
Comparison of condition indices and their seasonal variation in the freshwater clam, Egeria radiata...
11
AquacuUure and Fisheries Management 1993, 24, 603-612 Comparison of condition indices and their seasonal variation in the freshwater clam, Egeria radiata (Lamarck) (Tellinacea: Donacidae), from the Cross River, Nigeria L. ETIM & M. TAEGE Institute of Oceanography, University of Calabar, Calabar, Nigeria Abstract. From February 1987 to September 1989. the monthly variations of eight indices of condition which are commonly used in bivalve aquaculture were determined for Egeria radiata (Lam. 1804) (Bivalvia: Tellinacea: Donacidae) from the Cross River, Nigeria. The indices were compared and their suitability for determining the spawning season of Egeria radiata was assessed. In general, a fall in condition index was observed during the rainy season (June-October) and coincided exactly with minimum phytoplankton and pigment abundance. This decline in condition at the end of the wet season is as likely to be due to tissue depletion for maintenance under poor environmental condition as it is due to reproduction. The indices could be used as a quick and fast tool in assessing the spawning cycle of bivalves in the tropics. Introduction Condition has been defined, theoretically, as the 'ability of an animal to withstand an adverse environmental stress, be this physical, chemical or biological' (Bayne 1975). Within this context stress is 'a measurable alteration of a physiological, behavioural, biochemical or cytological steady-state which is induced by environmental change, and which renders the individual (or population, or community) more vulnerable to further environmental change' (Bayne 1975). In practice, condition index is a computed factor which is used in determining the condition, fatness, corpulence or well-being of aquatic animals such as finfish and bivalves. Infinfishit is usually calculated as the weight of thefishexpressed as a percentage of the cube of its length. This is the Fulton condition index, which shows the distribution of weight per unit volume of thefish(Etim, Etcheri & Umoren 1989). Clark's condition index is a variant of Fulton's and is determined as the weight of the gutted fish expressed as a percentage of the cube of its length. For bivalve molluscs, various indices of condition have been calculated by different workers. Mann (1978) selected a number of morphometric, biochemical and physiological parameters which he compared and evaluated the potential use tor computing condition indices in bivalve molluscs. A literature survey shows that a range of methods are used to study condition indices of bivalve molluscs (Mann 1978; Lucas & Beninger 1985). Some of these are difficult to determine because of the expensive and complex laboratory techniques required which are seldom available to aquaculturists or scientists in developing countries. Correspondence: Dr Lawrence Etim, Institute of Oceanography. University of Calabar, Calabar. Nigeria. 603
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparison of condition indices and their seasonal variation in the freshwater clam, Egeria radiata...

AquacuUure and Fisheries Management 1993, 24, 603-612
Comparison of condition indices and their seasonalvariation in the freshwater clam, Egeria radiata(Lamarck) (Tellinacea: Donacidae), from the CrossRiver, Nigeria
L. ETIM & M. TAEGE Institute of Oceanography, University of Calabar, Calabar,Nigeria
Abstract. From February 1987 to September 1989. the monthly variations of eight indices ofcondition which are commonly used in bivalve aquaculture were determined for Egeria radiata(Lam. 1804) (Bivalvia: Tellinacea: Donacidae) from the Cross River, Nigeria. The indices werecompared and their suitability for determining the spawning season of Egeria radiata wasassessed. In general, a fall in condition index was observed during the rainy season(June-October) and coincided exactly with minimum phytoplankton and pigment abundance.This decline in condition at the end of the wet season is as likely to be due to tissue depletion formaintenance under poor environmental condition as it is due to reproduction. The indicescould be used as a quick and fast tool in assessing the spawning cycle of bivalves in the tropics.
Introduction
Condition has been defined, theoretically, as the 'ability of an animal to withstand an adverseenvironmental stress, be this physical, chemical or biological' (Bayne 1975). Within thiscontext stress is 'a measurable alteration of a physiological, behavioural, biochemical orcytological steady-state which is induced by environmental change, and which renders theindividual (or population, or community) more vulnerable to further environmental change'(Bayne 1975). In practice, condition index is a computed factor which is used in determiningthe condition, fatness, corpulence or well-being of aquatic animals such as finfish andbivalves. In finfish it is usually calculated as the weight of the fish expressed as a percentage ofthe cube of its length. This is the Fulton condition index, which shows the distribution ofweight per unit volume of the fish (Etim, Etcheri & Umoren 1989). Clark's condition index isa variant of Fulton's and is determined as the weight of the gutted fish expressed as apercentage of the cube of its length.
For bivalve molluscs, various indices of condition have been calculated by differentworkers. Mann (1978) selected a number of morphometric, biochemical and physiologicalparameters which he compared and evaluated the potential use tor computing conditionindices in bivalve molluscs.
A literature survey shows that a range of methods are used to study condition indices ofbivalve molluscs (Mann 1978; Lucas & Beninger 1985). Some of these are difficult todetermine because of the expensive and complex laboratory techniques required which areseldom available to aquaculturists or scientists in developing countries.
Correspondence: Dr Lawrence Etim, Institute of Oceanography. University of Calabar, Calabar. Nigeria.
603

604 L. Etim & M. Taege
This paper describes some easily determined indices of condition and their suitability fordescribing the condition of Egeria radiata (Lamarck) in the Cross River, Nigeria.
Materials and methods
Monthly samples (Table 1) of E. radiata weit obtained from the Cross River (Nigeria) at ItuBridge (longitude 8° 4'E, latitude 5° l l 'N), which is about 100km from the mouth of theestuary. Clams with shell lengths beween 7-0 and 7-3cm were selected to reduce size variationto a minimum.
The clams were kept for 72 h in the laboratory aquaria containing filtered river water toenable them to remove all faeces and pseudofaeces. The aquaria water was changed dailyAfter the third day each clam was blotted dry and the following linear and morphometricdimensions determined: length, total weight, shell weight, soft tissue weight, dry tissueweight (obtained by drying the soft tissue in an oven at 60=C for 72 h), shell volume (obtainedby notmg the volume of water from a burette required to fill each of the shell valves). Britton& Morton (1982) contains more on the definitions and the methodsof determinations of thesedimensions.
The dried body tissues for each month's sample were pooled together and homogenizedto a powdery form.
For lipid determination, 4g of the powdery samples was wrapped in a filter paper andplaced in a Soxhlet apparatus and the extraction carried out for 6h using dlethyl ether. Thequantity of lipid extracted was determined gravimetrically after driving off the solvent fromthe fiask.
For protein determination, 0-5g of the homogenized sample was digested in a Kjeldahlflask using concentrated sulphuric acid. The reaction was catalysed with 8g of Kjeldahlcatalyst. The resulting nitrogen was distilled over as ammonia into 2% boric acid using amacro-Kjeldahl distillation apparatus. Sulphuric acid (OIN) was titrated against theammonia-in-boric acid solution. The titre value was converted to percentage nitrogen usingthe relationship:
% nitrogen = (^"'""^e of acid x 0-0014 x 100)weight of sample digested
The percentage nitrogen was then multiplied by the constant 6-25 to convert topercentage protein. Pearson (1975) gives more details ofthe Soxhlet and Kjeldahl processes.
Ash was determined by weighing the residue left after burning a known quantity of thehomogenized sample in a muffle furnace at 500°C for 12h (Ansell 1974; Etim 1990).
Table 1. Monthly sample size (numbers) of E. radiata obtained from the Cross River
Year
198719881989
January
2520
February
182221
March
221
April
202214
May
19
2
June
192315
July
172
13
August
191814
September
181515
October
14
November
1128
December
1919

Condition 0/Egeria 605
The following indices of conditions were then computed.
(1) Dry tissue weight x 100 DTW x 100Shell volume ^^ SV
This index has been used by Galtsoff (1964), Gabbott & Stephenson (1974), and Lutz,Inczs, Porter & Stotz (1980).
(2) Dry tissue weight X 100 DTW x 100Dry shell weight ^^ DSW
This has been used by Walne & Mann (1975) for Ostrea edulis (L.), Mann & Rhyther(1977) for six different species of bivalve and by Beninger & Lucas (1984) to monitor theseasonal variation in condition of two species of adult clams.
(3) Ash-free-dry weight x 100 AFDWor 3(Shell length)^ L
Wilde (1975) used this index in temperature and growth studies of the bivalve Macomabalthica (L.).
(4) Fresh tissue weight x 100 FTW x 100Live weight of organism LW
Nair & Nair (1987) used this to assess the quality of meat ('edibility index") of Crassostreamadrasensis (Preston).
(5) Ash-free dry weight AFDWDry shell weight °^ DSW
Walne & Mann (1975) used this during growth trials of two bivalve species while Beninger& Lucas (1984) used it to determine the seasonal variation in condition of two species of adultclam reared in a common habitat.
(6) CarbohydrateProtein -^
Mann (1978) recommended the use of this ratio as a biochemical index of condition.
(7) % organic content of flesh
This index is computed as 100 minus % ash content. It has been used by Mann (1978) asone of the biochemical indices of condition.
(8) % nitrogen free extract
The nitrogen free extract (NFE) is approximately equal to the carbohydrate content ofthe dry flesh and was determined by 'subtraction' as given by Pearson (1975), viz. % NFE =100 minus (% protein + % ash + % lipid).
Results
Eight different types of condition indices in E. radiata were chosen and their monthlyvariation monitored over a 32-month period. The data obtained are presented in Fig. 1. Each
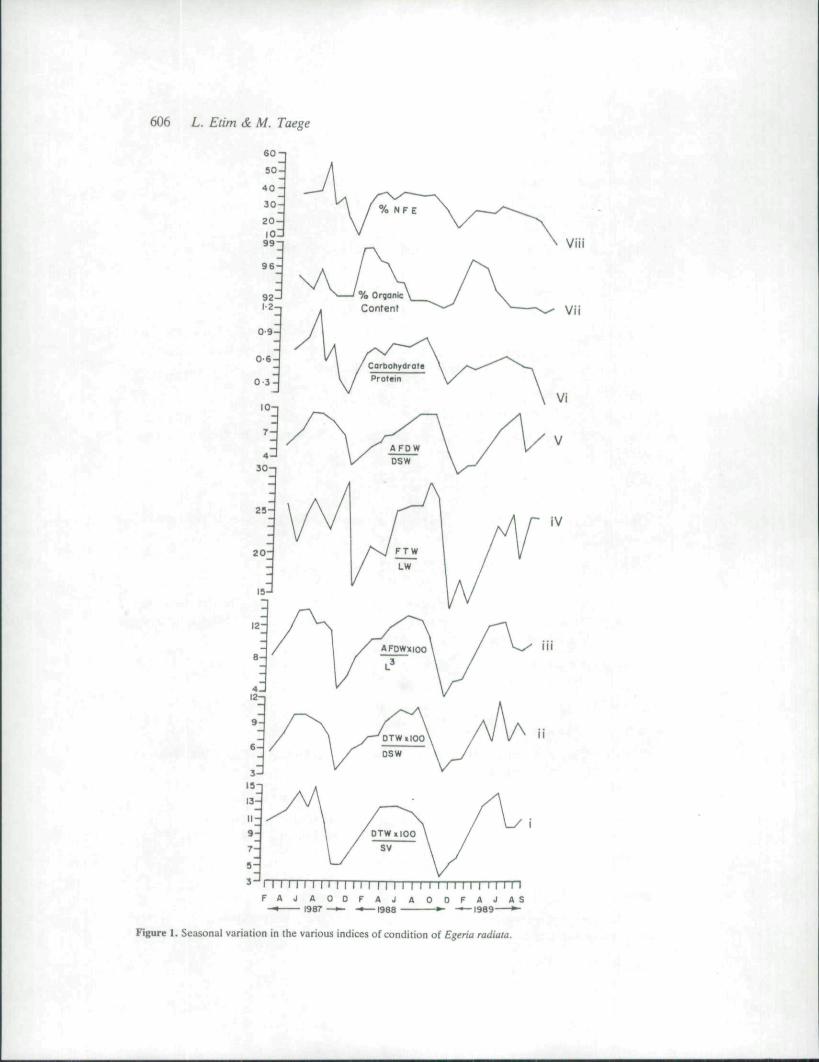
606 L. Etim & M. Taege
I I I
I I M I I 1 M I I I I I I I I I I M I I I I I I I I I I I
Figure I. Seasonal variation in Ihe various indices of condition of Egeria radiata.

Condition of Egeria 607
graph shows a pronounced seasonal cycle of maxima and minima with monthly changes inpattern and variation similar beween them. The highest values occurred between May andJune and the lowest between September and October of each year. The cycle for % organiccontent was out of phase with others.
Discussion
In aquaculture and ecology, the indices of condition in bivalves are usually determined fortwo main reasons: (1) economic reasons: in this case the index is used to designate the qualityof a marketed product; and (2) ecophysiological reasons: in this case the index is used for thepurpose of assessing the apparent health of a cultured or feral stock in order to characterizethe physiological activity (growth, reproduction, secretion, etc.) of the animal underprevailing environmental conditions (Lucas & Beninger 1985).
The characteristics of some of eight selected indices are as follows. Index 1 takes accountof the reduced tissue weight which occurs during spawning. Thus, it is suitable for use indifferentiating spawning from non-spawning periods (Mann 1978). The advantage of index 2is that both variables in the equation are determined gravimetrically, which facilitates theprocessing of large samples. This index incorporates the weight of the shell as the absoluteshell size but ignores the individual changes in shell cavity volume which might be due tovariation in overall shape or shell thickness of the animal. The numerator of index 3 does nottake into consideration the fluctuating effect of the ash content of the dry tissue. By using L̂for the denominator, the assumption is that the growth of the species in question isisometrical. This is hardly so for most species. The index also neglects any potential changesin shell thickness (Mann 1978). Index 4 is suitable for use with large samples because both thenumerator and the denominator are determined gravimetricaily. The disadvantage is that thedenominator is subject to much variation (see Dhamne & Mane 1979), thus the coefficient ofvariation of the index could be quite large. By using ash-free dry weight in index 5 instead ofthe dry weight, the fluctuation in the ash content of the dry weight is discounted. As thecarbohydrate decreases to critical levels, the animal utilizes more protein as a source ofenergy for maintenance. Hence the use of index 6 as a biochemical index. Index 8 is a usefulindex as it can show how the organism mobilizes its carbohydrate energy reserve at differentperiods during its reproductive cycle.
Several workers have incorporated shell volume in computing condition index forbivalves. The shell volume is usually determined by siniple displacement technique (Seed1973; Lutz et al. 1980). This method of measuring intervalval volume is not precise (Lucas &Beninger 1985), hence the precision of the computed index is often compromised. In thisstudy, the internal shell volume was determined by noting the volume of water from a buretterequired to fill each of the shell valves. Statistical tests (f-test: P > 0-05, d.f. = 95) showed thatE. radiata is volumetrically equivalved. The practical implication of this finding is that indetermining the volume of the specimen, a lot of time could be saved by measuring only onevalve and then multiplying the result by two.
The dry tissue weight/dry shell weight ratio is a widely used index. Shell valves can bedried by blotting them dry with a filter paper or by drying them in an oven to a constantweight. Statistically, the one is not significantly different from the other (r-test: P > 0-05, n =49). Hence the easier blotting-dry method should be preferred. For indices involving tissueweights, either the fresh tissue weight (Lutz etal. 1980), or the dry tissue weight (Walne 1970;

608 L. Etim & M. Taege
Beninger & Lucas 1984) or the ash-free dry weight (AFDW) (Walne & Mann 1975; Beninger& Lucas 1984) could be used. Lucas & Beninger (1985) stated that the index incorporatingAFDW is more accurate than the index with the dry tissue weight. The result of thisinvestigation is simiiar to that of Beninger & Lucas (1984); the pattern of variation of the twoindices is quite similar and they follow each other very closely. Therefore, because of ease ofdetermination the index incorporating dry tissue should be preferred to that involvingAFDW.
In measuring the fresh tissue weight, the mantle water must be allowed to drain from theshucked fresh tissue (Britton & Morton 1982); without this an erroneous estimate is bound tobe made of the fresh tissue weight of the organism. The shortcoming of the procedure is thatthe notion of 'drained' flesh is vague and is difficult to standardize in practice (Lucas &Beninger 1985) since the loss of fluid from the bivalve tissue is governed by so manyuncontrollable factors (Dhamne & Mane 1979). To a large extent, this probably accounts forwhy the fresh tissue weight: live weight ratio failed to exhibit a smooth curve.
The seasonal variation in dry tissue weight and condition indices of bivalves has beenshown to follow distinct pattems correlated with the sexual cycles of the organism. Forexample, the variations in the tissue and gonad weight of Chlamys opercularis (L.) have beenshown by Taylor & Venn (1979) to follow closely the reproductive cycle, with the maximumweight recorded just prior to spawning. The same pattern was found for Crassostreamadrasensis in which the condition of the fresh tissue weight was found to increase to amaximum level in October and thereafter declined to a minimum in subsequent months. Thiscoincided with the spawning period and continued until March/April when the oysters werefully spent (Nair & Nair 1987).
The seasonal variation in dry tissue weight and condition indices of the mussel, Mytilusedulis L., has been investigated (Lutz et al. 1980). A rapid fall in condition index and drytissue weight at the end of May was attributed to the release of gametes during late spawningin spring. A second decrease in condition index and tissue weight in August led Lutz et al.(1980) to infer a late summer spawning as a general feature of the population. The two typesof condition (volume of fresh tissue/volume of shell cavity, and weight of dried tissue/shellcavity volume) used by Lutz et al. (1980) exhibited closely similar patterns of variation andwere equally useful in elucidating the spawning period of M. edulis.
This seasonal variation in dry or fresh weight and condition index of bivalves in responseto annual reproductive cycles has been demonstrated for various species of bivalves by manyworkers like Gabbott & Walker (1971), Gabbott & Bayne (1973), Ansell (1974) Sundet &Vahl(1981).
A general pattern is that the condition indices of bivalve molluscs increase with the stageof sexual maturity of the species till a peak is reached, after which the condition and tissueweight fall due to spawning.
One area in which the measured condition indices will find useful application is in theelucidation of the spawning season of E. radiata. Figure 1 shows that the decline in conditionindices takes place during the rainy season, i.e. between July and October. It then followsthat the population of E. radiata under investigation spawns once in a year during the peak ofthe rainy season. This has been confirmed by Etim (1991), who elucidated the reproductivecycle of the same stock of £. radiata through histological study of the gonad and microscopicexamination of ctenidial smears. Table 2 shows the monthly changes in developmental stagesof the gonad of E. radiata.

Condition of Egeria 609
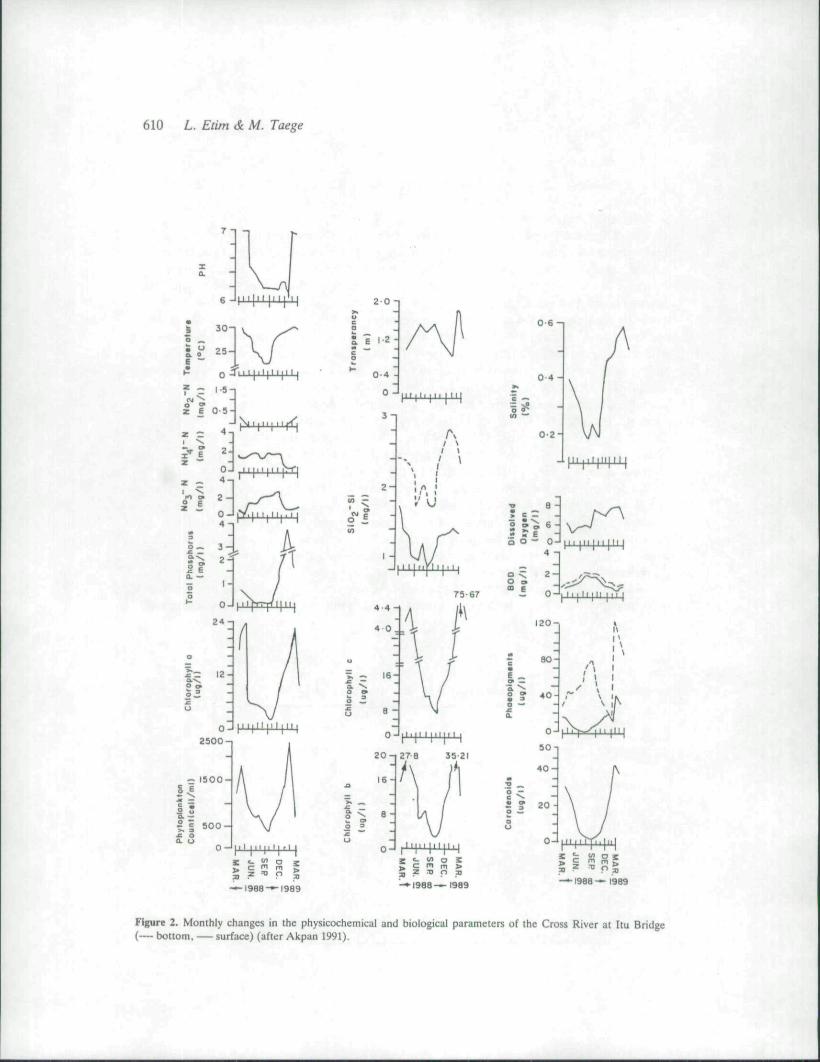
The limnology of the Cross River has been studied (Lowenberg & Kunzel 1986; Akpan1991) and its effect on the fisheries elucidated (Moses 1979, 1987). There are two climaticseasons in Nigeria — the dry season (November-March), and the rainy season (April-October) . Rainfall is one fundamental climatic factor with the most far-reaching effect on thehydrography of the river system. During the rains, an enormous amount of run-off watercarrying nutrients and debris from land flows in to the main river channel, resulting in theoverflow of the river and the flooding of the adjacent plains during the peak of the rainyseason (July-October). During this time there is an increase in water depth from 4 to 12m, alowering of conductivity, a fall in water temperature from 32°C in March to about 2O''C in July(Lowenberg & Kunzel 1987). During this period the flow is unidirectional (no longersubjected to tidal reversals), and current speed much faster (increasing from 0-6 m/s in the dryseason to 1-0 m/s in the rainy season). More detailed results of the limnological studies of theCross River have been given by Akpan (1991). The water body at Itu is verticallyhomogenous with regard to most physicochemical and biological parameters (the exceptionbeing phaeopigments, silicate and biological oxygen demand, BOD) (see Fig. 2). This figurealso shows that there is a decrease in phytoplankton biomass, temperature, salinity, totalphosphorus, NO2', dissolved oxygen pH during the rainy season. Conversely, there is anincrease in NO3", NHs"*", BOD during the rainy season.
The low pigment concentration and phytoplankton count in the river during the rains aredue to the attenuation in insolation and a reduction in nutrient concentration of the riverwater. This low nutrient concentration is mainly caused by a dilution effect from the flood andrun-off water. For a filter feeder like E. radiata, it is not surprising that low condition indexcoincides with such periods of low phytoplankton abundance. Thus, the condition of the clamis a reflection of the available food supply as well as the reproductive cycle. Consequently, thefall in condition of the clam recorded towards the end of the wet season is due to tissuedepletion for maintenance under poor environmental conditions and also to reproduction.
Table 2. Monthly changes in deveiopmeatal stages of the gonad of E. radiata
Stages of gonad development
Months Male Female
August 1988 Ripe/spawning Ripe/spawningSeptember 1988 Ripe/spawning Ripe/spawtiingOctober 1988 Spent/inactive Spent/inactiveNovember 1988 Spent/iniictive Spent/inactiveDecember 1988 Inactive InactiveJanuary 1989 . Inactive InactiveFebruary 1989 Immature ImmatureMarch 1989 Immature ImmatureApril 1989 Immature ImmatureMay 1989 Immature/ripe Immature/ripeJune 1989 Ripe/spawning Ripe/spawningJuly 1989 Ripe/spawning Ripe/spawningAugust 1989 Ripe/spawning Ripe/spawningSeptember 1989 Ripe/spawning Ripe/spawning
610 L. Etim & M. Taege
7 -1
6 -I
f - '-5-1Z E 0 5-
T
r- 1 2 -
0-12500-1
Ir 11 I I 11 I I II
0 - '
20-1
::H f̂- 16-
8 -
0- "
20-127 8 35 21
16 -
\ 8 -
0 - "
0 6 -
0 4 -
0 2-
• 1988 — 1989
Figure 2. Monthly changes in the physicochemical and biological parameters of the Cross River at Itu Bridge(— bottom, — surface) (after Akpan 1991),

Condition o/Egeria 611
Invariably, there is the ecological adaptation in which the clam gradually builds up bothsomatic and reproductive tissues during the dry season when food supply is abundant, and asubsequent drain on these during the ensuing rainy season months when the environmentalcondition is poor. Thus, the timing of spawning is such that the larvae stand to profit from thehigh phytoplankton abundance during the ensuing dry season months.
From the foregoing, it is difficult to describe a particular environmental parameter as the'proximate factor' whose changes stimulate the onset of spawning in E. radiata. The onset ofspawning, then, may be triggered off by the synergistic and/or antagonistic changes in all theenvironmental factors in the aquatic milieu which are in turn caused by the twin effect of rainand flood during the peak of the rainy season.
References
Akpan E.R. (1991) Seasonal variation in phytoplankton biomass and pigments in relation to physicochemical factorsin the Cross river system. PhD Thesis. University of Calabar.
Ansell A.D. (1974) Seasonal changes in biochemical composition of bivalve/It™ aftra from Clyde sea area. MarineBiology 25, 13-20.
Bayne B.L, (1975) Aspectsof the physiological condition in A/j'fi7i«e(iu/tr(L.), with special reference to the effectsof oxygen tension and salinity. In; Proceedings of the Ninth European Marine Biology Symposium (ed. by H.Barnes), pp. 213-238. Aberdeen University Press, Aberdeen.
Beninger P.G. & Lucas A. (1984) Seasonal variation in condition, reproductive activity and gross biochemicalcomposition of two species of adult clam reared in a common habitat: Tapes decussatus L. (Jeffreys) and Tapesphilippinarum (Adams & Reeve). Journal of Experimental Marine Biology and Ecology 79, 19-37.
Britton J.D. & Morton B. (1982) A dissection guide, field and laboratory manual for the introduced bivalveCorbicula fiuminea. Malacology Review (Suppl. 3), 82pp.
Dhamne K.P. & Mane U.H. (1979) Some factors affecting fluid loss in the edible clam fapAia/a/emu/ca (Lamarck).Journal of Molluscan Studies 45, 161-166.
Etim L. (1990) Annual variation in proximate composition and condition index of Egeria radiata (Bivalvia:Tellinacea: Donacidae) from the Cross river, Nigeria. Nigerian Journal of Technological Research 2, 95-98.
Etim L. (1991) Growth, biochemical composition and reproductive cycle in Egeria radiata (Lamarck 1804)(Lamellibranchia: Tellinacea: Donacidae) from the Cross River in Nigeria. PhD Thesis, University of Calabar.
Etim L., Etched I. & Umoren O. (1989) Aspects of the reproductive biology of Tilapia gulneensis (Perciformes:Cichlidae) in Editas ponds, Nigeria. Revue de Zoologie Africaine-Journal of African Zoology 103, 127-134.
Gabbott P. A. & Bayne B.L. (1973) Biochemical effects of temperature and nutritive stress on Mytilus edulis (L.),Journal of the Marine Biological Association of the United Kingdom.53, 269-368.
Gabbott P. A. &Stephenson R.R. (1974) A note on the relationship between the dry weight condition index and theglycogen content of adult oysters {Ostrea edulis (L.)) kept in the laboratory. Journal du Conseil Internationalpour I'Exploration de la Mer 35, 359-361.
Gabbott P. & Walker A. J.M- (1971) Changes in the condition index and biochemical content of adult oysters (Ostreaedulis (L.)) maintained under hatchery conditions. Journal du Conseil International pour I'Exploration de laMe/-34, 99-106.
Galtsoff P.S. (1964) The American oyster Crassosirea virginica Gmelin. Fishery Bulletin 64. US Fish and WildlifeService. Department of the Interior. Washington DC.
Holland D.L. & Hannant P.J. (1974) Biochemical changes during growth of the spat of the oyster, Ostrea edulis L.Journal of the Marine Biological Association of the United Kingdom 54, 1007-1116.
Lowenberg U. & Kunzel Th. (1986) Investigations on the hydrology of the lower Cross river, Nigeria. Mimeograph.University of Calabar.
Lucas A. & Beninger G. (1985) The use of physiological condition indices in marine bivalve aquacutture.Aquaculture 44, 187-200.
Lutz R.A.. Inc2s L.S., Porter B. & Stotz J.K. (1980) Seasonal variation in the condition of raft-cultivated mussels{Mytilus edulis (I.)). Proceedings of the World Mariculture Society U, 262-268.

612 L. Etim & M. Taege
Mann R. (1978) A comparison of morphometric, biochemical and physiological indices of condition in marinebivalve mollusc. \t\: Energy and Environmental Stress in Aqmtic Systems {fiti.by ^^i.T)^Q^^&i.'^.Q\hbon&),pp. 484-497, Technical Information Centre. USA.
MannR- & RhytherJ.H. (1977) Growth of six species of bivalve mollusc in a waste recyciing-aquaculture system.Aquaculture U, 231-245.
Moses B.S. (1979) The Cross river — its ecology and fisheries. In: Proceedings of the International Conference onKainji Lake and River Basin Development in Africa, New Bussa, pp. 355-371. Kalnji Lake Institute Press, NewBussa. Nigeria,
Moses B.S, (1987) The influence of Rood regime on fish catch and fish communities of the Cross river Hood plainecosystem, Nigeria. Environmetal Biology offish 18, 57-65.
Nair N.U. & Nair N.U. (1987) Condition index and percentage edibility of Crassoslrea madrasensis (Preston)inhabiting Cochin Harbour, Fishery Technology TA, 14-21,
Pearson R.D, (1975) Laboratory Techniques in Food Analysis. Butterworth, London.Seed R. (1976) Ecology, In: Marine Mussels: Their Ecology and Physiology {ed. by B.L. Bayne), pp. 13-65. IBP No.
10. Cambridge University Press. Cambridge.Sundet J.H. & Vahl D, (1981) Seasonal changes in dry weight and biochemical composition of the tissue of sexually
mature and immature Iceland scallops Chlamys islandica. Journal of the Marine Biological Association of theUnited Kingdom 61. 1001-1010.
Taylor A.C. & Venn T,J, (1979) Seasonal variation in weight and biochemical composition of the tissue of the queenscallop Chlamys opercularis, from Clyde sea area. Journal of the Marine Biological Association of the UnitedKingdom 59, 60^^21.
Walne P. R. (1970) The seasonal variation of meat and glycogen content of seven populations of Ostrea edults L. anda review of the literature. Fishery Investigations. London Series 2 26, 1-35,
Walne P.R. & Mann R. (1975) Growth and biochemical composition in Ostrea edulis and Crassostrea gigas. In:Proceedings of the Ninth European Marine Biological Symposium (ed, by H. Barnes), pp. 587-607. AberdeenUniversity Press, Aberdeen.
Wilde de P.AW.J, (1975) Influence of temperature on behaviour, energy, metabolism and growth of Macomabalthica (L.), In: Proceedings of the Ninth European Marine Biological Symposium (ed. by H. Barnes), pp.239-256. Aberdeen University Press, Aberdeen.













![Carbon and nitrogen assimilation, and growth of moongbean (Vigna radiata [L.] Wilczek) cultivars grown under sulfur regimes](https://static.fdokumen.com/doc/165x107/63254f6b545c645c7f09a25b/carbon-and-nitrogen-assimilation-and-growth-of-moongbean-vigna-radiata-l-wilczek.jpg)







