
Mineralogy and petrography of C asteroid regolith: The Sutter's Mill CM meteorite
Upload
independentCategory
view
3download
0
Arch Hydrobiol 157 2 213ndash225 Stuttgart May 2003
Feeding behaviour of the asteroid Meyenastergelatinosus in response to changes in abundanceof the scallop Argopecten purpuratus in northernChile
Marco Ortiz1 Sandra Jesse2 Wolfgang Stotz3 and MatthiasWolff2
With 5 figures and 2 tables
Abstract Between winter 1996 and autumn 1997 the prey spectrum and feedingbehaviour of the sea star Meyenaster gelatinosus (Meyen 1834) and the seasonalabundance of the dominant benthic macrofauna were analysed in (1) a shore-fringingseagrass meadow and (2) an adjacent sand-gravel benthic system of Tongoy Bay (cen-tral-north Chile) Of 524 sea stars collected 130 (25 ) had food in their stomachs orwere feeding on the scallop Argopecten purpuratus (Lamarck 1819) snails Calytraeatrochiformis (Born 1778) Tegula luctuosa (Orbigny 1841) and T eryomphala(Jones 1844) and small epifauna constituted by the polyplacophora Chiton spp gast-ropods such as Nassarius gayi (Kiener 1835) Nucula pisum (Sowerby 1835) Turri-tela cingulata (Sowerby 1825) and Mitrella unifasciata (Sowerby 1832) The scal-lop A purpuratus suffered an intensive commercial harvest during the study periodwhich decreased significantly its abundance in both habitats Before harvesting Apurpuratus constituted the principal prey for M gelatinosus in the seagrass habitatwhereas all prey types were equally distributed in the sea starrsquos diet in the sand-gravelhabitat After harvesting the frequency of occurrence of the scallop in the stomachs ofthe sea star in the seagrass habitat decreased and was substituted by small epifaunaFurthermore in the seagrass habitat after harvesting the breadth of the feeding-nicheof the sea star increased indicating that the sea star became more of a generalist fee-der In the sand-gravel habitat after harvesting the diet composition remained unal-
1 Authorsrsquo addresses Instituto de Investigaciones Oceanoloacutegicas Facultad de Recur-sos del Mar Universidad de Antofagasta PO Box 170 ChileE-Mail mortizuantofcl Corresponding author2 Zentrum fuumlr Marine Tropenoumlkologie (ZMT) Fahrenheitstrasse 6 28359 BremenGermany E-Mail jesseuni-bremende mwolffuni-bremende3 Grupo de Ecologiacutea y Manejo Universidad Catoacutelica del Norte Casilla 117 CoquimboChile E-mail wstotzucncl
DOI 1011270003-913620030157-0213 0003-9136030157-0213 $ 325atilde 2003 E Schweizerbartrsquosche Verlagsbuchhandlung D-70176 Stuttgart
214 Marco Ortiz et al
tered but M gelatinosus showed an inverse tendency specialising on the scallop Ad-ditionally M gelatinosus increased its feeding preferences on A purpuratus in bothhabitats (especially in sand-gravel) when the abundance of the scallop declined M ge-latinosus feeds on a wide size-range (4 to 14 cm) of the scallop with a marked domi-nance of larger size classes Our results confirm in part the observations of local fish-ermen of Puerto Aldea regarding the possible negative impact of M gelatinosus on thestanding stock of the commercially exploited scallop
Key words Argopecten feeding-niche breadth prey spectrum Ivlev index manage-ment polices subtidal communities
Introduction
Sea stars as top predators exert relevant impacts on coastal ecosystems andsome of them are considered keystone species (sensu Paine 1966 1974) Eventhough many studies had to be carried out to test the keystone-species hypo-thesis in different coastal ecosystems most of these works were conducted inintertidal communities (eg Phillips 1976 1977 Branch 1978 Phillips ampCastori 1982 Paine et al 1985 Tokeshi et al 1989 Arrontes amp Under-wood 1991 Espoz amp Castilla 2000) Instead few investigations have beenaddressed into subtidal communities and the role of these species thereinMost of them were essentially designed to estimate the effects of the sea starspecies on single prey species such as whelks clams or scallops (eg Angeret al 1977 Dayton et al 1977 Barbeau amp Scheibling 1994 a b Arse-nault amp Himmelmann 1996 Rochette et al 1996 Barbeau et al 1998Morissette amp Himmelmann 2000)
The asteroid Meyenaster gelatinosus is one of the dominant predators in-habiting benthic systems along the central-north Chilean coast (Madsen 1956Stotz 2001 pers comm) including the Tongoy Bay (Wolff amp Alarcoacuten1993) and the management area of Puerto Aldea (Ortiz et al unpubl Fig 1)Dayton et al (1977) described the feeding behaviour of M gelatinosus fromcommunities of central-south Chile where it inhabits exposed rocky shoresSince M gelatinosus occurs sympatrically with the economically importantscallop Argopecten purpuratus the fishermen of Puerto Aldea have suggestedimplementation of a management policy that intends to decrease the abun-dance of this ldquoundesiredrdquo asteroid Stotz amp Gonzalez (1997) proposed someguidelines for the development of a sustainable harvest strategy for the pop-ulation of A purpuratus in this management area based on the population dy-namic of the scallop However the proposed strategy does not take into ac-count its ecological linkages to other populations
Between June 1996 (winter) and May 1997 (autumn) a study was carriedout to estimate the abundance and production of dominant macrofauna species
Feeding behaviour of M gelatinosus 215
Fig 1 A Main topographical types along the Chilean coast 1 = dominated byexposed rocky shores 2 = dominated by exposed sandy shores 3 = mostly insularsystems B Principal bay systems of the IV Region of Coquimbo Chile C Study areaof Puerto Aldea located in southern part of Tongoy Bay
inhabiting the benthic community of Puerto Aldea During the study period Apurpuratus suffered two intensive harvests (September of 1996 and Decem-berndashJanuary of 1996ndash97) which reduced its biomass significantly (ca 50 Ortiz et al unpubl) The changes in scallop biomass due to harvesting maybe considered as a natural not replicated experiment Therefore the objectivesof this study were (1) to describe the prey spectrum of M gelatinosus beforeand after fishing on the scallop (2) to estimate the relative importance of A
216 Marco Ortiz et al
purpuratus in its diet and (3) to determine if M gelatinosus behaves as a gen-eralist or specialist feeder of A purpuratus
Material and methods
Description of the study area
The study was carried out in Tongoy Bay (30 15centS ndash 71 31centW) in the IV Region (Co-quimbo Chile Fig 1) The benthic area of Puerto Aldea corresponds to one of the 168territorial use rights for fisheries (management areas) established along the coastalChilean ecosystems (Castilla 2000) The management areas are assigned to fisher-men associations for the planning of sustainable manipulations of the systems in orderto increase the productivity of valuable resources In the study area four different hab-itats can be recognised (1) seagrass meadows from 0 to 4 m depth (2) sand-gravel be-tween 4 and 10 m (3) sand flats between 10 and 14 m and (4) mud flats gt14 m depth Acomplete description of these habitats was done by Jesse (2001) and Ortiz et al (un-publ) The entire Tongoy Bay and particularly the subtidal benthic system of PuertoAldea are protected from the prevailing south-west winds by Lengua de Vaca penin-sula The upwelling center near to Punta de Vaca (Daneri et al 2000) is one of themost important of northern Chile and supplies nutrients to the ecosystem and preventsthe establishment of a stable thermocline during summer Temperature of bottom waterranges between 13 C and 17 C between winter and summer respectively (Jesse 2001)
Sampling design
The analysis of the food spectrum of M gelatinosus was done in two habitats the sea-grass meadows composed exclusively of Heterozostera tasmanica (Martens exAscher) and the adjacent area constituted by sand and gravel
The seasonal biomass variation (between winter 1996 and autumn 1997) of M ge-latinosus A purpuratus and the gastropods Calyptraea trochiformis Tegula luctuosaand T euryonphala was determined in both habitats at 5 randomly chosen stationsThree random samples per station were collected by semi-autonomous diving(HOOKAH gear) for M gelatinosus and A purpuratus and 20 samples per station forC trochiformis and Tegula sp (Table 1) The prey species of M gelatinosus were de-termined daylight in situ as the sea star spreads out its stomach enveloping the preywhich allows easy recognition of its food Additionally predator and prey sizes wererecorded
Statistical analysis
Bartlett and Hartleyrsquos tests were used to evaluate the normality of data and Cochranrsquostest to assess homogeneity of variances (Underwood 1997) When non-normality andheterogeneity were significant (p lt 0005) data were transformed using square root orlog10 as most appropriate (Sokal amp Rohlf 1995 Underwood 1997) The lowerp-level (p lt0005) was used because parametric tests are robust with non-normality of
Feeding behaviour of M gelatinosus 217
data and non-severe heterogeneity of variance (Box 1953 Tiku et al 1986 Weera-handi 1995 Underwood 1997) T-test and one-way ANOVAs were then applied withp lt005 Post hoc analysis was done using Student-Newman-Keuls test (Steel amp Tor-rie 1988 Underwood 1997) with p lt007 (Type I error) according to the adjustmentrecommended by Day amp Quinn (1989) A posteriori statistical power analysis wascalculated in those situations where the null hypothesis was not rejected
Two analyses were used to estimate the feeding behaviour of M gelatinosus Thebreadth of feeding niche (B) was calculated using frequency () of occurrence (sensuLevins 1968) as follows
B =S pi
2 (1)
1
i =1
n
where pi denotes the wet weight proportion of the prey species i to the total prey bio-mass consumed by the sea star and n is the total number of different prey species Thisindex ranges from 1 when the predator concentrates its feeding on a single species(specialisation) to values gt 1 when different prey species are consumed in similar pro-portions (generalisation) The electivity or preference of M gelatinosus for a particularprey was estimated by Ivlevrsquos index (E Krebs 1989) which relates the proportion of aprey in the diet to its relative abundance in the environment as follows
Ei = vi ndash ni (2)vi + ni
where vi is the percentage of species i in the diet of the predator and ni is the percent-age of species i in the environment This index ranges from positive to negative valuesfor the preferred and the avoided preys respectively
Results
Of 524 individuals of M gelatinosus examined only 130 (~25 ) were feed-ing on different prey species Prey species included A purpuratus the gastro-pods Calyptraea trochiformis (Born 1778) Tegula luctuosa and T euryom-phala (grouped as Tegula sp) and smaller gastropods such as Nassarius spNucula sp Turritela sp and Mitrella sp and the polyplacophora Chiton spp(grouped as small epifauna Table 1) Note that these different species havedifferent mobilities the scallop is highly mobile compared to Tegula sp andsmall epifauna whereas the gastropod C trochiformis is semi-sessile
The index of niche breadth showed that in the seagrass meadow M gelati-nosus changed from a specialist (B = 105) to a generalist (B = 217) feeder asthe biomass of A purpuratus is decreased while in the sand-gravel habitat aninverse tendency seems to be observed (Table 2) The Ivlev index indicatedthat in sand-gravel habitat the preference of M gelatinosus for A purpuratus
218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

214 Marco Ortiz et al
tered but M gelatinosus showed an inverse tendency specialising on the scallop Ad-ditionally M gelatinosus increased its feeding preferences on A purpuratus in bothhabitats (especially in sand-gravel) when the abundance of the scallop declined M ge-latinosus feeds on a wide size-range (4 to 14 cm) of the scallop with a marked domi-nance of larger size classes Our results confirm in part the observations of local fish-ermen of Puerto Aldea regarding the possible negative impact of M gelatinosus on thestanding stock of the commercially exploited scallop
Key words Argopecten feeding-niche breadth prey spectrum Ivlev index manage-ment polices subtidal communities
Introduction
Sea stars as top predators exert relevant impacts on coastal ecosystems andsome of them are considered keystone species (sensu Paine 1966 1974) Eventhough many studies had to be carried out to test the keystone-species hypo-thesis in different coastal ecosystems most of these works were conducted inintertidal communities (eg Phillips 1976 1977 Branch 1978 Phillips ampCastori 1982 Paine et al 1985 Tokeshi et al 1989 Arrontes amp Under-wood 1991 Espoz amp Castilla 2000) Instead few investigations have beenaddressed into subtidal communities and the role of these species thereinMost of them were essentially designed to estimate the effects of the sea starspecies on single prey species such as whelks clams or scallops (eg Angeret al 1977 Dayton et al 1977 Barbeau amp Scheibling 1994 a b Arse-nault amp Himmelmann 1996 Rochette et al 1996 Barbeau et al 1998Morissette amp Himmelmann 2000)
The asteroid Meyenaster gelatinosus is one of the dominant predators in-habiting benthic systems along the central-north Chilean coast (Madsen 1956Stotz 2001 pers comm) including the Tongoy Bay (Wolff amp Alarcoacuten1993) and the management area of Puerto Aldea (Ortiz et al unpubl Fig 1)Dayton et al (1977) described the feeding behaviour of M gelatinosus fromcommunities of central-south Chile where it inhabits exposed rocky shoresSince M gelatinosus occurs sympatrically with the economically importantscallop Argopecten purpuratus the fishermen of Puerto Aldea have suggestedimplementation of a management policy that intends to decrease the abun-dance of this ldquoundesiredrdquo asteroid Stotz amp Gonzalez (1997) proposed someguidelines for the development of a sustainable harvest strategy for the pop-ulation of A purpuratus in this management area based on the population dy-namic of the scallop However the proposed strategy does not take into ac-count its ecological linkages to other populations
Between June 1996 (winter) and May 1997 (autumn) a study was carriedout to estimate the abundance and production of dominant macrofauna species
Feeding behaviour of M gelatinosus 215
Fig 1 A Main topographical types along the Chilean coast 1 = dominated byexposed rocky shores 2 = dominated by exposed sandy shores 3 = mostly insularsystems B Principal bay systems of the IV Region of Coquimbo Chile C Study areaof Puerto Aldea located in southern part of Tongoy Bay
inhabiting the benthic community of Puerto Aldea During the study period Apurpuratus suffered two intensive harvests (September of 1996 and Decem-berndashJanuary of 1996ndash97) which reduced its biomass significantly (ca 50 Ortiz et al unpubl) The changes in scallop biomass due to harvesting maybe considered as a natural not replicated experiment Therefore the objectivesof this study were (1) to describe the prey spectrum of M gelatinosus beforeand after fishing on the scallop (2) to estimate the relative importance of A
216 Marco Ortiz et al
purpuratus in its diet and (3) to determine if M gelatinosus behaves as a gen-eralist or specialist feeder of A purpuratus
Material and methods
Description of the study area
The study was carried out in Tongoy Bay (30 15centS ndash 71 31centW) in the IV Region (Co-quimbo Chile Fig 1) The benthic area of Puerto Aldea corresponds to one of the 168territorial use rights for fisheries (management areas) established along the coastalChilean ecosystems (Castilla 2000) The management areas are assigned to fisher-men associations for the planning of sustainable manipulations of the systems in orderto increase the productivity of valuable resources In the study area four different hab-itats can be recognised (1) seagrass meadows from 0 to 4 m depth (2) sand-gravel be-tween 4 and 10 m (3) sand flats between 10 and 14 m and (4) mud flats gt14 m depth Acomplete description of these habitats was done by Jesse (2001) and Ortiz et al (un-publ) The entire Tongoy Bay and particularly the subtidal benthic system of PuertoAldea are protected from the prevailing south-west winds by Lengua de Vaca penin-sula The upwelling center near to Punta de Vaca (Daneri et al 2000) is one of themost important of northern Chile and supplies nutrients to the ecosystem and preventsthe establishment of a stable thermocline during summer Temperature of bottom waterranges between 13 C and 17 C between winter and summer respectively (Jesse 2001)
Sampling design
The analysis of the food spectrum of M gelatinosus was done in two habitats the sea-grass meadows composed exclusively of Heterozostera tasmanica (Martens exAscher) and the adjacent area constituted by sand and gravel
The seasonal biomass variation (between winter 1996 and autumn 1997) of M ge-latinosus A purpuratus and the gastropods Calyptraea trochiformis Tegula luctuosaand T euryonphala was determined in both habitats at 5 randomly chosen stationsThree random samples per station were collected by semi-autonomous diving(HOOKAH gear) for M gelatinosus and A purpuratus and 20 samples per station forC trochiformis and Tegula sp (Table 1) The prey species of M gelatinosus were de-termined daylight in situ as the sea star spreads out its stomach enveloping the preywhich allows easy recognition of its food Additionally predator and prey sizes wererecorded
Statistical analysis
Bartlett and Hartleyrsquos tests were used to evaluate the normality of data and Cochranrsquostest to assess homogeneity of variances (Underwood 1997) When non-normality andheterogeneity were significant (p lt 0005) data were transformed using square root orlog10 as most appropriate (Sokal amp Rohlf 1995 Underwood 1997) The lowerp-level (p lt0005) was used because parametric tests are robust with non-normality of
Feeding behaviour of M gelatinosus 217
data and non-severe heterogeneity of variance (Box 1953 Tiku et al 1986 Weera-handi 1995 Underwood 1997) T-test and one-way ANOVAs were then applied withp lt005 Post hoc analysis was done using Student-Newman-Keuls test (Steel amp Tor-rie 1988 Underwood 1997) with p lt007 (Type I error) according to the adjustmentrecommended by Day amp Quinn (1989) A posteriori statistical power analysis wascalculated in those situations where the null hypothesis was not rejected
Two analyses were used to estimate the feeding behaviour of M gelatinosus Thebreadth of feeding niche (B) was calculated using frequency () of occurrence (sensuLevins 1968) as follows
B =S pi
2 (1)
1
i =1
n
where pi denotes the wet weight proportion of the prey species i to the total prey bio-mass consumed by the sea star and n is the total number of different prey species Thisindex ranges from 1 when the predator concentrates its feeding on a single species(specialisation) to values gt 1 when different prey species are consumed in similar pro-portions (generalisation) The electivity or preference of M gelatinosus for a particularprey was estimated by Ivlevrsquos index (E Krebs 1989) which relates the proportion of aprey in the diet to its relative abundance in the environment as follows
Ei = vi ndash ni (2)vi + ni
where vi is the percentage of species i in the diet of the predator and ni is the percent-age of species i in the environment This index ranges from positive to negative valuesfor the preferred and the avoided preys respectively
Results
Of 524 individuals of M gelatinosus examined only 130 (~25 ) were feed-ing on different prey species Prey species included A purpuratus the gastro-pods Calyptraea trochiformis (Born 1778) Tegula luctuosa and T euryom-phala (grouped as Tegula sp) and smaller gastropods such as Nassarius spNucula sp Turritela sp and Mitrella sp and the polyplacophora Chiton spp(grouped as small epifauna Table 1) Note that these different species havedifferent mobilities the scallop is highly mobile compared to Tegula sp andsmall epifauna whereas the gastropod C trochiformis is semi-sessile
The index of niche breadth showed that in the seagrass meadow M gelati-nosus changed from a specialist (B = 105) to a generalist (B = 217) feeder asthe biomass of A purpuratus is decreased while in the sand-gravel habitat aninverse tendency seems to be observed (Table 2) The Ivlev index indicatedthat in sand-gravel habitat the preference of M gelatinosus for A purpuratus
218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 215
Fig 1 A Main topographical types along the Chilean coast 1 = dominated byexposed rocky shores 2 = dominated by exposed sandy shores 3 = mostly insularsystems B Principal bay systems of the IV Region of Coquimbo Chile C Study areaof Puerto Aldea located in southern part of Tongoy Bay
inhabiting the benthic community of Puerto Aldea During the study period Apurpuratus suffered two intensive harvests (September of 1996 and Decem-berndashJanuary of 1996ndash97) which reduced its biomass significantly (ca 50 Ortiz et al unpubl) The changes in scallop biomass due to harvesting maybe considered as a natural not replicated experiment Therefore the objectivesof this study were (1) to describe the prey spectrum of M gelatinosus beforeand after fishing on the scallop (2) to estimate the relative importance of A
216 Marco Ortiz et al
purpuratus in its diet and (3) to determine if M gelatinosus behaves as a gen-eralist or specialist feeder of A purpuratus
Material and methods
Description of the study area
The study was carried out in Tongoy Bay (30 15centS ndash 71 31centW) in the IV Region (Co-quimbo Chile Fig 1) The benthic area of Puerto Aldea corresponds to one of the 168territorial use rights for fisheries (management areas) established along the coastalChilean ecosystems (Castilla 2000) The management areas are assigned to fisher-men associations for the planning of sustainable manipulations of the systems in orderto increase the productivity of valuable resources In the study area four different hab-itats can be recognised (1) seagrass meadows from 0 to 4 m depth (2) sand-gravel be-tween 4 and 10 m (3) sand flats between 10 and 14 m and (4) mud flats gt14 m depth Acomplete description of these habitats was done by Jesse (2001) and Ortiz et al (un-publ) The entire Tongoy Bay and particularly the subtidal benthic system of PuertoAldea are protected from the prevailing south-west winds by Lengua de Vaca penin-sula The upwelling center near to Punta de Vaca (Daneri et al 2000) is one of themost important of northern Chile and supplies nutrients to the ecosystem and preventsthe establishment of a stable thermocline during summer Temperature of bottom waterranges between 13 C and 17 C between winter and summer respectively (Jesse 2001)
Sampling design
The analysis of the food spectrum of M gelatinosus was done in two habitats the sea-grass meadows composed exclusively of Heterozostera tasmanica (Martens exAscher) and the adjacent area constituted by sand and gravel
The seasonal biomass variation (between winter 1996 and autumn 1997) of M ge-latinosus A purpuratus and the gastropods Calyptraea trochiformis Tegula luctuosaand T euryonphala was determined in both habitats at 5 randomly chosen stationsThree random samples per station were collected by semi-autonomous diving(HOOKAH gear) for M gelatinosus and A purpuratus and 20 samples per station forC trochiformis and Tegula sp (Table 1) The prey species of M gelatinosus were de-termined daylight in situ as the sea star spreads out its stomach enveloping the preywhich allows easy recognition of its food Additionally predator and prey sizes wererecorded
Statistical analysis
Bartlett and Hartleyrsquos tests were used to evaluate the normality of data and Cochranrsquostest to assess homogeneity of variances (Underwood 1997) When non-normality andheterogeneity were significant (p lt 0005) data were transformed using square root orlog10 as most appropriate (Sokal amp Rohlf 1995 Underwood 1997) The lowerp-level (p lt0005) was used because parametric tests are robust with non-normality of
Feeding behaviour of M gelatinosus 217
data and non-severe heterogeneity of variance (Box 1953 Tiku et al 1986 Weera-handi 1995 Underwood 1997) T-test and one-way ANOVAs were then applied withp lt005 Post hoc analysis was done using Student-Newman-Keuls test (Steel amp Tor-rie 1988 Underwood 1997) with p lt007 (Type I error) according to the adjustmentrecommended by Day amp Quinn (1989) A posteriori statistical power analysis wascalculated in those situations where the null hypothesis was not rejected
Two analyses were used to estimate the feeding behaviour of M gelatinosus Thebreadth of feeding niche (B) was calculated using frequency () of occurrence (sensuLevins 1968) as follows
B =S pi
2 (1)
1
i =1
n
where pi denotes the wet weight proportion of the prey species i to the total prey bio-mass consumed by the sea star and n is the total number of different prey species Thisindex ranges from 1 when the predator concentrates its feeding on a single species(specialisation) to values gt 1 when different prey species are consumed in similar pro-portions (generalisation) The electivity or preference of M gelatinosus for a particularprey was estimated by Ivlevrsquos index (E Krebs 1989) which relates the proportion of aprey in the diet to its relative abundance in the environment as follows
Ei = vi ndash ni (2)vi + ni
where vi is the percentage of species i in the diet of the predator and ni is the percent-age of species i in the environment This index ranges from positive to negative valuesfor the preferred and the avoided preys respectively
Results
Of 524 individuals of M gelatinosus examined only 130 (~25 ) were feed-ing on different prey species Prey species included A purpuratus the gastro-pods Calyptraea trochiformis (Born 1778) Tegula luctuosa and T euryom-phala (grouped as Tegula sp) and smaller gastropods such as Nassarius spNucula sp Turritela sp and Mitrella sp and the polyplacophora Chiton spp(grouped as small epifauna Table 1) Note that these different species havedifferent mobilities the scallop is highly mobile compared to Tegula sp andsmall epifauna whereas the gastropod C trochiformis is semi-sessile
The index of niche breadth showed that in the seagrass meadow M gelati-nosus changed from a specialist (B = 105) to a generalist (B = 217) feeder asthe biomass of A purpuratus is decreased while in the sand-gravel habitat aninverse tendency seems to be observed (Table 2) The Ivlev index indicatedthat in sand-gravel habitat the preference of M gelatinosus for A purpuratus
218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

216 Marco Ortiz et al
purpuratus in its diet and (3) to determine if M gelatinosus behaves as a gen-eralist or specialist feeder of A purpuratus
Material and methods
Description of the study area
The study was carried out in Tongoy Bay (30 15centS ndash 71 31centW) in the IV Region (Co-quimbo Chile Fig 1) The benthic area of Puerto Aldea corresponds to one of the 168territorial use rights for fisheries (management areas) established along the coastalChilean ecosystems (Castilla 2000) The management areas are assigned to fisher-men associations for the planning of sustainable manipulations of the systems in orderto increase the productivity of valuable resources In the study area four different hab-itats can be recognised (1) seagrass meadows from 0 to 4 m depth (2) sand-gravel be-tween 4 and 10 m (3) sand flats between 10 and 14 m and (4) mud flats gt14 m depth Acomplete description of these habitats was done by Jesse (2001) and Ortiz et al (un-publ) The entire Tongoy Bay and particularly the subtidal benthic system of PuertoAldea are protected from the prevailing south-west winds by Lengua de Vaca penin-sula The upwelling center near to Punta de Vaca (Daneri et al 2000) is one of themost important of northern Chile and supplies nutrients to the ecosystem and preventsthe establishment of a stable thermocline during summer Temperature of bottom waterranges between 13 C and 17 C between winter and summer respectively (Jesse 2001)
Sampling design
The analysis of the food spectrum of M gelatinosus was done in two habitats the sea-grass meadows composed exclusively of Heterozostera tasmanica (Martens exAscher) and the adjacent area constituted by sand and gravel
The seasonal biomass variation (between winter 1996 and autumn 1997) of M ge-latinosus A purpuratus and the gastropods Calyptraea trochiformis Tegula luctuosaand T euryonphala was determined in both habitats at 5 randomly chosen stationsThree random samples per station were collected by semi-autonomous diving(HOOKAH gear) for M gelatinosus and A purpuratus and 20 samples per station forC trochiformis and Tegula sp (Table 1) The prey species of M gelatinosus were de-termined daylight in situ as the sea star spreads out its stomach enveloping the preywhich allows easy recognition of its food Additionally predator and prey sizes wererecorded
Statistical analysis
Bartlett and Hartleyrsquos tests were used to evaluate the normality of data and Cochranrsquostest to assess homogeneity of variances (Underwood 1997) When non-normality andheterogeneity were significant (p lt 0005) data were transformed using square root orlog10 as most appropriate (Sokal amp Rohlf 1995 Underwood 1997) The lowerp-level (p lt0005) was used because parametric tests are robust with non-normality of
Feeding behaviour of M gelatinosus 217
data and non-severe heterogeneity of variance (Box 1953 Tiku et al 1986 Weera-handi 1995 Underwood 1997) T-test and one-way ANOVAs were then applied withp lt005 Post hoc analysis was done using Student-Newman-Keuls test (Steel amp Tor-rie 1988 Underwood 1997) with p lt007 (Type I error) according to the adjustmentrecommended by Day amp Quinn (1989) A posteriori statistical power analysis wascalculated in those situations where the null hypothesis was not rejected
Two analyses were used to estimate the feeding behaviour of M gelatinosus Thebreadth of feeding niche (B) was calculated using frequency () of occurrence (sensuLevins 1968) as follows
B =S pi
2 (1)
1
i =1
n
where pi denotes the wet weight proportion of the prey species i to the total prey bio-mass consumed by the sea star and n is the total number of different prey species Thisindex ranges from 1 when the predator concentrates its feeding on a single species(specialisation) to values gt 1 when different prey species are consumed in similar pro-portions (generalisation) The electivity or preference of M gelatinosus for a particularprey was estimated by Ivlevrsquos index (E Krebs 1989) which relates the proportion of aprey in the diet to its relative abundance in the environment as follows
Ei = vi ndash ni (2)vi + ni
where vi is the percentage of species i in the diet of the predator and ni is the percent-age of species i in the environment This index ranges from positive to negative valuesfor the preferred and the avoided preys respectively
Results
Of 524 individuals of M gelatinosus examined only 130 (~25 ) were feed-ing on different prey species Prey species included A purpuratus the gastro-pods Calyptraea trochiformis (Born 1778) Tegula luctuosa and T euryom-phala (grouped as Tegula sp) and smaller gastropods such as Nassarius spNucula sp Turritela sp and Mitrella sp and the polyplacophora Chiton spp(grouped as small epifauna Table 1) Note that these different species havedifferent mobilities the scallop is highly mobile compared to Tegula sp andsmall epifauna whereas the gastropod C trochiformis is semi-sessile
The index of niche breadth showed that in the seagrass meadow M gelati-nosus changed from a specialist (B = 105) to a generalist (B = 217) feeder asthe biomass of A purpuratus is decreased while in the sand-gravel habitat aninverse tendency seems to be observed (Table 2) The Ivlev index indicatedthat in sand-gravel habitat the preference of M gelatinosus for A purpuratus
218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 217
data and non-severe heterogeneity of variance (Box 1953 Tiku et al 1986 Weera-handi 1995 Underwood 1997) T-test and one-way ANOVAs were then applied withp lt005 Post hoc analysis was done using Student-Newman-Keuls test (Steel amp Tor-rie 1988 Underwood 1997) with p lt007 (Type I error) according to the adjustmentrecommended by Day amp Quinn (1989) A posteriori statistical power analysis wascalculated in those situations where the null hypothesis was not rejected
Two analyses were used to estimate the feeding behaviour of M gelatinosus Thebreadth of feeding niche (B) was calculated using frequency () of occurrence (sensuLevins 1968) as follows
B =S pi
2 (1)
1
i =1
n
where pi denotes the wet weight proportion of the prey species i to the total prey bio-mass consumed by the sea star and n is the total number of different prey species Thisindex ranges from 1 when the predator concentrates its feeding on a single species(specialisation) to values gt 1 when different prey species are consumed in similar pro-portions (generalisation) The electivity or preference of M gelatinosus for a particularprey was estimated by Ivlevrsquos index (E Krebs 1989) which relates the proportion of aprey in the diet to its relative abundance in the environment as follows
Ei = vi ndash ni (2)vi + ni
where vi is the percentage of species i in the diet of the predator and ni is the percent-age of species i in the environment This index ranges from positive to negative valuesfor the preferred and the avoided preys respectively
Results
Of 524 individuals of M gelatinosus examined only 130 (~25 ) were feed-ing on different prey species Prey species included A purpuratus the gastro-pods Calyptraea trochiformis (Born 1778) Tegula luctuosa and T euryom-phala (grouped as Tegula sp) and smaller gastropods such as Nassarius spNucula sp Turritela sp and Mitrella sp and the polyplacophora Chiton spp(grouped as small epifauna Table 1) Note that these different species havedifferent mobilities the scallop is highly mobile compared to Tegula sp andsmall epifauna whereas the gastropod C trochiformis is semi-sessile
The index of niche breadth showed that in the seagrass meadow M gelati-nosus changed from a specialist (B = 105) to a generalist (B = 217) feeder asthe biomass of A purpuratus is decreased while in the sand-gravel habitat aninverse tendency seems to be observed (Table 2) The Ivlev index indicatedthat in sand-gravel habitat the preference of M gelatinosus for A purpuratus
218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

218 Marco Ortiz et al
Table 1 Summary of the sampling design (number of samples [N] and area of sam-pling unit) assessing the abundance of prey species consumed by Meyenaster gelatino-sus in two different habitats after and before harvest of Argopecten purpuratus Tegulaluctuosa and T euryomphala are grouped as ldquoTegula sprdquo Chiton sp Nassarius spNucula sp Turritela sp and Mitrella sp are grouped as ldquosmall epifaunardquo
Habitats
Seagrass Sand-Gravel
Prey species N Area (m2) N Area (m2)
Argopecten purpuratus 15 20 15 20Megenaster gelatinosus 15 100 15 100Calyptraea trochiformis 60 025Tegula sp 60 025Small Epifauna
A sampling unit of 20 y 100 m2 means a transect of 10 m acute 2m and 50m acute 2 m respec-tively A sampling unit of 025 m2 means a cuadrate of 05 m acute 05 m Personal estimations (Ortiz pers obs) Abundance data taking from literature (Gonzaacutelez 1992 Leoacuten 2000)
increased after harvesting of scallop For most other prey species the electiv-ity index presented an inverse pattern (Table2)
Before the scallop harvest in winter-spring 1996 the scallop was the prin-cipal prey for the asteroid in the seagrass habitat (555 ) whereas no specificdominant prey species in stomachs was observed in the sand-gravel habitat(Table 2 Fig 2)
However after the harvesting period in summer 1997 this situation changedin the seagrass habitat but not in the sand-gravel habitat In the seagrass theimportance of the scallops declined (273 ) sea stars preyed more upon smallepifauna than on A purpuratus
Sea stars feeding on A purpuratus were significantly larger than sea starsfeeding on C trochiformis Tegula sp and small epifauna (F(3126) = 461p lt005 Fig 3) As well sea stars fed on larger scallops (ca 10 cm shell width)and not on small (lt 30 cm shell width Fig 4) The biomass of A purpuratusdecreased significantly in both habitats as consequence of the harvesting activ-ities (Fig 5b F(3102) = 1268 plt005) However sea stars did not show signif-icant changes in biomass in either habitat as a response to the decline in scal-lop abundance Only a significant difference between habitats was detected(ANOVA F(3116) = 918 p lt 005 SNK p lt007 Fig 5 a) As for other preyspecies C trochiformis presented significant seasonal differences in biomass(t(243) = 317 p lt005 Fig 5 c) whereas Tegula sp remained relatively con-stant (t(243) = 010 p = 092 Fig 5 d) nonetheless a rigorous conclusion doesnot seem justified due to the high level of type II error (b = 050) For the spe-cies grouped as small epifauna information of seasonal changes in abundance
Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 219
Table 2 Abundance in biomass (g wet weightm2) and in percentage () of prey spe-cies in the environment and in the stomachs of Meyenaster gelatinosus Feeding-nichebreadth and prey electivity of M gelatinosus Note the estimation of abundance ofsmall epifauna in the environment is based on Gonzaacutelez (1992) and Leoacuten (2000)
Item Seagrass
Biomass of prey Percentage of Biomass Breadth of Ivlevin environ- prey in stomachs of prey in feeding indexment () () (N) stomachs niche (B) (Ei)
Habitats
Before fishingArgopecten purpuratus 1908 (473) 555 (15) 22295 095 008Calyptraea trochiformis lt30 (065) 296 (8) 4287 00004 096Tegula sp lt 1900 (471) 37 (1) 291 0000002 ndash085Small epifauna lt 200 (495) 11 (3) 93 000002 038
105
After fishingA purpuratus 11831 (3571) 273 (3) 29325 031 013C trochiformis lt30 (09) 91 (1) 20174 015 082Tegula sp lt 1900 (5735) 91 (1) 1063 00004 ndash073Small epifauna lt 200 (604) 545 (6) 166 00012 08
217
Sand-gravel
Before fishingA purpuratus 10732 (1148) 228 (13) 13529 053 033C trochiformis 12195 (1305) 315 (18) 45149 0059 041Tegula sp 47892 (5123) 218 (12) 1384 000005 04Small epifauna ~2266 (2424) 239 (14) 434 00005 ndash00007
169
After fishingA purpuratus 3577 (326) 228 (8) 69317 066 075C trochiformis 26689 (2433) 257 (9) 9675 0013 003Tegula sp 56402 (5141) 314 (11) 4232 00024 ndash024Small epifauna ~2266 (210) 201 (7) 217 000064 ndash002
148
was not available but it is known that the abundance of small epifauna is con-siderably higher in the sand-gravel than the seagrass habitat (Gonzalez 1992Leon 2000)
Discussion
The present study described the food items and the feeding spectrum of adominant sea star predator in a subtidal system of the north-central Chilean
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002
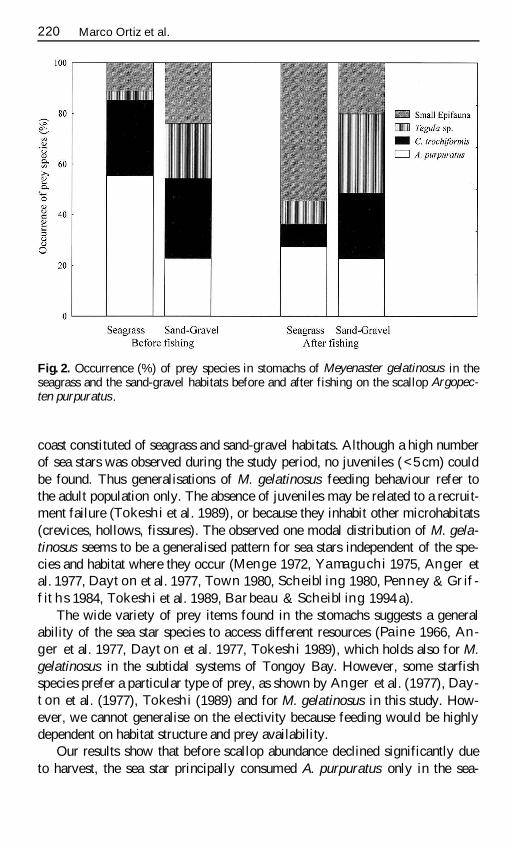
220 Marco Ortiz et al
Fig 2 Occurrence () of prey species in stomachs of Meyenaster gelatinosus in theseagrass and the sand-gravel habitats before and after fishing on the scallop Argopec-ten purpuratus
coast constituted of seagrass and sand-gravel habitats Although a high numberof sea stars was observed during the study period no juveniles (lt5 cm) couldbe found Thus generalisations of M gelatinosus feeding behaviour refer tothe adult population only The absence of juveniles may be related to a recruit-ment failure (Tokeshi et al 1989) or because they inhabit other microhabitats(crevices hollows fissures) The observed one modal distribution of M gela-tinosus seems to be a generalised pattern for sea stars independent of the spe-cies and habitat where they occur (Menge 1972 Yamaguchi 1975 Anger etal 1977 Dayton et al 1977 Town 1980 Scheibling 1980 Penney amp Grif-fiths 1984 Tokeshi et al 1989 Barbeau amp Scheibling 1994 a)
The wide variety of prey items found in the stomachs suggests a generalability of the sea star species to access different resources (Paine 1966 An-ger et al 1977 Dayton et al 1977 Tokeshi 1989) which holds also for Mgelatinosus in the subtidal systems of Tongoy Bay However some starfishspecies prefer a particular type of prey as shown by Anger et al (1977) Day-ton et al (1977) Tokeshi (1989) and for M gelatinosus in this study How-ever we cannot generalise on the electivity because feeding would be highlydependent on habitat structure and prey availability
Our results show that before scallop abundance declined significantly dueto harvest the sea star principally consumed A purpuratus only in the sea-
Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 221
Fig 3 Size structure of Meyenaster gelatinosus and the size of individuals duringfeeding activities on a) A purpuratus b) C trochiformis c) Tegula sp and d) SmallEpifauna (Note Size structure is pooled over time and habitats)
Fig 4 Size structure of Argopecten purpuratus and size of individuals consumed byMeyenaster gelatinosus (Note Size structure is pooled over time and habitats)
222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

222 Marco Ortiz et al
Fig 5 Average biomass (g wet weightm2) (plusmn standard deviation) of a) M gelatino-sus b) A pupuratus c) C trochiformis and d) Tegula sp pooled over time and habi-tats Small letters above the bars indicate where significant differences were detectedafter SNK test (similar letters indicate no differences)
grass habitat This may be explained by scallop dominance in the seagrassand therefore may reflect opportunistic feeding behaviour rather than trueelectivity After fishing the small epifauna replaced the scallop in the diet ofthe sea star which could be taken as a behavioural response of sea stars tochanges in prey availability
Only large individuals of M gelatinosus fed preferentially on A purpura-tus in comparison to smaller ones that consume other prey items This wouldsuggest some type of difficulty for sea stars to feed on small scallops possiblydue to a large handling time which increases the chance of scallops to escape(Barbeau amp Scheibling 1994 a) Scallops are highly mobile in comparisonto other prey species However further studies should be addressed to estimatethe energetic content of prey species to test an eventual energetic explanationfor the pattern feeding observed Likewise the fact that only small size-classscallops (lt30 cm) were not consumed by the sea star may be explained bynatural escape favoured by caves and bottom currents andor low energeticcontent
While the diet composition of M gelatinosus seems to reflect directly thefoodprey supply the feeding-niche breadth and the Ivlev indexes revealed un-expected insights Based on the breadth of the feeding-niche the significantdepletion of A purpuratus (by harvest) would be in itself a stimulus strong
Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 223
enough for the sea star to change its behaviour from a generalist to a specialistfeeder in the sand-gravel habitat The Ivlev index indicated an increase ofelectivity for the prey A pupuratus in both habitats (especially in sand-gravel)after harvesting These results show that M gelatinosus would be eventually afacultative specialist predator under certain circumstances This finding has asyet not been reported for subtidal sea star species possibly due to the diffi-culty to conduct experiments where changes in abundance of preys areneeded
These results confirm in part the notion of fishermen of Puerto Aldea thatM gelatinosus may exert a significant negative impact upon the abundanceand production of the commercially important scallop A purpuratus Never-theless further investigations are needed to (1) quantify consumption rate ofM gelatinosus on different prey species (2) determine energetic content of theprey species to assess the putative hypothesis that the preference for scallopsin sand-gravel habitat after harvesting could be a consequence of its compara-tively higher energetic contents
Acknowledgements
We thank two anonymous referees who improved substantially the overall quality ofthis paper
References
Anger K Rogal U Schreiver G amp Valentin G (1977) In-situ investigationson the echinoderm Asterias rubens as a predator of soft-bottom communities inthe western Baltic Sea ndash Helgolaumlnder wiss Meeresunters 29 439 ndash459
Arrontes J amp Underwood A J (1991) Experimental studies on some aspects ofthe feeding ecology of the intertidal starfish Patiriella exigua ndash J Exp Mar BiolEcol 148 255 ndash 269
Arsenault D J amp Himmelmann J H (1996) Size-related changes in vulnerabilityto predators and spatial refuge use by juvenil Iceland scallops Chlamys islandicandash Mar Ecol Prog Ser 140 115 ndash120
Barbeau M A amp Scheibling R E (1994 a) Behavioral mechanisms of prey sizeselection by sea stars (Asterias vulgaris Verril) and crabs (Cancer irroratus Say)preying on juvenile sea scallops (Placopecten magallanicus (Gmelin)) ndash J ExpMar Biol Ecol 180 103 ndash136
ndash ndash (1994 b) Temperature effects on predation of juvenile sea scallops [Placopec-ten magallanicus (Gmelin) by sea stars (Asterias vulgaris Verril) and crabs(Cancer irroratus Say)] ndash J Exp Mar Biol Ecol 182 27ndash 47
Barbeau M A Scheibling R E amp Hatcher B G (1998) Behavioural responsesof predatory crabs and sea stars to varying density of juvenile sea scallops ndashAquaculture 169 87ndash98
Box G (1953) Non-normality and test on variances ndash Biometrika 40 318 ndash335
224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

224 Marco Ortiz et al
Branch G M (1978) The responses of South African patellid limpets to invertebratepredators ndash Zool Afr 13 221 ndash232
Castilla J C (2000) Roles of experimental marine ecology in coastal managementand conservation ndash J Exp Mar Biol Ecol 250 3ndash21
Daneri G Dellarossa V Quintildeones R Jacob B Montero P amp Ulloa O(2000) Primary production and community respiration in the Humboldt CurrentSystem off Chile and associated oceanic areas ndash Mar Ecol Prog Ser 197 41ndash 49
Day R amp Quinn G (1989) Comparisons of treatments after an analysis of variancein ecology ndash Ecol Monogr 59 433 ndash463
Dayton P K Rosenthal R Mahen L amp Antezana T (1977) Populationstructure and foraging biology of the predaceous Chilean asteroid Meyenaster ge-latinosus and the escape biology of its preys ndash Mar Biol 39 361 ndash 370
Espoz C amp Castilla J C (2000) Escape responses of four Chilean intertidal lim-pets to seastars ndash Mar Biol 137 887 ndash892
Gonzalez S (1992) Heterozostera tasmanica (Martens en Aschers) den Hartog ycomunidad asociada en el norte de Chile ndash Tesis de Licenciatura UniversidadCatoacutelica del Norte Facultad de Cs del Mar Chile
Jesse S (2001) Comparative ecology of sympatric brachyuran crab species in theshallow subtidal of the Pacific coast of north Chile and their importance for the ar-tisanal fishery Puerto Aldea ndash PhD Dissertation University of Bremen Germany125 pp
Krebs Ch J (1989) Ecological methodology ndash Harper amp Row NY 654 ppLeon R (2000) Relaciones troacuteficas del Cancer polyodon (Poepping 1936) ndash Tesis
para obtener el tiacutetulo de Bioacutelogo Marino Universidad Catoacutelica del Norte sede Co-quimbo Chile
Levins R (1968) Evolution in changing environments ndash Princeton University PressPrinceton 120 pp
Madsen F J (1956) Asteroid Reports of the Lund University Chile expedition1948 ndash1949 ndash Acta Univ Lund (N 2) 67 1ndash53
Menge B A (1972) Competition for food between two intertidal starfish species andits effect on body size and feeding ndash Ecology 49 603 ndash619
Morissette S amp Himmelmann J H (2000) Decision of the asteroid Leptasteriaspolaris to abandon its prey when confronted with its predator the asteroid Asteriasvulgaris ndash J Exp Mar Biol Ecol 252 151 ndash157
Paine R T (1966) Food web complexity and species diversity ndash Amer Nat 10065ndash 75
ndash (1974) Intertidal community structure experimental studies on the relationshipbetween a dominant competitor and its principal predator ndash Oecologia 15 93ndash120
Paine R T Castilla J C amp Cancino J (1985) Perturbation and recovery pat-terns of starfish-dominated intertidal assemblages in Chile New Zealand andWashington State ndash Amer Nat 125 679 ndash 691
Penney A J amp Griffiths C L (1984) Prey selection and the impact of the starfishMarthasterias glacialis (L) and other predators on the mussel Choromytilus meri-dionalis (Krauss) ndash J Exp Mar Biol Ecol 75 19 ndash36
Phillips D W (1976) The effect of a species-specific avoidance response to pred-atory starfish distribution of two gastropods ndash Oecologia 23 83ndash94
Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002

Feeding behaviour of M gelatinosus 225
ndash (1977) Avoidance and escape responses of the gastropod mollusc Olivella bipli-cata (Sowerby ) to predatory asteroids ndash J Exp Mar Biol Ecol 28 77ndash86
Phillips D W amp Castori P (1982) Defensive responses to predatory seastars bytwo specialist limpets Notoacmaea insessa (Hinds) and Collisella instabilis(Gould) associated with marine algae ndash J Exp Mar Biol Ecol 59 23ndash30
Rochette R McNeil J N amp Himmelmann J H (1996) Inter-and-intra-popula-tion variations in the response of the whelk Buccinum undatum to the predatoryasteroid Leptasterias polaris ndash Mar Ecol Prog Ser 142 193 ndash201
Scheibling R E (1980) Abundance spatial distribution and size structure of pop-ulations of Oreaster reticulatus (Echinodermata Asteroidea) ion sand bottoms ndashMar Biol 57 107 ndash119
Sokal R amp Rohlf F (1995) Biometry 3rd ed ndash Freeman and Co San Francisco878 pp
Steel R amp Torrie H (1988) Bioestadiacutestica principios y procedimientos 1st edndashMcGraw-Hill 662 pp
Stotz W amp Gonzalez S (1997) Abundance growth and production of the seascallop Argopecten purpuratus (Lamarck 1819) bases for sustainable exploitationof natural scallop beds in north-central Chile ndash Fish Res 32 173 ndash183
Tiku M Tan W amp Balakrishnan N (1986) Robust Inference ndash Marcel Dekker321 pp
Tokeshi M Estrella C amp Paredes C (1989) Feeding ecology of a size-struc-tured predator population the South American sun-star Heliaster helianthus ndashMar Biol 100 495 ndash505
Town J C (1980) Movement morphology reproductive periodicity and some fac-tors affecting gonad production in the seastar Astrolole scabra (Hutton) ndash JExp Mar Biol Ecol 44 111 ndash132
Underwood A J (1997) Experiments in ecology their logical design and interpreta-tion using analysis of variance ndash Cambridge University Press 504 pp
Yamaguchi M (1975) Coral-reef asteroids of Guam ndash Biotropica 7 12ndash23Weerahandi S (1995) ANOVA under unequal error variances ndash Biometrics 51
589 ndash599Wolff M amp Alarcoacuten E (1993) Structure of a scallop Argopecten purpuratus (La-
marck 1819) dominated subtidal macro-invertebrate assemblage in northernChile ndash J Shell Res 12 295 ndash304
Submitted 2 January 2002 accepted 25 October 2002
Copyright © 2022 FDOKUMEN




















