 12859–12866
Liu J, Ishitani M, Halfter U, Kim C-S, Zhu J-K (2000) TheArabidopsis thaliana SOS2 gene encodes a protein kinasethat is required for salt tolerance. Proc Natl Acad SciUSA 79: 3730–3734
Liu J, Zhu J-K (1997) Proline accumulation and salt-stress-induced gene expression in a salt-hypersensitive mutantof Arabidopsis. Plant Physiol 114: 591–596
Liu J, Zhu J-K (1998) A calcium sensor homolog requiredfor plant salt tolerance. Science 280: 1943–1945
Luo J-H, Puc JA, Slosberg ED, Yao Y, Bruce JN, WrightTC, Becich MJ, Parsons R (1999) Differential subtractionchain, a method for identifying differences in genomicDNA and mRNA. Nucleic Acids Res 27: 24e
Mason HS, Mullet JE (1990) Expression of two soybeanvegetative storage protein genes during developmentand in response to water deficit, wounding, and jasmonicacid. Plant Cell 2: 569–579
Mendoza I, Quintero FJ, Bressan RA, Hasegawa PM, PardoJM (1996) Activated calcineurin confers high tolerance toion stress and alters the budding pattern and cell mor-phology of yeast cells. J Biol Chem 271: 23061–23067
Mendoza I, Rubio F, Rodriguez-Navarro A, Pardo JM(1994) The protein phosphatase calcineurin is essentialfor NaCl tolerance of Saccharomyces cerevisiae. J BiolChem 269: 8792–8796
Mikami K, Katagiri T, Iuchi S, Yamaguchi-Shinozaki K,Shinozaki K (1998) A gene encoding phosphatidylino-sitol-4-phosphate 5-kinase is induced by water stress andabscisic acid in Arabidopsis thaliana. Plant J 15: 563–568
Mizoguchi T, Irie K, Hirayama T, Hayashida N,Yamaguchi-Shinozaki K, Matsumoto K, Shinozaki K(1996) A gene encoding a mitogen-activated protein ki-nase kinase kinase is induced simultaneously with genesfor a mitogen-activated protein kinase and an S6 ribo-somal protein kinase by touch, cold, and water stress inArabidopsis thaliana. Proc Natl Acad Sci USA 84: 765–769
Murashige T, Skoog F (1962) A revised medium for rapidgrowth and bioassays with tobacco tissue culture.Physiol Plant 15: 473–479
Nelson DE, Shen B, Bohnert HJ (1998) The regulation ofcell-specific inositol metabolism and transport in plantsalinity tolerance. Plant Cell 10: 753–764
Niu X, Bressan RA, Hasegawa PM, Pardo JM (1995) Ionhomeostasis in NaCl stress environments. Plant Physiol109: 735–742
Ozcan S, Dover J, Johnston M (1998) Glucose sensing andsignalling by two glucose receptors in the yeast Saccha-romyces cerevisiae. EMBO J 17: 2566–2573
Piao HL, Pih KT, Lim JH, Kang SG, Jin JB, Kim SH,Hwang I (1999) An Arabidopsis GSK3/shaggy-like gene thatcomplements yeast salt stress-sensitive mutant is inducedby NaCl and abscisic acid. Plant Physiol 119: 1527–1534
Posas F, Chambers JR, Heyman JA, Hoeffler JP, NadalED, Arino J (2000) The transcriptional response of yeastto saline stress. J Biol Chem 275: 17249–17255
Rep M, Krantz M, Thevelein JM, Hohmann S (2000) Thetranscriptional response of Saccharomyces cerevisiae to os-motic shock. J Biol Chem 275: 8290–8300
Reymond P, Weber H, Damond M, Farmer EE (2000).Differential gene expression in response to mechanicalwounding and insect feeding in Arabidopsis. Plant Cell,12: 707–719
Ross JR, Nam KH, D’Auria JC, Pichersky E (1999) S-adenosyl-L-methionine: salicylic acid carboxyl methyl-transferase, an enzyme involved in floral scent produc-tion and plant defense, represents a new class of plantmethyltransferases. Arch Biochem Biophys 367: 9–16
Sabirov RZ, Azimov RR, Ando-Akatsuka, Miyoshi T,Okada Y (1999) Na1 sensitivity of ROMK1 K1 channel:role of the Na1/H1 antiporter. J Membr Biol 172: 67–76
Sanders D (2000) Plant biology: the salty tale of Arabidopsis.Curr Biol 10: R-486–R-488
Sheen J (1996) Ca21-dependent protein kinases and stresssignal transduction in plants. Science 274: 1900–1902
Shi H, Ishitani M, Kim C, Zhu J-K (2000) The Arabidopsisthaliana salt tolerance gene SOS1 encodes a putativeNa1/H1 antiporter. Proc Natl Acad Sci USA 97: 6896–6901
Shinozaki K, Yamaguchi-Shinozaki K (1997) Gene expres-sion and signal transduction in water-stress response.Plant Physiol 115: 327–334
Thomashow MF (1999) Plant cold Acclimation: freezingtolerance genes and regulatory mechanisms. Annu RevPlant Physiol Plant Mol Biol 50: 571–599
Utsugi S, Sakamoto W, Murata M, Motoyoshi F (1998)Arabidopsis thaliana vegetative storage protein (VSP)genes: gene organization and tissue-specific expression.Plant Mol Biol 38: 565–576
Winicov I (2000) Alfin1 transcription factor overespressionenhances plant root growth under normal and salineconditions and improves salt tolerance in alfalfa. Planta210: 416–22
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K, Shinozaki K (1997) Regulation of levels ofproline as an osmolyte in plants under water stress. PlantCell Physiol 38: 1095–1102
Zhu J-K (2000) Genetic analysis of plant salt toleranceusing Arabidopsis thaliana. Plant Physiol 124: 941–948
Zhu J-K, Hasegawa PM, Bressan RA (1997) Molecularaspects of osmotic stress in plants. Crit Rev Plant Sci 16:253–277
Salt and SOS-Regulated Genes
Plant Physiol. Vol. 126, 2001 375
Copyright © 2022 FDOKUMEN




















