
Full issue for TGLE Vol. 54 Nos. 1 & 2
77
The Great Lakes Entomologist The Great Lakes Entomologist Volume 54 Numbers 1 & 2 - Spring/Summer 2021 Numbers 1 & 2 - Spring/Summer 2021 Article 1 Full issue for TGLE Vol. 54 Nos. 1 & 2 Full issue for TGLE Vol. 54 Nos. 1 & 2 Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Recommended Citation . "Full issue for TGLE Vol. 54 Nos. 1 & 2," The Great Lakes Entomologist, vol 54 (1) Available at: https://scholar.valpo.edu/tgle/vol54/iss1/1 This Full Issue is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected].
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Full issue for TGLE Vol. 54 Nos. 1 & 2

The Great Lakes Entomologist The Great Lakes Entomologist
Volume 54 Numbers 1 & 2 - Spring/Summer 2021 Numbers 1 & 2 - Spring/Summer 2021
Article 1
Full issue for TGLE Vol. 54 Nos. 1 & 2 Full issue for TGLE Vol. 54 Nos. 1 & 2
Follow this and additional works at: https://scholar.valpo.edu/tgle
Part of the Entomology Commons
Recommended Citation Recommended Citation . "Full issue for TGLE Vol. 54 Nos. 1 & 2," The Great Lakes Entomologist, vol 54 (1) Available at: https://scholar.valpo.edu/tgle/vol54/iss1/1
This Full Issue is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected].

Vol. 54, Nos. 1 & 2 Spring/Summer 2021
THEGREAT LAKES
ENTOMOLOGIST
PUBLISHED BY
THE MICHIGAN ENTOMOLOGICAL
SOCIETY
1
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

THE MICHIGAN ENTOMOLOGICAL SOCIETY 2020–21
OFFICERSPresident Duke ElsnerPresident Elect Mark VanderWerpImmediate Pate President David HoughtonSecretary Adrienne O’Brien Treasurer Angie Pytel Member-at-Large Thomas E. Moore Member-at-Large Brian Scholtens Member-at-Large Crystal DaileyMember-at-Large Ralph GortonLead Journal Scientific Editor Kristi Bugajski Lead Journal Production Editor Alicia Bray Associate Journal Editor Anthony Cognato Associate Journal Editor Julie Craves Associate Journal Editor David HoughtonAssociate Journal Editor Ronald Priest Associate Journal Editor William Ruesink Associate Journal Editor William Scharf Associate Journal Editor Daniel Swanson Newsletter Editors Crystal Dailey and Duke ElsnerWebmaster Mark O’Brien
The Michigan Entomological Society traces its origins to the old Detroit Entomological Society and was organized on 4 November 1954 to “. . .promote the science of entomology in all its branches and by all feasible means, and to advance cooperation and good fellowship among persons interested in entomology.” The Society attempts to facilitate the exchange of ideas and information in both amateur and professional circles, and encourages the study of insects by youth. Membership in the Society, which serves the North Central States and adjacent Canada, is open to all persons interested in entomology.
There are five paying classes of membership:Active—annual dues $25.00
Student (to 12th grade)—annual dues $12.00 Institutional—annual dues $45.00
Sustaining—annual contribution $35.00 or more Life—$500.00
Dues are paid on a calendar year basis (Jan. 1–Dec. 31). Memberships accepted before July 1 shall begin on the preceding January 1; memberships accepted at a later date shall begin the following January 1 unless the earlier date is requested and the required dues are paid. All members in good standing receive the Newsletter of the Society. All active and sustaining members may vote in Society affairs. All dues and contributions to the Society are deductible for Federal income tax purposes.
SUBSCRIPTION INFORMATION The journal is published online and open access, there is no subscription needed. Articles are available in pdf format and can be printed from the website, free of charge. To view current and past volumes, please visit http://scholar.valpo.edu/tgle/ .
2
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 1
Influence of Age on Decision Making by Ovipositing Pieris rapae (Lepidoptera: Pieridae)
Brett Breitkreutz, Tracy L.E. Wagner, and Rodrigo J. Mercader*
Department of Biology, 1700 SW College Ave., Washburn University, Topeka, KS 66621*Corresponding author: (e-mail: [email protected]; phone: (785) 670-2091)
AbstractDue to its effect on the time available for host selection and learning, butterfly age is
expected to alter the degree of host specificity and potentially niche breadth. Here, we use the small cabbage white, Pieris rapae (L.) (Lepidoptera: Pieridae), to test the effect of age on ovipositional specificity and decision-making time. Specifically, we examined the ovipo-sitional behavior of P. rapae 4, 8, and 12 days post-emergence. Females were recorded in thirty-minute trials using leaves of two hosts, mustard leaves, Brassica juncea (L.) Czern., and collard greens, Brassica oleracea L. Acephala group, and the non-host common bean, Phaseolus vulgaris L.. Subsequently, we measured the duration of drumming events (a proxy for decision-making time) and whether the leaves were accepted or rejected as ovipositional substrates. As would be expected if prior experience influenced ovipositional behavior, we saw a reduction in the duration of drumming events as females aged. In particular, we saw a reduction in duration of drumming events when rejecting the non-host between days 4 and days 8 and 12. We also detected a decrease in drumming time between days 4 and 8 when accepting hosts, but an increase in drumming time between days 8 and 12 when accepting hosts. These results suggest both an increased ability to recognize hosts and an increase in selectivity with age.
Keywords: host selection, egg limitation, host specificity, oviposition behavior
Adult ovipositional behavior can determine the larval environment and con-sequently the realized host range of many plant-feeding insect species. Therefore, understanding the factors that affect oviposi-tional behavior can help us understand how host breadth may evolve. Over half a century ago Levins and Macarthur (1969) proposed that the costs imposed by decision-making on foraging herbivorous insects were a ma-jor component determining host breadth in plant feeding insects. Bernays and Wcislo (1994) formalized this concept into a “neural hypothesis”, which proposes that specializa-tion (e.g., narrower host range) may arise from limitations in the ability to gather and process information. In particular, foraging herbivorous insects with fewer number of choices (competing stimuli) require a lower attentional load, leading to decreased error rate and decision-making time (e.g., Bernays 2001, Egan and Funk 2006, Tosh et al. 2009).
For many butterfly species, oviposi-tional behavior of adult females determines larval resource use and therefore represents a foraging decision expected to experience high selective pressure. Host selection by ovipositing females under the “hierarchical threshold model” (Courtney et al. 1989) is
believed to be determined by genetically fixed preference hierarchies (Miller and Strickler 1984, Courtney et al. 1989, Mer-cader and Scriber 2007). Therefore, within the “hierarchical threshold model” each host plant is expected to have an intrinsic acceptability threshold determined by positive and negative stimuli and plants low in the acceptability threshold will only be accepted when the insect’s state has changed. Consequently, while the relative preference ranking between hosts is fixed, how tightly host preferences are adhered to (i.e. specificity) is believed to be plastic and may be modified by intrinsic factors such as physiological state, adult experience, age, and fecundity (Miller and Strickler 1984, Bossart and Scriber 1999, Mercader and Scriber 2005, Gamberale-Stille et al. 2019). These non-genetic intrinsic factors act as a form of phenotypic flexibility that can influ-ence the host breadth of species (Miller and Strickler 1984, Mercader and Scriber 2005, Fordyce 2006) and lead to within species variability.
Age has long been considered an im-portant factor affecting host selection (Gos-sard and Jones 1977) due to its effects on in-sect physiological state (Miller and Strickler
3
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

2 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
1984), time available for host selection (West and Cunningham 2002; Rosenheim 1999 a,b), and experience (e.g., Papaj and Prokopy 1989, Snell-Rood and Papaj 2009, Jones and Agrawal 2019). Models suggest that as ovi-positing insects age, a decrease in specificity is expected due to a heightened risk of not finding any suitable hosts, even towards the end of a lifetime for organisms initially limited by available eggs (Rosenheim, 1999 a,b). However, organisms that consume lim-ited resources as adults may experience egg limitations later in life, ultimately leading to increased specificity (Rosenheim 1999 a,b; Rosenheim et al. 2008; Rosenheim 2011). In addition, as adult females age, they also have the potential to become more efficient forag-ers due to prior experience (e.g. Papaj and Prokopy 1989, Snell-Rood and Papaj 2009, Jones and Agrawal 2019), thereby also in-creasing specificity. Therefore, as ovipositing adults age, specificity may decrease due to time limitation or increase due to experience and/or egg limitations.
Here we take advantage of the small cabbage white, Pieris rapae (L.) (Lepidop-tera: Pieridae), a specialist on members of the Brassicaceae family that has long been the subject of studies on ovipositional behavior (e.g., Richards 1940, Gossard and Jones 1977, Renwick and Radke 1988, Lund et al. 2019), to investigate the effect of aging on decision-making by ovipositing females. Specifically, we test a) the effect of age on ovipositional specificity and b) if ovipositing females exhibit reduced decision-making time with age/experience. Upon alighting on a plant, females of many butterfly species searching for oviposition sites will engage in a behavior known as “drumming”, which consists of rapidly moving their forelegs across the surface of the leaf (Schoonhoven et al. 2005). This behavior allows females to evaluate the ovipositional substrate, by exposing contact chemoreceptors in the tarsi to stimulant and deterrent compounds on the leaf surface. This behavior is well described (Schoonhoven et al. 2005) for P. rapae and provides an identifiable behavior linked to host plant evaluation. In addition, P. rapae are known to exhibit adult learning (Snell-Rood and Papaj 2009) and experience reduced egg laying with age (Gossard and Jones 1977), making them ideal candidates for this study.
Materials and Methods
Study organism: Pieris rapae in-dividuals were purchased as eggs from Carolina Biological Supply Co. and reared on green cabbage leaves Brassica oleracea L. var. capitata. Larvae were maintained at room temperature under an 18:6 L:D
photoperiod, allowed to pupate, transferred to individual emergence cages, and females were numbered on the front wings prior to introduction to a common cage along with males. Butterflies were provided an artificial nectar solution of 15 % honey-water solution and a lepidopteran Ringer’s solution (follow-ing Lederhouse et al. 1990) to ensure male fertility. Cabbage leaves were introduced daily for a one-hour period between 10 AM and 2 PM to allow females to oviposit. Fe-males that were a part of the ovipositional assays were removed from the common cage on days in which they were assayed as they were presented ovipositional substrates during the assays.
Three-choice oviposition assays: Three choice assay using leaves of two hosts, mustard leaves, Brassica juncea (L.) Czern., and collard greens, Brassica oleracea Aceph-ala group, and the non-host common bean, Phaseolus vulgaris L. were performed and filmed on 17 butterflies at 4, 8, and 12 days post emergence. B. juncea and B. oleracea were chosen as two hosts based on experience from prior assays using B. juncea and B. ol-eracea with P. rapae females from the same source. In particular, previous assays had indicated a preference for B. juncea under similar conditions, but not an overwhelming preference for B. juncea (B. juncea preferred in most, but not all assays). For each assay, a single fresh leaf of each of the three plants was placed individually over small glass cylinder (6 cm diameter and 7 cm height) and secured with Parafilm® exposing only the upper circular surface area for each assay; ensuring identical leaf surface area was exposed for each leaf and each assay. Cylinders with leaves were placed on the far side of a 60 × 30 × 30 cm (L × W × H) glass aquarium in a row with the sides and back walls covered in black cardboard. The order of each leaf in the row (i.e., left, middle, or right) was randomized between assays. A video recorder was placed on the front wall of the aquarium, and a light placed above the leaves. Prior to each assay butterflies were cooled for 2 minutes at 6-8 °C, placed in the enclosure, and allowed to heat up and then filmed for 30 minutes.
No-choice oviposition assays: To determine whether age decreased rejection time when non-hosts were encountered, we first conducted no-choice assays consisting of a leaf of the non-host P. vulgaris. Assays were performed and filmed prior to every three-choice trial on the same females as the three-choice trial. The setup was identical to the three-choice trial, with the exception that only a P. vulgaris leaf was available.
Behavior Coding: Films were viewed and the duration of each drumming event
4
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 3
and whether it led to egg laying or not were coded for 11 butterflies that laid eggs during at least two of the three choice trials using Solomon Coder (Péter 2011). Eleven but-terflies laid eggs during days 4 and 8, and 9 butterflies laid eggs during all three trial days. In total 1494 drumming events were recorded and 872 eggs were laid by these 11 butterflies during the trials.
Statistical analysis: All analyses conducted using R (R Core Team, 2019).
Host specificity: In our assays, very few eggs were laid on P. vulgaris, and we were solely interested in shifts in the proportion of eggs laid between the two hosts. Therefore, as the proportion of eggs laid per day for each butterfly are correlated all analyses were run on the proportion of eggs laid per trial on the preferred host, B. juncea. We used General Linear Mixed Effect Models (GLMM) with proportion of eggs laid on B. juncea as the response variable, individual butterfly as a random effect, and age as a fixed effect using the lme4 package (Bates et al. 2015). Proportion of eggs laid on B. juncea were arcsine transformed to meet distribution assumptions. Statistical significance was de-termined using likelihood ratio tests (LRT).
Decision making time: The effect of age/experience on decision-making time (duration of drumming) was tested in two main formats. We first tested effect of
host and age on the duration of drumming events as a GLMM with host, age, and their interaction as fixed effects and individual butterfly as a random effect. Subsequently, we analyzed the effect of age on duration of drumming events on the preferred host and non-host separately as GLMM’s with age as a fixed effect and butterfly ID as a random effect. Finally, we contrasted the duration of drumming events when the two host plants (B. juncea and B. olereacea) were rejected or accepted as GLMM’s with choice (rejected or accepted) and age as a fixed effect and butterfly ID as a random effect. Duration of drumming events were square root trans-formed to meet distribution assumptions and significant effects interactions were tested using likelihood ratio tests (LRT) and pairwise comparisons using Tukey’s HSD as implemented in the multcomp package (Hothorn et al. 2008).
Results
Three-choice oviposition assays: Host preference: The proportion of eggs laid on the preferred host B. juncea remained consistently higher throughout the trials and no significant effect of age was observed (LRT: χ2 = 2.554; df = 2; P = 0.279; Fig. 1).
Decision making time: We observed a significant reduction in the duration of drumming events with age overall (LRT:
Figure 1. Boxplots of the proportion of eggs laid on common bean, P. vulgaris, collard greens, B. oleracea Acephala group, and mustard greens, B. juncea by P. rapae females 4, 8, and 12 days post emergence. Boxes represent interquartile ranges, heavy lines medians, whiskers minimum and max-imum non-extreme outliers, and circles extreme outliers.
5
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

4 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
χ2 = 39.488; df = 2; P < 0.001; Fig. 2). More specifically, we observed an initial reduction in the duration of drumming events leading to hosts being accepted with age (LRT: χ2 = 39.464; df = 2; P < 0.001) and an overall re-duction in the duration of drumming events leading to non-host being rejected with age (LRT: χ2 = 17.989; df = 2; P < 0.001). How-ever, there was no change in the duration of drumming events with age for preferred hosts when rejected (LRT: χ2 = 1.024; df = 2; P = 0.599) and insufficient observations of acceptance of the non-host to analyze. Within drumming events leading to the acceptance
Figure 2. Mean duration of drumming events (seconds) by P. rapae females 4, 8, and 12 days post emergence a) when accepting hosts (B. ol-eracea Acephala group, and B. juncea), b) when rejecting hosts, or c) when rejecting the non-host (P. vulgaris). Means followed by the same letter are not significantly different at the P < 0.05 level (Tukey’s HSD).
of hosts, we saw an initial reduction in drum-ming duration for accepted hosts between 4-day old and 8-day old butterflies, followed by an increase between 8-day old and 12-day old butterflies (Fig. 2a). Within non-hosts we saw a reduction in drumming time in both 8- and 12-day old butterflies compared to 4-day old butterflies (Fig. 2c).
In addition, we observed an overall greater duration of drumming events when hosts were rejected than accepted (LRT: χ2 = 94.45; df = 1; P < 0.001). Suggesting, that within these assays when females were being more selective and rejecting a host plant, decision making times were increased.
No-choice oviposition assays: Very few drumming events took place in no-choice assays ( < 20 compared ~1500 in three choice assays) and none of the butterflies performed drumming behaviors during all three no-choice assay. In contrast, all butterflies that laid eggs during the three choice assays (described below) also drummed on the non-host P. vulgaris. These results suggest that P. rapae females foraging for ovipositional substrates are unlikely to meaningfully in-teract with non-host plants in the absence of neighboring host plants.
Discussion
Although an overall effect of age on ovipositional preference amongst hosts was not observed, we observed a distinct effect on the duration of drumming events. Tarsal drumming behavior is exceedingly common in butterflies and strongly linked with con-tact chemoreception (Renwick and Chew 1994), a key component of host plant recogni-tion and acceptance/rejection of hosts. Here we observed both an initial reduction in the duration of drumming events, as expected if prior experience influenced ovipositional behavior, and subsequently an increase in the duration of drumming events when ac-cepting hosts, as would be expected if female selectivity were to increase with age (Fig. 2).
Pieris rapae females exhibited an overall reduction in the duration of drum-ming events with age, indicating faster de-cision-making time with age/experience. In particular, there was a marked reduction in duration of drumming events when accepting hosts or rejecting non-hosts between days 4 and 8 (Fig. 2), suggesting an increased ability to recognize hosts from non-hosts. However, between days 8 and 12 we saw an increase in the duration of drumming events when hosts were accepted, but not when rejecting hosts or non-hosts (Fig. 2). This suggests that older females were capable of faster decision-making, yet decision-making time was longer when accepting hosts in
6
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 5
the final trial. Tradeoffs between speed and accuracy in foraging herbivores have been well-established (Bernays 2001) and a bias towards accuracy would be expected if an egg limitation were present.
It is worth noting that during no-choice assays using the non-host P. vulgaris very few drumming events occurred throughout all assays. This suggests that behaviors leading to oviposition are rare in the absence of suitable hosts. However, host searching in complex environments containing multiple non-hosts is common and increased neural capacity has been previously observed in P. rapae foraging in complex compared to sim-ple environments (Snell-Rood et al. 2009). Here we observed within P. rapae females primarily exposed to a simple environment (a single host present in non-testing arenas) a decrease in decision-making time when rejecting or accepting hosts in a previously experienced environment. However, we also found that as females age an increase in decision-making time could become present when accepting hosts. Although the design of this study does not allow us to separate the effects of experience and age, the observed patterns suggest that as females age, for-aging efficiency may be modulated by both experience and aging.
Acknowledgments
We would like to thank Tyler Brandon, Deysi de la Rosa, and Malcom S. Mikkelsen for help with data collection. We would also like to thank Benjamin Reed and Joshua Smith for comments on the manuscript. Funding for this project was provided by the Washburn University Biology Department.
Literature Cited
Bates, D., M. Maechler, B. Bolker, and S. Walker. 2015. lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1–7, http://CRAN.R-project.org/package=lme4.
Bernays, E.A., and W.T. Wcislo. 1994. Sensory capabilities, information processing, and re-source specialization. The Quarterly Review of Biology 69:187–204.
Bernays, E.A. 2001. Neural limitations in phytophagous insects: implications for diet breadth and evolution of host affiliation. Annual Review of Entomology 46: 703–727.
Bossart, J.L., J.M. Scriber. 1999. Preference variation in the polyphagous tiger swallow-tail butterfly (Lepidoptera: Papilionidae). Environmental Entomology 28: 628–637.
Courtney, S.P., G.K. Chen, and A. Gardner. 1989. A general-model for individual host selection. Oikos 55: 55–65.
Egan, S.P., and D.J. Funk. 2006. Individual advantages to ecological special-ization: insights on cognitive constraints from three conspecific taxa. Proceedings of the Royal Society B: Biological Sciences 273:843–848.
Fordyce, J.A. 2006. The evolutionary conse-quences of ecological interactions mediated through phenotypic plasticity. Journal of Experimental Biology 12: 2377–2383.
Gamberale-Stille, G., A. Schäpers, N. Janz, and S. Nylin. 2019. Selective attention by priming in host search behavior of 2 gen-eralist butterflies. Behavioral Ecology 30: 142–149.
Gossard, T.W., and R.E. Jones. 1977. The effects of age and weather on egg-laying in Pieris rapae L. Journal of Applied Ecology 14: 65–71.
Hothorn, T., F Bretz, P. Westfall, and R.M. Heiberger. 2008. Multcomp: Simultaneous Infer-ence in General Parametric Models—R Package Version 1.0–0. R Foundation for Statistical Computing. Vienna, Austria.
Jones, P.L., and A.A. Agrawal. 2019. Beyond preference and performance: host plant selection by monarch butterflies, Danaus plexippus. Oikos 128: 1092–1102.
Lederhouse, R.C., M.P. Ayres, J.M. Scriber. 1990. Adult nutrition affects male virility in Papilio glaucus L. Functional Ecology 1:743–751.
Levins, R., and R. MacArthur. 1969. An hypoth-esis to explain the incidence of monophagy. Ecology 50: 910–911.
Lund, M., D.C. Brainard, and Z. Szendrei. 2019. Cue hierarchy for host plant selection in Pieris rapae. Entomologia Experimentalis et Applicata 167: 330–340.
Mercader, R.J., and J.M. Scriber. 2005. Pheno-typic plasticity of host selection in adult tiger swallowtails; Papilio glaucus (L.). Pp 25–57. In Ananthakrishnan, T.N., and D. Whitman (eds.) Insects phenotypic plasticity: Diversity of responses. Science Publishers, Enfield, New Hampshire, USA.
Mercader, R.J., and J.M. Scriber. 2007. Di-versification of host use in two polyphagous butterflies: differences in oviposition speci-ficity or host rank hierarchy? Entomologia Experimentalis et Applicata 125: 89–101.
Miller, J.R., and K.L. Strickler. 1984. Finding and accepting host plants. Pp 127–158. In J.W. Bell and R.T. Carde [eds.] Chemical Ecology of Insects. Chapman and Hall, NY.
Papaj, D.R., and R.J. Prokopy. 1989. Ecolog-ical and evolutionary aspects of learning in
7
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

6 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
phytophagous insects. Annual Review of Entomology 34: 315–350.
Péter, A. 2011. Solomon Coder (version beta 11.01.22): a simple solution for behavior coding. http://solomoncoder.com/
R Core Team. 2019. R: a language and environ-ment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http.//www-R-project.org
Renwick, J.A.A., and C.D. Radke. 1988. Sen-sory cues in host selection for oviposition by the cabbage butterfly, Pieris rapae. Journal of Insect Physiology 34: 251–257.
Renwick, J. A. A., and F. Chew. 1994 Oviposi-tion behavior in Lepidoptera. Annual Review of Entomology 39: 377- 400.
Richards, O.W. 1940. The biology of the small white butterfly (Pieris rapae), with special reference to the factors controlling its abun-dance. Journal of Animal Ecology 9: 243–288.
Rosenheim, J.A. 1999a. The relative contribu-tions of time and eggs to the cost of reproduc-tion. Evolution 53: 376–385.
Rosenheim, J.A. 1999b. Characterizing the cost of oviposition in insects: a dynamic model. Evolutionary Ecology 13: 141–165.
Rosenheim, J.A. 2011. Stochasticity in reproduc-tive opportunity and the evolution of egg lim-itation in insects. Evolution 65: 2300–2312.
Rosenheim, J.A., S.J. Jepsen, C.E. Matthews, D.S. Smith, and M.R. Rosenheim. 2008. Time limitation, egg limitation, the cost of oviposition, and lifetime reproduction by an insect in nature The American Naturalist 172: 486–496.
Schoonhoven, L.M., B. Van Loon, J.J.A. van Loon, and M. Dicke. 2005. Insect-plant biology. Oxford University Press, Oxford.
Snell-Rood, E.C., D.R. Papaj, and W. Gronen-berg. 2009. Brain size: a global or induced cost of learning? Brain, Behavior and Evolu-tion 73: 111–128.
Snell-Rood, E.C., and D.R. Papaj. 2009. Pat-terns of phenotypic plasticity in common and rare environments: a study of host use and color learning in the cabbage white butterfly Pieris rapae. The American Naturalist 173: 615–631.
Tosh, C.R., J. Krause, and G.D. Ruxton. 2009. Theoretical predictions strongly support deci-sion accuracy as a major driver of ecological specialization. Proceedings of the National Academy of Sciences 106: 5698–5702.
West, S.A., and J. P. Cunningham. 2002. A general model for host plant selection in phytophagous insects. Journal of Theoretical Biology 214: 499–513
8
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 7
Susceptibility, Preference, and Suitability of Carpinus and Ostrya Taxa for Gypsy Moth Larvae (Lepidoptera: Lymantriidae)
Fredric Miller* and Susan WiegrefeThe Morton Arboretum, 4100 Illinois Route 53, Lisle, IL 60532
* Corresponding author: (e-mail: [email protected]; 630-719-2427)
AbstractTwenty Carpinus taxa and three Ostrya taxa, growing in the tree breeding produc-
tion area at The Morton Arboretum, Lisle, IL, were evaluated in laboratory bioassays for feeding susceptibility, preference, and suitability for gypsy moth (Lymantria dispar (L.)) (Lepidoptera: Lymantriidae) larvae. No-choice and multiple-choice laboratory feeding studies revealed that C. coreana, C. fargesii, C. laxiflora, and the hybrid C. caroliniana × C. orientalis were the least suitable for larval development and pupation, and were less preferred by gypsy moth larvae. Suitability rankings for gypsy moth larval development time were highly correlated with larval longevity, but the proportion of larvae pupating was not correlated with either larval longevity or with larval development time. Pupal fresh weights were not correlated with either larval longevity or with larval development time. However, larval dry frass weights were correlated with the proportion of larvae pupating, but not correlated with pupal fresh weights. Leaf toughness and leaf thickness do not appear to be factors in gypsy moth larval suitability and preference.
Keywords: Susceptibility, preference suitability, gypsy moth, Lymantria dispar, Carpinus, Ostrya
Since its introduction into the United States in the mid-19th century, the gypsy moth Lymantria dispar (L.) (Lepidoptera: Lymantriidae) is still considered one of the most destructive and persistent pest of nurs-ery crops, landscape plants, and rural and urban forest trees. Gypsy moth larvae feed on over 500 species of woody plant (Forbush and Fernald 1896, Liebhold et al. 1995) preferring oaks (Quercus spp.) (Barbosa and Krischik 1987), and other hardwood species (Elkinton and Liebhold 1990, Shields et al. 2003). Chronic defoliation can lead to trees that may be predisposed to abiotic factors such as drought, and biotic factors includ-ing lethal secondary wood-boring insects and pathogens, for example, the two-lined chestnut borer (Agrilus bilineatus) (Weber) and pathogens such as Armillaria root rot (Armillaria mellea). Common to most woody landscape and forest trees, host plant re-sistance for L. dispar has historically been compiled from anecdotal field studies and observations (Forbush and Fernald 1896, Mosher 1915) and while helpful, these studies require some level of interpretation as for host susceptibility, preference, and suitability. More recently, Liebhold et al. (1995) ranked over 600 North American angiosperm and gymnosperm tree species as to their susceptibility to the gypsy moth based on previous field and laboratory tests.
These susceptibility and suitability rankings were based on gypsy moth defoliation, larval abundance, growth and survival, larval foli-age preference, pupal weights, and ratio of pupal weight on host to the pupal weight on white oak (Q. alba) (standard host) (Liebhold et al. 1995). Concurrently, extensive studies have focused on the relative susceptibility and suitability of black oak (Q. velutina), bur oak (Q. macrocarpa), cherrybark oak (Q. pagoda), northern red oak (Q. rubra), pin oak (Q. palutris), swamp white oak (Q. bicolor), white oak (Q. alba), willow oak, (Q. phellos), northern pin oak (Q. ellipsoidalis), southern red oak (Q. falcata), water oak (Q. nigra), chestnut oak (Q. prinus), post oak (Q. stellata), sessile oak (Q. petraea), Turkey oak (Q. cerris), Hungarian oak (Q. frainetto), and Garry oak (Q. garryana) be-cause of the importance of the genus in the ecology and sustainability of eastern and mid-western North American and European forests (Barbosa and Capinera 1977; Barbosa and Greenblatt 1979; Barbosa et al. 1983; Lechowicz and Jobin 1983; Mauffette et al. 1983; Lechowicz and Mauffette 1986; Mill-er and Hanson 1989a,b; Miller et al. 1991; Montgomery 1991; Davidson et al. 1999; Foss and Rieske 2003; Rieske et al. 2003; Shields et al. 2003; Miller 2008; Milanovic et al. 2014). However, of the over 600 tree species listed in Liebhold et al. (1995), only
9
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

8 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Carpinus caroliniana, and Ostrya virginiana are listed with suitability rankings of 2 and 1, respectively. Additionally, Mauffette et al. (1983) found O. virginiana to be strongly pre-ferred in Quebec compared to New England where it is considered an intermediate host.
With the recent accessibility of China to foreign plant exploration, a wide variety of Asiatic woody plant material, including Carpinus and Ostrya taxa, has been discov-ered and developed for possible use in tree breeding programs and for increasing plant diversity in urban forests and landscapes (Ware 1992, 1995). With the recent loss of millions of North American ash (Fraxinus spp.) trees to the emerald ash borer (EAB) (Agrilus planipennis Fairmaire), and maples (Acer spp.) and other hardwoods to the Asian long-horned beetle (ALB) (Anoplophora gla-bripennis (Motschulsky)), there is an import-ant and critical need for the development, and availability of new urban tree species for replanting in EAB and/or ALB affected areas, and to minimize the economic impact on communities and municipalities for the need for chemical protection from gypsy moth defoliation (Poland and McCullough 2006, Raupp et al. 2006, Sydnor et al. 2007, Kovacs et al. 2010, Sadof et al. 2011, Van-natta et al. 2012, Hauer and Peterson 2017, Herms et al. 2019). With the exception of the aforementioned North American species of C. caroliniana and O. virginiana (Liebhold et al. 1995), to the best of our knowledge, no studies have been conducted on the relative susceptibility, preference, and suitability of recently acquired and developed Carpinus and Ostrya taxa of North American, Europe-an, and Asian parentage for the gypsy moth. Here, we report the results of a study to determine the relative susceptibility, feeding preference, and suitability of Carpinus and Osytra taxa in no-choice and multiple-choice laboratory feeding bioassays for gypsy moth larvae. Results from this study will contrib-ute to the use of additional taxa in landscape and urban forest plantings, which will min-imize the need for application of chemical insecticides, and contribute to more diverse landscapes and urban forest settings.
Materials and Methods
No-choice (NC) laboratory larval feeding trials. No-choice larval feeding trials were conducted using newly hatched first instar gypsy moth larvae. Twenty Carpinus taxa and three Ostrya taxa were evaluated for relative susceptibility, larval feeding preference, and suitability for larval development (refer to Table 1 for a listing of Carpinus and Osytra taxa tested). Candidate Carpinus and Ostrya taxa were growing in The Morton Arboretum (TMA) tree breeding
nursery production area at Lisle, IL, and ranged in height from 2 to 4 m and 8 to 10 cm dbh.
For the laboratory no-choice (NC) and multiple choice (MC) bioassay feeding tests, fully developed Carpinus leaves were randomly selected from the terminal 15 cm of each of four mid-canopy branches per tree from all four cardinal directions. The leaves were brought back to the entomology lab and held in cold storage in plastic bags at 5 °C for a maximum of 2 d. Leaves from each single tree replicate were combined for the laboratory bioassays. Depending on tree availability, one to three individual trees (replicates) each of either Carpinus or Ostrya taxon were evaluated. Bur oak (Q. macrocarpa) a highly preferred host of the gypsy moth, served as the standard.
First instar gypsy moth larvae, used in the NC and MC laboratory feeding studies, were reared from overwintering egg masses collected in late spring from infested trees on the grounds of TMA. Prior to use, the egg masses were held in brown Kraft paper bags in a refrigerator at approximately 4 °C. The egg masses were then placed in an incubator under a photoperiod of 16:8 (L:D) at approximately 25 °C. Within 24 hours of hatching, a cohort of 10 larvae was randomly selected and placed in a one liter (one quart) plastic container, with a screened lid to allow for ventilation, with foliage from the test Carpinus or Ostrya taxon. The containers were placed in plastic trays and were held in an incubator under a photoperiod of 16:8 (L:D) at approximately 25 °C. The containers were examined daily for larval mortality, evidence of feeding, and pupation. Foliage was replaced every two days. Depending on availability, one to three single tree repli-cates, of each Carpinus and Ostrya taxon, were assayed with 10 individual larvae for a total of 10 to 30 larvae per taxon. The bioassay for a given larva was terminated at adult emergence. Larval longevity was determined as the difference in the days from the date the larva was first introduced to the foliage until pupation or death. Larval development time was the difference in days from initial introduction to the foliage until pre-pupation. Within 12 hours of pupation, each individual pupa was weighed (nearest 0.001 g) to obtain the pupal fresh weight. Gypsy moth pupae were held at room tem-perature until adult emergence or until the pupa was determined to be dead due to lack of movement following repeated prodding with a probe. The proportion of larvae reach-ing pupation was calculated by recording the total number of larvae that pupated in each of the containers for all single tree replicates (1 to 3) for a given taxon. At the termination of the no-choice larval feeding bioassay trial,
10
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 9
the fecal pellets from each of the containers for each taxon were dried in an oven at 50 °C and then weighed (nearest 0.001 g).
Multiple-choice (MC) laboratory larval feeding bioassay. The larval mul-tiple-choice laboratory feeding bioassays were conducted as previously described by Miller et al. (2001a). For each of the three multiple-choice bioassays, ten first instar gypsy moth larvae were placed into each of 10 plastic petri dishes (0.6 × 15.0 cm). Each petri dish served as a replicate. Three leaf discs, 2.54 cm in diameter, representing one of each of the different Carpinus taxon choic-es, were placed into each dish and randomly arranged around the perimeter. The larvae had access to all foliage discs. The petri dish-es were placed in clear plastic bags to prevent drying of the leaf discs and were held in an incubator under a photoperiod of 16:8 (L:D) at ~25 °C. Condensation of water on the lid of the petri dish indicated a high relative humidity. The dishes were examined daily for three days. Each day, the foliage discs were removed from the dishes, replaced, and visually evaluated by two independent estimators, to the nearest 5%, and their estimates were averaged. The estimators used a defoliation template to determine the proportion of leaf tissue removed (Hall and Townsend 1987; Miller and Ware 1997, 1999a; Miller et al. 1999, 2001a, 2001b). New foliage discs were arranged randomly around the perimeter of each dish to elimi-nate possible bias.
Measuring Carpinus leaf tough-ness and thickness. Prior to using the leaves for the NC laboratory feeding trials, each leaf was measured for leaf thickness and inner and outer leaf toughness. Leaves were collected in the field as previously described. In the lab, leaf thickness was determined by measuring the leaf using a Vernier caliper to the nearest micrometer. For outer leaf toughness, the pentrometer (Chatillon™ digital force meter, Greensboro, N.C.) was applied within 0.5 cm from the leaf edge; for inner toughness, the pentrometer was applied to the center of the leaf adjoining the mid-rib.
Measures of larval susceptibility, preference, and suitability. Measures of susceptibility for gypsy moth larvae was quantified by the percent of leaf tissue removed in the NC feeding bioassays and the amount of larval frass production (dry weight). Host preference was a function of the percent of leaf tissue removed in the MC feeding bioassays. Host suitability was determined by larval development time to pupation (days), percent of larvae pupating, pupal fresh weight (mg), and the percent of adult emergence. In order to further define
host suitability for gypsy moth larvae, pu-pation, and adult emergence, we employed a suitability ranking system as described by Montgomery (1991), and summarized by Liebhold et al. (1995) by calculating the ratio of male and female pupal fresh weights for larvae feeding on Carpinus and Ostrya hosts evaluated in the NC feeding studies to the male and female pupal fresh weights (male = 0.339 g; female = 0.886 g) for larvae feeding on bur oak (Q. macrocarpa), a preferred stan-dard. For example, in our study, the mean suitability ranking (SR) for C. caroliniana was calculated to be 0.65 (0.221 g/0.339 g) for male pupae and 0.68 (0.720 g/0.886 g) for female pupae (where the numerator is the male/female pupal fresh weight for larvae feeding on C. caroliniana, and the denominator is the male/female pupal fresh weight for larvae feeding on Q. macrocarpa, respectively).
Statistical Analysis. Measures of susceptibility, preference, and suitability for gypsy moth larvae were subjected to analysis of variance (ANOVA) using taxon as the main effect. Means of significant effect (5%) were compared with the Dunn’s test. Coefficients of correlation were calculated by regression analysis for larval development time with percent pupation, percent adult emergence, pupal fresh weight, frass dry weight, leaf thickness, and leaf toughness; percent pupation was regressed with percent adult emergence; pupal fresh weight with frass dry weight, and leaf thickness and leaf toughness; and leaf thickness and toughness with mean frass dry weight. Host suitability rankings (SRs) were regressed with male and female pupal weights. The percent of leaf tissue removed, proportion of larvae pupat-ing, and proportion of adults emerging for each taxon were arcsine transformed before analysis to correct for non-normality. All data are presented as original means ± SEM. Data were analyzed using the SigmaStat for Windows (Jandel Scientific 1992).
Results
Gypsy moth larval development time to pupation. A summary of mean larval development time to pre-pupation is presented in Table 1. Overall, there was no significant difference in mean larval de-velopment time to pre-pupation with mean male and female larvae taking 46 and 47 days, respectively. Larvae failed to pupate when feeding on C. coreana, C. fargesii, C. orientalis, and the hybrid C. caroliniana × C. orientalis, living a mean of 11 days (range = 5 to 23 days). Male larvae feeding on C. betula, C. turczanianari, O. japonica, Q. macrocarpa (reference species), and the hybrids of C. car-oliniana × C. cordata, and C. caroliniana × C.
11
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

10 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2Ta
ble
1. G
ypsy
mot
h m
ean
larv
al d
evel
opm
ent,
perc
ent p
upat
ion,
per
cent
adu
lt e
mer
genc
e, m
ean
mal
e an
d fe
mal
e pu
pal w
eigh
ts, a
nd fr
ass
dry
wei
ghts
whe
n fe
edin
g on
sel
ecte
d C
arpi
nus
and
Ost
rya
taxa
.
Mea
n
La
rval
M
ean
%
Mea
n %
M
ean
Pup
al
Mea
n D
ry
Dev
elop
men
t2,3
P
upat
ion
Adu
lt E
mer
genc
e Fr
esh
Wei
ghts
Fr
ass
Wt.
Taxa
N
1 (d
ays)
M
ale
Fem
ale
Mal
e Fe
mal
e M
ale
(g)
Fem
ale
(g)
(g)
Car
pinu
s be
tulu
s 30
43
±2.9
b
2
7±0.
08ab
43
±0.0
9b
20±0
.07a
37
±0.0
9ab
0.31
9±0.
028a
b 0.
720±
0.07
8ab
11.3
91±1
.1b
C. c
arol
inia
na
10
45±1
.3b
10±
0.06
a 10
±0.0
6a
3±0.
03a
13±0
.06a
0
.221
±0.0
54a
0.60
6±0.
060a
b 17
.350
±1.6
bC
. cau
scis
a 30
46
±2.4
b
4
0±0.
07b
50±0
.09a
b 27
±0.0
8a
37±0
.09a
b 0.
342±
0.01
3b
0.77
6±0.
047a
b 13
.669
±1.2
bC
. cor
data
20
34
±6.3
ab
17±0
.07a
10
±0.0
6a
10±0
.07a
10
±0.0
6ab
0.26
7±0.
047a
b 0.
609±
0.17
2ab
6
.260
±6.3
aC
. cor
eana
10
5
±0.0
a
0±0.
00a
0±0.
00a
0±0.
00a
0±0.
00a
0.0
00±0
.0a
----
----
C. f
arge
sii
10
8±0
.0a
0±
0.00
a 0±
0.00
a 0±
0.00
a 0±
0.13
a 0
.000
±0.0
a ---
- ---
-C
. jap
onic
a 20
41
±0.4
b
2
0±0.
13a
20±0
.13a
20
±0.1
3a
20±0
.13a
b 0
.260
±0.0
a --
-- 7.
284±
6.9a
C. l
axifl
ora
10
18±0
.0ab
25
±0.1
1a
25±0
.13a
20
±0.1
3a
20±0
.10a
b 0
.286
±0.0
62ab
0
.693
±0.2
41ab
13
.698
±1.2
bC
. ori
enta
lis
10
23+0
.0a
0
±0.0
0a
0±0.
00a
0±0.
00a
0±0.
00a
0.0
00±0
.0a
----
----
C. t
scho
nosk
ii 20
44
±4.2
b
4
5±0.
11b
40±0
.11b
35
±0.1
1a
30±0
.10a
b 0
.355
±0.0
75b
0.5
55±0
.079
a 19
.826
±2.0
bC
. tur
czan
inar
i 20
42
±3.1
b
3
5±0.
11ab
10
±0.0
7a
3
0±0.
11a
5±0.
05a
0.3
22±0
.015
b --
---
13.6
68±2
.1b
Ost
rya
carp
inifo
lia
20
54±3
.1b
40±
0.11
b 25
±0.1
0ab
3
5±0.
11a
25±0
.10a
b 0
.258
±0.0
20a
0.7
85±0
.052
ab
14.5
16±1
.6b
O. j
apon
ica
20
38±3
.2b
50±
0.12
b 25
±0.1
0ab
3
0±0.
11a
20±0
.09a
b 0
.332
±0.0
31b
0.9
07±0
.061
b 9.
162±
1.0a
bO
. vir
gini
ana
10
34±0
.6ab
10
±0.1
0a
20±0
.13a
b 10
±0.1
0a
20±0
.13a
b ---
- ---
--
7.56
6±8.
1aC
. bet
ulus
x ts
chon
oski
i 30
45
±2.6
b
4
3±0.
09b
47±0
.09b
30
±0.0
9a
37±0
.09a
b 0.
290±
0.02
0ab
0.70
0±0.
058a
b 3.
852±
0.3a
C. c
arol
inia
na x
bet
ulus
30
51
±3.6
b
3
0±0.
09ab
37
±0.0
9ab
23±0
.08a
30
±0.0
9ab
0.30
4±0.
024a
b 0.
572±
0.04
9a
10.7
80±1
.0ab
C. c
arol
. x (b
etul
us x
tsch
onos
kii)
30
45±4
.1b
43±
0.09
b 30
±0.0
9ab
17±0
.07a
30
±0.0
9ab
0.22
7±0.
016a
0.
523±
0.08
6a
13.3
34±1
.2b
C. c
arol
inia
na x
cor
data
30
48
±3.9
b
4
0±0.
09b
27±0
.08a
b 30
±0.0
9b
27±0
.08a
b 0.
299±
0.02
0ab
0.67
3±0.
107a
b 18
.110
±2.0
bC
. car
olin
iana
x c
orea
na
30
44±0
.6b
20±0
.07a
27
±0.0
8ab
20±0
.07a
27
±0.0
8ab
0.25
7±0.
017a
0
.718
±0.0
44ab
11
.924
±1.5
bC
. car
olin
iana
x la
xiflo
ra
20
40±4
.6b
17±
0.08
a 7±
0.07
a 13
±0.0
8a
7±0.
07a
0.2
35±0
.028
a --
---
6.5
89±1
.0a
C. c
arol
inia
na x
ori
enta
lis
30
6±0
.0a
0±
0.00
a 0±
0.00
a
0±0.
00a
0±0.
00a
----
-----
---
--C
. car
olin
iana
x ts
chon
oski
i 30
38
±0.6
b
4
6±0.
09b
7±0.
05a
46±0
.09a
7±
0.05
a 0.
236±
0.02
7a
0.45
7±0.
001a
15
.047
±1.4
b‘C
. cor
data
x ja
poni
ca
20
34±0
.7ab
13
±0.0
8a
7±0.
07a
13±0
.08a
b 7±
0.07
a 0.
262±
0.03
1ab
0.5
09±0
.079
a 10
.417
±1.3
abQ
uerc
us m
acro
carp
a
30
57±2
.7b
30±0
.09a
b 70
±0.0
9b
30±0
.09a
b 70
±0.0
9b
0.33
9±0.
040b
0
.886
±0.0
51b
9.2
13±a
b
Sign
ifica
nce:
F=
109.
5 F=
34.6
F=
67.7
F=
111.
8 F=
60.7
F=
34
F=2.
7 F=
3.9
P<0.
001
P<0.
016
P<0.
001
P<0.
001
P<0.
001
P<0.
001
P=0.
002
P<0.
001
1 N =
Num
ber o
f sin
gle
petr
i dis
h re
plic
ates
per
Car
pinu
s tax
on
2 Mea
ns fo
llow
ed b
y th
e sa
me
lett
er a
re n
ot s
igni
fican
tly d
iffer
ent (
Dun
ns’ m
ultip
le co
mpa
riso
n te
st; P
<0.0
5)3 L
arva
l dev
elop
men
t (da
ys) i
s th
e tim
e fr
om w
hen
the
larv
ae w
ere
intr
oduc
ed to
the
Car
pinu
s fol
iage
unt
il pr
e-pu
pate
d4 M
ean
perc
ent p
upat
ion
is th
e pe
rcen
t of o
rigi
nal g
ypsy
larv
ae p
upat
ing
5 Mea
n pe
rcen
t adu
lt em
erge
nce
is th
e pe
rcen
t of o
rigi
nal l
arva
e em
ergi
ng a
s ad
ults
12
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 11
laxiflora completed pupation in significantly fewer days (mean = 56 days; range = 51 to 59 days) compared to larvae feeding on C. caroliniana, C. tschonoskii, and the hybrid C. caroliniana × C. tschonoskii (mean = 66 days; range = 65 to 67 days) (F = 31.1; P = 0.04). Larvae feeding on the remaining Carpinus and Ostrya taxa were intermediate in com-pleting pupation (mean = 60 days; range = 54 to 66 days). Female larvae, feeding on O. japonica, O. virginiana, and the reference species, Q. macrocarpa completed pupation in a significantly shorter time (mean = 54 days; range = 51 to 56 days) compared to larvae feeding on C. japonica, C. turcza-moamari., and the hybrid C. caroliniana × C. laxiflora taking a mean of 72 days (range = 71 to 73 days) (F = 2.8; P < 0.001). Female larvae feeding on the remaining taxa were intermediate in completing pupation (mean = 62 days; range = 57 to 67 days).
Frass production. Larval frass dry weight production is summarized in Table 1. Larvae feeding on C. betulus, C. caroliniana, C. causcica, C. laxiflora, C. tschonoskii,, C. turczaninari, and O. carpinifolia, and the hybrids of C. caroliniana × C. cordata, C. caroliniana × C. coreana, and C. caroliniana × C. tschonoskii produced significantly more frass (mean = 14.5 g; range = 11 to 18 g) compared to larvae feeding on C. cordata, C. japonica, O. virginiana, and the hybrids of C. betula × C. tschonoskii, C. caroliniana × C. laxiflora with less than 7.4 g of frass pro-duced (F = 3.9; P < 0.001). Larvae feeding on the other Carpinus and Ostrya taxa and the reference species, Q. macrocarpa produced intermediate amounts of frass (mean = 10.1 g; range 9.162 to 10.417 g). Larvae feeding on C. coreana, C. fargesii, C. orientalis, and the hybrid C. caroliniana × C. orientalis did not produce any frass (Table 1). The time for larval development to pupation was not reflective of frass production, (r2 = 0.09; P = 0.15) and frass production was not a good predictor of pupal fresh weight (r2 = 0.10; P = 0.19).
Mean pupal fresh weight. Male and female gypsy moth pupal fresh weights are summarized in Table 1. For all Caprinus and Osytra taxa, male pupae had significantly lower mean pupal fresh weights (0.293 g) compared to female pupae (0.727 g) (t = 22287; P < 0.001). Male pupal weight was significantly higher for larvae feeding on C. causisca, C. tschonoskii, C. turczamamari, O. japonica, and the reference species, Q. macrocarpa compared to larvae feeding on C. caroliniana, C. japonica, O. carpinifolia, and the hybrids C. caroliniana × (C. betula × C. tschonoskii), C. caroliniana × C. coreana, C. caroliniana × C. laxiflora, and C. caroliniana × C. tschonoskii (F = 35.4; P = 0.01). The remaining Carpinus taxa had intermediate
pupal fresh weights. Female pupal weights were significantly higher for larvae feeding on the preferred host of Q. macrocarpa (0.886 g) and the highly susceptible O. japonica (0.907 g) compared to larvae feeding on C. tschonoskii (0.555 g) and the hybrids C. car-oliniana × C. betula (0.572 g) C. caroliniana × (C. betula × C. tschonoskii) (0.227 g), C. caroliniana × C. tschonoskii (0.457 g) and C. cordata × C. japonica (0.509 g) (F = 2.7; P = 0.002). Female pupal weights for larvae feeding on the remaining Carpinus and Os-trya taxa had intermediate pupal weights. There was no relationship (r2 = 0.0; P = 0.94) between larval development time to pupation and pupal fresh weight.
Pupation and adult emergence. Mean percent gypsy moth pupation and adult emergence is summarized in Table 1. For all Carpinus and Ostyra taxa, there was no significant difference in mean male percent pupation (28%) versus female pupation (27%). Male larvae feeding on a majority of the Carpinus and Ostrya taxa had significantly higher percent pupation rates of (mean = 39%; range = 27% to 50%) compared to male larvae feeding on leaves of C. coreana, C. fargesii, C. orientalis, and the hybrid C. caroliniana × C. orientalis all of which failed to pupate (F = 6.1; P < 0.001). Male larvae feeding on C. caroliniana, C. japonica, C. turczamamari , and the hybrid C. cordata × C. japonica had intermediate percent pupation (mean = 11%; range 10 to 13%). Thirty percent of male larvae pu-pated when feeding on the reference host, Q. macrocarpa. Mean percent pupation for female larvae mirrored male pupation for larvae feeding on all three Ostrya taxa and eight Carpinus taxa with significantly higher mean percent pupation (mean = 33%; range = 20% to 47%) compared to remaining taxa (Table 1). Larval development time to pupation was a strong predictor of percent pupation (r2 = 0.83; P < 0.001).
There was no significant difference in percent male versus percent female adult emergence (23% versus 24%) with a male to female ratio of 1.2. Adult emergence was strongly correlated with both larval develop-ment time to pupation (r2 = 0.90; P < 0.001) and percent pupation (r2 = 0.83; P < 0.001) with adults emerging on all of the Carpinus and Ostrya taxa tested (Table 1).
Multiple-choice larval feeding preference bioassay. A summary of the three gypsy moth larval multiple-choice (MC) studies is presented in Table 2. In MC #1, gypsy moth larvae preferred C. betulus over C. coreana and C. fargesii (24%, 20% and 6% leaf tissue removed, respectively) (F = 19.6; P < 0.001). Carpinus caroliniana was highly preferred over C. coreana and
13
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

12 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
C. fargesii in MC #2 (26%, 8%, and 3%, re-spectively) (F = 28.7; P < 0.001). In MC #3, the hybrid C. caroliniana × C. orientalis was the least preferred (2% leaf tissue removed) compared to C. caroliniana and C. orientalis alone, both with 15% leaf tissue removed (F = 20.3; P < 0.001) (Table 2). In MC studies #1 and #2, feeding preference and susceptibility (NC studies) were highly correlated. In MC study #1, gypsy moth larvae preferred the susceptible C. betula over the less suscep-tible C. coreana and C. fargessi (r2 = 0.99; P = 0.04), and in MC study #2, the more susceptible C. caroliniana was much more preferred over the less susceptible C. cordata and C. fargesii (r2 = 0.99; P = 0.04). No such relationship was found in MC study #3, for gypsy moth larvae, when given a choice, be-tween C. caroliniana, C. orientalis, and the hybrid C. caroliniana × C. orientalis.
Carpinus leaf thickness and tough-ness. A summary of leaf thickness and inner and outer leaf toughness of Carpinus taxa is presented in Table 4. Carpinus betulus, C. caroliniana, C. orientalis, C. turczani-novii, and the hybrids of C. caroliniana × C. cordata, C. caroliniana × C. coreana, C. caroliniana × C. orientalis, C. cordata × C. japonica had significantly thicker leaves while C. japonica and C. tschonoskii had the thinnest leaves (F = 116.6; P < 0.001). Leaves of the hybrids of C. betulus × (C. bet-ulus × C. tschonoskii), and C. caroliniana ×
Table 2. Multiple-choice studies for gypsy moth larvae feeding on Carpinus taxa (2006). Mean % LeafTaxa Tissue Removed1
MC-1C. betulus 24±5.3bC. coreana 20±0.8aC. fargesii 6±2.5a
Significance: F=19.6 P<0.001
MC-2C. caroliniana 26±3.8cC. coreana 8±1.6bC. fargesii 3±1.0a
Significance: F=28.7 P<0.001
MC-3C. caroliniana 15±2.7bC. orientalis 15±3.7bC. caroliniana x C. orientalis 2±0.8a
Significance: F=20.3 P<0.0011Values within columns followed by the same letter are not significantly different (P<0.05; Dunn’s test)
Table 3. Suitability rankings (SRs) for selected Carpinus and Ostrya taxa for male and female gypsy moth pupae compared to the reference species, Quercus macrocarpa.Taxa Male Pupal Female Pupal Suitability Ranking1 Suitability Ranking1
Carpinus betulus 0.941±0.0829ab 0.813±0.0888abC. caroliniana 0.651±0.158a 0.684±0.0680abC. causcisa 1.009±0.0376b 0.876±0.0532abC. cordata 0.789±0.138ab 0.688±0.194abC. japonica 0.767±0.0257ab ----2
C. laxiflora 0.844±0183ab 0.782±0.272abC. tschonoskii 1.047±0.221b 0.626±0.0892aC. turczaninari 0.951±0.0458ab ----2
Ostrya carpinifolia 0.760±0.0584ab 0.709±0.183abO. japonica 0.978±0.0912ab 1.023±0.0693bC. betulus x tschonoskii 0.641±0.120a 0.790±0.0660abC. caroliniana x betulus 0.896±0.0726ab 0.646±0.0558aC. carolinian x (betulus x tschonoskii) 0.670±0.0489ab 0.590±0.0971aC. caroliniana x cordata 0.882±0.0594ab 0.633±0.160aC. caroliniana x coreana 0.758±0.0514ab 0.810±0.0512abC. caroliniana x laxiflora 0.693±0.0828ab ----2
C. caroliniana x tschonoskii 0.697±0.0788ab 0.516±0.00113aC. cordata x japonica 0.773±0.0919ab 0.574±0.0892aQuercus macrocarpa 0.999±0.119b 1.000±0.0572b Significance: F=1.7, P=0.049 F=2.5, P=0.0041Values within columns followed by the same letter are not significantly different (P<0.05; Dunn’s test)2Larvae failed to reach pupation
14
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 13
C. laxiflora, and the species, C. cordata, and C. laxiflora were intermediate in thickness. Inner leaf portions of the hybrids C. betulus × tschonoskii, C. caroliniana × coreana, C. caroliniana × orientalis, and species, C. betulus and C. cordata, were significantly tougher than the inner leaf portions of C. japonica leaves. Inner leaf toughness for the other taxa was intermediate (F = 62.2; P < 0.001). Outer leaf toughness was less variable with only the leaves of the hybrids C. caroliniana × cordata and C. caroliniana × orientalis being significantly tougher than C. japonica (F = 68.1; P < 0.001). Outer leaf toughness for the remaining Carpinus taxa was intermediate (Table 4). Leaf thickness was correlated with outer leaf toughness (r2 = 0.25; P = 0.03), but was not related to inner leaf toughness.
Effect of leaf thickness and tough-ness on the susceptibility and suitabil-ity of Carpinus taxa to larval feeding. Larval development time to pre-pupation was not correlated with Carpinus leaf thick-ness (r2 = 0.03; P=0.67) but was related to inner (r2 = 0.30; P = 0.03) and outer leaf thickness (r2 = 0.30; P = 0.03). Further, mean pupal fresh weight and leaf thickness (r2 = 0.12, P=0.18) were not related, but pupal fresh weight was related to inner (r2 = 0.31, 0.56; P = 0.04) and outer leaf toughness (r2
= 0.31, 0.56; P = 0.04). Leaf thickness and outer leaf toughness do not appear to be good predictors of frass production (thickness = r2 = 0.20; P = 0.08; outer toughness = r2 = 0.18; P = 0.10), but frass production was related to inner leaf toughness (r2 = 0.24; P = 0.05).
Suitability Rankings (SRs). Suit-able rankings (SRs), for male and female gypsy moth larvae feeding on Carpinus and Ostyra taxa, are summarized in Table 3. For all taxa tested, the overall mean male SR was 0.85 (range = 0.64 to 1.05) and 0.77 (range = 0.52 to 1.02) for female pupae, indicating that the majority of the Carpinus taxa tested in this study have a moderate to high host suitability for gypsy moth larvae. The SRs for C. caroliniana (male = 0.66 and female = 0.68), and for O. virginiana (male = 0.76 and female = 0.95) were within Liebhold’s et al. (1995) rating of 2 (SR range = 0.5 to 0.9) for a marginal to intermediate host (i.e. C. caroliniana), and a rating of 1.0 (SR great-er than 0.9) for an intermediate to highly preferred host (i.e. O. virginiana). Host suit-ability significantly affected male and female pupal weights differently (t = 22287.000; P = <0.001). Overall, female pupae were approximately 2.5 times heavier than male pupae (male = 0.293 g versus female = 0.721 g). Our findings, for these two taxa, are consistent with results reported by Mosher
Table 4. Summary of leaf thickness (microns), and inner and outer leaf toughness (grams) of selected Carpinus taxa. Thickness Toughness (grams)Taxa1 (microns) Inner OuterCarpinus betulus 250.00±25.0d 17.8286±5.9b 19.5657±1.0bCarpinus betulus × (C. betulus × C. tschonoskii) 145.00±13.0ab 16.3657±4.8ab 14.4457±2.1abCarpinus betulus × C. tschonoskii 180.00±18.0b 19.5657±5.7bc 17.0057±2.5abCarpinus caroliniana 202.50±14.0c 17.3714±4.9b 15.0857±3.2abCarpinus caroliniana × (C. betulus × C. tschonoskii) 175.00±17.5b 15.2686±6.1ab 13.8057±1.4abCarpinus caroliniana × C. cordata 202.50±40.2c 16.2743±5.2ab 16.9143±1.5abCarpinus caroliniana × C. coreana 222.50±44.1c 21.5771±5.6bc 23.3143±2.1cCarpinus caroliniana × C. laxiflora 162.50±10.7b 16.6857±1.2ab 15.0400±1.2abCarpinus caroliniana × C. orientalis 212.50±11.0c 48.8343±14.6c 45.0743±5.4dCarpinus carolinana × C. tschonoskii 190.00±9.5b 16.4571±4.2ab 14.7200±1.6abCarpinus cordata 145.00±11.6ab 21.0286±8.6bc 15.8171±2.1abCarpinus cordata × C. japonica 170.00±13.6b 16.0914±4.1ab 14.7200±2.0abCarpinus coreana 207.50±22.1c 19.4743±5.2bc 19.2000±1.8bCarpinus fargesii 180.00±10.8b 19.8400±5.3ab 18.5600±1.6abCarpinus japonica 117.50±8.9a 9.9657±2.1a 8.96000±1.0aCarpinus laxiflora 125.00±6.2ab 16.0000±4.0ab 14.9943±1.3abCarpinus orientalis 207.50±21.6c 27.8857±8.7bc 29.9886±3.2bcCarpinus tschonoskii 157.50±10.9ab 16.9143±4.8ab 17.0057±1.6abCarpinus turczaninovii 207.50±21.1c 17.0057±4.2ab 15.3600±1.6abSignificance: F=116.6 F=62.2 F=68.1 P<0.001 P<0.001 P<0.0011Values within columns followed by the same letter are not significantly different (P<0.05; Dunn’s test)
15
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

14 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
(1915), Mauffette et al. (1983), Montgomery (1991), Twery (1991), Liebhold et al. (1995), and Fite (2019). Carpinus causcica (SR = 1.00) and C. tschonoskii (SR = 1.05), and the reference species, Q. macrocarpa (SR = 1.00) were significantly more suitable hosts for male larval development compared to male larvae feeding on C. caroliniana (SR = 0.65), and the hybrid C. betulus × C. tschonoskii (SR = 0.64). All of the additional Carpinus and Osytra taxa (SR range = 0.67 to 0.98) were intermediately suitable for male gypsy moth larvae. Ostrya japonica (SR = 1.02) and the reference species, Q. macrocarpa (SR = 1.00) were significantly more suitable for female larval development compared to C. tschonoskii (SR = 0.63), and the hybrids, C. caroliniana × C. betula (SR = 0.65), C. caro-liniana × (C. betula × C. tschonoskii) (SR = 0.59), C. caroliniana × C. cordata (SR = 0.63), C. caroliniana × C. tschonoskii (SR = 0.52), and C. cordata × C. japonica (SR = 0.57). All of the additional Carpinus and Osytra taxa (SR range = 0.68 to 0.81) were intermediately suitable for female gypsy moth larval devel-opment (F = 2.5; P = 0.004) (Table 3). Both male and female pupal weights were strong predictors of suitability (male: r2 = 0.88; P < 0.001; female: r2 = 0.87; P < 0.001).
DiscussionIn this study, we examined the relative
susceptibility, preference, and suitability of Carpinus and Ostrya taxa for feeding and development by gypsy moth larvae, which will hopefully provide new insight into the potential use of these taxa in an overall tree breeding program.
Gypsy moth larvae feed on a wide va-riety of woody plant hosts. Variation in host susceptibility, preference, and suitability can be quite variable across and within genera (Mosher 1915, Barbosa and Capinera 1977, Barbosa and Greenblatt 1979, Mauffette et al. 1983, Lechowicz and Mauffette 1986, Berisford et al. 1990, Montgomery 1991, Liebhold et al. 1995, Foss and Rieske 2003, Miller 2008, Pearse 2011, Fite 2019).
Plants protect themselves from her-bivores in one of two ways; by using either physical plant traits, chemical defenses, and/or a combination of both. In all likelihood, that is the case here with Carpinus and Ostrya taxa (Hoxie et al. 1975; Meredith and Schuster 1979; Johnson et al. 1980a,b; Ryan et al. 1982; Matsuda and Senbo 1986; Tingey and Laubengayer 1986; Doss et al. 1987; Potter and Kimmerer 1989; Ranney and Walgenbach 1992; Spicer et al. 1995; Patton et al. 1997; Fulcher et al. 1998; Rowe and Potter 2000; Dalin and Bjorkman 2003). Gypsy moth host plant leaf chemistry has been widely studied on a variety of plant
hosts, and undoubtedly plays a major role in host plant defense against the gypsy moth (Thorsteinson 1960; Barbosa and Greenblatt 1979; Mattson 1980; Scriber and Slansky 1981; Schultz et al. 1982; Martinat and Barbosa 1987; Mattson and Scriber 1987; Schultz 1988; Foss and Rieske 2003; Rieske et al. 2003; Paluch et al. 2008, 2009), but was beyond the scope of this study. However, here we will discuss physical plant charac-teristics (i.e. leaf thickness, toughness, and pubescence) of Carpinus leaves, and their possible role in relative host susceptibility, preference, and suitability for gypsy moth larvae.
Relative susceptibility, preference and suitability of Carpinus taxa for gypsy moth larval development. The acceptability of a host plant by a herbivore is considered an indicator of host suscepti-bility and suitability (Martinat and Barbosa 1987, Montgomery 1994). In our study, host susceptibility was reflective of host suit-ability (r2 = 0.50; P < 0.001). Using larval frass production as a measure of feeding susceptibility, the vast majority of the taxa tested here appear to be moderately to highly susceptible to feeding by gypsy moth larvae; exceptions being C. coreana, C. fargesii, and C. orientalis and the hybrid C. caroliniana × C.orientalis where larvae failed to produce any measurable frass.
Physical plant traits, such as leaf toughness and thickness, can be main factors affecting invertebrate feeding, and usually correlate with leaf fiber and lignin content (Tanton 1962, Graca and Zimmer 2005). Agrawal and Fishbein (2006) found leaf toughness could be used to predict her-bivory of many plants including milkweeds (Asclepias spp.). Leaf thickness and tough-ness may also play a role in the relative susceptibility, preference, and suitability of Carpinus and Osytra taxa for feeding by and development of gypsy moth larvae. Pearse (2011) found that for leaf-feeding caterpil-lars, oak leaf toughness was one of the best predictors of tussock moth survival along with water content, and condensed and total tannin content. He also he found a negative correlation between caterpillar survival and leaf toughness. Conversely, Keathley and Potter (2008) found that fall webworm larvae preferred the thicker and tougher leaves of lilac (Syringa vulgaris), Bradford pear (Pyrus calleryana ‘Decaisne’), dogwood (Cornus florida), and tuliptree (Lirodendron tulipfera) over the thinner, and more tender leaves of sassafras (Sassafras albidum) and little leaf linden (Tilia cordata). Our results are similar to Keathley and Potter (2008) in that inner and outer leaf toughness was positively correlated with gypsy moth larval longevity, but larval longevity was not cor-
16
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 15
related with leaf thickness. Additionally, in our multiple choice feeding bioassays, when given a choice, gypsy moth larvae tended to prefer the thicker and tougher leaves of C. betulus, C. caroliniana, C. coreana, and C. orientalis over the thinner and less tough C. fargesii leaves.
Larval survival has been a common criteria used to document host suitability, with high first instar survival indicating high suitability and low first instar larval survival suggesting poor host susceptibility (Barbosa and Krischik 1987; Martinat and Barbosa 1987; Miller and Hanson 1989 a,b). In our study, larval development time and survival were not correlated with leaf thick-ness, but were correlated with inner and outer leaf toughness. Larvae feeding on the tougher leaves of C. coreana, C. orientalis, and the hybrid C. caroliniana × C. orientalis only lived 7 to 22 days, failed to pupate, and produced little or no frass. Additionally, only 20% and 27% of larvae feeding on the tough-er leaves of C. caroliniana and C. cordata pupated, respectively. Carpinus orientalis has thick and tough leaves, and is native to Eastern Europe and Asia Minor which most likely overlaps with the historical origins of the gypsy moth. Consequently, historical co-evolution may have contributed to the reduced suitability of C. orientalis as a host. Miller et al. (2001a) found that spring and fall cankerworm larvae feeding on the thicker and tougher Asian elm leaves lived a shorter period of time, had extended larval development time, failed to pupate or had low (less than 11% pupation) rates, and re-moved less than 15% of leaf tissue compared to larvae feeding on thinner and less tough North American elm taxa. However, co-evo-lution does not completely explain the lack of larval host suitability. For example, C. car-oliniana, which is native to North America, has tough leaves and is considered to be an intermediate to moderate gypsy moth larval host (Mosher 1915; Montgomery 1991, 1994; Fite 2019). Conversely, C. coreana, which is native to Korea, also has tough leaves, but when given a choice was not preferred by gypsy moth larvae. In all likelihood, neither of these aforementioned plant species would have co-evolved with the gypsy moth, but would be expected to be somewhat suitable hosts.
Gypsy moth larvae generally complete development between 35–40 days on highly suitable hosts (Barbosa and Capinera 1977, Barbosa and Greenblatt 1979, Miller and Hanson 1989a). In our study, there was no significant difference in male and female larval development time to pupation (males = 46 days; females = 47 days) on suitable hosts; development time was well within the range for known moderate to highly suitable
hosts (mean = 42 days; range = 34–51 days) (Barbosa and Capinera 1977, Barbosa and Greenblatt 1979, Miller and Hanson 1989a). In this study, larval development time to pupation was highly correlated with percent pupation and adult emergence (pupation: r2 = 0.83; P < 0.001; adult emergence (r2 = 0.81, P < 0.001), indicating that the longer the lar-vae fed the more likely they were to pupate and emerge as adults. Conversely, larvae feeding on unsuitable Carpinus hosts lived a mean of l3 days (range of 5-23 days) and none of the larvae completed larval development or pupated. However, longer larval develop-ment time to pupation did not necessarily reflect a higher pupal fresh weight or greater larval frass production, and frass production did not reflect pupal fresh weight. For exam-ple, larvae feeding on the highly preferred host, bur oak (Q. macrocarpa) took 43 days to reach the pupal stage, but produced sig-nificantly less frass (9.2 mg). Larvae feeding on the moderately suitable C. carolinana and C. causcisa hosts took a similar number of days (45 and 46 days, respectively) to reach the pupal stage, but produced significantly more frass. Consistent with our findings, Miller et al. (1991) found that larval devel-opment was not correlated with pupal fresh weight or frass production for gypsy moth larvae feeding on Garry oak (Q. garryana) and red alder (Alnus rubra). Additionally, Foss and Rieske (2003) found that bur oak (Q. macrocarpa) was the most preferred of five oak species tested, but gypsy moth larvae consumed relatively small amounts of foli-age and developed rapidly. Bur oak leaves in their study were the toughest, contained the highest levels of carbohydrates and tan-nins, and only moderate levels of nitrogen. Similarly, Miller, F. (in prep) also found that gypsy moth larvae feeding on thicker and tougher bur oak leaves had a relatively shorter development time and lower frass production compared to six other common oak species tested. Results from this study and other gypsy moth suitability studies suggest that there is considerable variation either in the nutritional value and/or host plant chemistry of Carpinus and Oystra taxa which may significantly affect larval devel-opment, but not enough to prevent pupation.
It is well-known that leaf trichomes help protect plants from insect herbivores specifically as it relates to feeding, growth, survival, and oviposition (Pillemer and Ting-ey 1976, Ramalho et al. 1984, Fordyce and Agrawal 2001). Feeding may be negatively correlated with trichome density, which is generally considered a “soft weapon” in plant defense compared to other plant traits (Levin 1973; Danielson et al. 1987; Potter et al. 1998; Miller and Ware 1999a,b; Miller et al. 1999; Miller 2000; Miller et al. 2001a,b;
17
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

16 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Dalin and Bjorkman 2003; Dalin et al. 2008). Plants have the ability to produce glandular (chemical-producing) and non-glandular tri-chomes. They may vary in morphology and genetics, and even within individual plant species (MacLean and Byers 1983; South-wood 1986; Agrawal 1999, 2000; Werker 2000; Dalin and Bjorkman 2003; Loe et al. 2007). Non-glandular trichomes function in structural defense and have low nutritional value while glandular trichomes provide both structural and chemical defense and may contain terpenes and alkaloids that act as feeding deterrents or toxins (Levin 1973, Rautio et al. 2002, War et al. 2012). Gypsy moth larvae feed by consuming the entire leaf, including leaf veins, in contrast with other insect herbivores, which skeletonize (i.e. Japanese beetle, Popillia japonica Newman), “windowpane” (i.e. leaf beetle larvae), or chew holes in the leaves (i.e. adult leaf beetles and flea weevils). In this study, pubescent and non-pubescent leaves were equally fed upon, indicating that leaf and veinal pubescence does not affect feeding susceptibility, preference, or suitability. For example, C. japonica and C. tschonoskii both have pubescent leaves, were moderately susceptible to larval feeding, and were highly suitable for larval development (Krussman 1976). Similarly, the leaves of C. caroliniana, C. cordata, C. orientalis, and C. turczaniowii all have thin pubescence on the veins and midrib on the leaf underside (Krussman 1976), and were also found to be moderately to highly susceptible and preferred, but not very suitable. Miller et al. (2001a) found, in multiple-choice bioassays feeding studies, that there was no consistent feeding prefer-ence pattern for leaf consuming spring and fall cankerworm larvae feeding on pubescent elm leaves.
Using the suitability rankings (SRs), as previously described by Montgomery (1991) and Liebhold et al. (1995), we found a strong correlation between both male and female pupal weights and host suitability, (male pupae: r2 = 0.87; P < 0.001; female pupae: r2 = 0.88; P < 0.001). Suitability appears to have a significant effect on both male and female pupal weights with females weighing nearly 2.5 times more than males. A mean suitability ranking (SR) of 0.85 for male pupae (range = 0.64 to 1.05) and female pupae 0.77 (range = 0.52 to 1.02), for all taxa, indicate that the vast majority of Carpinus and Ostrya taxa have moderate to high host suitability for gypsy moth lar-vae. More specifically, our SRs for male and female gypsy moth pupae for C. caroliniana (male = 0.66; female = 0.68) and SRs of 0.76 (male) and 0.95 (female) for O. virginiana, are comparable to suitability rankings iden-tified by Liebhold et al. (1995) suggesting C.
caroliniana represents an intermediate host and O. virginiana an intermediate to highly preferred host (Mosher 1915, Montgomery 1991, Twery 1991, Liebhold et al. 1995, Fite 2019).
Hybridization, feeding suscepti-bility and preference, and suitability of Carpinus taxa for gypsy moth larvae. In this study, it appears that hybridization may have a either a stabilizing, positive, or negative influence on host susceptibility, preference, and suitability for gypsy moth larvae, may be polygenic, and may not be the result of single gene expression (Paige and Capman 1993, Orions 2000, Cheng et al. 2011). It is well known that secondary metabolites are a common chemical defense employed by plants against herbivorous insects and hybridization may increase the variation of secondary metabolites affecting herbivore resistance (Rieseberg and Ell-strand 1993, Patton et al. 1997, Fulcher et al. 1998, Orians 2000, Cheng et al. 2011). Fur-ther, most secondary metabolites (SMs) in hybrids may also be present in the parents, but hybrids may also miss some parental secondary metabolites or have novel ones (Paige and Capman 1993, Cheng et al. 2011, Lopez-Caamal and Tovar-Sanchez 2014). Hybridization appeared to have a stabilizing effect on the moderate suitability of the indi-vidual species of C. betulus, C. caroliniana, C. cordata, and C. tschonoskii and their as-sociated hybrids of C. betulus × tschonoskii, C. carolinana × betulus, C. caroliniana × (betulus × tschonoskii, and C. caroliniana × cordata) which were also moderately suitable for gypsy moth larval development. Further, when the moderately suitable C. caroliniana (SR = 0.65) was hybridized with the unsuitable (SR = 0, no larvae reached pu-pation) species of C. coreana, the SR for the resulting hybrid (C. caroliniana × C. cordata) increased (SR = 0.81). In contrast, when the highly preferred (SR = 0.80) C. cordata was hybridized with the unsuitable C. japonica (SR = 0), the SR value for the hybrid (C. cordata × C. japonica) decreased (SR = 0.57). Apparently, the unsuitable traits associated with C. coreana were not preserved in the hybrid leading to higher suitability, but were preserved with C. japonica resulting in lower suitability. More dramatically, when C. caroliniana was crossed with the unsuitable species of C. orientalis (SR = 0), the resulting C. caroliniana × C. orientalis hybrid was also unsuitable (SR = 0) for gypsy moth larval development. These findings are further supported in multiple-choice study #3 (MC-3), where, when given a choice, gypsy moth larvae removed significantly more leaf tissue from C. caroliniana and C. orientalis leaves compared to the C. caroliniana × C. orientalis hybrid. In contrast, hybridization
18
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 17
rendered the C. caroliniana × C. laxiflora hy-brid unsuitable compared to the moderately suitable parents of C. carolinana (SR = 0.65) and C. laxiflora (SR = 0.79).
In conclusion, based on the finding in this study, there does not appear to be a large pool of Carpinus or Ostrya taxa suitable for future tree breeding programs and for use in areas where gypsy moth outbreaks are frequent. However, there are a few Carpinus taxa (i.e. C. coreana, C. fargesii, C. orientalis, and the hybrid C. carolinana × C. orientalis) on which gypsy moth larvae were not able to complete development, pu-pate, or reach the adult stage. The genera of Carpinus and Ostrya consist of a number of small to medium-sized deciduous trees which are distributed across the temperate regions of the northern hemisphere. Most are slow growing and can tolerate a variety of soil conditions (i.e. moisture, pH, and texture), and require minimal maintenance (i.e. pruning) making them good candidates for use in urban settings as hedges, screen-ing and group plantings, and parkway and landscape trees particularly in areas where chronic gypsy moth feeding pressure is ab-sent or rare, reducing the need for intensive chemical protection (Krussman 1976, Raupp et al. 1992, Dirr 2009, Sjoman et al. 2019). In order to make better use of the genotypic and phenotypic attributes of Carpinus and Osytra taxa in urban landscapes, where chronic insect defoliation is common, further studies are needed to determine the factor(s) responsible for feeding susceptibility and preference, and host suitability of Carpinus and Ostrya taxa for gypsy moth larvae.
Acknowledgments
The authors would like to express their sincere thanks to members of The Morton Arboretum entomology lab and grounds staff for their assistance in conducting this study. A special note of thanks is extended to the J. Frank Schmidt Family Charitable Foundation for their generosity in providing on-going financial support for this project. A special thanks to K. Shearer for her helpful comments on an earlier draft of the manu-script, and to H. Anderson and L. Prazak for their assistance in editing and formatting the manuscript.
Literature Cited
Agrawal, A.A., 1999. Induced responses to herbivory in wild radish: effects on several herbivores and plant fitness. Ecology 80: 1713–1723.
Agrawal, AA. 2000. Specificity of induced re-sistance in wild radish: causes and conse-
quences for two specialist and two generalist caterpillars. Oikos 89: 493–500.
Agrawal, A.A. and M. Fishbein. 2006. Plant defense syndromes. Ecology 87: S132–S149.
Barbosa, P. and J.L. Capinera. 1977. The influ-ence of food on developmental characteristics of the gypsy moth, Lymantria dispar (L.). Ca-nadian Journal of Zoology. 55(9): 1424–1429.
Barbosa, P. and J. Greenblatt. 1979. Suitabil-ity, digestibility, and assimilation of various host plants of the gypsy moth, Lymantria dispar L. Oecologia 43: 111–119.
Barbosa, P. and V.A. Krischik. 1987. Influence of alkaloids on feeding preference of eastern deciduous forest trees by gypsy moth, Lyman-tria dispar. American Naturalist 130: 53–69.
Barbosa, P., M. Waldvogel, P. Martinat, and L.W. Douglass. 1983. Developmental and reproductive performance of the gypsy moth (Lymantria dispar (L.) (Lepidoptera: Lymantridae), on selected hosts common to the mid-Atlantic and southern forests. Environmental Entomology 12: 1858–1862.
Berisford, C.W., T.J. Lanigan, and M.E. Mont-gomery. 1990. Performance of gypsy moth larvae on hosts from the deep south: survival, development, and host preferences. USDA Gypsy Moth Research Review. Pg. 48.
Cheng, D., K. Vrieling, and P.G.L. Kinkhamer. 2011. The effect of hybridization on second-ary metabolites and herbivore resistance: implications for the evolution of chemical diversity in plants. Phytochemical Review 10: 107–117.
Dalin, P. and C. Bjorkman. 2003. Adult beetle grazing induces willow trichome defense against subsequent larval feeding. Oecologia 134: 112–118.
Dalin, P., J. Agren, C. Bjorkman, P. Hut-tumen, and K. Karkkainen. 2008. Leaf trichome formation and plant resistance to herbivory. Chapter 4 in Induced plant resis-tance to herbivory. Pages 89–105.
Danielson, S.D., G.R. Manglitz, and E.L. Sorensen. 1987. Resistance of perenni-al glandular-haired Medicago species to oviposition by alfalfa weevils (Coleoptera: Curculionidae). Environmental Entomology 16: 195–197.
Davidson, C.B., K.W. Gottschalk, and J.E. Johnson. 1999. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. Forest Science 45(1): 74–83.
Dirr, M.A., 2009. Manual of woody landscape plants: Their identification, ornamental char-acteristics, culture, propagation, and uses. Stipes Publishing Company, Champaign, Illinois. 826 pp.
19
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

18 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Doss, R.P., C.H. Shanks, Jr., J. D. Chamber-lain, and J.K.L. Garth. 1987. Role of leaf hairs in resistance of a clone of strawberry, Fragaria chiloensis, to feeding by adult black vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae). Environmental Entomology 16: 764–766.
Elkinton, J.S. and A.M. Liebhold. 1990. Pop-ulations dynamics of gypsy moth in North America. Annual Review of Entomology 35: 571–596.
Fite, K. 2019. Gypsy moth host preferences. Bartlett Research Lab Technical Report. 2 pp.
Forbush, E.H. and C.H. Fernald. 1896. The gypsy moth. Porthetria dispar (Linn.). Bos-ton: Wright and Potter. 495 p.
Fordyce, J.A. and A.A. Agrawal. 2001. The role of plant trichomes and caterpillar group size on growth and defense of the pipevine swal-lowtail Battus philenor. Journal of Animal Ecology 70: 997–1005.
Foss, L.K. and L.K. Rieske. 2003. Species-specif-ic differences in oak foliage affect preference and performance of gypsy moth caterpillars. Entomologia Experimentalis et Applicata 108: 87–93.
Fulcher, A.F., T.G. Ranney, and J.D. Burton. 1998. Role of foliar phenolics in host plant resistance of Malus taxa to adult Japanese beetles. HortScience 233: 862–865.
Graca, M.A.S. and M. Zimmer. 2005. Leaf toughness (Ch-18). Methods to study litter decomposition: A practical guide, 121–125. M.A.S. Graca, F. Barlocher, and M.O. Gess-ner eds.
Hall, R.W. and A.M. Townsend. 1987. Suitabil-ity of Ulmus wilsoniana, the ‘Urban elm’, and their hybrids for the elm leaf beetle, Xanthogaleruca luteola (Muller) (Coleoptera: Chrysomelidae). Environmental Entomology 16: 1042–1044.
Hauer, R.J., and W.D. Peterson. 2017. Effects of emerald ash borer on municipal forestry budgets. Landscape Urban Planning 157: 98–105.
Herms D.A., D.G. McCullough, C.S. Sadof, D.R. Smitley. F.D. Miller, and C.W. Cranshaw. 2019. Insecticide options for protecting ash trees from emerald ash bor-er. North Central IPM Center Bulletin. 3rd Edition. 16 pp.
Hoxie, R.P., S.G. Wellso, and J.A. Webster. 1975. Cereal leaf beetle response to wheat trichome length and density. Environmental Entomology 4: 365–370.
Jandel Scientific. 1992. SigmaStat for Windows. San Rafael, CA.
Johnson, K.J.R., E.L. Sorenson, and E.K. Horber. 1980a. Resistance in glandular
haired annual Medicago species to feeding by adult alfalfa weevils. Environmental Entomology 9: 133–136.
Johnson, K.J.R., E.L. Sorenson, and E.K. Horber. 1980b. Resistance of glandular haired annual Medicago species to oviposi-tion by adult alfalfa weevils. Environmental Entomology 9: 241–245.
Keathley, C.P. and D.A. Potter. 2008. Quantita-tive resistance traits and suitability of woody plant species for a polyphagous Scarab, Popillia japonica Newman. Environmental Entomology 37(6): 1548–1557.
Kovacs, K.F., R.G. Haight, D.G. McCullough, R.J. Mercader, N.W. Siegert, and A.M. Liebhold. 2010. Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecological Economics 69: 569–578.
Krussman, G. 1976. Manual of cultivated broad-leaved trees and shrubs (Vol. I: A-D). Translated by M.E. Epp. Technical Editor G.S. Daniels. 448 pp.
Lechowicz, M.J. and L. Jobin. 1983. Esti-mating the susceptibility of tree species to attack by the gypsy moth, Lymantria dispar. Ecological Entomology 8: 171–183.
Lechowicz, M.J. and Y. Mauffette. 1986. Host preferences of the gypsy moth in eastern North American versus European forests. Revue d’Entomologie du Quebec 31: 43–51.
Levin, D.A. 1973. The role of trichomes in plant defense. Quarterly Review of Biology 48: 3–15.
Liebhold, A.M., K.W. Gottschalk, R.M. Muz-ika, M.E. Montgomery, R. Young, K. O’Day, and B. Kelley. 1995. Suitability of North American tree species to the gypsy moth: A summary of field and laboratory tests. USDA-Forest Service. GTR-NE-211. 34 pp.
Loe, G., P. Torang, M. Gaudeul, and J. Agren. 2007. Trichome production and spatiotemporal variation in herbivory in the perennial herb, Arabidospsis lyrata. Oikos 116: 134–142.
Lopez-Caamal, A. and E.Tovar-Sanchez. 2014. Genetic, morphological, and chemical pat-terns of plant hybridization. Revista Chilena de Historia Natural 87:16.
MacLean, P.S., and R.A. Byers. 1983. Ovi-positional preferences of the alfalfa blotch leafminer (Diptera: Agromyzidae) among some simple and glandular-haired Medica-go species. Environmental Entomology 12: 1083–1086.
Martinat, P.J. and P. Barbosa. 1987. Rela-tionship between host plant acceptability and suitability in newly enclosed first-instar gypsy moths, Lymantria dispar (L.) (Lepi-
20
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 19
doptera: Lymantriidae). Annual of the En-tomological Society of America 80: 141–147.
Matsuda, K. and S. Senbo. 1986. Chlorogenic acid as a feeding deterrent for the salicace-ae-feeding leaf beetle, Lochmateae capreae cribrata (Coleoptera: Chrysomelidae) and other species of leaf beetles. Applied Ento-mology and Zoology 21: 411–416.
Mattson, W.J. 1980. Herbivory in relation to plant nitrogen content. Annual Review of Ecological Systems. 11: 119–161.
Mattson, W.J., and J.M. Scriber. 1987. Nutri-tional ecology of insect foliovores of woody plants: nitrogen, water, fiber, and mineral considerations. Pp. 105–146. In J.G. Rodri-guez and F. Slansky, Jr. (eds.). Nutritional ecology of insects, mites, and related inver-tebrates. Wiley, New York.
Mauffette, Y., M.J. Lechowicz, and L. Jobin. 1983. Host preferences of the gypsy moth, Lymantria dispar (L.) in southern Quebec. Canadian Journal of Forest Research 13: 53–60.
Meredith, W.R., Jr. and M.F. Schuster. 1979. Tolerance of glabrous and pubescent cottons to tarnished plant bug. Crop Science 19: 484–488.
Milanovic, S., J. Lazarevic, Z. Popovic, Z. Miletic, M. Kostic, Z. Radulovic, D. Karadzic, and A. Vuleta. 2014. Preference and performance of the larvae of Lymantria dispar (Lepidoptera: Lymantridae) on three species of European oaks. European Journal of Entomology 111 (3): 1–8.
Miller, F. 2000. Insect resistance of elm geno-types. [Proceedings of the First International Elm Conference.] Entitled: The Elms: Breed-ing; Conservation, and Disease Management. ed. C. Dunn. pp. 137–154.
Miller, F. 2008. Preliminary report on the rela-tive susceptibility of Quercus taxa for gypsy moth development. International Oaks: The Journal of the International Oak Society #19: 63–64.
Miller, J.C. and P.E. Hanson. 1989a. Labo-ratory feeding tests on the development of gypsy moth larvae with reference to plant taxa and allelochemicals. Station Bulletin #674. Oregon State University, Agriculture Experiment Station, Corvallis, Oregon. 63 p.
Miller, J.C. and P.E. Hanson. 1989b. Labo-ratory feeding tests on the development of gypsy moth, Lymantria dispar (L.) (Lepi-doptera: Lymantriidae) larvae on foliage of gymnosperms. Canadian Entomologist 121(6): 425–429.
Miller, F. and G. Ware. 1997. Preference for and suitability of Asian elm species and hybrids for the adult elm leaf beetle (Coleoptera:
Chrysomelidae). Journal of Economic Ento-mology 90(6): 1641–1645.
Miller, F., and G. Ware. 1999a. Feeding pref-erences for selected Tilia spp. and cultivars by the adult Japanese beetle (Coleoptera: Scarabaeidae). Journal of Arboriculture 25: 168–173.
Miller, F. and G. Ware. 1999b. Resistance of elms of the Ulmus davidiana complex to defoliation by the adult elm leaf beetle (Cole-optera: Chrysomelidae). Journal of Economic Entomology 92(5): 1147–1150.
Miller, F., S. Jerdan, and G. Ware. 1999. Feeding preference of adult Japanese beetles (Coleoptera: Scarabaeidae) for Asian elm spe-cies and their hybrids. Journal of Economic Entomology 92(2): 421–426.
Miller, F., K. Malmquist, and G. Ware. 2001a. Evaluation of Asian, European, and North American elm (Ulmus spp.) biotypes to feed-ing by spring and fall cankerworms. Journal of Environmental Horticulture 19: 216–221.
Miller, F., G.Ware, and J. Jackson. 2001b. Preference of temperate Chinese elms (Ulmus spp.) for the adult Japanese beetle (Cole-optera:Scarabaeidae). Journal of Economic Entomology 94(2): 445–448.
Miller, J.C., P.E. Hanson, ad D.M. Kimber-ling. 1991. Development of the gypsy moth (Lepidoptera: Lymantriidae) on Garry oak and red alder in western North America. En-vironmental Entomology 20(4): 1097–1101.
Montgomery, M.E. 1991. Variation in the suit-ability of tree species for gypsy moth. USDA gypsy moth research review. Pgs. 1–13.
Montgomery, M.E. 1994. Comparison on several tree species of growth and survival of newly hatched larvae of gypsy moth from three continents. Northeastern Forest Experiment Station, Hamden, CT. 2 p.
Mosher, F. H. 1915. Food plants of the gypsy moth in America. USDA Bull. No. 250, 39p.
Orians, C.M. 2000. The effects of hybridization in plants on secondary chemistry: implications for the ecology and evolution of plant-her-bivore interactions. American Journal of Botany 87:1749–1756.
Paige, K.N. and W.C. Capman. 1993. The Ef-fects of Host-Plant Genotype, Hybridization, and Environment on Gall-Aphid Attack and Survival in Cottonwood: The Importance of Genetic Studies and the Utility of RFLPS. Evolution 47(1): 36–45.
Paluch, G., F. Miller, J. Zhu, and J. Coats. 2008. Influence of elm foliar chemistry for the host suitability of the Japanese beetle, Popillia japonica, and the gypsy moth Ly-mantria dispar. Journal of Agriculture and Urban Entomology 23 (4): 209–223.
21
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

20 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Paluch, G., F. Miller, J. Zhu, and J. Coats. 2009. Total phenolic content of Asian elm leaves and host plant suitability for gypsy moth. Journal of Environmental Horticulture 27 (2): 105–108.
Patton, C.A., T.G. Ranney, J.D. Burton, and J.F. Walgenbach. 1997. Feeding response of Japanese beetle to naturally occurring me-tabolites found in rosaceous plants. Journal of Environmental Horticulture 15: 222–227.
Pearse, I.S. 2011. The role of leaf defensive traits in oaks on the preference and performance of a polyphagous herbivore, Orgyia vetusta. Ecological Entomology 36: 635–642.
Pillemer, E.A. and W.M. Tingey. 1976. Hooked trichomes: a physical barrier to a major agri-cultural pest. Science 193: 482–484.
Poland, T.M. and D.G. McCullough. 2006. Emerald ash borer: invasion of the urban forest and the threat to North American ash resources. Journal of Forestry 104: 118–124.
Potter, D.A. and T.W. Kimmerer. 1989. Inhi-bition of herbivory on young holly leaves: evidence for the defensive role of saponins. Oecologia 78: 322–329.
Potter, D.A., P.G. Spicer, D. Held, and R.E. McNiel. 1998. Relative susceptibility of cultivars o flowering crabapples, lindens, and roses to defoliation by Japanese beetles. Journal of Environmental Horticulture 16: 105–110.
Ramalho, F.S., W.L. Parrott, J.N. Jenkins, and J.D. McCarty, Jr. 1984. Effects of cot-ton leaf trichomes on the mobility of newly hatched tobacco budworms (Lepidoptera: Noctuidae). Journal of Economic Entomology 77: 619–621.
Ranney, T.G. and J. F. Walgenbach. 1992. Feeding preference of Japanese beetles for taxa of birch, cherry and crabapple. Journal of Environmental Horticulture 10: 177–180.
Raupp, M. J., A. B. Cumming, and E. C. Raupp. 2006. Street tree diversity in eastern North American and its potential for tree loss to exotic borers. Arboriculture and Urban Forestry 32: 297–304.
Raupp, M. J., C. S. Koehler, and J. A. Da-vidson. 1992. Advances in implementing integrated pest management for woody land-scape plants. Annual Review of Entomology 37: 561–586.
Rautio, P., A. Markkola, M. Martgel, J. Tu-omi, E. Harma, K. Kuikka, A. Siitonen, IL Riesco, M. Roitto. 2002. Developmental plasticity in birch leaves: defoliation causes a shift from glandular to non-glandular tri-chomes. Oikos 98: 437–446.
Rieseberg L.H. and N.C. Ellstrand. 1993. What can molecular and morphological markers
tell us about plant hybridization? Critical Review in Plant Science 12: 213–241.
Rieske, L.K., C.c. Rhoades, and S.P. Miller. 2003. Foliar chemistry and gypsy moth, Lymantria dispar (L.), herbivory on pure American chestnut, Castanea dentate (Fam. Fagaceae), and a disease resistant hybrid. Environmental Entomology 32(2): 359–365.
Rowe, W.J. and D.A. Potter. 2000. Shading effects on susceptibility of Rosa spp. to de-foliation by Popillia japonica (Coleoptera: Scarabaeidae). Environmental Entomology 29(3): 502–508.
Ryan, J.D., P. Gregory, and W.M. Tingey. 1982. Phenolic oxidase activities in glandular trichomes of Solanum berthaultii. Phyto-chemistry 21: 1885–1887.
Sadof, C. S., L. Purcell, F. J. Bishop, C. Que-sada, and Z. Zhang. 2011. Evaluating restoration capacity and costs of managing the emerald ash borer with a web-based cost calculator in urban forests. Arboriculture and Urban Forestry 37: 74–83.
Schultz, J.C. 1988. Many factors influence the evolution of herbivore diets, but plant chem-istry is essential. Ecology 69: 896–897.
Schultz, J.C., P.J. Nothnagle, and I.T. Bald-win. 1982. Individual and seasonal variation in leaf quality of two northern hardwood tree species. American Journal of Botany 69: 753–759.
Scriber, J.M. and F. Slansky. 1981. The nutri-tional ecology of immature insects. Annual Review of Entomology 26: 183–211.
Shields, V.D.C., B.P. Broomell, and J.O.B. Sal-ako. 2003. Host selection and acceptability of selected tree species by gypsy moth larvae, Lymantria dispar (L.). Annuals of the Ento-mological Society of America 96: 920–926.
Sjoman, H., S. Hannus, P. Bellan, T. Barblish-vili, T. Darchidze, and S. Sikharulidze. 2019. Hunting for a larger diversity of urban trees in Western Europe – A case study from the southern Caucasus. Arboriculture and Urban Forestry 45(5): 221–235.
Southwood, S.R. 1986. Plant surfaces and insects – an overview. In: Juniper B. South-wood SR (eds). Insects and the plant surface. Arnold, London. Pp 1–22.
Spicer, P.G., D.A. Potter, and R.E. McNiel. 1995. Resistance of flowering crabapple cul-tivars to defoliation by the Japanese beetle (Coleoptera: Scarabaeidae). Journal of Eco-nomic Entomology 88: 979–985.
Sydnor, T.D., M. Bumgardner, and A. Todd. 2007. The potential economic impacts of emerald ash borer (Agrilus planipennis) on Ohio communities. Arboriculture and Urban Forestry 33(1): 48–54.
22
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 21
Tanton, M.T. 1962. The effect of leaf “toughness” on the feeding of larvae of the mustard bee-tle Phaedon cochleariae Fab. Entomologia Experimentalis et Applicatation 5: 74–78.
Thorsteinson, A.J. 1960. Host selections in phytophagous insects. Annual Review of Entomology 5: 193–218.
Tingey, W.M. and J.E. Laubengayer. 1986. Glandular trichomes of a resistant hybrid potato alter feeding behavior of the potato leafhopper (Homoptera: Cicadellidae). Jour-nal of Economic Entomology 79: 1230–1234.
Twery, M.J. 1991. Effects of defoliation by gypsy moth. Proceedings of USDA Interagency Gypsy Moth Research Review 1990 (ed. by Gottschalk, K.W., Twery, M.J., and Smith S.I.) pp. 27–39. USDA Forest Service Techni-cal Bulletin NE-146, East Windsor CT, USA.
Vannatta, A.R., R. H. Hauer, and N.M. Schuettpelz. 2012. Economic analysis of emerald ash borer (Coleoptera: Buprestidae) management options. Journal of Economic Entomology 105: 196-206.
War, A.R., M.G. Paulraj, T.Ahmad, A.A.Buh-roo, B. Hussain, S.Ignacimuthu, and H.C.Sharma. 2012. Mechanisms of plant defense against insect herbivores. Plant Sig-naling and Behavior 7(10): 1306–1320. Avail-able from https://doi.org/10.4161/psb.21663.
Ware, G. 1992. Elm breeding and improvement at The Morton Arboretum. The Morton Ar-boretum Quarterly 28: 846–849.
Ware, G. 1995. Little-known elms from China: Landscape tree possibilities. Journal of Ar-boriculture 21: 284–288.
Werker, E. 2000. Trichome density and develop-ment. Advanced Botanical Research 31: 1–35.
23
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

22 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
The monarch butterfly (Danaus plexippus L.) (Lepidoptera: Nymphalidae) and its migratory behavior have been in-vestigated for well over 70 years. Indeed, since 1975, the eastern cohort of the North American monarch butterfly has become known throughout the world for its spectac-ular annual migration from the provinces of southern Canada and the northern tier states of the United States to fewer than a dozen well-defined overwintering roosts in the montane forests of Michoacan, Mex-ico (National Geographic 1976). Migrating monarchs in Michigan have an extended migratory period, somewhat variable in timing and success from year to year, but temporally wave-like in occurrence, and most pronounced between 15 August and 15 Oc-tober (Douglas 2019). Outside of Michigan, individual monarchs also collectively form successive waves of migrating butterflies that leave their natal locations commencing mid- to late August and continuing through the end of October in most years (Brower 1995). Recent research suggests that a “mi-gration window” exists when the Sun Angle
at Solar Noon (SASN) is about 57° at the leading edge of the migration and 46° at the trailing edge. Ninety percent of the tagged monarchs (Monarch Watch) recovered in Mexico were from monarchs tagged within this window (Taylor et al. 2019).
Despite the current knowledge con-cerning the environmental cues and the variety of possible biophysical and physio-logical receptors involved in inducing the annual migration, the monarch’s pre-migra-tory flight orientation under field conditions remain a poorly understood biological phe-nomenon of seemingly unlimited complexity. One important aspect of monarch migratory behavior that has been ignored under field conditions is a description of any flight orientation behavior that then becomes the directional flight used by migrating butter-flies during the late summer and early fall migrations. In the experiment reported here, I show how migratory butterflies orient when released over open water at a central location equidistant from the continuous forested shoreline (so that the shoreline environment
Spiraling Flight Behavior May Integrate the Biological Compass Systems of Migratory North American
Monarch Butterflies, (Danaus plexippus L.)Matthew M. Douglas
Grand Rapids Community CollegeSenior Adjunct Research Scientist, The University of Kansas
(e-mail: [email protected]).
AbstractHere I report a unique spiraling flight and orientation behavior (comprising multiple
clockwise and counterclockwise circles coupled by extended figure-8 patterns) observed in free-flying “late” migratory monarchs released under sunny ambient field conditions from a location with an unobstructed view of the sky. Following this spiraling flight, migrants continue to fly at very high altitudes until a final orientation and migratory flight direc-tion is established with vanishing bearings statistically concentrated to the SW/W. These results provide preliminary evidence for the possible calibration and use of an inclination magnetic compass that can be used under all local free-flying field conditions. During this spiraling flight, migrating monarch butterflies are proposed to be measuring the direction and inclination of the continuously varying local magnetic field, in a manner analogous to the figure-8 pattern performed to calibrate digital compasses. This behavioral research focuses on the vital pre-migratory orientation and flight behavior that may provide a key behavioral link explaining how migrating monarch butterflies using a time-compensated sun compass system could also integrate the biophysical input from the polarized UV sensitive dorsal rim area (DRA) and the cellular cryptochrome (CRY) system linked to the proposed inclination-based magnetic compass system residing in the distal end of the antennae.
Keywords: monarch migratory orientation, monarch migratory flight, monarch migratory compass systems, ultraviolet sensitive DRA, CRY, inclination-based magnetic compass system in monarchs, orientation flight dance of monarchs.
24
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 23
could not provide immediate unambiguous visual directional cues). Immediately follow-ing release, each butterfly’s flight behavior and vanishing bearing were recorded and analyzed.
Materials and Methods
Wild-caught gravid female monarchs captured in the vicinity of Grand Rapids, Michigan (42.96 N, 856.67 W, USA) in Au-gust 2015 were allowed to oviposit on mature native Asclepias syriaca L. established in a large hexagonal mesh tent (diameter of 5 m and peak dome height of 3 m; mesh of 0.3 cm). The tent was exposed to ambient environmental conditions throughout the study. First instar larvae were removed from the ovipositional plants and reared on fresh milkweed growing within the tent. After pupation, chrysalides were removed and hung on foam supports in the meshed tent. Following eclosion, newly emerged adults fed on nectariferous plants (e.g., cultivars of Lantana spp, Verbena spp, Buddleja spp) as well as at “nectar stations” filled with raw (star thistle) honey and water solution (1:10 ratio), all of which provided adult nutrition, still under ambient conditions within the tent enclosure, until the butterflies were re-leased. In the experimental results reported here, butterflies were released on 7 October 2015 from the middle of a post-glacial kettle lake (Toft (Rome’s) Lake, Newaygo County, Michigan, 43° N, 85° W), which comprises ap-proximately 10 hectares of surface water and surrounding wetlands that terminate in a completely forested, undeveloped shoreline.
Prior to release, butterflies were transferred to large cylindrical mesh cages (64 cm high, 40 cm diameter) composed of a translucent white mesh (with mesh open-ings of .3 cm) and cinch knot tie at the top. They were then transported via a row boat to open water approximately 700 meters from shore in all directions with a clear and unobstructed view of the sky. In all cases, vegetation was reasonably uniform in the background, and immediately unavailable to monarchs after taking flight. Ambient conditions were the following: negligible wind to 4 mps (to avoid butterfly drift), sun-ny skies (to provide adequate UV exposure to the DRA), ambient temperatures of 25°C (to permit controlled flight) and release time between 1:00 and 2:00 PM (an optimal diurnal migratory window for late migratory monarchs in Michigan (unpublished data)). Monarchs were allowed to exit of their own accord from the narrow opening at the top of the mesh cage created when the cinch knot was loosened. The monarchs readily flew up without provocation and, because of the lake surrounding them and the lack of nectarifer-
ous plants at the lakeshore, were compelled to take an orientation. The monarchs were observed until they could no longer be seen with a Nikon 10 × 42, 5.5-degree binocular, at which point the “vanishing bearing” was recorded.
Results
After release, 68 of 71 released mon-archs (96 percent) flew rapidly upward using “powered flight” (e.g., Gibo and Pallett 1979); at first in very tight clockwise circles, but then quickly expanding clockwise circles. At this point, many releases made an extended figure-8 as they circled counter-clockwise, and often repeated this scenario a number of times (Fig. 1), until at very high altitudes (greater than 300m), the individuals flew directionally between 180° and 330° with a mean of 247° (SW/W). (Fig. 2). Monarchs released without any nearby physical struc-ture such as trees, such as along the eastern shore of Lake Michigan, will continue to fly directionally until they are completely out of view. Anecdotally, monarchs have been observed flying over Lake Michigan, over tall buildings such as the Empire State Building, as well as by glider pilots at more than 300 meters (Gibo 1981). The remaining 4 percent of released butterflies exhibited more direct flight, elevating quickly after several small circles, and then flying more directly to high elevation without consistent circling. This is in contrast to “summer monarchs” released during June and July, under similar condi-tions, when only 54 percent of butterflies re-leased exhibited circling behavior (Douglas, unpublished data).
It should be understood that each individual monarch flight orientation is somewhat unique (for example, some first flew counter-clockwise, then clockwise, and some repeated the reversal patterns a num-ber of times prior to maintaining a specific direction). For that reason, the behavior cannot be specifically quantified.
Discussion
With these data of flight behavior and vanishing orientation in mind, it is possible to create a reasonable hypothesis concerning their significance in the much larger picture of monarch butterfly migration. We know that all life on Earth has evolved against a backdrop of environmental cues that entrain an organism’s biological rhythms to the external biophysical cues around it. These biophysical cues, or “zeitgebers” include latitude and longitude, the azimuthal change and passage of the sun across the sky (sun path/day arc); the type of radiation (such as infrared and polarized ultraviolet light) that
25
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

24 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
can be sensed and measured; the strength, declination, inclination, and regional and temporal variation of the magnetic field; the prevailing winds of land and the currents of oceans; gravity, and the rotation of the earth, its inclination, and its annual passage around the sun. This list is not exhaustive. Navigation in many organisms, whether diurnally or nocturnally local, or truly mi-gratory, is dependent on the reception and interpretation of these biophysical cues and their variation over time and location.
Because of the different zeitgebers available, multiple modes of orientation are possible, depending on the group of
organisms. In support of this statement, Barrie (2019) points out that “Much of the confusion that surrounds bird navigation, probably arises from the fact that birds (like many other animals) use a range of different navigational mechanisms and make their choice among them, according to the precise circumstances in which they find themselves. They may well have some way of assessing the quality of information available from each source, before “deciding” which system is likely to be the most reliable, and they probably use different navigational tools at different stages in their journeys.”
Figure 1: Depiction of Spiraling Pre-Migratory Flight Behavior Under Field Conditions (video available online at https://www.youtube.com/watch?v=IcRmcI9v7b4). SASN estimated to be about 45 degrees N.
Toft (Rome’s) Lake: October 7; Clear skies, 25° C, variable winds less than 4mps, butterflies released from central location over the lake and vanishing directions noted: Ninety-six percent of butterflies exhibited circling counterclockwise and clockwise prior to powered directional flight until visibly out of sight, at which point the vanishing direction was taken. Releases took place between 1:00 and 2:00 pm, with SASN elevation at approximately 45 degrees.
26
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1
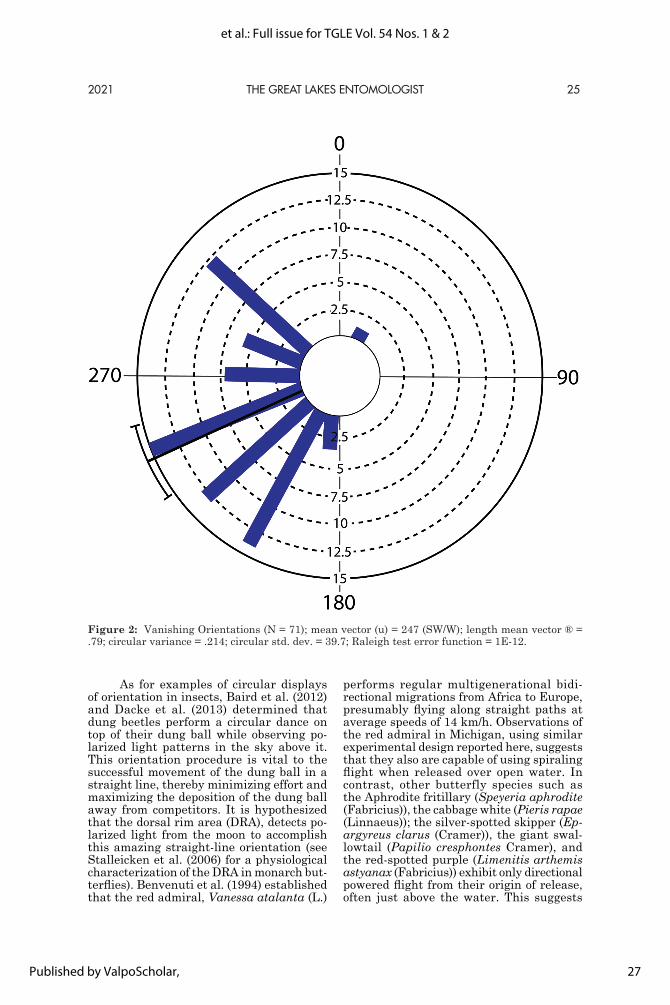
2021 THE GREAT LAKES ENTOMOLOGIST 25
As for examples of circular displays of orientation in insects, Baird et al. (2012) and Dacke et al. (2013) determined that dung beetles perform a circular dance on top of their dung ball while observing po-larized light patterns in the sky above it. This orientation procedure is vital to the successful movement of the dung ball in a straight line, thereby minimizing effort and maximizing the deposition of the dung ball away from competitors. It is hypothesized that the dorsal rim area (DRA), detects po-larized light from the moon to accomplish this amazing straight-line orientation (see Stalleicken et al. (2006) for a physiological characterization of the DRA in monarch but-terflies). Benvenuti et al. (1994) established that the red admiral, Vanessa atalanta (L.)
performs regular multigenerational bidi-rectional migrations from Africa to Europe, presumably flying along straight paths at average speeds of 14 km/h. Observations of the red admiral in Michigan, using similar experimental design reported here, suggests that they also are capable of using spiraling flight when released over open water. In contrast, other butterfly species such as the Aphrodite fritillary (Speyeria aphrodite (Fabricius)), the cabbage white (Pieris rapae (Linnaeus)); the silver-spotted skipper (Ep-argyreus clarus (Cramer)), the giant swal-lowtail (Papilio cresphontes Cramer), and the red-spotted purple (Limenitis arthemis astyanax (Fabricius)) exhibit only directional powered flight from their origin of release, often just above the water. This suggests
Figure 2: Vanishing Orientations (N = 71); mean vector (u) = 247 (SW/W); length mean vector ® = .79; circular variance = .214; circular std. dev. = 39.7; Raleigh test error function = 1E-12.
27
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

26 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
that these latter-mentioned species do not have the compassing capabilities exhibited by monarch butterflies.
Late summer and mid-fall migrating butterflies from the eastern cohort of the North American monarch have a number of mechanisms they can use alone or in com-bination (depending on the environmental zeitgebers available) to orient themselves and navigate their journey to the overwin-tering roosts of Mexico. (These roosts may be over 4,000 km away from their natal locations; monarchs from the most north-ern areas complete this flight in about 75 days traveling a rough average of 50 kms per day, according to multiyear data from Monarch Watch.) For example, migratory monarchs may use a time-compensated sun compass (Perez et al. 1997), polarized light (Reppert et al. 2004, Stalleicken et al. 2005), antennal circadian clocks that coordinate the sun compass orientation (Merlin et al. 2009), the integration of skylight cues via the sun compass (Heinze and Reppert 2011), a magnetic compass measuring the intensity of the local magnetic field as well as the inclination (Guerra et al. 2014). A basic un-derstanding of the neurobiology of monarch butterfly migration has been established by Reppert et al. (2016), although Mouritsen et al. (2013) have suggested that monarch butterflies are not true navigators, and that their displacement from their natal origins a thousand miles west did not affect their ability to orient and choose a consistent flight direction that can be measured directly by their vanishing bearings. In addition, Guilford and Taylor (2014), suggest that a solar heading (of migrating monarchs) need not require time compensation at all. They conclude, “. . . that clock shift experiments alone are neither necessary nor sufficient to identify the occurrence of all conceivable use of solar information in animal orientation, so that a predictable response to clock shift should not be regarded as an acid test of the use of solar information in navigation.”
It seems certain, at least, that mon-archs have a time-compensated sun compass and can measure the seasonal position, azimuth, and height of the sun in the sky; respond to the e-vector produced by the polarization of ultraviolet light; likely use geomagnetic strength and inclination cues as well as “channelizing” geographic barriers such as mountains and oceans. Alone or in combination, migrating monarchs are able to locate fewer than a dozen overwintering roosts at 3,000 m in the oyamel and false white pine forests of the Transvolcanic Mountains of central Mexico in the states of Michoacan and Mexico.
Recent anatomical, neural, and genetic research suggests that sunlight is perceived by the retina and records the azimuthal an-gle of the light. Light polarization is detected by the specialized cells of the dorsal rim area (e.g., Barta et al. 2004, Labhart et al. 2009) and is interpreted by the central complex of the brain, where single neurons “combine the azimuthal location of the sun and the e-vector angle” of the polarized light (Rep-pert 2004). Although the central complex processes the neural input from the sun and its polarized ultraviolet light, the distal tip of the antennae contains the receptive sensilla that control the circadian clock (Merlin et al. 2009). This circadian clock relies on a tran-scriptional-translational auto-regulatory negative feedback loop that drives rhythms in the mRNA and protein levels of core cir-cadian clock components, which involves two cryptochrome proteins, CRY 1 and CRY 2. CRY 1 apparently functions as a blue light photoreceptor that “sets” the circadian clock whereas CRY 2 is similar to the mammalian CRY in that it functions as one of the major repressors in the feedback loop (Reppert et al. 2016). In effect, cryptochromes may define a circadian clock mechanism in mon-arch butterflies that may itself underlie sun compass navigation (Kyriacou 2009). This would not be unexpected as Gegear et al. (2008) determined that cryptochromes mediate light-dependent magnetosensitivy in Drosophila.
As an integral part of their migratory machinery, migrating monarchs can also de-tect the magnetic field of the earth (Zhu et al. 2008). This would likely involve measuring its intensity as well as the inclination—both of which are available but often ambiguous because the inclination signal, for example, can easily be swamped by the daily and the local variations of the magnetic field. Mag-netic sensors have not been found, and they could occur anywhere or everywhere on the body because the earth’s magnetic field easi-ly penetrates living tissue, which means that the presence of magnetite is not necessarily an indicator that an organism possesses the ability to “see” the magnetic field. If so many different zeitgebers are available, and so many mechanisms of detection for orien-tating and maintaining migratory flight are available, how do the monarch butterflies integrate them to commence and maintain migratory orientation?
In 1974, during a graduate ecology class at the University of Kansas, Dr. Orley (Chip) Taylor (founder of Monarch Watch) exposed his students to the strange spiraling flight pattern of some migratory monarchs when release over a semi-open field (1976 personal notes, University of Kansas). At the time we assumed, as had Kanz (1977) and
28
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 27
others, that this flight pattern was a defen-sive strategy in which escaping butterflies flew toward the sun to escape predation. Lin-coln Brower also observed this flight pattern (personal communication, 2001), as has Gibo (1986) who attributed it to gliding and altitu-dinal gain by circling during migration, when they can fly well over 1 km above the ground. And interestingly, released butterflies not exhibiting directional flight (e.g., those “spiraling upwards on thermal currents”) were excluded from analyses by Perez et al. (1997). According to Jonathan Weiner, even Fred Urquhart pondered his observations of spiraling flight: “They form a ring, or circle, and follow each other around and around, like children on a carrousel, drifting in the wind. Caged monarchs, when released, often fly straight up and form this magic circle.” Why, Urquhart does not speculate—perhaps it is an exercise in orientation.” (Weiner 1983). In effect, early reports of “spiraling behavior” were discounted or explained in terms of other possible phenomena.
It should be noted, however, that a specific flight behavior may serve more than one function, just as with the colors of butterfly wings, which serve multiple functions (e.g., thermoregulation, cryptic coloration, warning coloration, and mate attraction). In this case, the unusual and stereotypical flight orientation of migratory monarch butterflies upon release could pro-vide a reliable means of integrating all of the potential mechanisms used by migrating monarch butterflies both for orientation and maintaining a preferred SE-SW flight toward the overwintering roosts in Mexico. This flight pattern cannot be an escape pattern because the final orientation direction during migration, regardless of when or where the butterflies are released, is consistently between SE and SW (Douglas, unpublished data). In addition, released migratory but-terflies do not fly toward the sun; their final vanishing direction is established within a few minutes of flight and is consistent and independent of the time of when they were released. Finally, this flight is active powered flight comprising reversals and figure-8’s—hardly an efficient way to locate thermal updrafts. At first it is invariably a rapid up-ward “explosive” flight (usually less than 30 seconds in duration) and comprises constant and vigorous powered spiraling flight until a specific orientation and direction is achieved, at which point, often thousands of feet above ground, the butterflies continue to intermit-tently use a mix of power flight and gliding, usually in a linear direction, as long as wind direction and strength remain stable. The upward spiraling flight pattern exposes the migratory butterflies to the polarized light patterns of the sun, and the circling and
reversals could conceivably measure the constantly changing local magnetic field of the earth at the release location.
In effect, spiraling flight may be a behavioral means of exposing migratory monarchs to all of the zeitgebers they need to migrate in approximately the right direction toward the overwintering roosts, regardless of the environmental conditions (e.g., as long as ambient temperatures allow flight, monarchs will migrate when skies are sun-ny, partly cloudy, and completely overcast; even during light rain (Floyd Preston, Uni-versity of Kansas; personal observations), when thermals are very weak or absent. As migratory monarchs approach large open expanses of water, such as Lake Michigan, they may continue powered flight direc-tionally over the lake, or perform spiraling formations, singly or in small groups, on sunny and partly cloudy days when thermals may form along the lakeshore. However, monarchs may also fly over Lake Michigan with robust directionality on completely overcast days—when thermals are rare, weak, or unpredictable. Thus, these initial spiraling flights are likely not due to modes of escape, or necessarily to sun location, nor to riding the thermals because there is virtually no temperature variation at their level of flight over the numerous large lakes and wetlands in Michigan during completely overcast days that could provide thermal uplift. However, there is always a constant but temporally variable magnetic field that can provide information, despite the possi-ble lack of a distinct polarized light pattern due to its absorbance by the atmosphere. It makes more sense that these migratory monarchs, forced to orient over open water with few visible landmarks such as trees or buildings, are flying in tight circles and fig-ure-8 patterns in an attempt to calibrate the magnetoreception mechanism—analogous to the figure-8 pattern some digital compasses require for calibration.
In summary, these results may pro-vide preliminary evidence for the possible calibration and use of a monarch inclination magnetic compass that can be used under all local free-flying field conditions in which migrating monarch butterflies are proposed to be measuring the direction and inclination of the local magnetic field, in a manner anal-ogous to the figure-8 pattern performed to calibrate digital compasses. This behavioral research reported here is but a single study of those conducted over variable environ-mental conditions over many years, which focuses on the vital pre-migratory orienta-tion and flight behavior. This “orientation flight dance” may provide a key behavioral link explaining how migrating monarch butterflies using a time-compensated sun
29
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

28 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
compass system could also integrate the biophysical input from the polarized UV sensitive dorsal rim area (DRA) and the cellular cryptochrome (CRY) system linked to the proposed inclination-based magnetic compass system residing in the distal end of the antennae. The orientation- and pre-mi-gratory- “orientation flight dance” performed by migratory monarchs may calibrate and in-tegrate the time-compensated sun compass, the plane-polarized light detection system in the DRA, as well as the magnetic compass of migratory monarchs.
Acknowledgment
Thanks to Brandy Van Zalen for her illustration of monarch spiraling flight behavior.
Literature Cited
Barrie, D. 2019. In Super Navigators: Exploring the wonders of how animals find their way. The Experiment, NY.
Baird, E., Byrne, M.J., Smolka, J., Warrant, E.J. and M. Dacke. 2012 The dung beetle dance: An orientation behavior? PLos ONE. Available from https://doi.org/10.1371/jour-nal.pone.0030211.
Barta, A., and G. Horvath. 2004. Why is it advantageous for animals to detect celestial polarization in the ultraviolet? Skylight polarization under clouds and canopies is strongest in the UV. Journal of Theoretical Biology 226: 429–437.
Benvenuti, S., P. Dall’Antonia, and P.L Ioale. 1994. Migration pattern of the red admiral, Vanessa atalanta L. (Lepidoptera, Nymph-alidae), in Italy. Italian Journal of Zoology 61(3): 343–351.
Brower, L.P. 1995. Understanding and misun-derstanding the migration of the monarch butterfly (Nymphalidae) in North America: 1857–1995. Journal of the Lepidopterists Society 49: 304–385 (1995).
Brower, L.P. 2001. Department of Biology, Sweet Briar College; Personal communication. (TGLE unable to verify).
Dacke, M., Baird, E., Byrne, M., Scholtz, C.H., and E.J. Warrant. 2013. Dung beetles use the Milky Way for orientation. Current Bi-ology 23(4): 298–300.
Douglas, M.M. 2019. Use of Spotted Knapweed/Star Thistle (Asterales: Asteraceae) as the Primary Source of Nectar by early Migrating Monarch Butterflies (Lepidoptera: Nymph-alidae) from Beaver Island, Michigan. The Great Lakes Entomologist 51(3–4): 35–41.
Gegear, R.J., Casselman, A., Waddell, S., and S.M. Reppert. 2008. Cryptochrome
mediates light-dependent magnetosensitivity in Drosophila. Nature 454: 1014–1018. Doi: 10.1038/nature07183.
Gibo D.L. 1981. Altitudes attained by migrating Danaus p. plexippus (Lepidoptera: Da-naidae), as reported by glider pilots. Canadi-an Journal of Zoology 59: 571–572.
Gibo, D.L. 1986. Flight Strategies of Migrating Monarch Butterflies (Danaus plexippus L.) in Southern Ontario. In: Danthanarayana W. (eds) Insect Flight, pp 172–184. Pro-ceedings in Life Sciences. Springer, Berlin, Heidelberg. Available from https://doi.org/10.1007/978-3-642-71155-8_12.
Gibo D.L and M.J. Pallett. 1979. Soaring flight of monarch butterflies Danaus plexippus (Lepidoptera: Danaidae) during the late summer migration in southern Ontario. Canadian Journal of Zoology 57: 1393–1401.
Guerra, P.A., Gegear, R.J., Reppert, S.M. 2014. A magnetic compass aids monarch butterfly migration. Nature Communica-tions 5:4164. Available from https://doi.org/10.1038/ncomms5164
Guilford, T. and G. K. Taylor. 2014. The sun compass revisited. Animal Behaviour 97: 135–143.
Heinze, S., and S.M. Reppert. 2011. Sun compass integration of skylight cues in migratory monarch butterflies. Neuron 69: (345–358).
Kanz J.E. 1977. The orientation of migrant and non-migrant monarch butterflies, Danaus plexippus (L.). Psyche 84(2): 120–141.
Kyriacou, C.P. 2009. Clocks, cryptochromes and monarch migrations. Journal of Bi-ology 8: #55. Available from https://doi.org/10.1186/jbiol153.
Labhart, T., Baumann, F., and G.D. Ber-nard. 2009. Specialized ommatidia of the polarization-sensitive dorsal rim area in the eye of monarch butterflies have non-functional reflecting tapeta. Cell and Tissue Research 338: #391. Available from https://doi.org/10.1007/s00441-009-0886-7.
Merlin, C., Gegear, R.J., Reppert, S.M. 2009. Antennal circadian clocks coordinate sun compass orientation in migratory monarch butterflies. Science. 325: 1700–1704.
Mouritsen, H., Derbyshire, R., Stalleicken, J., Mouritsen, O., Frost, B.J., and D.R. Norris. 2013. An experimental displace-ment and over 50 years of tag-recoveries show that monarch butterflies are not true navigators. Proceedings of the National Academy of Sciences 110 (18): 7348–53.
National Geographic. 1976. Discovering the monarch’s Mexican haven. 150 (2): 161–174.
30
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 29
Perez, S.M., O.R. Taylor, R. Jander. 1997. A sun compass in monarch butterflies. Nature 387: 29.
Preston, F. University of Kansas; unpublished correspondence. (TGLE unable to verify).
Reppert, S.M., Zhu, H., and R. White. 2004. Polarized light helps monarch butterflies navigate. Current Biology 14: 155–158.
Reppert, S.M., P.A. Guerra, and C. Merlin. 2016. Neurobiology of Monarch Butterfly Migration. Annual Review of Entomology 61: 25–42.
Stalleicken, J., Mukhida, M., Labhart, T., Wehner, R., Frost, B., and H. Mouritsen. 2005. Do monarch butterflies use polarized skylight for migratory orientation? Journal of Experimental Biology 208: 2399–2408.
Stalleicken, J., Labhart, T., and H. Mourit-sen. 2006. Physiological characterization of the compound eye in monarch butterflies
with focus on the dorsal rim area. Journal of Comparative Physiology 192 (3): 321–31.
Taylor, O.R. Lovett, J.P, Gibo, D.L., Weis-er, E.L., Thogmartin, W.E., Semmens, D.J., Diffendorfer, J.E., Pleasants, J.M., Pecoraro, S.D., and R. Grundel. 2019. Is the timing, pace, and success of the monarch migration associated with sun angle? Frontiers in Ecology and Evolution 7: 442 Available from https://doi.org/10.3389/fevo.2019.00442
Weiner, J. 1983. Field Notes, A Flight of But-terflies. The Sciences 23: 57–58.
Zhu, H., Sauman, I., Yuan, Q., Casselman, A., Emery-Le, M., Emery P., and S.M. Reppert, 2008. Cryptochromes define a novel circadian clock mechanism in mon-arch butterflies that may underlie sun compass navigation. PLos Biol. 6, e4. Avail-able from https://doi.org/10.1371/journal.pbio.0060004
31
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

30 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
The push towards agricultural inten-sification over the past several decades has resulted in simplified cropping systems and landscapes, as is highlighted by an increase in monocultures and a decrease in natural and semi-natural habitats in surrounding areas. This environmental simplification is generally found to result in compromised eco-system services, such as decreased pollina-tion, eroded soil nutrient and water supplies, and diminished pest control (Millennium Ecosystem Assessment 2005, Tscharntke et al. 2005, Foley et al. 2011). In terms of pest control, environmental simplification can decrease the abundance, diversity, or effectiveness of natural enemies of crop pests (Root 1973, Andow 1991, Perfecto et al. 2003, Bianchi et al. 2006, Vandermeer and Perfecto 2007, Letourneau et al. 2011), which may, in turn, push farmers to be more reliant on synthetic pesticides. In addition to causing environmental harm, this chemi-cal-based pest control is vulnerable to losing its effectiveness through the development of pesticide resistance. These limitations, coupled with a growing demand for organic produce and increasing costs of synthetic
inputs, have generated a large interest in controlling pests through biological control (Lewis et al. 1997, Landis et al. 2000, Simon et al. 2010).
Understanding how natural enemies are influenced by their environment is criti-cal for developing strategies to augment the effectiveness of biological control services. To date, much research has shown that hetero-geneity at both a local and landscape scale in agricultural areas commonly correlates with higher densities of natural enemies (for syntheses, see: Andow 1991, Bengtsson et al. 2005, Bianchi et al. 2006, Letourneau et al. 2011, Chaplin-Kramer et al. 2011), though a recent synthesis reveals the role of land-scape heterogeneity to be mixed (Karp et al. 2018). Despite these advances, the relative importance of local vs. landscape environ-mental heterogeneity on natural enemies is not well understood. In the current review, we address this research gap.
The importance of landscape hetero-geneity on biological control is evident in the literature, as the home range of many arthropod individuals extends far beyond
Agricultural Natural Enemies Benefit Predominantly from Broader Scales of Environmental Heterogeneity: A Quantitative Review
Aaron L. Iverson1.*, Robyn J. Burnham2, John H. Vandermeer21 Environmental Studies Department, St. Lawrence University, Canton, NY, USA
2 Department of Ecology and Evolutionary Biology, University of Michigan, USA* Corresponding author: (e-mail: [email protected]; phone: (315) 229-5970).
AbstractNaturally occurring predator and parasitoid communities are well known to respond
to multiple scales of environmental heterogeneity within and around agroecoystems, yet our understanding of which scales are most influential on different functional guilds of enemies is limited. Using vote-counting methodology, we synthesized the results from 40 empirical studies that observed how natural enemy richness, diversity, or parasitism rate is affected by environmental heterogeneity at a local scale (e.g. a focal field), an intermediate scale (e.g. habitat in immediate proximity of a focal field), and landscape scale (e.g. habitat within >200 m radius around focal field). Heterogeneity at all scales was more commonly beneficial than antagonistic to natural enemies as a whole, where positive responses were always significantly larger than negative responses. However, when using a conservative approach of comparing the positive and ‘non-positive’ (combined neutral and negative re-sponses), landscape heterogeneity was the only scale where positive responses significantly outweighed non-positive responses. The same trend held for natural enemy guilds; though all guilds had more positive than negative responses to all scales of heterogeneity, inter-mediate and landscape scales were the only scales where any guild had significantly more positive than non-positive responses. These results suggest the importance of incorporating geographically large-scale strategies when seeking to conserve natural enemies and enhance or sustain conservation biological control services.
Keywords: agroecosystem, biological control, landscape, parasitoid, predator
32
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 31
the scale of a crop field (Tscharntke et al. 2007, Rusch et al. 2010), and may reach up to several kilometers (Roschewitz et al. 2005). Studies investigating the role of landscape heterogeneity often focus on the habitat composition extending from a cou-ple hundred meters to several kilometers beyond a focal field (e.g., percent of non-crop area within a given radius of a field). Many studies highlight the importance of providing natural, semi-natural, or perennial habitat that natural enemies can inhabit when conditions in the agricultural area make survival difficult (Thies and Tscharntke 1999, Clough et al. 2005, Attwood et al. 2008, Rusch et al. 2010). These refuges—forests, hedgerows, field margins, fallows, meadows, or wetlands—may function as source habi-tats for predator or parasitoid populations and provide alternative resources (e.g. prey, pollen, nectar), permanent vegetation for reproduction and overwintering, or protec-tion during disturbances (Rusch et al. 2010, Morandin and Kremen 2013). Landscapes with more natural/semi-natural areas may also provide benefits in terms of connectivity, allowing organisms a conduit for migration (Benton et al. 2003).
Similarly, local (within-field) heteroge-neity has repeatedly been shown to positively influence the natural enemy community in agroecosystems (Andow 1991, Simon et al. 2010, Letourneau et al. 2011, Iverson et al. 2014). Studies of local heterogeneity usually compare fields of different planned (e.g., crop species) or sometimes associated diversity (e.g. weeds). Many studies compare mono-cultural to polycultural cropping systems. Two dominant theories are postulated to help explain why pest regulation in agro-ecosystems often results from higher local floristic diversity: the resource concentration hypothesis and the natural enemies hypoth-esis (Root 1973). The resource concentration hypothesis proposes that specialized herbi-vores will be better able to persist in areas where their food source is concentrated (e.g., monocultures) compared to polycultures, where they will be less efficient at locating acceptable food plants. This phenomenon results from one or more different mecha-nisms: (1) polycultures may cause chemical interference by collectively containing more plant volatiles which confuse or repel herbi-vores relying on olfactory cues in their search for host plant species; (2) herbivores may be visually confused when navigating through multiple plant species to reach their host; (3) a difference in host quality between polycul-ture and monoculture systems may result from changes in inter-plant competition; (4) the increased amount of non-host surface area in polycultures may inhibit herbivores through increasing search times for locating
host plants; and (5) abiotic factors, such as differences in shade, humidity, wind, and mid-day temperatures between the two cultural practices (Andow 1991). Although these same mechanisms could also decrease the efficiency of natural enemies, evidence suggests natural enemies may not be as inhibited, and some even have enhanced search efficiencies in polycultures (Perfecto and Vet 2003).
The natural enemies hypothesis (Root 1973) proposes that natural enemies will be present in higher numbers in more complex habitats via at least two mechanisms. First, complex habitats will likely host a greater diversity of prey due to a greater diversity of host plants and microhabitats. Second, com-plex habitats offer other food resources, such as nectar and pollen, which are especially im-portant for enemies (e.g., parasitoid wasps) whose different life stages require different foods. Both of these mechanisms result in increased temporal stability and availability of resources for the natural enemies.
The response of arthropods to envi-ronmental simplification at different scales can vary by organism, and often depends on the organism’s trophic position and dis-persal ability, which are often a function of body size (Tscharntke et al. 2005, Gabriel et al. 2010, Gonthier et al. 2014). Higher trophic-level organisms, and especially spe-cialists (e.g., many parasitoids), are often more susceptible to habitat fragmentation than herbivorous pests (Kruess and Tschar-ntke 2000, Tscharntke et al. 2005, Klein et al. 2006). Many parasitoids may also be particularly sensitive to local heterogeneity due to their often limited dispersal abilities and narrow host ranges (van Nouhuys 2005, Shaw 2006), whereas natural enemy species that have high dispersal potential, such as ballooning spiders, might be less influenced by local habitat heterogeneity and more in-fluenced by landscape heterogeneity (Clough et al. 2005, Schmidt and Tscharntke 2005). Furthermore, the influence of heterogeneity may be highly context-dependent, where the interaction between local and landscape het-erogeneity is important. For example, local heterogeneity can be more influential for organisms in simplified (e.g., high proportion of cropped area) rather than in complex land-scapes, as simple landscapes may not have as many natural enemies dispersing into farms from the surrounding landscape, and farms in these landscapes thus benefit relatively more from local management improvements (Thies and Tscharntke 1999, Tscharntke et al. 2005, Gabriel et al. 2010, Geiger et al. 2010, Batáry et al. 2011, Winqvist et al. 2011, Concepción et al. 2012, Tuck et al. 2014). Relatedly, local heterogeneity may be particularly important in landscapes of
33
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

32 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
intermediate complexity where overly sim-plified landscapes are not able to support a pool of natural enemies from which to draw (Tscharntke et al. 2005).
It is well established that environ-mental heterogeneity at local and landscape scales is important for natural enemies, yet we do not have a clear understanding of which scale may be most commonly important among natural enemy guilds. Understanding the most influential scale for different organisms is critical for farmers, land managers, conservation practitioners, and policy makers in order to prioritize the scale of management that leads to the most effective and efficient biological control. In the present review, we provide a quantitative analysis based on a vote-counting methodol-ogy of 40 studies to determine which scale of environmental heterogeneity is most influen-tial for natural enemy diversity, abundance, and parasitism rate. We also explore wheth-er the response to scale varies depending on the natural enemy functional guild.
Methods
Literature search and study se-lection: To collect relevant studies, we reviewed the first 60 results from all cross-field combinations of the following two fields in addition to the term “agroecosystem OR agriculture”: 1) local, landscape, manage-ment, intensification or scale, and 2) natural enemies, predator, parasitoid, parasitism, biodiversity, biocontrol or biological control. For example, one query included the terms “agroecosystem OR agriculture” plus one term from group 1 (e.g. “local”) plus one term from group 2 (e.g. “natural enemies”). From these search results (N = 2100), we first eliminated studies that were clearly unrelated to the topic based on the title. We then read the abstracts and, if still deemed to be relevant, the full content of the remaining studies that were selected based on their title. From these, we included only studies that consisted of field experiments or surveys that investigated how natural enemy abun-dance, richness, diversity, size/condition, or parasitism rate differed between agricultur-al areas of differing management intensities at a local, intermediate, or landscape scale (for our categorization of scales, see below). We did not constrain by biogeographic re-gion. Our search yielded a total of 40 studies (Kruess and Tscharntke 1994; Marino and Landis 1996; Murphy et al. 1996; Bommar-co 1998; Elliott et al. 1998; Murphy et al. 1998; Carmona and Landis 1999; Menalled et al. 1999; Thies and Tscharntke 1999; Kruess and Tscharntke 2000; Nicholls et al. 2001; Östman et al. 2001; Elliott et al. 2002; Armbrecht and Perfecto 2003; Harmon
et al. 2003; Kruess 2003; Menalled et al. 2003; Thies et al. 2003; Weibull et al. 2003; Costamagna et al. 2004; Pfiffner and Wyss 2004; Prasifka et al. 2004; Tylianakis et al. 2004; Bianchi et al. 2005; Clough et al. 2005; Purtauf et al. 2005a, 2005b; Roschewitz et al. 2005; Schmidt and Tscharntke 2005; Schmidt et al. 2005; Thies et al. 2005; Gi-anoli et al. 2006; Klein et al. 2006; Wilby et al. 2006; Aroga and Ambassa-Kiki 2007; Cai et al. 2007; Gardiner et al. 2009; Meyer et al. 2009; Thies and Tscharntke 2010). We conducted literature searches in Aug 2010.
Data compilation: We compiled the response of natural enemy diversity (in-cluding species richness and other diversity metrics), such as Simpson’s and Shannon’s diversity indices, abundance, and parasitism rate to three scales of environmental hetero-geneity (local, intermediate, landscape; see below). Multiple observations were possible within a given study. If a study assessed multiple metrics (e.g., both richness and abundance) for a single species or a single group, each metric was considered as a separate observation. If a study considered multiple scales, one observation (and only one) was recorded for each of the three scales per natural enemy metric. For example, if a study calculated landscape diversity at 1 km, 2 km, and 3 km radii, all of which fit into our category of ‘landscape diversity’, we distilled the information into one observation. To do so, if the response to at least one scale was positive and there were no negative respons-es, we recorded the observation as positive (and did the same for a negative response). If all responses were neutral or if there were discordant responses (positive and negative), we recorded the observation as neutral. We did the same if there were multiple mea-sures of environmental heterogeneity at the same scale (e.g. percent non-crop area and landscape habitat diversity at 1km). If studies reported natural enemy responses for individual species and for larger groupings (e.g., by guild or for all natural enemies), we used the most inclusive grouping that was presented.
We grouped observations into three distinct environmental heterogeneity scales: local, intermediate, and landscape. Lo-cal-scale heterogeneity was characterized by the within-field planned or associated diversity of plants. Most often, these studies compared monoculture to polyculture crop-ping systems, but some included within-field weed diversity (Clough et al. 2005, Purtauf et al. 2005b, Roschewitz et al. 2005). We categorized intermediate-scale heteroge-neity as structural diversity located in the immediate surroundings of a field. This cat-egory included measures such as proximity to field edges, presence of refuge strips, or
34
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 33
field perimeter-to-area ratios. For example, fields with higher perimeter-to-area ratios had relatively more field margins per unit area and were considered more complex. Landscape-scale heterogeneity pertained to regions incorporating multiple fields and/or habitat patches or beyond (minimum 200 m radius but up to 6 km radius from sampling location). More heterogeneous landscapes had a greater diversity of habitat types or a larger proportion of semi-natural, natural, or non-crop area.
We then categorized the observations according to the functional guild of the natural enemy, permitting comparisons of the relative importance of environmental heterogeneity to different functional guilds. In one analysis, we coarsely divided the observations into parasitoids or predators. In another analysis, we further divided the predators into either ground-foraging species or plant-foraging species (Table 1, Appendix 1). Although most species of natural enemies are capable of foraging on both the ground and on plants, we separated them by the habitat in which they spend the majority of their foraging time (if known), or by where they were captured in the study based on the capturing method, e.g. sweep netting vegetation vs. pitfall traps. Plant-foraging species included primarily enemies that are strong fliers (e.g., coccinellid beetles, pompilid and sphecid wasps, and most beneficial insects in the orders Neuroptera and Hemiptera) or species that are almost exclusively plant-dwelling (e.g., syrphid fly larvae). Ground-foraging species included primarily ground-foraging ants (Formici-dae), ground-foraging beetles (Carabidae and Staphylinidae), and ground-dwelling spiders (Araneae). Although they are often plant-foragers too, spiders were grouped as ground-foraging because nearly all re-searchers in our included studies collected these using pitfall traps (Harmon et al. 2003, Weibull et al. 2003, Pfiffner and Wyss 2004, Clough et al. 2005, Schmidt and Tscharntke 2005). The category of parasitoids included several families of Hymenoptera and, to a lesser extent, Diptera.
Data analysis: Each observation was recorded as positive, negative, or neutral depending on whether natural enemy di-versity, abundance, size, or parasitism rate significantly (p < 0.05) increased (positive), decreased (negative), or showed no signifi-cant effect (neutral) in the more heteroge-neous environment. Using these tallies, we calculated the effect on the natural enemy community of 1) environmental heteroge-neity (all three scales combined), 2) scale of heterogeneity (each scale considered sepa-rately), and 3) functional guild of natural enemy. Furthermore, we observed if the type
of natural enemy metric (i.e., abundance, diversity, or parasitism rate) affected these outcomes. We grouped the metrics of species richness and diversity indices under the category ‘diversity’. Only two studies com-pared the sizes or condition of the natural enemies (Bommarco 1998, Östman et al. 2001); these observations were included in the ‘abundance’ category. For all analyses, we determined whether the observed fre-quency of positive responses compared to the combined neutral and negative responses was significantly different (p < 0.05) from the expectation of a binomial distribution, where the probability of success was 0.5 for either outcome.
ResultsBenefits of environmental het-
erogeneity: Overall effect and effect by scale: Our literature search yielded 40 pertinent studies and 130 observations. Overall, we found that the number of positive responses (54.6% of observations) by natural enemies to a heterogeneous environment with all scales combined far outweighed the number of negative responses (3.8% of obser-vations; Table 1, Fig. 1). However, the num-ber of neutral responses was also relatively large (41.5% of all observations). The extent of the benefit of heterogeneity on natural enemies depended on the scale at which the heterogeneity was observed, and only at the landscape scale were the positive responses (61.9%) of natural enemies significantly larger than the combined neutral (34.9%) and negative responses (3.2%; Table 1, Fig. 1). Intermediate-scale heterogeneity still had a majority (55.6%) of observations returning a positive response, while local-scale het-erogeneity had the lowest percentage, with 38.7% of total responses being positive (and 58% neutral responses), though negative responses to local heterogeneity were still low (3%).
Effect by natural enemy guild: When we subdivided results by natural en-emy guild, we found that positive outcomes from increased heterogeneity were also al-ways much greater than negative outcomes for all guilds. However, the positive outcomes were only significantly greater than the combined neutral and negative effects for plant-foraging predators at intermediate and landscape scales and for parasitoids at a landscape scale (Table 1, Fig. 1). No guild showed a significantly positive response (compared to combined neutral and negative responses) at the local scale.
Effect by natural enemy metric: When the results were dissected according to the reporting metric (i.e., abundance, diversity, or parasitism rate), for all scales
35
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

34 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
combined and at each individual scale, again positive outcomes far outweighed negative outcomes. However, parasitism rate at a landscape scale was the only metric where positive outcomes (68.2%) were significantly higher than the combined neutral and neg-ative outcomes, although species diversity was marginally significant (p = 0.055) at a landscape scale (Table 1).
Discussion
Overall response to environmental heterogeneity: We show that the natural enemy community consistently benefits from environmental heterogeneity, both within and around agroecosystems. In all scale and natural enemy functional guild catego-ries, we observed a much greater number
Table 1. Proportion positive, neutral, and negative responses of natural enemies, in-cluding separate enemy guilds and metrics (diversity, abundance, parasitism rate), to increasing environmental heterogeneity. Positive Neutral Negative p* NAll scales combined All natural enemies combined 0.55 0.42 0.04 0.127 130Parasitoids 0.58 0.40 0.02 0.084 53Predators combined 0.52 0.43 0.05 0.324 77 Ground-foraging predators 0.38 0.55 0.08 0.923 40 Plant-foraging predators 0.67 0.30 0.03 0.018 33Species diversitya 0.57 0.37 0.07 0.181 30Species abundance 0.52 0.44 0.05 0.356 66Parasitism rate 0.59 0.41 0.00 0.115 34 Landscape scaleb All natural enemies combined 0.62 0.35 0.03 0.021 63Parasitoids 0.66 0.34 0.00 0.031 29Predators combined 0.59 0.35 0.06 0.115 34 Ground-foraging predators 0.47 0.47 0.07 0.500 15 Plant-foraging predators 0.72 0.22 0.06 0.015 18Species diversitya 0.70 0.20 0.10 0.055 10Species abundance 0.55 0.42 0.03 0.237 31Parasitism rate 0.68 0.32 0.00 0.026 22 Intermediate scale All natural enemies combined 0.56 0.39 0.06 0.203 36Parasitoids 0.50 0.43 0.07 0.395 14Predators combined 0.59 0.36 0.05 0.143 22 Ground-foraging predators 0.42 0.50 0.08 0.613 12 Plant-foraging predators 0.80 0.20 0.00 0.011 10Species diversitya 0.60 0.30 0.10 0.172 10Species abundance 0.58 0.37 0.05 0.180 19Parasitism rate 0.43 0.57 0.00 0.500 7 Local scale All natural enemies combined 0.39 0.58 0.03 0.859 31Parasitoids 0.50 0.50 0.00 0.377 10Predators combined 0.33 0.62 0.05 0.905 21 Ground-foraging predators 0.23 0.69 0.08 0.954 13 Plant-foraging predators 0.20 0.80 0.00 0.813 5Species diversitya 0.40 0.60 0.00 0.623 10Species abundance 0.38 0.56 0.06 0.773 16Parasitism rate 0.40 0.60 0.00 0.500 5* Bold numbers indicate values where the frequency of positive responses compared to the combined neutral and negative responses was significantly different from a binomial distribution (p < 0.05).aDiversity metric includes species richness and other diversity measures (e.g., Simpson’s and Shan-non’s diversity indices).bFor a definition of scales (landscape, intermediate, local), see ‘Methods’.
36
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 35
of positive than negative responses, where positive responses on average outweighed negative responses by a factor of 14. These results corroborate the growing body of evidence showing beneficial responses of natural enemy communities to environ-mental heterogeneity at local or landscape scales (Andow 1991, Langellotto and Denno 2004, Bianchi et al. 2006, Poveda et al. 2008, Simon et al. 2010 Chaplin-Kramer et al. 2011, Letourneau et al. 2011, Gonthier et al. 2014). Despite a clear pattern of positive responses outweighing negative responses, we also observed many neutral impacts of heterogeneity on the natural enemy commu-nity, corroborating other studies which show mixed results and highlighting the ecologi-cal complexities associated with assessing impacts of heterogeneity (Karp et al. 2018). Our results help to disentangle the varied responses of environmental heterogeneity by separately assessing the impact of three different scales of heterogeneity on different guilds of natural enemies.
Scales of heterogeneity: Although positive responses outweighed negative responses at all scales, beneficial effects were especially pronounced at landscape scales. These results suggest that broad-scale heterogeneity is extremely valuable
in maintaining natural enemy populations, supporting the idea that many arthropod species interact with their environment at a larger-than-local level (Thies and Tschar-ntke 1999), with important implications regarding regional planning and manage-ment processes. These results also provide support for a density-mediated mechanism (the enemies hypothesis), i.e., a top-down mechanism of herbivore control (Hairston et al. 1960). Here, natural enemies likely benefit from increased food and habitat re-sources in non-crop areas surrounding farm fields. However, trait-mediated effects (the resource concentration hypothesis) often function simultaneously and complementari-ly to the enemies hypothesis, and may still be an important factor in many of these studies.
Although the effects of local hetero-geneity can often be as important as, or more important than, effects of landscape heterogeneity in agroecosystems (Puech et al. 2014), our results are consistent with the findings of other vote-count studies that have focused on single scales, where higher positive responses of natural enemies to environmental heterogeneity appear to be found at landscape scales. For instance, Bianchi et al. (2006) found that 74% of their observations showed a positive response to
Figure 1. Percent of all observations returning a positive response of natural enemy abundance or diversity as a result of environmental heterogeneity at local, intermediate, and landscape scales, as well as at all scales combined. *Indicates cases where the frequency of positive responses compared to the combined neutral and negative responses was significantly different from a binomial distribution (P < 0.05).
37
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

36 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
landscape heterogeneity, whereas studies of local heterogeneity showed (surprisingly consistent) positive responses barely to ex-ceed 50% of all responses for natural enemies (52.7% in Andow 1991; 52% in Poveda et al. 2008; 53.3% in Simon et al. 2010). Our results are also consistent with recently reported declines in arthropod biodiversity in natural areas, where intensified land use (agricultural cover) at a landscape-scale, more so than local-scale variables, was a primary driver of declines in species abun-dance and richness in grasslands (Seibold et al. 2019).
Arthropod guilds and environ-mental heterogeneity: The positive responses to increased broad-scale het-erogeneity observed in the parasitoid and especially plant-foraging predator commu-nities may reflect their particular sensitiv-ity to environmental disturbance at these scales. Parasitoids and many plant-foraging predators, such as syrphid flies, predatory wasps, and some predatory beetles, are reliant on alternative food sources, such as pollen and nectar, at some point in their life cycles (Langellotto and Denno 2004). Although some crops or weeds within crops may provide these resources, they are often most abundant in non-crop areas. Further-more, the small size and high prevalence of prey specialization in parasitoids may also contribute to lower dispersal abilities and increased sensitivity to a simplified environ-ment (Roland and Taylor 1997).
On the other hand, ground-foraging predators, such as ground spiders and ca-rabid beetles, may be relatively less reliant on resources in non-crop habitat and more sensitive to soil management practices, such as tilling (Sharley et al. 2008), which could mask any differences in vegetation diversi-ty or structure. Many of these species rely on stable soil habitats for protection (e.g., overwintering) or for oviposition (Rusch et al. 2010). For instance, Langellotto and Denno (2004) observed a large impact, especially on spiders, from enhancing the structural complexity of soil detritus. Other studies have shown that structural diver-sity, rather than plant species diversity, in the landscape physically inhibits carabid movement between fields (Frampton et al. 1995, Mauremooto et al. 1995). Additionally, some spiders are able to avoid size-dispersal limitations through long-distance windborne dispersal (ballooning), which may allow them to be less affected by intensively managed landscapes (Weyman et al. 2002).
When the metrics of parasitism rate, abundance, and diversity were considered independently, parasitism rate at a land-scape scale was the only category where
observations of enemies benefitting from heterogeneity were significantly more fre-quent than the combined neutral and nega-tive observations. These results may again reflect how the often small and specialized parasitoids may be particularly sensitive to environmental disturbance (see above).
Management implications: Our findings support the value of increasing environmental heterogeneity for promoting natural enemy presence in agroecosystems. Our results suggest that farmers can en-hance their local natural enemy community through increases in within-field diversity (e.g., polycultures), but especially through improvements in broader-scale, and especial-ly landscape-scale, heterogeneity. Although landscape-scale heterogeneity will often, but not always, apply beyond the scope of an individual farm, intermediate-scale en-hancement may be provided, for example, by increasing the size of vegetated (not bare) field margins, decreasing the size of fields, or including vegetation strips (e.g., floral strips, beetle banks) within fields. These vegetation strips may be especially effective if specific plants that provide resources for natural enemies, but do not simultaneously attract pests, are included (Pfiffner and Wyss 2004). Intentional set-aside conservation areas around crop fields need not trade-off with crop productivity, where benefits to yield from biocontrol may outweigh small losses in cultivated area (Pywell et al. 2015). Furthermore, vegetation strips or weedy field margins offer additional benefits, such as habitat for biodiversity, including polli-nators, and erosion control (Wratten et al. 2012, Morandin and Kremen 2013).
The benefit of landscape-scale het-erogeneity emphasizes the importance of region-wide land management or collective, community-scale initiatives. Currently, many management suggestions that seek to enhance biological control have focused solely on increasing local diversity (Gurr et al. 2000). We suggest, therefore, that govern-ment agencies and organizations should not only encourage farmers to make local-scale changes for biodiversity enhancement but should provide incentives for individual landholders and communities to make landscape-level management decisions that will positively impact biodiversity. Govern-ment-supported or certification-based (sensu Tscharntke et al. 2014) economic incentives could play an important role in promoting landscape heterogeneity. Although planning at large geographic scales is challenging, additive effects are common, where the land-use changes of individual farmers scale up to landscape-level effects (Holzschuh et al. 2008, Gabriel et al. 2010).
38
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 37
Our results suggest the importance of focusing management strategies on a particular pest or group of pests, as we show that the scale of management can have differential effects based on the natural enemy guild. For example, if the dominant pest spends all or part of its life cycle in the soil, ground-foraging predators, such as ground spiders and carabid beetles, may be the most effective biocontrol agents. As we show that these predators generally respond less to environmental heterogeneity and likely more to direct soil management, it may be important to vary a farm’s cultural techniques to optimize the survival and growth of the predators. However, caution must be exercised to avoid a “one-problem, one-species” approach, as it is clear that the consortium of natural enemies is important for biological control given the inherent complexities of food webs even in simplified agroecosystems (Altieri 1999, Tscharntke et al. 2007, Vandermeer et al. 2010).
Limitations and further research: Enhancements in the natural enemy com-munity may not necessarily translate into enhanced crop health (Symondson et al. 2002). However, although we did not include the effect of environmental heterogeneity on direct biocontrol, crop yield, or pest abun-dance, other studies have shown that natural enemy species often respond more strongly to heterogeneity than do pest species (Lang-ellotto and Denno 2004, Chaplin-Kramer et al. 2011). Similarly, Risch et al. (1983) showed in a review of 150 studies that her-bivores respond in an opposite manner to local heterogeneity, where 53% of herbivore species were significantly less abundant on more diverse farms. These studies suggest that an improved natural enemy community often translates into improved crop health.
Further research is needed to clarify the complex ecological interactions that underpin effective biocontrol and the influ-ence of spatial scale on these interactions. A growing number of studies highlight indirect effects, both density- and trait-mediated (Werner and Peacor 2003), and the potential effects of land management, as well as in-trinsic (self-organized) factors, in structuring them (Vandermeer and Perfecto 2008, Hsieh et al. 2012, Liere et al. 2014). Furthermore, the high proportion of neutral responses of natural enemies to heterogeneity may reflect the large variance in response, indicating that we may not be considering the appropri-ate metrics for understanding natural enemy distributions. The relatively high occurrence of neutral responses may also be reflective of the vote-counting method we employed, which is not sensitive to subtle effects.
Although mechanisms of local-level influences on natural enemies are better understood, mechanisms at a landscape scale are less clear, undoubtedly due to the difficulty of landscape-scale manipulation or comparison. To address this gap, we en-courage research that characterizes the land-scape in terms of traits or resources, rather than relying solely on coarse landscape met-rics (e.g., non-crop area) (sensu Schellhorn et al. 2015). Additionally, research on bio-control should report the dispersal abilities, if known, of each of the organisms studied, allowing for a clearer consensus on the role of dispersal in an organism’s sensitivity to environmental heterogeneity.
ConclusionsThe simplification of agricultural lands
threatens the health of many ecosystems worldwide, impacting both humans and the biodiversity on which we depend (IAASTD 2008, IPBES 2019). It is therefore critical to understand how we can implement agro-ecosystems that provide important services, such as pest control, with an eye on reducing reliance on pesticides and other practices that simplify rather than diversify agricul-ture. Boosting natural enemy populations through habitat enhancement is one way to achieve this goal. Our results suggest that environmental heterogeneity, especially at broader (i.e., intermediate and particularly landscape) scales, is important for increas-ing the diversity and abundance of natural enemies. These findings highlight the impor-tance of not only individual landowners, but also collective land management practices, in maximizing potential biocontrol services from natural enemies.
AcknowledmentsWe thank Louis Iverson for providing
useful comments on this paper and the Uni-versity of Michigan Department of Ecology and Evolutionary Biology for funding.
Literature CitedAltieri, M. A. 1999. The ecological role of bio-
diversity in agroecosystems. Agriculture, Ecosystems & Environment. 74: 19–31.
Andow, D. A. 1991. Vegetational diversity and arthropod population response. Annual Re-view of Entomology. 36: 561–86.
Armbrecht, I., and I. Perfecto. 2003. Lit-ter-twig dwelling ant species richness and predation potential within a forest frag-ment and neighboring coffee plantations of contrasting habitat quality in Mexico. Agriculture Ecosystems and Environment. 97: 107–115.
39
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

38 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Aroga, R., and R. Ambassa-Kiki. 2007. Effects of maize–cowpea intercropping on maize stem borers and their natural enemies, pp. 314–328. In Badu-Apraku, B., Fakorede, M., Lum, A., Menkir, A., Ouedraogo, M. (eds.), Demand-Driven Technol. Sustain. Maize Prod. West Cent. Africa. International Insti-tute of Tropical Agriculture, Ibadan, Nigeria.
Attwood, S. J., M. Maron, A. P. N. House, and C. Zammit. 2008. Do arthropod assemblag-es display globally consistent responses to intensified agricultural land use and man-agement? Global Ecology and Biogeography. 17: 585–599.
Batáry, P., A. Báldi, D. Kleijn, and T. Tscharntke. 2011. Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proceedings of the Royal Society B. 278: 1894–902.
Bengtsson, J., J. Ahnstrom, and A.-C. Weibull. 2005. The effects of organic agriculture on biodiversity and abundance: a meta-analysis. Journal of Applied Ecology. 42: 261–269.
Benton, T. G., J. A. Vickery, and J. D. Wilson. 2003. Farmland biodiversity: is habitat het-erogeneity the key? Trends in Ecology and Evolution. 18: 182–188.
Bianchi F.J., W.K.R.E. van Wingerden, A.J. Griffioen, M. van der Veen, M.J.J. van der Straten, R.M.A. Wegman, and H.A.M. Meeuwsen. 2005. Landscape factors affect-ing the control of Mamestra brassicae by nat-ural enemies in Brussels sprout. Agriculture Ecosystems and Environment. 107:145–150.
Bianchi, F. J. J. A., C. J. H. Booij, and T. Tscharntke. 2006. Sustainable pest regu-lation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proceedings of the Royal Society B. 273: 1715–27.
Bommarco, R. 1998. Reproduction and energy reserves of a predatory carabid beetle rela-tive to agroecosystem complexity. Ecological Applications. 8: 846–853.
Cai, H.-J., Z.-S. Li, and M.-S. You. 2007. Im-pact of habitat diversification on arthropod communities: A study in the fields of Chinese cabbage, Brassica chinensis. Insect Science. 14: 241–249.
Carmona, D. M., and D. A. Landis. 1999. In-fluence of refuge habitats and cover crops on seasonal activity- density of ground beetles (Coleoptera : Carabidae) in field crops. Bio-logical Control. 28: 1145–1153.
Chaplin-Kramer, R., M. E. O’Rourke, E. J. Blitzer, and C. Kremen. 2011. A me-ta-analysis of crop pest and natural enemy response to landscape complexity. Ecology Letters. 922–932.
Clough, Y., A. Kruess, D. Kleijn, and T. Tscharntke. 2005. Spider diversity in cereal fields: comparing factors at local, landscape and regional scales. Journal of Biogeography. 32: 2007–2014.
Concepción, E. D., M. Díaz, D. Kleijn, A. Bál-di, P. Batáry, Y. Clough, D. Gabriel, F. Herzog, A. Holzschuh, E. Knop, E. J. P. Marshall, T. Tscharntke, and J. Verhulst. 2012. Interactive effects of landscape context constrain the effectiveness of local agri-envi-ronmental management. Journal of Applied Ecology. 695–705.
Costamagna, A. C., F. D. Menalled, and D. A. Landis. 2004. Host density influences parasitism of the armyworm Pseudaletia unipuncta in agricultural landscapes. Basic and Applied Ecology. 5: 347–355.
Elliott, N. C., R. W. Kieckhefer, J. Lee, and B. W. French. 1998. Influence of within-field and landscape factors on aphid predator populations in wheat. Landscape Ecology. 14: 239–252.
Elliott, N. C., R. W. Kieckhefer, and G. J. Mi-chels. 2002. Predator abundance in alfalfa fields in relation to aphids, within-field vege-tation, and landscape matrix. Environmental Entomology. 31: 253–260.
Foley, J. A., N. Ramankutty, K. A. Brauman, E. S. Cassidy, J. S. Gerber, M. Johnston, N. D. Mueller, C. O’Connell, D. K. Ray, P. C. West, C. Balzer, E. M. Bennett, S. R. Carpenter, J. Hill, C. Monfreda, S. Polasky, J. Rockstrom, J. Sheehan, S. Siebert, D. Tilman, and D. P. M. Zaks. 2011. Solutions for a cultivated planet. Na-ture. 478: 337–342.
Frampton, G., T. Cilgi, G. Fry, and S. D. Wratten. 1995. Effects of grassy banks on the dispersal of some carabid beetles (Cole-optera: Carabidae) on farmland. Biological Conservation. 71: 347–355.
Gabriel, D., S. M. Sait, J. a Hodgson, U. Schmutz, W. E. Kunin, and T. G. Benton. 2010. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecology Letters. 13: 858–69.
Gardiner, M., D. Landis, C. Gratton, C. Di-Fonzo, M. O’Neal, J. Chacon, M. Wayo, N. Schmidt, E. Mueller, and G. Heimpel. 2009. Landscape diversity enhances biolog-ical control of an introduced crop pest in the north-central USA. Ecological Applications. 19: 143–154.
Geiger, F., G. R. de Snoo, F. Berendse, I. Guerrero, M. B. Morales, J. J. Oñate, S. Eggers, T. Pärt, R. Bommarco, and J. Bengtsson. 2010. Landscape composition influences farm management effects on farmland birds in winter: A pan-European
40
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 39
approach. Agriculture, Ecosystems & Envi-ronment. 139: 571–577.
Gianoli, E., I. Ramos, A. Alfaro-Tapia, Y. Valdéz, E. R. Echegaray, and E. Yábar. 2006. Benefits of a maize–bean–weeds mixed cropping system in Urubamba Valley, Peru-vian Andes. International Journal of Pest Management. 52: 283–289.
Gonthier, D. J., K. K. Ennis, S. Farinas, H.-Y. Hsieh, A. L. Iverson, P. Batáry, J. Ru-dolphi, T. Tscharntke, B. J. Cardinale, and I. Perfecto. 2014. Biodiversity conser-vation in agriculture requires a multi-scale approach. Proceedings of the Royal Society B. 281: 20141358.
Gurr, G. M., S. D. Wratten, and P. Barbosa. 2000. Success in conservation biological control of arthropods, pp. 105–132. In Gurr, G.M., Wratten, S.D. (eds.), Biological Control Measures of Success. Kluwer Academic Pub-lishers, Dordrecht, The Netherlands.
Hairston, N. G., F. E. Smith, and L. B. Slobod-kin. 1960. Community structure, population control, and competition. American Natural-ist. 94: 421–425.
Harmon, J. P., E. E. Hladilek, J. L. Hinton, T. J. Stodola, and D. a. Andow. 2003. Herbivore response to vegetational diversity: spatial interaction of resources and natural enemies. Population Ecology. 45: 75–81.
Holzschuh, A., I. Steffan-Dewenter, and T. Tscharntke. 2008. Agricultural landscapes with organic crops support higher pollinator diversity. Oikos. 117: 354–361.
Hsieh, H.-Y., H. Liere, E. J. Soto, and I. Per-fecto. 2012. Cascading trait-mediated inter-actions induced by ant pheromones. Ecology and Evolution. 2: 2181–91.
IAASTD. 2008. Agriculture at a Crossroads. Island Press, Washington, D.C.
IPBES. 2019. Global assessment report on bio-diversity and ecosystem services of the In-terngovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. IPBES Secretariat, Bonn, Germany.
Iverson, A. L., L. E. Marín, K. K. Ennis, D. J. Gonthier, B. T. Connor-Barrie, J. L. Remfert, B. J. Cardinale, and I. Perfecto. 2014. Do polycultures promote win-wins or trade-offs in agricultural ecosystem services? A meta-analysis. Journal of Applied Ecology. 51: 1593–1602.
Karp, D. S., R. Chaplin-Kramer, T. D. Mee-han, E. A. Martin, F. DeClerck, H. Grab, C. Gratton, L. Hunt, A. E. Larsen, A. Martínez-Salinas, M. E. O’Rourke, A. Rusch, K. Poveda, M. Jonsson, J. A. Ro-senheim, N. A. Schellhorn, T. Tscharnt-ke, S. D. Wratten, W. Zhang, A. L. Iverson, L. S. Adler, M. Albrecht, A. Alignier, G. M.
Angelella, M. Zubair Anjum, J. Avelino, P. Batáry, J. M. Baveco, F. J. J. A. Bian-chi, K. Birkhofer, E. W. Bohnenblust, R. Bommarco, M. J. Brewer, B. Caballe-ro-López, Y. Carrière, L. G. Carvalheiro, L. Cayuela, M. Centrella, A. Ćetković, D. C. Henri, A. Chabert, A. C. Costamagna, A. De la Mora, J. de Kraker, N. Desneux, E. Diehl, T. Diekötter, C. F. Dormann, J. O. Eckberg, M. H. Entling, D. Fiedler, P. Franck, F. J. Frank van Veen, T. Frank, V. Gagic, M. P. D. Garratt, A. Getachew, D. J. Gonthier, P. B. Goodell, I. Graziosi, R. L. Groves, G. M. Gurr, Z. Hajian-Fo-rooshani, G. E. Heimpel, J. D. Herrmann, A. S. Huseth, D. J. Inclán, A. J. Ingrao, P. Iv, K. Jacot, G. A. Johnson, L. Jones, M. Kaiser, J. M. Kaser, T. Keasar, T. N. Kim, M. Kishinevsky, D. A. Landis, B. Lavandero, C. Lavigne, A. Le Ralec, D. Lemessa, D. K. Letourneau, H. Liere, Y. Lu, Y. Lubin, T. Luttermoser, B. Maas, K. Mace, F. Madeira, V. Mader, A. M. Cortesero, L. Marini, E. Martinez, H. M. Martinson, P. Menozzi, M. G. E. Mitch-ell, T. Miyashita, G. A. R. Molina, M. A. Molina-Montenegro, M. E. O’Neal, I. Opatovsky, S. Ortiz-Martinez, M. Nash, Ö. Östman, A. Ouin, D. Pak, D. Paredes, S. Parsa, H. Parry, R. Perez-Alvarez, D. J. Perović, J. A. Peterson, S. Petit, S. M. Philpott, M. Plantegenest, M. Plećaš, T. Pluess, X. Pons, S. G. Potts, R. F. Pywell, D. W. Ragsdale, T. A. Rand, L. Raymond, B. Ricci, C. Sargent, J.-P. Sar-thou, J. Saulais, J. Schäckermann, N. P. Schmidt, G. Schneider, C. Schüepp, F. S. Sivakoff, H. G. Smith, K. Stack Whitney, S. Stutz, Z. Szendrei, M. B. Takada, H. Taki, G. Tamburini, L. J. Thomson, Y. Tricault, N. Tsafack, M. Tschumi, M. Valantin-Morison, M. Van Trinh, W. van der Werf, K. T. Vierling, B. P. Werling, J. B. Wickens, V. J. Wickens, B. A. Wood-cock, K. Wyckhuys, H. Xiao, M. Yasuda, A. Yoshioka, and Y. Zou. 2018. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Pro-ceedings of the National Academy of Sciences. 115: E7863–E7870.
Klein, A. M., I. Steffan-Dewenter, and T. Tscharntke. 2006. Rain forest promotes tro-phic interactions and diversity of trap-nest-ing hymenoptera in adjacent agroforestry. Journal of Animal Ecology. 75: 315–323.
Kruess, A. 2003. Effects of landscape structure and habitat type on a plant-herbivore-para-sitoid community. Ecography. 26: 283–290.
Kruess, A., and T. Tscharntke. 1994. Habitat fragmentation, species loss, and biological control. Science (80-. ). 264: 1581–4.
Kruess, A. and T. Tscharntke. 2000. Species richness and parasitism in a fragmented
41
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

40 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
landscape : Experiments and field studies with insects on Vicia sepium. Oecologia 122: 129–37.
Landis, D. A., S. D. Wratten, and G. M. Gurr. 2000. Habitat management to conserve natural enemies of arthropod pests in agri-culture. Annual Review of Entomology. 45: 175–201.
Langellotto, G. A., and R. F. Denno. 2004. Responses of invertebrate natural enemies to complex-structured habitats: a meta-analyt-ical synthesis. Oecologia. 139: 1–10.
Letourneau, D. K., I. Armbrecht, B. S. Rivera, J. M. Lerma, E. J. Carmona, M. C. Daza, S. Escobar, V. Galindo, C. Gutiérrez, S. D. López, J. L. Mejía, A. M. A. Rangel, J. H. Rangel, L. Rivera, C. A. Saavedra, A. M. Torres, and A. R. Trujillo. 2011. Does plant diversity benefit agroecosystems? A synthetic review. Ecological Applications. 21: 9–21.
Lewis, W. J., J. C. Van Lenteren, S. C. Phatak, and J. H. Tumlinson. 1997. A total system approach to sustainable pest management. Proceedings of the National Academy of Sciences. 94: 12243–12248.
Liere, H., I. Perfecto, and J. Vandermeer. 2014. Stage-dependent responses to emer-gent habitat heterogeneity: consequences for a predatory insect population in a coffee agroecosystem. Ecology and Evolution. 4: 3201–3209.
Lundgren, J.G., K. Wyckhuys, and N. Desneux. 2009. Population responses by Orius insidiosus to vegetational diversity. BioControl. 54(1): 135-142.
Marino, P. C., and D. A. Landis. 1996. Effect of landscape structure on parasitoid diversity and parasitism in agroecosystems. Ecological Applications. 6: 276–284.
Mauremooto, J. R., S. D. Wratten, S. P. Worner, and G. L. A. Fry. 1995. Permeabil-ity of hedgerows to predatory carabid beetles. Agriculture, Ecosystems & Environment. 52: 141–148.
Menalled, F. D., A. C. Costamagna, P. C. Marino, and D. A. Landis. 2003. Temporal variation in the response of parasitoids to agricultural landscape structure. Agri-culture, Ecosystems & Environment. 96: 29–35.
Menalled, F. D., P. C. Marino, S. H. Gage, and D. A. Landis. 1999. Does agricultural landscape structure affect parasitism and parasitoid diversity? Ecological Applications. 9: 634–641.
Meyer, B., F. Jauker, and I. Steffan-De-wenter. 2009. Contrasting resource-de-pendent responses of hoverfly richness and
density to landscape structure. Basic and Applied Ecology. 10: 178–186.
Millennium Ecosystem Assessment. 2005. Ecosystems and Human Well-Being: Health Synthesis. Washington, D.C.: Island Press.
Morandin, L. and C. Kremen. 2013. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecological Applications 23(4): 829–39.
Murphy, B. C., J. A. Rosenheim, and J. Granett. 1996. Habitat diversification for improving biological control: abundance of Anagrus epos (Hymenoptera: Mymaridae) in grape vineyards. Environmental Entomology. 25: 495–504.
Murphy, B., Rosenheim, J., Dowell, R., and J. Granett. 1998. Habitat diversification tactic for improving biological control: Parasitism of the western grape leafhopper. Entomologia Experimentalis et Applicata 87(3): 225–35.
Nicholls, C. I., M. Parrella, and M. A. Altieri. 2001. The effects of a vegetational corridor on the abundance and dispersal of insect biodi-versity within a northern California organic vineyard. Landscape Ecology. 16: 133–146.
Östman Ö, B. Ekbom, J. Bengtsson, and A. Weibull. 2001. Landscape complexity and farming practice influence the condition of polyphagous carabid beetles. Ecological Applications. 11: 480–488.
Perfecto, I., A. Mas, T. Dietsch, and J. Vander-meer. 2003. Conservation of biodiversity in coffee agroecosystems: a tri-taxa comparison in southern Mexico. Biodiversity and Conser-vation. 12: 1239–1252.
Perfecto, I., and L. E. M. Vet. 2003. Effect of a nonhost plant on the location behavior of two parasitoids: The tritrophic system of Cotesia spp. (Hymenoptera: Braconidae), Pieris rapae (Lepidoptera: Pieridae), and Brassica oleraceae. Environmental Ento-mology. 32: 163–174.
Pfiffner, L., and E. Wyss. 2004. Use of sown wildflower strips to enhance natural enemies of agriculture pests, pp. 165–186. In Gurr, G.M., Wratten, S.D., Altieri, M.A. (eds.), Ecological Engineering for Pest Management. CSIRO Publishing, Collingwood, Austrailia.
Poveda, K., M. I. Gomez, and E. Martinez. 2008. Diversification practices: their effect on pest regulation and production. Revista Colombiana de Entomologia. 34: 131–144.
Prasifka, J. R., K. M. Heinz, and R. R. Minzen-mayer. 2004. Relationships of landscape, prey and agronomic variables to the abun-dance of generalist predators in cotton (Gos-sypium hirsutum) fields. Landscape Ecology. 19: 709–717.
Puech, C., S. Poggi, J. Baudry, and S. Aviron. 2014. Do farming practices affect natural
42
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 41
enemies at the landscape scale? Landscape Ecology. 125–140.
Purtauf, T., J. Dauber, and V. Wolters. 2005a. The response of carabids to landscape sim-plification differs between trophic groups. Oecologia. 142: 458–464.
Purtauf, T., I. Roschewitz, J. Dauber, C. Thies, T. Tscharntke, and V. Wolters. 2005b. Landscape context of organic and conventional farms: Influences on carabid beetle diversity. Agriculture, Ecosystems & Environment. 108: 165–174.
Pywell, R. F., M. S. Heard, B. A. Woodcock, S. Hinsley, L. Ridding, M. Nowakowski, and J. M. Bullock. 2015. Wildlife-friendly farming increases crop yield: Evidence for ecological intensification. Proceedings of the Royal Society B. 282.
Risch, S. J., D. A. Andow, and M. A. Altieri. 1983. Agroecosystem diversity and pest control: Data, tentative conclusions, and new research directions. Environmental Entomol-ogy. 12: 625–629.
Roland, J., and P. D. Taylor. 1997. Insect parasitoid species respond to forest struc-ture at different spatial scales. Nature. 386: 710–713.
Root, R. 1973. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecological Monographs. 43: 95–124.
Roschewitz, I., M. Hücker, T. Tscharntke, and C. Thies. 2005. The influence of landscape context and farming practices on parasitism of cereal aphids. Agriculture, Ecosystems & Environment. 108: 218–227.
Rusch, A., M. Valantin-Morison, J. P. Sar-thou, and J. Roger-estrade. 2010. Biocon-trol-based integrated management of oilseed rape pests. Springer Netherlands, Dordrecht.
Schellhorn, N. A., V. Gagic, and R. Bommarco. 2015. Time will tell: Resource continuity bol-sters ecosystem services. Trends in Ecology and Evolution. 30: 524–530.
Schmidt, M. H., I. Roschewitz, C. Thies, and T. Tscharntke. 2005. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. Journal of Applied Ecology. 42: 281–287.
Schmidt, M. H., and T. Tscharntke. 2005. Landscape context of sheetweb spider (Ara-neae: Linyphiidae) abundance in cereal fields. Journal of Biogeography. 32: 467–473.
Seibold, S., M. M. Gossner, N. K. Simons, N. Blüthgen, J. Müller, D. Ambarlı, C. Am-mer, J. Bauhus, M. Fischer, J. C. Habel, K. E. Linsenmair, T. Nauss, C. Penone, D. Prati, P. Schall, E. D. Schulze, J. Vogt, S. Wöllauer, and W. W. Weisser. 2019.
Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature. 574: 671–674.
Sharley, D. J., A. A. Hoffmann, and L. J. Thomson. 2008. The effects of soil tillage on beneficial invertebrates within the vine-yard. Agricultural and Forest Entomology. 10: 233–243.
Shaw, M. R. 2006. Habitat considerations for parasitic wasps (Hymenoptera). Journal of Insect Conservation. 10: 117–127.
Simon, S., Bouvier, J., Debras, J., and B. Sauphanor. 2010. Biodiversity and pest management in orchard systems: A review. Agronomy for Sustainable Development. 30(1): 139–52.
Symondson, W. O. C., K. D. Sunderland, and M. H. Greenstone. 2002. Can generalist predators be effective biocontrol agents? Annual Review of Entomology. 47: 561–594.
Thies, C., I. Roschewitz, and T. Tscharnt-ke. 2005. The landscape context of cereal aphid-parasitoid interactions. Proceedings of the Royal Society B. 272: 203–10.
Thies, C., I. Steffan-dewenter, and T. Tschar-ntke. 2003. Effects of landscape context on herbivory and parasitism at different spatial scales. Oikos. 101: 18–25.
Thies, C., and T. Tscharntke. 1999. Landscape structure and biological control in agroecosys-tems. Science (80). 285: 893–895.
Thies, C., and T. Tscharntke. 2010. Biological Rape Pest Control in Spatio-Temporally Changing Landscapes, pp. 273–284. In Wil-liams, I.H. (ed.), Biocontrol-Based Integrated Management of Oilseed Rape Pests. Springer Netherlands, Dordrecht.
Tscharntke, T., R. Bommarco, Y. Clough, T. O. Crist, D. Kleijn, T. a. Rand, J. M. Ty-lianakis, S. Van Nouhuys, and S. Vidal. 2007. Conservation biological control and enemy diversity on a landscape scale. Biol. Control. 43: 294–309.
Tscharntke, T., A. M. Klein, A. Kruess, I. Steffan-Dewenter, and C. Thies. 2005. Landscape perspectives on agricultural intensification and biodiversity - ecosystem service management. Ecology Letters. 8: 857–874.
Tscharntke, T., J. C. Milder, G. Schroth, Y. Clough, F. DeClerck, A. Waldron, R. Rice, and J. Ghazoul. 2014. Conserving biodiversity through certification of tropical agroforestry crops at local and landscape scales. Conservation Letters. 1–10.
Tuck, S. L., C. Winqvist, F. Mota, J. Ahnström, L. a. Turnbull, and J. Bengtsson. 2014. Land-use intensity and the effects of organ-ic farming on biodiversity: a hierarchical
43
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

42 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
meta-analysis. Journal of Applied Ecology. 51: 746–755.
Tylianakis, J. M., R. K. Didham, and S. D. Wratten. 2004. Improved fitness of aphid parasitoids receiving resource subsidies. Ecology. 85: 658–666.
Vandermeer, J., and I. Perfecto. 2007. The agricultural matrix and a future paradigm for conservation. Conservation Biology. 21: 274–277.
Vandermeer, J., and I. Perfecto. 2008. Spatial pattern and ecological process in the coffee agroforestry system. Ecology. 89: 915–920.
Vandermeer, J., I. Perfecto, and S. Philpott. 2010. Ecological complexity and pest control in organic coffee production: uncovering an autonomous ecosystem service. Bioscience. 60: 527–537.
van Nouhuys, S. 2005. Effects of habitat frag-mentation at different trophic levels in insect communities. Annales Zoologici Fennici. 42: 433–447.
Weibull, A.-C., Ö. Östman, and A. Granqvist. 2003. Species richness in agroecosystems: the effect of landscape, habitat and farm management. Biodivers. Conserv. 12: 1335–1355.
Werner, E. E., and S. D. Peacor. 2003. A review of trait-mediated indirect interactions in eco-logical communities. Ecology. 84: 1083–1100.
Weyman, G., K. Sunderland, and P. Jepson. 2002. A review of the evolution and mech-anisms of ballooning by spiders inhabiting arable farmland. Ethology, Ecology & Evo-lution. 14: 306–326.
Wilby, A., L. P. Lan, K. L. Heong, N. P. D. Huyen, N. H. Quang, N. V. Minh, and M. B. Thomas. 2006. Arthropod diversity and community structure in relation to land use in the Mekong Delta, Vietnam. Ecosystems. 9: 538–549.
Winqvist, C., J. Bengtsson, T. Aavik, F. Ber-endse, L. W. Clement, S. Eggers, C. Fisch-er, A. Flohre, F. Geiger, J. Liira, T. Pärt, C. Thies, T. Tscharntke, W. W. Weisser, and R. Bommarco. 2011. Mixed effects of organic farming and landscape complexity on farmland biodiversity and biological control potential across Europe. Journal of Applied Ecology. 48: 570–579.
Wratten, S. D., M. Gillespie, A. Decourtye, E. Mader, and N. Desneux. 2012. Pollinator habitat enhancement: Benefits to other eco-system services. Agriculture, Ecosystems & Environment. 159: 112–122.
44
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 43A
ppen
dix
1. S
umm
ary
of a
rtic
les
incl
uded
in a
naly
sis,
sho
win
g ca
tego
ries
of n
atur
al e
nem
y gu
ild, n
atur
al e
nem
y cl
assi
ficat
ion,
num
ber
of o
bser
vati
ons
show
ing
a po
siti
ve, n
eutr
al, o
r ne
gati
ve r
espo
nse
to e
nvir
onm
enta
l het
erog
enei
ty (a
t lan
dsca
pe, l
ocal
, and
inte
rmed
iate
sc
ales
), an
d ty
pe o
f met
ric
asse
ssed
.
Land
scap
e Lo
cal
Inte
rmed
iate
A
utho
r G
uild
* N
atur
al e
nem
ies
Pos
N
eu N
eg
Pos
N
eu
Neg
P
os
Neu
Neg
M
etri
c#
Arm
brec
ht &
Per
fect
o 20
03
Gro
und-
fora
ging
H
ymen
opte
ra: F
orm
icid
ae
1
1
1 D
Arog
a &
Am
bass
a-K
iki 2
007
Plan
t-for
agin
g D
erm
apte
ra: F
orfic
ulid
ae;
1
A
Cole
opte
ra: C
occi
nelli
dae;
Bl
atto
dea:
Bla
ttel
lidae
; Ar
anea
e: A
rane
idae
; H
ypm
enop
tera
: For
mic
idae
; H
emip
tera
: var
ious
Bian
chi e
t al.
2005
Pl
ant-f
orag
ing,
Co
leop
tera
: Sta
phyl
inid
ae,
3 3
A,
P
Pa
rasi
toid
s Ca
rabi
dae;
Hym
enop
tera
:
Br
acon
idae
, Tri
chog
ram
ma;
N
euro
pter
a: C
hrys
opid
ae
Bom
mar
co 1
998
Gro
und-
fora
ging
Co
leop
tera
: Car
abid
ae
1
A
(C
ondi
tion)
Cai e
t al.
2007
Al
l Va
riou
s
3 1
A, D
Carm
ona
& L
andi
s 19
99
Gro
und-
fora
ging
Co
leop
tera
: Car
abid
ae
1
A
Clou
gh e
t al.
2005
G
roun
d-fo
ragi
ng
Aran
eae:
var
ious
1
1
2
2
A, D
Cost
amag
na e
t al.
2004
Pa
rasi
toid
s H
ymen
opte
ra: B
raco
nida
e
1
A
Ellio
t et a
l. 19
98
Plan
t-for
agin
g Co
leop
tera
: Coc
cine
llida
e;
6
A
N
euro
pter
a: C
hrys
opid
ae;
Hem
ipte
ra: N
abid
ae
Ellio
t et a
l. 20
02
Plan
t-for
agin
g Co
leop
tera
: Coc
cine
llida
e;
2 3
A
Neu
ropt
era:
Chr
ysop
idae
;
Hem
ipte
ra: N
abid
ae
Gar
dine
r et a
l. 20
09
All
Vari
ous
1
A
Gia
noli
et a
l. 20
06
All
Vari
ous
1
A
Cont
inue
d on
nex
t pag
e
45
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,
44 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2A
ppen
dix
1. C
onti
nued
.
Land
scap
e Lo
cal
Inte
rmed
iate
A
utho
r G
uild
* N
atur
al e
nem
ies
Pos
N
eu N
eg
Pos
N
eu
Neg
P
os
Neu
Neg
M
etri
c#
Har
mon
et a
l. 20
03
Gro
und-
fora
ging
Co
leop
tera
: Car
abid
ae,
2 1
A
St
aphy
linid
ae; A
rane
ae
Kle
in e
t al.
2006
Pl
ant-f
orag
ing
Hym
enop
tera
: Eum
enid
ae,
2
1 1
A,
D
Pom
pilid
ae, S
phec
idae
Kru
ess
& T
scha
rntk
e 19
94
Para
sito
ids
Hym
enop
tera
: Bra
coni
dae,
2
3
A,D
,P
Eulo
phid
ae, E
upel
mid
ae,
Pter
omal
idae
, Tor
ymid
ae
Kru
ess
& T
scha
rntk
e 20
00
Para
sito
ids
Hym
enop
tera
: Pte
rom
alid
ae,
2
1 1
A,
D,P
Eu
pelm
idae
, Bra
coni
dae,
Ic
hneu
mon
idae
Kru
ess
2003
Pa
rasi
toid
s H
ymen
opte
ra: B
raco
nida
e,
2 2
3
1
A,
D, P
Pt
erom
alid
ae, E
ucoi
lidae
Lund
gren
et a
l. 20
09
Plan
t-for
agin
g H
emip
tera
: Ant
hoco
rida
e
1
A
Mar
ino
& L
andi
s 19
96
Para
sito
ids
Dip
tera
: Tac
hini
dae;
1
1
A
Hym
enop
tera
: Bra
coni
dae,
Ic
hneu
mon
idae
Men
alle
d e
t al.
2003
Pa
rasi
toid
s H
ymen
opte
ra: B
raco
nida
e 3
2
P
Men
alle
d et
al.
1999
Pa
rasi
toid
s H
ymen
opte
ra: B
raco
nida
e,
1 2
P
Eu
loph
idae
, Ich
neum
onid
ae
Mey
er e
t al.
2009
Pl
ant-f
orag
ing
Dip
tera
: Syr
phid
ae
1
1
D
, A
Mur
phy
et a
l. 19
96
Para
sito
ids
Hym
enop
tera
:Mym
arid
ae
1
A
Mur
phy
et a
l. 19
98
Para
sito
ids
Hym
enop
tera
:Mym
arid
ae
1
P
Nic
holls
et a
l. 20
01
Plan
t-for
agin
g,
Cole
opte
ra: C
occi
nelli
dae;
4
4 1
A, P
Para
sito
ids
Neu
ropt
era:
Chr
ysop
idae
; H
emip
tera
: Nab
idae
; D
ipte
ra: S
yrph
idae
; H
ymen
opte
ra: M
ymar
idae
46
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 45A
ppen
dix
1. C
onti
nued
.
Land
scap
e Lo
cal
Inte
rmed
iate
A
utho
r G
uild
* N
atur
al e
nem
ies
Pos
N
eu N
eg
Pos
N
eu
Neg
P
os
Neu
Neg
M
etri
c#
Öst
man
et a
l. 20
01
Gro
und-
fora
ging
Co
leop
tera
: Car
abid
ae
1
1
1
A
(Con
ditio
n)
Pfiffn
er &
Wys
s 20
04
Gro
und/
Co
leop
tera
: Car
abid
ae;
2 4
A,
D
pl
ant-f
orag
ing
Aran
eae;
var
ious
oth
er
pred
ator
s
Pras
ifka
et a
l. 20
04
Gro
und/
Ar
anea
e; C
oleo
pter
a:
1
1
A
pl
ant-f
orag
ing
Cocc
inel
lidae
; Hem
ipte
ra:
Anth
ocor
idae
, Nab
idae
,
Ly
gaei
dae
(s
ubfa
mily
Geo
cori
nae)
Purt
auf e
t al.
2005
a G
roun
d-fo
ragi
ng
Cole
opte
ra: C
arab
idae
1
1
A, D
Purt
auf e
t al.
2005
b G
roun
d-fo
ragi
ng
Cole
opte
ra: C
arab
idae
1
1
2
A, D
Rosc
hew
itz e
t al.
2005
Pa
rasi
toid
s H
ymen
opte
ra: v
ario
us
1
1
P
Schm
idt e
t al.
2005
G
roun
d-fo
ragi
ng
Aran
eae:
var
ious
1
1
1 1
A, D
Schm
idt
& T
scha
rntk
e 20
05
Gro
und-
fora
ging
Ar
anea
e: L
inyp
hiid
ae
2 1
A
Thie
s &
Tsc
harn
tke
1999
Pa
rasi
toid
s H
ymen
opte
ra: I
chne
umon
idae
1
1
1
P
Thie
s et
al.
2003
Pa
rasi
toid
s H
ymen
opte
ra: I
chne
umon
idae
1
P
Thie
s et
al.
2005
Pa
rasi
toid
s H
ymen
opte
ra: A
phid
iidae
3
P
Thie
s an
d Ts
char
ntke
201
0 Pa
rasi
toid
s H
ymen
opte
ra: I
chne
umon
idae
1
P
Tylia
naki
s et
al.
2004
Pa
rasi
toid
s H
ymen
opte
ra: A
phid
iidae
1
1
P
Wei
bull
et a
l. 20
03
Gro
und-
fora
ging
Ar
anea
e; C
oleo
pter
a:
Cara
bida
e, S
taph
ylin
idae
1 1
2
1
1
D
Wilb
y et
al.
2006
Al
l Va
riou
s
2
1 1
A, D
* “Al
l” re
fers
to s
tudi
es co
ntai
ning
all
men
tione
d gu
ilds
(par
asito
ids,
gro
und
fora
gers
, and
pla
nt fo
rage
rs)
# A=a
bund
ance
, D=d
iver
sity
(spe
cies
rich
ness
or d
iver
sity
inde
x), P
=par
asiti
sm r
ate.
Tw
o st
udie
s (in
dica
ted
by th
e de
sign
atio
n ‘C
ondi
tion’
) ass
esse
d th
e co
nditi
on o
f nat
ural
en
emie
s as
an
indi
cato
r of n
atur
al e
nem
y st
atus
; due
to a
low
sam
ple
size
, the
se o
bser
vatio
ns w
ere
grou
ped
with
the
A (a
bund
ance
) stu
dies
.
47
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

46 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Since the early work done by Roark (1947), more than 1200 plant species have been listed as having potential insecticidal value. Bioactivities from plant species against insects, parasites, bacteria and fungi used for pest control have been abun-dantly reported in the literature (Ganjian et al. 1983, Pereira et al. 1997, Adedire and Akinneye 2004, Koona et al. 2007, Ambró-sio et al. 2008, Chukwujekwu et al. 2009, Chenniappan et al. 2011, Tesch et al. 2011, Chagas-Paula et al. 2012, Nhamo et al. 2013, Utono et al. 2014, Green et al. 2017). Despite this emergence of plant products used for pest control, there has been an increased need for an environmentally safe insecti-cide because of the rise of pestilent disease vectors and the emergent disease pathogens they transmit (Isman and Greineisen 2013, Tembo et al. 2018). In addition to being a significant biting pest species, mosquitoes are well-known vectors of pathogens that cause diseases such as malaria, yellow fever, dengue, and many types encephalitides that affect millions of people worldwide (World Health Organization 1996, CDC 2007, Ghosh et al. 2012, Rohring 2013, Mudin 2015).
Due to the multi-pronged anti-herbiv-ory chemical defense systems plants possess
to deter phytophagous insects that include feeding deterrent toxins, growth regulators as well as repellents, many of these primary defense functions are also effective against controlling or repelling mosquitoes and other biting Diptera (Pichersky and Gershenzon 2002). The use of plants to repel mosquitoes, specifically anopheline mosquitoes has been thoroughly reviewed by Maia and Moore (2011) and more recently by Asadollahi et al. (2019) where they concluded that knowl-edge on traditional practices using plants as repellents may be a significant resource from which new natural products could be utilized as alternatives to synthetic chemical repellents. Since the 1920’s, the application of phytochemicals has been used to control mosquito populations (Shaalan et al. 2005). Much of this research has been a bio-pros-pecting exploration into which plants repel mosquitoes through air-borne volatiles from these plants without controlling them (Maia and Moore 2011). However, the attractive toxic sugar bait (ATSB) approach has been well developed and tested on Anopheles, Ae-des and Culex adult and larval mosquitoes (Müller and Schlein 2006, 2008; Müller et al. 2008; Müller et al. 2010a,b,c).
Plant Extract Efficacy on Mosquito Mortality: Preliminary Studies on the Effect of Ailanthus altissima Extract
on Adult Aedes aegypti and Culex quinquefasciatusJohn R. Wallace*, Calen D. Wylie, Ryan L. Wagner
Department of Biology, Millersville University, Millersville, PA 17551* Corresponding author: (e-mail: [email protected];
tel: 717-871-4318, Fax 717-871-7964).
AbstractDue to the negative environmental impact and resistance to synthetic insecticides,
the development of biological control has increased significantly over the past half century with the potential of plant extracts only recently attracting attention. The purpose of this preliminary study was to examine the potential of Ailanthus altissima (Mill.) Swingle extract as a botanical insecticide on adult mosquitoes. Two species of mosquitoes (Aedes aegypti (Linnaeus) and Culex quinquefasciatus Say) (Diptera: Culicidae) and a non-target lepidopteran species, Painted Lady butterfly (Vanessa cardui (Linnaeus) (Lepidoptera: Nymphalidae) were treated with A. altissima extract from new, mature, and senesced leaflets using serial dilutions (0, 25, 50, 75, 100%) of extract via two application methods (aerosol and sugar feeding). We found that application method and leaf age had significant effects on mosquito mortality at high concentrations. These findings indicate that while mortality was not comparatively high to commercial products, there may be potential to use an in-vasive plant extract as a bio-control tool for mosquito vectors of human disease pathogens.
Keywords: Ailanthus altissima, Tree of Heaven, Culex quinquefasciatus, Aedes aegypti, Vanessa cardui, plant extract
48
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 47
Approximately 344 plant species have been described as having mosquitocidal ac-tivity on biting adult mosquitoes (Sukumar et al. 1991, Tawatsin et al. 2001, Phasomku-solsil and Soonwera 2010) while many others have been reviewed as potential larvicidal properties (Shaalan et al 2005). An example of one of these plants is a non-native tree species, Ailanthus altissima (Mill.) Swingle in North America and was originally found in China but introduced to Europe and North America in the mid-late 1700s (Ding et al. 2006). Biochemically, A. altissima has been well documented to possess the quas-sanoid, ailanthone, that is associated with herbidical as well as insecticidal activities (Heisey 1996, De Feo et al. 2003, Alves et al. 2014). Several investigators have noted that leaf and root extract from A. altissima has demonstrated variable insecticidal activity against hematophagic and phytophagic in-sects (Tsao et al. 2002, De Feo et al. 2009). Ailanthus altissima has been used as an insecticide to control crop pests such as Spodoptera littoralis (Boisduval) (Lepidop-tera: Noctuidae) (Pavela et al. 2014, Pavela 2016). The extract is well documented in terms of an herbicide (Heisey and Heisey 2003) but has not been widely tested as an insecticide against insect vectors of human pathogens (Tsao et al. 2002). Recently, we have conducted evaluations of A. altissima food supplements on several species of lepi-dopteran larvae e.g., Spodoptera frugiperda J.E. Smith and also found that A. altissima reduced larval relative growth rates and adult size and also affected adult oviposition behavior (Wagner and Card 2020, Wagner et al. 2020). These findings suggested that A. altissima might be a source of pesticide metabolites that might be useful to control other pest insect species. The purpose of this preliminary study was to determine the efficacy of A. altissima extract on the adult survivorship of two mosquito species, Culex quinquefasciatus Say and Aedes ae-gypti (Linnaeus) (Diptera: Culicidae) and a non-target lepidopteran species, Vanessa cardui (Linnaeus) (Lepidoptera: Nymphali-dae). The objectives for this study included: 1) determine the concentration effect of A. altissima extract on target and non-target insect mortality; 2) compare the extract type (leaf age) on insect mortality and; 3) compare application method (aerosol v. sugar fed) on insect mortality.
Materials and Methods
Two adult mosquito species were used in this study, Cx. quinquefasciatus and Ae. aegypti based on their involvement in vec-toring several viral pathogens worldwide (Liu et al. 2017) and were purchased as
third instar larvae from Benzon Research, Inc. in Carlisle, PA and maintained at 23°C and 14:10 L:D schedule. Larvae were fed approximately 1oz of Tetramin® fish flakes twice/day until pupation as suggested in Imam et al. (2014). Pupae were transferred to rearing chambers and maintained until adult emergence. Adult mosquitoes were fed on a 10% sucrose solution until assayed with A. altissima extract. Adult male and female mosquitoes (n = 25 mosquitoes/cage) were kept in a cage (n = 3 replicate cages per experiment) to avoid crowding. The Painted Lady butterfly, V. cardui was selected as the non-target species to represent an adult in-sect that sugar feeds as a primary food source and has well documented migratory patterns that may overlap with both culicid species chosen for this study (Abbot 1951, Stefanes-cu et al. 2013). Vanessa cardui larvae were purchased from Folk’s Butterfly Farm in Nescopeck, PA. Larvae were maintained at the same temperature and light regimes as mosquitoes and fed artificial food provided by the vendor until pupation. Once V. cardui adults emerged, they were maintained 10% sucrose solution soaked in cotton balls.
We designed two experiments with a 3 × 2 × 5 factorial design in order to obtain a response to A. altissima leaf extract testing three leaf ages (new, mature and senesced leaves) with two application methods (aerosol and sugar fed) across five different concentrations (100%, 75%, 50%, 25%, and 0%). Leaves were collected from live trees between May–October, 2015 representing three different ages (100 leaves/age): newly emerged (May), mature (July) and senesced leaves (October) to assess the effect of leaf age on insect mortality (n = 3 replicate cages/leaf age). Leaves were stored at 0°C until they were used for extractions. Leachate from leaf extract was obtained by using a leaflet pack of 25 g wrapped in cheesecloth that was compressed to a ball-like shape and placed in 1L of distilled water to steep for 24 hours at room temperature (21°C). Dilutions were prepared from a concentrate of 100% extract that was diluted to 75, 50, 25 and 0% (0% represented the control) to assess extract concentration effect insect on mortality (n = 3 replicate cages/treatment with 25 mosquitoes/cage and 12 V. cardui/cage). Distilled water was used as a con-trol solution. Two delivery or application methods were tested (aerosol and sugar fed solution). The aerosol application ex-periment consisted of using a spray bottle to apply three squirts (approximately 3 ml of spray) at the fine spray setting of each extract sprayed evenly across the cage and directly on to the individual mosquitoes and/or butterflies. Aerosol droplet size from the commercial spray bottle was estimated
49
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

48 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
to be approximately 50 microns (Kripena 2020). The sugar fed (ATSB) application experiment was conducted by applying 1 mL of extract solution and 1 mL of 10% sucrose to the cotton ball placing the cotton ball solu-tion inside the each cage. All treated cages were checked 24 hours post extract exposure and mortality was recorded. All percentage data were arcsine transformed for statistical analysis. All comparisons were made using a two-way ANOVA and a Tukey’s Range test to compare mean differences.
Results
We found that both species of mos-quitoes were affected by both extract applications, whereas mortality effects on the non-target species was minimal (Fig. 1A,B,C). In general, the highest mean per-centage mortality after 24 hours neared 60% for both mosquito species.
For example, Ae. aegypti mortality was significantly higher as a result of aerosol ap-plications (F = 30.8; df = 1, 4; P < 0.01); and concentration (F = 70.9; df = 1, 4; P < 0.01) compared to ATSB applications (Fig. 1A). There was no interaction effect between leaf age and concentration for the aerosol treat-ment (F = 1.61; P > 0.05); however, there was an interaction effect between leaf age and concentration for the sugar fed application (F = 52.2; P < 0.01). But, leaf age had mixed effects, that is, no significant difference was observed on mortality with aerosol (F = 2.69; df = 2, 4; P > 0.05) compared to significant effects with ATSB applications (F = 9.42; df = 2, 4; P < 0.01) (Fig. 1A). Whereas, Cx. quinquefaciatus mortality was significantly different for the ATSB application method (F = 6.91; df = 1, 4; P < 0.05), leaf age (F = 4.11; df = 1, 2; P < 0.05) and concentration (F = 18.0; df = 1, 4; P < 0.01) (Fig. 1B). There was an interaction effect between leaf age and concentration for the aerosol application (F = 5.34; P < 0.01) and no interaction effect between leaf age and concentration for the ATSB application (F = 1.85; P > 0.05). While there was no difference on the non-target, V. cardui mortality between application methods (F = 0.75; df = 1, 4; P < 0.05) (Fig. 1C), there was significantly greater mortality with 100% concentration (F = 8.4; df = 1, 4; P < 0.01); however, this mortality % (10.8%) was significantly lower than the mortality for both culicid species (Fig. 1A,B). Leaf age had no significant effect on mortality of V. cardui (F = 0.88; df = 2, 4; P < 0.05). There were no interaction effects between leaf age and concentration with either application method for V. cardui.
Discussion
Both aerosol and ATSB application methodologies have been well tested and demonstrated to successfully control several genera of mosquitoes (Müller et al. 2008, 2010b; Maia and Moore 2011). We report in this preliminary study that the method of application of A. ailanthus extract had different effects on the two mosquito taxa studied. We found significant effects on Ae. aegypti mortality especially when extracts from mature and senesced leaves were used at high concentrations with an aerosol application. Whereas, the ATSB application had a greater impact on Cx. quinquefascia-tus mortality compared to Ae. aegypti. This difference suggests the mode of application may be taxon-specific (e.g., Culicid taxa) suggesting multiple approaches in the use of plant extracts as mosquitocidal agents. While mortality in this study was not as high as commercial synthetic insecticides (not tested in this experiment) to control vectors of major arboviral pathogens, such as dengue viruses, in a region where A. altissi-ma is native, it may provide an inexpensive and simple approach to maintain enough sustainable efforts to reduce breeding sites (Mudin 2015). Using A. altissima, an inva-sive plant species in the United States, as a leaflet extract could provide a cost effective alternative to reducing mosquito population and disease cases while at the same time find an application to reduce populations of a noxious invasive tree such as A. altissi-ma. Preliminary work on larval and pupal survival indicates that concentration may have similar effects (unpublished data). Future work will address leaf age and other extract sources such as bark and roots on larval survival.
With leaf age having a significant impact on mosquito mortality on both appli-cation methods, if A. altissima extract were to be used to control culicid vectors, concen-trations of the allelopathic compounds would need to be adjusted over the growing season. Publication on the insecticidal attributes of the allelochemistry found within the plant leaf tissue has been sparse to nonexistent (Pavela 2014, Pavela and Sedlak 2018). Our findings suggest a broader ecological selec-tive advantage to maintain such herbicidal compounds as a biocontrol strategy for insect pest species, such as mosquitoes. The leaf extract had a greater impact on mosquito mortality with little to no mortality on Painted Lady butterflies suggesting that this product could be used in field applications where these two families of insects coincide. We did find that the ATSB method of applica-tion did have a slightly greater impact on V. cardui mortality, this method of application
50
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 49
Figure 1. Comparative analysis of A. altissima extract concentration and leaf age (A. Aedes aegypti; B. Culex quinquefasciatus; C. Vanessa cardui) from leachate on mean % mortality using the aerosol and sugar fed (ATSB) applications. Error bars represent SEM.
A.
B.
C.
51
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

50 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
would not benefit those non-targets that nectar feed. While the use of A. altissima extract has had detrimental effects on the mortality of other lepidopteran species, e.g., S. littoralis and S. frugiperda (Pavela et al. 2014, Wagner et al. 2020, Wagner and Card 2020), additional work on the impacts of other non-target taxa is warranted.
The effects of A. altissima against target species such as mosquitoes or other dipteran and/or pest species, the safe use, and the overall socio-economic and agro-eco-logical benefit requires additional work. Be-cause natural compounds are not as effective as synthetic pesticides (Casida 1980), in order to adopt more widespread natural pest control products, the novel use of a readily available resource, such as the invasive A. altissima to control pest populations can only be attained through their evaluation under lab as well as field conditions (Tembo 2018). Using an invasive plant such as A. altissi-ma to control pest insect species could have significant effects on both invasive species and pest management while having little impact on non-target nectar feeding taxa as long as the ATSB method of application is not employed.
Acknowledgments
We would like to thank the Niemey-er-Hodgson Undergraduate Research Board, the MU Student Research Grant, and The Millersville University Biological Student Investigators grant for funding this project. We would also like to thank Elizabeth Miller and John Malachowski for their assistance in the collection of A. altissima leaves.
Literature Cited
Abbot, C.H.. 1951. A quantitative study of the migration of the Painted Lady butterfly, Vanessa cardui (L.). Ecology 32(2): 155–171.
Adedire, C.O., and Akinneye, J.O. 2004. Bi-ological activity of tree marigold, Tithonia diversifolia, on cowpea seed bruchid, Calloso-bruchus maculatus (Coleoptera: Bruchidae). Annals of Applied Biology 144: 185–189. doi: 10.1111/j.1744-7348.2004.tb00332.x
Alves, I.A.B. S., Miranda, H.M., Soares, L.A.L., Randau, K.P. 2014. Simaroubaceae family: botany, chemical composition and biological activities. Revista Brasileira de Farmacog-nosia 24: 481–501.
Ambrósio, S.R., Oki, Y., Heleno, V.C.G., Chaves, J.S., Nascimento, P.G.B.D., Lichston, J.E., Constantino, M.G., Varan-da, E.M. Da Costa, F.B. 2008. Constituents of glandular trichomes of Tithonia diversifo-lia: relationships to herbivory and antifeed-
ant activity. Phytochemistry 69: 2052–2060. doi: 10.1016/j.phytochem.2008.03.019.
Asadollahi, A., Khoobdel, M., Zahraei-Ra-mazani, A., Azarmi, S., Mosawi, S.H. 2019. Effectiveness of plant-based repel-lents against different Anopheles species: a systematic review. Malaria Journal 18 #436 Available from https://doi.org/10.1186/s12936-019-3064-8.
Casida, J.E. 1980. Pyrethrum flowers and pyre-throid insecticides. Environmental Health Perspectives 34: 189–202. doi: 10.1289/ehp.8034189.
CDC Working Group in the Mosquito Trans-mitted Diseases Workforce (US). 2007. Infectious Disease Information: Mosqui-to-Borne Diseases. Atlanta (GA).
Chagas-Paula, D.A., Oliveira, R.B., Rocha, B.A., and Da Costa, F.B. 2012. Ethnobot-any, chemistry, and biological activities of the genus Tithonia (Asteraceae). Chemistry & Biodiversity 9(2): 210–235. doi: 10.1002/cbdv.201100019.
Chenniappan, K, Chellamuthu, V., Dhan-dapani, A., Kadarkarai, M. 2011. Syn-ergistic activity of Andrographis paniculata Nees with Bacillus thuringiensis var israel-ensis against malarial vector, Anopheles stephensi Liston (Diptera: Culicidae). Journal of the Entomological Research Society 13(3): 71–86.
Chukwujekwu, J.C., Lategan, C.A., Smith, P.J., Van Heerden, F.R., Van Staden, J. 2009. Antiplasmodial and cytotoxic activity of isolated sesquiterpene lactones from the acetone leaf extract of Vernonia colorata. South African Journal of Botany 75: 176–179. doi: 10.1016/j.sajb.2008.10.001.
De Feo, V., De Martino, L., Quaranta, E. and Pizza, C. 2003. Isolation of phytotoxic compounds from Tree-of-Heaven (Ailanthus altissima Swingle). Journal of Agricultural and Food Chemistry 51: 1177–1180.
De Feo, V., Mancini, E., Voto, E., Curini, M., and Digilio, M.C. 2009. Bioassay-oriented isolation of an insecticide from Ailanthus altissima. Journal of Plant Interactions 4(2): 119–123.
Ding, J., Yun, W., Hao, Z., Weidong, F., Rear-don, R. Min.L. 2006. Assessing potential biological control of the invasive plant, tree-of-heaven, Ailanthus altissima. Biocontrol Science and Technology 16(5/6): 547–566.
Ganjian, I., Kubo, I., and Fludzinski, P. 1983. Insect anti-feedant elemanolide lactones from Vernonia amygdalina. Phytochem-istry 22: 2525–2526. doi: 10.1016/0031-9422(83)80154-x.
Ghosh, A., Chowdhury, N., Chandra, G. 2012. Plant extracts as potential mosquito larvi-
52
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 51
cides. Indian Journal of Medical Research 135(5): 581–598.
Green, P.W.C., Belmain, S.R., Ndakidemi, P.A., Farrell, I.W., Stevenson, P.C. 2017. Insecticidal activity of Tithonia diversifolia and Vernonia amygdalina. Industrial Crops and Products 110: 15–21. doi: 10.1016/j.ind-crop.2017.08.021
Heisey, R.M. 1996. Identification of an allelo-pathic compound from Ailanthus altissima (Simaroubaceae) and characterization of its herbicidal activity. American Journal of Botony 83(2): 192–200.
Heisey, R.M., Heisey, T.K. 2003. Herbicidal effects under field conditions of Ailanthus altissima bark extract, which containes ail-anthone. Plant and Soil 256: 85–99.
Imam, H.Z., Sofi, G., Aziz, S. 2014. The basic rules and methods of mosquito rearing (Aedes aegypti). Tropical Parasitology 4: 53–55.
Isman, M. B., Grieneisen, M. L. 2013. Botanical insecticide research: many publications, lim-ited useful data. Trends in Plant Science. 19: 140–145. doi: 10.1016/j.tplants.2013.11.005
Koona, P., Tatchago, V., and Malaa, D. 2007. Impregnated bags for safer storage of legume grains in West and Central Africa. Journal of Stored Products Research 43: 248–251. doi: 10.1016/j.jspr.2006.06.005
Kripena, K. 2020. Fogger droplet size and effec-tivity, compared. Insect Cop. Available from https://insectcop.net/droplet-sizes-of-differ-ent-mosquito-foggers-and-repellents/.
Liu, Z., Zhou, T., Lai, Z., Zhang, Z., Jia, Z., Zhou, G., Williams, T., Xu, J., Gu, J. Zhou, X., Lin, L., Yan, G., Chen, X-G. 2017. Com-petence of Aedes aegypti, Ae. Albopictus, and Culex quinquefasciatus mosquitoes as Zika virus vectors, China. Emerging Infectious Diseases 23(7): 1085–1091.
Maia, M.F. and Moore, S.J. 2011. Plant-based insect repellents: a review of their efficacy, development and testing. Malaria Journal 10(Suppl 1):S11. doi:10.1186/1475-2875-10-S1-S11.
Mudin, R.N. 2015. Dengue incidence and the prevention and control program in Malay-sia. International Medical Journal Malaysia 14(1): 5–9.
Müller, G.C. and Schlein, Y. 2006. Sugar questing mosquitoes in arid areas gather on scarce blossoms that can be used for con-trol. International Journal for Parasitology 36:1077–1080.
Müller, G.C. and Schlein, Y. 2008. Efficacy of toxic sugar baits against adult cistern-dwell-ing Anopheles claviger. Transactions of the Royal Society of Tropical Medicine & Hygiene 102(5): 480–484.
Müller, G.C., Kravchenko, V.D., Schlein, Y. 2008. Decline of Anopheles sergentii and Ae-des caspius populations following presenta-tion of attractive, toxic (Spinosad), sugar bait stations in an oasis. Journal of the American Mosquito Control Association 24(1):147–149.
Müller, G.C., Beier, J.C., Traore, S.F., Toure, M.B., Traore, M.M., Bah, S., Doumbia, S., Schlein, Y. 2010a. Successful field trial of at-tractive toxic sugar bait (ATSB) plant-spray-ing methods against malaria vectors in the Anopheles gambiae complex in Mali, West Africa. Malaria Journal 9: 210.
Müller, G.C., Junnila, A., Qualls, W., Revay, E.E., Kline, D.L., Allan, S., Schlein, Y., Xue, R.D. 2010b. Control of Culex quinque-fasciatus in a storm drain system in Florida with attractive toxic sugar baits (ATSB). Medical and Veterinary Entomology 24(4): 346–351.
Müller, G.C., Junnila, A., and Schlein, Y. 2010c. Effective control of adult Culex pip-iens by spraying an attractive toxic sugar bait solution in the vegetation near larval developmental sites. Journal of Medical Entomology 47(1): 63–66.
Nhamo, N., Rodenburg, J., Zenna, N., Ma-kombe, G., and Luzi-Kilhupi, A. 2013. Narrowing the rice yield gap in East and Southern Africa: using and adapting exist-ing technologies. Agricultural Systems 131: 45–55. doi: 10.1016/j.agsy.2014.08.003.
Pavela, R. 2014. Acute, synergistic and antago-nistic effects of some aromatic compounds on the Spodoptera littoralis Boisd. (Lep., Noctu-idae) larvae. Industrial Crops and Products 60: 247–258.
Pavela, R., Zabka, M., Tylova, T., and Kresi-nova, Z. 2014. Insecticidal activity of com-pounds from Ailanthus altissima against Spodoptera littoralis larvae. Pakistan Jour-nal of Agricultural Sciences 51(1): 101–112.
Pavela, R. 2016. History, presence and perspec-tive of using plant extracts as commercial botanical insecticides and farm products for protection against insects-a review. Plant Protection Science 52: 229–241. doi: 10.17221/31/2016-PPS.
Pavela, R. and Sedlak, P. 2018. Post-application temperature as a factor influencing the insec-ticidal activity of essential oil from Thymus vulgaris. Industrial Crops and Products 113: 46-49.
Pereira, P. S., Dias, D. A., Vichnewski, W., Tucci Nasi, A. M. T., and Herz, W. 1997. Sesquiterpene lactones from Brazilian Titho-nia diversifolia. Phytochemistry 45: 1445–1448. doi: 10.1016/S0031-9422(97)00142-8.
Pichersky, E. and Gershenzon J. 2002. The formation and function of plant volatiles: per-
53
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

52 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
fumes for pollinator attraction and defense. Current Opinion in Plant Biology 5: 237–243.
Phasomkusolsil S. and Soonwera M. 2010. In-sect repellent activity of medicinal plant oils against Aedes aegypti (Linn.), Anopheles min-imus (Theobald) and Culex quinquefasciatus Say based on protection time and biting rate. Southeast Asian Journal of Tropical Medicine and Public Health 41: 831–840.
Roark, R.C. 1947. Some promising insecticidal plants. Economic Botany 1: 437–445.
Roehrig, J. 2013. West Nile Virus in the United States-a historical perspective. Viruses 5: 3088–3108.
Shaalan, E.A.S., Canyonb, D., Younesc, M.W.F., Abdel-Wahaba, H., Mansoura A.H. 2005. A review of botanical phytochem-icals with mosquitocidal potential. Environ-ment International 3: 1149–1166.
Stefanescu, C., Paramo, F., Åkesson, S., Alar-con, M., Avila, A., Brereton, T., Carnicer, J., Cassar, L.F., Fox, R., Heliola, J., Hill, J.K., Hirneisen, N. 2013. Multi- generation-al long-distance migration of insects: study-ing the painted lady butterfly in the Western Palaearctic. Ecography 36(4): 474–486.
Sukumar, K, Perich, M.J. and Boobar, L.R. 1991. Botanical derivatives in mosquito control: a review. Journal of the American Mosquito Control Association 7: 210–237.
Tawatsin, A, Wratten, S.D., Scott, R.R., Thavara, U., Techadamrongsin Y. 2001. Repellency of volatile oils from plants against three mosquito vectors. Journal of Vector Ecology 26(1):76–82. PMID: 11469188.
Tembo, Y, Mkindi, A.G., Mkenda, Mpumi, N., Mwanauta, R., Stevenson, P.C., Ndaki-demi, P.A., and Belmain, S.R. 2018. Pesti-cidal plant extracts improve yield and reduce
insect pests on legume crops without harming beneficial arthropods. Frontiers in Plant Science 9: 1425. doi: 10.3389/fpls.2018.01425
Tesch, N. R., Mora, F., Rojas, L., Díaz, T., Vel-asco, J., Yánez, C., Rios, N., Carmona, J., Pasquale, S. 2011. Chemical composition and antibacterial activity of the essential oil of Lantana camara var. moritziana. Natural Product Communications 6: 1031–1034.
Tsao, R., Romanchuk, F.E., Peterson, C.J., and Coats, J.R. 2002. Plant growth regu-latory effect and insecticidal activity of the extracts of the Tree of Heaven (Ailanthus altissima L.). BMC Ecology 2: Article 1. Available from http://www.biomedcentral.com/1472-6785/2/1).
Utono, I. M., Coote, C., and Gibson, G. 2014. Field study of the repellent activity of ‘Lem-ocimum’-treated double bags against the insect pests of stored sorghum, Tribolium castaneum and Rhyzopertha dominica, in northern Nigeria. Journal of Stored Prod-ucts Research 59: 222–230. doi: 10.1016/j.jspr.2014.03.005.
World Health Organization. 1996. Report of the WHO informal consultation on the evaluation on the testing of insecticides, CTD/WHO PES/IC/96.1. Geneva: WHO; 1996. World Health Organization; p. 69.
Wagner, R.L. and Card, J.A. 2020. Ailanthus altissima aqueous extract deters Spodoptera frugiperda oviposition. The Great Lakes Entomologist 53(1-2): 67–71.
Wagner, R.L., Leach, M.E., and Wallace, J.R. 2020. Leaf extract from Ailanthus altissima negatively impacts life history aspects in Spodoptera frugiperda. Journal of the Kansas Entomological Society 93(2): 140–152.
54
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 53
True albinism (i.e., the complete lack of pigmentation) is rare in insects (McCafferty and Bloodgood 1985). Whereas melanin is the major pigment responsible for surface color in vertebrates (Riley 1997), insect col-oration is more complex, with a diversity of (variously synthesized or acquired) pigments and cuticular structure involved in color production (Ghiradella 2009). Thus, partially albinic insects (lacking in one pigment or some but not all pigments) may be difficult to recognize as such, and cases of full or partial albinism are rarely reported among insects (Olosutean 2015). Yet there are known ex-amples of albinic specimens from various orders, including well-documented cases in Lepidoptera and Orthoptera (Putnam 1958, Shapiro 1977, Gall and Schweitzer 1983, Tanaka 1993, González-Estébanez and Manceñido-González 2011).
Here, I describe cases of anomalous pale pubescence recently discovered in three genera of cleptoparasitic bees (Hymenoptera: Apidae: Nomadinae): Brachymelecta Linsley, Epeolus Latreille, and Triepeolus Robertson. Specifically, specimens were found with pale (off-white to pale-yellow) hairs on parts of the body (most conspicuously the discs of the metasomal terga) where in most conspe-cifics there are dark (brown or black) hairs. Curiously, the integument in the aberrant specimens is still primarily dark brown/black, as it is normally. Thus, these individuals appear to exhibit a form of partial albinism in which only cuticular hairs lack the normal pigmentation. Most specimens were captured in recent years (2015 and 2019) and are in good condition, showing little wing wear and no signs of integument discoloration (speci-
mens subjected to prolonged exposure to the elements and/or certain killing agents tend to fade to a reddish-brown color). In the absence of these and other signs of degradation (the DNA of the only sequenced specimen was of a high-enough quality to yield a DNA barcode sequence 630 base pairs in length), I do not attribute the observed atypical pubescence coloration to age- or preservation-related factors.
In two cases, partially albinic spec-imens were mistaken for and erroneously described as new species (one was even placed in its own genus). Hence, it is im-portant that bee taxonomists be made aware of this phenomenon and its potential to result in taxonomic errors. To this effect, differences between anomalously pale-haired specimens and conspecifics exhibiting the usual pubescence coloration are discussed and illustrated below, following Michener (2007) and Onuferko (2018) for terminology. High-quality digital photographs were taken (at the Canadian Museum of Nature Natural Heritage Campus in Gatineau, Quebec, Can-ada (CMNC)) using the Leica Z16 APO A im-aging system (Leica Microsystems, Wetzlar, Germany), which were focus stacked with the accompanying LAS software and assembled into figure plates in Adobe Photoshop 2020 (Adobe Inc.).
Instances of Atypical Pubescence Coloration in Cleptoparasitic Apidae
Brachymelecta californica (Cresson)The holotype of Melecta? mucida
Cresson was recently recognized as an unusual specimen of a widespread species,
Anomalously Pale-Haired Specimens in Three Genera of Cleptoparasitic Bees (Hymenoptera: Apidae: Nomadinae)
Thomas M. OnuferkoResearch Division, Canadian Museum of Nature, P.O. Box 3443, Station D, Ottawa, Ontario, K1P
6P4, Canada. (e-mail: [email protected]).
AbstractCases of partially albinic specimens of cleptoparasitic bees (Hymenoptera: Apidae:
Nomadinae) in which the pubescence lacks the usual pigmentation are presented and discussed. Anomalously pale-haired individuals within the following three genera are known: Brachymelecta Linsley, Epeolus Latreille, and Triepeolus Robertson. Since two of the aberrant specimens were mistaken for and erroneously described as new taxa, the present paper draws attention to this phenomenon within bees and its potential to cause taxonomic confusion.
Keywords: Brachymelecta, Epeolus, partial albinism, pigments, Triepeolus
55
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

54 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
B. californica (Cresson), which until very recently (Onuferko et al. 2021) was known as Xeromelecta californica (Cresson). The specimen, which is housed in the Academy of Natural Sciences of Drexel University in Philadelphia, Pennsylvania, USA (ANSP), was assigned to its own genus (Brachymelec-ta) by Linsley (1939) due to the presence of two (as opposed to three) submarginal cells on each fore wing and pale pubescence cov-ering almost the entire dorsal surface of the metasoma. However, the number of submar-ginal cells is known to be variable within B. californica and can differ even between the left and right sides of the same specimen (Scarpulla 2018, Onuferko et al. 2021). In the M.? mucida holotype, the pubescence is entirely pale except on the axillae, each of which bears a tuft of brown hairs. An anomalously pale-haired specimen of what is now understood to be the same species but completely lacking in dark hairs (including on the axillae) except on T6 was discovered among some cleptoparasitic bees borrowed from the Packer Collection at York Univer-sity (PCYU) in Toronto, Ontario, Canada. The specimen was DNA barcoded as part of the taxonomic revision by Onuferko et al. (2021) and its sequence matched with 99.5% similarity to a sequence of a specimen of B. californica exhibiting the usual pubescence coloration. Although in both the M.? muci-da holotype and sequenced specimen pale hairs cover the discs of the metasomal terga, they are sparser than those comprising the metasomal fasciae, such that the metasoma still appears to be fasciate (see Fig. 1A–B). For these reasons, the primary types of M.? mucida, which was described in 1879, and Melecta californica Cresson, which was described in the preceding year, were recog-
nized as belonging to the same species, and the name Brachymelecta mucida (Cresson) was thus rendered invalid. However, since the “species” was the first among its conge-ners to be removed from Melecta and placed in its own genus, Brachymelecta takes prece-dence as the generic name over Xeromelecta Linsley, so the accepted name for the species is now B. californica. This extreme example of how the erroneous recognition of an aber-rant specimen as a new species can result in a major nomenclatural change illustrates the importance of describing species from series of specimens whenever possible.
Material examined: USA: Nevada: ♂, M.? mucida holotype, H.K. Morrison leg. (ANSP 2294); New Mexico: 1 ♀, NM 15 (16.7 mi N of Silver City), Gila National For-est (32.9462° N, 108.1978° W), 05.ix.2015, R.R. Ferrari leg. (Barcode of Life Data System (BOLD) sample ID: CCDB-34570 A03, PCYU).
Epeolus ainsliei CrawfordAlthough rare in Epeolus compactus
Cresson and uncommon in E. minimus (Robertson) (vide infra), in some species of Epeolus, especially E. ainsliei Crawford, it is not uncommon to find extensively or entirely pale-haired individuals, which are otherwise morphologically indistinguishable from con-specifics with abundant dark brown/black hairs. This suggests that the phenotype has some adaptive value or is at the very least not detrimental for the survival of individ-uals of certain species/populations. On the discs of the metasomal terga, E. ainsliei exhibits continuous variation in pubescence coloration, which ranges from entirely pale to predominantly dark brown/black (see Fig.
Figure 1. Brachymelecta californica (Cresson), metasoma of female, dorsal view: (A) anomalously pale-haired specimen (BOLD sample ID: CCDB-34570 A03, PCYU); (B) specimen exhibiting the usual pubescence coloration (BOLD sample ID: LRBBC1015, PCYU LRB-445). DNA barcode sequences are available for both specimens, which show a high degree of overlap (98.5% similarity) and were assigned the same barcode index number: BOLD:AAC6481.
56
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 55
2A–B), with the differences in hair color not corresponding to any particular geographic pattern. I collected predominantly to entirely pale-haired specimens of E. ainsliei at an active sand dune complex in Spruce Woods Provincial Park, Manitoba, Canada in July 2019, in an area dominated by white prai-rie clover, Dalea candida Michx. ex Willd. (Fabaceae), and where other conspicuously
pale/pale-haired aculeates (Hymenoptera: Aculeata), including Bembix pallidipicta Smith (Bembicidae: Bembicinae), Colletes wilmattae Cockerell (Colletidae: Colleti-nae), Neolarra vigilans (Cockerell) (Apidae: Nomadinae), Perdita perpallida Cockerell (Andrenidae: Panurginae), and Philanthus albopilosus Cresson (Philanthidae: Philan-thinae), were observed and collected.
Figure 2. Epeolines, habitus, dorsal view: (A–B) Epeolus ainsliei Crawford; (C–D) E. minimus (Rob-ertson); (E–F) Triepeolus helianthi (Robertson); (A) extensively pale-haired female (CMNC); (B) female with abundant dark brown/black hairs on the discs of the metasomal terga (C. P. Gillette Museum of Arthropod Diversity, Fort Collins, Colorado, USA); (C) anomalously pale-haired male (CMNC); (D) male exhibiting the usual pubescence coloration (CMNC); (E) anomalously pale-haired female (WRME WRME0501596); (F) female exhibiting the usual pubescence coloration (WRME WRME0508674).
57
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

56 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Material examined: CANADA: Man-itoba: 1 ♀, 1 ♂, Spirit Sands, Spruce Woods Provincial Park, active sand dune complex (49.6637° N, 99.2858° W), 16.vii.2019, T.M. Onuferko leg. (CMNC); 3 ♀♀, same collection data as for preceding, except 30.vii.2019, T.M. Onuferko leg. (CMNC).
Epeolus compactus CressonIn a recent revision of Epeolus for all
known species occurring in North America north of Mexico (Onuferko 2018), the holo-type of E. crucis Cockerell was recognized as an unusual specimen of E. compactus, and the former name was synonymized un-der the latter. Interestingly, the specimen, which is housed in the Smithsonian National Museum of Natural History U.S. National Entomological Collection (USNM) in Wash-ington, D.C., USA, was correctly identified as E. compactus previously by William J. Fox and still bears his original identification label. However, Cockerell (1904) disagreed with the identification and described the specimen under a new name, E. crucis. The specimen lacks dark hairs entirely, and the pale hairs on the discs of the metasomal terga are sparser than those comprising the metasomal fasciae, such that the metasoma still appears to be fasciate (see fig. 38 in Onuferko 2018).
Material examined: USA: New Mex-ico: ♀, E. crucis holotype, Las Cruces, C.H.T. Townsend leg. (USNM 534043).
Epeolus minimus (Robertson)Eight specimens of E. minimus were
caught in pan traps that I set out for ap-proximately six hours on an active sand dune in southwestern Saskatchewan on 4 August 2019. Of these, a male (one of seven collected there and then) has pale hairs on the discs of the metasomal terga, although they are sparser and slightly darker than those comprising the metasomal fasciae, and thus apparently not completely lacking in pigments (Fig. 2C). In this species, the hairs on the discs of the metasomal terga are usually black except for the T1 basal fascia and lateral longitudinal bands on T1 and T2 (Fig. 2D).
Material examined: CANADA: Sas-katchewan: 1 ♂, N of Bitter Lake, Tunstall, active sand dune (50.1456° N, 109.8062° W), 04.viii.2019, ex white pan trap, T.M. Onuferko leg. (CMNC).
Triepeolus helianthi (Robertson)An unusual specimen of Triepeolus
helianthi (Robertson) was discovered among representatives of the genus borrowed from the Wallis-Roughley Museum of Entomology
(WRME) at the University of Manitoba in Winnipeg, Manitoba, Canada. The specimen was identified as T. helianthi by Prof. Jason Gibbs and agrees with the current taxon concept for the species (see Rightmyer 2008) but lacks dark hairs entirely and instead has only pale-yellow pubescence. As is the case with the partially albinic specimens of Bra-chymelecta and Epeolus, the metasoma in this specimen is still distinctly fasciate due to the greater density of hairs comprising the metasomal fasciae (see Fig. 2E–F).
Material examined: CANADA: Man-itoba: 1 ♀, Linden, Rural Municipality of Taché (49.6790° N, 96.8890° W), 09.viii.2019, S. Shukla-Bergen (WRME WRME0501596).
Concluding Remarks
Although herein documented within cleptoparasitic Apidae, this phenomenon may be more widespread among bees. Ito and Sakagami (1980) reported variation in pubescence coloration in Bombus crypta-rum (Fabricius) (as B. florilegus) (Apidae: Apinae), with varying degrees of albinism observed in workers and males on different islands in the western Pacific. Unusually pale-haired individuals of common eastern bumble bees, Bombus impatiens Cresson, have also been observed on occasion (Z. Portman, personal communication, 2021). In species of Bombus Latreille, pubescence coloration is known to be produced by two pigments, eumelanin and pheomelanin, the absence of both of which is presumed to produce white hair coloration (Polidori et al. 2017). Whether the same pigments are responsible for pubescence coloration in the cleptoparasitic apids treated in the present paper remains to be determined. Future discoveries of albinic specimens should be documented to assess the extent of this phenomenon within bees and investigate the potential importance of teratologous mutants in producing advantageous phe-notypes via directional selection. It has been suggested that in various sand wasps, extensive pale-yellow pigmentation (or xanthochroism), which is a common feature among psammophiles, may help with preda-tor avoidance (by providing cryptic coloration against pale substrates) and/or thermoreg-ulation (by slowing down the rate at which specimens heat up while perched on sand) (Hilchie 1982, O’Neill and Evans 1983). This might explain why multiple conspicuously pale-haired individuals of E. ainsliei were observed on active sand dunes. At the same time, when investigating potential cases of albinism it is important to consider possible discoloration effects from various collection and preservation techniques. For instance, just as vane traps used for long periods of
58
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 57
time and pan traps used repeatedly will suf-fer some bleaching from prolonged exposure to direct sunlight, insects too may fade and become discolored under such conditions (Packer and Darla-West 2021).
AcknowledgmentsI thank Laurence Packer (PCYU) and
Jason Gibbs (WRME) for sending me the aberrant specimens of B. californica and T. helianthi, respectively, for study. I also thank Laurence Packer for his constructive comments on this note. Additionally, I am grateful for the research permits issued to me under which I collected bees on various sand dunes in Manitoba (permit number: PP-PHQ-19-004) and Saskatchewan (permit number: 19AR006). Those particular speci-mens were collected as part of a Beaty Post-doctoral Fellowship for Species Discovery, awarded to me by the Canadian Museum of Nature, and I thank both the Ross Beaty fam-ily and museum for supporting my research. Lastly, I thank three anonymous reviewers for their constructive feedback on this note.
Literature CitedCockerell, T. D. A. 1904. II.—Some par-
asitic bees. Annals and Magazine of Natural History 13: 33–42. https://doi.org/10.1080/00222930409487052
Gall, L. F. and D. F. Schweitzer. 1983. Albinic variants of Chlosyne nycteis from Connecticut (Nymphalidae). Journal of the Lepidopterists’ Society 37: 85–86.
Ghiradella, H. 2009. Coloration, pp. 213–220. In Resh, V. H. and R. T. Cardé (eds), Encyclope-dia of insects, second edition. Academic Press (Elsevier), USA.
González-Estébanez, F. J. and D. C. Manceñi-do-González. 2011. Descripción de algunas teratologías en Lepidoptera ibéricos. Boletín de la Sociedad Entomológica Aragonesa 49: 247–252.
Hilchie, G. J. 1982. Evolutionary aspects of geo-graphical variation in color and of prey in the beewolf species Philanthus albopilosus Cres-son. Quaestiones Entomologicae 18: 91–126.
Ito, M. and S. F. Sakagami. 1980. The bumble-bee fauna of the Kurile Islands (Hymenop-tera: Apidae). Low Temperature Science. Series B, Biological Science 38: 23–51.
Linsley, E. G. 1939. A revision of the Nearctic Melectinae (Hymenoptera, Anthophoridae). Annals of the Entomological Society of America 32: 429–468. https://doi.org/10.1093/aesa/32.2.429
McCafferty, W. P. and D. W. Bloodgood. 1985. A case of albinism in Ephemeroptera. Ento-mological News 96: 119–120.
Michener, C. D. 2007. The Bees of the World, 2nd edition. The Johns Hopkins University Press, Baltimore. Maryland, USA.
O’Neill, K. M. and H. E. Evans. 1983. Alter-native male mating tactics in Bembecinus quinquespinosus (Hymenoptera: Sphecidae): correlations with size and color variation. Be-havioral Ecology and Sociobiology 14: 39–46. https://doi.org/10.1007/BF00366654
Olosutean, H. 2015. An albino water strider (He-miptera: Gerridae) from the Danube Delta. Entomological News 124: 287–294. https://doi.org/10.3157/021.124.0407
Onuferko, T. M. 2018. A revision of the clep-toparasitic bee genus Epeolus Latreille for Nearctic species, north of Mexico (Hymenop-tera, Apidae). ZooKeys 755: 1–185. https://doi.org/10.3897/zookeys.755.23939
Onuferko, T. M., L. Packer, and J. A. Genaro. 2021. Brachymelecta Linsley, 1939, previ-ously the rarest North American bee genus, was described from an aberrant specimen and is the senior synonym for Xeromelecta Linsley, 1939. European Journal of Taxon-omy 754(1): 1–51. https://doi.org/10.5852/ejt.2021.754.1393
Packer, L. and G. Darla-West. 2021. Bees: how and why to sample them, pp. 55–83. In San-tos, J. C. and G. W. Fernandes (eds), Measur-ing arthropod biodiversity. Springer, Cham. https://doi.org/10.1007/978-3-030-53226-0_3
Polidori, C., A. Jorge, and C. Ornosa. 2017. Eumelanin and pheomelanin are predomi-nant pigments in bumblebee (Apidae: Bom-bus) pubescence. PeerJ 5: e3300. https://doi.org/10.7717/peerj.3300
Putnam, L. 1958. Albinism in the migra-tory grasshopper, Melanoplus bilitura-tus (Wlk.). Nature 182: 1529. https://doi.org/10.1038/1821529a0
Rightmyer, M. G. 2008. A review of the clepto-parasitic bee genus Triepeolus (Hymenop-tera: Apidae).—Part I. Zootaxa 1710: 1–170. https://doi.org/10.11646/zootaxa.1710.1.1
Riley, P. A. 1997. Melanin. The International Journal of Biochemistry & Cell Biology 29: 1235–1239. https://doi.org/10.1016/S1357-2725(97)00013-7
Scarpulla, E. J. 2018. Four submarginal cells on a forewing of Melitoma taurea (Say) (Hyme-noptera: Apidae), and a summary of known records of atypical and variable numbers of submarginal cells. Insecta Mundi 0667: 1–28.
Shapiro, A. M. 1977. An “albinic” Pieris sisym-brii (Pieridae) from the California Sierras. Journal of the Lepidopterists’ Society 31: 134.
Tanaka, S. 1993. Hormonal deficiency causing albinism in Locusta migratoria. Zoological Science 10: 467–471.
59
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

58 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Two of the earliest major reports on birch-infesting insects in the United States were those of Packard (1890) and Felt (1905). The bronze birch borer, Agrilus anxius Gory (Coleoptera: Buprestidae), was recognized as a major pest of birch since the late 1800s (Chittenden 1898), and still today often reaches outbreak levels especially when trees are stressed by drought and defoliation (Haack and Petrice 2019). Several species of bark- and wood-infesting insects, often called borers, have been associated with various species of birch in North America, including Coleoptera (e.g., Buprestidae, Cerambyci-dae, and Scolytinae), Hymenoptera (e.g., Siricidae and Xiphydriidae), and Lepidop-tera (e.g., Sesiidae) (Smith 1976, Taft et al. 1991, Solomon 1995, Smith and Schiff 2001).
This paper reports on borers and par-asitoids reared from yellow birch (Betula alleghaniensis Britton) at the same Michigan location where Haack (2020) reported rear-ing records from shagbark hickory [Carya ovata (Mill.) K. Koch] and slippery elm (Ul-mus rubra Muhl.). The study site was near Dansville in Ingham County, where I lived for about 30 years. This property was over 13 ac (5.3 ha) in size, and along with the neighboring properties contained over 60 ac (24 ha) of contiguous mature forest of the
beech-maple type (Cohen 2008). The most common tree species growing in this Dans-ville woodlot were listed in Haack (2020). Although these rearing records are decades old, they still provide valuable new informa-tion with respect to larval host plants, adult seasonal emergence, and new state records for Michigan insects.
Methods. On 19 April 1986, I cut two stems (12- and 15-cm-diam) of an apparently healthy, multi-stemmed yellow birch tree near ground level in the Dansville woodlot (N 42.5481° Lat, W 84.3189° Long). The two stems were placed upright and left leaning against the trunk of the tree from which they were cut for the next 12 months. On 26 April 1987, I cut the two stems into more than 20 sections, mostly between 40–55 cm long, and down to a final branch diameter of about 3 cm. When examining the cut ends of each section it was clear that the greatest borer activity was in the sections that were 3 to 10 cm in diameter. Therefore, I collected 16 trunk and branch sections that were in this diameter range (about 7 m total length for all 16 sections) and placed them all in a single rearing cage (ca. 60 cm wide, 45 cm deep, and 45 cm tall). The cage had a wood floor, a sliding Plexiglas front panel, and fine screening on the other three side walls and
Bark- and Wood-Infesting Insects (Coleoptera and Hymenoptera) and Associated Parasitoids Reared from Yellow Birch (Betula alleghaniensis) in Ingham County, Michigan
Robert A. HaackUSDA Forest Service, Northern Research Station, 3101 Technology Blvd., Suite F, Lansing, MI 48910
(e-mail: [email protected] (emeritus)).
AbstractFour species of bark- and wood-infesting borers (two Coleoptera and two Hyme-
noptera) and six parasitoid species (Hymenoptera) were reared from yellow birch (Betula alleghaniensis Britton) one year after the trees were cut and left standing in a woodlot in Ingham County, Michigan in 1986–87. The borers were species of Cerambycidae (Coleop-tera) and Xiphydriidae (Hymenoptera), and hymenopteran parasitoid species of Aulacidae, Braconidae, Chalcididae, and Pteromalidae. Xiphydriophagus meyerinckii (Ratzeburg) (Pteromalidae) is a new Michigan state record. For the borers, yellow birch is a new host record for the cerambycid Sternidius alpha (Say) and the xiphydriid Xiphydria tibialis Say. Seasonal emergence data are presented for each insect, as well as sex ratio data for the xiphydriids and aulacid parasitoids. Mean (± SE) exit hole density for all borers and parasitoids combined was 2.3 ± 0.7 exit holes/dm2 for all branch and trunk samples, but significantly higher (4.7 ± 0.8 exit holes/dm2) on smaller branches (3–5 cm diam) vs. larger branches (0.3 ± 0.1 exit holes/dm2, 5–10 cm diam). Dry weight data for the most common borer (Xiphydria mellipes Harris) and most common parasitoid [Aulacus pallipes Cresson; Aulacidae] demonstrated wide intraspecific variation in adult size.
Keywords: Borers, parasitoids, host range, state records, adult size, yellow birch
60
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 59
upper surface. The cage was kept on a count-er, inside a covered shed (ca. 8 ×12 ft or 2.4 × 3.7 m), that had double doors at opposite ends that remained open to allow air flow. The shed received direct sunlight during the early morning hours, but later was shaded, and therefore the indoor temperatures of the shed were similar to ambient conditions.
The cage was generally checked ev-ery 1–2 days for recently emerged insects through August 1987, and then less fre-quently through October 1987. After each collection, all insects were placed in labeled vials and then frozen. Later, once individuals of each morphospecies had been identified by experts (see Table 1 and acknowledgments), the insects were totaled by species and emer-gence date. Specimens of each species were retained by the identifiers in their personal or institutional collections. All parasitoids were identified by staff at the US Depart-ment of Agriculture, Systematic Entomology Laboratory in Beltsville, MD.
In late summer 1987, after all emer-gence had apparently stopped, the emer-gence density of all borers and parasitoids was calculated for each branch. The insect emergence values were based simply on a count of all exit holes seen on the bark sur-face. At first, I attempted to classify the exit holes as large (about 2.5 mm in diameter or larger) or small (less than 2.5 mm), thinking that the larger holes would represent bor-ers and the smaller holes would represent parasitoids. However, this was not possible because there was considerable overlap between some the larger parasitoids (e.g., female Aulacidae) and the smaller borers (e.g., male Xiphydriidae). The bark surface area of each trunk or branch section was calculated as a cylinder, based on the aver-age diameter of each cut end and the section length. As a proxy for borer size, the adult dry weight of selected borers and parasitoids was determined by drying undamaged (i.e., no missing body parts) insects to a constant weight, using an analytical balance. The intraspecific size variation value, I, was cal-culated for these selected species as the ratio of the dry weight of the heaviest individual to the weight of the lightest individual in a method similar to that computed by Anders-en and Nilssen (1983) based on insect length.
Statistical analyses. Mean differenc-es in attack density and adult dry weight were tested for significance using a two-tailed t-test. An alpha level of 0.05 was used to test for significance.
Results. In 1987, which was assumed the first year after infestation of the two birch stems that were cut in 1986, 190 in-dividual borers, representing four species were reared, including two species of Ceram-
bycidae [Coleoptera: Sternidius alpha (Say) and Xylotrechus colonus (Fabricius)] and two species of Xiphydriidae (Hymenoptera: Xiphydria mellipes Harris and Xiphydria tibialis Say) (Table 1). The xiphydriid X. mellipes was the most common borer reared, representing 95% of all borers (Table 1). The sex ratio (male:female) of the reared X. mellipes was 1:1.4. Considering all borers, individuals were collected from 17 May to 19 July 1987 (Table 1).
Similarly, 36 individual parasitoids, representing six species (all Hymenoptera), were reared from birch in 1987, including one species of Aulacidae (Aulacus pallipes Cresson), three Braconidae (Atanycolus impressifrons Shenefelt, Cenocoelius sp., and Coeloides rossicus betulae Mason), one Chalcididae (Haltichella sp.), and one Pteromalidae [Xiphydriophagus meyerinckii (Ratzeburg)] (Table 1). The most common parasitoid reared was the aulacid A. pallipes, representing 81% of all parasitoids collected (Table 1). The male:female sex ratio of the reared A. pallipes was 1:2.6. Considering all parasitoids, individuals were collected from 10 May to 21 July 1987 (Table 1).
Overall, 225 exit holes were counted on the 16 trunk and branch sections, which corresponds closely to the 226 borers and parasitoids that were collected. Considering all exit holes, the mean (± SE) density was 2.2 ± 0.7 exit holes/dm2 of bark surface area for all 16 sections. However, if the sections were divided into two groups based on aver-age diameter then it was clear that densities were significantly higher (4.7 ± 0.8 exit holes/dm2) on smaller branches (3–5 cm diam; N = 7 sections) as compared with larger branch and trunk sections (0.3 ± 0.1 exit holes/dm2; 5–10 cm diam; N = 9 sections; t-value = 6.35, P < 0.0001).
The mean dry weights for X. mellipes adults were 7.6 ± 0.5 mg for males (N = 20; range 4.8–12.1 mg) and 21.4 ± 0.5 mg for fe-males (N = 38; range 13.3–27.2 mg); females were significantly larger than males (t-value = 17.8, P < 0.0001). Similarly, the mean dry weights for A. pallipes adults were 3.8 ± 0.5 mg for males (N = 5; range 2.1–4.8 mg) and 6.4 ± 0.5 mg for females (N = 12; 4.0–8.9 mg); females were significantly larger than males (t-value = 3.7, P < 0.003). Using the range of dry weights for the above species, I-ratios were 2.5 and 2.0 for X. mellipes males and females, respectively, and similarly 2.3 and 2.2 for A. pallipes males and females.
Discussion. There is a succession of borers, parasitoids and other associated in-sects that colonize the woody tissues of trees as they decline, die and decay (Blackman and Stage 1924, Savely 1939, Haack and Slansky 1987). For the four borers reared
61
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

60 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
in the present study, yellow birch is a new host record for the cerambycid S. alpha and the xiphydriid X. tibialis, although both have been reared from other Betula species, e.g., river birch (Betula nigra L.) (Patton 1879, Smith 1976, MacRae and Rice 2007). For the other two borers, Gosling (1986) reared X. colonus from yellow birch in Michigan, and Smith (1976) reports that X. mellipes is a birch specialist, having been reared from at least five birch species including yellow birch. Of the four parasitoids that were iden-tified to the species level, only the braconid C. rossicus betulae has been reared from yellow birch (Mason 1978); however, the au-lacid A. pallipes has been reared from other birch species (Townes 1950, Smith 2001), and the pteromalid X. meyerinckii, which is a European species, has been reared from birch in Europe as well as other hardwoods (Ferrière 1951).
All four borers reared in this study have previously been reported from Mich-igan (Gosling 1973, Smith 1976, Gosling and Gosling 1977). Similarly, using the online SCAN database (https://scan-bugs.
org/) in March 2021, which contains collec-tion records of insects from over 100 North American arthropod collections, Michigan specimens of all four borers currently reside in various collections throughout the United States. With respect to the four parasitoids identified to species, Michigan records have been published for A. pallipes (Townes 1950, Smith 2001), A. impressifrons (Shenefelt 1943), and C. rossicus betulae (Mason 1978). Of these three species, Michigan specimens are reported in SCAN for only A. pallipes.
In the case of X. meyerinckii, no pub-lished papers or SCAN records have been found stating that this insect has been previously collected in Michigan, and it is therefore considered a new state record. This pteromalid has been reported in Indiana, parasitizing Xiphydria maculata Say in maple (Acer) (Deyrup 1984), and reared from larch (Larix) in Newfoundland that were infested by the bark beetle Dendroctonus simplex LeConte (Coleoptera: Curculionidae: Scolytinae) (Langor 1991). In Europe, X. meyerinckii has been reared from at least two species of Xiphydria (Schimitschek 1974).
Table 1. Collection data for adult bark- and wood-infesting borers (Coleoptera and Hyme-noptera) and parasitoids (Hymenoptera) reared from yellow birch (Betula alleghaniensis) trunk and branch sections held outdoors in 1987 in Ingham County, Michigan. Number collected Range of collection datesFAMILY Calendar Julian days
Species Total Males Female days (mean) Identifier2
Borers CERAMBYCIDAE Sternidius alpha 1 n.a.1 n.a. VI 4 155 (155) DCLGXylotrechus colonus 4 n.a. n.a. VI 28–VII 11 179–192 (185) DCLG XIPHYDRIIDAE Xiphydria mellipes 181 76 105 V 17–VII 19 137–199 (167) DRSXiphydria tibialis 4 1 3 VI 17–VII 3 169–184 (176) DRS Parasitoids AULACIDAE Aulacus pallipes 29 8 21 V 28–VII 21 148–202 (168) DRS BRACONIDAE Atanycolus impressifrons 3 0 3 V 10–V 21 130–141 (136) PMMCenocoelius sp. 2 1 1 V 10–V 21 130–141 (136) PMMCoeloides rossicus betulae 2 1 1 VI 22–VI 28 173–178 (176) PMM CHALCIDIDAE Haltichella sp. 1 0 1 V 28 148 (148) EEG PTEROMALIDAE Xiphydriophagus meyerinckii 1 0 1 V 28 148 (148) EEG1 Cerambycid adults were not sexed.2 Identifiers: DCLG = David C. L. Gosling, DRS = David R. Smith, EEG = Eric E. Grissell, and PMM = Paul M. Marsh.
62
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 61
I did not attempt to associate the para-sitoids reared in this study with their insect hosts; however, Deyrup (1984) did make such associations through detailed dissec-tions of infested branches. For the braconid A. impressifrons, Marsh (1979) reports that it has been reared from cerambycids. Similarly, the Cenocoelius species reared in the present study likely had a cerambycid host. In fact, the braconid Cenocoelius ash-meadii Dalla Torre has been reared from the cerambycid S. alpha, as well as other cerambycids (Marsh 1979), and has been previously collected in Michigan based on SCAN records. Members of the chalcidid genus Haltichella are known to be primary parasitoids of Lepidoptera as well as hyper-parasitoids of Braconidae and Tachinidae (Diptera) (Halstead 1990). The only species of Haltichella reported in SCAN to be from Michigan is Haltichella xanticles (Walker), which is recognized as both a primary para-sitoid and hyperparasitoid (Halstead 1990).
Life history studies have been conduct-ed on a few species of Xiphydria. Deyrup (1984) studied X. maculata and its parasit-oids on maple in Indiana, and reported two of the same parasitoids as found in this study (C. rossicus betulae and X. meyerinckii), as well as two other distinct Aulacus species, one Braconidae, one Ichneumonidae, and one Orussidae (Hymenoptera). In addition, Deyrup (1984) reported univoltine life cycles for the borers and parasitoids in his study, adult flight for most species between late May and early August, and that X. maculata mostly infested maple branches and stems that were 2.5 to 9 cm in diameter. Working in New York, Blackman and Stage (1924) reported that Xiphydria hicoriae Rohwer, which infests hickory (Carya), was univol-tine, had adult emergence mostly during July and early August, favored host material 5–9 cm in diameter, and would reinfest the same host material at least once. Smith and Schiff (2001) reported that Xiphydria decem Smith and Schiff was reared from a recently dead branch (2.8–4.1 cm diam) of river birch in southern Illinois in April and May. In studies of Xiphydria picta Konow on alder (Alnus) in the North Caucasus re-gion of southern Russia, Kravchenko (1972) noted that infestation occurs soon after tree death, oviposition is primarily on stems and branches 4–40 cm in diameter, borer densities are highest on smaller diameter trunk and branch sections, the flight season extends from July to early September, and the sex ratio is female biased.
Deyrup (1984) noted that the Aulacus females in his study oviposited on Xiphydria eggs. Worldwide, several Aulacus species use xiphydriids as hosts, especially in the northern hemisphere, as well wood-boring
Coleoptera, especially in the southern hemi-sphere (Evenhuis and Vlug 1975, Smith 2001, Jennings and Austin 2004).
Andersen and Nilssen (1983) reported that intraspecific size variation (I) was gen-erally lower for free-living groups (usually families) of beetles (family medians of 1.1 to 1.4) compared with those that developed in woody tissues [medians of 1.2 for ambrosia beetles (Platypodinae and some Scolytinae) to 2.1 for wood-borers of the cerambycid subfamily Spondylidinae (= Aseminae)]. The highest I value for individual species was 3.3 for two Cerambycinae cerambycids. Haack and Slansky (1987) calculated mean I values of 2.8 for 12 Siricidae and 2.8 for 9 Xiphydriidae, all of which are wood-in-festing Hymenoptera. The I index value for X. mellipes, combining data for males and females, is 2.7 based on length data given in Smith (1976) and 5.9 based on dry weights from the present study. For the A. pallipes parasitoids reared in the present study, the I index value for males and females com-bined is 4.2 based on dry weight. Such wide variation in intraspecific size in wood-boring insects likely reflects, in part, differences in nutritional quality and water content of the woody tissues (Andersen and Nilssen 1983, Haack and Slansky 1987, Shibata 1998, Torres-Vila et al. 2018). Similarly, the broad size variation noted in the borer X. mellipes is likely the main reason for size variation in the parasitoid A. pallipes. Others have found that parasitoid size tends to increase with the size of the larval host in various bupres-tid and cerambycid borers (Urano and Hijii 1995, Paine et al. 2004, Wang et al. 2008).
Acknowledgments
The author thanks David C. L. Gos-ling (Glen Oaks Community College, MI), Eric E. Grissell [Systematic Entomology Laboratory (SEL), Beltsville, MD], Paul M. Marsh (SEL), and David R. Smith (SEL) for providing identification services; and Leah S. Bauer and Toby R. Petrice (both at USDA Forest Service, Northern Research Station, Lansing, MI), E. Richard Hoebeke (University of Georgia, Athens, GA), and two anonymous reviewers for commenting on an earlier draft of this paper. The affiliations listed above were those of the researchers at the time when taxonomic assistance was provided. This research was supported in part by the USDA Forest Service.
Literature Cited
Andersen, J. and A. C. Nilssen. 1983. In-trapopulation size variation of free-living and tree-boring Coleoptera. The Canadian Entomologist 115: 1453–1464.
63
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

62 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Blackman, M. W. and H. H. Stage. 1924. On the succession of insects living in the bark and wood of dead, dying, and decaying hickory. New York State College of Forestry, Techni-cal Bulletin 17: 3–269.
Chittenden, F. H. 1898. A destructive borer enemy of birch trees, with notes on related species. USDA Division of Entomology Bul-letin New Series18: 44–51.
Cohen, J. G. 2008. Natural community ab-stract for mesic southern forest. Mich-igan Natural Features Inventory, Lan-sing, MI. 12 pp. https://web.archive.org/web/20090105155916/http://web4.msue.msu.edu/mnfi/abstracts/ecology/Mesic_south-ern_forest.pdf
Deyrup, M. A. 1984. A maple wood wasp, Xi-phydria maculata, and its insect enemies (Hymenoptera: Xiphydriidae). The Great Lakes Entomologist 17: 17–28.
Evenhuis, H. H. and H. J. Vlug. 1975. Aulacus striatus parasiet van Xiphydria camelus. Entomologische Berichten 35(4): 58. (in German)
Felt, E. P. 1905. Insects affecting park and wood-land trees. New York State Museum Memoir 8: Vol 1. The University of the State of New York, Albany, NY. 459 pp.
Ferrière, C. 1951. A new genus of Pteromalidae (Hym.), parasitic on the alder wood-wasp, Xiphydria camelus L., found in Norfolk, England. Entomologist’s Monthly Magazine 87: 322–324.
Gosling, D. C. L. 1973. An annotated list of the Cerambycidae of Michigan (Coleoptera) Part I, Introduction and the subfamilies Parandri-nae, Prioninae, Spondylinae, Aseminae, and Cerambycinae. The Great Lakes Entomolo-gist 6: 65–84.
Gosling, D. C. L. 1986. Ecology of the Ceramby-cidae (Coleoptera) of the Huron Mountains in northern Michigan. The Great Lakes Entomologist 19: 153–162.
Gosling, D. C. L., and N. M. Gosling. 1977. An annotated list of the Cerambycidae of Mich-igan (Coleoptera). Part II, The subfamilies Lepturinae and Lamiinae. The Great Lakes Entomologist 10: 1–37.
Haack, R. A. 2020. Bark- and wood-infesting Coleoptera and associated parasitoids reared from shagbark hickory (Carya ovata) and slippery elm (Ulmus rubra) in Ingham Coun-ty, Michigan. The Great Lakes Entomologist 53: 185–191.
Haack, R. A. and T. R. Petrice. 2019. Historical population increases and related inciting factors of Agrilus anxius, Agrilus bilineatus, and Agrilus granulatus liragus (Coleoptera: Buprestidae) in the Lake States (Michigan,
Minnesota, and Wisconsin). The Great Lakes Entomologist 52: 21–33.
Haack, R. A. and F. Slansky. 1987. Nutritional ecology of wood-feeding Coleoptera, Lepidop-tera, and Hymenoptera, pp. 449–486. In: F. Slansky and J. G. Rodriguez (eds.), Nutri-tional ecology of insects, mites, spiders and related invertebrates. John Wiley, New York.
Halstead, J. A. 1990. Review of Haltichella Spinola in the Nearctic Region (Hymenop-tera: Chalcididae). Proceedings of the Ento-mological Society of Washington 92: 153–159.
Jennings, J. T. and A. D. Austin. 2004. Biology and host relationships of aulacid and gaster-uptiid wasps (Hymenoptera: Evanioidea): a review, pp. 187–215. In: K. Rajmohana, K. SudheerGirish, P. Kumar, and S. Santhosh (eds.), Perspectives on Biosystematics and Biodiversity. University of Calicut, Kerala, India.
Kravchenko, M. A. 1972. Ecology of the variegat-ed wood wasp Xiphydria picta Knw. in North Caucasia. Soviet Journal of Ecology 3: 77–79.
Langor, D. W. 1991. Arthropods and nematodes co-occurring with the eastern larch beetle, Dendroctonus simplex [Col.: Scolytidae], in Newfoundland. Entomophaga 36: 303–313.
MacRae, T. C. and M. E. Rice. 2007. Biological and distributional observations on North American Cerambycidae (Coleoptera). The Coleopterists Bulletin 61: 227–263.
Marsh, P. M. 1979. Braconidae, pp. 144–215. In: K. V. Krombein, P. D. Hurd, D. R. Smith, and B. D. Burks (eds). Catalog of the Hyme-noptera in America North of Mexico. Vol. 1. Smithsonian Institution Press, Washington, D.C.
Mason, W. R. M. 1978. A synopsis of the Nearctic Braconini, with revisions of Nearctic species of Coeloides and Myosoma (Hymenoptera: Braconidae). The Canadian Entomologist 110: 721–768.
Packard, A. S. 1890. Fifth report of the United States Entomological Commission: being a revised and enlarged edition of Bulletin No. 7, on insects injurious to forest and shade trees. US Department of Agriculture, Washington, DC. 957 pp.
Paine, T. D., A. L. Joyce, J. G. Millar and L. M. Hanks. 2004. Effect of variation in host size on sex ratio, size, and survival of Syngaster lepidus, a parasitoid of eucalyptus longhorned beetles (Phoracantha spp.): II. Biological Control 30: 374–381.
Patton W. H. 1879. A rare Xiphydria and its par-asite. The Canadian Entomologist 11: 12–15.
Savely, H. E. 1939. Ecological relations of certain animals in dead pine and oak logs. Ecological Monographs 9: 321–385.
64
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 63
Schimitschek, E. 1974. Contributions to the ecology of coniferous and leaf wood Siricidae in Europe. Journal of Applied Entomology 75: 225–247. (in German)
Shenefelt, R. D. 1943. The genus Atanycolus Fo-erster in America north of Mexico. Research Studies of the State College of Washington 11: 51–163.
Shibata, E. 1998. Effects of Japanese cedar inner bark nutritional quality on development of Semanotus japonicus (Coleoptera: Ceram-bycidae). Environmental Entomology 27: 1431–1436.
Smith, D. R. 1976. The xiphydriid woodwasps of North America (Hymenoptera: Xiphydriidae). Transactions of the American Entomological Society 102: 101–131.
Smith, D. R. 2001. World catalog of the family Aulacidae (Hymenoptera). Contributions on Entomology, International. 4: 263–319.
Smith, D. R. and N. M. Schiff. 2001. A new species of Xiphydria Latreille (Hymenoptera: Xiphydriidae) reared from river birch, Betula nigra L., in North America. Proceedings of the Entomological Society of Washington. 103: 962–967.
Solomon, J. D. 1995. Guide to insect borers in North American broadleaf trees and shrubs.
U.S. Department of Agriculture, Forest Service, Agriculture Handbook, AH-706. Washington, D.C. 735 pp.
Taft, W. H., D. Smitley and J. W. Snow. 1991. A guide to the clearwing borers (Sesiidae) of the north central United States. Michigan State University, North Central Regional Extension Publication 394. East Lansing, MI.
Torres-Vila, L., F. J. Mendiola-Díaz, and Ã. Sánchez-González. 2018. Adult size and sex ratio variation of Cerambyx welensii (Co-leoptera: Cerambycidae) in Mediterranean oak (Fagaceae) woodlands. The Canadian Entomologist 150: 334–346.
Townes, H. 1950. The Nearctic species of Gas-teruptiidae (Hymenoptera). Proceedings of the United States National Museum. 100: 85–145.
Urano, T. and N. Hijii. 1995. Resource utiliza-tion and sex allocation in response to host size in two ectoparasitoid wasps on subcor-tical beetles. Entomologia Experimentalis et Applicata 74: 23–35.
Wang, X. Y, Z. Q. Yang, H. Wu and J. R. Gould. 2008. Effects of host size on the sex ratio, clutch size, and size of adult Spathius agrili, an ectoparasitoid of emerald ash borer. Bio-logical Control 44: 7–12.
65
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

64 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
Idana marginata (Say 1830) (Diptera: Tephritoidea: Ulidiidae) (Fig. 1) is one of the largest ulidiids in North America (Marshall 2012) and the only species of Idana Loew. Little is known regarding its natural history although Marshall (2012) indicated larvae develop in compost. The species has previ-ously been recorded from southeastern Can-ada and the northeastern United States from Michigan and Indiana east to Massachusetts and south to Virginia (Steyskal 1965, 1987). However, Brimley (1938) recorded I. mar-ginata from Linville Falls, North Carolina during May.
Methods
Specimen data and SpecimensLabel data from the new state record
reported below are presented verbatim. The specimens were field collected into 70–80% ethanol. However, since most Diptera are prone to excessive tissue distortion during the normal dehydration process associated with pinning, the HMDS technique (Nation 1983) was used in specimen preparation to minimize exoskeletal collapsing and shriv-eling.
The specimens of I. marginata reported herein are vouchered in the Insect Research Collection (WIRC) of the Department of En-tomology, University of Wisconsin-Madison.
Study Site
Habitat images (Fig. 2a, b), albeit from different points in time, reference the locality where the Lindgren trap was set from which the specimens were captured.
New Record of Idana marginata (Say) (Diptera: Tephritoidea: Ulidiidae) from Wisconsin
Daniel K. YoungDepartment of Entomology, University of Wisconsin; Madison, WI 53706.
(e-mail: [email protected]; ORCiD ID: 0000-0002-5711-2519).
AbstractIdana marginata (Say) (Diptera: Tephritoidea: Ulidiidae) was previously known from
northeastern North America: Canada and the United States, west to northeastern Illinois and south to North Carolina. Herein, collection events are reported from Richland County in southwestern Wisconsin. This new state record significantly increases the known geographical distribution of the species to the west. The specimens were recovered from an unbaited Lindgren funnel trap during two intervals between 26 June and 28 July 2019.
Keywords: Ulidiidae, Idana marginata, distribution, Wisconsin
Figure 1. Idana marginata (Say). Habitus, dorsal view. Focus-stacked digital image: Craig Brabant.
The site is located near the north shoreline of the Cruson Slough of the Wisconsin River in the southeastern corner of Richland County. This site is part of the Lone Rock Unit of the Lower Wisconsin State Riverway (LWSR) in southwestern Wisconsin. The habitat was historically a Quercus-Pinus sand barren immediately to the north and the slough bottomland to the south. Most of this area has recently been highly modified with much of the oak-pine removed. The extent to which these management practices may have im-
66
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 65
Figure 2. General habitats where Idana marginata (Say) specimens were collected. [Images were captured April 2014 (a) and April 2015 (b).] Digital images: DKY.
67
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

66 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
pacted the habitat of I. marginata remains to be determined.
Results and Discussion
One male and two females of I. margin-ata (Fig. 1) were recovered from an unbaited Lindgren funnel trap: [1st label] USA: WI: Richland Co. // LWSR – Lone Rock Unit // 43.19356°N/-90.23950°W // WGS84; 26 June –5 July 2019 // Jacquelyn R. Whisenant; [2nd label] unbaited Lindgren trap on // tree 10 meters from river// 20190626_JRW_LoneR. An additional three males and four females were recovered from the same trap during the subsequent sampling interval: [1st label] USA: WI: Richland Co. // LWSR – Lone Rock Unit // 43.19356°N/-90.23950°W // WGS84; 5–28 July 2019 // Jacquelyn R. Whisenant; [2nd label] unbaited Lindgren trap on // tree 10 meters from river// 20190705_JRW_Lon-eR.
It should be noted the “river” described on the collection event labels is actually an outflow of the slough that passes beneath a nearby earthen levy. The discharge even-tually flows back into the Wisconsin River approximately one mile from the levy. Al-though the microhabitat is in proximity to the discharge of the Cruson Slough, no ref-erence to I. marginata was made by Keiper et al. (2002).
Prior to the current discovery, the western-most record for I. marginata was from Indiana (Steyskal 1965). More spe-cifically, data from the Global Biodiversity Information Facility (GBIF) (https://www.gbif.org/species/1523861) indicate records from as far west as northwestern Indiana. An additional record has also been validated from the internet: St. Charles, DuPage Co., IL, 5 June 2019 (https://bugguide.net/node/view/1671858/bgimage). The Wisconsin re-cords reported here are approximately 170 miles west-northwest of the Illinois location and the land between the two locations has become highly altered by human activity, largely agricultural. Thus, it would be in-teresting to explore whether any gene flow remains between what might now represent disjunct populations. Interestingly, my students and I have collected at this site numerous times in the past 20+ years by
hand, Malaise traps, and Lindgren funnel traps without previously encountering this large, conspicuous and colorful ulidiid fly.
AcknowledgmentsI am grateful to the collector, Jacki
Whisenant, for specific information relating to the collection site. My thanks to Craig Brabant for his kind assistance in creating the habitus image of I. maginata and for reviewing the manuscript. Thanks are ex-tended to two reviewers for their time and suggestions.
Literature CitedBrimley, C. S. 1938. The insects of North Car-
olina being a list of the insects of North Carolina and their close relatives. North Carolina Department of Agriculture Division of Entomology, Raleigh. 560 p.
Global Biodiversity Information Facility. Idana marginata (Say, 1830). Source: Cata-logue of Life. Map showing 78 georeferenced occurrence records.https://www.gbif.org/species/1523861 (accessed 02 January 2020).
Keiper, J. B, W. E. Walton and B. A. Foote. 2002. Biology and ecology of higher Diptera from freshwater wetlands. Annual Reviews of Entomology 47: 207–232.
Marshall, S. A. 2012. Flies: The Natural History and Diversity of Diptera. Firefly Books, Buf-falo, NY. 616 p.
Nation, J. L., 1983. A new method using Hexam-ethyldisilazane for preparation of soft insect tissues for scanning electron microscopy. Biotechnic & Histochemistry 58: 347–351.
Steyskal, G. C. 1965. Family Otitidae (Ortalidae, Ulidiidae, Pterocallidae). p. 642-654. In A. Stone, C. W. Sabrosky, W. W. Wirth, R. H. Foote, and J. R. Coulson (eds.), A Catalog of the Diptera of America North of Mexico. Agriculture Handbook No. 276. United States Department of Agriculture. Agricultural Re-search Service, Washington, D. C.
Steyskal, G. C. 1987. 63. Otitidae. p. 799-808. In J. F. McAlpine (ed.), Manual of Nearctic Diptera. Volume 2. Biosystematics Research Centre, Agriculture Canada. Monograph No. 28 Canada Communication Group – Publish-ing, Ottawa, Canada.
68
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 67
Lestes australis Walker (commonly known as southern spreadwing) (Odonata: Lestidae) is an enigmatic, uncommon, ear-ly-season species in Wisconsin, where it is at the northern edge of its eastern North American range (Paulson 2011). It is enig-matic, in part, because its identification has long been confused with two morphologically similar species, L. forcipatus Rambur (sweet-flag spreadwing) and L. disjunctus Selys (northern spreadwing, formerly common spreadwing). Prior to the late 1940s, ento-mologists were unaware that the australis form existed. Montgomery (1941) warned that determinations of L. disjunctus and L. forcipatus had been so badly confused that published records of the two species should be disregarded. Walker (1952) named the australis form as a new subspecies of L. disjunctus and he stated that this form “has caused all the confusion between L. disjunc-tus and forcipatus.” A measure of confusion concerning the identification of L. australis persists to this day.
It remains unclear if L. australis is a valid species or a subspecies of L. disjunctus. Since Walker (1952) named it as a subspe-cies, most references on Odonata have treat-ed it as such (see Westfall and May 2006). However, Donnelly (2003) urged that L. australis should be elevated to species status because of widely overlapping ranges and morphological and life history differences between it and the nominate L. disjunctus,
and he noted that the most difficult diag-nosis in the complex is in distinguishing the males of L. forcipatus and L. australis. Paulson (2004) cited the arguments given by Donnelly (2003) and stated that L. australis and L. disjunctus “must be recognized as a more northerly and a more southerly species distinct from one another.” Still, Westfall and May (2006) provisionally retained L. australis as a subspecies of L. disjunctus be-cause of a lack of genetic distinction between the two forms. Paulson (2011) acknowledged that the genetic differences between L. aus-tralis and L. disjunctus are less than those between most species of Lestes, but he again advocated that L. australis was deserving of full species rank because of structural differences and a different flight season. Abbott (2011) also considered L. australis to be deserving of full species status, as do we in this note.
Current identification tools (Walker 1952, 1953; Catling 2003; Donnelly 2003; Lam 2004; Westfall and May 2006; Abbott 2011; Paulson 2011; DuBois 2019) allow clear separation of males of L. australis and L. disjunctus, but distinguishing males of L. australis and L. forcipatus remains somewhat uncertain due to subtle mor-phologic and color- and pruinosity-pattern differences, and probable overlap in some of the frequently used character states. There-fore, when determining these species, field guide authors have recommended looking
The Puzzling Presence of Lestes australis (Odonata: Lestidae) in Wisconsin—Does This Species Migrate?
Robert B. DuBois1,* and Kenneth J. Tennessen2
1 Department of Natural Resources, Bureau of Natural Heritage Conservation, 1701 North 4th Street, Superior, Wisconsin 54880 (retired).
2 P.O. Box 585, Wautoma, Wisconsin 54982*Corresponding author: (e-mail: [email protected], phone: 715-817-2631).
AbstractLestes disjunctus australis Walker (Odonata: Lestidae), 1952 was described as a sub-
species of Lestes disjunctus Selys, 1862. In recent decades it has been considered deserving of full species status by most specialists. The core of its eastern North American range is south of Wisconsin, but during April through June of some years, mature individuals, and occasionally reproductive behavior, are observed at shallow ponds and wetlands mostly in the southern half of the state. Since first recorded in Wisconsin in 2002, it has been detected in 13 of Wisconsin’s 72 counties. However, there has been no unequivocal documentation of successful reproduction in the state. Various possibilities regarding long-range dispersal or facultative migration of this species and other species of Zygoptera are discussed.
Keywords: Zygoptera; Lestes australis; Lestes disjunctus australis; migration; dis-persal
69
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

68 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
for associated pairs because females of L. forcipatus have a long and easily recognized ovipositor that will help rule out L. australis (Lam 2004, Paulson 2011, DuBois 2019), but they also warn that males of species of Lestes will sometimes form tandem with the wrong species.
The first Wisconsin record for L. aus-tralis was a male collected on 26 June 2002, at a beaver pond in northern Douglas County (one of Wisconsin’s northern-most counties; Fig. 1). It was determined as L. australis by RBD, but because it was so far north of its known range, it was sent to T. W. Donnelly, an expert with the group, for verification. Donnelly confirmed the specimen as L. aus-tralis. Subsequent visits to the site did not detect more specimens, so it was presumed to be a vagrant. Five years later, another male was collected in Monroe County on 28 April 2007. When adult odonates in Wisconsin are seen as early in the flight season as April, they are usually migrants, such as Anax junius (Drury) and Sympetrum corruptum (Hagen). May (2013) noted that the mean date of first appearance of migrants was at least six weeks earlier than the mean date of first emergence of resident species in Maryland and New York. We therefore considered the possibility that L. australis might be a migrant. We are not the first to voice the possibility of migration in this species. Donnelly (1992) noted that in some years, numbers of L. australis were taken at several sites around Binghamton, New York, which he called episodic irruptions. These observations led Soltesz et al. (1995) to pose the question, “Do Zygoptera migrate?”
Another five years passed without additional observations of L. australis, but the lack of observations could have been due to low levels of early season sampling effort rather than the absence of the species in the state. In 2012, unassociated males of L. australis and some pairs in copula and ovipositing were observed on four occasions from mid-May to mid-June in a small, fre-quently visited retention pond in the Town of Holmen, La Crosse County (D. Jackson, pers. comm.). Since then, at least a few ear-ly-season individuals of L. australis have been observed most years at this retention pond, including many males and repro-ductive behavior in 2015 and 2018. Lestes australis was not seen at the pond in 2013 or 2019, and in May 2020 only a single male was found. However, two teneral Lestes, a male and a female, were collected along a short shoreline of the pond on 24 July 2020. Both were tentatively determined as L. aus-tralis, but because of their teneral condition, including some key body parts being twisted or shriveled, and the lack of full color or
any pruinosity, their identity could not be confirmed with certainty.
At another frequently visited site, a man-made wildlife pond in the Buena Vista State Natural Area in Portage County, we observed L. australis adults on two occasions in 2016 and 2017. We made four visits to the pond in May 2017 to collect Lestes nymphs, hoping to document survival of L. australis through the winter. However, all 17 nymphs reared from the pond were L. eurinus Say (amber-winged spreadwing), and 74 F-0 and F-1 nymphs that were preserved and identified were also all L. eurinus.
Over the last decade, individual males or in a few cases multiple specimens have been observed or collected at a handful of additional sites so that L. australis has now been observed in 13 of Wisconsin’s 72 counties, mostly in the southern half of the state (Fig. 1; https://wiatri.net/inventory/odonata/SpeciesAccounts/SpeciesDetail.cfm?TaxaID=168). However, the only hint of successful reproduction remains the un-certain determinations of two tenerals at the Holmen retention pond in 2020.
In central and southern Wisconsin, the habitats where L. australis has been found include permanent, but usually shallow ponds and marshes with abundant emergent vegetation and probably an absence of fish (at least lacking centrarchids). Elsewhere in the range of L. australis it is also found in lakes and slow streams, but we have not found it in those habitats in Wisconsin. Here it has a short, early flight period that peaks in late May and early June. Any Lestes seen in Wisconsin in April or the first half of May are most likely to be L. australis. In the core of its range in the U.S. south of Wisconsin, L. australis is known to have a long flight season that extends well into autumn (Paulson 2011), but in Wisconsin it has not been confirmed later than the end of June. Lestes eurinus is also an early season, resident Wisconsin species commonly found in the same habitats as L. australis, but it is larger and distinctively marked, and while there is overlap in the flight periods of the two species, the onset of the flight season of L. australis is earlier by more than a week. Specimens of L. australis arrive suddenly and fully mature, as would be expected with migration or extreme vagrancy. However, we cannot dismiss the possibility of some successful reproduction in Wisconsin, or even the existence of some resident populations. Successful reproduction is expected to be hard to confirm because of the previously noted difficulty of identifying the species, especially when teneral, or as nymphs or exuviae.
70
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1
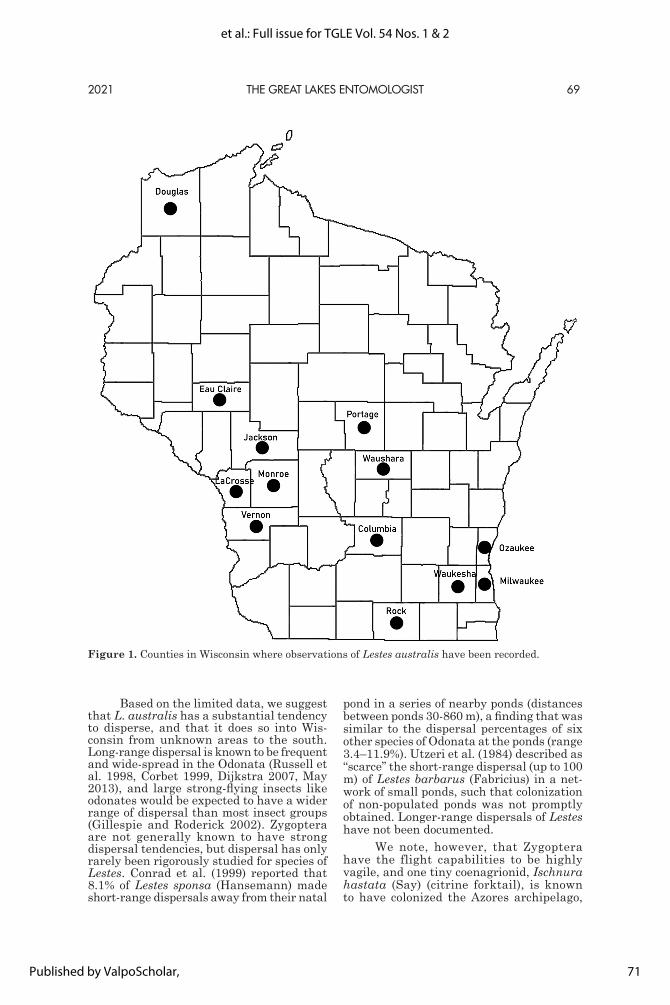
2021 THE GREAT LAKES ENTOMOLOGIST 69
Based on the limited data, we suggest that L. australis has a substantial tendency to disperse, and that it does so into Wis-consin from unknown areas to the south. Long-range dispersal is known to be frequent and wide-spread in the Odonata (Russell et al. 1998, Corbet 1999, Dijkstra 2007, May 2013), and large strong-flying insects like odonates would be expected to have a wider range of dispersal than most insect groups (Gillespie and Roderick 2002). Zygoptera are not generally known to have strong dispersal tendencies, but dispersal has only rarely been rigorously studied for species of Lestes. Conrad et al. (1999) reported that 8.1% of Lestes sponsa (Hansemann) made short-range dispersals away from their natal
pond in a series of nearby ponds (distances between ponds 30-860 m), a finding that was similar to the dispersal percentages of six other species of Odonata at the ponds (range 3.4–11.9%). Utzeri et al. (1984) described as “scarce” the short-range dispersal (up to 100 m) of Lestes barbarus (Fabricius) in a net-work of small ponds, such that colonization of non-populated ponds was not promptly obtained. Longer-range dispersals of Lestes have not been documented.
We note, however, that Zygoptera have the flight capabilities to be highly vagile, and one tiny coenagrionid, Ischnura hastata (Say) (citrine forktail), is known to have colonized the Azores archipelago,
Figure 1. Counties in Wisconsin where observations of Lestes australis have been recorded.
71
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

70 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
which is located about 1,500 km from the nearest mainland, and it has been captured at an altitude of 300 m in nets deployed from aircraft (Cordero Rivera et al. 2005). We sug-gest that the movements of Zygoptera might often go undetected because they are small and hard to see at most distances, and some evidently move at altitudes where observers are not usually looking. Holt (2003) noted the probability in most species of animals of genetic variation in niche requirements and in dispersal tendencies among individuals. While dispersal confers advantages for colo-nizing new habitats and maintaining genetic diversity (Smith et al. 2009), it also leads to individuals temporarily occupying habitats in which their niche requirements are not ad-equately met (known as “sink” populations). This could be the case when L. australis adults disperse into Wisconsin, because the eggs or nymphs might not be adapted to survive the long, cold winters. If the tenerals found at the Holmen pond on 24 July 2020 were indeed L. australis as is likely the case, then egg and nymph development could have been completed in as little as about 8 weeks. It is possible that L. australis is expanding its range northward in response to ecological factors like climate change or spatial varia-tions in environmental conditions. Grewe et al. (2013) found that lentic species of some European Odonata have expanded their range boundaries northward by an average of 115 km per decade, but that lotic species in the same southern European group have not, on average, changed their boundaries. They concluded that lentic species are bet-ter adapted to disperse than lotic species because their habitats are less persistent in time and space. Dispersal could also result from shifts in abundances of interacting species in source habitats, or because of internal traits like tendencies to disperse or recent adaptations to niche characteristics (Holt 2003).
Lestes australis is not the only species of Zygoptera having some individuals that make occasional northern forays into Wis-consin from primary breeding areas to the south, as I. hastata evidently behaves in similar fashion, although with the difference that successful reproduction in Wisconsin of that species has been documented (Tennes-sen 2011). The term “facultative migrant” has sometimes been used to describe odo-nates that do not have annual, directed mi-gration flights in the classical sense, but that disperse north only in years when drought conditions affect habitats within their usual breeding ranges, or when prolonged souther-ly winds facilitate long-distance northward transport (Soltesz et al. 1995, Russell et al. 1998). Short-distance flights of odonates have been called seasonal refuge flights (Corbet
1999, May 2013) and occasional or irregular flights have been called irruptions (Donnelly 2003, May 2013). In any case, gaining more knowledge about the movement patterns of widely dispersing or transient species of Odonata could benefit our ability to protect any habitats that might be valuable in their life history. We therefore urge observers to report Wisconsin records of Odonata to the Wisconsin Odonata Survey http://wiatri.net/inventory/odonata/ and to OdonataCentral https://www.odonatacentral.org.
Acknowledgments
We thank J. & C. Anderson, J. Cal-laghan, R. Chrouser, K. Huebner, D. Jack-son, S. LaValley, C. Mankiewicz, and F. Van den Broek for reporting their observations of L. australis to the Wisconsin Odonata Survey. The Bureau of Natural Heritage Conservation of the Wisconsin Department of Natural Resources supplied research funding for RBD.
Literature Cited
Abbott, J. C. 2011. Damselflies of Texas: a field guide. University of Texas Press, Austin.
Catling, P. M. 2003. The Austral Spreadwing, Lestes australis, in Ontario. Ontario Odonata 4: 18–22.
Conrad, K. F., K. H. Willson, I. F. Harvey, C. J. Thomas, and T. N. Sherratt. 1999. Dispersal characteristics of seven odonate species in an agricultural landscape. Ecog-raphy 22: 524–531.
Corbet, P. S. 1999. Dragonflies: Behavior and Ecology of Odonata. Cornell University Press, Ithaca, NY.
Cordero Rivera, A., M. O. Lorenzo Carballa, C. Utzeri, and V. Vieira. 2005. Partheno-genetic Ischnura hastata (Say), widespread in the Azores (Zygoptera: Coenagrionidae). Odonatologica 34: 1–9.
Dijkstra, K.-D. B. 2007. Gone with the wind: westward dispersal across the Indian Ocean and island speciation in Hemicordulia dragonflies (Odonata: Corduliidae). Zootaxa 1438: 27–48.
Donnelly, T. W. 1992. The Odonata of New York. Bulletin of American Odonatology 1(1): 1–27.
Donnelly, N. 2003. Lestes disjunctus, forcipatus, and australis: a confusing complex of North American damselflies. Argia 15(3): 10–13.
DuBois, R. 2019. Damselflies of Minnesota, Wisconsin & Michigan. Kollath+Stensaas Publishing, Duluth, MN.
Gillespie, R. G. and G. K. Roderick. 2002. Arthropods on islands: colonization, specia-
72
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

2021 THE GREAT LAKES ENTOMOLOGIST 71
tion, and conservation. Annual Review of Entomology 47: 595–632.
Grewe, Y., C. Hof, D. M. Dehling, R. Brandl, and M. Brändle. 2013. Recent range shifts of European dragonflies provide support for an inverse relationship between habitat predictability and dispersal. Global Ecology and Biogeography 22: 403–409.
Holt, R. D. 2003. On the evolutionary ecology of species’ ranges. Evolutionary Ecology Research 5: 159–178.
Lam, E. 2004. Damselflies of the Northeast. Bio-diversity Books, Forest Hills, NY.
May, M. L. 2013. A critical review of progress in studies of migration of dragonflies (Odo-nata: Anisoptera), with emphasis on North America. Journal of Insect Conservation 17(1): 1–15.
Montgomery, B. E. 1941. Records of Indiana dragonflies. X. 1937–1940. Proceedings of the Indiana Academy of Science 50: 229–241.
Paulson, D. 2004. New common names for some North American Odonata. Argia 16(3): 29–30.
Paulson, D. 2011. Dragonflies and Damselflies of the East. Princeton University Press, Princeton, NJ.
Russell, R. W., M. L. May, K. L. Soltesz, and J. W. Fitzpatrick. 1998. Massive swarm migrations of dragonflies (Odonata) in
eastern North America. American Midland Naturalist 140: 325–342.
Smith, R. F., L. C. Alexander, and W. O. Lamp. 2009. Dispersal by terrestrial stages of stream insects in urban watersheds: a synthesis of current knowledge. Journal of the North American Benthological Society 28(4): 1022–1047.
Soltesz, K., B. Barber, and G. Carpenter. 1995. A spring dragonfly migration in the Northeast. Argia 7(3): 10–14.
Tennessen, K. 2011. Follow-up search for Ischn-ura hastata (Citrine Forktail) in Wisconsin (or “What a difference a year makes”). Argia 23(4): 20–22.
Utzeri, C., G. Charchini, E. Falchetti, and C. Belfiore. 1984. Philopatry, homing and dispersal in Lestes barbarus (Fabricius) (Zygoptera: Lestidae). Odonatologica 13(4): 573–584.
Walker, E. M. 1952. The Lestes disjunctus and forcipatus complex (Odonata: Lestidae). Transactions of the American Entomological Society 78: 59–74.
Walker, E. M. 1953. The Odonata of Canada and Alaska, Volume I. University of Toronto Press, London, Ontario.
Westfall, M. J. and M. L. May. 2006. Damselflies of North America (revised edition). Scientific Publishers, Inc., Gainesville, FL.
73
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

72 THE GREAT LAKES ENTOMOLOGIST Vol. 54, Nos. 1–2
74
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1

INSTRUCTIONS FOR AUTHORSSUBJECTS
Papers dealing with any aspect of entomology will be considered for publication in The Great Lakes Entomologist. Appropriate subjects are those of interest to professional and amateur entomologists in the North Central States and Canada, as well as general papers and revisions directed to a larger audience while retaining an interest to readers in our geographic area. All manuscripts are refereed by at least two reviewers.
SUBMISSION REQUIREMENTSManuscripts must be typed in English with line numbers, double-spaced, with 1” margins and uploaded. Please use italics rather than underline.
Use subheadings sparingly and set them into paragraphs in boldface.Footnotes (except for authors’ addresses, which must be on the title page, and treated as a footnote), legends, and captions of illustrations
should be on separate pages.Titles should be concise, identifying the order and family discussed. The author of insect species must be given fully at least once in the abstract
and text, but not in the title. If a common name is used for a species or group, it should be in accordance with the common names published by the Entomological Society of America, or the Entomology Society of Canada.
The format for references must follow that described in the style guidelines used by the Entomological Society of America, except that journal titles are not abbreviated.
Please make sure that all references cited in the text appear in the Literature Cited section, and that all references in the Literature Cited appear in the body of the paper.
FIGURES AND TABLESPhotographs should be high resolution digital files (300 dpi). Drawings, charts, graphs, and maps must be scaled to permit proper reduction
without loss of detail.Figures must meet the following criteria: (1) gray-scale images must be submitted with a resolution of at least 300 dpi ; (2) line art or graphs
must be sent as 600 dpi.Scanned images should be saved in the native application. Never embed the images in a word-processing document.Captions for figures should be numbered consecutively and typed in order at the end of the manuscript. Captions should not be attached to
illustrations.Tables should be kept as uncluttered as possible, and should fit normally across a page when typeset.Tables cannot be submitted as Excel files or graphics, but only as text. Contributors should follow the Council of Biology Editors Style Manual,
7th ed., and examine recent issues of The Great Lakes Entomologist for proper format of manuscripts.The organizational format for a manuscript is as seen in the recent issues of the journal: TITLE, Author(s), Abstract, Introduction, Methods &
Materials, Results, Discussion, Acknowledgments, Literature Cited, Tables, and a List of Figures.Do not use extra spaces between paragraphs or references in the Lit. Cited.The columns of text in tables should be aligned with TABS, not spaces.Some symbols may not translate properly from one computer system to another. Do not use extbols.
PAGE CHARGES Papers published in The Great Lakes Entomologist are subject to a page charge of $20 per published page for non-members and $5 per published
page for members. Color images can be included at no additional charge. Members of the Society, who are authors without funds from grants, institutions, or industry, and are unable to pay costs from personal funds, may apply to the Society for financial assistance. Application for subsidy must be made at the time a manuscript is initially submitted for publication. Authors will receive a page proof, together with a page charge form. Reprints will be provided as PDF files. Extensive changes to the proof by the author will be billed at a rate of $1.00 per line.
COVER ARTWORK Cover art or photographs are desired for upcoming issues. They are published free of charge. We only require that they be suitably prepared
as described for images above, and that the subject be identified as accurately as possible.
ATTRIBUTION AND USE POLICIESReproduction, posting, transmission or other distribution or use of the article or any material therein, in any medium as permitted by a person-
al-use exemption or by written agreement of The Great Lakes Entomologist, requires credit to The Great Lakes Entomologist as original publisher (e.g., The Great Lakes Entomologist © 2017).
Personal-use Exceptions: The following uses are always permitted to the author(s) and do not require further permission from ValpoScholar provided the author does not alter the format or content of the articles, including the copyright notification:
• Posting on ResearchGate, Academia.edu, or a similar website;• Storage and back-up of the article on the author’s computer(s) and digital media (e.g., diskettes, back-up servers, Zip disks, etc.), provided
that the article stored on these computers and media is not readily accessible by persons other than the author(s);• Posting of the article on the author(s) personal website, provided that the website is non-commercial;• Posting of the article on the internet as part of a non-commercial open access institutional repository or other non-commercial open access
publication site affiliated with the author(s)’s place of employment (e.g., a Entomology professor at the University of the Great Lakes can have her article appear in the University of the Great Lakes’s Department of Entomology online publication series); and
• Posting of the article on a non-commercial course website for a course being taught by the author at the university or college employing the author.People seeking an exception, or who have questions about use, should contact the editors.
GENERAL TERMS AND CONDITIONS OF USEUsers of the ValpoScholar website and/or software agree not to misuse the ValpoScholar service or software in any way.The failure of ValpoScholar to exercise or enforce any right or provision in the policies or the Submission Agreement does not constitute a waiver
of such right or provision. If any term of the Submission Agreement or these policies is found to be invalid, the parties nevertheless agree that the court should endeavor to give effect to the parties’ intentions as reflected in the provision, and the other provisions of the Submission Agreement and these policies remain in full force and effect. These policies and the Submission Agreement constitute the entire agreement between ValpoScholar and the Author(s) regarding submission of the Article.
EDITOR’S ADDRESS All manuscripts for The Great Lakes Entomologist should be submitted via the online system at http://scholar.valpo.edu/tgle/ . Any questions
regarding submissions should be sent to the Lead Scientific Editor, Kristi Bugajski, Valparaiso University, 1610 Campus Drive East Valparaiso, IN 46383. (email: [email protected]).
OTHER BUSINESS Other correspondence should be directed to the Secretary, Michigan Entomological Society, 2104 Needham Rd., Ann Arbor, MI 48104.
75
et al.: Full issue for TGLE Vol. 54 Nos. 1 & 2
Published by ValpoScholar,

The Great Lakes EntomologistPublished by the Michigan Entomological Society
Volume 54 Nos. 1 & 2ISSN 0090-0222
Table of Contents
Influence of Age on Decision Making by Ovipositing Pieris rapae (Lepidoptera: Pieridae)
Brett Breitkreutz, Tracy L.E. Wagner, and Rodrigo J. Mercader ...................... 1Susceptibility, Preference, and Suitability of Carpinus and Ostrya Taxa for
Gypsy Moth Larvae (Lepidoptera: Lymantriidae)Fredric Miller and Susan Wiegrefe ............................................................. 7
Spiraling Flight Behavior May Integrate the Biological Compass Systems of Migratory North American Monarch Butterflies, (Danaus plexippus L.)
Matthew M. Douglas ............................................................................... 22Agricultural Natural Enemies Benefit Predominantly from Broader Scales of
Environmental Heterogeneity: A Quantitative ReviewAaron L. Iverson, Robyn J. Burnham, and John H. Vandermeer .................. 30
Plant Extract Efficacy on Mosquito Mortality: Preliminary Studies on the Effect of Ailanthus altissima Extract on Adult Aedes aegypti and Culex quinquefasciatus
John R. Wallace, Calen D. Wylie, and Ryan L. Wagner ..SCIENTIFIC NOTE 46Anomalously Pale-Haired Specimens in Three Genera of Cleptoparasitic
Bees (Hymenoptera: Apidae: Nomadinae)Thomas M. Onuferko ...................................................SCIENTIFIC NOTE 53
Bark- and Wood-Infesting Insects (Coleoptera and Hymenoptera) and Associated Parasitoids Reared from Yellow Birch (Betula alleghaniensis) in Ingham County, Michigan
Robert A. Haack .........................................................SCIENTIFIC NOTE 58New Record of Idana Marginata (Say) (Diptera: Tephritoidea: Ulidiidae)
from WisconsinDaniel K. Young ...............................................ENTOMOLOGICAL NOTE 64
The Puzzling Presence of Lestes australis (Odonata: Lestidae) in Wisconsin—Does This Species Migrate?
Robert B. DuBois and Kenneth J. Tennessen ........ENTOMOLOGICAL NOTE 67
Cover photoIdana marginata (Say) (Focus-stacked digital image)
Photo by Craig Brabant
76
The Great Lakes Entomologist, Vol. 54, No. 1 [], Art. 1
https://scholar.valpo.edu/tgle/vol54/iss1/1




















