
Gastrointestinal nematode infections in roe deer (Capreolus capreolus) from the NW of the Iberian...
26
1 Gastrointestinal nematode infections in roe deer (Capreolus capreolus) from 1 the NW of the Iberian Peninsula: assessment of some risk factors 2 3 Pato FJ 1 , Vázquez L 1 , Díez-Baños N 2 , López C 1 , Sánchez-Andrade R 1 , Fernández G 1 , Díez- 4 Baños P 1 , Panadero R 1 , Díaz, P 1 , Morrondo P 1 5 6 7 1 Department of Animal Pathology (INVESAGA Group). Faculty of Veterinary Sciences. 8 University of Santiago de Compostela. 27002 Lugo, Spain. 9 10 2 Department of Animal Health. Faculty of Veterinary Sciences. University of León, 24071 11 León, Spain. 12 13 Corresponding author: Tel.: +34 982822102; fax +34 982822001 E-mail adress : patrocinio.morrondo @usc.es. (P. Morrondo) *Manuscript Click here to view linked References
Transcript of Gastrointestinal nematode infections in roe deer (Capreolus capreolus) from the NW of the Iberian...

1
Gastrointestinal nematode infections in roe deer (Capreolus capreolus) from 1
the NW of the Iberian Peninsula: assessment of some risk factors 2
3
Pato FJ1, Vázquez L
1, Díez-Baños N
2, López C
1, Sánchez-Andrade R
1, Fernández G
1, Díez-4
Baños P1, Panadero R
1, Díaz, P
1, Morrondo P
1 5
6
7
1Department of Animal Pathology (INVESAGA Group). Faculty of Veterinary Sciences. 8
University of Santiago de Compostela. 27002 Lugo, Spain. 9
10
2Department of Animal Health. Faculty of Veterinary Sciences. University of León, 24071 11
León, Spain. 12
13
Corresponding author: Tel.: +34 982822102; fax +34 982822001
E-mail adress: patrocinio.morrondo @usc.es. (P. Morrondo)
*ManuscriptClick here to view linked References

2
Abstract 14
Intestinal contents of 218 roe deer hunted in the northwest (NW) of the Iberian 15
Peninsula during the 2008-2009 hunting seasons were examined in order to provide information 16
on the gastrointestinal (GI) nematode prevalence and intensity of infection and the possible 17
influence of some environmental and intrinsic factors such as climatic conditions, age and 18
sex. 19
All the animals studied harboured GI nematodes, and a total of 20 different species 20
belonging to ten genera were identified. Spiculopteragia spiculoptera/Spiculopteragia 21
mathevossiani, Ostertagia leptospicularis/Ostertagia kolchida and Nematodirus filicollis were 22
the most common. This is the first citation for Chabertia ovina, Cooperia pectinata, C. 23
punctata, C. oncophora, Haemonchus contortus, Nematodirus spathiger, Oesophagostomum 24
venulosum, Teladorsagia trifurcata, Trichostrongylus capricola, T. colubriformis, T. vitrinus 25
and Trichuris capreoli in roe deer from the Iberian Peninsula. Prevalence and intensity were 26
significantly higher in the abomasum, where infections with more than one GI nematode 27
species were the most common; in the other intestinal segments infections with only one GI 28
nematode species were the most prevalent. 29
When considering the influence of the different risk factors on the prevalence of GI 30
nematodes, the highest prevalence for most of the genera were observed in roe deer from 31
coastal areas, where climatic conditions are more favourable for the development and survival 32
of third stage larvae in the environment. Regarding the sex of the animals, the prevalence was, 33
in general, higher in males than in females, probably due to behavioural and physiological 34
sex-related differences. On the contrary, no differences were found in relation to the age of 35
the animals. 36

3
This study reveals that roe deer from the NW of the Iberian Peninsula are widely and 37
intensely infected with gastrointestinal nematodes, which probably affect the health status of 38
these ungulates. 39
40
41
Key words: gastrointestinal nematode species; roe deer; abomasum; intestine; risk factors; 42
Iberian Peninsula. 43
44

4
1. Introduction 45
The distribution of the European roe deer (Capreolus capreolus, Linnaeus, 1758) 46
populations is influenced by forest structure, vegetation characteristics and human 47
disturbance. Roe deer live in woodlands or farmland providing permanent cover, although 48
they are increasingly found on open areas. They prefer humid and cold environmental 49
conditions, which may explain the lower densities recorded in Spain in relation to other 50
European areas (Centenera, 2005; Burbaitė and Csányi, 2009). However, the progressive 51
abandonment of agricultural lands in the northwest (NW) of the Iberian Peninsula has caused 52
the appearance of alternating grazing and forest areas, representing an ideal environment for 53
roe deer, and the great diversity of plant resources in this area has led to a considerable 54
increase in the census of that ungulate in the last three decades (Centenera, 2005). 55
The NW of the Iberian Peninsula has climatic conditions which favour the survival of 56
parasitic infective forms in the environment, especially gastrointestinal (GI) nematodes, thus 57
enabling the possibility for infection throughout the year (Mezo-Menéndez et al., 1995; 58
Nogareda et al., 2006). There are a number of reports of domestic ruminants (Díaz et al., 59
2005; Díez-Baños et al., 1992, 2008) and roe deer (Vázquez et al., 2009) parasitized with GI 60
nematodes in this region. The significance of GI nematode infections in roe deer has been 61
previously reported in different European countries (Jansen, 1975; Borgsteede et al., 1990; 62
Kochko, 1997; Rossi et al., 1997; Ferté et al., 1999; Rehbein et al., 2000; Balicka-Ramisz et 63
al., 2003; Pilarczyk et al., 2005; Bolukbas et al., 2012). 64
In previous studies it was also found that roe deer from the NW of the Iberian 65
Peninsula presents a high prevalence of protozoa of the genera Sarcocystis (López et al., 66
2003), Eimeria (Díaz et al., 2010), Neospora and Toxoplasma (Panadero et al., 2010), 67
bronchopulmonary nematodes (Panadero et al., 2001) and ixodid ticks (Vázquez et al., 2011). 68

5
However, in relation to the prevalence of gastrointestinal nematode infection, only a few 69
studies have been conducted on roe deer from south and central Spain (Navarrete et al., 1990; 70
Reina et al., 1992; Ramajo et al. 2007). 71
The main objective of this study was to identify the species of GI nematodes affecting 72
roe deer in the NW of the Iberian Peninsula as well as to determine their prevalence and 73
intensity of infection, taking into account their location in the different parts of the digestive 74
system. This study also reports the effect of some environmental (climatic conditions) and 75
intrinsic (age, sex) factors on the GI nematode roe deer infection. 76
77
2. Material and methods 78
2.1. Study area 79
The NW of the Iberian Peninsula (43º47'-41º49' N, 6º42'-9º18' W) is an important 80
livestock-rearing area with an oceanic climate characterized by mild temperature and high 81
precipitation (Díaz et al., 2007), maintaining a high level of humidity; however, the existence 82
of climatic differences allow a division into three zones, previously described by Vázquez et 83
al. (2011): a Coastal area situated from sea level to 200 m, with moderate mean annual 84
precipitation (1,300-1,500 mm) and temperature (14ºC); a Central area situated at 200-650 m 85
with lower precipitation (900-1,300 mm) and temperature (11.5ºC); and a mountainous area 86
situated at 650-2,100m), characterized by the lowest temperature (10ºC) and the highest 87
precipitation (> 1,500 mm). 88
In this region there are large areas of woodland, mainly composed of Quercus robur 89
and Castanea sativa. In winter, roe deer eat mostly shrubs (Genista tridentata, Vaccinium 90
myrtillus, Daboecia cantabrica), whereas in spring and summer months they feed on grass 91
(Poa pratensis, Lolium perenne and Festuca pratensis) and tender shoots of deciduous trees 92
(Salix alba, Betula pendula, Alnus glutinosa, Populus alba) (Ortega, 2009). 93

6
94
2.2. Animals 95
A total of 218 roe deer killed during two consecutive hunting seasons (April-October 96
2008-2009) in the NW of the Iberian Peninsula were examined in this study. According to 97
hunting policy, only males older than two years-old should be killed; females are hunted 98
under special conditions. For that reason most roe deer were male (n=187) and only 31 were 99
females. Animals were also divided in two age-groups, young (< 3 years old; n=30) and 100
adults (≥ 3 years old; n=188), according to their teeth features. 101
Animals were eviscerated after killing and the different organs were individually 102
bagged up, identified and frozen at -20 °C until processed in the laboratory. 103
104
2.3. Parasitological procedures 105
Abomasum and small and large intestine contents were individually examined. In the 106
large intestine, the cecum was also independently considered. The different areas of the 107
intestine were identified and sealed with ligatures at the pylorus, ileocecal junction and the 108
rectum to ensure that the nematodes remained in each of the different parts of the GI tract. 109
Once the different sections were opened, the contents and washings of mucosa were rinsed 110
using a set of three sieves of meshes 1.5 mm, 400 µm and 150 µm. After the filtration of each 111
segment, the material retained on the sieves was collected and stored in 5% formaldehyde. A 112
50% aliquot was examined for worms; adult nematodes were extracted under 113
stereomicroscope at 40x, counted and classified according to their sex. For species 114
identification, males were mounted on semipermanent preparations and identified on the basis 115
of reported morphological descriptions (Skrjabin et al., 1954; Yamaguti, 1961; Gibbons and 116
Khalil, 1982; Durette-Desset, 1983, 1989; Taylor et al., 2007). Moreover, it was also taken 117

7
into consideration the existence of Ostertagiinae male polymorphism that affects the 118
morphology and length of spicules and the genital cone (Dróźdź, 1995). Several approaches 119
have supported the existence of the following pairs: Teladorsagia circumcincta (minor morph 120
Teladorsagia trifurcata; Lancaster et al., 1983); Ostertagia leptospicularis (minor morph 121
Ostertagia kolchida; Dróźdź, 1995, Ortiz et al., 1996) and Spiculopteragia spiculoptera 122
(minor morph Spiculopteragia mathevossiani; Liénard et al., 2006). However, the different 123
morphotypes were considered as different species for data analysis. 124
125
2.4. Statistical analysis 126
Data were analysed using PASW Statistics (version 18.0) for Windows. The Chi-127
square test was applied to determine significant differences (p≤ 0.05) in the prevalence of GI 128
nematode species between the different digestive segments (abomasum, small intestine, large 129
intestine and cecum). For the intensity of infection (Bush et al., 1997), the arithmetic mean 130
( x ) was used as a central tendency measure and the standard deviation (SD) as a dispersion 131
measure. Non parametric Kruskal-Wallis and Mann-Whitney tests were used to compare the 132
intensity of infection between the different digestive segments. 133
Due to the low prevalence observed for some species and morphs, only GI genera 134
were included in the analysis of the extrinsic and intrinsic factors. The existence of significant 135
differences in both the prevalence and intensity of infection by GI nematode genera 136
considering the different variables studied (climatic area, sex and age) were analysed using 137
Chi-square and Kruskal-Wallis tests, respectively. Odds ratios (OR) with 95% confidence 138
intervals (95% CI) were also calculated. 139
140
3. Results 141

8
All the roe deer examined (100%) were infected by GI nematodes and the number of 142
worms per animal ranged from 3 to 10,072 ( x = 1,070; SD= 1,020). Moreover, 127,140 out of 143
217,786 adult nematodes recovered were females and 90,646 males, giving a female/male 144
ratio of 1.4. 145
The prevalence of infection was very high in the abomasum (100%) and small intestine 146
(91.8%) and lower in the large intestine (58.6%) and cecum (53.5%); these differences were 147
significant (p <0.001). The mean intensity in the abomasum ( x = 954; SD= 769) was higher 148
than in the small intestine ( x = 85; SD= 143), large intestine ( x = 3; SD= 2) and cecum ( x = 149
3; SD= 2); Kruskal-Wallis test showed that there were significant differences (p <0.001). 150
Mann-Whitney test also showed significant differences between the intensity of infection in 151
the abomasum and that of the rest of the GI segments (p <0.001 for all groups). 152
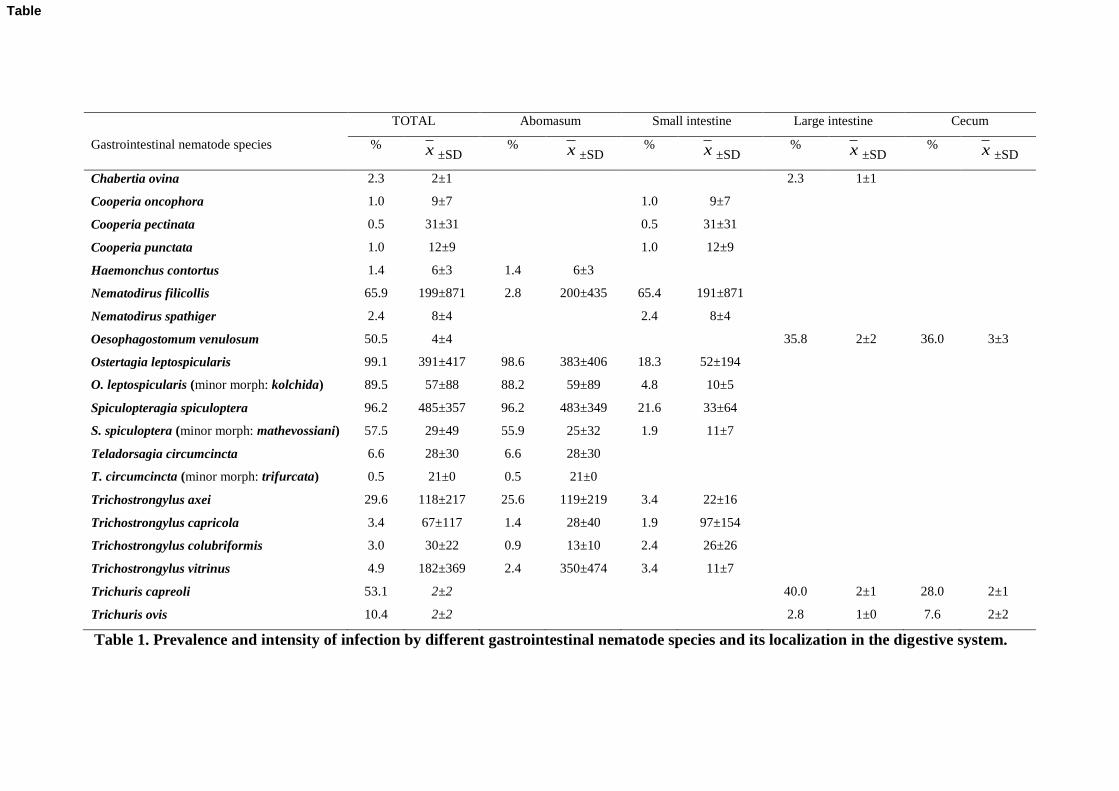
A total of 20 different species of GI nematodes were identified in this study, mainly in the 153
abomasum and in the small intestine. The percentage and mean intensity of infection for each 154
species, as well as its localization in the digestive system, are summarized in Table 1. 155
Ostertagia leptospicularis/O. kolchida, S. spiculoptera/S. mathevossiani and Nematodirus 156
filicollis were the dominant species. In the abomasum 12 species of GI nematodes were 157
identified. Ostertagia leptospicularis and S. spiculoptera were the species with the highest 158
prevalence and intensity of infection. In the small intestine 13 species were identified, and the 159
dominant species were N. filicollis, S. spiculoptera and O. leptospicularis. Abomasum and 160
small intestine shared nine GI species. In the large intestine and cecum Trichuris capreoli, 161
Trichuris ovis and Oesophagostomum venulosum were observed; Chabertia ovina was only 162
identified in the large intestine. 163
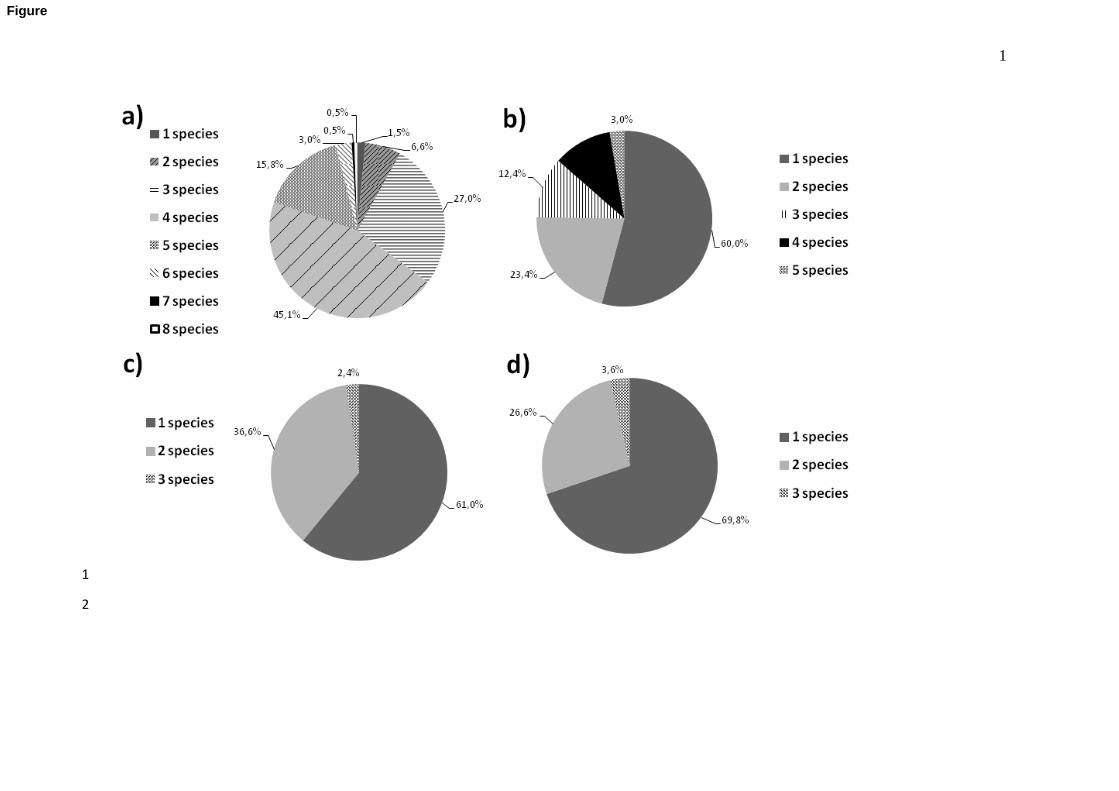
Infections with more than one GI nematode species were the most common in the 164
abomasum, whereas infections with one species were the most prevalent in the other segments 165

9
(Figure 1 a-d). In the abomasum (Fig 1a), infections by four species were the most frequent 166
(45.1%), being O. leptospicularis/O. kolchida and S. spiculoptera/S. mathevossiani the most 167
prevalent associations. Only one GI nematode species was identified in 60% of small 168
intestines, with a predominance of N. filicollis; associations of two species (23.4%) -with a 169
predominance of N. filicollis and S. spiculoptera- were detected in less proportion (Fig 1b). 170
Infection with T. capreoli was the most frequent in the large intestine (61%), followed by the 171
association O. venulosum and T. capreoli (Fig 1c). Finally, O. venulosum was the most 172
common GI nematode species in the cecum, followed by the association O. venulosum and T. 173
capreoli (Fig. 1d). 174
When considering the influence of the different risk factors on the GI prevalence and 175
intensity of infection (Table 2), roe deer from coastal areas presented the highest values for 176
most of the genera, and significant differences were found for Oesophagostomum (p= 0.012 177
and p <0.001, respectively); differences in the prevalence for Spiculopteragia and Trichuris 178
were also significant (p= 0.030 and p= 0.042 for prevalence and intensity, respectively). 179
Moreover, odds-ratio values showed that roe deer from coastal areas have a higher risk of 180
infection by Trichostrongylus (OR= 2.0; CI 95% 1.01-3.9), Oesophagostomum (OR= 2.0; CI 181
95% 1.01-3.8) and Trichuris (OR= 2.2; CI 95% 1.1-4.4). Regarding the sex of the animals 182
(Table 2), it was observed that the prevalence and intensity of infection were generally higher 183
in males than in females, with significant differences for Oesophagostomum (p= 0.016 and p< 184
0.001, respectively) and Trichuris (p< 0.001 and p< 0.001) and only in the intensity for 185
Spiculopteragia (p= 0.012). Moreover, the percentage of roe deer infected with Nematodirus 186
were significantly higher in males (p= 0.028) and the intensity in females (p= 0.014). Odds-187
ratio values showed that males have a higher risk of infection than females for Nematodirus 188
(OR= 2.3; CI 95% 1.1-5.0), Oesophagostomum (OR= 2.7; CI 95% 1.2-6.2) and Trichuris (OR 189

10
5.0; CI 95% 2.1-12.2). In contrast, no significant differences were found in relation to the age 190
of the animals (Table 2). 191
192
4. Discussion 193
In the last decades, the health status of wild animals has been gradually gaining 194
importance, and in this sense it has been reported that the presence of GI nematodes has a 195
negative impact on both the nutritional status and growth of roe deer (Zaffaroni et al., 1997; 196
Segonds-Pichon et al., 2000). The current study has revealed that GI nematodes are prevalent 197
parasites in roe deer in the NW of the Iberian Peninsula, since all animals examined 198
harboured at least one nematode species. Our data correlates with several studies conducted in 199
roe deer from Central and Southern Europe (Vetýška, 1980; Kochko, 1997; Rossi et al., 1997; 200
Rehbein et al., 2000; Balicka-Ramisz et al., 2003; Togni et al., 2004; Pilarczyk et al., 2005), 201
confirming the widespread occurrence of strongylate nematodes in the European roe deer. 202
The present work is one of the largest parasitological studies carried out in roe deer in 203
Spain; the high number of animals analysed and the extent of the study area (29.574 km2) may 204
explain the high number of GI nematode genera (n= 10) and species (n= 20) identified. 205
Although all the species detected have been previously reported in roe deer from other 206
European countries (Nickel et al., 1978; Bernard et al., 1988; Kutzer et al., 1988; Rossi et al., 207
1997; Zaffaroni et al., 2000; Bolukbas et al., 2012), this is the first report of C. ovina, 208
Cooperia pectinata, Cooperia punctata, Cooperia oncophora, Haemonchus contortus, 209
Nematodirus spathiger, O. venulosum, T. trifurcata, Trichostrongylus capricola, 210
Trichostrongylus colubriformis, Trichostrongylus vitrinus and T. capreoli in roe deer from the 211
Iberian Peninsula. 212

11
In addition, the dominant species found in this work (O. leptospicularis/kolchida and 213
S. spiculoptera/mathevossiani) have been frequently reported as the most common in 214
European roe deer (Borgsteede et al., 1990; Rossi et al., 1997; Rehbein et al., 2000; Zaffaroni 215
et al., 2000; Balicka-Ramisz et al., 2003; Togni et al., 2004; Bolukbas et al., 2012). Those 216
species are primarily considered pathogens of wild ruminants (Hoberg et al., 2001; Liénard et 217
al., 2006), even though they have been sporadically reported in domestic ruminants (Suarez 218
and Cabaret, 1992; Rickard et al., 1993; Stevenson et al., 1996; Silvestre et al., 2000). In 219
contrast, the presence of several species primarily parasites of livestock, such as those 220
belonging to the genera Chabertia, Cooperia, Haemonchus and Teladorsagia (Hoberg et al., 221
2001), suggests that roe deer grazed pasture previously grazed by domestic ruminants. The 222
NW of the Iberian Peninsula is an important livestock breeding area where cattle and sheep 223
are mainly reared in a semiextensive system, grazing every day and brought into the paddocks 224
at night (Pedreira et al., 2006; Díaz et al., 2007). Consequently, cross infection from livestock 225
to roe deer can occur in common grazing pastures. Previous studies have concluded that 226
infective larvae of the genera Ostertagia, Oesophagostomum, Trichostrongylus and Cooperia 227
are the most frequent in pastures from the northwest of Spain (Mezo-Menéndez et al., 1995; 228
Nogareda et al., 2006). 229
There was considerable variation in the nematode community between the different 230
segments of the GI tract. The highest prevalence and intensity were detected in the 231
abomasum, coinciding with previous reports in roe deer (Bernard et al., 1988; Rossi et al., 232
1997; Rehbein et al., 2000). Ostertagia leptospicularis/O. kolchida and S. spiculoptera/S. 233
mathevossiani were the most prevalent species in this segment, as found by several authors in 234
roe deer from other European countries (Jansen, 1975; Vetýška, 1980; Borgsteede et al., 235
1990; Dróźdź et al., 1992; Rossi et al., 1997; Ferté et al., 1999; Zaffaroni et al., 2000; Togni 236

12
et al., 2004; Bolukbas et al., 2012). Ostertagia kolchida and S. mathevosiani presented lower 237
prevalence and intensity of infection than their major morphs. 238
Nematodirus spp. were the most common parasites of the small intestine, and in the 239
current study N. filicollis was the dominant nematode species. Although several investigations 240
have found the roe deer-specific N. europaeus as the most prevalent Nematodirus species in 241
the small intestine of this wild ungulate (Borgsteede et al., 1990; Dróźdź et al., 1992; Jansen, 242
1992; Rossi et al., 1997), previous studies carried out in roe deer from Spain have reported 243
only N. filicollis within this genus (Navarrete et al., 1990; Reina et al., 1992; Ramajo et al., 244
2007). In addition, our data also coincide with that observed in roe deer from other European 245
countries, such as the United Kingdom (Dunn, 1965), France (Bernard et al., 1988) and 246
Turkey (Bolukbas et al., 2012). 247
The lowest prevalence and intensity of infection were detected in the large intestine 248
and the cecum; these results are consistent with that observed by other authors (Bernard et al., 249
1988; Rossi et al., 1997; Bolukbas et al., 2012). Trichuris ovis and O. venulosum were the 250
dominant nematode species; a number of studies have reported that these two species have a 251
high prevalence in European roe deer (Borgsteede et al., 1990; Rehbein et al., 2000; Bolukbas 252
et al., 2012). Chabertia ovina was only found in the large intestine of a small number of 253
animals, confirming that this species is rare or incidental in cervids (Borgsteede et al., 1990; 254
Hoberg et al., 2001). 255
Most of the discrepancies in prevalence values reported in the literature may be as a 256
result of different climatic conditions in the areas where roe deer were captured, which can 257
influence the life cycle of several species of GI nematodes. Prevalence and intensity of 258
infection were, in general, higher in roe deer from the coastal area, where temperature and 259
humidity conditions are more favourable for the development and survival of third stage 260

13
larvae of strongylate nematodes in the environment (Mezo-Menéndez et al., 1995, 1997; 261
Uriarte et al., 2003; Nogareda et al., 2006). 262
Behavioural and physiological sex-related differences may be the cause of the higher 263
prevalence and intensity observed in male roe deer. Males are more active than females, and 264
territorial bucks exhibit a pronounced territorial behaviour, aggressively expelling other males 265
(Melis et al., 2005). The expelled males are then forced to disperse, increasing the chance of 266
infection. Moreover, it has been also suggested that certain sexual hormones, such as sex 267
steroids, modulate several aspects of host immunity, so males could become more susceptible 268
than females to many infectious pathogens, including helminths (Poulin, 1996; Klein, 2000). 269
Several authors have reported that adult roe deer present lower GI nematode 270
prevalence and intensity than both fawns and old individuals (Borgsteede et al., 1990; 271
Segonds-Pichon et al., 2000; Body et al., 2011). In domestic ruminants, a general increase in 272
resistance to strongylid infection with age is observed (Kloosterman et al., 1991; Ploeger et 273
al., 1994; Bowman, 1999). Nevertheless, in the present study no significant differences on 274
both the prevalence and intensity of GI nematodes depending on the age of the animals were 275
detected, that could be related to the Spanish hunting policy, since it is only allowed to shoot 276
roe deer older than two years-old. Consequently, the group of young animals was mainly 277
composed by a low number of animals close to three years-old that have already had the 278
opportunity to get infected several times with GI nematode infective larvae and consequently, 279
may have developed a partial protective immune response. 280
In conclusion, this study reveals that roe deer from the NW of the Iberian Peninsula 281
are widely and intensely infected with GI nematodes. Significant differences were detected 282
when considering the climatic area and the sex of the animals. Infections by GI nematodes are 283
caused in this region by a high number of nematode species -12 were first cited in the Iberian 284

14
Peninsula- that may affect negatively the health status of these ungulates; on the other hand, 285
roe deer may act as potential reservoirs for other domestic or wild ruminants. 286
287
Acknowledgments 288
This work was supported by the Research Projects FAU2006-00006-00-00 and 289
07MRU034261PR and by a grant for consolidating and structuring competitive research 290
groups (CN2012/326, Xunta de Galicia). 291
292
References 293
Balicka-Ramisz, A., Cisek, A., Ramisz, A., Pilarczyk, B., 2003. Investigation of the lung, 294
stomach and intestine helminth infections of roe deer in North-West Poland. Tierarztl. 295
Umschau. 58, 489-491. 296
Bernard, J., Biesemans, W., Mathy, P., 1988. Nematodes parasites gastro-intestinaux des 297
Ongules gibier dans les Ardennes belges. Schweiz. Arch. Tierheilk. 130, 77-103. 298
Body, G., Ferté, H., Gaillard, J.M., Delorme, D., Klein, F., Gilot-Fromont, E., 2011. 299
Population density and phenotypic attributes influence the level of nematode 300
parasitism in roe deer. Oecologia 167, 635-646. 301
Bolukbas, C.S., Gurler, A.T., Beyhan, Y. E., M. Acici., Umur. S., 2012. Helminths of roe deer 302
(Capreolus capreolus) in the Middle Black Sea Region of Turkey. Parasitol. Int. 61, 303
729–730. 304
Borgsteede, F.H.M., Jansen, J., Van Nispen Tot Pannerden, H.P.M., Van Der Burg, W.P.J, 305
Noorman, N., Poutsma, J., Kotter, J.F., 1990. An investigation of the endoparasitic 306

15
helminth fauna of roe deer (Capreolus capreolus L.) in the Netherlands. Z. Jagdwiss. 307
36, 104-109. 308
Bowman, D.D., 1999. Georgis' parasitology for veterinarians. Saunders, Philadelphia, 414 pp. 309
Burbaitė, L., Csányi, S., 2009. Roe deer population and harvest changes in Europe. Eston. J. 310
Ecol. 58, 169-180. 311
Bush, A.O., Lafferty, K.D., Lotz, J.M., Shostak, A.W., 1997. Parasitology meets ecology on 312
its own terms: Margolis et al. revisited. J. Parasitol. 83, 575-583. 313
Centenera, R., 2005. El corzo: acercamiento a una realidad: expansión, caza y gestión. La 314
Trébere, Madrid, 192 pp. 315
Díaz, P., Pedreira, J., Arias, M., Lomba, C., Suárez, J.L., Paz, A., Morrondo, P., 2005. 316
Infecciones parasitarias en vacas de raza Rubia Gallega de la provincia de Lugo: 317
influencia de la edad. Buiat. Esp. 10, 231-234. 318
Díaz, P., Paz-Silva, A., Sánchez-Andrade, R., Suárez, J.L., Pedreira, J., Arias, M., Díez-319
Baños, P., Morrondo, P. (2007). Assessment of climatic and orographic condictions on 320
the infection by Calicophoron daubneyi and Dicrocoelium dendriticum in grazing beef 321
cattle. Vet. Parasitol. 149, 285-289. 322
Díaz, P., Painceira, A., Dacal, V., Vázquez, L., Cienfuegos, S., Pato, F.J., Paz-Silva, A., 323
Panadero, R., Sánchez-Andrade, R., López, C., Díez-Baños, P., Morrondo, P., 2010. 324
Eimeria infections in wild ruminants (Capreolus capreolus) and extensive reared 325
domestic ruminant from Galicia (N.W. Spain). Rev. Ibero-latinoam. Parasitol. 69, 83-326
89. 327
Díez-Baños, N., Cabaret, J., Díez-Baños, P., 1992. Interspecific interactions in naturally 328
acquired nematode communities from sheep abomasum in relation to age of host and 329
season in four areas of León (Spain). Int. J. Parasitol. 22, 327-334. 330

16
Díez-Baños, P., Pedreira, J., Sánchez-Andrade, R., Francisco, I., Suárez, J.L., Diaz, P., 331
Panadero, R., Arias, M.S., Painceira, A., Paz-Silva, A., Morrondo, P., 2008. Field 332
evaluation for anthelmintic resistant ovine gastrointestinal nematodes by in vitro and 333
in vivo assays. J. Parasitol. 94, 925-928. 334
Dróźdź, J., Demiaszkiewicz, A. W., Lachowicz, J., 1992. The helminth fauna of the roe deer 335
Capreolus capreolus (L.) in a hunting area inhabited by red deer, elk and European 336
bison (Borecka Forest, Poland) over the yearly cycle. Acta Parasitol. 37, 83–88. 337
Dróźdź, J., 1995. Polymorphism in the Ostertagiinae Lopez-Neyra, 1947 and comments on 338
the systematics of these nematodes. Syst. Parasitol. 32, 91-99. 339
Dunn, A.M., 1965. The gastro-intestinal helminths of wild ruminants in Britain. I. Roe deer, 340
Capreolus capreolus capreolus. Parasitology 55, 739-745. 341
Durette-Desset, M.C., 1983. Key to the genera of the Superfamily Trichostrongylidea. In: 342
Anderson, R.C., Chabaud, A.G. (Eds.), ClH keys to the nematode parasites of 343
vertebrates. Commonwealth Agricultural Bureaux, Farnham Royal, Bucks, England, 344
pp. 1-86. 345
Durette-Desset, M.C., 1989. Nomenclature proposée pour les espèces décrites dans la sous-346
famille des Ostertagiinae López-Neyra, 1947. Ann. Parasit. Hum. Comp. 56, 297-312. 347
Ferté, H., Clevá, D., Depaquit, J., Gobert, S., Léger, N., 1999. Status and origin of 348
Haemonchinae (Nematoda: Trichostrongylidae) in deer: a survey conducted in France 349
from 1985 to 1998. Parasitol. Res. 86, 582-587. 350
Gibbons, L.M., Khalil, L.F., 1982. A key for the identification of the nematode family 351
Trichostrongylidae, Leiper 1912. J. Helminthol. 56, 185-223. 352

17
Hoberg, E.P., Kocan A.A., Lora G.R., 2001. Gastrointestinal strongyles in wild ruminants. In: 353
Samuel, W.M., Pybus, M.J., Kocan, A.A. (Eds.), Parasitic diseases of wild mammals. 354
Iowa State University Press. Ames, United States, pp. 193-227. 355
Jansen, J., 1975. On the helminth fauna of the moufflon (Ovis aries musimon) compared with 356
those of domestic sheep (Ovis aries dom.) and deer (Capreolus capreolus, Cervus 357
elaphus) in the Netherlands. 3rd
International Wildlife Disease Conference. Viena, 358
Austria, pp. 589-613. 359
Jansen, J. (1992). On the nematode parasite fauna of Friesian roe deer (Capreolus capreolus). 360
In: Hernández, S. (Ed.), “In Memoriam” al Profesor Dr. Francisco de Paula Martínez 361
Gómez. Servicio de Comunicaciones de la Universidad de Córdoba, Córdoba, Spain, 362
pp. 301-307. 363
Klein, S.L., 2000. The effects of hormones on sex differences in infection: from genes to 364
behavior. Neurosci. Biobehav. Rev. 24, 627-638. 365
Kloosterman, A., Ploeger, H.W., Frankena, K., 1991. Age resistance in calves to Ostertagia 366
ostertagi and Cooperia oncophora. Vet. Parasitol. 39, 101-113. 367
Kochko, Y.P., 1997. The principal helminthiases of ruminant ungulates in Belovezhskaya 368
Pushcha. In: Luchkov, A., Tolkach, V., Berwick, S., Brylski, P. (Eds.), Belovezhskaya 369
Pushcha Forest Biodiversity Conservation. Belorusskiĭ dom pechati, Minsk, pp. 224-370
235. 371
Kutzer, E., Sugár, L., Buchacher-Tonitz, S., 1988. Beitrage zur parasitenfauna der 372
wildlebenden wiederkäuer ungarns. II. Aufbauentwicklung des parasitenbefalles bei 373
rehen (Capreolus c. capreolus). Parasit. Hung. 21. 85-97 374

18
Lancaster, M.B., Hong, C., Michel, J.F., 1983. Polymorphism in trichostrongylidae. In: Stone, 375
E.R., Platt, H.L., Khalil, L.F. (Eds.), Concepts in Nematode Systematics. Academic 376
Press, London/New York, pp. 293–302. 377
López, C., Panadero, R., Bravo, A., Paz, A., Sánchez-Andrade,
R., Díez-Baños,
P., Morrondo,
378
P. (2003). Sarcocystis spp. infection in roe deer (Capreolus capreolus) from the 379
North-West of Spain. Z. Jagdwiss. 49, 211-218. 380
Liénard, E., Depaquit, J., Ferté, H., 2006. Spiculopteragia mathevossiani Ruchliadev, 1948 is 381
the minor morph of Spiculopteragia spiculoptera (Gushanskaya, 1931): molecular 382
evidence. Vet. Res. 37, 683–694. 383
Melis, C., Hoem, S.A., Linnell, J.D.C., Andersen, R., 2005. Age-specific reproductive 384
behaviours in male roe deer Capreolus capreolus. Acta Theriol. 50, 445-452. 385
Mezo-Menéndez, M., Díez-Baños, P., Morrondo Pelayo, P., Díez-Baños, N., 1995. Faecal egg 386
output, contamination of pastures and serum pepsinogen concentration in heifers with 387
natural gastrointestinal nematode infections in North-West Spain. J. Helminthol. 69, 388
53-58. 389
Mezo-Menéndez, M., Morrondo, P., Díez-Baños, N., Díez-Baños, P., 1997. Ciclo biológico y 390
epidemiología. In: Rojo, F.A. (Ed.), Bovis. Gastroenteritis parasitarias. Luzán 5, 391
Madrid, Spain, pp. 29-41. 392
Navarrete, I., Reina, D., Habela, M., Nieto, C.G., Serrano, F., Pérez, E., 1990. Parasites of roe 393
deer (Capreolus capreolus) in Cáceres province, Spain. 32 Internationalen 394
Symposiums über die Erkrankungen der Zoo- und Wildtiere. Eskilsuna, Sweden. 395
Nickel, S., Hiepe, T., Ness, H., Pingel, H., 1978. Beiträge zur parasitenfauna der DDR 2. 396
Mitteilung. Untersuchungen zum helminthenvorkommen beim reh (Capreolus 397
capreolus). Angew. Parasitol. 19, 194-202. 398

19
Nogareda, C., Mezo, M., Uriarte, J., Lloveras, J., Cordero Del Campillo, M., 2006. Dynamics 399
of infestation of cattle and pasture by gastrointestinal nematodes in an Atlantic 400
Temperate ambient. J. Vet. Med. 53, 439-444. 401
Ortega, P., 2009. Cosas de corzos: Apuntes de biología y caza en España. Editorial Caïrel, 402
Madrid, Spain, 216 pp. 403
Ortiz, J.M., Goyena, M, Alonso F., 1996. First report of two polymorphic species of 404
Ostertagia (Nematoda:Trichostrongyloidea) in Cervus elaphus in Spain: O. 405
leptospicularis and O. kolchida. Res. Rev. Parasitol. 56, 221-223 406
Panadero, R., Carrillo, E. B., López, C., Díez-Baños, N., Díez-Baños, P., Morrondo, M.P., 407
2001. Bronchopulmonary helminths of roe deer (Capreolus capreolus) in the 408
northwest of Spain. Vet. Parasitol. 99, 221-229. 409
Panadero, R., Painceira, A., López, C., Vázquez, L., Paz, A., Diaz, P., Dacal, V., Cienfuegos, 410
S.; Fernández, G.; Lago, N.; Díez-Baños, P.; Morrondo, P. (2010). Seroprevalence of 411
Toxoplasma gondii and Neospora caninum in wild and domestic ruminants sharing 412
pastures in Galicia (Northwest Spain). Res. Vet. Sci. 88, 111-115. 413
Pedreira, J., Paz-Silva, A., Sánchez-Andrade, R., Suárez, J.L., Arias, M., Lomba, C., Díaz, P., 414
López, C., Díez-Baños, P., Morrondo, P., 2006. Prevalences of gastrointestinal 415
parasites in sheep and parasite-control practices in NW Spain. Prev. Vet. Med. 75, 56-416
62. 417
Pilarczyk, B., Balicka-Ramisz, A., Ramisz, A., Lachowska, S., 2005. The occurrence of 418
intestinal parasites of roe deer and red deer in the Western Pomerania volvodeship. 419
Wiad. Parazytol. 51, 397-310. 420

20
Ploeger, H.W., Kloosterman, A., Rietveld, F.W., Berghen, P., Hilderson, H., Hollanders W., 421
1994. Quantitative estimation of the level of exposure to gastrointestinal nematode 422
infection in first-year calves. Vet. Parasitol. 55, 287-315. 423
Poulin, R., 1996. Helminth growth in vertebrate hosts: does host sex matter?. Int J Parasitol. 424
26, 1311-1315. 425
Ramajo, V., Pérez, R., Ramajo, A., Oleaga, A., 2007. Preliminary data about the parasitism 426
caused by Protozoa, Helminths and Ticks in cervids and wild bovids from Salamanca 427
(Western Spain). Res. Rev. Parasitol. 67, 69-77. 428
Rehbein, S., Lutz, W., Visser, M., Winter, R., 2000. Investigation of the parasite fauna of 429
wildlife in Northrhine-Westfalia. 1. Endoparasites of roe deer. Z. Jagdwiss. 46, 248-430
269. 431
Reina, D., Habela, M., Serrano, F., Nieto, C.G., Breña, M., Pérez, E., Navarrete, I., 1992. 432
Contribución al conocimiento de la parasitofauna de los animales silvestres y de vida 433
libre en la provincia de Cáceres (España). In: Hernández, S. (Ed.), “In Memoriam” al 434
Profesor Dr. Francisco de Paula Martínez Gómez. Servicio de Comunicaciones de la 435
Universidad de Córdoba, Córdoba, Spain, pp. 407-428. 436
Rickard, L.G., Hoberg, E.P., Allen, N.M., Zimmerman, G.L., Craig, T.M., 1993. 437
Spiculopteragia spiculoptera and S. asymmetrica (Nematoda: Trichostrongyloidea) 438
from red deer (Cervus elaphus) in Texas. J. Wildl. Dis. 29:512-5. 439
Rossi, L., Eckel, B., Ferroglio, E., 1997. A survey of the gastro-intestinal nematodes of roe 440
deer (Capreolus capreolus) in a mountain habitat. Parassitologia 39, 303-312. 441
Segonds-Pichon, A., Ferté, H., Gaillard, J.M., Lamarque, F., Duncan, P. 2000. Nematode 442
infestation and body condition in roe deer (Capreolus capreolus). Game Wildl. Sci. 443
17, 241-258. 444

21
Silvestre, A., Chartier, C., Sauvé, C., Cabaret, J., 2000. Relationship between helminth 445
species diversity, intensity of infection and breeding management in dairy goats. Vet. 446
Parasitol. 94:91-105. 447
Skrjabin, K.I., Shikhobalova, N.P., Schulz, R.S., 1954. Trichostrongylids of animals and man. 448
In: Skrjabin, K.I. (Ed.), Essentials of Nematodology, vol. 3. Israel program for 449
scientific translations, Jerusalem, 704 pp. 450
Stevenson, L.A., Gasser, R.B. & Chilton, N.B., 1996. The ITS-2 rDNA of Teladorsagia 451
circumcincta, T. trifurcata and T. davtiani (Nematoda: Trichostrongylidae) indicates 452
that these taxa are one species. Int. J. Parasitol. 26, 1123-1126. 453
Suarez, V.H., Cabaret, J., 1992. Interbreeding in the subfamily Ostertagiinae (Nematoda: 454
Trichostrongylidae) of ruminants. J. Parasitol. 78, 402-405. 455
Taylor, M.A., Coop, R.L., Wall, R.L. (Eds.), 2007. Veterinary Parasitology. Ed. Blackwell 456
Publishing, Oxford. 600 pp. 457
Togni, T., Manfredi, M.T., Di Cerbo, A.R., Zanzani, S., Gioppo, S., Piccolo, G., Bregoli, M., 458
Trevisiol, K., 2004. Abomasal nematodes community in Cervidae (Cervus elaphus and 459
Capreolus capreolus) from the Trentino Alto Adige (North Italy). Parassitologia 46, 460
71. 461
Uriarte, J., Llorente, M.M., Valderrábano, J., 2003. Seasonal changes of gastrointestinal 462
nematode burden in sheep under an intensive grazing system. Vet. Parasitol. 118, 79-463
92. 464
Vázquez, L., Dacal, V., Pato, F.J., Paz-Silva, A., Diez-Baños, N., López, C., Panadero, R., 465
Sánchez-Andrade, R., Diez-Baños, P., Morrondo, P., 2009. The occurrence of 466
endoparasites of roe deer (Capreolus capreolus) in two different areas from NW 467
Spain. Rev. Ibero-latinoam. Parasitol. 68, 25-33. 468

22
Vázquez, L., Panadero, R., Dacal, V., Pato, F.J., López, C., Díaz, P., Arias, M.S., Fernández, 469
G., Diez-Baños, P., Morrondo, P., 2011. Tick infestation (Acari: Ixodidae) in roe deer 470
(Capreolus capreolus) from northwestern Spain: population dynamics and risk 471
stratification. Exp. Appl. Acarol. 53, 399-409. 472
Vetýška, V., 1980. Endoparasites of roe deer in the Strakonice Region. Acta Vet. Brno. 49, 473
91-103. 474
Yamaguti, S., 1961. Systema Helminthum. Vol. III The nematods of vertebrates part I- II. 475
Interscience Publishers Inc, New York. 1135 pp. 476
Zaffaroni, E., Citterio, C., Sala, M., Lauzi, S., 1997. Impact of abomasal nematodes on roe 477
deer and chamois body condiction in an alpine environment. Parassitologia 39, 313-478
317. 479
Zaffaroni, E., Manfredi M.T., Citterio, C., Sala, M., Piccolo, G., Lanfranchi, P., 2000. Host 480
specificity of abomasal nematodes in free ranging alpine ruminants. Vet. Parasitol. 90, 481
221-30. 482
483

23
Figure caption 484
485
Figure 1. Percentage of plurispecific and monospecific gastrointestinal nematode infections in 486
roe deer from NW Spain. a) abomasum b) small intestine c) large intestine d) cecum. 487
488
489
TOTAL Abomasum Small intestine Large intestine Cecum
Gastrointestinal nematode species % x ±SD % x ±SD
% x ±SD % x ±SD
% x ±SD
Chabertia ovina 2.3 2±1 2.3 1±1
Cooperia oncophora 1.0 9±7 1.0 9±7
Cooperia pectinata 0.5 31±31 0.5 31±31
Cooperia punctata 1.0 12±9 1.0 12±9
Haemonchus contortus 1.4 6±3 1.4 6±3
Nematodirus filicollis 65.9 199±871 2.8 200±435 65.4 191±871
Nematodirus spathiger 2.4 8±4 2.4 8±4
Oesophagostomum venulosum 50.5 4±4 35.8 2±2 36.0 3±3
Ostertagia leptospicularis 99.1 391±417 98.6 383±406 18.3 52±194
O. leptospicularis (minor morph: kolchida) 89.5 57±88 88.2 59±89 4.8 10±5
Spiculopteragia spiculoptera 96.2 485±357 96.2 483±349 21.6 33±64
S. spiculoptera (minor morph: mathevossiani) 57.5 29±49 55.9 25±32 1.9 11±7
Teladorsagia circumcincta 6.6 28±30 6.6 28±30
T. circumcincta (minor morph: trifurcata) 0.5 21±0 0.5 21±0
Trichostrongylus axei 29.6 118±217 25.6 119±219 3.4 22±16
Trichostrongylus capricola 3.4 67±117 1.4 28±40 1.9 97±154
Trichostrongylus colubriformis 3.0 30±22 0.9 13±10 2.4 26±26
Trichostrongylus vitrinus 4.9 182±369 2.4 350±474 3.4 11±7
Trichuris capreoli 53.1 2±2 40.0 2±1 28.0 2±1
Trichuris ovis 10.4 2±2 2.8 1±0 7.6 2±2
Table 1. Prevalence and intensity of infection by different gastrointestinal nematode species and its localization in the digestive system.
Table

1
GENERA OVERALL
PREVALENCE
AND INTENSITY
CLIMATIC AREA SEX AGE
COAST CENTER MOUNTAIN MALES FEMALES YOUNG ADULTS
Chabertia 2.3
(2±1)
2.3
(2±0)
3.2
(2±1)
1.4
(1±0)
2.2
(2±1)
3.4
(1±0)
3.7
(1±0)
2.2
(2±1)
Cooperia 1.9
(18±21)
2.3
(6±0)
3.2
(23±24)
0.0
-
1.7
(8±6)
3.3
(49±0)
3.6
(4±0)
1.6
(23±23)
Haemonchus 1.4
(1±0)
2.3
(1±0)
2.1
(1±0)
0.0
-
1.6
(1±0)
0.0
-
3.3
(1±0)
1.1
(1±0)
Nematodirus 66.2
(199±871)
60.0
(198±368)
58.6
(107±118)
70.3
(301±1,384)
65.8
(123±192)
45.2
(863±2,657)
56.7
(675±2,408)
63.8
(131±223)
Oesophagostomum 50.5
(6±5)
62.2
(7±5)
38.4
(6±7)
55.4
(5±4)
52.4
(6±5)
29.6
(2±0)
46.7
(4±2)
49.5
(6±5)
Ostertagia 99.1
(443±489)
100.0
(539±583)
96.0
(397±529)
89.2
(444±350)
96.3
(458±509)
93.5
(350±339)
100
(364±247)
95.2
(456±518)
Spiculopteragia 96.7
(497±367)
97.8
(507±473)
94.9
(462±318)
95.9
(542±350)
95.7
(513±368)
87.1
(391±348)
93.3
(483±326)
94.7
(500±374)
Teladorsagia 6.6
(29±33)
2.3
(8±0)
8.4
(12±10)
6.9
(62±37)
7.7
(29±33)
0.0
-
13.3
(12±12)
5.5
(36±37)
Trichostrongylus 34.5
(129±252)
44.4
(158±288)
29.3
(104±166)
28.4
(134±318)
34.2
(130±257)
19.4
(109±214)
26.7
(119±184)
33.0
(130±261)
Trichuris 55.9
(3±3)
68.9
(3±4)
46.5
(2±2)
55.4
(2±2)
59.4
(3±3)
22.6
(2±1)
46.7
(4±2)
55.3
(2±3)
2
Table 2. Prevalence (%) and intensity ( x ±SD; in brackets) by different gastrointestinal nematode genera when considering the climatic 3
area, sex and age of the animals. 4
5
Table
1
1
2
Figure




















