
Effect of immersion time of cultch on spatfall of the scallop Argopecten purpuratus (Lamarck 1819)...
17
Effect of immersion time of cultch on spatfall of the scallop Argopecten purpuratus (Lamarck 1819) in the Marine Reserve at La Riconada, Antofagasta, Chile MIGUEL AVENDAN ˜ O DI ´ AZ 1, *, MARCELA CANTILLA ´ NEZ SILVA 1 and JUAN PEN ˜ A FORNER 2 1 Departamento de Acuicultura, Universidad de Antofagasta, Casilla 170, Antofagasta, Chile; 2 Instituto de Acuicultura de Torre la Sal (CSIC), E-12595, Ribera de Cabanes, Castello ´n, Spain; *Author for correspondence (e-mail: [email protected]; fax: +56-55-637804) Received 1 April 2004; accepted in revised form 17 October 2005 Key words: Antofagasta, Argopecten purpuratus, Chile, Collectors, Environmental parameters, Japanese method, Scallops, Spat settlement Abstract. A pilot program to evaluate the feasibility of collecting juvenile (seed) scallops using Japanese technology was carried out from October 2001 to April 2002 in the Marine Reserve at La Rinconada, Antofagasta, Chile. The continuous presence of larvae in waters in the reserve allowed for collection of 400–15,340 spat per collector and showed the high reproductive capability of local scallops and the relationship between numbers of larvae in the water and amount of spatfall. Highest rates of settlement were proportional to high numbers of umbone larvae in the water at >5000 larvae m )3 when collectors were first immersed. Absence of a relationship between larval numbers and postlarval settlement with trophic factors such as chlorophyll and particulate organic matter in the water suggests that productivity was not limited by these factors. Harvest of spat from collectors immersed for four and five months had recovery efficiencies of 5.5% and 59.4% com- pared with settlement in collectors after immersion for one month. Cohort analyses of recovered spat suggested that settlement occurred over the lengthy immersion period and was due to the continuous presence of larvae in the water. The loss of a high percentage of spat over the 4–5 immersion period compared with settlement in the first month can be attributed to intraspecific competition of spat density and growth as well as clogging of the collectors by sediment and marine fouling that impeded a flow of water through the nets. Introduction The scallop, Argopecten purpuratus (Lamarck 1819), occurs from Paita, Peru (5ŶS, 81ŶW) to Tongoy, Chile (30ŶS, 71ŶW) and is one of the most important marine species in aquaculture in Chile and Peru (Akaboshi and Illanes 1983; Ysla and Vargas 1989; Avila et al. 1994; Bandin and Mendo 1999). Culture methods used in Chile are similar to those developed in Japan for culture of Patinopecten yessoensis (Jay 1857) (Ventilla 1982). The Japanese method to capture spat in the ocean is more advantageous than producing seed in hatcheries (Le Pennec 1997). Adaptation of the Japa- nese technique to capture A. purpuratus spat began in Chile in 1977 (Hojas Aquaculture International (2006) 14:267–283 ȑ Springer 2005 DOI 10.1007/s10499-005-9033-y
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effect of immersion time of cultch on spatfall of the scallop Argopecten purpuratus (Lamarck 1819)...

Effect of immersion time of cultch on spatfall
of the scallop Argopecten purpuratus (Lamarck 1819)
in the Marine Reserve at La Riconada, Antofagasta, Chile
MIGUEL AVENDANO DIAZ1,*, MARCELA CANTILLANEZSILVA1 and JUAN PENA FORNER2
1Departamento de Acuicultura, Universidad de Antofagasta, Casilla 170, Antofagasta, Chile;2Instituto de Acuicultura de Torre la Sal (CSIC), E-12595, Ribera de Cabanes, Castellon, Spain;
*Author for correspondence (e-mail: [email protected]; fax: +56-55-637804)
Received 1 April 2004; accepted in revised form 17 October 2005
Key words: Antofagasta, Argopecten purpuratus, Chile, Collectors, Environmental parameters,
Japanese method, Scallops, Spat settlement
Abstract. A pilot program to evaluate the feasibility of collecting juvenile (seed) scallops using
Japanese technology was carried out from October 2001 to April 2002 in the Marine Reserve at La
Rinconada, Antofagasta, Chile. The continuous presence of larvae in waters in the reserve allowed
for collection of 400–15,340 spat per collector and showed the high reproductive capability of local
scallops and the relationship between numbers of larvae in the water and amount of spatfall.
Highest rates of settlement were proportional to high numbers of umbone larvae in the water at
>5000 larvae m)3 when collectors were first immersed. Absence of a relationship between larval
numbers and postlarval settlement with trophic factors such as chlorophyll and particulate organic
matter in the water suggests that productivity was not limited by these factors. Harvest of spat from
collectors immersed for four and five months had recovery efficiencies of 5.5% and 59.4% com-
pared with settlement in collectors after immersion for one month. Cohort analyses of recovered
spat suggested that settlement occurred over the lengthy immersion period and was due to the
continuous presence of larvae in the water. The loss of a high percentage of spat over the 4–5
immersion period compared with settlement in the first month can be attributed to intraspecific
competition of spat density and growth as well as clogging of the collectors by sediment and marine
fouling that impeded a flow of water through the nets.
Introduction
The scallop, Argopecten purpuratus (Lamarck 1819), occurs from Paita, Peru(5�S, 81�W) to Tongoy, Chile (30�S, 71�W) and is one of the most importantmarine species in aquaculture in Chile and Peru (Akaboshi and Illanes 1983;Ysla and Vargas 1989; Avila et al. 1994; Bandin and Mendo 1999). Culturemethods used in Chile are similar to those developed in Japan for culture ofPatinopecten yessoensis (Jay 1857) (Ventilla 1982).
The Japanese method to capture spat in the ocean is more advantageousthan producing seed in hatcheries (Le Pennec 1997). Adaptation of the Japa-nese technique to capture A. purpuratus spat began in Chile in 1977 (Hojas
Aquaculture International (2006) 14:267–283 � Springer 2005
DOI 10.1007/s10499-005-9033-y

1982) when initial attempts to culture this species began (Avendano 1984;Illanes 1986). In recent years naturally caught spat has formed the basis of thelarge scallop culture industry that produces nearly 20,000 t annually (wholeweight), making Chile the third largest producer of pectinids in the world(Avendano et al. 2001). Indiscriminate harvest drastically reduced naturalpopulations (Wolff and Alarcon 1993; Avendano and Cantillanez 1996; Stotzand Gonzalez 1997; Stotz and Mendo 2001) and affected the ability of thesepopulations to produce sufficient spat for commercial scallop operations(Gajardo et al. 1996).
The La Rinconada Marine Reserve was established in northern Chile inSeptember 1997 by the Chilean government as a reserve for Argopecten pur-puratus populations. This resulted from observations that this species wasbeing over exploited in Chile but consistent breeding occurred in the MarineReserve because of favourable biological conditions (Avendano 1993; Aven-dano and Le Pennec 1996, 1997; Avendano and Cantillanez 1996, 1997, 2003).The Purple scallop culture in Chile produces the third largest harvest in theworld, after China and Japan, where scallop spat are caught from naturalpopulations in some bays, like Mutsu Bay in Japan (Ventilla 1982). Theobjective of this work was to investigate scallop spat collection in Chile. TheMarine Reserve was proposed as an experimental area where government andother institutions could conduct studies to determine the best method toconserve scallop populations that were being used for collection of spat sincethe populations had; (a) a high genetic variability; (b) maintained a largepopulation over time; and (c) had a high recovery potential.
Establishment of the reserve included implementation of a large program tocollect seed to re-populate over-exploited ‘‘Management areas’’ in the artisanfishery and to meet the demands of the commercial scallop culture industry. Apilot scale spat collection experiment was initiated with the ‘‘Conservation andProtection Branch of the La Rinconada Marine Reserve, Antofagasta, Chile’’program to collect scallop seed with a mean size greater than 20 mm shellheight. To achieve this objective it was necessary to extend the period whencollectors were in the water compared to previous experiments since growthrate of juvenile scallops in the collectors was found to be 143 lm/day whenwater temperatures were 15.3 �C (Cantillanez 2000; Cantillanez et al. 2001).The present study presents results of work when purple scallop spat collectorswere held in the water for periods of four and five months in order to producejuveniles greater than 20 mm shell height for restocking programme.
Methods
Study area
The study area was in the La Rinconada part of Antofagasta Bay (23�28¢ S;70�30¢ W), about 20 km N of the city of Antofagasta, Chile (Figure 1). It has
268

an area of about 271 ha and has one of the most important populations ofArgopecten purpuratus in Chile (Avendano and Cantillanez 1996). Two distinctzones have been identified in the area, one with a high density of scallops with amean of about 11 scallopsÆm2 the other, a more peripheral area, with a meandensity of about 3 scallopsÆm2 (Avendano et al. 2000). Scallop populations inthis bay are in a continuous breeding condition but the peak breeding period isbetween September and April of the following year (Avendano and Le Pennec1996, 1997; Cantillanez et al. 2001).
Recent studies have shown that themarine reserve area is subject to alternatingflows of current that produce a net northward flow that is reflected by the northshore, producing cyclic reversals every few days. This permits a high degree ofinterchange and mixing of water masses and also a high degree of retention ofparticulate matter within a radius of 5 km inside the bay. This is conducive toretention of A. purpuratus larvae throughout the year and permits continuousspat settlement on collectors although there can be high seasonal and interannualvariability (Avendano et al. 2000; Cantillanez 2000; Cantillanez et al. 2001).
Plankton sampling
In previous studies in La Rinconada Marine Reserve we detected naturalspawnings of A. purpuratus every year from September to April (Cantillanez
Figure 1. Geographic location of the study area in the La Rinconada Marine Reserve, Anto-
fagasta, Chile.
269

2000; Cantillanez et al. 2005). Reproduction of this scallop population wasstudied by the authors during the last 15 years and plankton sampling and spatcollection was programmed according to previous results.
Quantitative plankton sampling for A. purpuratus larvae was carried outafter intense spawnings were detected between October 2001 and April 2002.Collections were made throughout the water column to a depth of 17 m at astation identified by Cantillanez et al. (2001) as being an important seed col-lecting area. Vertical hauls were made with a plankton net having a mesh of53 lm. Samples were fixed in 70% ethanol, well mixed and split into ten rep-licate subsamples in the laboratory using a ten-cell plankton separator. Larvaein two of the 1/10 subsamples were counted and larval length measured usingan ocular micrometer (Le Pennec 1978). Total number of larvae per m3 wascalculated from the average number of larvae found in the two samples ·10and the volume (V) of water filtered by the plankton net where:
V ¼ pr2h ð2Þwhere r=radius of the mouth of the net (0.125 m) and h=depth of the watercolumn sampled.
Spat collection
When large numbers of umbone larvae (>170 lm) were observed at thesampling station indicating the potential of a significant spatfall (Cantillanezet al. 2001), groups of spat collectors were placed out at the same station usinga longline system (Thouzeau 1991a). Sometimes collectors were not submergedafter plankton sampling because there were collectors installed some weekspreviously. Groups of 200 collectors were set out in October and Novemberand 150 in December 2001. In 2002, 100 collectors were set out in January andMarch and 200 in April. All collectors were within 1–5 m above the bottomsince larvae settle at deeper levels in the water column (Illanes et al. 1985;Avendano and Cantillanez 1992). In order to estimate spatfall in the collectorsafter one month of exposure, three collectors from each group were collectedfrom 1 m above the bottom. Each collector was cleaned carefully in the lab-oratory to recover all settled spat. Each sample was well mixed and placed inthe ten-cell plankton separator. Scallop spat from two sub samples werecounted and shell height measured with the aid of a stereoscopic microscopefitted with an ocular micrometer. A mean value for the number of scallop spatper collector was obtained for the three collectors of each group.
Groups of collectors were retrieved after four or five months of submer-gence. Three collectors from each group that were 1 m above the bottom weretaken to the laboratory and all the spat in each collector counted. All seed fromone collector from each group were measured for shell height using callipersaccurate to 0.01 mm.
270

Histograms of the size distribution of spat from collectors exposed for onemonth as well as from collectors exposed for extended periods were analysedfor identification of cohorts using the MIX 3.1a program (MacDonald andPitcher 1979). This was done to determine any relationship between initialsettlement of spat and the presence of larvae >170 lm at the time the collectorswere immersed and also between numbers of spat in collectors after one monthcompared to numbers after 4–5 months.
Measurement of physical and chemical parameters in the area
Bottom water temperature was measured using data loggers (VEMCO, TRmodel) installed at a depth of 16 m that recorded the temperature every 6 hduring the entire period. Monthly measurements of water samples from asimilar depth obtained with a Niskin sampler determined chlorophyll ‘‘a’’(Chla) and pheo-pigments levels using the methods of Strickland and Parsons(1968) and particulate organic matter (POM) levels using the method ofChauvaud and Thouzeau (1995).
Data analysis
Dendrogram and non-metric multidimensional scaling (NMDS) were appliedto data matrices of all measurements including mean size of larvae when col-lectors were deployed, spat in collectors after one month, daily averages oftemperature and trophic parameters recorded during each period. The datawere first transformed using the double-root (data)1/4.
Results
Results of vertical plankton hauls showed that scallop larvae were present inthe water column throughout the entire study period (Table 1). High larvaldensities were found throughout the period of highest reproductive activity ofA. purpuratus at La Rinconada and were often over 1000 larvae per m3.Highest densities were 21,700 larvae per m3 in December 2001 and 29,802larvae per m3 in March 2002. Large numbers of umbone larvae (>170 lm)were present in all months, exceeding 5000 m)3 throughout the study periodexcept for February and April 2002.
Table 2 shows that collectors submerged for one month had spat that variedin size from 277 lm to 4.7 mm. The number of cohorts identified showed arelationship between umbone larval numbers in the water column prior toinstallation of the collectors and spatfall (Figure 2).
The 950 collectors placed in the water between October 2001 and April 2002and retrieved between March and August 2002 had a total of 785,500 juvenile
271
scallops with a mean size ranging from 8.2 to 23.4 mm shell height that werethen seeded in the Reserve (Table 3). Individual cohorts identified for eachgroup of collectors that were submerged for 4–5 months are shown in Figure 3.Some of these collectors showed the presence of new cohorts compared tothose observed in collectors after one month of immersion. Mean shell heightreached by the first cohort that settled (C1) in each group of collectors (im-mersed for 4–5 months) varied between 7.67 (2102 individuals) and 30.08 mm(167 individuals).
The percent recovery of juveniles from collectors immersed for 4–5 monthscompared with numbers of spat in collectors after one month varied from 5.5%to 59.4%. The lower percentage of recovery was from collectors installed inOctober 2001 that had an initial settlement of 15,340 spat per collector. Fivemonths later the mean recovery was only 837 individuals per collector. Incontrast, a high recovery of 59.4% was observed in collectors placed out inNovember 2001 that had an initial set of 890 spat per collector and a recoveryafter 4 months of 529 juveniles. However, the presence of two other cohortswas identified in these collectors in addition to the one present after the firstmonth. In the present study growth of the different cohorts were not analysed
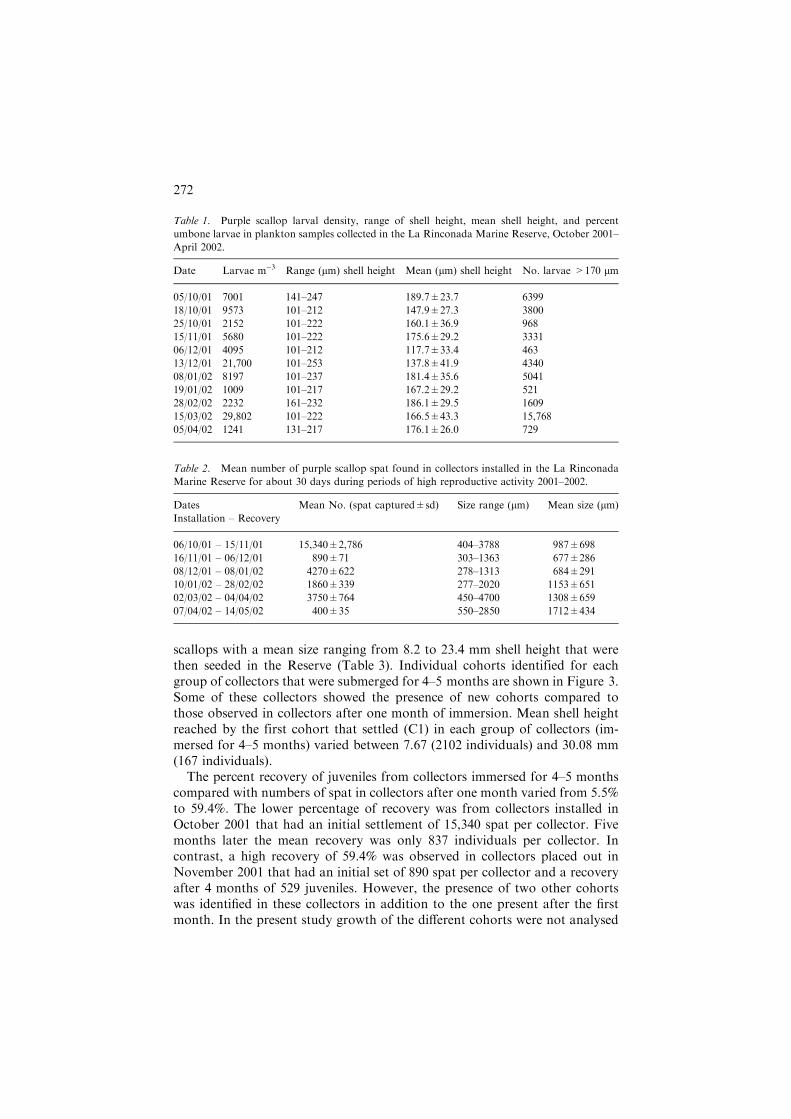
Table 1. Purple scallop larval density, range of shell height, mean shell height, and percent
umbone larvae in plankton samples collected in the La Rinconada Marine Reserve, October 2001–
April 2002.
Date Larvae m)3 Range (lm) shell height Mean (lm) shell height No. larvae >170 lm
05/10/01 7001 141–247 189.7±23.7 6399
18/10/01 9573 101–212 147.9±27.3 3800
25/10/01 2152 101–222 160.1±36.9 968
15/11/01 5680 101–222 175.6±29.2 3331
06/12/01 4095 101–212 117.7±33.4 463
13/12/01 21,700 101–253 137.8±41.9 4340
08/01/02 8197 101–237 181.4±35.6 5041
19/01/02 1009 101–217 167.2±29.2 521
28/02/02 2232 161–232 186.1±29.5 1609
15/03/02 29,802 101–222 166.5±43.3 15,768
05/04/02 1241 131–217 176.1±26.0 729
Table 2. Mean number of purple scallop spat found in collectors installed in the La Rinconada
Marine Reserve for about 30 days during periods of high reproductive activity 2001–2002.
Dates Mean No. (spat captured±sd) Size range (lm) Mean size (lm)
Installation – Recovery
06/10/01 – 15/11/01 15,340±2,786 404–3788 987±698
16/11/01 – 06/12/01 890±71 303–1363 677±286
08/12/01 – 08/01/02 4270±622 278–1313 684±291
10/01/02 – 28/02/02 1860±339 277–2020 1153±651
02/03/02 – 04/04/02 3750±764 450–4700 1308±659
07/04/02 – 14/05/02 400±35 550–2850 1712±434
272
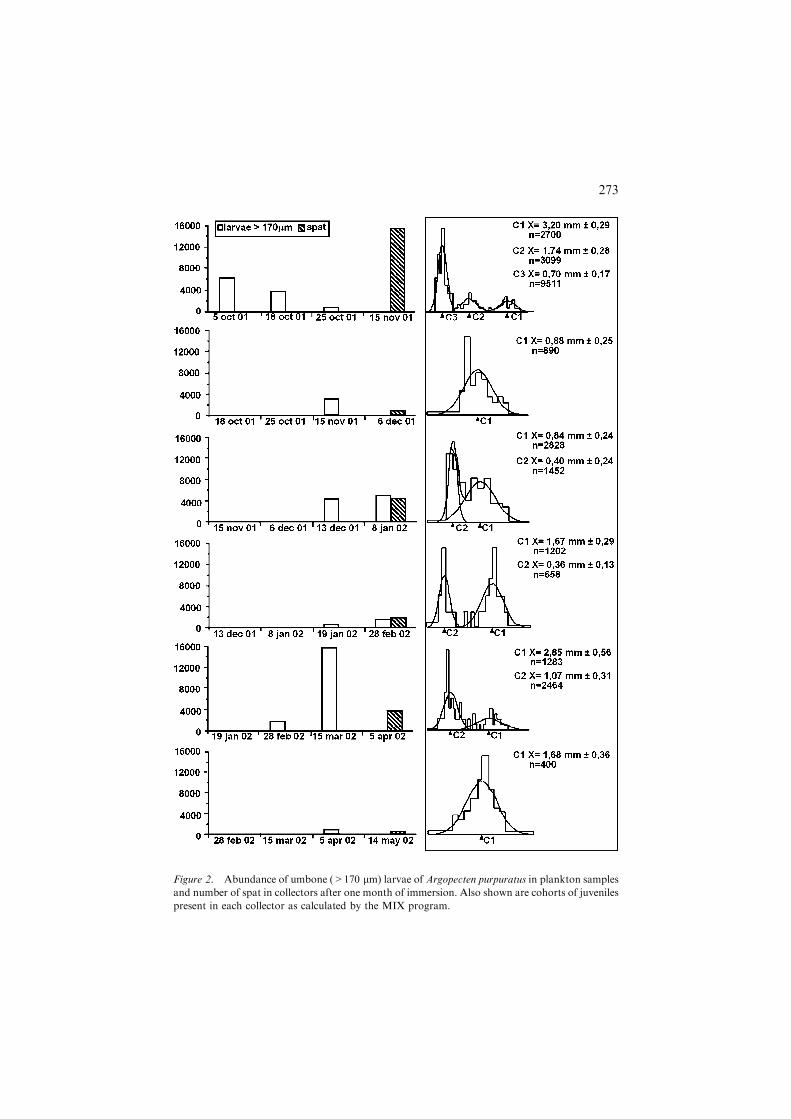
Figure 2. Abundance of umbone (>170 lm) larvae of Argopecten purpuratus in plankton samples
and number of spat in collectors after one month of immersion. Also shown are cohorts of juveniles
present in each collector as calculated by the MIX program.
273

in detail because we tried to identify the cohort in collectors after one month ofimmersion and evaluate the presence of scallop spat after 4–5 months. Highestrecovery occurred in collectors placed out in December 2001, where initially4270 spat belonging to two cohorts occurred, resulting in 2102 juveniles(49.2%) that were recovered in 4 months from a single cohort.
Table 3. Numbers and shell height of A. purpuratus spat found in collectors in the La Rinconada
Marine Reserve during periods of maximum reproductive activity 2001–2002.
Dates Immersion
time (days)
No. of
collector
Mean spat
per collector
Total
capture
Size
range (mm)
Mean
size (mm)Installation – Recovery
06/10/01 – 14/03/02 159 200 837±390 167,400 2.2–35.7 20.6±5.6
16/11/01 – 02/04/02 137 200 529±91 105,800 10.5–36.4 23.4±6.1
08/12/01 – 02/04/02 115 150 2102±732 315,300 3.3–17.2 8.2±2.9
10/01/02 – 03/06/02 144 100 979±211 97,900 8.0–33.1 19.7±6.8
02/03/02 – 07/07/02 127 100 543±88 54,300 4.6–31.0 20.3±6.5
07/04/02 – 23/08/02 138 200 224±79 44,800 6.4–26.8 17.0±5.3
Figure 3. Distribution of shell height of A. purpuratus spat in collectors that were immersed for
4–5 months at the La Riconada Marine Reserve, Antofagasta, Chile.
274

Figure 4 shows environmental parameters recorded during the study period.Bottom water temperature (16 m) showed significant daily variations (Fig-ure 4a) with extremes ranging from 12.9 to 18 �C that occurred during thesummer months (Dec–Mar). Trophic parameters (Figure 4b) that were mea-sured concurrently with water temperatures showed the highest values for Chlain the spring and fall months, varying between 2.47 and 8.89 lg l)1. Duringsummer and winter these concentrations varied between 1.08 and 4.69 lg l)1.The pheo-pigments varied between 0 and 3.5 lg l)1 in spring and summer andbetween 1.15 and 2.84 lg l)1 in fall and winter months. Concentration of POMranged from 1.2 to 6.8 mg l)1.
Figure 4. Physical and chemical parameters of water at a depth of 16 m in the La Rinconada
Marine Reserve, Antofagasta, Chile, between October 2001 and August 2002. (a) water tempera-
ture; (b) trophic parameters (CL=chlorophyll a; POM=particulate organic matter; PH=pheo-
pigments).
275

Results of the dendrogram analysis are shown in Figure 5a. The only rela-tionship found was between number of larvae when collectors were immersedand number of spat one month later. There was no relationship betweennumbers of larvae in the water and spatfall with water temperature or other
Figure 5. Dendrogram analysis (a) and non-metric multidimensional scaling (nMDS) (b), applied
to measurements taken in the present study (LT=total larvae; LU=umbone larvae; PL=post
larvae; CL=chlorophyll a; POM=particulate organic matter; PH=pheo-pigments; and T=water
temperature).
276

trophic parameters. This same relationship was also shown by multivariateanalysis (NMDS) where the best relationship occurred between numbers ofspat and umbone larvae (Figure 5b).
Discussion
The continuous presence of A. purpuratus larvae found in the Marine Reservearea with high densities during October 2001 and March 2002 confirms earlierobservations by Avendano and Le Pennec (1996, 1997) and Cantillanez (2000).These authors reported that in addition to finding a continuous degree ofreproductive activity in this scallop population, intensive spawning generallyoccurred between September and April of the following year. A prolongedreproductive period was also reported by DiSalvo et al. (1984) for otherpopulations of this species in Chile (30�S). In the Mediterranean, Pena et al.(1996) found that Aequipecten opercularis (L. 1758), spawned throughout theyear, with peaks in April and December. However, other species, such asMimachlamys varia (L. 1758) and Pecten jacobaeus (L. 1758) showed restrictedreproductive and spawning cycles, the former spawning in April and June andthe latter between February and May. In spring, spawning of the three speciesoccurred concurrently and larvae and spat may have competed for space andfood.
Results from collectors immersed for one month confirmed that A. purpu-ratus larvae were retained in the marine reserve area (Avendano et al. 2000;Cantillanez 2000; Cantillanez et al. 2001). It also showed that major spat set-tlement occurred after the highest reproductive activity in October (Pereiraet al. 1989) and there was a relationship between the number of larvae presentwhen the collectors were immersed and settlement one month later. Thisrelationship was most evident between the presence of umbone larvae andspatfall in collectors immersed in October 2001 and January and March 2002when numbers of umbone larvae exceeded 5000 larvae m)3. A relationshipbetween numbers of umbone larvae and spatfall in collectors in this bay wasalso reported by Cantillanez (2000). Ventilla (1982) reported that 100–200umbone larvae m)3 in waters of Mutsu Bay, Japan generally produced 1000Patinopecten yessoensis spat per collector while the presence of 1000 lar-vae m)3 produced 30,000–60,000 spat per collector. Such high densities of spatwere not observed in the present study even when larval densities of15,000 larvae m)3 occurred in the water.
A lack of a consistent relationship between abundance of larval cohorts andspat settlement was described by Boucher (1985) for Pecten maximus L. 1758.He postulated that the lack of settlement of many larval cohorts was due tohigh mortality that was independent of larval density. Thouzeau (1989, 1991b,c, 1995) suggested that abundance of post-metamorphic stages of this specieswas the result of larval survival that was regulated principally by water cir-culation, water temperature and trophic quality of the water.
277

The effect of water circulation on distribution of A. purpuratus larvae in themarine reserve was observed by Cantillanez (2000) who found that the pre-dominant currents tended to concentrate the larvae in the central area of thereserve but an unusual change in direction of the prevailing S-SW wind couldmove larvae from the usual area of settlement to other areas. The prevailingwind in the study area causes upwelling near the coast that sustains highproductivity throughout the year (Guillen 1983; Rodrıguez et al. 1991; Marınet al. 1993; Escribano et al. 1995; Cantillanez et al. 2001). This high produc-tivity that has Chla levels ranging from 1.08 to 8.89 lg l)1 suggests that food isnot a limiting factor for larval survival (Thouzeau 1991b, c), and is probablythe explanation for a lack of correlation between trophic parameters and thepresence of larvae and post-larvae in the present study. It should be noted thatthe above author worked in areas of low productivity where Chla varied from0.2 to 1.0 lg l)1. In the present study no relationship was found between thelife stages of A. purpuratus and water temperature although a previous study byCantillanez (2000) in the same area showed that temperature had a stronginfluence on larval growth and growth rates of cohorts ranged from 4.29 to7.3 lm day)1 in water temperatures that ranged from 13.2 to 19 �C at a depthof 16 m.
Based of results by Cantillanez (2000), the predominance of water temper-atures lower than 15.5 �C in the present study suggests that each larval cohortwould have required 30 days to reach settlement and this would explain therelationship of the number of larval cohorts present and the number of post-larval cohorts in the collectors after one month.
Declines in the number of juveniles recovered compared to initial levels ofspat settlement observed after one month may be related to settlement densityand growth of the spat acting independently or synergistically to decrease theviable number of juveniles within the collectors over prolonged periods. Forexamples, the high density of initial settlement in October 2001 (15,340 percollector) and the prolonged period of time (5 months), together with growthof juveniles during this period could have generated a high degree of intra-specific competition for space and food producing high mortalities (Mason1983). Initial cohorts that settled in collectors in this group were reduced from2700 to 375 spat in cohort 1, from 3099 to 427 individuals in cohort 2 and from9511 to 35 individuals in cohort 3 with respective mean shell heights per cohortof 24.5 mm, 18 mm and 8.5 mm. The reverse situation, low levels of initialsettlement produced high recovery of larger sized juveniles, e.g. in November2001 (890 spat per collector) and January 2002 (1860 spat per collector) hadrecoveries of 59.4% and 52.6% of juveniles after 4 and 5 months of immersion.However, at the end of the immersion period the group of collectors installed inNovember had new cohorts that were not present after the first month ofimmersion and numbers of the first cohort were reduced from 890 to 167 (meansize 30.1 mm). Similar results were observed for other cohorts. Shorter periodsof immersion favoured survival of spat in collectors although the seed recov-ered was smaller.
278

Collectors immersed for 4–5 months in a breeding area of Aequipecten teh-uelchus d’Orbigny 1846 in the Gulf of San Matias, Argentina, had settlement of400–1600 spat per collector, with shell heights ranging from 10.1 to 11.0 mm(Narvarte 2001). She suggested that differences in growth of spat of this speciesin the collectors was due to the small size of the mesh of the exterior bag thatfailed to allow a sufficient flow of water thus impeding growth. Various otherauthors have suggested that prolonged immersion of collectors used to collectspat of Placopecten magellanicus Gmelin 1791, P. maximus, and P. yessoensisresulted in clogging of the mesh with sediment and organic matter that inter-feres with settlement and survival (Yamamoto 1964; Gruffydd and Beaumont1972; Gruffydd et al. 1975; Minchin 1976; Paul 1978; Naidu and Scaplen 1979;Brand et al. 1980; Fegan 1983; Thouzeau 1989; 1991a, b; Chauvaud et al.1996). Fegan (1983) stated that clogging of collectors by fouling limited thecapture efficiency of collectors after 15 days immersion while Wilson (1987)found no settlement of P. maximus in collectors after one month’s immersion.In the present study, settlement of new cohorts of A. purpuratus continued tooccur in collectors after one month’s immersion and this confirmed similarprevious observations by Cantillanez (2000). Roman and Cano (1987) alsoobserved increasing spatfall with extended immersion time of collectors.Continued spat settlement after the first month of immersion suggests that thepercent recovery obtained in the present study was less than recorded for initialsettlement and suggests significant mortalities occurred after the first month.Slow growth of the cohort in collectors installed in December and recovered inApril (mean 7.7 mm shell height) could have been due to clogging of thecollectors and by high spat density (2102 spat per collector) that prevented anadequate flow of water through the collectors.
Although large mortalities of juveniles occurred in the collectors held for longperiods, yields of about 200–2000 spat per collector allowed us to distribute 0.75million juveniles scallops into the reserve in 2002. Results of spatfall in thisstudy were comparable with those reported for Bahia Independencia, Peru(Table 4) but greater than those reported for Tongoy Bay, Chile.
Table 4. Comparison of the amount of spatfall of A. purpuratus in the present study with results
of scallop spatfall in other studies.
Place Time of
immersion (months)
No. of spat
per bag
Authors
La Rinconada, Chile 4–5 224–2102 This study
Bahıa. Independencia, Peru 2 1580 Ysla and Vargas (1989)
Bahıa. Independencia, Peru 2 1428 Bandin and Mendo (1999)
Bahıa. Tongoy, Chile ? 40.8 Akaboshi and Illanes (1983)
Bahıa. Tongoy, Chile ? 130–580 Illanes et al. (1985)
Bahıa. Tongoy, Chile ? 230–280 Pereira et al. (1989)
Bahıa. Mejillones, Chile 2 140–155 Avendano and Cantillanez
(1989, 1992)
Captures comercial, Chile ? 25–180 Avila et al. (1994)
279

In recent years A. purpuratus culture has been based primarily on collectionof juveniles from the natural environment, often the larvae originated fromculture operations. However, there is little information on the number andsizes of wild-captured juveniles used by the industry. Some juveniles are pro-duced in hatcheries but survival of settled spat through culture is only about2–6% (Abarca 2001). Development of techniques to capture juveniles from theopen environment has permitted a three-fold expansion in production of cul-tured scallops (Stotz and Mendo 2001).
Results of this study show that A. purpuratus in this area has an extensivereproductive period that may be termed a ‘‘stepping or dribble’’ effect and thisis different from the generally brief and discrete spawning period of otherspecies. When environmental conditions are favourable the latter strategy canproduce a large cohort of spat but under unfavourable conditions few andperhaps no spat may be produced (Paulet et al. 1988; Paulet 1990). The formerstrategy, stepping effect, was previously observed by Cantillanez et al. (2001)who found larvae in the plankton and spatfall throughout the year. Resultsshow that the Marine Reserve could be used as a major source to collectjuvenile scallops for culture operations or to re-populate overexploited areas inChile.
It should be emphasized that to reduce mortalities when juveniles are used tore-populate areas they should be at least 30 mm shell height as recommendedby Ventilla (1982) and Brand (1991). The objective of the pilot programme wasto produce scallop spat for re-populating some areas. There are no previous re-population experiments in Chile and in the present study, average juvenile shellheight in collectors after 4–5 months immersion was only about 20 mm. It isnot recommended to leave collectors in the ocean for extended periods.Juveniles should be harvested prior to three months immersion (Avendano andCantillanez 1989) and re-distributed in pearl nets for intermediate culture untilthe juveniles attain an appropriate size for out planting (Avendano et al. 2001).
Acknowledgements
The present study was developed within the framework of FNDR Project codeBIP No. 20127869-0. Thanks are due to Dr. Neil Bourne for constructivecomments and for improving the English on the manuscript.
References
Abarca A. 2001. Scallop hatcheries of Argopecten purpuratus (Lamarck, 1819) in Chile: a survey of
the present situation. 13th International Pectinid Workshop, Coquimbo Chile, April 18–24, 2001,
pp. 74–75.
Akaboshi S. and Illanes J.E. 1983. Estudio experimental sobre la captacion, pre-cultivo y cultivo,
en ambiente natural de Chlamys (Argopecten) purpurata, Lamarck 1819, en Bahıa Tongoy, IV
Region, Coquimbo. Symposium Internacional de Acuacultura, Coquimbo-Chile, pp. 233–254
Septiembre 1983.
280

Avendano M. 1984. Una tecnica para el cultivo de Argopecten purpuratus (Lamarck, 1819) en la
Bahıa de Mejillones del Sur. Chile. Rev. Latinoam. Acuicult. 22: 18–22.
Avendano M. 1993. Donnees sur la biologie de Argopecten purpuratus (Lamarck, 1819), Mollusque
Bivalve du Chili. These de doctorat, Universite de Bretagne Occidentale, 167 pp.
Avendano M. and Cantillanez M. 1989. Observaciones sobre captacion de semillas de Argopecten
purpuratus (Lamarck, 1819) en la Bahıa de Mejillones del Sur. Chile. Estud. Oceanol. 8: 51–59.
Avendano M. and Cantillanez M. 1992. Colecta artificial de semilla de A. purpuratus (Lamarck,
1819) en la Bahıa de Mejillones, Chile. II.- Observaciones sobre niveles optimos de captacion.
Estud. Oceanol. 11: 39–43.
Avendano M. and Cantillanez M. 1996. Efecto de la pesca clandestina, sobre Argopecten purpu-
ratus (Lamarck, 1819), en el banco de La Rinconada, II Region. Cienc. Tec. Mar. CONA. 19:
57–65.
Avendano M. and Cantillanez M. 1997. Necesidad de crear una reserva marina en el banco de
ostiones de La Rinconada – II Region. Estud. Oceanol. 16: 109–113.
Avendano M. and Cantillanez M. 2003. Population estimates, extraction, and translocation of the
pectinid Argopecten purpuratus within Mejillones bay, Chile. Sci. Mar. 67(3): 285–292.
Avendano M., Cantillanez M., Rodrıguez L., Zuniga O., Escribano R. and Oliva, M. 2000.
Conservacion y proteccion Reserva Marina La Rinconada Antofagasta-Chile. Informe Final
Proyecto FNDR Cod. BIP N� 20127869-0, 187 pp.
Avendano M., Cantillanez M., Le Pennec M., Lodeiros C. and Freites L. 2001. Cultivo de
pectınidos Iberoamericanos en suspension. In: Maeda-Martınez A.N. (ed.), Los Moluscos
Pectınidos de Iberoamerica: Ciencia y Acuicultura, Editorial Limusa, Mexico, pp. 193–211.
Avendano M. and Le Pennec M. 1996. Contribucion al conocimiento reproductivo de Argopecten
purpuratus en dos poblaciones de la II Region - Chile. Estud. Oceanol. 15: 1–10.
Avendano M. and Le Pennec M. 1997. Intraspecific variation in gametogenesis in two populations
of the Chilean mollusc bivalve, Argopecten purpuratus (Lamarck). Aquacult. Res. 28: 175–183.
Avila M., Seguel M., Plaza H., Bustos E. and Otaiza R. 1994. Estado de situacion y Perspectivas de
la acuicultura en Chile. Informe SGI-IFOP 94/1, 166 pp.
Bandin R. and Mendo J. 1999. Asentamiento larval de la concha de abanico (Argopecten purpu-
ratus) en colectores artificiales en la Bahıa Independencia, Pisco, Peru. Invest. Mar., Valparaiso
27: 3–13.
Boucher J. 1985. Caracteristiques dynamiques du cycle vital de la coquille Saint-Jacques (Pecten
maximus): hypotheses sur les stades critiques pour le recrutement. Cons. Int. Explor. Mer, C.M.
1985/K23/Sess.Q, 10 pp.
Brand A.R. 1991. Scallop ecology: Distribution and behaviour. In: Shumway S. (ed.), Scallop:
Biology, Ecology and Aquaculture, Elsevier, Amsterdam, pp. 517–583.
Brand A., Paul J. and Hoogester J. 1980. Spat settlement of scallops Chlamys opercularis (L) and
Pecten maximus (L) on artificial collectors. J. Mar. Biol. Assoc. UK 60: 379–389.
Cantillanez M. 2000. Reproduction, vie larvaire et pre-recrutement du Pectinidae Argopecten
purpuratus (Lamarck, 1819) dans la baie d’Antofagasta (Chili). These doctorat Ph. D., Univ.
Brest. France, 168 pp.
Cantillanez M., Thouzeau G. and Avendano, M. 2001. Reproductive cycle in Argopecten purpu-
ratus during El Nino and la Nina conditions: a case study in the Rinconada Bay (Chile). 13th
International Pectinid Workshop, Coquimbo Chile, April 18–24, 2001, pp. 86–88.
Chauvaud L. and Thouzeau G. 1995. Captage des post-larves d’invertebres benthiques en relation
avec la variabilite spatio-temporelle des facteurs environnementaux en rade de Brest. Actes de
colloque 3emes Rencontres Scientifiques Internationales, Programme Rade, 14–16 mars 1995,
Brest, vol. 2, pp. 29–41.
Chauvaud L., Thouzeau G. and Grall J. 1996. Experimental collection of great scallop postlarvae
and other benthic species in the Bay of Brest: settlement patterns in relation to spatio-temporal
variability of environmental factors. Aquacult. Int. 4: 263–288.
DiSalvo L.H., Alarcon E., Martınez E. and Uribe E. 1984. Progress in mass culture of Argopecten
purpuratus with notes on its natural history. Rev. Chil. Hist. Nat. 57: 33–45.
281

Escribano R., Rodrıguez L. and Irribaren C. 1995. Temporal variability of sea temperature in Bay
of Antofagasta, northern Chile (1991–1995). Estud. Oceanol. 14: 39–47.
Fegan D. 1983. Scallop spat collection in the Obvan area, 1982. 4th Inter. Pectinid Workshop,
Aberdeen, May 1983, 20 pp.
Gajardo G., Coutteau P., Cure K., Sorgeloos P. and Beardmore J.A. 1996. Nutritional improve-
ment of the commercial production of marine aquaculture species through application of
innovative biotechniques. In: Gajardo G. and Coutteau P. (eds), Improvement of the Com-
mercial Production of Marine Aquaculture Species. Proceedings of a Workshop on Fish and
Mollusc Larviculture, pp. 7–12.
Guillen O. 1983. Condiciones Oceanograficas y sus fluctuaciones en el Pacıfico Sur-oriental. In:
Sharp G.D. and Csirke J. (eds), Actas de la consulta de expertos para examinar cambios en la
abundancia y composicion por especies de recursos de peces nerıticos, San Jose, Costa Rica,
1983. FAO inf. Pesca (291) 3: 607–658.
Gruffydd L.D. and Beaumont A. 1972. A method for rearing Pecten maximus larvae in the lab-
oratory. Mar. Biol. 15: 350–355.
Gruffydd L., Lane W. and Beaumont A. 1975. The glands of the larval foot in Pecten maximus (L.),
and possible homologues in other bivalves. J. Mar. Biol. Assoc. UK 55: 463–476.
Hojas F. 1982. Antecedentes Sobre El Cultivo Del Ostion del Norte, En Antofagasta, Chile. Chile
Pesquero, Marzo 1982, pp. 10–12.
Illanes J.E. 1986. Situacion actual del cultivo del ostion Chlamys (Argopecten) purpurata y ostra
(Crassostrea gigas) en el norte de Chile. En: La Pesca en Chile. Valparaiso, (Chile). Univ.
Catolica de Valparaiso, pp. 305–314.
Illanes J.E., Akaboshi S. and Uribe E. 1985. Efectos de la Temperatura en la reproduccion del
Ostion del Norte Argopecten purpuratus en la Bahıa de Tongoy durante el Fenomeno El Nino.
Invest. Pesq. (Chile). 32: 167–173.
Le Pennec M. 1978. Genese de la coquille larvaire et postlarvaire chez divers Bivalves marins. These
Doctorat Sciences Naturelles, Brest, 229 pp. 108 pl.
Le Pennec M. 1997. Les ecloseries de Mollusques Bivalves: mode d’emploi. Bull. Aquacult. Assoc.
Can. 97(3): 31–37.
MacDonald P.D.M. and Pitcher T.J. 1979. Age-groups from size-frequency data: a versatile and
efficient method of analyzing distribution mixtures. J. Fish. Res. Board Can. 36: 987–1001.
Marın V., Rodrıguez L., Vallejo L., Fuenteseca J. and Oyarce E. 1993. Efectos de la surgencia
costera sobre la productividad primaria primaveral de Bahia Mejillones del Sur (Antofagasta,
Chile). Rev. Chile. Hist. Nat. 66: 479–491.
Mason J. 1983. Scallop and queen fisheries in the British Isles. Fishing News Books Ltd, England,
p. 147.
Minchin D. 1976. Pectinid Settlement. Scallop Workshop. Baltimore, Ireland 11–16 May 1976.
Naidu K.S. and Scaplen R. 1979. Settlement and survival of giant scallop Placopecten magellanicus
larvae, on enclosed polyethylene film collectors. In: Pillay T.V.R and Dill W.A. (eds.), Advances
in Aquaculture, Fishing news (Books) Ltd., England, pp. 379–381.
Narvarte M.A. 2001. Settlement of tehuelche scallop, Aequipecten tehuelchus D’Orb., larvae on
artificial substrata in San Matıas Gulf (Patagonia, Argentina). Aquaculture 196: 55–65.
Paul J.D. 1978. Settlement and growth of scallops on collectors. 2nd Intern. Pectinid Workshop,
Brest.
Paulet Y.M. 1990. Role de la reproduction dans le determinisme du recrutement chez Pecten
maximus (L) de la baie de Saint-Brieuc. These de Doctorat, Universite de Bretagne Occidentale,
Brest France, 194 pp.
Paulet Y.M., Lucas A. and Gerard A. 1988. Reproduction and larval development in two Pecten
maximus (L) populations from Brittany. J. Exp. Mar. Biol. Ecol. 119: 145–156.
Pena J.B., Canales J., Adsuara J.M. and Sos M.A. 1996. Study of seasonal settlements of five
scallop species in the western Mediterranean. Aquacult. Int. 4: 253–261.
282

Pereira L., Illanes J.E. and Akaboshi S. 1989. Captacion de semilla en ambiente natural del ostion
del norte Argopecten purpuratus en Bahıa Tongoy IV Region, Chile. Anales Cientıficos UNALM
(Lima, Peru), Vol Extr. Memorias del 2� COLACMAR, 17–21 agosto 1987: 281–292.
Rodrıguez L., Marın V., Farıas M. and Oyarce E. 1991. Identification of an upwelling zone by
remote sensing and in situ measurements. Mejillones del Sur Bay (Antofagasta-Chile). Sci. Mar.
55(3): 467–473.
Roman G. and Cano J. 1987. Pectinid settlement on collectors in Malaga, S.E. Spain, in 1985. Sixth
International Pectinid Workshop. Menai- Bridge, Wales, 9–14 April 1987, 32 pp.
Stotz W.B. and Gonzalez S.A. 1997. Abundance, growth, and production of sea scallop Argopecten
purpuratus (Lamarck, 1819): bases for sustainable exploitation of natural scallop beds in north-
central Chile. Fish. Res. 32: 173–183.
Stotz W.B. and Mendo J. 2001. Pesquerıa, repoblamiento y manejo de bancos naturales de
pectınidos en Iberoamerica: su interaccion con la acuicultura. In: Maeda-Martınez A.N. (ed.),
Los Moluscos Pectınidos de Iberoamerica: Ciencia y Acuicultura, Editorial Limusa, Mexico,
pp. 357–374.
Strickland J.D.H. and Parsons T.R. 1968. A practical handbook of seawater analysis. Fish. Res.
Board Can., Bull. 167: 1–310.
Thouzeau G. 1989. Determinisme du pre-recrutement de Pecten maximus (L.) en baie de Saint-
Brieuc. These de Doctorat d’Universite., Univ. Bretagne Occidentale, Brest, 545 pp.
Thouzeau G. 1991a. Experimental collection of postlarvae of Peceten maximus (L.) and other
benthic macrofaunal species in the Bay of Saint-Brieuc, France. I Settlement patterns and biotic
interaction among the species collected. J. Exp. Mar. Biol. Ecol. 148: 159–179.
Thouzeau G. 1991b. Experimental collection of postlarvae of Peceten maximus (L.) and other
benthic macrofaunal species in the Bay of Saint-Brieuc, France. II Reproduction patterns and
postlarval growth of five mollusk species settlement patterns and biotic interaction among the
species collected. J. Exp. Mar. Biol. Ecol. 148: 181–200.
Thouzeau G. 1991c. Determinisme du pre-recrutement de Pecten maximus (L.) en baie de Saint-
Brieuc: processus regulateurs de l’abondance, de la survie et de la croissance des post-larves et
juveniles. Aquat. Living Resour. 4: 77–99.
Thouzeau G. 1995. Aspects de la dynamique spatio-temporelle du pre –recrutement de Pecten
maximus L., en baie de Saint-Brieuc. ICES Mar. Sci. Symp. 199: 31–39.
Ventilla R.F. 1982. The scallop industry in Japan. Adv. Mar. Biol. 20: 310–380.
Wilson J. 1987. Reproduction of Pecten maximus (L) in Kikiesan bay, Co. Galway, Ireland. 6th
Inter. Pectinid Workshop, Menai-Bridge.
Wolff M. and Alarcon E. 1993. Structure of scallop Argopecten purpuratus (Lamarck, 1819)
dominated subtidal macro-invertebrate assemblage in Northern Chile. J. Shellfish Res. 2:
295–304.
Yamamoto G. 1964. Studies on the propagations of the scallop Patinopecten yessoensis (Jay) in
Mutsu bay. Fish. Res. Board Can. Trans. Ser. 1054: 68.
Ysla L. and Vargas J. 1989. Informe sobre el proyecto de captacion de larvas de concha de abanico
(Argopecten purpuratus) en la zona de Pisco. Bol. Fac. Pesq., Univ. Nac. Agrar. Peru 28: 41–67.
283




















