BAHIA ANÁLISE & DADOS @BULLET v.22 @BULLET n.4 @BULLET OUT./DEZ. 2012 ISSN 0103 8117
Upload
independentCategory
view
0download
0
FL44CH15-Siggers ARI 18 November 2011 14:2
Fluid Mechanics of the EyeJennifer H. Siggers and C. Ross EthierDepartment of Bioengineering, Imperial College London, London SW7 2AZ,United Kingdom; email: [email protected], [email protected]
Annu. Rev. Fluid Mech. 2012. 44:347–72
First published online as a Review in Advance onOctober 17, 2011
The Annual Review of Fluid Mechanics is online atfluid.annualreviews.org
This article’s doi:10.1146/annurev-fluid-120710-101058
Copyright c© 2012 by Annual Reviews.All rights reserved
0066-4189/12/0115-0347$20.00
Keywords
porous medium, buoyancy-driven flow, deformed sphere, glaucoma, drugdelivery
Abstract
Fluid mechanical processes are an intrinsic part of several aspects of thephysiology and pathology of the human eye. In this article we describe se-lected phenomena that are amenable to particularly interesting mathemati-cal, experimental, or numerical analyses. We initially focus on glaucoma, acondition often associated with raised intraocular pressure. The mechanicsin this disease is by no means fully understood, but we present some of themodeling work that provides a partial explanation. We next focus on otherfeatures of the dynamics of the two specialized ocular fluids: the aqueousand vitreous humors. With regard to the aqueous humor, we discuss prob-lems concerning the transport of heat and proteins and the hydration ofthe cornea. With regard to the vitreous humor, we discuss the possibility offlow, which occurs primarily as a result of saccades or motions of the eyeball.Finally, we describe a model of the degradation of Bruch’s membrane in theretina.
347
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Intraocular pressure(IOP): the pressurewithin the eye, whichis generated by theproduction anddrainage of theaqueous humor
1. INTRODUCTION
The eye is a remarkable organ, capable of transducing photons into neural signals with highefficiency under a wide range of operating conditions. The retina, containing the specializedcells that carry out this transduction process, is aided in its function by many supporting tissues.The development and proper function of this complex system depend critically on biomechanicalfactors, as has been summarized elsewhere (Ethier et al. 2004a). Here we focus specifically onthe fluid mechanical aspects of the eye, of which there are many. We begin with an overview ofrelevant anatomy and physiology.
There are a number of processes within the eye in which fluid flow is important. Perhaps themost evident of these are the production, circulation, and drainage of aqueous humor, a clear,colorless fluid that is secreted at a flow rate of 2 to 2.5 μL min−1 (Brubaker et al. 1989) by aspecialized tissue known as the ciliary processes, located just posterior to the iris (the colored partof the eye) (Figure 1). In view of the very small flow rates and modest dimensions, the flow ofaqueous humor is creeping and inertia can be neglected. The aqueous humor itself is a very diluteprotein solution and thus can be treated as Newtonian with viscosity nearly identical to that ofsaline (Beswick & McCulloch 1956, Moses 1979). It flows into and fills a small region anterior tothe lens but behind the iris, known as the posterior chamber (see Figure 2a), then passes anteriorlythrough the pupil (the aperture in the central part of the iris), and enters the anterior chamber,where it circulates while bathing the iris and the inner surface of the cornea (the clear part of theeye). Eventually the aqueous humor drains from the eye via specialized tissues located in the angleof the anterior chamber, where the iris, cornea, and sclera (the white part of the eye) meet (seealso Figure 2 and Sections 2.1 and 2.2). These specialized tissues have a significant hydrodynamicflow resistance, and the drainage of the aqueous humor out of the eye therefore requires that therebe a positive pressure within the eye itself, the so-called intraocular pressure (IOP).
Vortex vein
Sclera
Choroid
RPE
Retinal vessels
RetinaLens
Iris
Cornea
Opticnerve
Zonules
Ciliary processesCiliary muscle
Anteriorchamber
Figure 1Overview of a human eye with major anatomical structures identified. Abbreviation: RPE, retinal pigmentepithelium. Figure modified from Krey & Brauer (1998), copyright c© MSD SHARP & DOHME GMBHGermany with kind permission.
348 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
a b
Trabecularmeshwork
Ciliarybody
Iris
Posteriorchamber
CiliaryprocessesCiliarymuscle
Trabecularmeshwork
Schlemm’scanal
Schlemm’s canal
Collector channelsArterioles
Aqueous veins
Schlemm’scanal
Cornea
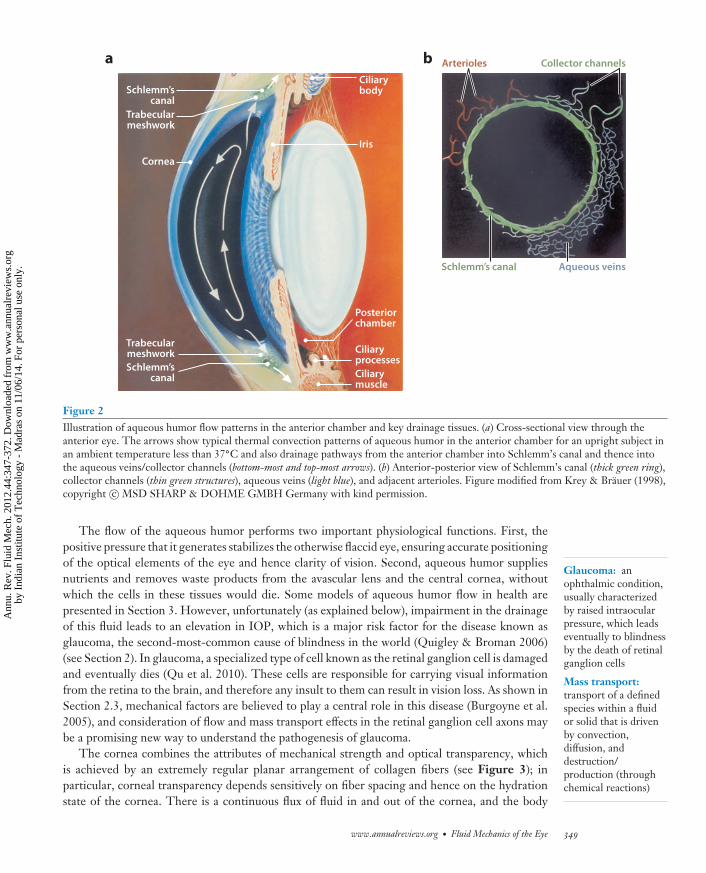
Figure 2Illustration of aqueous humor flow patterns in the anterior chamber and key drainage tissues. (a) Cross-sectional view through theanterior eye. The arrows show typical thermal convection patterns of aqueous humor in the anterior chamber for an upright subject inan ambient temperature less than 37◦C and also drainage pathways from the anterior chamber into Schlemm’s canal and thence intothe aqueous veins/collector channels (bottom-most and top-most arrows). (b) Anterior-posterior view of Schlemm’s canal (thick green ring),collector channels (thin green structures), aqueous veins (light blue), and adjacent arterioles. Figure modified from Krey & Brauer (1998),copyright c© MSD SHARP & DOHME GMBH Germany with kind permission.
Glaucoma: anophthalmic condition,usually characterizedby raised intraocularpressure, which leadseventually to blindnessby the death of retinalganglion cells
Mass transport:transport of a definedspecies within a fluidor solid that is drivenby convection,diffusion, anddestruction/production (throughchemical reactions)
The flow of the aqueous humor performs two important physiological functions. First, thepositive pressure that it generates stabilizes the otherwise flaccid eye, ensuring accurate positioningof the optical elements of the eye and hence clarity of vision. Second, aqueous humor suppliesnutrients and removes waste products from the avascular lens and the central cornea, withoutwhich the cells in these tissues would die. Some models of aqueous humor flow in health arepresented in Section 3. However, unfortunately (as explained below), impairment in the drainageof this fluid leads to an elevation in IOP, which is a major risk factor for the disease known asglaucoma, the second-most-common cause of blindness in the world (Quigley & Broman 2006)(see Section 2). In glaucoma, a specialized type of cell known as the retinal ganglion cell is damagedand eventually dies (Qu et al. 2010). These cells are responsible for carrying visual informationfrom the retina to the brain, and therefore any insult to them can result in vision loss. As shown inSection 2.3, mechanical factors are believed to play a central role in this disease (Burgoyne et al.2005), and consideration of flow and mass transport effects in the retinal ganglion cell axons maybe a promising new way to understand the pathogenesis of glaucoma.
The cornea combines the attributes of mechanical strength and optical transparency, whichis achieved by an extremely regular planar arrangement of collagen fibers (see Figure 3); inparticular, corneal transparency depends sensitively on fiber spacing and hence on the hydrationstate of the cornea. There is a continuous flux of fluid in and out of the cornea, and the body
www.annualreviews.org • Fluid Mechanics of the Eye 349
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
0.5 µm
Figure 3Normal human cornea of a 62-year-old male patient, showing the regular arrangement of collagen fiberswithin the corneal stroma. Figure taken from Langham, Maurice E., ed. The Cornea: MacromolecularOrganization of a Connective Tissue. Papers from a Symposium Held in Kyoto, Japan, 1967, under the Auspices of theDepartment of Ophthalmology, Osaka University, p. 124, figure 7-1. Copyright c© 1969 by The Johns HopkinsPress, reprinted with permission.
therefore has developed sophisticated mechanisms to control this transport. Specifically, water isactively and continually pumped out of the corneal stroma (the central layer of the cornea) bythe corneal endothelium, a specialized layer of cells lining the interior corneal surface. The neteffect is that the stroma is in a continual state of thermodynamic disequilibrium with respect toits adjacent bathing fluids, namely the tear film anteriorly and aqueous humor posteriorly (seeSection 3.4).
The majority of the ocular globe is filled by a clear, colorless, gel-like material known asvitreous humor, which occupies the vitreous chamber of the eye. The chamber is surrounded bytwo tissues, the retina and the choroid, the former of which comprises many layers (see Figure 4).Vitreous humor has complex viscoelastic properties, and although there have been several attemptsto characterize its properties experimentally (Lee et al. 1992, Nickerson et al. 2008, Swindle et al.2008, Zimmerman 1980), its rheology is not fully understood. It is known that the vitreous humorbecomes progressively liquefied with age; approximately 20% of the vitreous humor is liquid in14–18-year-olds, and this rises to more than 50% in subjects aged 80–90 years (Bishop 2000).In approximately 25%–30% of subjects, liquefaction can lead to a process in which the retinadetaches, risking loss of sight. In this context, it is also important to note that the eye is notnormally stationary, even when apparently focused on a fixed point. Instead, the eye constantlyexecutes a series of extremely rapid angular rotations (300◦ s−1 or more) known as saccades (Rayner
350 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
Viscoelastic fluid:a fluid whose stresstensor depends onboth the deformationand the rate ofdeformation of itsparticles
50 µm
Vitreous humor
Ganglion cell layer
Inner plexiform layer
Outer plexiform layer
Inner nuclear layer
Outer nuclear layer
Inner segments of photoreceptors
Outer segments of photoreceptorsRetinal pigment epithelium
Choroid
Sclera
Direction of light travel
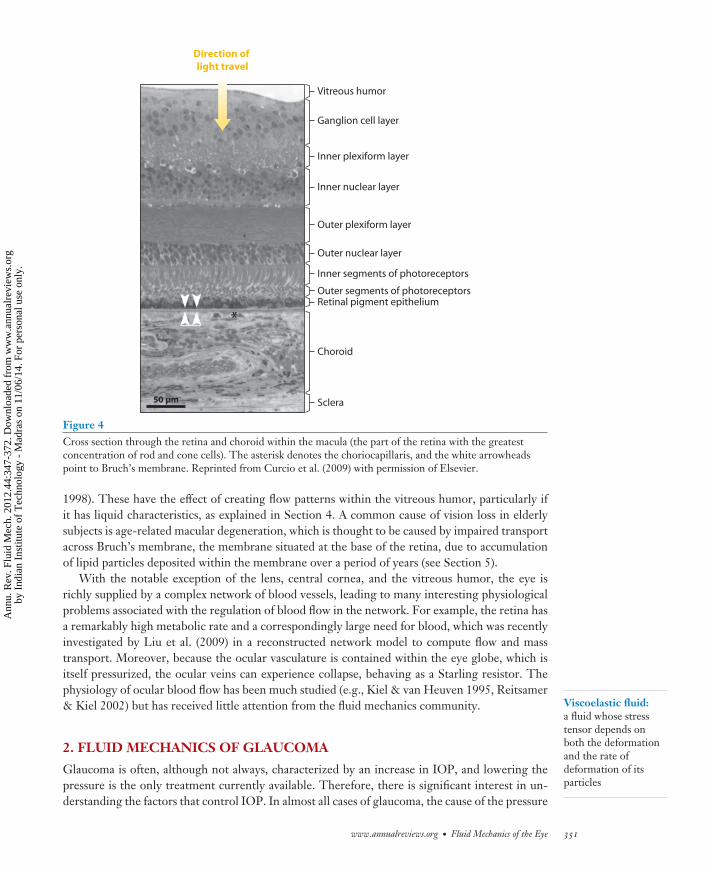
Figure 4Cross section through the retina and choroid within the macula (the part of the retina with the greatestconcentration of rod and cone cells). The asterisk denotes the choriocapillaris, and the white arrowheadspoint to Bruch’s membrane. Reprinted from Curcio et al. (2009) with permission of Elsevier.
1998). These have the effect of creating flow patterns within the vitreous humor, particularly ifit has liquid characteristics, as explained in Section 4. A common cause of vision loss in elderlysubjects is age-related macular degeneration, which is thought to be caused by impaired transportacross Bruch’s membrane, the membrane situated at the base of the retina, due to accumulationof lipid particles deposited within the membrane over a period of years (see Section 5).
With the notable exception of the lens, central cornea, and the vitreous humor, the eye isrichly supplied by a complex network of blood vessels, leading to many interesting physiologicalproblems associated with the regulation of blood flow in the network. For example, the retina hasa remarkably high metabolic rate and a correspondingly large need for blood, which was recentlyinvestigated by Liu et al. (2009) in a reconstructed network model to compute flow and masstransport. Moreover, because the ocular vasculature is contained within the eye globe, which isitself pressurized, the ocular veins can experience collapse, behaving as a Starling resistor. Thephysiology of ocular blood flow has been much studied (e.g., Kiel & van Heuven 1995, Reitsamer& Kiel 2002) but has received little attention from the fluid mechanics community.
2. FLUID MECHANICS OF GLAUCOMA
Glaucoma is often, although not always, characterized by an increase in IOP, and lowering thepressure is the only treatment currently available. Therefore, there is significant interest in un-derstanding the factors that control IOP. In almost all cases of glaucoma, the cause of the pressure
www.annualreviews.org • Fluid Mechanics of the Eye 351
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
increase is known to be an increase in the hydrodynamic resistance to aqueous humor drainage,necessitating a higher pressure to drive the outflow. Glaucoma can be further classified intoclosed-angle glaucoma and open-angle glaucoma. In closed-angle glaucoma, the iris moves ante-riorly from its normal position, reducing or eliminating the gap between it and the cornea. Thisis an interesting fluid-structure interaction problem and frequently leads to a complete occlu-sion of outflow with attendant dramatic increases in IOP; a mathematical model of this is brieflydiscussed in Section 3.1. In open-angle glaucoma, the drainage tissues remain accessible to theaqueous humor, but they present an elevated flow resistance. There are a number of different typesof open-angle glaucoma, the most common of which is known as primary open-angle glaucoma.This condition has puzzled researchers for many years, as there are no evident structural changesin the drainage tissues that could explain their elevated resistance. Here we discuss some of themodeling work that has attempted to shed light on the source of the resistance. Section 2.1 dis-cusses changes in the trabecular meshwork, and Section 2.2 focuses on changes in the mechanicsof flow into and through Schlemm’s canal. Section 2.3 discusses a possible mechanism for thedeath of retinal ganglion cells, which is the cause of vision loss in glaucoma.
2.1. Increase in Resistance Across the Trabecular Meshwork
To understand the fluid mechanics of aqueous humor drainage, we must provide further detailson the anatomy of the drainage tissues located in the angle of the anterior chamber (see alsoFigure 2). As the aqueous humor leaves the eye, it first passes through the trabecular meshwork,which can be represented as a biological porous material. It then enters a collecting duct known asSchlemm’s canal, notable for its unusual endothelial cellular lining containing a number of smallmicrometer-sized openings (known as pores). As discussed below, the hydrodynamic interactionof the flows through the pores and that through the trabecular meshwork are of great interest.The aqueous humor then flows along the canal and out through a drainage structure known as acollector channel, from which it eventually mixes with venous blood in the sclera and returns tothe right heart.
A significant amount of work has been devoted to understanding the drainage of aqueoushumor through these tissues. Originally, attention focused on analysis of the trabecular meshworkas a porous material (Ethier 1986, McEwen 1958, Seiler & Wollensak 1985). Micrographs ofnormal and diseased tissue were analyzed to estimate the tissue porosity, εTM , and the specificsurface, STM , and the hydraulic permeability, KTM , of the trabecular meshwork was estimatedusing classical Carman-Kozeny theory (Bear 1988),
KT M = ε3T M
kS2T M
, (1)
for suitable values of the Kozeny constant, k. The resulting computed permeabilities were com-pared with estimates from experimental measurements, with the surprising finding that the com-puted permeability was one to two orders of magnitude higher than the best experimental data.The conclusion was that either the trabecular meshwork had little hydraulic flow resistance or,more likely, the fundamental assumptions underlying the calculation were incorrect.
Some evidence indicates that the apparently open spaces within the trabecular meshwork are ac-tually filled with a gel-like biopolymer consisting of proteins and long-chain carbohydrates. It wastherefore appropriate to include the hydrodynamic effects of this gel material, achieved throughthe use of the fiber matrix model (Weinbaum 1998), in which the individual biopolymer strands areapproximated by long, randomly oriented cylinders. Through the use of classical low–Reynoldsnumber results (Happel 1959, Spielman & Goren 1968), it was possible to make good estimates
352 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Porous medium:a solid material whosesmall-scale structure ischaracterized by poresfilled with fluid; flowin such materials isusually governed bythe Darcy equation
of the hydraulic permeability of a pure biopolymer gel or of tissues with well-characterized mi-crostructure (Ethier 1986). Application of this theory to the trabecular meshwork required theconsideration of a two-level hierarchical porous medium, in which a material (the gel) with charac-teristic length scales on the order of angstroms to nanometers and hydraulic permeability Kgel wasembedded within a second material (the trabecular meshwork) having pores with dimensions onthe order of micrometers to tens of micrometers. Accounting for steric hindrance and tortuosityeffects leads to a prediction of the overall permeability of the composite material as (Ethier 1986)
K = KgelεT M
2 − εT M. (2)
This produced a much more satisfactory agreement between theory and result, leading to whatis now known as the gel-filled meshwork theory. However, subsequent experimental data havecalled this model into question; for example, treatment of the trabecular meshwork with enzymesthat are known to degrade the biopolymer gel seems to have a much smaller effect on hydraulicresistance than would be expected (Hubbard et al. 1997).
The fluid mechanics of aqueous humor drainage through the small pores in the cellular liningof Schlemm’s canal is also important (Bill & Svedbergh 1972, Eriksson & Svedbergh 1980). Byexploiting the fact that these pores are small and relatively isolated, one needs only to consider flowthrough a single pore and treat the entire cellular lining as a parallel network of hydrodynamicallynoninteracting pores. The flow resistance of a single pore of radius R can be calculated fromSampson’s theory in low–Reynolds number hydrodynamics (Happel & Brenner 1983), whichrelates the pressure drop, �p, across a thin surface to the volumetric flow through the pore, q,through the surface via
q�p
= R3
3μ, (3)
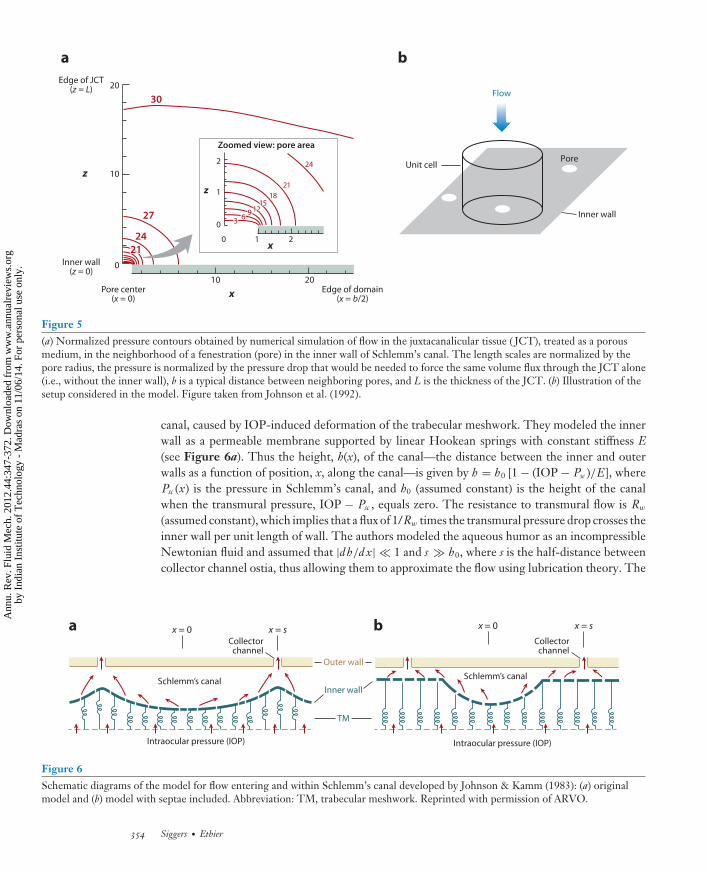
where μ is the dynamic viscosity of the aqueous humor, and �p is the pressure difference acrossthe surface between locations infinitely far from the surface on either side of the surface. Thesurprising finding from this calculation was that it predicts an extremely low flow resistance ofthe endothelial lining of Schlemm’s canal, certainly much lower than the observed resistance ofthe entire system. This led to a paradox: Neither the lining of Schlemm’s canal nor the trabecularmeshwork alone seemed to offer sufficient flow resistance to agree with experimental evidence.A possible resolution of this paradox was put forward by Johnson et al. (1992), who noted aninteresting hydrodynamic interaction between the endothelial lining of Schlemm’s canal and theupstream porous material of the trabecular meshwork. Specifically, because the pores are few andaccount for only a small fraction of the total area of the inner wall of Schlemm’s canal, they mustact to hydrodynamically focus λ (or funnel λ) aqueous humor drainage through the trabecularmeshwork (see Figure 5). The situation was modeled as a porous slab bounded on one surface bya plate pierced by hydrodynamically isolated pores. By considering a simplified unit-cell model,consisting of a single pore and the upstream region of the porous medium drained by that pore,one can calculate the hydraulic resistance of the ensemble structure in a straightforward manner.
The theory predicts an overall flow resistance that is generally consistent with experimentalmeasurements (Overby et al. 2009). Furthermore, it reconciles observations that both pore densityand the composition of the trabecular meshwork have an effect on the overall resistance to flowin this tissue.
2.2. Flow Within Schlemm’s Canal
In a different study aimed at identifying the source of outflow resistance, Johnson & Kamm (1983)developed a model to consider the hydrodynamic effects of the partial or total collapse of Schlemm’s
www.annualreviews.org • Fluid Mechanics of the Eye 353
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
20
30
27
24
Edge of JCT(z = L)
Inner wall(z = 0)
Pore center(x = 0)
Zoomed view: pore area
Edge of domain(x = b/2)
x
x
z
z
10
0
0
18
24
21
0 1 2
1
2
10 20
21
69 122
15
3
Pore
Inner wall
Unit cell
Flow
a b
Figure 5(a) Normalized pressure contours obtained by numerical simulation of flow in the juxtacanalicular tissue ( JCT), treated as a porousmedium, in the neighborhood of a fenestration (pore) in the inner wall of Schlemm’s canal. The length scales are normalized by thepore radius, the pressure is normalized by the pressure drop that would be needed to force the same volume flux through the JCT alone(i.e., without the inner wall), b is a typical distance between neighboring pores, and L is the thickness of the JCT. (b) Illustration of thesetup considered in the model. Figure taken from Johnson et al. (1992).
canal, caused by IOP-induced deformation of the trabecular meshwork. They modeled the innerwall as a permeable membrane supported by linear Hookean springs with constant stiffness E(see Figure 6a). Thus the height, h(x), of the canal—the distance between the inner and outerwalls as a function of position, x, along the canal—is given by h = h0 [1 − (IOP − Psc )/E], wherePsc (x) is the pressure in Schlemm’s canal, and h0 (assumed constant) is the height of the canalwhen the transmural pressure, IOP − Psc , equals zero. The resistance to transmural flow is Rw
(assumed constant), which implies that a flux of 1/Rw times the transmural pressure drop crosses theinner wall per unit length of wall. The authors modeled the aqueous humor as an incompressibleNewtonian fluid and assumed that |dh/d x| � 1 and s � h0, where s is the half-distance betweencollector channel ostia, thus allowing them to approximate the flow using lubrication theory. The
Collectorchannel
Schlemm’s canal
TM
Outer wall
Inner wall
Intraocular pressure (IOP)
x = 0 x = s
Schlemm’s canal
Intraocular pressure (IOP)
x = 0 x = sa bCollectorchannel
Figure 6Schematic diagrams of the model for flow entering and within Schlemm’s canal developed by Johnson & Kamm (1983): (a) originalmodel and (b) model with septae included. Abbreviation: TM, trabecular meshwork. Reprinted with permission of ARVO.
354 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
governing equations reduced to a single equation for h:
12μ
wRw
(h − h0) = dd x
(h3 dh
d x
), (4)
where w is the width of the cross section of Schlemm’s canal in the anterior-posterior direction.Boundary conditions arise from prescribing the pressure at the collector channels, which, usingthe spring condition given above, leads to
h = h0
(1 − IOP − Pc c
E
), (5)
at x = s, where Pcc is the pressure in the collector channels. They solved the governingEquation 4 numerically over the half-distance between neighboring collector channels (i.e., theregion 0 ≤ x ≤ s in Figure 6), imposing a symmetry condition at x = 0.
The model predicts a nonlinear dependence between the total outflow and the pressure drop,IOP− Pc c , due to an increase in resistance to outflow. The nonlinearity occurs because the heightof Schlemm’s canal decreases when the pressure drop is large, increasing the canal’s resistanceand hence the overall resistance. Thus, to maintain a constant outflow to meet physiologicaldemands, the IOP must increase more than it would if the dependence were linear. Another causeof increased IOP could be that the inner wall resistance, Rw, increases, while the other parametersmaintain constant values, which would mean the IOP must also increase to maintain the outflow.For a normal or slightly raised IOP (up to approximately 25 mmHg), and using parameter valuessuitable for the human eye, the predicted height of the canal is approximately spatially uniform, andthe predicted resistance to outflow is approximately constant as IOP increases. The approximatelyuniform height of Schlemm’s canal implies an approximately spatially uniform luminal pressure,which shows that the majority of total outflow resistance does not derive from the resistance to flowwithin Schlemm’s canal. For higher IOPs, the canal begins to collapse near the collector channels,and at IOP = 29 mmHg, there is complete collapse. However, complete or almost-completecollapse is unrealistic because Schlemm’s canal contains septae, short structures modeled as beingof height hs, which protrude into Schlemm’s canal from the outer wall to prevent complete collapse.
With a normal or slightly raised IOP, these protrusions make no difference to the model resultsbecause the minimum width of the canal is greater than hs. For higher IOPs, there is partial collapseof the canal with the septae supporting the collapsed region (see Figure 6b). For still-higher IOPs,the channel is completely supported by the septae and has constant height hs, and under theseconditions the total outflow resistance is constant. In a further extension, the authors consideredcompliant septae, meaning that the height can drop below hs in the collapsed region, increasingthe resistance of Schlemm’s canal compared with the rigid septae model. A comparison of modelresults with experimental data suggests that Schlemm’s canal collapse does not occur in glaucoma,implying that glaucoma cannot be caused by weakening of the trabecular meshwork alone.
A related question concerns the flow within the lumen of Schlemm’s canal, as opposed to thatacross the endothelial lining of the canal. In the vascular system, which shares many biologicalsimilarities with Schlemm’s canal, the caliber of vessels is strongly influenced by the shear stressexerted by the blood flowing within the vessel. However, because the volumetric flow rates inthe lumen of Schlemm’s canal are minuscule, shear stresses might, a priori, also be expectedto be too small to have physiological impact. However, a simple calculation in which the crosssection of Schlemm’s canal was treated as an ellipse showed that the estimated shear stresseswere similar to those observed in the vascular system (Ethier et al. 2004b), suggesting that thebiological mechanisms for caliber regulation in the two systems could be similar. This is potentiallyimportant, as Schlemm’s canal is observed to be shorter in the anterior-posterior direction in
www.annualreviews.org • Fluid Mechanics of the Eye 355
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
glaucomatous eyes, even though its height h is unaffected (Allingham et al. 1996). This increasesoutflow resistance, and thus dysregulation of the mechanisms controlling Schlemm’s canal calibermay play a role in glaucoma.
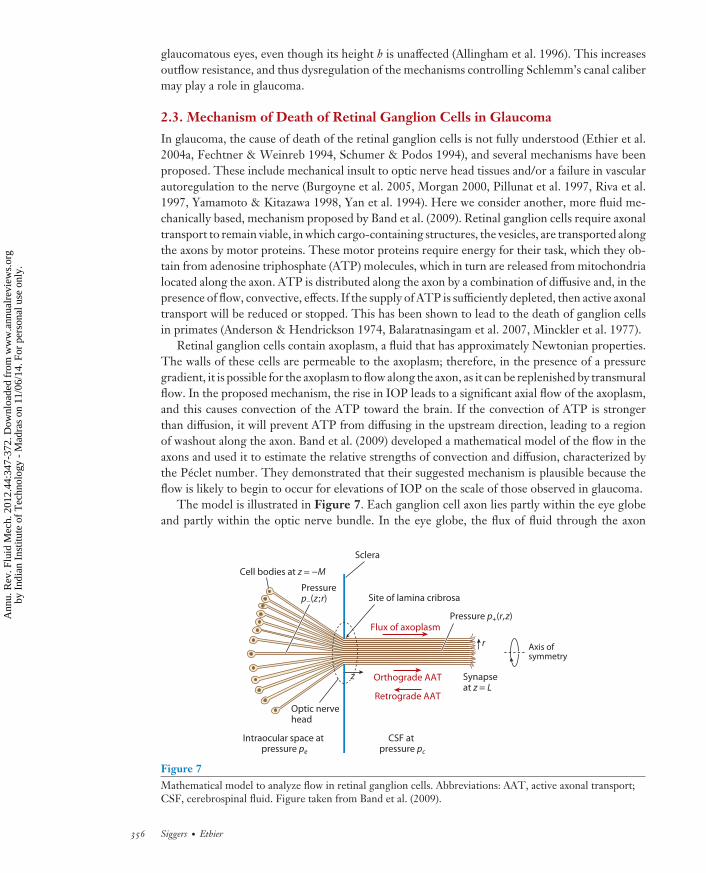
2.3. Mechanism of Death of Retinal Ganglion Cells in Glaucoma
In glaucoma, the cause of death of the retinal ganglion cells is not fully understood (Ethier et al.2004a, Fechtner & Weinreb 1994, Schumer & Podos 1994), and several mechanisms have beenproposed. These include mechanical insult to optic nerve head tissues and/or a failure in vascularautoregulation to the nerve (Burgoyne et al. 2005, Morgan 2000, Pillunat et al. 1997, Riva et al.1997, Yamamoto & Kitazawa 1998, Yan et al. 1994). Here we consider another, more fluid me-chanically based, mechanism proposed by Band et al. (2009). Retinal ganglion cells require axonaltransport to remain viable, in which cargo-containing structures, the vesicles, are transported alongthe axons by motor proteins. These motor proteins require energy for their task, which they ob-tain from adenosine triphosphate (ATP) molecules, which in turn are released from mitochondrialocated along the axon. ATP is distributed along the axon by a combination of diffusive and, in thepresence of flow, convective, effects. If the supply of ATP is sufficiently depleted, then active axonaltransport will be reduced or stopped. This has been shown to lead to the death of ganglion cellsin primates (Anderson & Hendrickson 1974, Balaratnasingam et al. 2007, Minckler et al. 1977).
Retinal ganglion cells contain axoplasm, a fluid that has approximately Newtonian properties.The walls of these cells are permeable to the axoplasm; therefore, in the presence of a pressuregradient, it is possible for the axoplasm to flow along the axon, as it can be replenished by transmuralflow. In the proposed mechanism, the rise in IOP leads to a significant axial flow of the axoplasm,and this causes convection of the ATP toward the brain. If the convection of ATP is strongerthan diffusion, it will prevent ATP from diffusing in the upstream direction, leading to a regionof washout along the axon. Band et al. (2009) developed a mathematical model of the flow in theaxons and used it to estimate the relative strengths of convection and diffusion, characterized bythe Peclet number. They demonstrated that their suggested mechanism is plausible because theflow is likely to begin to occur for elevations of IOP on the scale of those observed in glaucoma.
The model is illustrated in Figure 7. Each ganglion cell axon lies partly within the eye globeand partly within the optic nerve bundle. In the eye globe, the flux of fluid through the axon
Intraocular space at pressure pe
CSF at pressure pc
Pressurep−(z;r)
Pressure p+(r,z)
r
Site of lamina cribrosa
Sclera
Synapse at z = L
Cell bodies at z = −M
Flux of axoplasm
Retrograde AAT
Orthograde AAT
Axis ofsymmetry
Optic nervehead
z
Figure 7Mathematical model to analyze flow in retinal ganglion cells. Abbreviations: AAT, active axonal transport;CSF, cerebrospinal fluid. Figure taken from Band et al. (2009).
356 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Homogenization:a technique used toanalyze multiple-scalesproblems; quantitiesare averaged over thesmall scale, leading tosimplified large-scaleequations
wall is assumed to be proportional to the local pressure drop across the axon wall. In the opticnerve, the axons are treated as a bundle of fibers whose cross sections form a hexagonal lattice,and flux between neighboring axons is also proportional to the pressure difference between thefluids in them. Flow along the length of the axon is driven by the axial pressure gradient. Theseassumptions were used to homogenize the model, leading to a relationship between the axial fluxand pressure gradient along the axons, which can be solved to find the flow and pressure in theform of sums of an infinite series of Bessel functions.
If the IOP is elevated to levels commonly seen in glaucoma, the Peclet number for ATPpredicted by the model is greater than one within substantial extraocular regions of the axons.This suggests there will be significant depletion of ATP in these regions, illustrating the potentialimportance of the proposed mechanism.
3. FLOW IN THE ANTERIOR CHAMBER
3.1. Thermal Transport
In the anterior chamber, the aqueous humor flows radially outward toward the trabecular mesh-work in the normal course of its drainage from the eye. In addition to this flow, there is also athermally driven flow, as temperature gradients exist between the anterior and posterior surfacesof the anterior chamber (the back of the cornea and the front of the iris). The posterior surface isclose to body temperature, but the anterior surface is closer to atmospheric temperature (usuallycooler). Convection is thought to increase the efficiency of nutrient delivery, but it is also likelyto give rise to significant clinical effects if there is particulate matter within the aqueous humor,such as blood cells or pigment particles (Canning et al. 2002).
Canning et al. (2002) and Fitt & Gonzalez (2006) developed a model of the fluid flow inthe anterior chamber of the eye, using the simplified geometry illustrated in Figure 8. Theytreated the fluid as incompressible and Newtonian and used the Boussinesq approximation todescribe the variations in the density of the fluid. The posterior surface consisted of a disc rep-resenting the pupil in the center and an annulus surrounding it representing the iris, which wasassumed to be at a fixed temperature T1 close to body temperature. The anterior surface of thechamber was assumed to be at a fixed cooler temperature T0. The velocity at the pupil was assumed
z
T = T0
T = T1
x
y
a
a
−a
−aPupilaperture
Iris: z = 0
Cornea: z = h(x,y)
Flow w0(x,y)
Figure 8Sketch of the model developed by Canning et al. (2002) to investigate flow in the anterior chamber.Reprinted with permission of Oxford University Press.
www.annualreviews.org • Fluid Mechanics of the Eye 357
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Buoyancy-drivenflow: flow driven byspatial densitygradients in the fluid,frequently due totemperature variationsin the fluid
to be purely normal to the plane of the iris and given by a prescribed function w0(x, y), with no-slipvelocity applied on the other boundaries, except that drainage occurs at the angles of the domain.
Canning et al. (2002) used experimental measurements to argue that the aspect ratio of themodel is small (h � a) (see Figure 8) and also estimated the reduced Reynolds number to besmall. In the formal mathematical limit in which these quantities are small, and also neglectingviscous dissipation, time dependence, and convection of heat, they derived a simplified system ofequations. They were able to manipulate these and reduce them to the single differential equation
∇H · (h3∇H P
) = −12μw0, (6)
where ∇H = ex∂/∂x + ey∂/∂y in Cartesian coordinates, P is the deviation in the pressure fromthe hydrostatic pressure profile, and μ is the viscosity at the temperature T0.
The authors estimated the size of w0 and concluded that, typically, it is likely to be smallcompared to the thermally driven velocity, and therefore they considered the case w0 = 0, whichallows an exact solution of Equation 6. Estimation of the stress induced by this flow suggests it isunlikely to be strong enough to cause particle or cell detachment from the iris. A comparison ofthe transit time of the solution with the time it would take a particle in the aqueous humor to settleunder gravity shows that the convective velocity is several times faster than the settling velocity.Calculation of the Stokes drag allows a full model of particle transport to be developed. The modelshowed that particles remain in the vertical plane that contains them and that is also perpendicularto the iris. The authors solved the model numerically and used their results to comment onfeatures that would be present in a number of clinical conditions. This work was extended byFitt & Gonzalez (2006) to include inflow (w0 �= 0), to consider other directions of gravity withrespect to the model (e.g., a supine patient), to investigate vibrations of the lens as the head oreye moves (phakodenesis), and to investigate the flow produced by rapid eye movements duringsleep. Their results showed that the buoyancy-driven flow typically exceeds the flow driven byother mechanisms by orders of magnitude and plays a dominant role in several medical conditionsof the anterior chamber.
3.2. Fluid-Structure Interaction Models of the Iris and Aqueous Humor
Other studies of aqueous humor flow include those by Heys et al. (2001) and Heys & Barocas(2002b). These authors developed an axisymmetric model of the flow in both the anterior andposterior chambers, in which they modeled the aqueous humor as a Newtonian viscous fluidand the iris as an incompressible neo-Hookean solid. The iris deforms as a result of the stressexerted by the flow, and the authors calculated the steady-state position of the iris tissue. Heys& Barocas (2002a) considered a fully three-dimensional model and included thermal convection.Their results showed that convection effects in the flow are dominant; that is, the calculated velocitymagnitude predicted by the equations with the convection terms added is many times larger thanthat predicted by the equations without convection included. Their results were consistent withclinical observations of Krukenberg’s spindle, a condition in which pigment from the iris becomesattached to the posterior surface of the cornea in characteristic vertical stripes. A similar model wasused to investigate the flow and deformation produced by small oscillations of the position of theiris (Huang & Barocas 2006) and the recovery from an indentation in the cornea or sclera (Amini& Barocas 2010), which was done by imposing an initial rotation in the iris position at its root.
In other work, Huang & Barocas (2004) adapted the model by adding an active term intoits stress tensor to represent contraction of the sphincter muscle in the normal circumferentialdirection. They tuned the geometry to model both normal eyes and eyes with features that arethought to be risk factors in closed-angle glaucoma. Their results predicted that the further forward
358 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
the lens position is, the greater the likelihood of iris-lens contact, which leads to a greater pressuredifference between the posterior and anterior chambers. Conversely, decreasing the diameter ofthe anterior chamber leads to a smaller angle between the iris and the cornea, which is likely toincrease the flow resistance of the aqueous outflow pathway. However, testing their model fordifferent pupil diameters suggests that the condition is most severe when the pupil is small. Thiscontradicts clinical tests that suggest the condition is most serious in dark environments when thepupil is large.
3.3. Transport of Proteins
As noted above, the aqueous humor is produced behind the iris, passes through the pupil, andfills the anterior chamber before draining from the eye. It is therefore natural to assume that theproteins within the aqueous humor would follow the same route. However, this is not the case;protein mass transport in the anterior chamber proceeds in a manner for which the details haveonly been elucidated over the past decade or so, despite the fact that the circulation of aqueoushumor has been reasonably well understood for a century.
The essential fact driving the difference between water and protein transport in the anterioreye is that the lining epithelium of the ciliary processes, the tissue responsible for secretion ofthe aqueous humor, is highly impermeable to proteins. Therefore, proteins naturally presentin the ciliary body are prevented from entering the posterior chamber and instead build up inthe extravascular space of the ciliary body to create a reservoir of plasma proteins. Proteins thendiffuse anteriorly from this reservoir, leaking into the anterior chamber from the anterior iris. Thismechanism has been confirmed by an elegant series of tracer studies in various species (Barsottiet al. 1992, Bert et al. 2006, Freddo et al. 1990), complemented by theoretical models of proteinmass transport with predictions that show a reasonable agreement with experimental data (Barsottiet al. 1992).
3.4. Dynamics of the Cornea
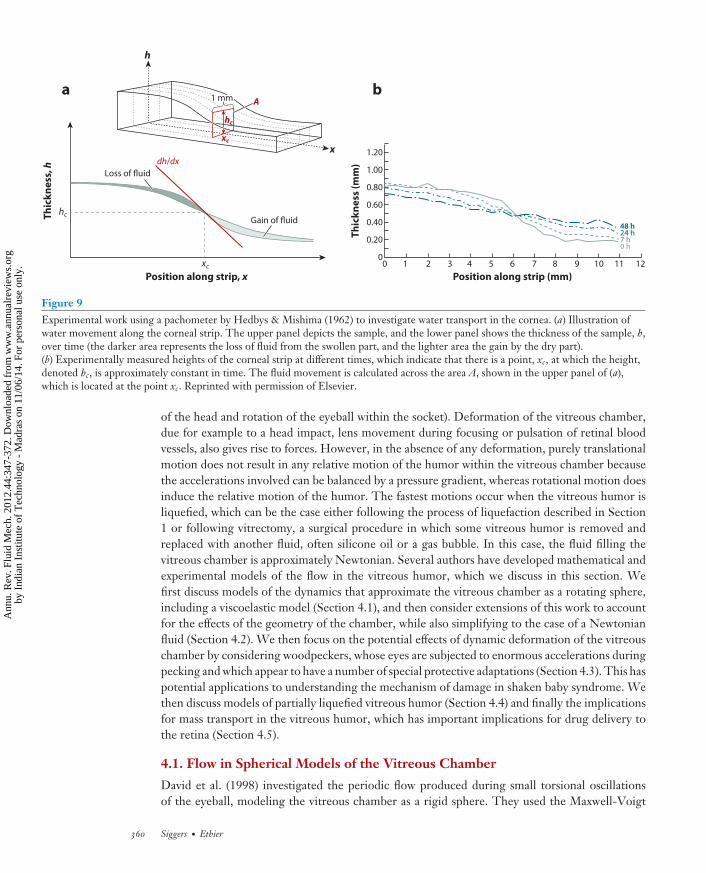
Hedbys & Mishima (1962) carried out early quantitative studies of fluid transport in the cornea.Their work is notable because it investigated water transport both across and in the plane ofthe corneal stroma and because of their clever experimental design for measuring transport inthe tangent plane of the stroma. They developed an optical pachometer capable of measuringdynamic thickness profiles of corneas and applied this to corneal samples in which the water hadbeen partially expelled from part of the cornea (see Figure 9). Conserving mass, relating cornealhydration to the local swelling pressure, and using Darcy’s law, they were able to deduce stromalpermeability as a function of hydration. They observed that the transport properties of the corneaare anisotropic, with a lower permeability for flow normal to the stroma than for that in the tangentplane, and this difference becomes more pronounced as the corneal hydration decreases.
These phenomena are understandable when the ultrastructure of the cornea is considered,in which arrays of collagen fibers are oriented largely parallel to the plane of the cornea (seeFigure 9). With the use of classical results for flow in porous media in the limit of vanishingfiber solid fraction, the permeability is predicted to be approximately twofold lower for flow inthe normal direction than that in the plane of the fibers (Happel & Brenner 1983), a result thatis quantitatively in agreement with the experimental data.
4. FLUID MECHANICS OF THE VITREOUS HUMOR
The vitreous cavity has an approximately spherical shape and contains the vitreous humor, whichis subject to mechanical forces as a result of the motion of the eyeball (due primarily to the motion
www.annualreviews.org • Fluid Mechanics of the Eye 359
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
xc
hc
h
xdh/dx
Position along strip, x
Thic
knes
s, h
hc
A
xc
1 mm1 mm1 mm
Loss of fluid
Gain of fluid
1.20
1.00
0.80
0.60
0.40
0.20
00 1 2 3 4 5 6
Position along strip (mm)
Thic
knes
s (m
m)
7 8 9 10 11 12
48 h24 h7 h0 h
a b
Figure 9Experimental work using a pachometer by Hedbys & Mishima (1962) to investigate water transport in the cornea. (a) Illustration ofwater movement along the corneal strip. The upper panel depicts the sample, and the lower panel shows the thickness of the sample, h,over time (the darker area represents the loss of fluid from the swollen part, and the lighter area the gain by the dry part).(b) Experimentally measured heights of the corneal strip at different times, which indicate that there is a point, xc, at which the height,denoted hc, is approximately constant in time. The fluid movement is calculated across the area A, shown in the upper panel of (a),which is located at the point xc. Reprinted with permission of Elsevier.
of the head and rotation of the eyeball within the socket). Deformation of the vitreous chamber,due for example to a head impact, lens movement during focusing or pulsation of retinal bloodvessels, also gives rise to forces. However, in the absence of any deformation, purely translationalmotion does not result in any relative motion of the humor within the vitreous chamber becausethe accelerations involved can be balanced by a pressure gradient, whereas rotational motion doesinduce the relative motion of the humor. The fastest motions occur when the vitreous humor isliquefied, which can be the case either following the process of liquefaction described in Section1 or following vitrectomy, a surgical procedure in which some vitreous humor is removed andreplaced with another fluid, often silicone oil or a gas bubble. In this case, the fluid filling thevitreous chamber is approximately Newtonian. Several authors have developed mathematical andexperimental models of the flow in the vitreous humor, which we discuss in this section. Wefirst discuss models of the dynamics that approximate the vitreous chamber as a rotating sphere,including a viscoelastic model (Section 4.1), and then consider extensions of this work to accountfor the effects of the geometry of the chamber, while also simplifying to the case of a Newtonianfluid (Section 4.2). We then focus on the potential effects of dynamic deformation of the vitreouschamber by considering woodpeckers, whose eyes are subjected to enormous accelerations duringpecking and which appear to have a number of special protective adaptations (Section 4.3). This haspotential applications to understanding the mechanism of damage in shaken baby syndrome. Wethen discuss models of partially liquefied vitreous humor (Section 4.4) and finally the implicationsfor mass transport in the vitreous humor, which has important implications for drug delivery tothe retina (Section 4.5).
4.1. Flow in Spherical Models of the Vitreous Chamber
David et al. (1998) investigated the periodic flow produced during small torsional oscillationsof the eyeball, modeling the vitreous chamber as a rigid sphere. They used the Maxwell-Voigt
360 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Steady streaming:the time average of afluctuating flow,arising because of anonconservative bodyforce, Reynoldsstresses, or boundaryeffects
viscoelastic model proposed by Lee et al. (1992) to characterize the rheological behavior of thevitreous humor. The angular displacement was modeled as ε cos ωt, and the assumption of smalloscillations allowed them to linearize the model and seek solutions proportional to e iωt . This led to alinear relationship between the shear stress and the shear strain, whose constant of proportionalitywas the complex modulus, G, dependent on ω. In terms of spherical polar coordinates (r, θ ,φ), centered on the sphere and with the axis parallel to the axis of the oscillations, the velocityfield is
u = − iεR3ω[sin(ar/R) − (ar/R) cos(ar/R)]2r2(sin a − a cos a)
sin θ eiωt eφ + c.c., (7)
where R is the radius of the sphere; a = αc e−iπ/4; αc is the complex Womersley number, given by
αc =√
iρω2 R2
G; (8)
ρ is the fluid density; eφ is the unit vector in the direction of increasing φ; and c.c. denotes thecomplex conjugate. Thus for small values of |αc|, the fluid moves almost as a rigid body, whereasfor large |αc|, the motion becomes confined to a Stokes boundary layer of width |αc|−1 andthe fluid in the center of the sphere remains stationary. Their results show that for myopic eyes,which usually have a larger radius, the shear stress generated by the vitreous humor on the retinais typically larger than for nonmyopic eyes.
Repetto et al. (2005) studied vitreous fluid dynamics experimentally by creating an enlargedmodel of the vitreous chamber in the form of a perspex cylinder containing a spherical cavity.They mounted the cylinder on a motor that could perform prescribed torsional rotations aboutthe vertical axis and observed the resulting motion of the fluid on the horizontal mid-plane of themodel using particle image velocimetry. Under periodic forced rotations of prescribed amplitudeand frequency, the behavior was characterized by two dimensionless parameters: the Womersleynumber, α, and the angular amplitude of the oscillations, ε. Their results were shown to agree well,both qualitatively and quantitatively, with the theoretical predictions of David et al. (1998). Theauthors also considered angular displacements based on measurements of realistic saccades, thatis, a single rotation through a fixed angle starting from an initially stationary fluid. At each pointin space, they measured the maximum over time of the absolute value of the azimuthal componentof the fluid velocity and also the timescale over which it was achieved. By decomposing thetime dependence of the angular displacement into a linear superposition of Fourier modes, theycompared these measurements with the theory of David et al. (1998), finding good agreementeven though the flow is not periodic, whereas David et al.’s model assumes periodicity. They usedtheir results to show that the shear stress is not strongly dependent on the angle through whichthe eye moves in a saccade. Thus, because small-angle saccades are much more frequent thanlarge-angle saccades, small-angle saccades are responsible for generating the majority of the shearstress on the retina when integrated over time.
In addition to the behavior just described, there is also a steady component of flow in thevitreous humor [steady streaming (e.g., see Riley 2001)]. For small-amplitude oscillations, thiscomponent is much smaller in magnitude than the leading-order oscillatory flow, but even soit can play an important role in mass transport because the transport it induces does not tendto cancel over a period of the oscillatory motion. Therefore, Repetto et al. (2008) studied thissteady streaming flow analytically in a similar system, i.e., a torsionally oscillating sphere filledwith a Newtonian viscous and incompressible fluid, assuming rotations of small angular amplitudeε. They formulated the solution as a series expansion in powers of the small parameter ε : u =εu1 + ε2u2 + · · · , p = εp1 + ε2 p2 + · · · . The leading-order solutions u1 and p1 have frequencyω and are also given by Equation 7, but with αc replaced by the Womersley number α. The first
www.annualreviews.org • Fluid Mechanics of the Eye 361
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
corrections u2 and p2 are driven by the nonlinear term u1 · ∇u1 in the Navier-Stokes equationand are thus a superposition of a solution with frequency 2ω and a steady solution, denoted u(2)
2 ,p (2)
2 and u(0)2 , p (0)
2 , respectively [thus u2 = u(0)2 + u(2)
2 and p2 = p (0)2 + p (2)
2 ]. Because the steadysolution is more important in terms of its implication for mass transport, the authors calculatedu(0)
2 and p (0)2 , but neglected u(2)
2 and p (2)2 . The solution took the form of a sum of terms whose
dependence on θ and φ was found exactly, but whose dependence on r was an integral that hadto be computed numerically. In the limit α � 1, the integral could be calculated analytically,in which case the velocity can be shown to be proportional to α6 and thus grows very slowlyas α increases. The integral can also be found analytically in the limit α � 1, and in this casethe velocity tends to a constant value. The authors also performed experiments using the sameapparatus as Stocchino et al. (2007) but taking images only once per period to reveal the average,rather than the instantaneous, velocity. The theoretical and experimental results showed goodagreement for small amplitudes ε (within 10% for ε = 0.0885) over the whole range of α (from 3.1to 15.9).
4.2. Flow in Models that Account for the Real Shape of the Vitreous Chamber
The vitreous chamber is not perfectly spherical, and the most prominent feature is an indentationinto the chamber caused by the presence of the lens. To investigate the effect of the shape,Stocchino et al. (2007) used an experimental model similar to the spherical model of Repettoet al. (2005), but with a modified shape. Based on their analysis of several ultrasound and magneticresonance scans, the authors modeled the lens as a spherical indentation into the sphere, with bothspheres having the same radius. This introduces a further nondimensional parameter, δ, equal tothe maximum depth of the indentation divided by the vitreous cavity radius R. Again they subjectedthis apparatus to periodic, torsional rotations and, approximately at each of the times when theangular velocity reached its maximum absolute value, observed a circulation structure generated atthe back of the indentation. This structure then traveled toward the center of the sphere and wasannihilated. The path taken by the structure depended on the value of the Womersley number,α. For small α, it traveled approximately in a straight line to the center of the vitreous cavity. Forlarge α, the circulation structure initially took the same path as in the low-α case but then divergedfrom the low-α track as it moved away from the lens. This experimental work was extended byStocchino et al. (2010), who used particle image velocimetry with images separated by a multipleof the oscillation period to find a steady streaming flow on the plane of symmetry orthogonal to theaxis of rotation. For moderate α, this revealed two large, counter-rotating steady circulation cells.As α was increased, a complicated sequence of topological changes took place in the flow, and forthe largest value of α considered (α = 45.7), the most obvious circulations were a counter-rotatingpair with the a sense of rotation opposite to those visible for small α.
There has also been analytical progress on this problem. Repetto (2006) assumed the flow tobe incompressible and irrotational. Thus the governing equations reduce to Laplace’s equationfor the velocity potential, subject to no-penetration boundary conditions, and time enters theproblem only as a parameter. In a perfect sphere, the velocity equals zero because there is no stressat the boundary to drive a flow. Motivated by this, the author assumed the indentation to be small,δ � 1, and linearized the problem. He found the potential as a sum of the spherical harmonicfunctions each multiplied by a function of the radial coordinate. The linearized unsteady Bernoulliequation was used to find the pressure.
However, this solution did not reproduce the circulations seen in the experiments, as they arenot irrotational. To model these circulations, Repetto et al. (2010) dropped the assumption ofpotential flow and considered Newtonian viscous flow in an indented sphere. They also assumed
362 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
that δ is small and expanded the velocity as a double series u = ε(δ0u10 + δu11 + · · ·) + ε2(δ0u20 +δu21 + · · ·) + · · ·, and similarly for the pressure. The component u10 and the steady streamingcomponent of u20, denoted u(0)
20 , equal the components u1 and u(0)2 of the solution for the flow
in a true sphere described in Section 4.1. The calculation of u11 and u(0)21 (the steady streaming
component of u21) is performed in terms of vector spherical harmonics, which are a basis ofpairwise orthogonal, vector-valued functions of θ and φ. The components u11 and u(0)
21 are writtenas a sum of an unknown function of r times a vector spherical harmonic times a known functionof t. The analysis also shows that u11 and u(0)
21 , which arise as a result of the deformed geometry,grow rapidly as α increases and become increasingly important in the overall flow structure. Thusthe method is not expected to predict the velocity accurately for large α.
Plotting u10 + δu11 reveals a circulation that forms every half-period behind the indentation,moves to the center of the sphere, and is annihilated. This reproduces the features of the exper-imentally observed circulations for low α but not the path of the circulations for high α, whichis to be expected as the series expansion is not accurate for large α. Examination of the steadycomponent arising because of the deformation, u21, reveals that there are two large steady circu-lations on the horizontal mid-plane (the plane of symmetry perpendicular to the axis of rotation)just inside the indentation. The wall shear stress is maximal on the apex of the indentation andhas two additional smaller maxima on either side of this point.
4.3. Protective Mechanisms in the Eyes of Woodpeckers
Wygnanski-Jaffe et al. (2007) observed that, during pecking, the eyes of woodpeckers undergovery large accelerations and decelerations that, if scaled up correctly to the human eye, would causesignificant damage and loss of sight, yet the woodpecker eyes seem to be unharmed. They thereforeaimed to understand the physiological adaptations protecting the woodpecker eye. This could berelevant to shaken baby syndrome, a condition caused by violent shaking of a small child, whichis usually characterized by retinal hemorrhage, subdural hematoma, and acute encephalopathy.The mechanisms causing retinal hemorrhage are currently unknown, but investigation of theprotective features of the woodpecker eye could give insight into the particular mechanism offailure in human eyes when subjected to large accelerations and decelerations.
The authors identified a number of anatomical specializations in the woodpecker eye that pre-sumably confer protection against large accelerations. This work highlights that dynamic motionof the eye will lead to deformation of the eye globe, which has not been incorporated into previousstudies of vitreous flow, but which will undoubtedly lead to much interesting fluid mechanics.
4.4. Models of Partially Liquefied Vitreous Humor
Repetto et al. (2004) considered a spherical model of the vitreous chamber of radius R containingan elastic membrane dividing the chamber into two equal hemispherical parts. They consideredboth free membrane motions, in which the sphere remains stationary but the membrane andfluid start from a nonequilibrium configuration, and periodically forced motions, in which thesphere performs torsional oscillations about a diameter whose end points are points of attachmentof the membrane. In both cases, they assume the membrane displacement and amplitude of thevelocity are small, allowing them to linearize the system. They also assume that the membranedisplacement from equilibrium, η(r, φ, t), is proportional to sin φ, where (r, θ, φ) is a system ofspherical coordinates that has its axis normal to the equilibrium plane of the membrane (in theforced case, these coordinates rotate in time). The assumption of a separable solution allows them
www.annualreviews.org • Fluid Mechanics of the Eye 363
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
to expand the membrane displacement as
η =∞∑
m=1
J1
(αmrR
)sin φ em(t), (9)
where αm is the m-th positive zero of the Bessel function J1 of the radial coordinate, and em(t) arefunctions to be determined. The velocity potential satisfies Laplace’s equation, and they expandit as
ϕ =∞∑
m=1
ψm(r, θ ) sin φdem(t)
dt+ [χ (r, θ ) sin φ eiωt + c.c.], (10)
where the second term involving the function χ is only needed in the case of forced oscillations.The analysis for free motions yields the natural frequencies of the system, which are the frequenciesassociated with each of the functions em. These are found to be substantially lower than the naturalfrequencies of the membrane in the absence of fluid. With forced oscillations, there is an infiniteresponse at each of the natural frequencies, suggesting that, for a viscous fluid, there will be a largebut finite response at the natural frequencies. Such a response could in turn lead to the generationof large shear stresses on the retina, potentially leading to damage and subsequent detachment.
Repetto et al. (2011) studied a circular model of the vitreous chamber filled with partiallyliquefied vitreous humor. They modeled the liquefied component as a Newtonian incompress-ible fluid and the gel component as a homogeneous isotropic viscous elastic incompressible solid,characterized by a Mooney-Rivlin strain energy function, and assumed that the two componentswere separated by an elastic membrane. They solved a numerical model to find the solid deforma-tion and fluid flow. Their results showed oscillations of the vitreous humor for sufficiently largevalues of the elastic modulus of the solid. The stresses were particularly high near the points ofattachment of the membrane to the retina, which could account for the increased risk of retinaldetachment at these locations.
4.5. Mass Transport in the Vitreous Humor
Direct injection into the vitreous humor is commonly used to deliver large quantities of a drugto the retina (Maurice 2001). The instantaneous distributions at various times after injection andthe timescales associated with uptake of the drug have been investigated by a number of authors.
Xu et al. (2000) investigated the distribution of a drug after injection using a numerical model.They included both diffusion of the drug and convection due to the slow flow that exists because ofa pressure drop between the anterior and posterior of the vitreous chamber and/or by active uptakeby the retina. The flow was assumed to be governed by Darcy’s law. The authors performed in vitroexperiments with a small sample of bovine vitreous humor to determine the diffusion coefficientof a model compound representing the drug. They also determined the hydraulic conductivityby performing compression experiments on a sample of vitreous humor and then numericallysolved the equation for mass conservation and a governing equation for the network phase. Theyused their results to estimate the Peclet numbers in human and mouse eyes, finding these to beapproximately 0.41 and 0.024, respectively. Thus they concluded that the slow anterior-posteriorflow does not typically play the dominant role in transport in the vitreous humor, at least for themodel compound considered.
Once injected, various mechanisms can lead to nondelivery of the drug to the retina. Theseinclude convection due to choroidal blood flow, active transport by the retinal pigment epithe-lium, and convective losses due to collecting vessels outside the sclera. Balachandran & Barocas(2008) developed a model to investigate typical loss rates due to these three mechanisms. They
364 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
considered a model consisting of three regions: the vitreous chamber, the retinal pigment epithe-lium (surrounding the vitreous chamber on its posterior surface), and the choroid (surroundingthe retinal pigment epithelium). They used Darcy’s law and the convection-diffusion equationto model the fluid flow and the drug transport, respectively (with different diffusivities in eachregion). In the vitreous humor, there was assumed to be no source or uptake of the drug, while, tomodel the active transport within the retina, there was an additional transport term kact ·∇c , wherec is the drug concentration and kact is a vector in the radially outward direction. In the choroid,there was no additional transport, but they added a rate-of-uptake term γ (c − c bl), where cbl isthe drug concentration in the blood and γ is constant. The boundary conditions were as follows:At the lens, they applied no penetration of fluid and no mass flux of drug; at the anterior hyaloidmembrane (the anterior surface of the vitreous humor immediately posterior to the lens) and atthe surface of the sclera, they set the pressures (with an approximate drop of 5 mmHg betweenthem driving the flow); and at both the hyaloid membrane and at the sclera, they assumed a rateof uptake proportional to the amount of drug available, but with different constants of propor-tionality in the two regions. The authors solved the system numerically to obtain concentrationprofiles of the drug and compared the loss rates by each of the three mechanisms.
Repetto et al. (2010) also used their calculated flows to estimate a Peclet number quantifying thedegree of mixing that occurs because of convective mass transport in the vitreous humor. Becausethe flow components u10, u11, and u(0)
20 all consist of closed streamlines, these components do notinduce mixing, and thus the estimate of the Peclet number is based on the maximum magnitudeof u(0)
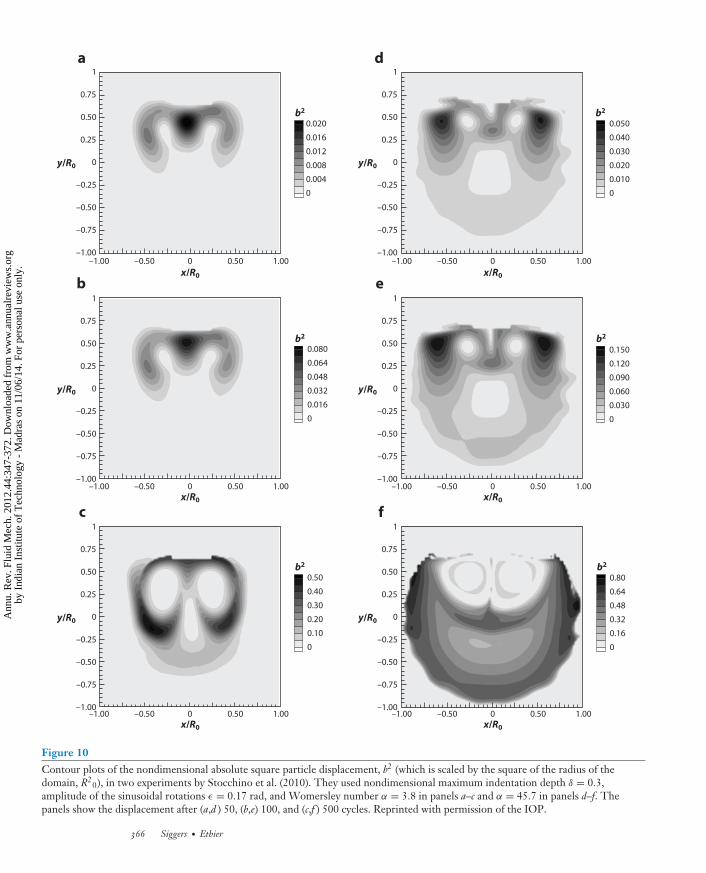
21 . For fluorescein, a commonly used tracer in ophthalmology, this gives an estimated Pecletnumber of approximately 1,000. This would suggest that the strength of the convection inducedby saccades is typically much greater than diffusion, and thus convection should not be neglectedin a model of drug transport. Stocchino et al. (2010) calculated the particle trajectories associatedwith the steady component of the flow and used these to find typical distances traveled by a particleover time. They found that the value of the Womersley number has a significant effect on masstransport, with flows at high Womersley numbers transporting the fluid significantly further aftera fixed number of periods (see Figure 10).
5. TRANSPORT ACROSS BRUCH’S MEMBRANE
Among the elderly of the industrialized world, age-related macular degeneration is the mostcommon cause of vision loss. Bruch’s membrane is the innermost layer of the choroid, and it issituated immediately outside the retinal pigment epithelium, which is the outer layer of the retina(see Figure 4). The macula is an approximately circular region of the retina situated close to theoptic nerve and has the highest density of photoreceptors.
Age-related macular degeneration is thought to be caused by a buildup of lipids within Bruch’smembrane, which reduces mass transport across the membrane in a process that bears somesimilarities to atherosclerosis, the main cause of arterial disease. The reduction in mass transportleads to injury to the photoreceptors because it both reduces the nutrients supplied and decreasesthe removal rate of metabolites, which causes vision loss (Curcio et al. 2009).
The effect of lipid accumulation on fluid flow was studied by McCarty et al. (2008) boththeoretically and experimentally. In the theoretical model, they assumed that the fluid crossing themembrane is Newtonian and incompressible and treated the membrane as a porous medium withspecific hydraulic conductivity Km. Thus the mechanics was governed by Darcy’s equation and thecontinuity equation, which together reduce to Laplace’s equation for the pressure, ∇2 P = 0. Theytreated the lipid as being composed of identical rigid spheres, each of radius ra, and developed twomodels to estimate the effective specific hydraulic conductivity, K, of the porous medium when
www.annualreviews.org • Fluid Mechanics of the Eye 365
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
FL44CH15-Siggers ARI 18 November 2011 14:2
–1.00 –0.50 0 0.50 1.00
–1.00 –0.50 0 0.50 1.00
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
b2
b2
b2
b2
b2
b2
0.020
0.016
0.012
0.008
0.004
0
a
0.080
0.064
0.048
0.032
0.016
0
b
0.50
0.40
0.30
0.20
0.10
0
c
0.050
0.040
0.030
0.020
0.010
0
d
0.150
0.120
0.090
0.060
0.030
0
e
0.80
0.64
0.48
0.32
0.16
0
f
y/R0
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
y/R0
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
y/R0
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
y/R0
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
y/R0
–1.00
–0.75
–0.50
–0.25
0
0.25
0.50
0.75
1
y/R0
x/R0
x/R0
–1.00 –0.50 0 0.50 1.00
x/R0
–1.00 –0.50 0 0.50 1.00
–1.00 –0.50 0 0.50 1.00
x/R0
x/R0
–1.00 –0.50 0 0.50 1.00
x/R0
Figure 10Contour plots of the nondimensional absolute square particle displacement, b2 (which is scaled by the square of the radius of thedomain, R2
0), in two experiments by Stocchino et al. (2010). They used nondimensional maximum indentation depth δ = 0.3,amplitude of the sinusoidal rotations ε = 0.17 rad, and Womersley number α = 3.8 in panels a–c and α = 45.7 in panels d–f. Thepanels show the displacement after (a,d ) 50, (b,e) 100, and (c,f ) 500 cycles. Reprinted with permission of the IOP.
366 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
embedded, approximately uniformly, by lipid spheres with volume fraction ϕ (volume of spheresper unit total volume).
In the first model, they used a unit-cell approach, in which a single rigid sphere was surroundedby a larger concentric spherical volume of the porous medium, such that the volume fraction ofthe rigid sphere equaled ϕ. They assumed that the velocity on the outer surface of the poroussphere was the average velocity in the medium. In this assumption, the outer surface is sufficientlyfar from the rigid sphere that the velocity on it is approximately uniform, and therefore ϕ mustbe small for it to be valid. The resulting model can be solved exactly to find the pressure field,and a comparison of the spatially averaged pressure gradient with Darcy’s law yielded the effectivehydraulic conductivity, K:
K = Km1 − ϕ
1 + ϕ/2. (11)
The second way to estimate K started with the rigid sphere embedded in the porous sphere ofthe first model and used the calculated pressure distribution to find the total force on the rigidsphere, which was
43ϕ
πr3a μV 0
(1K
− 1Km
). (12)
Comparing this with the formula derived by Brinkman for the force on a sphere in a porousmedium, they obtained the relationship
1K
= 1Km
+ 9ϕ
2r2a (1 − ϕ)
(1 + ra√
K+ r2
a
3K
), (13)
which agrees with Equation 11 to first order in ϕ in the limit K � r2a (which was relevant for their
experiments).The authors tested these theoretical results by conducting experiments. They used Matrigel,
a material that has similar properties to those of Bruch’s membrane. After the addition of latexnanospheres to the Matrigel, the measured values of the effective hydraulic conductivities agreedwell with those predicted by the theory. However, with embedded spheres of low-density lipopro-tein instead of latex nanospheres, the effective hydraulic conductivity decreased significantly morethan the theory would predict, a phenomenon that has not been satisfactorily explained.
6. DISCUSSION
Our aim in writing this article was to show that the eye presents a wealth of interesting andchallenging problems in fluid mechanics. Several of these problems have been tackled; however,there remain many outstanding unsolved fluid mechanical problems. We recommend this area tothe reader as a source of interesting and accessible research questions that have potential impactson our most important sense.
SUMMARY POINTS
1. The combined resistance of the trabecular meshwork filled with biopolymer togetherwith the inner lining of Schlemm’s canal are estimated to be sufficient to be the primarysource of resistance to the outflow of aqueous humor in health. The observed increase inIOP during glaucoma could result partially from faulty caliber regulation in Schlemm’scanal but does not result from the collapse of Schlemm’s canal per se.
www.annualreviews.org • Fluid Mechanics of the Eye 367
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
2. Using a mathematical model, it is possible to show that typical raised IOP values inglaucoma can drive a sufficiently large flow along the axons of the retinal ganglion cellsto cause washout of energy-providing ATP in the cells, which could promote cell deathand vision loss.
3. Thermal convection is typically the dominant mechanism driving flow in the anteriorchamber.
4. The iris deforms as a result of the mechanical forces acting on it, leading to a complicatedfluid-structure interaction problem, which is relevant for closed-angle glaucoma andrecovery after a transient deformation of the iris position.
5. The transport of proteins within the anterior eye does not follow the same path as theflow of aqueous humor itself.
6. Fluid transport in the cornea is anisotropic owing to the arrangement of fibers withinthe corneal stroma.
7. If the vitreous humor is treated as viscoelastic, the vitreous chamber is assumed to bespherical, and movements of the eyeball are assumed to be torsional and sinusoidal, thelinearized equations can be solved exactly to find the primary azimuthal component ofthe flow. If the fluid is additionally assumed to be Newtonian, then a secondary streamingcomponent of flow can be found semianalytically.
8. The departure from perfect sphericity in the real shape of the vitreous cavity has asignificant effect on the flow, leading to additional circulation structures in both theprimary flow and in the steady streaming flow. Transient temporal deformations in theshape of the vitreous cavity are likely to have a large effect on the flow and pressure,which is not fully understood.
9. Nonhomogeneous properties of the vitreous humor can lead to additional stresses. Amembrane separating the cavity into two regions could lead to the possibility of resonanceat particular frequencies of oscillation. Alternatively, if the vitreous cavity is occupied bya hemispherical region of elastic solid and a hemispherical region of viscous fluid, withthe two parts separated by a membrane, then the stress is particularly high at the pointsof attachment of the membrane.
10. In the case of liquefied vitreous humor, mass transport typically primarily results fromconvection induced by flow due to eye movements. The steady streaming componentof the flow plays one of the dominant roles in transport. In addition, mass transport issignificantly affected by both the shape of the chamber and the frequency of oscillation.
11. Impaired transport through Bruch’s membrane, thought to be responsible for maculardegeneration, can be partially understood by considering a homogenized mathematicalmodel of lipid particles embedded in a membrane.
FUTURE ISSUES
1. Open-angle glaucoma is known to be caused by increased resistance in the outflow path-way of the aqueous humor. However, the exact locations and causes of the change inresistance are not understood.
368 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
2. Closed-angle glaucoma results from the iris physically blocking the outflow of aqueoushumor. The mechanisms underlying this condition have not been fully resolved.
3. Glaucoma results in the death of retinal ganglion cells and subsequent vision loss. Themechanism of cell death has not been conclusively proven.
4. The rheological properties of the vitreous humor need to be characterized and incorpo-rated into a model of vitreous flow.
5. It is important to explore the effects of transient deformation of the vitreous cavity on thevitreous pressure and flow, which may be important to understand the effect of impactsand retinal hemorrhage in shaken baby syndrome.
6. Possible mechanical causes of retinal detachment and strategies for treatment need to beexplored.
7. Convective drug transport in the vitreous humor needs to be investigated, in particularunderstanding the timescales involved and locations of delivery.
8. With regard to the transport across Bruch’s membrane, the reasons for the increasein resistance to transport when the membrane contains embedded lipids are not fullyresolved.
DISCLOSURE STATEMENT
The authors are not aware of any biases that might be perceived as affecting the objectivity of thisreview.
ACKNOWLEDGMENTS
C.R.E. acknowledges financial support from a Royal Society Wolfson Research Merit Award.
LITERATURE CITED
Allingham RR, de Kater AW, Ethier CR. 1996. Schlemm’s canal and primary open glaucoma: correlationbetween Schlemm’s canal dimensions and outflow facility. Exp. Eye Res. 62:101–9
Amini R, Barocas VH. 2010. Reverse pupillary block slows iris contour recovery from corneoscleral indenta-tion. J. Biomech. Eng. 132:071010
Anderson DR, Hendrickson A. 1974. Effect of intraocular pressure on rapid axoplasmic transport in monkeyoptic nerve. Invest. Ophthalmol. Vis. Sci. 13:771–83
Balachandran RK, Barocas VH. 2008. Computer modeling of drug delivery to the posterior eye: effect ofactive transport and loss to choroidal blood flow. Pharm. Res. 25:2685–96
Balaratnasingam C, Morgan WH, Bass L, Matich G, Cringle SJ, Yu DY. 2007. Axonal transport and cy-toskeletal changes in the laminar regions after elevated intraocular pressure. Invest. Ophthalmol. Vis. Sci.48:3632–44
Demonstrated theplausibility of aproposed mechanismfor cell death inglaucoma.
Band LR, Hall CL, Richardson G, Jensen OE, Siggers JH, Foss AJE. 2009. Intracellular flow inoptic-nerve axons: a mechanism for cell death in glaucoma. Invest. Ophthalmol. Vis. Sci. 50:3750–58
Barsotti MF, Bartels SP, Freddo TF, Kamm RD. 1992. The source of protein in the aqueous humor of thenormal monkey eye. Invest. Ophthalmol. Vis. Sci. 33:581–95
Bear J. 1988. Dynamics of Fluids in Porous Media. New York: Dover
www.annualreviews.org • Fluid Mechanics of the Eye 369
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Bert RJ, Caruthers SD, Jara H, Krejza J, Melhem ER, et al. 2006. Demonstration of an anterior diffusionalpathway for solutes in the normal human eye with high spatial resolution contrast-enhanced dynamicMR imaging. Invest. Ophthalmol. Vis. Sci. 47:5153–62
Beswick JA, McCulloch C. 1956. Effect of hyaluronidase on the viscosity of the aqueous humour. Br. J.Ophthalmol. 40:545–48
Bill A, Svedbergh B. 1972. Scanning electron microscopic studies of the trabecular meshwork and the canal ofSchlemm: an attempt to localize the main resistance to outflow of aqueous humor in man. Acta Ophthalmol.50:295–320
Bishop PN. 2000. Structural macromolecules and supramolecular organisation of the vitreous gel. Prog. Retin.Eye Res. 19:323–44
Brubaker RF, Ritch R, Shields MB, Krupin T. 1989. Measurement of aqueous flow by fluorophoto-metry. In The Glaucomas, Vol. 1, ed. R Ritch, MB Shields, T Krupin, pp. 337–44. St Louis: C.V.Mosby
Burgoyne CF, Downs JC, Bellezza AJ, Suh JKF, Hart RT. 2005. The optic nerve head as a biome-chanical structure: a new paradigm for understanding the role of IOP-related stress and strainin the pathophysiology of glaucomatous optic nerve head damage. Prog. Retin. Eye Res. 24:39–73
Canning CR, Greaney MJ, Dewynne JN, Fitt AD. 2002. Fluid flow in the anterior chamber of a human eye.IMA J. Math. Appl. Med. 19:31–60
Curcio CA, Johnson M, Huang JD, Rudolf M. 2009. Aging, age-related macular degeneration, andthe response-to-retention of apolipoprotein B-containing lipoproteins. Prog. Retin. Eye Res. 28:393–422
Calculated a closed-form solution for theflow of vitreous in amodel of eyemovements.
David T, Smye S, Dabbs T, James T. 1998. A model for the fluid motion of vitreous humour of thehuman eye during saccadic movement. Phys. Med. Biol. 43:1385–99
Eriksson A, Svedbergh B. 1980. Trans-cellular aqueous-humor outflow: theoretical and experimental study.Graefes Arch. Clin. Exp. Ophthalmol. 212:187–97
Improved estimates ofthe resistance of thetrabecular meshwork toaqueous outflow using ahierarchical model.
Ethier CR. 1986. The hydrodynamic resistance of hyaluronic acid: estimates from sedimentationstudies. Biorheology 23:99–113
Ethier CR, Johnson M, Ruberti J. 2004a. Ocular biomechanics and biotransport. Annu. Rev. Biomed. Eng.6:249–73
Ethier CR, Read AT, Chan D. 2004b. Biomechanics of the Schlemm’s canal enthothelial cells: influence onthe F-actin architecture. Biophys. J. 87:2828–37
Fechtner RD, Weinreb RN. 1994. Mechanisms of optic nerve damage in primary open angle glaucoma. Surv.Ophthalmol. 39:23–42
Demonstrated thatthermal convection istypically the dominantdriver of flow in theanterior chamber.
Fitt AD, Gonzalez G. 2006. Fluid mechanics of the human eye: aqueous humour flow in the anteriorchamber. Bull. Math. Biol. 68:53–71
Freddo TF, Bartels SP, Barsotti MF, Kamm RD. 1990. The source of proteins in the aqueous humor of thenormal rabbit. Invest. Ophthalmol. Vis. Sci. 31:125–37
Happel J. 1959. Viscous flow relative to arrays of cylinders. AIChE J. 5:174–77Happel J, Brenner H. 1983. Low Reynolds Number Hydrodynamics. The Hague: Martinus Nijhoff
Developed a newexperimental techniqueand a theoretical modelto investigate flow inthe cornea.
Hedbys BO, Mishima S. 1962. Flow of water in the corneal stroma. Exp. Eye Res. 1:262–75Heys JJ, Barocas VH. 2002a. A Boussinesq model of natural convection in the human eye and formation of
Krunberg’s spindle. Ann. Biomed. Eng. 30:392–401Heys JJ, Barocas VH. 2002b. Computational evaluation of the role of accommodation in pigmentary glaucoma.
Invest. Ophthamol. Vis. Sci. 43:700–8Heys JJ, Barocas VH, Taravella MJ. 2001. Modeling passive mechanical interaction between aqueous humor
and iris. J. Biomech. Eng. 123:540–47Huang EC, Barocas VH. 2004. Active iris mechanics and pupillary block: steady-state analysis and comparison
with anatomical risk factors. Ann. Biomed. Eng. 32:1276–85Huang EC, Barocas VH. 2006. Accommodative microfluctuations and iris contour. J. Vis. 6:653–60Hubbard WC, Johnson M, Gong H, Gabelt BT, Peterson JA, et al. 1997. Intraocular pressure and outflow
facility are unchanged following acute and chronic intracameral chondroitinase ABC and hyaluronidasein monkeys. Exp. Eye Res. 65:177–90
370 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Developed a model toinvestigate thecombined resistance ofthe trabecularmeshwork and innerwall of Schlemm’s canalto the outflow ofaqueous humor.
Johnson M, Shapiro A, Ethier CR, Kamm RD. 1992. Modulation of outflow resistance by the poresof the inner wall endothelium. Invest. Ophthamol. Vis. Sci. 33:1670–75
Developed amathematical model offlow in and throughSchlemm’s canal toinvestigate the source ofincreased aqueousoutflow resistance inglaucoma.
Johnson MC, Kamm RD. 1983. The role of Schlemm’s canal in aqueous outflow from the human eye.Invest. Ophthamol. Vis. Sci. 24:320–25
Kiel JW, van Heuven WAJ. 1995. Ocular perfusion pressure and choroidal blood flow in the rabbit. Invest.Ophthalmol. Vis. Sci. 36:579–85
Krey HF, Brauer H. 1998. Chibret Augenatlas: Eine Repetition fur Artze mit Zeigetafeln fur Patienten. Munich:Chibret Med. Serv.
Lee B, Litt M, Buchsbaum G. 1992. Rheology of the vitreous body. Part I: viscoelasticity of human vitreous.Biorheology 29:521–33
Liu D, Wood NB, Witt N, Hughes AD, Thom SA, Xu XY. 2009. Computational analysis of oxygen transportin the retinal arterial network. Curr. Eye Res. 34:945–56
Maurice D. 2001. Review: practical issues in intravitreal drug delivery. J. Ocul. Pharmacol. Ther. 17:393–401
Developed amathematical model toinvestigate the effect ofembedded lipidparticles on thetransport throughBruch’s membrane.
McCarty WJ, Chimento MF, Curcio C, Johnson M. 2008. Effects of particulates and lipids on thehydraulic conductivity of Matrigel. J. Appl. Physiol. 105:621–28
McEwen WE. 1958. Application of Poiseuille’s law to aqueous outflow. AMA Arch. Ophthal. 60:290–94Minckler DS, Bunt AH, Johanson GW. 1977. Orthograde and retrograde axoplasmic transport during acute
ocular hypertension in the monkey. Invest. Ophthalmol. Vis. Sci. 16:426–41Morgan JE. 2000. Optic nerve head structure in glaucoma: astrocytes as mediators of axonal damage. Eye
14:437–44Moses RA. 1979. Circumferential flow in Schlemm’s canal. Am. J. Ophthalmol. 88:585–91Nickerson CS, Park J, Kornfield JA, Karageozian H. 2008. Rheological properties of the vitreous and the role
of hyaluronic acid. J. Biomech. 41:1840–46Overby DR, Stamer WD, Johnson M. 2009. The changing paradigm of outflow resistance generation: towards
synergistic models of the JCT and inner wall endothelium. Exp. Eye Res. 88:656–70Pillunat LE, Anderson DR, Knighton RW, Joos KM, Feuer WJ. 1997. Autoregulation of human optic nerve
head circulation in response to increased intraocular pressure. Exp. Eye Res. 64:737–44Qu J, Wang D, Grosskreutz CL. 2010. Mechanisms of retinal ganglion cell injury and defense in glaucoma.
Exp. Eye Res. 91:48–53Quigley HA, Broman AT. 2006. The number of people with glaucoma worldwide in 2010 and 2020. Br. J.
Ophthalmol. 90:262–67Rayner K. 1998. Eye movements in reading and information processing: 20 years of research. Psychol. Bull.
124:372–422Reitsamer HA, Kiel JW. 2002. A rabbit model to study orbital venous pressure, intraocular pressure, and
ocular hemodynamics simultaneously. Exp. Eye Res. 43:3728–34Repetto R. 2006. An analytical model of the dynamics of the liquefied vitreous induced by saccadic eye
movements. Meccanica 41:101–17Repetto R, Ghigo I, Seminara G, Ciurlo C. 2004. A simple hydro-elastic model of the dynamics of a vitreous
membrane. J. Fluid Mech. 503:1–14Repetto R, Siggers JH, Stocchino A. 2008. Steady streaming within a periodically rotating sphere. J. Fluid
Mech. 608:71–80Repetto R, Siggers JH, Stocchino A. 2010. Mathematical model of flow in the vitreous humor induced by
saccadic eye rotations: effect of geometry. Biomech. Model. Mechanobiol. 9:65–76Repetto R, Stocchino A, Cafferata C. 2005. Experimental investigation of vitreous humour motion within a
human eye model. Phys. Med. Biol. 50:4729–43Repetto R, Tatone A, Testa A, Colangeli E. 2011. Traction on the retina induced by saccadic eye movements
in the presence of posterior vitreous detachment. Biomech. Model. Mechanobiol. 10(2):191–202Riley N. 2001. Steady streaming. Annu. Rev. Fluid Mech. 33:43–65Riva CE, Hero M, Titze P, Petrig B. 1997. Autoregulation of human optic nerve head blood flow in response
to acute changes in ocular perfusion pressure. Graefes Arch. Clin. Exp. Ophthalmol. 235:618–26Schumer RA, Podos SM. 1994. The nerve of glaucoma! Arch. Ophthalmol. 112:37–44Seiler T, Wollensak J. 1985. The resistance of the trabecular meshwork to aqueous-humor outflow. Graefes
Arch. Clin. Exp. Ophthalmol. 223:88–91
www.annualreviews.org • Fluid Mechanics of the Eye 371
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44CH15-Siggers ARI 18 November 2011 14:2
Spielman L, Goren SL. 1968. Model for predicting pressure drop and filtration efficiency in fibrous media.Environ. Sci. Technol. 2:279–87
Stocchino A, Repetto R, Cafferata C. 2007. Eye rotation induced dynamics of a Newtonian fluid within thevitreous cavity: the effect of the chamber shape. Phys. Med. Biol. 52:2021–34
Stocchino A, Repetto R, Siggers JH. 2010. Mixing processes in the vitreous chamber induced by eye rotations.Phys. Med. Biol. 55:453–67
Swindle KE, Hamilton PD, Ravi N. 2008. In situ formation of hydrogels as vitreous substitutes: viscoelasticcomparison to porcine vitreous. J. Biomed. Mater. Res. A 87:656–65
Weinbaum S. 1998. 1997 Whitaker Distinguished Lecture: models to solve mysteries in biomechanics at thecellular level; a new view of fiber matrix layers. Ann. Biomed. Eng. 26:627–43
Wygnanski-Jaffe T, Murphy CJ, Smith C, Kubai M, Christopherson P, et al. 2007. Protective ocular mecha-nisms in woodpeckers. Eye 21:83–89
Xu J, Heys JJ, Barocas VH, Randolph TW. 2000. Permeability and diffusion in vitreous humor: implicationsfor drug delivery. Pharm. Res. 17:664–69
Yamamoto T, Kitazawa Y. 1998. Vascular pathogenesis of normal-tension glaucoma: a possible pathogeneticfactor, other than intraocular pressure, of glaucomatous optic neuropathy. Prog. Retin. Eye Res. 17:127–43
Yan DB, Coloma FM, Metheetrairut A, Trope GE, Heathcote JG, Ethier CR. 1994. Deformation of thelamina cribrosa by elevated intraocular pressure. Br. J. Ophthalmol. 78:643–48
Zimmerman RL. 1980. In vivo measurements of the viscoelasticity of the human vitreous humor. Biophys. J.29:539–44
372 Siggers · Ethier
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44-FrontMatter ARI 1 December 2011 22:45
Annual Review ofFluid Mechanics
Volume 44, 2012Contents
Aeroacoustics of Musical InstrumentsBenoit Fabre, Joel Gilbert, Avraham Hirschberg, and Xavier Pelorson � � � � � � � � � � � � � � � � � � � � 1
Cascades in Wall-Bounded TurbulenceJavier Jimenez � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �27
Large-Eddy-Simulation Tools for Multiphase FlowsRodney O. Fox � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �47
Hydrodynamic Techniques to Enhance Membrane FiltrationMichel Y. Jaffrin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �77
Wake-Induced Oscillatory Paths of Bodies Freely Risingor Falling in FluidsPatricia Ern, Frederic Risso, David Fabre, and Jacques Magnaudet � � � � � � � � � � � � � � � � � � � � � �97
Flow and Transport in Regions with Aquatic VegetationHeidi M. Nepf � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 123
Electrorheological Fluids: Mechanisms, Dynamics,and Microfluidics ApplicationsPing Sheng and Weijia Wen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 143
The Gyrokinetic Description of Microturbulence in Magnetized PlasmasJohn A. Krommes � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 175
The Significance of Simple Invariant Solutions in Turbulent FlowsGenta Kawahara, Markus Uhlmann, and Lennaert van Veen � � � � � � � � � � � � � � � � � � � � � � � � � � 203
Modern Challenges Facing Turbomachinery AeroacousticsNigel Peake and Anthony B. Parry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 227
Liquid Rope CoilingNeil M. Ribe, Mehdi Habibi, and Daniel Bonn � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 249
Dynamics of the Tear FilmRichard J. Braun � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 267
Physics and Computation of Aero-OpticsMeng Wang, Ali Mani, and Stanislav Gordeyev � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 299
v
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

FL44-FrontMatter ARI 1 December 2011 22:45
Smoothed Particle Hydrodynamics and Its Diverse ApplicationsJ.J. Monaghan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 323
Fluid Mechanics of the EyeJennifer H. Siggers and C. Ross Ethier � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 347
Fluid Mechanics of Planktonic MicroorganismsJeffrey S. Guasto, Roberto Rusconi, and Roman Stocker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 373
Nanoscale Electrokinetics and Microvortices: How MicrohydrodynamicsAffects Nanofluidic Ion FluxHsueh-Chia Chang, Gilad Yossifon, and Evgeny A. Demekhin � � � � � � � � � � � � � � � � � � � � � � � � � 401
Two-Dimensional TurbulenceGuido Boffetta and Robert E. Ecke � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 427
“Vegetable Dynamicks”: The Role of Water in Plant MovementsJacques Dumais and Yoel Forterre � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 453
The Wind in the Willows: Flows in Forest Canopies in Complex TerrainStephen E. Belcher, Ian N. Harman, and John J. Finnigan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 479
Multidisciplinary Optimization with Applicationsto Sonic-Boom MinimizationJuan J. Alonso and Michael R. Colonno � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 505
Direct Numerical Simulation on the Receptivity, Instability,and Transition of Hypersonic Boundary LayersXiaolin Zhong and Xiaowen Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 527
Air-Entrainment Mechanisms in Plunging Jets and Breaking WavesKenneth T. Kiger and James H. Duncan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 563
Indexes
Cumulative Index of Contributing Authors, Volumes 1–44 � � � � � � � � � � � � � � � � � � � � � � � � � � � � 597
Cumulative Index of Chapter Titles, Volumes 1–44 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 606
Errata
An online log of corrections to Annual Review of Fluid Mechanics articles may be foundat http://fluid.annualreviews.org/errata.shtml
vi Contents
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.

AnnuAl Reviewsit’s about time. Your time. it’s time well spent.
AnnuAl Reviews | Connect with Our expertsTel: 800.523.8635 (us/can) | Tel: 650.493.4400 | Fax: 650.424.0910 | Email: [email protected]
New From Annual Reviews:
Annual Review of Statistics and Its ApplicationVolume 1 • Online January 2014 • http://statistics.annualreviews.org
Editor: Stephen E. Fienberg, Carnegie Mellon UniversityAssociate Editors: Nancy Reid, University of Toronto
Stephen M. Stigler, University of ChicagoThe Annual Review of Statistics and Its Application aims to inform statisticians and quantitative methodologists, as well as all scientists and users of statistics about major methodological advances and the computational tools that allow for their implementation. It will include developments in the field of statistics, including theoretical statistical underpinnings of new methodology, as well as developments in specific application domains such as biostatistics and bioinformatics, economics, machine learning, psychology, sociology, and aspects of the physical sciences.
Complimentary online access to the first volume will be available until January 2015. table of contents:•What Is Statistics? Stephen E. Fienberg•A Systematic Statistical Approach to Evaluating Evidence
from Observational Studies, David Madigan, Paul E. Stang, Jesse A. Berlin, Martijn Schuemie, J. Marc Overhage, Marc A. Suchard, Bill Dumouchel, Abraham G. Hartzema, Patrick B. Ryan
•The Role of Statistics in the Discovery of a Higgs Boson, David A. van Dyk
•Brain Imaging Analysis, F. DuBois Bowman•Statistics and Climate, Peter Guttorp•Climate Simulators and Climate Projections,
Jonathan Rougier, Michael Goldstein•Probabilistic Forecasting, Tilmann Gneiting,
Matthias Katzfuss•Bayesian Computational Tools, Christian P. Robert•Bayesian Computation Via Markov Chain Monte Carlo,
Radu V. Craiu, Jeffrey S. Rosenthal•Build, Compute, Critique, Repeat: Data Analysis with Latent
Variable Models, David M. Blei•Structured Regularizers for High-Dimensional Problems:
Statistical and Computational Issues, Martin J. Wainwright
•High-Dimensional Statistics with a View Toward Applications in Biology, Peter Bühlmann, Markus Kalisch, Lukas Meier
•Next-Generation Statistical Genetics: Modeling, Penalization, and Optimization in High-Dimensional Data, Kenneth Lange, Jeanette C. Papp, Janet S. Sinsheimer, Eric M. Sobel
•Breaking Bad: Two Decades of Life-Course Data Analysis in Criminology, Developmental Psychology, and Beyond, Elena A. Erosheva, Ross L. Matsueda, Donatello Telesca
•Event History Analysis, Niels Keiding•StatisticalEvaluationofForensicDNAProfileEvidence,
Christopher D. Steele, David J. Balding•Using League Table Rankings in Public Policy Formation:
Statistical Issues, Harvey Goldstein•Statistical Ecology, Ruth King•Estimating the Number of Species in Microbial Diversity
Studies, John Bunge, Amy Willis, Fiona Walsh•Dynamic Treatment Regimes, Bibhas Chakraborty,
Susan A. Murphy•Statistics and Related Topics in Single-Molecule Biophysics,
Hong Qian, S.C. Kou•Statistics and Quantitative Risk Management for Banking
and Insurance, Paul Embrechts, Marius Hofert
Access this and all other Annual Reviews journals via your institution at www.annualreviews.org.
Ann
u. R
ev. F
luid
Mec
h. 2
012.
44:3
47-3
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by I
ndia
n In
stitu
te o
f T
echn
olog
y -
Mad
ras
on 1
1/06
/14.
For
per
sona
l use
onl
y.
Copyright © 2022 FDOKUMEN