
First come, last primed: FN400 reflects post-encoding editing of the memory trace
14
Behavioural Brain Research 266 (2014) 63–76 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Research report First come, last primed: FN400 reflects post-encoding editing of the memory trace Giulio Pergola a,b,∗ , Christian Bellebaum c,d , Boris Suchan c a Department of Basic Medical Science, Neuroscience and Sense Organs, University of Bari – ‘Aldo Moro’, Piazza Giulio Cesare, 11, I-70100 Bari, Italy b Cognitive Neuroscience Sector, International School for Advanced Studies (SISSA), via Bonomea 265, I-34136 Trieste, Italy c Department of Neuropsychology, Ruhr-University Bochum, D-44780 Bochum, Germany d Institute of Experimental Psychology, Heinrich Heine University Düsseldorf, Universitaetsstrasse 1, 40225 Düsseldorf, Germany highlights • Aim: the role of the FN400 component in conceptual priming and associative memory. • Encoding: learning same- and different-category associations between picture pairs. • Retrieval: participants performed a recognition and recall test. • 260–350 ms: frontal category effect selectively for items shown first within a pair. • Exploratory analyses: earlier effects in the time windows 40–90 ms and 150–200 ms. article info Article history: Received 10 September 2013 Received in revised form 23 February 2014 Accepted 28 February 2014 Available online 11 March 2014 Keywords: Event-related potential Conceptual priming FN400 Associative encoding Frontal negativity abstract Explicit associative memory relies on different neural substrates depending on similarity of the items associated. Extant literature dissociates old/new effects elicited by homogeneous and heterogeneous pairs. However, current results scarcely address potential priming effects induced by the presentation sequence. In the present ERP study, participants learned associations between two pictures, which either belonged to the same semantic category (e.g., animal–animal), or to different categories (e.g., animal–scene). Pic- tures forming a pair were shown sequentially, allowing for investigation of the different neural processes related to presentation of the first and the second item. After the study phase, participants performed a recognition judgment. After recognition, participants were asked to recall the associated picture. During retrieval, between 260 and 350 ms post-stimulus there was a significant frontal effect of cat- egory (i.e., same-category versus different-category), but only for items shown first within a pair. In the 350–600 ms time window the parietal old/new effect was unaffected by semantic category, but was modulated by presentation order. Exploratory analyses revealed even earlier effects in the time windows 40–90 ms and 150–200 ms. This evidence supports the priming account of the FN400 and highlights the importance of sequence effects in electrophysiological activity during episodic retrieval. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Explicit memory is thought to consist of an episodic and a semantic component [1] intermingled with each other [2]. Episodic memory allows encoding of information after a single exposure and ∗ Corresponding author at: Department of Basic Medical Science, Neuroscience and Sense Organs, University of Bari – ‘Aldo Moro’, Piazza Giulio Cesare, 11, I-70100 Bari, Italy. Tel.: +39 3408223991; fax: +39 080 5593204. E-mail address: [email protected] (G. Pergola). is dependent on the activity of the medial temporal lobe (MTL; for reviews, see Refs. [3,4]. Other forms of memory are spared when the MTL is damaged, for example various forms of implicit memory [5,6], including priming [7]. Priming is the perceptual or conceptual alteration of a stimulus processing after exposure to the same or another stimulus [8] and might operate in parallel with recognition memory when stimuli are processed [8,9]. Recognition memory, i.e., the ability to judge previous occurrence of an event or item, is increasingly described using a dual model [10,11], which proposes the existence of two components: an explicit one – recollection – and a more implicit-like cognitive module, called familiarity. http://dx.doi.org/10.1016/j.bbr.2014.02.050 0166-4328/© 2014 Elsevier B.V. All rights reserved.
Transcript of First come, last primed: FN400 reflects post-encoding editing of the memory trace

R
Fm
Ga
b
c
d
h
•••••
a
ARRAA
KECFAF
1
sm
aB
h0
Behavioural Brain Research 266 (2014) 63–76
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
esearch report
irst come, last primed: FN400 reflects post-encoding editing of theemory trace
iulio Pergolaa,b,∗, Christian Bellebaumc,d, Boris Suchanc
Department of Basic Medical Science, Neuroscience and Sense Organs, University of Bari – ‘Aldo Moro’, Piazza Giulio Cesare, 11, I-70100 Bari, ItalyCognitive Neuroscience Sector, International School for Advanced Studies (SISSA), via Bonomea 265, I-34136 Trieste, ItalyDepartment of Neuropsychology, Ruhr-University Bochum, D-44780 Bochum, GermanyInstitute of Experimental Psychology, Heinrich Heine University Düsseldorf, Universitaetsstrasse 1, 40225 Düsseldorf, Germany
i g h l i g h t s
Aim: the role of the FN400 component in conceptual priming and associative memory.Encoding: learning same- and different-category associations between picture pairs.Retrieval: participants performed a recognition and recall test.260–350 ms: frontal category effect selectively for items shown first within a pair.Exploratory analyses: earlier effects in the time windows 40–90 ms and 150–200 ms.
r t i c l e i n f o
rticle history:eceived 10 September 2013eceived in revised form 23 February 2014ccepted 28 February 2014vailable online 11 March 2014
eywords:vent-related potentialonceptual primingN400ssociative encodingrontal negativity
a b s t r a c t
Explicit associative memory relies on different neural substrates depending on similarity of the itemsassociated. Extant literature dissociates old/new effects elicited by homogeneous and heterogeneouspairs. However, current results scarcely address potential priming effects induced by the presentationsequence.
In the present ERP study, participants learned associations between two pictures, which either belongedto the same semantic category (e.g., animal–animal), or to different categories (e.g., animal–scene). Pic-tures forming a pair were shown sequentially, allowing for investigation of the different neural processesrelated to presentation of the first and the second item. After the study phase, participants performed arecognition judgment. After recognition, participants were asked to recall the associated picture.
During retrieval, between 260 and 350 ms post-stimulus there was a significant frontal effect of cat-egory (i.e., same-category versus different-category), but only for items shown first within a pair. In
the 350–600 ms time window the parietal old/new effect was unaffected by semantic category, but wasmodulated by presentation order. Exploratory analyses revealed even earlier effects in the time windows40–90 ms and 150–200 ms.This evidence supports the priming account of the FN400 and highlights the importance of sequenceeffects in electrophysiological activity during episodic retrieval.
© 2014 Elsevier B.V. All rights reserved.
. Introduction
Explicit memory is thought to consist of an episodic and aemantic component [1] intermingled with each other [2]. Episodicemory allows encoding of information after a single exposure and
∗ Corresponding author at: Department of Basic Medical Science, Neurosciencend Sense Organs, University of Bari – ‘Aldo Moro’, Piazza Giulio Cesare, 11, I-70100ari, Italy. Tel.: +39 3408223991; fax: +39 080 5593204.
E-mail address: [email protected] (G. Pergola).
ttp://dx.doi.org/10.1016/j.bbr.2014.02.050166-4328/© 2014 Elsevier B.V. All rights reserved.
is dependent on the activity of the medial temporal lobe (MTL; forreviews, see Refs. [3,4]. Other forms of memory are spared whenthe MTL is damaged, for example various forms of implicit memory[5,6], including priming [7]. Priming is the perceptual or conceptualalteration of a stimulus processing after exposure to the same oranother stimulus [8] and might operate in parallel with recognitionmemory when stimuli are processed [8,9]. Recognition memory,
i.e., the ability to judge previous occurrence of an event or item, isincreasingly described using a dual model [10,11], which proposesthe existence of two components: an explicit one – recollection– and a more implicit-like cognitive module, called familiarity.
6 l Brain
Retpcowod[f
pbpcpiddfd
MraPiocvpfeoop
bcBatiwetcat
wuw
aEtbcwimtrn
4 G. Pergola et al. / Behavioura
ecollection is thought to be a form of recall [12]. Familiaritynables recognition but not retrieval of information associated tohe item at hand. However, not all scholars agree on the dualrocessing model. The main alternative view, the “single pro-ess account”, explains the dichotomy between subcomponentsf recognition memory as the difference between stronger andeaker memory traces (i.e., stronger memory traces elicit a feeling
f recollection [4,13]). Proponents of this view acknowledge thatifferent behavioral outcomes reflect dissociable neural processing14]. However, the techniques used to separate recollection fromamiliarity are debated (for reviews, see Refs. [11,12,15–18]).
One way to examine the putatively different neural under-innings of recognition memory has been the assessment ofrain activity during associative encoding and retrieval. The dualrocessing model holds that familiarity supports memory for per-eptual associations, i.e., recognition of previous co-occurrence oferceptual features [19]. Mayes, Montaldi [20] proposed that famil-
arity enables associations between items that share a commonomain (within-domain associations), but not between items thato not share a domain (between-domain associations). Hence dif-erent explicit associations would recruit the MTL cortices to aifferent extent.
This theory is supported by evidence provided by Jager,ecklinger [9], who showed a double dissociation between event-
elated potentials (ERP) elicited by successful recognition ofssociations between perceptually similar versus different items.articipants studied pairs of faces that either belonged to differentndividuals (inter-item condition), or to the same individual (butne of the images had been morphed with another face; intra-itemondition). During retrieval, a frontal old/new effect in the inter-al 300–400 ms was specific for intra-item associations, whereas aarietal old/new effect in the interval 400–700 ms was only foundor inter-item associations [9]. The authors interpreted the formerffect as a correlate of familiarity and the latter effect as a correlatef recollection. Interestingly, the ERP also showed earlier frontalld/new effects, which was interpreted as reflecting perceptualriming (see also Ref. [21]).
The frontal old/new effect found by Jager, Mecklinger [9] resem-les a well-known component, called frontal N400 (FN400). Thisomponent has also been associated to conceptual priming [22].ased on behavioral evidence, it has been proposed that primingnd familiarity are correlated [23], but current evidence on thisopic is still controversial [24]. Paller, Lucas [24] suggested that onlyn restricted circumstances conceptual priming closely co-varies
ith familiarity. The two processes are more difficult to separatespecially for meaningful stimuli, such as words and nameable pic-ures. It is therefore unclear whether the processing underlyingonceptual priming may contribute to familiarity for some itemsnd not others, e.g., depending on meaningfulness of the stimuli orhe presentation sequence.
Notably, in the study by Jager, Mecklinger [9], retrieval cuesere always the first members of the pairs studied. It is currentlynknown whether their results also apply to cues shown secondithin a pair during encoding.
During associative encoding, the order of presentation mayffect the potency of a cue in eliciting recall. For instance, differentRP are elicited depending on whether the second member of a pairo encode does or does not carry associative information inducedy the presentation sequence [25]. Specifically, Kim, Vallesi [25]ould show that the semantic context in which pairs of associatesere studied modulated the amplitude of the N400 during encod-
ng. In previous work [26], we found that the onset of the second
ember of the pair, during associative encoding, triggered a sus-ained positivity, which in other studies appeared to specificallyelate to the process of associating information [27,28]. Unfortu-ately, no results have been reported by Kim, Vallesi [25] on the
Research 266 (2014) 63–76
retrieval phase of their study. We have shown that during retrieval,ERP amplitudes elicited by recognition cues differ based on theassociative load carried at encoding [26]. Hence, previous evidencereveals that associative information during encoding affects ERPat retrieval, and that during encoding, the onset of the secondmember of a pair elicits specific electrophysiological responses.However, previous results indicating the FN400 as a correlate offamiliarity during the retrieval phase of memory were limited toitems that did not carry – at the time of encoding – associativeinformation. In fact, these items were studied before appearanceof the second member of the pair. Extant evidence appears want-ing concerning the link between associative encoding and ERP atretrieval.
Surprisingly, few studies addressed the effect of presentationorder on associative encoding and on ERP during retrieval. Thebulk of the literature on order effects refers to memory for order,rather than addressing how stimuli presentation sequence affectsmemory for items. Caplan, Glaholt [29], however, reported distinc-tive electrophysiological activity for cued recall of associative pairs,compared to order memory for sequences of three words–namely,a left parietal and fronto-polar effect around 500 ms post-stimulusonset. Notably, Caplan, Glaholt [29] reported similar behavioralaccuracy for backward and forward cued recall (i.e., when the cuewas the first or the second member of the pair, respectively). Thisis not the case for free recall of item lists, in which forward recallis definitely facilitated compared to backward recall [30]. Further-more, Yang, Zhao [31] have shown that recognition memory showsa similar advantage for forward associative recognition, while con-firming the symmetry of cued recall. Another study relevant tothe effect of order on recognition memory has been reported byWiegand, Bader [32]. They found that the FN400 was specific ofassociative recognition for reversed-order pairs, and was not foundwith same-order pairs. However, in both studies [31,32] stimuliwere presented together on the screen, and not sequentially, andit is possible that ERP components at least partly reflected object-place associations or paired recognition.
Hence, although presentation order differentially affects differ-ent memory processes, its relevance to conceptual priming andrecognition memory has not yet been explored.
In our view, if the early frontal old/new effect reflects famil-iarity, but not priming, a greater effect could be predicted forlow-integration associations compared to high-integration asso-ciations [20]. However, the effect should equally apply to the firstand second member of the pair. To the best of our knowledge, nostudies addressed whether putative ERP correlates of familiarity,recollection or conceptual priming vary depending on presentationorder in associative memory tasks. Note that conceptual primingmay also be associative [33]. Since priming is normally operational-ized as the modification of the cognitive processing of a stimulusinduced by previous stimulation, it is expected that it is affectedby the sequence of presentation. Hence, we hypothesized that ERPeffects reflecting conceptual priming are modulated by presenta-tion order, which is not expected of electrophysiological substratesof familiarity.
We set up an ERP study to investigate the electrophysiologi-cal basis of presentation sequence in associative encoding, usingdifferently assorted pairs. In the study by Jager, Mecklinger [9]the similarity was perceptually defined, i.e., the two members ofa pair shared some perceptual features. We reasoned that usingsemantic categories could reduce the likelihood that recognitionis accomplished through familiarity (recollection being prevalentin this condition [34]). Conceptual priming effects depend on prior
access to conceptual information, hence semantic associations areexpected to modulate them. Finding an FN400 in such conditionswould strengthen the notion that it reflects conceptual priming,rather than familiarity, also for meaningful stimuli.
l Brain
oatcsr
mesrt[
ts
rprcosteer
2
2
tafPvtfToi1
2
4(p[
dAt3cpr3aaw
G. Pergola et al. / Behavioura
We used stimuli including pictures of animate and inanimatebjects (animals and tools) as well as scenes. Different-categoryssociations were picture pairs representing items belongingo different semantic categories (e.g., tool-scene), and same-ategory associations were composed by items belonging to theame semantic category (e.g., tool–tool). We analyzed separatelyetrieval cues shown first and second within a pair.
Based on the differential involvement of the hippocampus inemorizing different kinds of association [20], the associative cat-
gory could be expected to affect memory scores, particularlyhowing an advantage in recall for different-category items. Asegards presentation order, previous behavioral evidence suggestshat no difference should be found in cued recall performance30,31].
At encoding, we expected that a sequence of pictures belongingo different categories would elicit a larger N400, compared to aequence of pictures belonging to the same category [25,35,36].
At retrieval, we tested whether the FN400 characterized bothecognition cues shown first and second within a same-categoryair (as expected if FN400 reflects familiarity) or was specific ofecognition cues shown first [9]. We predicted that both same-ategory and different-category associations would elicit a parietalld/new effect, because the late parietal positivity [34] is thought toupport recall independent on the stimulus material. We also hadhe opportunity to test the effect of presentation sequence duringncoding on the late parietal positivity. Additionally, we conductedxploratory analyses to find effects earlier than 200 ms that mighteflect implicit memory processing.
. Methods
.1. Participants
Twenty-two healthy, right-handed German native speakers par-icipated in the study. Participants were recruited by means ofdvertisements. One participant was excluded because of technicalailure. Another one was excluded because of excessive movement.articipants with a minimum of seven artifact-free trials in the rele-ant experimental conditions were included in the analysis, so thathe final sample included fifteen subjects (eight females; age rangedrom 21 to 30 years, mean and standard deviation: 23.5 ± 2.2 years).he study was approved by the Ethics Committee of the Facultyf Medicine of the Ruhr University Bochum. All participants gavenformed written consent. Subjects received either course credit or5D upon completion of the task.
.2. Stimuli
Stimuli were 304 nameable color photographs (resolution:00 × 400 pixels) depicting common tools (120 pictures), animals120 pictures) or indoor and outdoor scenes (64 pictures). Thereparation of the stimuli database has been described elsewhere37].
Two hundred and fifty-six pictures were paired and were shownuring the study phases (see below). Pairs were divided in two sets.“same-category” set included object-object associations in which
he semantic category of the items was held constant (64, of which2 animal-animal and 32 tool–tool associations); a “different-ategory” set included object-scene associations, in which oneicture represented either an animal or a tool, and the otherepresented an indoor/outdoor scene (64 picture pairs, including
2 animal-scene and 32 tool-scene associations). There were nonimal–tool pairs. All pairs featured semantically and perceptu-lly unrelated images, as assessed through a piloting procedureith an independent sample of 12 participants [37]. Twenty-fourResearch 266 (2014) 63–76 65
pictures of animals and twenty-four pictures of tools served as“new” stimuli. They were only shown during the test phases of theexperiment (see below).
2.3. Recognition memory task
The experiment was divided into 8 blocks. Each block includeda study phase, a delay and a test phase. During the study phase, 16stimuli pairs per block appeared sequentially (i.e., one picture afterthe other) in the center of the screen. The experimental trial startedwith a white screen (500–1000 ms), followed by the text “nextpair” (1100–2000 ms). Afterwards, a black fixation cross appearedin the center of the screen (1000 ms), and then the first picture ofthe pair was shown (each picture was presented for 2000 ms). Afixation cross (1000 ms) separated the two pictures within a pair.Sample stimuli have been shown elsewhere [26]. Participants wereinstructed to encode the information for later recall testing andwere explicitly told that recall of associative information was themain goal of the task. Encoding instructions thus pointed at maxi-mizing recollection in the retrieval phase.
After a delay (∼70 s), during which participants rested, 16 stud-ied and 6 unstudied (i.e., “new”) single pictures per block werepresented. Only pictures of tools and animals (one per pair) wereshown in the test phase. Since tools and animals could be pre-sented either first or second during encoding, there were twice asmany trials for same-category relative to different-category pairsfor the order comparisons (because scenes were not analyzed). Thepictures were shown in the center of the screen for 2000 ms. Partic-ipants were instructed to refrain from responding during this time.Afterwards two black boxes with the texts (in capital letters) “old”and “new” appeared in the lower section of the screen, at the leftand the right, respectively. This screen was shown for 5000 ms, oruntil participant’s response. Participants responded with the righthand using the left (old) or right (new) arrow on a computer key-board. Failure to respond within the given time was followed bya message prompting participants to react faster. After a positiverecognition judgment (i.e., “Old” response), participants were askedto what category the associated picture belonged (this screen lasted4500 ms or until subjects’ response). Participants pressed the but-tons “1” (animal), “2” (tool), “3” (scene) on the computer keyboardto respond. Afterwards participants were prompted to recall theassociated picture and name the stimulus (deadline for responsewas 4000 ms), and answers were recorded using a portable voicerecorder.
Before starting the experiment, participants performed a train-ing block with different stimuli.
2.4. EEG recording
Subjects were seated approximately 70 cm in front of a com-puter monitor in a sound shielded room. Stimuli were presentedusing the software “Presentation” (http://www.neuro-bs.com).
EEG was recorded using 30 silver–silver chloride electrodesmounted on an elastic cap according to the International 10–20System at the following electrode sites: F7, F3, Fz, F4, F8, FT7, FC3,FCz, FC4, FT8, T7, C3, Cz, C4, T8, TP7, CP3, CPz, CP4, TP8, P7, P3, Pz,P4, P8, PO7, PO3, POz, PO4, PO8. All electrodes were referencedto the linked mastoids and re-referenced offline to the averagesignal, following previous literature on ERP elicited by associa-tive memory tasks [27,28,38–40]. EEG signal was amplified using
a Brain Products BrainAmp Standard amplifier (Brain Products,Munich, Germany) and the vision recorder software provided byBrain Products. Data have been sampled at 500 Hz with impedance<5 k�.
6 l Brain
2
waip
2
iefsttnpb
tt(srtw
2
atlnebrptTibbiw5
A(d
mFo
2
ldcdim2aB
6 G. Pergola et al. / Behavioura
.5. Data analysis
Only trials in which an animal or tool was shown and thatere correctly endorsed as old or new in the test phase entered
nalysis (i.e., no scenes included, no misses and false alarmsncluded). This restriction applied to both the study and testhase.
.5.1. Stimuli visual featuresFor the explorative analyses regarding retrieval effects it is crit-
cal to control for basic visual features of the stimuli used. Wextracted stimuli size, brightness and high spatial frequency poweror the 128 “old” images depicting animals and tools that werehown during the test phase of the experiment. The resolution ofhe images was constant, hence “size” was defined as the fraction ofhe image not occupied by the white background. To assess bright-ess we used the mean intensity of all pixels of the images. Therocedure to obtain high spatial frequency power estimates haseen described elsewhere [41].
We tested the estimates obtained for normality of the dis-ribution and then performed a multivariate ANOVA (becausehe variables considered are interdependent) with factors TYPEanimal, tool) and CATEGORY (same, different). Note that the pre-entation sequence within each pair during the study phase wasandomized across participants. This implies that basic visual fea-ures were randomized across the factor ORDER, which thereforeas not included in this analysis.
.5.2. Behavioral dataDescriptive analysis of recognition performance included hit
nd false alarm rates. Cued recall performance was estimated ashe correct associative recall rate, i.e., the number of hits fol-owed by correct recall of the paired associate over the totalumber of hits [26]. To ensure that participants had made anffort to remember at least the category to which the associateelonged, we performed a paired t-test including the categoryecall rate and the guess baseline. Category recall rate was com-uted as the percentage of correctly recalled categories over theotal of hits for which the associated pictures were not recalled.he guess baseline was computed as the total of recognizedtems not followed by correct recall of the associate multipliedy chance level. Participants chose between three categories,ut since there were no animal–tool pairs, the choice was lim-
ted, within each trial, to animal/tool (if an animal or a toolere shown, respectively) or scene, so chance performance was
0%.Further insight on task performance was sought by three-way
NOVAs on the hit-rates and cued recall rates with the factors TYPEanimal, tool), ORDER (the sequence of presentation within a pairuring encoding: first, second), and CATEGORY (same, different).
Medians of reaction times were analyzed by means of repeatedeasures ANOVA with the factor RESPONSE (hit, correct rejection).
inally, we computed three way ANOVAs (same factors as above)n reaction times to hits and complete recall trials.
.5.3. ERP analysisEEG data were analyzed off-line using the Brain Vision Ana-
yzer Software package (Brain Products, Munich, Germany). Rawata were inspected to mark bad recordings using the followingriteria: maximum allowed voltage step: 150 �V; maximal allowedifference of values within 200 ms: 600 �V; lowest allowed activ-
ty within 200 ms: 0.37 �V. Intervals matching the criteria were
arked as bad starting 200 ms before the artifact and ending00 ms after the artifact. These data were not included in the furthernalyses. A high- (0.5 Hz) and a low-pass filter (25 Hz) were applied.link artifacts were removed using an independent component
Research 266 (2014) 63–76
analysis procedure. Independent components were identified ashigh-energy components with a symmetric frontal positive topog-raphy [42] and were discarded in the back-transformation to therecording sites space. After the independent component analy-sis, data were then re-referenced to the average of all recordingsites.
Epochs were defined reflecting the 2 × 2 × 2 design of theexperiment (TYPE × ORDER × CATEGORY), starting 200 ms beforestimulus presentation and lasting for 1000 ms. Trial classificationwas performed using custom-written Matlab® (MathWorks: Nat-ick, MA, USA) codes.
Data were baseline corrected using the time window from−200 ms to 0 ms prior to stimulus onset. Automatic artifact rejec-tion excluded trials exceeding the following criteria: maximumallowed gradient: 50 �V/ms; maximal allowed difference within1200 ms: 200 �V; lowest allowed activity within 100 �V: 0.5 �V.Artifacts were marked as bad starting 200 ms prior to the artifactand ending 200 ms after the end of the artifact. Trials contain-ing artifacts were excluded from further analyses. Segments werethen averaged and the difference between waves reflecting hitsand waves reflecting correct rejections was computed. The analysisemployed difference waves because the study focused on differ-ences between hits. We were interested in the different magnitudeof old/new effects, rather than on old/new effects per se. Workingwith difference waves, old/new effects are simply obtained by test-ing the electrophysiological index of interest (e.g., mean amplitudeof the different wave in the given interval) against zero. We testedthe significance of old/new effects whenever we found a significantdifference between hits.
In the analysis of the study phase we were particularly inter-ested in the effect of CATEGORY, so we selected events in whichthe second item was shown and that resulted in successful recogni-tion during retrieval. When the first item is shown, the associationhas not yet been completed, therefore it would be meaninglessto refer to same- or different-category items. We considered theinterval 300–500 ms, the typical time window for the N400 com-ponent, because we expected that when the second item of thepair differed by category from the first item the waveform wouldbe more negative-going compared to the same-category associa-tions [35,36]. The effect of presentation order on ERP of associativeencoding has been analyzed in more details in an independentstudy [26].
During retrieval participants performed an old/new assessment.To identify memory-related processing, we compared hits withcorrect rejections. During retrieval, we considered for the anal-ysis the time windows highlighted by Jager, Mecklinger [9], i.e.,300–400 ms and 400–700 ms, adjusting them by visual inspection.In our experiment the effects occurred slightly earlier and there-fore we analyzed the time window 260–350 ms for frontal old/neweffects (consistent with previous literature [8,21,43]). The timewindow 350–600 ms was analyzed to find parietal effects duringretrieval [9,34]. For each condition we extracted and analyzed themean amplitude of the difference waves in the target time windowat the electrode sites used by Jager, Mecklinger [9] (F3, Fz, F4; C3,Cz, C4; P3, Pz, P4).
Additionally, we performed exploratory analyses on effects ear-lier than 200 ms. We computed individual participants’ t-test wavesof the old/new differences found in the time window 0–200 ms. Wethen averaged the t-test waveforms across participants to obtaingrand averages of the statistical effects. On these waveforms, weidentified a left frontal negative peak (hits more negative thancorrect rejections) in the t-waves with a latency of 56–60 ms anda midline parietal positive peak (hits more positive than correctrejections) with latencies varying from 152 to 180 ms. We there-
fore submitted mean amplitudes in the intervals 40–90 ms and150–200 ms to statistical analysis.
G. Pergola et al. / Behavioural Brain
Table 1Behavioral indices and results of recognition memory and recall performance. Ratesare computed over the total number of hits. Abbreviations: FA, false alarms; H+, hitsfollowed by successful recall of the uniquely paired associate; H0+, hits followedby successful recall of the category which the item paired with the recognition cuebelonged to; H0− , hits followed by no recall of either the item associated to therecognition cue, or its category.
Index Average performance Standard error
RecognitionHit-rate 0.967 0.006FA-rate 0.008 0.003D′ 4.7 0.19
Cued recall+
2
ummsto
riredlcwor
trCaA
3
3
bfbsm
tev[
o
3
b
p
H rate 0.48 0.05H0+ rate 0.28 0.02H0− rate 0.25 0.03
.5.4. Statistical analysesFor the analysis of the encoding phase data, mean amplitude val-
es extracted from the time windows of interest entered a repeatedeasures ANOVA with ROW (frontal, central, parietal), LINE (left,idline, right), TYPE (animal, tool; this factor refers to the type of
timulus shown on the screen), and CATEGORY (same, different;his factor refers to the type of the stimulus associated with thene shown on the screen).
In the retrieval phase of the current experiment pictures rep-esented either animals or tools (factor TYPE). Any differencesnvolving the factor TYPE could arise from semantic processingelated to the picture at hand, and therefore would not imply differ-nt memory processing related to animals and tools. However, theifference waves hits-correct rejections were computed within the
evels of the factor TYPE, i.e., we subtracted waveforms of animalsorrect rejections from waveforms of animals hits and the sameas done for tools. In the analysis of the resulting waves an effect
f TYPE implies larger or smaller old/new effects for animals withespect to tools.
ANOVAs (˛ = .05) on data collected during retrieval includedhe factors: LINE (frontal, central, parietal), ROW (left, midline,ight), TYPE (animal, tool), ORDER (shown first, shown second), andATEGORY (same, different). Greenhouse-Geißer corrections werepplied where appropriate. Interactions were resolved by multipleNOVAs or paired t-tests, where appropriate.
. Results
.1. Stimuli visual features
We found that the distribution of the values of the variablesrightness and high spatial frequency power was significantly dif-erent from the normal distribution [Kolmogorov–Smirnov test;rightness: P = .03; frequency: P = .011]. For this reason, we boot-trapped (10,000 resamplings) the coefficients of the general linearodel and computed significance estimates on these values.We found no significant effects of DOMAIN and TYPE in the mul-
ivariate analysis [F3,122 < 2.6, P > .05, partial �2 < .1]. Specifically, theffects of DOMAIN and TYPE on the single variables all yielded P-alues > .1 except for a non-significant trend of TYPE on brightnessbootstrap: P = .093].
We found no significant evidence that could support an effectf basic visual features on the ERP findings reported.
.2. Behavioral data
Table 1 summarizes the descriptive statistics computed onehavioral performances in the recognition and the cued recall task.
Data show that recognition was close to ceiling, but only twoarticipants reached ceiling. Deep encoding of the associations
Research 266 (2014) 63–76 67
was successful, as shown by the cued recall rate (Table 1: noticethat about 76% of hits were followed by recall of the paired item,or at least the category it belonged to). Even when subjects didnot recall the item, category recall occurred significantly morefrequently than it could be expected by chance [guess baselinewas 0.5; mean ± standard error of the mean (s.e.m.): 0.56 ± 0.04;|t14| = 3.3, P = .005].
A three-way ANOVA on hit-rates with factors TYPE (animal,tool), ORDER (shown first, shown second), and CATEGORY (same,different) yielded a significant main effect of TYPE [F1,14 = 11.8,P = .004, partial �2 = .46]: tools were recognized more often thananimals (mean ± s.e.m. for tools: 0.963 ± 0.007; mean ± s.e.m. foranimals: 0.93 ± 0.01). No other effects were significant [P > .05].
Fig. 1 illustrates the results of the ANOVA on cued recallrates, which yielded a significant TYPE × CATEGORY interaction[F1,14 = 5.2, P = .04, partial �2 = .27], as well as significant main effectsof ORDER [F1,14 = 5.5, P = .034, partial �2 = .28], TYPE [F1,14 = 7.5,P = .016, partial �2 = .35], and CATEGORY [F1,14 = 5.5, P = .034, par-tial �2 = .28]. The ORDER main effect consisted of higher recall rateswhen the cue shown during retrieval had appeared second dur-ing encoding. The main effect of TYPE was similar to the effectfound on recognition, with tools more often followed by success-ful recall than animals. The main effect of CATEGORY consistedof higher recall rates for different-category than same-categoryitems. After averaging across the ORDER factor, resolution of theTYPE × CATEGORY interaction revealed that the CATEGORY maineffect was driven by pictures of animals, which elicited a sig-nificantly higher recall performance for different-category pairs[|t14| = 2.7, P = 0.016]. This was not the case of tools [|t14| = 0.75,P = .47].
The analysis of reaction times is reported in the supplementarymaterial. In short, responses to hits were slower than responses tocorrect rejections; responses to tools were faster than responses toanimals.
3.3. ERP data
The following paragraphs report the outcome of the ERP anal-ysis during encoding and retrieval; the analysis of effects duringretrieval is further split in planned analysis and exploratory analy-sis. Interactions only involving the factors LINE and ROW, referringto electrode position, are not reported. Interactions involving thefactor LINE, but not the factor ROW, are further analyzed in thesupplementary material, because the main text focuses on antero-posterior localization of the effects. Effects with P < .05 are reported.See the supplementary material for additional figures representingthe waveforms separated by ORDER, TYPE, and CATEGORY.
3.3.1. EncodingThe analysis on data collected during encoding involved images
that were shown second within a pair and were subsequentlyrecognized as old. Mean amplitudes were analyzed for thetime window 300–500 ms, reflecting the time window to assessN400 commonly elicited by congruency assessment of subse-quent stimuli [35]. Fig. 2 shows sample waveforms of the effectsdescribed.
Repeated measures ANOVA with factors ROW (frontal, cen-tral, parietal), LINE (left, midline, right), TYPE (animal, tool),and CATEGORY (same, different) yielded a significant maineffect of CATEGORY [F1,14 = 16, P = .001, partial �2 = .54] and thesignificant interactions: ROW × TYPE [F1.2,17 = 17, P < .001, par-tial �2 = .55]; LINE × TYPE [F1.5,21 = 11, P < .008, partial �2 = .33];
ROW × CATEGORY [F1.5,21 = 5.8, P < .016, partial �2 = .29].The main effect of CATEGORY consisted of overall more nega-tive waveforms for different-category pairs (mean ± s.e.m. same:−.69 ± .22 �V; different: −1.2 ± .24 �V).

68 G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76
Fig. 1. Behavioral results. Recall performance was computed as the successful recall rate on the number of hits. Bars represent 95% confidence intervals for the mean (onlyupward bars represented). Tools were recalled more often than animals and recognition cues shown second within a pair during the study phase yielded higher recallr y assos atego
c
tMmGR
[P
Fsah
ates. An effect of category was only significant for animals, with different-categorame-category animal picture; A B, different-category animal picture; T W, same-c
Hence, there was an overall larger N400 for different-categoryompared to same-category pairs.
In order to analyze the two interactions involving the ROW fac-or, data were pooled across the factor LINE. See the Supplementary
aterial for the interaction involving the LINE factor. Repeatedeasures ANOVAs with factors TYPE (animal, tools), and CATE-ORY (same, different) were performed for the three levels of the
OW factor.Pooled frontal sites yielded significant main effects of TYPEF1,14 = 8.3, P = .012, partial �2 = .37] and CATEGORY [F1,14 = 9.8,= .007, partial �2 = .41]. The main effect of TYPE consisted of
ig. 2. Effects of category during encoding at midline frontal, central and parietalites. The waveforms represent the electrophysiological activity elicited by toolsnd animals shown second in same- and different-category pairs. The time windowighlighted yields a significant effect of category.
ciations eliciting higher recall rates than same-category ones. Abbreviations: A W,ry tool picture; T B, different-category tool picture.
more negative waveforms elicited by animals (mean ± s.e.m. tools:−3.1 ± .5 �V; animals: −3.6 ± .5 �V). The main effect of CATE-GORY reflected more negative potentials for different-categoryitems (mean ± s.e.m. same: −3.0 ± .5 �V; different: −3.7 ± .5 �V).At central sites a significant main effect of CATEGORY was alsofound [F1,14 = 7.1, P = .003, partial �2 = .48], with the same direc-tion as at frontal sites (mean ± s.e.m. same: −1.3 ± .3 �V; different:−2.0 ± .4 �V). At parietal sites we found a significant main effect ofTYPE [F1,14 = 24, P < .001, partial �2 = .63], with more positive poten-tials elicited by animals (mean ± s.e.m. tools: 1.8 ± .3 �V; animals:2.7 ± .3 �V). In other words, CATEGORY affected ERPs with a fronto-central topography, with higher amplitudes of the N400 for objectsfollowing a scene (different-category items) compared to objectsfollowing objects of the same category. TYPE effects showed morenegative frontal ERP for animals compared to tools and the reverseat parietal electrodes.
To summarize, ERP of associative encoding differed between 300and 500 ms post-stimulus presentation as a function of the categoryboth of the item at hand and of the item previously shown (the firstmember of the pair).
3.3.2. Retrieval: planned analyses3.3.2.1. 260–350 ms. Fig. 3 illustrates the effects found in the timewindows: 260–350 ms and 350–600 ms. Figs. 4–7 show the differ-ence waveforms between hits and correct rejections during the testphase of the memory task in all experimental conditions.
Repeated measures ANOVA included the mean amplitudesof the hit-correct rejections difference waves with factors ROW(frontal, central, parietal), LINE (left, midline, right), TYPE (animal,tool), ORDER (first, second), and CATEGORY (same, different). Thefollowing interactions were significant: ROW × TYPE [F1.4,20 = 6.4,P = .013, partial �2 = .32]; LINE × TYPE [F1.8,25 = 3.9, P = .037, par-
tial �2 = .22]; LINE × ORDER [F1.6,22 = 5.6, P = .015, partial �2 = .29];ROW × ORDER × CATEGORY [F1.7,24 = 4.4, P = .046, partial �2 = .24].To resolve the interactions involving the factor ROW we pooleddata across the factor LINE. Separate ANOVAs for each level of the

G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76 69
Fig. 3. Waveforms at midline frontal and parietal electrodes during retrieval. Shaded areas highlight the effects discussed in the text. The x-axis reports time in ms and they-axis reports potential in �V.
Fig. 4. Difference waveforms at all electrode sites for animal pictures shown first. The figures illustrate the CATEGORY effect. The waves represent the difference betweenhits and correct rejections in the given condition. The time scale starts at 200 ms pre-stimulus and ends at 800 ms post-stimulus. Positivity points downwards.

70 G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76
F . Theh e-stim
fn
splwesPei−cfsiEofi
ig. 5. Difference waveforms at all electrode sites for animal pictures shown secondits and correct rejections in the given condition. The time scale starts at 200 ms pr
actor ROW yielded significant effects at frontal and parietal, butot central sites [all F < 3, all P > .05].
At frontal sites, the interaction ORDER × CATEGORY wasignificant [F1,14 = 4.7, P = .047, partial �2 = .25]. Paired t-tests com-aring same- and different-category items separately per each
evel of ORDER revealed that, only for pictures shown firstithin a pair, old/new effects significantly dissociated the cat-
gory of the associations [|t14| = 2.6, P = .022]; no effect wasignificant for items shown second within a pair [|t14| = .76,= .46]. Different-category associations elicited larger old/newffects (mean ± s.e.m. of the difference for different-categorytems shown first: .62 ± .2 �V; same-category items shown first:.091 ± .2 �V). Next we asked if the old/new effects were signifi-
ant by computing a t-test against zero. The result was significantor different-category items [|t14| = −2.6, P = .022] but not forame-category items [|t14| < 2, P > 05]. These results show that
n this time window the CATEGORY factor modulated frontalRP only for items shown first within a pair. A significantld/new effect was only found for different-category cues shownrst.figures illustrate the CATEGORY effect. The waves represent the difference betweenulus and ends at 800 ms post-stimulus. Positivity points downwards.
At parietal sites a significant main effect of TYPE [F1,14 = 6.3,P = .025, partial �2 = .31] consisted of more positive ERP for hits thanfor correct rejections in the case of animals (mean ± s.e.m. of thedifference for tools: −.20 ± .2 �V; animals: .40 ± .2 �V). Old/neweffects, however, were not significant (animals [|t14| = 2.1, P = .054];tools [|t14| = .20, P = .30]).
In summary, in the 260–350 ms time window old/new effectsdiffered at frontal sites as a function of order and category. Onlyfor items shown first did ERP show a significant old/new effectfor different-category items. At parietal sites ERP discriminatedold/new effects for animals from old/new effects for tools. Atright sites old/new effects differed between items shown first andsecond, without significant old/new effects (see SupplementaryMaterial).
3.3.2.2. 350–600 ms. Repeated measures ANOVA on the mean
amplitudes of the hit-correct rejections difference waves involvedthe factors ROW (frontal, central, parietal), LINE (left, midline,right), TYPE (animal, tool), ORDER (shown first, shown second),and CATEGORY (same, different). The interactions ROW × TYPE
G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76 71
F s illusc us and
[P[
Cft��io.rlts.P
P
ig. 6. Difference waveforms at all electrode sites for tools shown first. The figureorrect rejections in the given condition. The time scale starts at 200 ms pre-stimul
F1.3,18 = 4.2, P = .048, partial �2 = .23], LINE × ORDER [F2,27 = 11,< .001, partial �2 = .44], and ROW × LINE × ORDER × TYPE
F2.8,39 = 3.3, P = .033, partial �2 = .19] were significant.To resolve the interactions, we pooled data across the factor
ATEGORY, and analyzed separately the three levels of the ROWactor. No significant effects were found at frontal sites. At cen-ral sites, interactions LINE × ORDER [F1.9,27 = 6.7, P = .005, partial2 = .33] and a main effect of ORDER [F1,14 = 5.6, P = .043, partial2 = .26] were significant. Recognition cues shown second dur-
ng encoding elicited more positive old/new effects (mean ± s.e.m.f the difference for pictures shown first: .50 ± .15 �V; second:
25 ± .13 �V). Post hoc tests revealed that the effect was driven byight-sided potentials [|t14| = 4.4, P = .001] and was not significant ateft and midline sites [|t14| < 2, P > .05]. At central right sites (elec-rode C4) the old/new effect was significant for recognition cueshown second during encoding [|t14| = 2.4, P = .029; mean ± s.e.m.:
51 ± .21 �V] and not for recognition cues shown first [|t14| < 2,> .05; mean ± s.e.m.: .016 ± .19 �V].Parietal ERPs revealed a significant TYPE main effect [F1,14 = 12,= .004, partial �2 = .46] as well as significant LINE × ORDER
trate the CATEGORY effect. The waves represent the difference between hits andends at 800 ms post-stimulus. Positivity points downwards.
[F1.5,22 = 4.3, P = .035, partial �2 = .24] and LINE × ORDER × TYPE[F1.7,24 = 4.2, P = .032, partial �2 = .23] interactions. Animals elicitedlarger old/new effects compared to tools (mean ± s.e.m. of thedifference for animals: 1.0 ± .21 �V; second: .24 ± .17 �V). At leftparietal sites we found a significant main effect of TYPE [F1,14 = 14,P = .002, partial �2 = .49] consistent with the general main effect(mean ± s.e.m. of the difference for animals: 1.1 ± .17 �V; tools:.53 ± .21 �V). Notably, the left parietal old/new effects was sig-nificant for both animals [|t14| = 6.4, P < .001] and tools [|t14| = 2.5,P = .023].
The same main effect of TYPE was also found at midline sites[F1,14 = 9.4, P = .008, partial �2 = .40; mean ± s.e.m. of the differ-ence for animals: 1.3 ± .20 �V; tools: .21 ± .29 �V], together witha significant ORDER × TYPE interaction [F1,14 = 6.5, P = .023, partial�2 = .32]. Analysis of this interaction indicated that the differencebetween animals and tools was significant in the case of recognition
cues shown first during encoding [|t14| = 3.7, P = .002] and not forcues shown second [|t14| = 1.6, P = .14]. Regardless of ORDER, mid-line parietal old/new effects were significant for animals [shownfirst: |t14| = 7.0, P < .001; shown second: |t14| = 5.6, P < .001], but not
72 G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76
F res illuc us and
fP
[oo.P
etwOwe
3
et
ig. 7. Difference waveforms at all electrode sites for tools shown second. The figuorrect rejections in the given condition. The time scale starts at 200 ms pre-stimul
or tools [shown first: |t14| = −.38, P = .71; shown second: |t14| = 2.0,= .066].
Right parietal ERPs yielded a significant main effect of ORDERF1,14 = 5.5, P = .034, partial �2 = .28]. Recognition cues shown sec-nd during encoding showed larger old/new effects (mean ± s.e.m.f the difference for pictures shown first: .17 ± .32 �V; second:
50 ± .30 �V), which however failed to reach significance [|t14| < 2,> .05].
In summary, a centro-parietal positivity characterized old/newffects, consistently with the expectation on the late parietal posi-ivity. Left parietal ERPs discriminated hits from correct rejections,ith larger differences elicited by images of animals. The factorRDER modulated old/new effects at right centro-parietal sites,ith maximal amplitude for recognition cues shown second during
ncoding. No effect of CATEGORY was found.
.3.3. Retrieval: exploratory analysesLike for planned analyses, the mean amplitudes of differ-
nce waves hits-correct rejection were extracted in the targetime intervals (40–90 ms and 150–200 ms) and entered repeated
strate the CATEGORY effect. The waves represent the difference between hits andends at 800 ms post-stimulus. Positivity points downwards.
measures ANOVA with factors ROW (frontal, central, parietal),LINE (left, midline, right), TYPE (animal, tool), ORDER (first, sec-ond), and CATEGORY (same, different). Fig. 4 illustrates samplewaveforms.
3.3.3.1. 40–90 ms. The ANOVA yielded a significant main effectof ORDER [F1,14 = 5.6, P = .033, partial �2 = .28] and the significantinteractions: ROW × TYPE [F1.1,16 = 4.7, P = .043, partial �2 = .25];ROW × ORDER × CATEGORY [F1.1,15 = 7.3, P = .014, partial �2 = .34];LINE × ORDER × CATEGORY [F1.6,22 = 4.4, P = .032, partial �2 = .24].
The main effect of ORDER consisted of more negative wave-forms for hits of items shown first compared to items shownsecond (mean ± s.e.m. of the difference wave for shown first:−.14 ± .08 �V; shown second: .01 ± .09 �V). The old/new effect perse was not significant [first-shown: |t14| = −1.8, P = .091 second-shown: |t14| < 2, P > .05].
To resolve the interactions involving the ROW factor data werepooled across the LINE factor. Repeated measures ANOVA com-puted separately per each level of the factor ROW included factorsORDER, TYPE, and CATEGORY.

G. Pergola et al. / Behavioural Brain Research 266 (2014) 63–76 73
F ring rt
waEtPci.ofc
cfi(os
Ppoo
dtcle
3RpTsaw�tfoPwpw
ig. 8. Enlarged view of the waveforms at midline frontal and parietal electrodes duime in ms and the y-axis reports potential in �V.
At frontal sites, a significant ORDER × CATEGORY interactionas found [F1,14 = 8.1, P = .013, partial �2 = .37]. After pooling
cross the factor TYPE, we found a marginally significant CAT-GORY effect for items shown first [|t14| = 1.9, P = .078], whilehe difference was significant for items shown second [|t14| = 2.2,= .048]. The effect consisted of more negative waveforms for hitsompared to correct rejections in the case of different-categorytems (mean ± s.e.m. of the difference wave for same-category:09 ± .2 �V; different-category: −.43 ± .2 �V). For items shown sec-nd, hits significantly differed from correct rejections selectivelyor different-category items [|t14| = 2.5, P = .024], and not for same-ategory items [|t14| = .39, P = .70].
At central sites a main effect of ORDER was the only signifi-ant finding [F1,14 = 4.9, P = .04, partial �2 = .26], with items shownrst eliciting more negative waveforms than items shown secondmean ± s.e.m. of the difference wave for first shown: −.17 ± .1; sec-nd shown: .02 ± .1). The old/new effects per se, however, were notignificant [both |t14| < 2, P > .05].
At parietal sites a main effect of TYPE was significant [F1,14 = 5.2,= .039, partial �2 = .27]. Hits to animals elicited significantly moreositive waveforms compared to correct rejections (mean ± s.e.m.f the difference wave for tools: −.19 ± .2 �V; animals: .47 ± .2 �V;ld new effect for animals: |t14| = 2.2, P = .042).
In summary, between 40 and 90 ms post-stimulus frontal ERPiscriminated different-category from same-category items selec-ively in the case of items shown second. The order of presentationould also be dissociated based on ERP at central sites. The stimu-us type modulated ERP at parietal sites, with a significant old/newffect only found for animals (Fig. 8).
.3.3.2. 150–200 ms. The ANOVA yielded a significantOW × ORDER × CATEGORY interaction [F1.4,20 = 6.2, P = .014,artial �2 = .31]. After pooling data across the factors LINE andYPE, we computed ANOVAs with factors ORDER and CATEGORYeparately per each level of ROW. At frontal sites no effectspproached significance [F1,14 < 3, P > .05]. At central sites thereas a significant main effect of ORDER [F1,14 = 11, P = .006, partial
2 = .43], with hits more positive going for items shown secondhan for those shown first (mean ± s.e.m. of the difference waveor first-shown: −.06 ± .1 �V; second-shown: .31 ± .2 �V). Theld/new effect was reliable only for items shown second [|t14| = 2.3,
= .036]. At parietal sites the interaction ORDER × CATEGORYas significant [F1,14 = 6.5, P = .023, partial �2 = .32]. Follow-upaired t-tests revealed that only for items shown first thereas a significant effect of CATEGORY [|t14| = 2.2, P = .042].etrieval. Shaded areas highlight the effects discussed in the text. The x-axis reports
Different-category items yielded more negative potentials com-pared to same-category items (mean ± s.e.m. of the difference wavefor same-category: −.16 ± .1 �V; different-category: −.21 ± .2 �V).The effect was not significant for items shown second [|t14| = −.92,P = .38]. No significant old/new effect was found [both |t14| < 2,P > .05].
To summarize, between 150 and 200 ms post-stimulus centraland parietal ERP dissociated hits as a function of the order of pre-sentation and the category of the association. In particular, themagnitude of old/new effects differed at parietal sites selectivelyfor items shown first within a pair.
Fig. 5 illustrates a synopsis of the effects detected on pooledfrontal and pooled parietal sites earlier than 350 ms in all differentexperimental conditions, with the aim to summarize the findingspossibly related to priming effects. The qualitative pattern is verysimilar in all time windows considered (40–90 ms; 150–200 ms;260–350 ms). This implies that the relationship between condi-tions in terms of old/new effects did not change dramatically inthe interval 40–350 ms. Note, for instance, that frontal ERPs ofdifferent-category items were regularly more positive-going thanERPs of same-category items when recognition cues were shownfirst during encoding, but not when they were shown second(Fig. 5). Frontal ERPs discriminated different-category hits fromcorrect rejections in the 40–90 ms time window for items shownsecond and in the 260–350 ms time window for items shown first(highlighted with rectangles in Fig. 9). At parietal sites, old/neweffects elicited by animals were regularly more positive-going com-pared to those elicited by tools (notice the pointy pattern in thealternation animal/tool).
4. Discussion
The aim of this study was testing hypotheses on the func-tional role of the FN400 in the context recognition memory byinvestigating the effect of presentation sequence on the FN400. Inkeeping with results obtained by modulating the perceptual asso-ciation between faces, we found that a frontal old-new effect inthe time window 260–350 ms was only significant for items pre-sented first within a pair. Interestingly, in the current experimentdifferent-category associations, and not same-category associa-
tions, showed significant old/new effects. The parietal old/neweffect, thought to reflect recall/recollection, was modulated bystimulus type and order of presentation, but not by semanticcategory.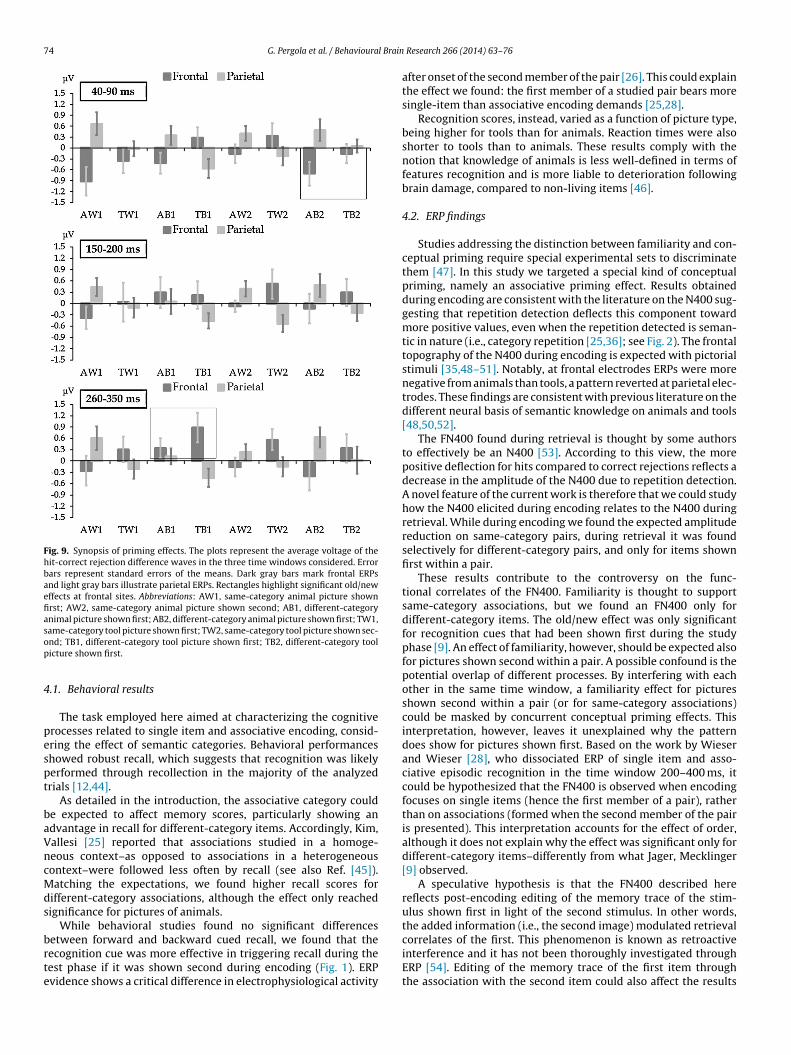
74 G. Pergola et al. / Behavioural Brain
Fig. 9. Synopsis of priming effects. The plots represent the average voltage of thehit-correct rejection difference waves in the three time windows considered. Errorbars represent standard errors of the means. Dark gray bars mark frontal ERPsand light gray bars illustrate parietal ERPs. Rectangles highlight significant old/neweffects at frontal sites. Abbreviations: AW1, same-category animal picture shownfirst; AW2, same-category animal picture shown second; AB1, different-categoryanimal picture shown first; AB2, different-category animal picture shown first; TW1,same-category tool picture shown first; TW2, same-category tool picture shown sec-op
4
pespt
baVncMds
brte
nd; TB1, different-category tool picture shown first; TB2, different-category toolicture shown first.
.1. Behavioral results
The task employed here aimed at characterizing the cognitiverocesses related to single item and associative encoding, consid-ring the effect of semantic categories. Behavioral performanceshowed robust recall, which suggests that recognition was likelyerformed through recollection in the majority of the analyzedrials [12,44].
As detailed in the introduction, the associative category coulde expected to affect memory scores, particularly showing andvantage in recall for different-category items. Accordingly, Kim,allesi [25] reported that associations studied in a homoge-eous context–as opposed to associations in a heterogeneousontext–were followed less often by recall (see also Ref. [45]).atching the expectations, we found higher recall scores for
ifferent-category associations, although the effect only reachedignificance for pictures of animals.
While behavioral studies found no significant differences
etween forward and backward cued recall, we found that theecognition cue was more effective in triggering recall during theest phase if it was shown second during encoding (Fig. 1). ERPvidence shows a critical difference in electrophysiological activityResearch 266 (2014) 63–76
after onset of the second member of the pair [26]. This could explainthe effect we found: the first member of a studied pair bears moresingle-item than associative encoding demands [25,28].
Recognition scores, instead, varied as a function of picture type,being higher for tools than for animals. Reaction times were alsoshorter to tools than to animals. These results comply with thenotion that knowledge of animals is less well-defined in terms offeatures recognition and is more liable to deterioration followingbrain damage, compared to non-living items [46].
4.2. ERP findings
Studies addressing the distinction between familiarity and con-ceptual priming require special experimental sets to discriminatethem [47]. In this study we targeted a special kind of conceptualpriming, namely an associative priming effect. Results obtainedduring encoding are consistent with the literature on the N400 sug-gesting that repetition detection deflects this component towardmore positive values, even when the repetition detected is seman-tic in nature (i.e., category repetition [25,36]; see Fig. 2). The frontaltopography of the N400 during encoding is expected with pictorialstimuli [35,48–51]. Notably, at frontal electrodes ERPs were morenegative from animals than tools, a pattern reverted at parietal elec-trodes. These findings are consistent with previous literature on thedifferent neural basis of semantic knowledge on animals and tools[48,50,52].
The FN400 found during retrieval is thought by some authorsto effectively be an N400 [53]. According to this view, the morepositive deflection for hits compared to correct rejections reflects adecrease in the amplitude of the N400 due to repetition detection.A novel feature of the current work is therefore that we could studyhow the N400 elicited during encoding relates to the N400 duringretrieval. While during encoding we found the expected amplitudereduction on same-category pairs, during retrieval it was foundselectively for different-category pairs, and only for items shownfirst within a pair.
These results contribute to the controversy on the func-tional correlates of the FN400. Familiarity is thought to supportsame-category associations, but we found an FN400 only fordifferent-category items. The old/new effect was only significantfor recognition cues that had been shown first during the studyphase [9]. An effect of familiarity, however, should be expected alsofor pictures shown second within a pair. A possible confound is thepotential overlap of different processes. By interfering with eachother in the same time window, a familiarity effect for picturesshown second within a pair (or for same-category associations)could be masked by concurrent conceptual priming effects. Thisinterpretation, however, leaves it unexplained why the patterndoes show for pictures shown first. Based on the work by Wieserand Wieser [28], who dissociated ERP of single item and asso-ciative episodic recognition in the time window 200–400 ms, itcould be hypothesized that the FN400 is observed when encodingfocuses on single items (hence the first member of a pair), ratherthan on associations (formed when the second member of the pairis presented). This interpretation accounts for the effect of order,although it does not explain why the effect was significant only fordifferent-category items–differently from what Jager, Mecklinger[9] observed.
A speculative hypothesis is that the FN400 described herereflects post-encoding editing of the memory trace of the stim-ulus shown first in light of the second stimulus. In other words,the added information (i.e., the second image) modulated retrieval
correlates of the first. This phenomenon is known as retroactiveinterference and it has not been thoroughly investigated throughERP [54]. Editing of the memory trace of the first item throughthe association with the second item could also affect the results
l Brain
oliecdeidrtc
dgn(tbceewts
datamtifwccdnocd
e4papcftvcioIotth4a
cfa
[
[
[
[
[
[
G. Pergola et al. / Behavioura
btained by Jager, Mecklinger [9]. Since pictures shown first wereess effective recognition cues compared to pictures shown secondn the current study, there is no evidence that the effect fostersxplicit association, i.e., recollection, thereby supporting the con-eptual priming account of the FN400. As regards the fact that onlyifferent-category recognition cues yielded significant old/newffects, we tentatively propose that the N400 effect observed dur-ng encoding of same-category items, but not during encoding ofifferent-category items, could modulate the FN400 found duringetrieval, therefore resulting in absent FN400 effects during theest phase. With respect to this working hypothesis, however, theurrent results remain preliminary.
On the other hand, later potentials between 350 and 600 msid not discriminate different categories of association. This sug-ests that the recollection component of recognition memory wasot affected by the manipulation of this experimental conditionconsistent with Ref. [44]), although this evidence is at odds withhe finding that different-category items were followed more ofteny successful recall. It is possible that a greater statistical powerould detect more subtle differences with respect to the late pari-tal old/new effect. The finding that recognition cues shown secondlicited larger putative correlates of recollection [34] is consistentith their greater associative load. In this respect, it is important
o stress the sequential presentation of the images in the currenttudy, which differs from other experimental approaches [33,55].
Given the absence of priming effects for items shown seconduring the study phase, we searched for earlier effects of categorynd order. ERPs in the 150–200 ms time window with a parietalopography are thought to index perceptual categorization [56]nd perceptual priming [40,57]. Repeated items are expected to beore positive-going than novel items [21,58]. Current results show
hat in the 150–200 ms time window the putative electrophysiolog-cal correlates of priming were larger for items shown second thanor items shown first; within recognition cues shown first, theyere more positive-going for same-category than for different-
ategory items. The lack of an old/new effect could relate to theonceptual demands of the task: Voss, Schendan [57] selectivelyetected a significant P170 repetition effect in a perceptual, andot in a conceptual, memory task. Nevertheless, the presentationf the second member of the pair modulated this early component,ausing a more negative deflection at parietal sites selectively forifferent-category items.
Repetition effects have also been reported in the literature asarly as 50 ms after stimulus onset [59], consistently with the0–90 ms time window reported in the current study. Such earlyotentials have been typically interpreted as the result of thectivity of visual areas and have been assumed to reflect visualrocessing of the stimuli [39,60]. However, we found no signifi-ant effects of CATEGORY or TYPE in the analysis of basic visualeatures of the stimuli we used. Activity in the visual cortex is alsohought to support priming [43]. The 40–90 ms time window isery early, yet the fact that the old/new effects in this time windowharacterized recognition cues shown second during encoding isnteresting because it offers complementary evidence to the findingf an FN400 for items shown first in the 260–350 ms time window.t seems that the items shown second present earlier correlatesf priming compared to items shown first. The homogeneous pat-ern of effects found in the 40–350 ms time window (Fig. 9) suggesthat priming effects might take place in this “wider” time window,ence including the FN400, with a differentiation emerging in the0–90 ms and the 260–350 ms windows between items shown firstnd second during encoding, that yield different associative load.
Caution in such a conclusion is invited by the need to repli-ate the findings with robust testing to exclude the possibility ofalse positives. It is especially important to separate recognition tri-ls followed by successful recall from trials not followed by recall
[
Research 266 (2014) 63–76 75
[26] to ascertain how explicit associative memory affects the ERPcurrently reported. Formal testing of priming effects could addi-tionally improve the operational definition of the processes that weproposed to underlie the electrophysiological effects detected. Theearly associative effects described here for the first time, nonethe-less, suggest that associative memory studies need to take intoaccount the presentation sequence whenever pairs are presentedsequentially, and yield promise for future experimental scrutiny.
4.3. Conclusions
We showed FN400 effects depending on the order of presen-tation in a task featuring sequential presentation of the items. Inaccord with previous literature we only found an old/new effect inthe time window 260–350 ms for items shown first within a pairduring encoding. Our working hypothesis for future investigationis that the FN400 found here was a correlate of retroactive editingof the memory trace. Earlier effects could be a correlate of primingfor items shown second within a pair. We conclude that currentresults more strongly support the conceptual priming account ofthe FN400, than the familiarity account.
Funding
This work was supported by the NovoBrain Programme [MarieCurie Actions FP6, Early Stage Training]. GP has been also partly sup-ported by the “Foodcast” grant awarded to Prof. Raffaella Rumiatiand by the “Fondazione Con Il Sud” grant awarded to Prof. Alessan-dro Bertolino.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.bbr.2014.02.050.
References
[1] Tulving E. Episodic memory: from mind to brain. Annu Rev Psychol2002;53:1–25.
[2] Greenberg DL, Verfaellie M. Interdependence of episodic and semantic mem-ory: evidence from neuropsychology. J Int Neuropsychol Soc 2010;16:748–53.
[3] Eichenbaum H, Yonelinas AP, Ranganath C. The medial temporal lobe and recog-nition memory. Annu Rev Neurosci 2007;30:123–52.
[4] Squire LR, Wixted JT, Clark RE. Recognition memory and the medial temporallobe: a new perspective. Nat Rev Neurosci 2007;8:872–83.
[5] Schacter DL. Implicit expressions of memory in organic amnesia: learning ofnew facts and associations. Hum Neurobiol 1987;6:107–18.
[6] Gabrieli JD. Cognitive neuroscience of human memory. Annu Rev Psychol1998;49:87–115.
[7] Schacter DL, Buckner RL. Priming and the brain. Neuron 1998;20:185–95.[8] Voss JL, Paller KA. Fluent conceptual processing and explicit memory for faces
are electrophysiologically distinct. J Neurosci 2006;26:926–33.[9] Jager T, Mecklinger A, Kipp KH. Intra- and inter-item associations doubly disso-
ciate the electrophysiological correlates of familiarity and recollection. Neuron2006;52:535–45.
10] Ranganath C. A unified framework for the functional organization of the medialtemporal lobes and the phenomenology of episodic memory. Hippocampus2010;20:1263–90.
11] Brown MW, Warburton EC, Aggleton JP. Recognition memory: material, pro-cesses, and substrates. Hippocampus 2010;20:1228–44.
12] Montaldi D, Mayes AR. The role of recollection and familiarity in the functionaldifferentiation of the medial temporal lobes. Hippocampus 2010;20:1291–314.
13] Slotnick SD. The nature of recollection in behavior and the brain. Neuroreport2013;24:663–70.
14] Wixted JT, Mickes L, Squire LR. Measuring recollection and familiarity in themedial temporal lobe. Hippocampus 2010;20:1195–205.
15] Aggleton JP, O’Mara SM, Vann SD, Wright NF, Tsanov M, Erichsen JT.
Hippocampal-anterior thalamic pathways for memory: uncovering a networkof direct and indirect actions. Eur J Neurosci 2010;31:2292–307.16] Rugg MD, Vilberg KL, Mattson JT, Yu SS, Johnson JD, Suzuki M. Item mem-ory, context memory and the hippocampus: fMRI evidence. Neuropsychologia2012;50:3070–9.

7 l Brain
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
6 G. Pergola et al. / Behavioura
17] Brown MW, Barker GR, Aggleton JP, Warburton EC. What pharmacologicalinterventions indicate concerning the role of the perirhinal cortex in recog-nition memory. Neuropsychologia 2012;50:3122–40.
18] Pergola G, Suchan B. Associative learning beyond the medial temporal lobe:many actors on the memory stage. Front Behav Neurosci 2013;7:162.
19] Yonelinas AP, Aly M, Wang WC, Koen JD. Recollection and familiarity:examining controversial assumptions and new directions. Hippocampus2010;20:1178–94.
20] Mayes A, Montaldi D, Migo E. Associative memory and the medial temporallobes. Trends Cogn Sci 2007;11:126–35.
21] Schendan HE, Kutas M. Time course of processes and representations suppor-ting visual object identification and memory. J Cogn Neurosci 2003;15:111–35.
22] Yovel G, Paller KA. The neural basis of the butcher-on-the-bus phe-nomenon: when a face seems familiar but is not remembered. Neuroimage2004;21:789–800.
23] Wang WC, Yonelinas AP. Familiarity is related to conceptual implicit memory:an examination of individual differences. Psychon Bull Rev 2012;19:1154–64.
24] Paller KA, Lucas HD, Voss JL. Assuming too much from ‘familiar’ brain potentials.Trends Cogn Sci 2012;16:313–5, discussion 5–6.
25] Kim AS, Vallesi A, Picton TW, Tulving E. Cognitive association formation inepisodic memory: evidence from event-related potentials. Neuropsychologia2009;47:3162–73.
26] Pergola G, Trotta M, Suchan B. Asymmetric hemispheric contribution to ERPs inassociative memory indexes goal relevance and quantity of information. BehavBrain Res 2013;241:7–16.
27] Mangels JA, Picton TW, Craik FI. Attention and successful episodic encoding: anevent-related potential study. Brain Res Cogn Brain Res 2001;11:77–95.
28] Wieser S, Wieser HG. Event-related brain potentials in memory: cor-relates of episodic, semantic and implicit memory. Clin Neurophysiol2003;114:1144–52.
29] Caplan JB, Glaholt MG, McIntosh AR. EEG activity underlying successful studyof associative and order information. J Cogn Neurosci 2009;21:1346–64.
30] Kahana MJ. The cognitive correlates of human brain oscillations. J Neurosci2006;26:1669–72.
31] Yang J, Zhao P, Zhu Z, Mecklinger A, Fang Z, Li H. Memory asymmetry of forwardand backward associations in recognition tasks. J Exp Psychol Learn Mem Cogn2013;39:253–69.
32] Wiegand I, Bader R, Mecklinger A. Multiple ways to the prior occurrence of anevent: an electrophysiological dissociation of experimental and conceptuallydriven familiarity in recognition memory. Brain Res 2010;1360:106–18.
33] Yang J, Meckingler A, Xu M, Zhao Y, Weng X. Decreased parahippocampal activ-ity in associative priming: evidence from an event-related fMRI study. LearnMem 2008;15:703–10.
34] Rugg MD, Curran T. Event-related potentials and recognition memory. TrendsCogn Sci 2007;11:251–7.
35] Duncan CC, Barry RJ, Connolly JF, Fischer C, Michie PT, Naatanen R, et al.Event-related potentials in clinical research: guidelines for eliciting, recor-ding, and quantifying mismatch negativity, P300, and N400. Clin Neurophysiol2009;120:1883–908.
36] Kutas M, Federmeier KD. Thirty years and counting: finding meaning in the
N400 component of the event-related brain potential (ERP). Annu Rev Psychol2011;62:621–47.37] Pergola G, Gunturkun O, Koch B, Schwarz M, Daum I, Suchan B. Recall deficits instroke patients with thalamic lesions covary with damage to the parvocellularmediodorsal nucleus of the thalamus. Neuropsychologia 2012;50:2477–91.
[
[
Research 266 (2014) 63–76
38] Ranganath C, Paller KA. Neural correlates of memory retrieval and evaluation.Brain Res Cogn Brain Res 2000;9:209–22.
39] VanRullen R, Thorpe SJ. Is it a bird? Is it a plane? Ultra-rapid visual categorisa-tion of natural and artifactual objects. Perception 2001;30:655–68.
40] Voss JL, Lucas HD, Paller KA. Conceptual priming and familiarity: differentexpressions of memory during recognition testing with distinct neuro-physiological correlates. J Cogn Neurosci 2010;22:2638–51.
41] Foroni F, Pergola G, Argiris G, Rumiati RI. The FoodCast research image database(FRIDa). Front Hum Neurosci 2013;7:51.
42] Peterburs J, Pergola G, Koch B, Schwarz M, Hoffmann KP, Daum I, et al. Alterederror processing following vascular thalamic damage: evidence from an anti-saccade task. PLoS ONE 2011;6:e21517.
43] Groh-Bordin C, Zimmer HD, Mecklinger A. Feature binding in perceptual prim-ing and in episodic object recognition: evidence from event-related brainpotentials. Brain Res Cogn Brain Res 2005;24:556–67.
44] Pergola G, Ranft A, Mathias K, Suchan B. The role of the thalamic nuclei inrecognition memory accompanied by recall during encoding and retrieval: anfMRI study. Neuroimage 2013;74:195–208.
45] Bower GH, Thompson-Schill S, Tulving E. Reducing retroactive interference: aninterference analysis. J Exp Psychol Learn Mem Cogn 1994;20:51–66.
46] Sartori G, Lombardi L. Semantic relevance and semantic disorders. J Cogn Neu-rosci 2004;16:439–52.
47] Paller KA, Voss JL, Boehm SG. Validating neural correlates of familiarity. TrendsCogn Sci 2007;11:243–50.
48] Barrett SE, Rugg MD. Event-related potentials and the semantic matching ofpictures. Brain Cogn 1990;14:201–12.
49] Holcomb PJ, McPherson WB. Event-related brain potentials reflect semanticpriming in an object decision task. Brain Cogn 1994;24:259–76.
50] McPherson WB, Holcomb PJ. An electrophysiological investigation of semanticpriming with pictures of real objects. Psychophysiology 1999;36:53–65.
51] West WC, Holcomb PJ. Event-related potentials during discourse-level seman-tic integration of complex pictures. Brain Res Cogn Brain Res 2002;13:363–75.
52] Sitnikova T, West WC, Kuperberg GR, Holcomb PJ. The neural organization ofsemantic memory: Electrophysiological activity suggests feature-based segre-gation. Biol Psychol 2006;71:326–40.
53] Voss JL, Federmeier KD. FN400 potentials are functionally identical to N400potentials and reflect semantic processing during recognition testing. Psy-chophysiology 2011;48:532–46.
54] Tendolkar I, Doyle MC, Rugg MD. An event-related potential study of retroactiveinterference in memory. Neuroreport 1997;8:501–6.
55] Quamme JR, Yonelinas AP, Norman KA. Effect of unitization on associativerecognition in amnesia. Hippocampus 2007;17:192–200.
56] Schendan HE, Ganis G, Kutas M. Neurophysiological evidence for visual per-ceptual categorization of words and faces within 150 ms. Psychophysiology1998;35:240–51.
57] Voss JL, Schendan HE, Paller KA. Finding meaning in novel geometric shapesinfluences electrophysiological correlates of repetition and dissociates percep-tual and conceptual priming. Neuroimage 2010;49:2879–89.
58] Joyce C, Rossion B. The face-sensitive N170 and VPP components manifest thesame brain processes: the effect of reference electrode site. Clin Neurophysiol
2005;116:2613–31.59] George N, Jemel B, Fiori N, Renault B. Face and shape repetition effects inhumans: a spatio-temporal ERP study. Neuroreport 1997;8:1417–23.
60] VanRullen R, Thorpe SJ. The time course of visual processing: from early per-ception to decision-making. J Cogn Neurosci 2001;13:454–61.




















