
First description of the adult stage of Clinostomum cutaneum Paperna, 1964 (Digenea: Clinostomidae)...
8
MORPHOLOGICAL AND MOLECULAR DIFFERENTIATION OF CLINOSTOMUM COMPLANATUM AND CLINOSTOMUM MARGINATUM (DIGENEA: CLINOSTOMIDAE) METACERCARIAE AND ADULTS Monica Caffara, Sean A. Locke*, Andrea Gustinelli, David J. Marcogliese*, and Maria L. Fioravanti Department of Veterinary Medical Sciences, Alma Mater Studiorum Universita ` di Bologna, Via Tolara di Sopra 50, 40064 Ozzano Emilia (BO), Italy. e-mail: [email protected] ABSTRACT: The separation of Clinostomum complanatum Rudolphi, 1814 and Clinostomum marginatum Rudolphi, 1819 has long been unclear. Recent data confirm the validity of the junior species, C. marginatum, by ,1% differences in its 18S rDNA sequences. We collected adults and metacercariae of C. complanatum and C. marginatum and found reliable morphological differences in the genital complex at both developmental stages. In addition, we identified basic morphometrics (distance between suckers, body width) in metacercariae that may be useful for discriminating the species. The morphological differences were supported by the comparison of sequences of internal transcribed spacers of ribosomal DNA and of the mitochondrial gene cytochrome c oxidase I (COI) from 39 specimens. In 36 specimens, the average divergence between the species was 7.3% in ITS and 19.4% in COI sequences. Two specimens from North America and 1 from Europe had sequences that did not allow them to be clearly allied with either species. Species of Clinostomum Leidy, 1856 (Digenea: Clinostomidae) have been subjected to several taxonomic revisions because of the high degree of morphological variability within species and similarities between different species (Ukoli, 1966; Yamaguti, 1971; Feizullaev and Mirzoeva, 1983). In particular, the validity and priority of 2 widely reported species of Clinostomum, i.e., Clinosto- mum complanatum Rudolphi, 1814 and Clinostomum marginatum Rudolphi, 1819, have been disputed for nearly 200 yr. In redescribing the specimens studied by Rudolphi, Braun (1901) accepted the validity of C. marginatum (the ‘‘American type’’) and C. complanatum (the ‘‘European type’’), but concluded that the species were primarily distinguished by their geographical distribution. Subsequently, other authors included morphological evidence in the separation of C. marginatum from C. complanatum (e.g., Osborne, 1911, 1912; Cort, 1913; Smallwood, 1914; Hopkins, 1933; Maccagno, 1934; Amin, 1969; Travassos et al., 1969; Lunaschi and Drago, 2009). However, other authors found no consistent morphological differences between the 2 species and considered C. marginatum a junior synonym of C. complanatum (e.g., Baer, 1933; Yamaguti, 1933; Manter, 1938; Price, 1938; Dollfus, 1950; Ukoli, 1966; Yamaguti, 1971; Dowsett and Lubinsky, 1980; Feizullaev and Mirzoeva, 1986; Larson et al., 1988; McAllister, 1990). Despite this lack of consensus, in recent decades, C. marginatum has been widely employed for Nearctic specimens, whereas C. complanatum was used for specimens originating from the Palearctic. New evidence for the validity of C. marginatum was recently presented by Dzikowski et al. (2004), who published sequences of 18S rRNA from isolates of C. complanatum from Israel and C. marginatum from the United States that differ at 22 of 1,892 positions (based on GenBank accessions AY245760 and AY245701). However, the evolutionary conservation of 18S (Hillis and Dixon, 1991) could reduce the utility of these markers for discriminating congeneric species, especially without morpho- logical support. For example, Clinostomum cutaneum Paperna, 1964 and Clinostomum phalacrocoracis Dubois, 1930 show marked morphological differences, but differ at only 3 positions over 1,913 base pairs (bp) of 18S rRNA (Gustinelli et al., 2010). Indeed, the results of Dzikowski et al. (2004) were questioned by McAllister et al. (2010), and both names, C. complanatum and C. marginatum, continue to be used for specimens collected in North America (McAllister et al., 2007, 2010; Forest and Cone, 2011). Additional molecular data are, therefore, desirable to resolve the status of C. marginatum and should be linked to morphological information (Perkins et al., 2011). The internal transcribed spacer (ITS1 and ITS2) regions are more suitable for taxonomic studies at the species level and have been used to discriminate species in at least 19 digenean families (Nolan and Cribb, 2005). Notably, C. cutaneum and C. phalacrocoracis are better resolved by ITS rRNA (16 differences over 1,030 bp of ITS rRNA) than by 18S rRNA (see above). Some authors have also used a ,300-bp fragment of the mitochondrial gene cytochrome c oxidase I (COI) beginning at about 800 bp from the 59 end, using primers developed by Bowles et al. (1995). Recently, Locke, McLaughlin, Dayanandan, et al. (2010) and Locke, McLaughlin, and Marcogliese (2010) discriminated species of digeneans with the use of the initial 650 bp of COI, a region that is currently the focus of the DNA barcoding initiative (www. barcodinglife.org), a large-scale effort to standardize molecular identification of species. Here, we use sequences of ITS and the barcode region of COI, as well as morphology, to test whether C. complanatum and C. marginatum are valid species, and attempt to identify practical molecular and morphological characters for distinguishing between them. MATERIALS AND METHODS Morphological study of specimens Specimens putatively identified as C. marginatum on the basis of molecular data and geographical distribution were taken from both definitive and second intermediate hosts collected across North America. Adults were collected from 3 great blue herons (Ardea herodias L.) from 2 localities in Quebec, Canada (46.124N, 73.028W; 45.422N, 73.583W) and from cattle egrets (Bubulcus ibis L.) from Florida and Texas. Metacercar- iae were obtained from largemouth bass (Micropterus salmoides Lace ´- pe `de), smallmouth bass (M. dolomieu Lace ´pe `de), pumpkinseed (Lepomis gibbosus L.), yellow perch (Perca flavescens Mitchill), golden shiner (Notemigonus crysoleucas Mitchill), johnny darter (Etheostoma nigrum Rafinesque), and round goby (Neogobius melanostomus Pallas) from the St. Lawrence River and its tributaries in Quebec and Ontario (45.128N, 74.404W; 45.316N, 73.879W; 45.433N, 73.734W) and from a boreal toad (Anaxyrus boreas Baird & Girard) in Pleasanton Ridge Regional Park, Received 11 February 2011; revised 20 June 2011; accepted 24 June 2011. * Fluvial Ecosystem Research Section, Aquatic Ecosystem Protection Research Division, Water Science and Technology Directorate, Science and Technology Branch, Environment Canada, St. Lawrence Centre, 105 McGill Street, 7th floor, Montre ´al, QC H2Y 2E7, Canada. DOI: 10.1645/GE-2781.1 J. Parasitol., 97(5), 2011, pp. 884–891 F American Society of Parasitologists 2011 884
-
Upload
mehmetakif -
Category
Documents
-
view
0 -
download
0
Transcript of First description of the adult stage of Clinostomum cutaneum Paperna, 1964 (Digenea: Clinostomidae)...

MORPHOLOGICAL AND MOLECULAR DIFFERENTIATION OF CLINOSTOMUM
COMPLANATUM AND CLINOSTOMUM MARGINATUM (DIGENEA: CLINOSTOMIDAE)
METACERCARIAE AND ADULTS
Monica Caffara, Sean A. Locke*, Andrea Gustinelli, David J. Marcogliese*, and Maria L. FioravantiDepartment of Veterinary Medical Sciences, Alma Mater Studiorum Universita di Bologna, Via Tolara di Sopra 50, 40064 Ozzano Emilia (BO), Italy.e-mail: [email protected]
ABSTRACT: The separation of Clinostomum complanatum Rudolphi, 1814 and Clinostomum marginatum Rudolphi, 1819 has long beenunclear. Recent data confirm the validity of the junior species, C. marginatum, by ,1% differences in its 18S rDNA sequences. Wecollected adults and metacercariae of C. complanatum and C. marginatum and found reliable morphological differences in the genitalcomplex at both developmental stages. In addition, we identified basic morphometrics (distance between suckers, body width) inmetacercariae that may be useful for discriminating the species. The morphological differences were supported by the comparison ofsequences of internal transcribed spacers of ribosomal DNA and of the mitochondrial gene cytochrome c oxidase I (COI) from 39specimens. In 36 specimens, the average divergence between the species was 7.3% in ITS and 19.4% in COI sequences. Two specimensfrom North America and 1 from Europe had sequences that did not allow them to be clearly allied with either species.
Species of Clinostomum Leidy, 1856 (Digenea: Clinostomidae)
have been subjected to several taxonomic revisions because of the
high degree of morphological variability within species and
similarities between different species (Ukoli, 1966; Yamaguti, 1971;
Feizullaev and Mirzoeva, 1983). In particular, the validity and
priority of 2 widely reported species of Clinostomum, i.e., Clinosto-
mum complanatum Rudolphi, 1814 and Clinostomum marginatum
Rudolphi, 1819, have been disputed for nearly 200 yr. In redescribing
the specimens studied by Rudolphi, Braun (1901) accepted the
validity of C. marginatum (the ‘‘American type’’) and C. complanatum
(the ‘‘European type’’), but concluded that the species were primarily
distinguished by their geographical distribution. Subsequently, other
authors included morphological evidence in the separation of C.
marginatum from C. complanatum (e.g., Osborne, 1911, 1912; Cort,
1913; Smallwood, 1914; Hopkins, 1933; Maccagno, 1934; Amin,
1969; Travassos et al., 1969; Lunaschi and Drago, 2009). However,
other authors found no consistent morphological differences between
the 2 species and considered C. marginatum a junior synonym of C.
complanatum (e.g., Baer, 1933; Yamaguti, 1933; Manter, 1938; Price,
1938; Dollfus, 1950; Ukoli, 1966; Yamaguti, 1971; Dowsett and
Lubinsky, 1980; Feizullaev and Mirzoeva, 1986; Larson et al., 1988;
McAllister, 1990). Despite this lack of consensus, in recent decades,
C. marginatum has been widely employed for Nearctic specimens,
whereas C. complanatum was used for specimens originating from
the Palearctic.
New evidence for the validity of C. marginatum was recently
presented by Dzikowski et al. (2004), who published sequences of
18S rRNA from isolates of C. complanatum from Israel and C.
marginatum from the United States that differ at 22 of 1,892
positions (based on GenBank accessions AY245760 and
AY245701). However, the evolutionary conservation of 18S
(Hillis and Dixon, 1991) could reduce the utility of these markers
for discriminating congeneric species, especially without morpho-
logical support. For example, Clinostomum cutaneum Paperna,
1964 and Clinostomum phalacrocoracis Dubois, 1930 show
marked morphological differences, but differ at only 3 positions
over 1,913 base pairs (bp) of 18S rRNA (Gustinelli et al., 2010).
Indeed, the results of Dzikowski et al. (2004) were questioned by
McAllister et al. (2010), and both names, C. complanatum and C.
marginatum, continue to be used for specimens collected in North
America (McAllister et al., 2007, 2010; Forest and Cone, 2011).
Additional molecular data are, therefore, desirable to resolve the
status of C. marginatum and should be linked to morphological
information (Perkins et al., 2011). The internal transcribed spacer
(ITS1 and ITS2) regions are more suitable for taxonomic studies
at the species level and have been used to discriminate species in at
least 19 digenean families (Nolan and Cribb, 2005). Notably, C.
cutaneum and C. phalacrocoracis are better resolved by ITS rRNA
(16 differences over 1,030 bp of ITS rRNA) than by 18S rRNA
(see above). Some authors have also used a ,300-bp fragment of
the mitochondrial gene cytochrome c oxidase I (COI) beginning at
about 800 bp from the 59 end, using primers developed by Bowles
et al. (1995).
Recently, Locke, McLaughlin, Dayanandan, et al. (2010) and
Locke, McLaughlin, and Marcogliese (2010) discriminated species
of digeneans with the use of the initial 650 bp of COI, a region
that is currently the focus of the DNA barcoding initiative (www.
barcodinglife.org), a large-scale effort to standardize molecular
identification of species. Here, we use sequences of ITS and the
barcode region of COI, as well as morphology, to test whether C.
complanatum and C. marginatum are valid species, and attempt to
identify practical molecular and morphological characters for
distinguishing between them.
MATERIALS AND METHODS
Morphological study of specimens
Specimens putatively identified as C. marginatum on the basis ofmolecular data and geographical distribution were taken from bothdefinitive and second intermediate hosts collected across North America.Adults were collected from 3 great blue herons (Ardea herodias L.) from 2localities in Quebec, Canada (46.124N, 73.028W; 45.422N, 73.583W) andfrom cattle egrets (Bubulcus ibis L.) from Florida and Texas. Metacercar-iae were obtained from largemouth bass (Micropterus salmoides Lace-pede), smallmouth bass (M. dolomieu Lacepede), pumpkinseed (Lepomisgibbosus L.), yellow perch (Perca flavescens Mitchill), golden shiner(Notemigonus crysoleucas Mitchill), johnny darter (Etheostoma nigrumRafinesque), and round goby (Neogobius melanostomus Pallas) from theSt. Lawrence River and its tributaries in Quebec and Ontario (45.128N,74.404W; 45.316N, 73.879W; 45.433N, 73.734W) and from a boreal toad(Anaxyrus boreas Baird & Girard) in Pleasanton Ridge Regional Park,
Received 11 February 2011; revised 20 June 2011; accepted 24 June 2011.* Fluvial Ecosystem Research Section, Aquatic Ecosystem Protection
Research Division, Water Science and Technology Directorate, Scienceand Technology Branch, Environment Canada, St. Lawrence Centre,105 McGill Street, 7th floor, Montreal, QC H2Y 2E7, Canada.DOI: 10.1645/GE-2781.1
J. Parasitol., 97(5), 2011, pp. 884–891
F American Society of Parasitologists 2011
884

California (37.620N, 121.891W). Voucher specimens have been depositedin the U.S. National Parasite Collection, Beltsville, Maryland (USNPCNos. 104561.00–104568.00), including 9 subsectioned specimens fromwhich DNA sequences were obtained (see below). Specimens putativelyidentified as C. complanatum were obtained from both definitive andsecond intermediate hosts in Italy. Adults were archival stained specimenscollected from the esophagus of little egret (Egretta garzetta L.) andherons (Ardea cinerea L., Ardea purpurea L.) collected in Italy. Meta-cercariae were taken from barbel (Barbus barbus L., B. meridionalis Risso)and chub (Squalius cephalus L.) from the Sillaro (44.393N, 11.591E) andSanterno Rivers (44.296N, 11.943E) in Emilia-Romagna and chub fromthe Soligo River (45.900N, 12.174E) in Veneto, Italy. Voucher specimenshave been deposited in the U.S. National Parasite Collection, Beltsville,Maryland (USNPC Nos. 104551.00–104560.00). All specimens were fixedin 70% ethanol. Specimens used for morphological work were cleared inAmman’s lactophenol, or stained with Mayer’s acid carmine or usingMalzacher’s method (Pritchard and Kruse, 1982), prior to being mountedon slides in Canada balsam. In some cases, tissue subsamples wereremoved for molecular analysis, and the remainder of the specimen servedas a morphological voucher for sequence data. Some specimens in poorcondition were not studied morphologically and were used solely formolecular work. Line drawings were made with the aid of a drawing tube.Measurements are given in micrometers unless otherwise stated and weretaken following Matthews and Cribb (1998).
We attempted to identify morphological characteristics that reliablydistinguish C. marginatum from C. complanatum as metacercariae becausethese stages are more frequently encountered and existing diagnosticcharacters are based on adults (Maccagno, 1934). Principal-componentsanalysis (PCA) and discriminant function analysis (DFA) were conductedon measurements that were log-transformed so that data would be ona similar scale, with the use of PRIMER v.6 and PERMANOVA+(PRIMER-E, Plymouth, United Kingdom) and SPSS 12.0 (SPSS Inc.,Chicago, Illinois), respectively. Morphometric ratios were not included inanalyses because they are not statistically independent from measurements.
Molecular analysis
Total DNA was extracted from specimens of C. complanatum andsome specimens of C. marginatum using a QIAamp DNA Mini Kit(Qiagen, Valencia, California) following the manufacturer’s protocol,and the internal transcribed spacers (ITS) rRNA gene was amplified asreported by Gustinelli et al. (2010). Extraction and PCR of COI and ITSfrom most specimens of C. marginatum took place at Canadian Centrefor DNA Barcoding (Guelph, Canada) (see Moszczynska et al., 2009).For all specimens, the amplification of the COI fragment was conductedwith the use of the protocol and degenerate primers of Moszczynska et al.(2009). The PCR products were sequenced at 1, or more, of 3 laboratoriesthat all employ the same technology (ABI 3730 DNA Analyzer): CCDB,PRIMM (Milan, Italy), and StarSEQ GmbH (Mainz, Germany). TheCOI PCR product of specimen 26 (see Results) was sequenced once atPRIMM and a second time at StarSEQ. The COI PCR product ofspecimen 26 was also cloned into a pCR2.1 TOPO cloning vector withthe use of the TOPO TA Cloning Kit (Invitrogen, Carlsbad, California).Plasmid DNA was isolated from each clone using Quantum Prep-Plasmid Miniprep Kit (Biorad, Hercules, California) following themanufacturer’s instructions. Insertions of expected size were confirmedby restriction digestion with EcoRI enzyme. Ten clones were sequencedin 1 direction with an M13 universal primer at PRIMM. DNA trace fileassembly was carried out with the use of Vector NTI AdvanceTM 11software (Invitrogen) and sequences were aligned and analyzed inMEGA 5.0 (Tamura et al., 2011). Sequences have been deposited onGenBank (JF718584-JF718619 for COI; JF718620-JF718643 andJN108032 for ITS). Specimen records (including sequencing institution,primers, chromatograms, sequences, collection data, host data, specimenimages, and voucher accession information) are available in projectCLINO at www.barcodinglife.org.
REDESCRIPTION
Clinostomum marginatum
Adult (Fig. 1) (based on 10 specimens) (min–max [mean ± SD] mm):Body oval, elongated, with truncated forebody, wider in gonadal region,
5.9–8.2 (7.0 ± 0.791) mm long, 1.3–2.8 (2 ± 0.387) mm wide. Oral suckersmall, 171–394 (311 ± 58) long 3 252–501 (399 ± 67) wide, surrounded bya well-developed oral collar. Pharynx not observed; esophagus very short,bifurcates immediately posterior to level of oral sucker, esophageal bulbpresent. Ventral sucker larger than oral, 601–918 (764 ± 106) long 3 583–966 (756 ± 120) wide. Intestinal ceca run to posterior end of body, withslightly indented margins both in pre- and postacetabular regions.Excretory ducts in complex web draining into excretory system, notclearly visible. Testes between middle and posterior third of body; anteriortestis, 290–743 (519 ± 149) long 3 569–1,141 (802 ± 190) wide, triangular,slightly lobed, compressed laterally by cirrus sac and dorsoventrally byuterus, when filled with eggs; posterior testis, 166–587 (461 ± 115) long 3379–1,414 (790 ± 259) wide, triangular and median, stretching acrossintracecal space. In some specimens efferent ducts were visible, originatingfrom middle part of posterior margin of anterior testis and from middlepart of anterior margin of posterior testis, both ducts reaching posteriorend of cirrus sac. Cirrus sac, 151–795 (327 ± 183) long 3 166–741 (497 ±
212) wide, anterior to ovary, surrounding laterodorsally right margin ofanterior testis and displacing its anterior lobe to left. Genital pore anteriorto anterior testis, not clearly visible in most specimens. Ovary small, 118–306 (175 ± 61) long 3 101–267 (186 ± 53) wide, ovoid or rounded inshape, in some specimens slightly lobed, located in intertesticular space onright side of body. Vitellaria extensive, extending from posterior margin ofventral sucker to almost reaching end of ceca. Uteroduct runs around leftmargin of anterior testis and opens into uterine sac, which occupies almostall space between ventral sucker and anterior testis when filled with eggs.Metraterm muscular, connecting uterus to genital atrium. Mature eggs,101–109 (105 ± 3) long 3 63–79 (69 ± 5.8) wide, in uterine sac, some inuteroduct and in ootype complex space.
Metacercaria (Fig. 2) (based on 11 specimens, measurements inTable I): Similar to adult, but slightly smaller. Oral collar well developed.Ventral sucker larger than oral. Forebody with glandular structure,anterior to ventral sucker. Intestinal ceca lateral to ventral sucker andgenital primordia, characterized by visible intestinal pouches especially inpostacetabular part. Testes less conspicuous and with greater digitationthan in adults. Cirrus sac well developed, crescent shaped, compressedagainst left anterior margin of anterior testis. Genital pore opening lateralto right anterior margin of anterior testis. Ovary irregular in shape,smaller than cirrus sac, located dextrally in intertesticular space. Vitellarianot evident. Tubular proximal uterus follows similar pathway to that inadult; muscular metraterm visible. Excretory ducts connecting at posteriorextremity just posterior to cecal ends. Cuticular surface covered by thinspines.
Clinostomum complanatum
Adult (Fig. 3)(based on 5 specimens) (min–max [mean ± SD] mm):Body stout, oval, elongated, with slightly truncated anterior extremityand flattened posterior end, wider in gonadal region, 3.4–6.3 (4.9 ± 0.9)mm long, 1.5–2.7 (1.9 ± 0.3) mm wide. Oral sucker small 190–570 (422 ±
121) long 3 320–850 (557 ± 161) wide, with presence of oral collar.Pharynx not observed. Esophageal bulb well developed. Ventral suckerlarger than oral, 600–900 (760 ± 78) long 3 620–900 (737 ± 72) wide.Intestinal ceca run laterally to posterior end of body with indentedmargins more evident in postacetabular regions. Testes grossly triangu-lar, facing each other with concave face, slightly lobed. Anterior testis,550–750 (694 ± 67) long, 360–600 (456 ± 82) wide, in posterior end ofmiddle third of body, strongly offset to left of median line. Posteriortestis, 600–940 (791 ± 99) long, 300–510 (410 ± 81) wide, in posteriorthird of body, with axis on median line. Cirrus sac, 350–400 (382 ± 19)long, 100–200 (162 ± 45) wide, extending from intertesticular space toposterior right margin of anterior testis, opening directly into genitalatrium. Ovary oval, 220–310 (256 ± 32) long, 140–300 (213 ± 62) wide,located dextrally in intertesticular space. Uterine sac tubular withmetraterm extending along right margin of anterior testis, which is offsetto left. Vitellaria extending from posterior margin of ventral suckeralmost reaching posterior end of ceca. Eggs yellowish, elliptical 100–125(114 ± 5.8) long, 65–90 (74 ± 4.7) wide.
Metacercaria (Fig. 4) (based on 10 specimens measurements in Table I):Similar to adult, slightly longer and less stout, with inward flexure at levelof acetabulum. Oral collar visible. Ventral sucker larger than oral.Glandular structure present in forebody, anterior to ventral sucker.Intestinal ceca lateral to ventral sucker and genital complex, withprominent lateral pouches. Testes smaller than in adult, with more
CAFFARA ET AL.—DISTINGUISHING CLINOSTOMUM SPP. 885
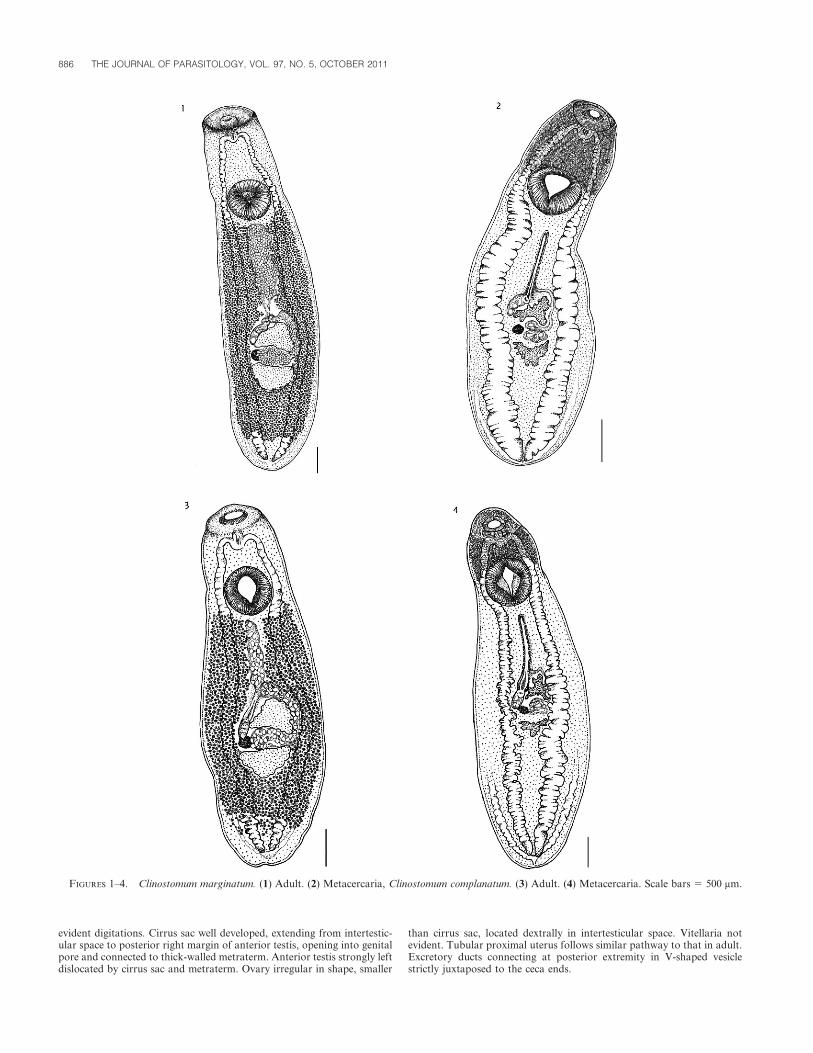
evident digitations. Cirrus sac well developed, extending from intertestic-ular space to posterior right margin of anterior testis, opening into genitalpore and connected to thick-walled metraterm. Anterior testis strongly leftdislocated by cirrus sac and metraterm. Ovary irregular in shape, smaller
than cirrus sac, located dextrally in intertesticular space. Vitellaria notevident. Tubular proximal uterus follows similar pathway to that in adult.Excretory ducts connecting at posterior extremity in V-shaped vesiclestrictly juxtaposed to the ceca ends.
FIGURES 1–4. Clinostomum marginatum. (1) Adult. (2) Metacercaria, Clinostomum complanatum. (3) Adult. (4) Metacercaria. Scale bars 5 500 mm.
886 THE JOURNAL OF PARASITOLOGY, VOL. 97, NO. 5, OCTOBER 2011

Remarks
Morphological differentiation of C. complanatum and C. marginatum: Incleared or stained specimens of C. marginatum, only the anterior lobe ofthe anterior testis is compressed to the left by the cirrus sac, whereas in C.complanatum, the entire anterior testis is displaced to the left by both thecirrus sac and the uterine sac. The genital pore in C. marginatum opensanteriorly to the anterior testis, close to the midline, whereas in C.complanatum it opens at the right external margin of the anterior testis.The genital pore was in the same position both in adults and metacercariaeof C. marginatum and C. complanatum. Among 21 metacercariae analyzed,11 of 17 morphometrics differed significantly in C. marginatum and C.complanatum (Table I). However, because many of these were highlycorrelated (data not shown), a stepwise discriminant function analysis wasperformed to identify which were the best predictors of membership inC. marginatum and C. complanatum. A single discriminant function wascalculated in 2 steps, with morphometrics allowed to enter the model if theprobability of F resulting from the reduction in Wilks’s lambda, waslambda P , 0.05. Body width entered the model in the first step (F1,19 5
43.072, Wilks’s lambda 5 0.306, P , 0.0005) and distance betweensuckers entered in the second step (F2,18 5 87.763, Wilks’s lambda 50.093, P , 0.0005), yielding a function explaining 100% of the variabilitybetween the species (x2 5 42.751, P , 0.0005). The measurement closest tosubsequent entry was ovary width (P 5 0.065). In cross-validatinganalyses, each specimen was then classified using a function of body widthand distance between suckers calculated from the remaining 20 specimens,and all 21 specimens were assigned to the correct species. Essentially, thissuggests that C. marginatum metacercariae are narrower and have morewidely spaced oral and ventral suckers than C. complanatum, and thatthese characters can reliably distinguish between the 2 species. Otherdifferences, although significant, are correlated among themselves andwith these 2 characters, such that they do not yield additional information.
A 2-dimensional principal components analysis provided a reasonablygood representation of the multivariate relationships of the 21 metacer-cariae of C. marginatum and C. complanatum in the 17-dimensional spaceoriginating from morphometric data (Fig. 5). The first axis explained43.3% of the total variance, and the second explained 14.2%. Notably,both species were perfectly separated by the first axis, and the majority of
C. marginatum were more negative on axis 2 than C. complanatum. On thefirst axis, body width was the third strongest loading component (afteranterior testis length and oral sucker width) and distance between suckerswas unimportant. On the second axis, body width loaded weakly anddistance between suckers was the third strongest component (after ovarywidth and distance between testes). This shows that not only do these 2metacercarial characters distinguish between these 2 species, as shown bythe discriminant analysis; they are also important in explaining the totalmorphometric variance of the specimens, regardless of species.
Molecular analysis: In total, ITS sequences were obtained from 25specimens and COI sequences were obtained from 36 specimens, com-prising 10 metacercariae of C. complanatum and 23 metacercariae and 3adults of C. marginatum. Sequences of both markers were obtained from 5C. complanatum metacercariae, and from 2 C. marginatum adults and 12 C.marginatum metacercariae as well as from 3 additional specimens(specimens 26, 27, 28) that could not be allied with either species. COI
TABLE 1. Metacercariae of Clinostomum marginatum and Clinostomum complanatum: measurements and statistical comparisons. Values in bold indicatesignificant differences between the species.
Clinostomum marginatum (n 5 10)
min–max (mean ± SD) mm
Clinostomum complanatum (n 5 11)
min–max (mean ± SD) mm
Equality of means
(test statistic [t2(0.05),19], P-value)
Oral collar width 502–874 (721 ± 100) 686–1,030 (820 ± 110) 2.214, 0.039
Body length 4,215–6,346 (5,402 ± 672) 4,495–7,874 (5,741 ±1,223) 0.891, 0.389
Body width 979–1,532 (1,329 ± 173) 1,635–2,434 (1,934 ±239) 6.635, ,0.0005
Body length/width 3.050–4.403 (4.03 ± 0.422) 2.200–4.369 (2.994 ± 0.661) .
Oral sucker (OS) length 238–599 (312 ± 104) 259–337 (294 ± 27) 20.392, 0.699
OS width 194–586 (290 ± 106) 284–507 (401 ± 74) 2.928, 0.009
OS width/body width 0.816–1.053 (0.925 ± 0.071) 1.065–1.666 (1.358 ± 0.181) .
Ventral sucker (VS) length 510–757 (669 ± 64) 637–910 (795 ± 77) 3.449, 0.003
VS width 571–804 (708 ± 60) 766–952 (839 ± 60) 3.617, 0.002
VS width/OS width 0.987–1.118 (1.060 ± 0.043) 1.781–2.694 (2.140 ± 0.318) .
VS width/body width 0.468–0.616 (0.529 ± 0.045) 0.391–0.494 (0.436 ± 0.027) .
Distance between suckers 959–1,491 (1,243 ± 142) 860–1,115 (1,020 ± 84) 24.348, ,0.0005
Anterior testis (AT) length 231–413 (307 ± 53) 316–957 (484 ± 178) 3.185, 0.005
AT width 264–498 (389 ± 77) 273–559 (412 ± 74) 0.765, 0.454
AT width/length 0.681–1.720 (1.295 ± 0.294) 0.457–1.217 (0.905 ± 0.216) .
Posterior testis (PT) length 233–457 (327 ± 57) 245–441 (328 ± 63) 20.062, 0.951
PT width 295–492 (405 ± 56) 408–602 (493 ± 56) 3.633, 0.002
PT width/length 0.891–1.521 (1.254 ± 0.171) 1.089–1.876 (1.544 ± 0.286) .
Distance between testes 196–393 (320 ± 56) 214–527 (353 ± 91) 1.115, 0.279
Ovary length 85–142 (115 ± 16) 135–164 (149 ± 11) 5.505, ,0.0005
Ovary width 62–97 (80 ± 13) 97–178 (129 ± 24) 5.713, ,0.0005
Ovary width/length 0.434–0.968 (0.710 ± 0.168) 0.589–1.086 (0.866 ± 0.138) .
Cirrus sac (CS) length 172–343 (256 ± 48) 209–405 (296 ± 59) 1.633, 0.119
CS width 119–153 (137 ± 14) 124–197 (157 ± 24) 2.385, 0.032
CS length/body length 0.027–0.061 (0.048 ± 0.010) 0.034–0.071 (0.053 ± 0.013) .
FIGURE 5. Principal components analysis of 17 morphometrics in 21metacercariae of Clinostomum complanatum and Clinostomum margina-tum. The orientation and length of overlaid vectors is indicative of andproportionate to the importance of morphometrics along both axes (2 of17 are labeled).
CAFFARA ET AL.—DISTINGUISHING CLINOSTOMUM SPP. 887

sequences only were obtained from 10 metacercariae and 1 adult of C.marginatum, and 3 metacercariae of C. complanatum. ITS only was obtainedfrom a single metacercaria of C. marginatum and 2 of C. complanatum. Thequality of chromatograms for both mtDNA and rDNA targets wasgenerally good. However, chromatograms of ITS were either obtained inonly 1 direction or were of poorer quality in 6 of 23 cases (6, in 1 direction,but high quality); for COI chromatograms, this occurred in 17 of 36specimens (7, in 1 direction, 6 of medium quality, and 4 of low quality).
Sequences of the ITS region were 1,035 bp (588 bp ITS1, 159 bp 5.8S, and288 bp ITS2) in C. marginatum and 1,026 bp (584 bp ITS1, 159 bp 5.8S, and283 bp ITS2) in C. complanatum. The amplified COI fragments in C.marginatum and C. complanatum were about 600 bp in length in both species.
Except for 3 specimens (specimens 26, 27, 28) discussed further below,there were 65 differences between C. marginatum and C. complanatum inthe alignment of ITS sequences, 41 bp in ITS1, and 24 bp in ITS2. Therewas a 4-bp deletion in ITS1 and a 6-bp deletion in ITS2 in C.complanatum. At the 59 end of ITS1, 6 short tandem TA repeats occurredin the ITS sequences of C. marginatum, but not in C. complanatum.Neither C. marginatum nor C. complanatum showed differences in 5.8Ssequences. The rDNA sequences obtained from C. complanatum areidentical to those reported by Dzikowski et al. (2004) for this species; thosewe obtained from C. marginatum are not contiguous with the 18S rDNAsequence of those authors. There were 166 differences between the 2species in the 661-bp alignment of COI nucleotide sequences and 15differences in translated amino acids.
The K2P distance between the species was 7.3% over 579 positionsamong all 22 ITS sequences (Fig. 6). Sequences of COI diverged by amean of 19.4% (range 18.3–21.5% over 168 positions, n 5 33 sequences)
between the species. There was no variation within C. marginatum in ITSsequences over 579 positions (n 5 17). The mean intraspecific variation inCOI sequences of this species was 0.6% (range 0–1.7% over 366 positions,n 5 25). In analyses limited to 21 bidirectional COI sequences $420 bp,mean intraspecific divergence in C. marginatum COI was 0.4% (range 0–1.1%). Most of the COI nucleotide variation within C. marginatum wasnot reflected in sequences of translated amino acids, which were identicalexcept at 1 position, where 21 of 25 specimens had isoleucine, 3 had valine,and 1 had threonine. At this position, C. complanatum had valine. WithinC. complanatum, mean intraspecific distance was 0% (over 745 positions, n5 9) in ITS and 0.8% (range 0–1.9% over 318 positions, n 5 8) in COI.There was no variation in translated amino acids COI within C.complanatum. Metacercariae and adults identified as C. marginatum hadidentical, or highly similar, ITS and COI sequences.
The 2 species were not reciprocally monophyletic in neighbor-joininganalysis (Fig. 6). This was due to 3 specimens showing indeterminategenetic affinity (Table II, Fig. 6). One metacercaria collected from B.barbus in Italy (specimen 26) had ITS sequences identical to C.complanatum and was morphologically indistinguishable from this species.However, the COI sequence initially obtained from this specimen wasmore similar, but still quite divergent (,10%), from C. marginatum(Fig. 6). A second sequencing of the COI PCR product of specimen 26, aswell as a cloned sequence, both yielded a C. complanatum haplotype.Specimen 27, collected from Lithobates pipiens Schreber, in NorthAmerica, had the same unique COI haplotype initially sequenced forspecimen 26, but its ITS was identical to C. marginatum; this specimen wasmorphologically similar to C. marginatum, but possessed thick cuticularspines typical of Clinostomum attenuatum, a species not included in this
Figure 6. Neighbor-joining analyses of(A) nucleotide and (B) amino acidsequences of cytochrome c oxidase I(COI) and of (C) nucleotide sequencesof internal transcribed spacer (ITS) from39 Clinostomum specimens and selectedsequences from other studies. (A) Ki-mura 2-parameter distances among COInucleotide sequences using 168 basescommon to all specimens. (B) Dayhoffdistances among translated COI aminoacids with the use of 56 residues commonto all specimens. (C) Kimura 2-parameterdistances among ITS sequences with theuse of 557 bases common to all speci-mens. (A–C) are on the same scale.Specimens 1–25 and 37 are Clinostomummarginatum and specimens 29–36 and38–39, are Clinostomum complanatum.Specimens 26–28 are potential hybrids(see text, Table II). AY245701 is C.complanatum from Dzikowski et al.(2004) and HQ439578–HQ439579 arefrom Bonett et al. (2011). Host speciesare denoted by Amru 5 Ambloplitesrupestris, Arhe 5 Ardea herodias, Baca5 Barbus canis, Baba 5 Barbus barbus,Bame 5 Barbus meridionalis, Bubo 5Anaxyrus (Bufo) boreas, Etni 5 Etheos-toma nigrum, Euty 5 Eurycea tynerensis,Legi 5 Lepomis gibbosus, Mido 5Micropterus dolomieu, Misa 5 Micro-pterus salmoides, Neme 5 Neogobiusmelanostomus, Nocr 5 Notemigonus cry-soleucas, Pefl 5 Perca flavescens, Pino 5Pimephales notatus, Racl 5 Lithobates(Rana) clamitans, Rapi 5 Lithobates(Rana) pipiens, Sqce 5 Squalius cephalus.
888 THE JOURNAL OF PARASITOLOGY, VOL. 97, NO. 5, OCTOBER 2011

study. Specimen 28, from Lithobates clamitans in Quebec, had the sameunique COI haplotype as specimens 26 and 27 and ITS sequence wasidentical to C. marginatum; the morphological features of the specimen 28were identical to 27, showing thick cuticular spines. The translated aminoacids of the unique COI haplotype common to specimens 26, 27, and 28were identical to those of most (21 of 25) C. marginatum specimens. One ofthe COI sequences from a heretoplasmous Clinostomum sp. specimen fromEurycea tynerensis from Oklahoma studied by Bonett et al. (2011)clustered with the main C. marginatum haplotype, whereas the othershowed intermediate similarity to both C. marginatum and the specimens26–28 sequence cluster (Fig. 6).
DISCUSSION
Morphological differences in both adults and metacercariae of
C. complanatum and C. marginatum provided strong evidence of
the validity of the latter species. Three important morphological
differences were observed between metacercariae in the 2 species,
i.e., the organization of the genital complex, body width, and
distance between the suckers. Although the genital complex did
not emerge from statistical analysis because simple measurements
do not reflect the complex structural relationships of this
character, it is probably a more reliable diagnostic feature. The
shape of cirrus sac changes during development (Vianna et al.,
2003), but this does not affect the position of this diagnostic
structure (Ukoli, 1966), which is easily seen providing that the
uterus is not completely filled with eggs. Like Ukoli (1966), we
also found that the position of the genital pore is a useful
character for discriminating between these Clinostomum species,
although other authors have argued that this character can vary
with different fixation or flattening techniques (Baer, 1933), or
during development (Vianna et al., 2003).
Thus, our morphologic study identified several anatomic features
useful for distinguishing these 2 species both at metacercariae and
adult stages. Our redescription of C. marginatum is strongly
consistent with those of Braun (1901), Osborne (1911, 1912), Cort
(1913), Maccagno (1934), Ukoli (1966), and Lunaschi and Drago
(2009). Our statistical analysis also suggests that metacercariae of C.
complanatum and C. marginatum can be distinguished by the
distance between the ventral and oral sucker and by the width of the
body. These characters have appealing simplicity and accessibility,
and can be recognized in unstained specimens by nonexperts.
However, there are several reasons why further work is necessary to
verify their utility. First, we did not investigate different stages of
metacercaria development. Second, these traits may vary in
specimens fixed or flattened with techniques other than those
employed here. Third, these traits may vary in different host species
(e.g., Blankespoor, 1974; Perkins et al. 2011).
Except for 3 specimens (discussed below), the morphological
differences of C. marginatum and C. complanatum were well
supported by nuclear and mitochondrial sequences that provide
independent estimates of species boundaries. Both ITS and COI
are commonly used to infer species boundaries and relationships
in flatworms (Morgan and Blair, 1995, 1998; Leon-Regagnon
et al., 1999; Tkach et al., 2000; Kostadinova et al., 2003; Scholz
et al., 2004; Vilas et al., 2005).
In digeneans, ITS is relatively conserved, with divergence
generally ranging from ,1 to 20% between species and variation
less than 1% within species (Morgan and Blair, 1998; Nolan and
Cribb, 2005). In most species studied to date, ITS1 is more
variable than ITS2, possibly because of evolutionary constraints
on the secondary structure of ITS2 (Nolan and Cribb, 2005;
Coleman, 2007). Tandem repeats similar to those we observed in
ITS1 in C. marginatum have been reported in diverse digeneans
(e.g., Morgan and Blair, 1995; van Herwerden et al., 1999; Tkach
et al., 2000), but are notably absent in C. complanatum, C.
cutaneum, and C. phalacrocoracis (Gustinelli et al., 2010; present
study).
A number of other studies of species of digeneans have employed
a fragment of COI downstream from the region that we studied and
have generally obtained better resolution of species than those using
rDNA (Vilas et al., 2005). The 600-bp span of COI we sequenced
was more variable both within, and between, C. complanatum and
C. marginatum than ITS. In general, the signal of within versus
between species variation was clearer in sequences of COI than ITS.
Although the COI fragments contained less information than ITS,
i.e., 600 versus 1,000 bp, the divergence between the species in COI
was over 3 times greater than in ITS. Indeed, the low level of
variation in ITS between 2 other Clinostomum species reported by
Gustinelli et al. (2010) (1.2–2.1%) indicates these data may not be
sufficient for discriminating species of Clinostomum without
morphological support. With smaller numbers of differences in
sequences of greater length, base-calling errors become more of a
problem. Since almost all (33/36) specimens were better resolved by
COI than by ITS data, the mitochondrial markers seem sufficient
for routine discrimination of these species in most contexts.
However, the specimens with indeterminate genetic attributes
should not be ignored. We speculate that these could be
descendants of hybridization between the 2 species. Maccagno
(1934) reported C. marginatum (consistent with our description)
TABLE 2. Summary of genetic and morphological attributes of 3 specimens of Clinostomum examined from North America and Europe. Specimens arenumbered as in Figure 5.
Specimen Collection Morphology
c oxidase I (COI)
nucleotides
COI amino
acids
Cloned COI
nucleotides/repeated
sequencing
Internal
transcribed spacer
26 .Barbus barbus, Santerno
River, Italy, 2010
.Clinostomum
complanatum
.Unique haplotype .Clinostomum
marginatum
.C. complanatum .C. complanatum
27 .Lithobates pipiens,
North America, 2009*
.Clinostomum
attenuatum/
C. marginatum
.Unique haplotype .C. marginatum – .C. marginatum
28 .Lithobates clamitans, Quebec,
Canada, 2010
.C. attenuatum/
C. marginatum
.Unique haplotype .C. marginatum – .C. marginatum
* A live frog was obtained from Boreal Northwest (St. Catherine’s, Ontario, Canada), a biological supply company that could not provide specific geographic information.
CAFFARA ET AL.—DISTINGUISHING CLINOSTOMUM SPP. 889

in Italy; the 2 species may not be as geographically isolated as
commonly supposed. Hybridization could explain the identical
COI sequences of 3 specimens on 2 continents, which are more
similar to C. marginatum sensu stricto and identical to the latter
when translated to amino acids. Matrilineal descendants of
hybrids could retain C. marginatum mtDNA, and successive
backcrosses would quickly lead to sweeps of nDNA from locally
abundant species. Thus, the ITS sequences of the North American
specimens are those of C. marginatum, and that of the European
specimen is C. complanatum. Because nDNA encodes the overall
phenotype, these lineages would be expected to show greater
morphological resemblance to the locally abundant species. The
mitochondrial heteroplasmy of the European specimen is also
consistent with hybridization and is intriguingly similar to what
Bonett et al. (2011) observed in a specimen of C. marginatum in
Oklahoma. Similar discordance between nuclear and mitochon-
drial genes has been used to demonstrate introgressive hybridiza-
tion in diverse animals (Ballard and Whitlock, 2004), including
digeneans (reviewed by Detwiler and Criscione, 2010). However,
the pattern we observed could also arise from incomplete lineage
sorting and historical introgression. Ultimately, further work,
including larger-scale sampling and other molecular markers, is
needed to clarify the status of these lineages and, therefore, the
utility of these markers for distinguishing Clinostomum species in
these problem cases. It is notable that the use of any of the 4 data
types alone (COI, ITS, morphology, geographic origin) could
have led to an erroneous conclusion with regard to the taxonomic
status of these 3 specimens, and that their specific identity is still
uncertain when all data are considered together.
In the present study, we reconfirmed the validity of C. marginatum
with novel genetic data and morphological analyses of 2 life-cycle
stages. We evaluated multiple molecular and morphological charac-
ters for discriminating C. marginatum from C. complanatum, linked
our findings to historical taxonomic work, and provide further
evidence of low levels of hybridization in Clinostomum species (see
Bonett et al., 2011). Our molecular data linked metacercariae and
adults of C. marginatum from naturally infected hosts. We hope our
results will stimulate further taxonomic work that unites both
morphology and DNA on these and other species of Clinostomum.
To this end, a COI sequence library from diverse Clinostomum
species, together with complete morphological descriptions, would be
fruitful for the reorganization of the taxonomy of this genus.
ACKNOWLEDGMENTS
The research was supported by the Italian Ministry of Education,University and Research (MIUR–PRIN08) and by the Natural Sciencesand Engineering Research Council of Canada, through funding to theCanadian Barcode of Life Network, and by Environment Canada’sSTAGE funding to David Marcogliese. We thank Mike Kinsella (Missoula,Montana), Gary Hendrickson (Department of Fisheries Biology, Hum-boldt State University, Arcata, California), Louise Champoux (CanadianWildlife Service), Andree Gendron (Environment Canada), BrandonBallengee (McGill University), Daniel McLaughlin (Concordia University),and Pieter Johnson (University of Colorado), who generously providedClinostomum specimens, or host animals, or both. We are grateful forcritical input by Ron Bonett (University of Tulsa), Chris Blanar (NovaSoutheastern University), and an anonymous reviewer.
LITERATURE CITED
AMIN, O. M. 1969. Helminth fauna of suckers (Catostomidae) of the GilaRiver system, Arizona. II. Five parasites from Catostomus spp.American Midland Naturalist 82: 429–443.
BAER, J. G. 1933. Note sur un nouveau trematode, Clinostomumlophophallum sp. nov., avec quelques considerations generales sur lafamille des Clinostomidae. Revue Suisse de Zoologie 40: 317–342.
BALLARD, J. W. O., AND C. WHITLOCK. 2004. The incomplete naturalhistory of mitochondria. Molecular Ecology 13: 729–744.
BLANKESPOOR, H. D. 1974. Host-induced variation in Plagiorchis nobleiPark 1936 (Plagiorchiidae: Trematoda). American Midland Natural-ist 92: 415–433.
BONETT, R. M., M. A. STEFFEN, A. L. TRUJANO-ALVAREZ, S. D. MARTIN, C.R. BURSEY, AND C. T. MCALLISTER. 2011. Distribution, abundance,and genetic diversity of Clinostomum spp. metacercariae (Trematoda:Digenea) in a modified Ozark stream system. Journal of Parasitology97: 177–184.
BOWLES, J., D. BLAIR, AND D. P. MCMANUS. 1995. A molecular phylogenyof the human schistosomes. Molecular Phylogenetics and Evolution4: 103–109.
BRAUN, M. 1901. Die arten der gattung Clinostomum Leidy. ZoologischeJahrbucher. Abteilung fur Systematik, Okologie und Geographie derTiere 14: 1–48.
COLEMAN, A. W. 2007. Pan-eukaryote ITS2 homologies revealed by RNAsecondary structure. Nucleic Acids Research 35: 3322–3329.
CORT, W. W. 1913. Notes on the trematode genus Clinostomum.Transactions of the American Microscopical Society 32: 169–183.
DETWILER, J. T., AND C. D. CRISCIONE. 2010. An infectious topic inreticulate evolution: Introgression and hybridization in animalparasites. Genes 1: 102–123.
DOLLFUS, R. P. 1950. Trematodes recoltes au Congo Belge par leProfesseur Paul Brien (mai-aout, 1937). Annales du Musee du CongoBelge. C. Zoologie 1: 1–136.
DOWSETT, J. A., AND G. LUBINSKY. 1980. Clinostomum complanatum orClinostomum marginatum? (Trematoda: Clinostomatidae). A problemof priority and synonymy. Canadian Journal of Zoology 58: 680–682.
DZIKOWSKI, R., M. G. LEVY, M. F. POORE, J. R. FLOWERS, AND I. PAPERNA.2004. Clinostomum complanatum and Clinostomum marginatum(Rudolphi, 1819) (Digenea: Clinostomidae) are separate speciesbased on differences in ribosomal DNA. Journal of Parasitology90: 413–414.
FEIZULLAEV, N. A., AND S. S. MIRZOEVA. 1983. Revision of the superfamilyClinostomoidea and analysis of its system. Parazitologiya 17: 3–11.
FOREST, J. J. H., AND D. K. CONE. 2011. Helminth parasites of goldenshiner Notemigonus crysoleucas (Pisces: Cyprinidae) from Ontarioand Nova Scotia, Canada. Comparative Parasitology 78: 220–222.
GUSTINELLI, A., M. CAFFARA, D. FLORIO, E. O. OTACHI, E. M. WATHUTA,AND M. L. FIORAVANTI. 2010. First description of the adult stage ofClinostomum cutaneum Paperna, 1964 (Digenea: Clinostomidae) fromgrey herons Ardea cinerea L. and a redescription of the metacercariafrom the Nile tilapia Oreochromis niloticus niloticus (L.) in Kenya.Systematic Parasitology 76: 39–51.
HILLIS, D. M., AND M. T. DIXON. 1991. Ribosomal DNA: Molecularevolution and phylogenetic inference. Quarterly Review of Biology66: 411–453.
HOPKINS, S. H. 1933. Note on the life history of Clinostomum marginatum(Trematoda). Transactions of the American Microscopical Society52: 147–149.
KOSTADINOVA, A., E. A. HERNIOU, J. BARRETT, AND D. T. LITTLEWOOD. 2003.Phylogenetic relationships of Echinostoma rudolphi, 1809 (Digenea:Echinostomatidae) and related genera re-assessed via DNA andmorphological analysis. Systematic Parasitology 54: 159–176.
LARSON, O. R., G. L. UGLEM, AND J. L. KOOK. 1988. Fine structure andpermeability of the metacercarial cyst wall of Clinostomum margina-tum (Digenea). Parasitology Research 74: 352–355.
LEON-REGAGNON, V., D. R. BROOKS, AND G. PEREZ-PONCE DE LEON. 1999.Differentiation of Mexican species of Haematoloechus loos, 1899(Digenea: Plagiorchiformes): Molecular and morphological evidence.Journal of Parasitology 85: 935–946.
LOCKE, S. A., J. D. MCLAUGHLIN, S. DAYANANDAN, AND D. J.MARCOGLIESE. 2010. Diversity and specificity in Diplostomum spp.metacercariae in freshwater fishes revealed by cytochrome c oxidase Iand internal transcribed spacer sequences. International Journal forParasitology 40: 333–343.
———, ———, AND D. J. MARCOGLIESE. 2010. DNA barcodes showcryptic diversity and a potential physiological basis for hostspecificity among Diplostomoidea (Platyhelminthes: Digenea) para-
890 THE JOURNAL OF PARASITOLOGY, VOL. 97, NO. 5, OCTOBER 2011

sitizing freshwater fishes in the St. Lawrence River, Canada.Molecular Ecology 19: 2813–2827.
LUNASCHI, L. I., AND F. B. DRAGO. 2009. Digenean parasites of six speciesof bird from Formosa province, Argentina. Revista Mexicana deBiodiversidad 80: 39–46.
MACCAGNO, T. 1934. Clinostomum marginatum Rud. In Europa. Bollettinodei Musei di Zoologia e Anatomia Comparata Torino, XLIV–serieIII 44: 1–8.
MANTER, H. W. 1938. A collection of trematodes from Florida Amphibia.Transactions of the American Microscopical Society 57: 26–37.
MATTHEWS, D., AND T. H. CRIBB. 1998. Digenetic trematodes of the genusClinostomum Leidy, 1856 (Digenea: Clinostomidae) from birds ofQueensland, Australia, including C. wilsoni n. sp. from Egrettaintermedia. Systematic Parasitology 39: 199–208.
MCALLISTER, C. T. 1990. Metacercaria of Clinostomum complanatum(Rudolphi, 1814) (Trematoda: Digenea) in a Texas salamander,Eurycea neotenes (Amphibia: Caudata), with comments on C.marginatum (Rudolphi, 1819). Journal of the HelminthologicalSociety of Washington 57: 69–71
———, C. R. BURSEY, J. A. CRAWFORD, A. R. KUHNS, C. SHAFFER, AND
S. E. TRAUTH. 2010. Metacercariae of Clinostomum (Trematoda:Digenea) from three species of Ambystoma (Caudata: Ambystoma-tidae) from Arkansas and Illinois, U.S.A. Comparative Parasitology77: 25–30.
———, C. R. BURSEY, M. L. NEIMILLER, AND B. T. MILLER. 2007. Anoteworthy infection of Clinostomum complanatum (Digenea: Clin-ostomatidae) in the cave salamander Eurycea lucifuga (Caudata:Plethodontidae) from north central Tennessee. Texas Journal ofScience 59: 321–326.
MORGAN, J. A. T., AND D. BLAIR. 1995. Nuclear rDNA ITS sequence variationin the trematode genus Echinostoma: An aid to establishing relationshipswithin the 37-collar-spine group. Parasitology 111: 609–615.
———, AND ———. 1998. Relative merits of nuclear ribosomal internaltranscribed spacer and mitochondrial CO1 and ND1 genes fordistinguishing among Echinostoma species (Trematoda). Parasitology116: 289–297.
MOSZCZYNSKA, A., S. A. LOCKE, J. D. MCLAUGHLIN, D. J. MARCOGLIESE,AND T. J. CREASE. 2009. Development of primers for the mitochon-drial cytochrome c oxidase I gene in digenetic trematodes illustratesthe challenge of barcoding parasitic helminths. Molecular EcologyResources 9: 75–82.
NOLAN, M. J., AND T. H. CRIBB. 2005. The use and implications ofribosomal DNA sequencing for the discrimination of digeneanspecies. Advances in Parasitology 60: 101–163.
OSBORN, H. L. 1911. On the distribution and mode of occurrence in theUnited States and Canada of Clinostomum marginatum, a trematodeparasitic in fish, frogs and birds. Biological Bulletin 20: 350–366.
———. 1912. On the structure of Clinostomum marginatum, a trematodeparasite of the frog, bass and heron. Journal of Morphology 23: 189–229.
PERKINS, S. L., E. S. MARTINSEN, AND B. G. FALK. 2011. Do moleculesmatter more than morphology? Promises and pitfalls in parasites.Parasitology. (In press).
PRICE, E. W. 1938. A redescription of Clinostomum intermedialis Lamont(Trematoda: Clinostomidae) with a key to the species of the genus.Proceedings of the Helminthological Society of Washington 5: 11–13.
PRITCHARD, M. H., AND G. KRUSE. 1982. The collection and preservationof animal parasites. University of Nebraska Press, Lincoln,Nebraska, 141 p.
SCHOLZ, T., A. SKERIKOVA, T. SHIMAZU, AND M. J. GRYGIER. 2004. Ataxonomic study of species of Bothriocephalus Rudolphi, 1808(Cestoda: Pseudophyllidea) from eels in Japan: Morphological andmolecular evidence for the occurrence of B. claviceps (Goeze, 1782)and confirmation of the validity of B. japonicus Yamaguti, 1934.Systematic Parasitology 57: 87–96.
SMALLWOOD, W. M. 1914. Preliminary report on diseases of fish in theAdirondacks: Field notes on Clinostomum marginatum with acontribution to its life history, preliminary report. New York StateCollege of Forestry at Syracuse University Technical PublicationNo. 1, Series 14 2: 8–27.
TAMURA, K., D. PETERSON, N. PETERSON, G. STECHER, M. NEI, AND S.KUMAR. 2011 MEGA5: Molecular evolutionary genetics analysisusing maximum likelihood, evolutionary distance, and maximumparsimony methods. Molecular Biology and Evolution. (In press).
TKACH, V. V., J. PAWLOWSKI, AND V. P. SHARPILO. 2000. Molecular andmorphological differentiation between species of the Plagiorchisvespertilionis group (Digenea, Plagiorchiidae) occurring in Europeanbats, with a re-description of P. vespertilionis (Muller, 1780).Systematic Parasitology 47: 9–22.
TRAVASSOS, L., J. F. TEIXEIRA DE FREITAS, AND A. KOHN. 1969.Trematodeos do Brasil. Memorias do Instituto Oswaldo Cruz 67:1–886.
UKOLI, F. M. A. 1966. On Clinostomum tilapiae n. sp., and C.phalacrocoracis Dubois, 1931 from Ghana, and a discussion of thesystematics of the genus Clinostomum Leidy, 1856. Journal ofHelminthology 40: 187–214.
VAN HERWERDEN, L., D. BLAIR, AND T. AGATSUMA. 1999. Intra- andinterindividual variation in ITS1 of Paragonimus westermani (Trem-atoda: Digenea) and related species: Implications for phylogeneticstudies. Molecular Phylogenetics and Evolution 12: 67–73.
VIANNA, R. T., JR., J. PEREIRA, AND D. A. BRANDAO. 2003. Ontogeneticvariation of metacercariae of Clinostomum complanatum (Rudolphi,1814) (Digenea: Clinostomidae). Comunicacoes do Museu deCiencias e Tecnologia da PUCRS, Serie Zoologia, Porto Alegre 16:223–243.
VILAS, R., C. D. CRISCIONE, AND M. S. BLOUIN. 2005. A comparisonbetween mitochondrial DNA and ribosomal internal transcribedregions in prospecting for cryptic species of platyhelminth parasites.Parasitology 131: 839–846.
YAMAGUTI, S. 1933. Studies on the helminth fauna of Japan. Part 1.Trematodes of birds, reptiles and mammals. Japanese Journal ofZoology 5: 1–134.
———. 1971. Synopsis of digenetic trematodes of vertebrates, vols. 1 and2. Keigaku, Tokyo, Japan, 1074 p.
CAFFARA ET AL.—DISTINGUISHING CLINOSTOMUM SPP. 891




















