
Expression and localization of ghrelin and its functional receptor in corpus luteum during...
12
Expression and localization of ghrelin and its functional receptor in corpus luteum during different stages of estrous cycle and the modulatory role of ghrelin on progesterone production in cultured luteal cells in buffalo M. Gupta a , S.S. Dangi a , V.S. Chouhan a , I. Hyder a , V. Babitha a , V.P. Yadav a , F.A. Khan c, d , A. Sonwane b , G. Singh a , G.K. Das e , A. Mitra b , S. Bag a , M. Sarkar a, * a Division of Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar 243122, India b Animal Genetics Division, Indian Veterinary Research Institute, Izatnagar 243122, India c Department of Animal Sciences, University of Florida, Gainesville, FL 32611 d D.H. Barron Reproductive and Perinatal Biology Research Program, University of Florida, Gainesville, FL 32611 e Division of Animal Reproduction, Indian Veterinary Research Institute, Izatnagar 243122, India article info Article history: Received 14 November 2013 Received in revised form 9 January 2014 Accepted 10 January 2014 Keywords: Ghrelin GHS-R1a Buffalo Corpus luteum Progesterone abstract Evidence obtained during recent years provided has insight into the regulation of corpus luteum (CL) development, function, and regression by locally produced ghrelin. The pre- sent study was carried out to evaluate the expression and localization of ghrelin and its receptor (GHS-R1a) in bubaline CL during different stages of the estrous cycle and inves- tigate the role of ghrelin on progesterone (P4) production along with messenger RNA (mRNA) expression of P4 synthesis intermediates. The mRNA and protein expression of ghrelin and GHS-R1a was significantly greater in mid- and late luteal phases. Both factors were localized in luteal cells, exclusively in the cytoplasm. Immunoreactivity of ghrelin and GHS-R1a was greater during mid- and late luteal phases. Luteal cells were cultured in vitro and treated with ghrelin each at 1, 10, and 100 ng/mL concentrations for 48 h after obtaining 75% to 80% confluence. At a dose of 1 ng/mL, there was no significant difference in P4 secretion between control and treatment group. At 10 and 100 ng/mL, there was a decrease (P < 0.05) in P4 concentration, cytochrome P45011A1 (CYP11A1), and 3-beta- hydroxysteroid dehydrogenase mRNA expression and localization. There was no difference in mRNA expression of steroidogenic acute regulatory protein between control and treatment group. In summary, the present study provided evidence that ghrelin and its receptor are expressed in bubaline CL and are localized exclusively in the cell cytoplasm and ghrelin has an inhibitory effect on P4 production in buffalo. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction Buffalo (Bubalus bubalis) is an important livestock spe- cies, concentrated mostly in the tropical and subtropical regions of the world. Asian buffaloes, which represent 97.2% of the world’s buffalo, lead the worldwide population of 189.8 million [1]. In India, the population of buffalo is half compared with the cattle population but contributes 55% of the total milk production (62.8 MT in 2010) [1]. A large number of buffaloes (30%–40%) remain unproductive as the result of reproductive problems such as subestrus, anestrus, and infertility, all of which have hormonal etiologies [2,3], incurring an estimated loss of 19 to 20 million tons of milk each year. Infertility may result from poor development and * Corresponding author. Tel.: þ91 0581 231045; fax: þ91 0581 2301327. E-mail address: [email protected] (M. Sarkar). Contents lists available at ScienceDirect Domestic Animal Endocrinology journal homepage: www.domesticanimalendo.com 0739-7240/$ – see front matter Ó 2014 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.domaniend.2014.01.004 Domestic Animal Endocrinology 48 (2014) 21–32
Transcript of Expression and localization of ghrelin and its functional receptor in corpus luteum during...

ilable at ScienceDirect
Domestic Animal Endocrinology 48 (2014) 21–32
Contents lists ava
Domestic Animal Endocrinology
journal homepage: www.domest icanimalendo.com
Expression and localization of ghrelin and its functionalreceptor in corpus luteum during different stages of estrouscycle and the modulatory role of ghrelin on progesteroneproduction in cultured luteal cells in buffalo
M. Gupta a, S.S. Dangi a, V.S. Chouhan a, I. Hyder a, V. Babitha a, V.P. Yadav a, F.A. Khan c,d,A. Sonwane b, G. Singh a, G.K. Das e, A. Mitra b, S. Bag a, M. Sarkar a,*
aDivision of Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar 243122, IndiabAnimal Genetics Division, Indian Veterinary Research Institute, Izatnagar 243122, IndiacDepartment of Animal Sciences, University of Florida, Gainesville, FL 32611dD.H. Barron Reproductive and Perinatal Biology Research Program, University of Florida, Gainesville, FL 32611eDivision of Animal Reproduction, Indian Veterinary Research Institute, Izatnagar 243122, Indiaa r t i c l e i n f o
Article history:Received 14 November 2013Received in revised form 9 January 2014Accepted 10 January 2014
Keywords:GhrelinGHS-R1aBuffaloCorpus luteumProgesterone
* Corresponding author. Tel.: þ91 0581 231045; faE-mail address: [email protected] (M. Sarka
0739-7240/$ – see front matter � 2014 Elsevier Inchttp://dx.doi.org/10.1016/j.domaniend.2014.01.004
a b s t r a c t
Evidence obtained during recent years provided has insight into the regulation of corpusluteum (CL) development, function, and regression by locally produced ghrelin. The pre-sent study was carried out to evaluate the expression and localization of ghrelin and itsreceptor (GHS-R1a) in bubaline CL during different stages of the estrous cycle and inves-tigate the role of ghrelin on progesterone (P4) production along with messenger RNA(mRNA) expression of P4 synthesis intermediates. The mRNA and protein expression ofghrelin and GHS-R1a was significantly greater in mid- and late luteal phases. Both factorswere localized in luteal cells, exclusively in the cytoplasm. Immunoreactivity of ghrelin andGHS-R1a was greater during mid- and late luteal phases. Luteal cells were cultured in vitroand treated with ghrelin each at 1, 10, and 100 ng/mL concentrations for 48 h afterobtaining 75% to 80% confluence. At a dose of 1 ng/mL, there was no significant differencein P4 secretion between control and treatment group. At 10 and 100 ng/mL, there was adecrease (P < 0.05) in P4 concentration, cytochrome P45011A1 (CYP11A1), and 3-beta-hydroxysteroid dehydrogenase mRNA expression and localization. There was no differencein mRNA expression of steroidogenic acute regulatory protein between control andtreatment group. In summary, the present study provided evidence that ghrelin and itsreceptor are expressed in bubaline CL and are localized exclusively in the cell cytoplasmand ghrelin has an inhibitory effect on P4 production in buffalo.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Buffalo (Bubalus bubalis) is an important livestock spe-cies, concentrated mostly in the tropical and subtropicalregions of the world. Asian buffaloes, which represent 97.2%
x: þ91 0581 2301327.r).
. All rights reserved.
of the world’s buffalo, lead the worldwide population of189.8 million [1]. In India, the population of buffalo is halfcompared with the cattle population but contributes 55% ofthe total milk production (62.8 MT in 2010) [1]. A largenumber of buffaloes (30%–40%) remain unproductive as theresult of reproductive problems such as subestrus, anestrus,and infertility, all of which have hormonal etiologies [2,3],incurring an estimated loss of 19 to 20 million tons of milkeach year. Infertility may result from poor development and

M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3222
incomplete vascularization of the corpus luteum (CL), lead-ing to a deranged luteal function. A thorough understandingof the endocrine, autocrine, and paracrine factors thatregulate luteal development and function would be helpfulin elucidating the mechanisms underlying luteal insuffi-ciency, thereby facilitating development of approaches foralleviation of infertility resulting from luteal dysfunction.
Reproductive cycles in mammals represent a process ofchronological proliferation, differentiation, and trans-formation of ovarian follicular cells followed by formationand regression of the CL in a cyclic manner. The CL is aremarkable, transiently functioning organ and plays acentral role in the regulation of the estrous cycle and in themaintenance of pregnancy. This function is carried outlargely by progesterone (P4), which is the main steroidhormone synthesized by this gland [4]. This process is wellregulated by pituitary gonadotropins. However, recent re-ports also provide evidence of an essential modulatory roleon luteal dynamics by locally produced factors [5].
Data obtained in recent years have demonstrated a closerelationship between ghrelin and reproductive functions infemales. Ghrelin is a novel growth hormone–releasing acyl-ated peptide, isolated from the stomach. It is the endogenousligand for the growth hormone secretagogue receptor (GHS-R1a), a 7-transmembrane G protein–coupled receptor [6].Activity of ghrelin is dependent on a unique n-octanoylationat Ser-3 [6]. Acylation of ghrelin is necessary for binding andactivating the GHS-R1a [7]. A striking feature of ghrelin is itswidespread pattern of expression [8,9]. Expression of ghrelinhas been demonstrated in an array of tissues and cell types,including the stomach, small intestine, pancreas, lympho-cytes, placenta, kidney, lung, pituitary, and brain [9]. Althoughthe general view has been that the principal effects of ghrelinare on the neuroendocrine component of reproduction[10–13], evidence has emerged to indicate direct involvementof ghrelin in ovarian function. Expression of ghrelin and itscognate receptor has been demonstrated in the ovary ofhuman [14], rat [15], sheep [16], goat [17], pig [18], and cattle[19]. In human and pigs, it is reported that expression ofghrelin and its active receptor GHS-R1a paralleled folliculardevelopment [14,20]. In pig, sheep, and goat ovaries, it hasbeen demonstrated that ghrelin and its receptor expression isestrous-cycle dependent,withmaximal expressionduring thedevelopment of the CL [16–18]. Plasma ghrelin concentrationranges from 100 to 150 pg/mL in fed cattle to 600 to 1,000 pg/mL in fasted cows [21], whereas ovarian follicular fluid (FF)ghrelin concentrations vary from 109 to 148 pg/mL [20].
Several studies have shown that ghrelin plays a crucialrole in steroidogenesis. Key factors involved in P4 synthesisin the CL of various species include steroidogenic acuteregulatory protein (StAR), cholesterol side chain cleavageenzyme complex P450scc or cytochrome P45011A1(CYP11A1), and 3-beta-hydroxysteroid dehydrogenase (3b-HSD) [22–24]. Both stimulatory and inhibitory effects ofghrelin on the secretion of P4 by cultured rat, porcine,rabbit, human, and chicken ovarian cells have been re-ported [18,25,26], although such actions have not beenfound consistently [27,28]. Given their role in controllingovarian function, ghrelin and its receptor are hypothesizedto be involved in CL formation, function, development, andsteroidogenesis during the estrous cycle in an autocrine/
paracrine manner in buffalo. To test this hypothesis, weevaluated (a) messenger RNA (mRNA) and protein expres-sion and localization of ghrelin and its receptor in bubalineCL during different stages of estrous cycle and (b) thein vitro effects of ghrelin on P4 secretion, StAR, CYP11A1,and 3b-HSDmRNA expression and localization in luteal cellculture.
2. Materials and methods
2.1. Collection of CL during the estrous cycle
Entire reproductive tracts from buffalo cows werecollected at a local slaughterhouse within 10 to 20 min afterslaughter and were transported on ice to the laboratory. Thestage of the estrous cycle was defined by macroscopicobservation of the ovaries (color, consistency, CL stage,number, and size of follicles) and the uterus (color, consis-tency, andmucus) as described previously [29]. Forty ovaries,each with CL, were used to extract 10 CL per group for RNAextraction, Western blotting, and immunohistochemistrystudies. The CL was assigned to the following stages: earlyluteal phase (days 1–4), mid-luteal phase (days 5–10), lateluteal phase (days 11–16), and regressed CL (days >17) ofestrous cycle. Luteal tissue was frozen in liquid nitrogen andstored at �80�C until RNA and protein isolation.
2.2. Collection of follicles during final follicular growth
Only follicles (n ¼ 10) which appeared healthy (ie, wellvascularized and having transparent follicular wall andfluid) and whose diameters were>13mmwere used. Largefollicles (>13 mm) were collected only after CL regression,with signs of mucus production in the uterus and cervixand were assumed to be preovulatory. For RNA extraction,follicles were dissected from the ovary. The surroundingtissue (theca externa) was removed with forceps under astereomicroscope as previously described [29]. FF aspiratedfrom follicles was stored at �20�C until determination ofP4. As healthy follicles have relatively constant P4 levels inFF, only follicles with P4 <100 ng/mL FF were used for theevaluation to exclude atretic follicles [29]. Follicles weresnap-frozen in liquid nitrogen and stored at �80�C untilRNA and protein isolation. The preovulatory follicles (PF)were used as a control/calibrator in the quantitative real-time–polymerase chain reaction (qRT-PCR) experiments.
2.3. Hormone determination
Concentrations of P4, determined in the FF and in thespent media of luteal cell culture, were estimated by P4125IRIA kit (IM1188) supplied by Immunotech, Czech Republicas per manufacturer’s instruction. The FF was diluted 1:5with phosphate-buffered saline (PBS). The measurablerange was 0.05 to 50 ng/mL. The intra- and interassay co-efficients of variation were 6.5% and 7.2%, respectively.
2.4. Primers
A primer for ghrelin was designed using the Fast PCR(Version: 6.2.73) software. Published primers were used for
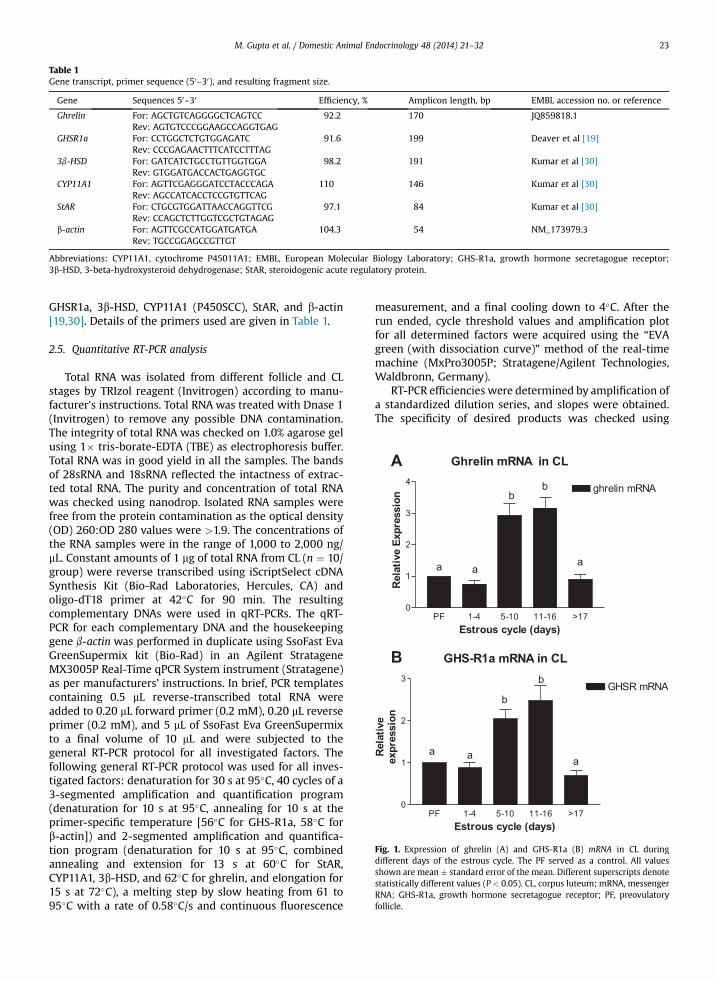
Table 1Gene transcript, primer sequence (50–30), and resulting fragment size.
Gene Sequences 50–30 Efficiency, % Amplicon length, bp EMBL accession no. or reference
Ghrelin For: AGCTGTCAGGGGCTCAGTCCRev: AGTGTCCCGGAAGCCAGGTGAG
92.2 170 JQ859818.1
GHSR1a For: CCTGGCTCTGTGGAGATCRev: CCCGAGAACTTTCATCCTTTAG
91.6 199 Deaver et al [19]
3b-HSD For: GATCATCTGCCTGTTGGTGGARev: GTGGATGACCACTGAGGTGC
98.2 191 Kumar et al [30]
CYP11A1 For: AGTTCGAGGGATCCTACCCAGARev: AGCCATCACCTCCGTGTTCAG
110 146 Kumar et al [30]
StAR For: CTGCGTGGATTAACCAGGTTCGRev: CCAGCTCTTGGTCGCTGTAGAG
97.1 84 Kumar et al [30]
b-actin For: AGTTCGCCATGGATGATGARev: TGCCGGAGCCGTTGT
104.3 54 NM_173979.3
Abbreviations: CYP11A1, cytochrome P45011A1; EMBL, European Molecular Biology Laboratory; GHS-R1a, growth hormone secretagogue receptor;3b-HSD, 3-beta-hydroxysteroid dehydrogenase; StAR, steroidogenic acute regulatory protein.
Fig. 1. Expression of ghrelin (A) and GHS-R1a (B) mRNA in CL duringdifferent days of the estrous cycle. The PF served as a control. All valuesshown are mean � standard error of the mean. Different superscripts denotestatistically different values (P < 0.05). CL, corpus luteum; mRNA, messengerRNA; GHS-R1a, growth hormone secretagogue receptor; PF, preovulatoryfollicle.
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–32 23
GHSR1a, 3b-HSD, CYP11A1 (P450SCC), StAR, and b-actin[19,30]. Details of the primers used are given in Table 1.
2.5. Quantitative RT-PCR analysis
Total RNA was isolated from different follicle and CLstages by TRIzol reagent (Invitrogen) according to manu-facturer’s instructions. Total RNA was treated with Dnase 1(Invitrogen) to remove any possible DNA contamination.The integrity of total RNA was checked on 1.0% agarose gelusing 1� tris-borate-EDTA (TBE) as electrophoresis buffer.Total RNA was in good yield in all the samples. The bandsof 28sRNA and 18sRNA reflected the intactness of extrac-ted total RNA. The purity and concentration of total RNAwas checked using nanodrop. Isolated RNA samples werefree from the protein contamination as the optical density(OD) 260:OD 280 values were >1.9. The concentrations ofthe RNA samples were in the range of 1,000 to 2,000 ng/mL. Constant amounts of 1 mg of total RNA from CL (n ¼ 10/group) were reverse transcribed using iScriptSelect cDNASynthesis Kit (Bio-Rad Laboratories, Hercules, CA) andoligo-dT18 primer at 42�C for 90 min. The resultingcomplementary DNAs were used in qRT-PCRs. The qRT-PCR for each complementary DNA and the housekeepinggene b-actin was performed in duplicate using SsoFast EvaGreenSupermix kit (Bio-Rad) in an Agilent StratageneMX3005P Real-Time qPCR System instrument (Stratagene)as per manufacturers’ instructions. In brief, PCR templatescontaining 0.5 mL reverse-transcribed total RNA wereadded to 0.20 mL forward primer (0.2 mM), 0.20 mL reverseprimer (0.2 mM), and 5 mL of SsoFast Eva GreenSupermixto a final volume of 10 mL and were subjected to thegeneral RT-PCR protocol for all investigated factors. Thefollowing general RT-PCR protocol was used for all inves-tigated factors: denaturation for 30 s at 95�C, 40 cycles of a3-segmented amplification and quantification program(denaturation for 10 s at 95�C, annealing for 10 s at theprimer-specific temperature [56�C for GHS-R1a, 58�C forb-actin]) and 2-segmented amplification and quantifica-tion program (denaturation for 10 s at 95�C, combinedannealing and extension for 13 s at 60�C for StAR,CYP11A1, 3b-HSD, and 62�C for ghrelin, and elongation for15 s at 72�C), a melting step by slow heating from 61 to95�C with a rate of 0.58�C/s and continuous fluorescence
measurement, and a final cooling down to 4�C. After therun ended, cycle threshold values and amplification plotfor all determined factors were acquired using the “EVAgreen (with dissociation curve)” method of the real-timemachine (MxPro3005P; Stratagene/Agilent Technologies,Waldbronn, Germany).
RT-PCR efficiencies were determined by amplification ofa standardized dilution series, and slopes were obtained.The specificity of desired products was checked using

A
B
C
Estrous cycle (days)
Estrous cycle (days)
Fig. 2. Relative protein expression (mean � standard error of the mean,band densitometry) in CL tissue on different days of the estrous cycle inbuffalo, with the PF serving as the control (A) Western blot bands of ghrelin,GHS-R1, and b-actin in buffalo CL; b-actin used as reference protein. Relativechange in expression of ghrelin (B) and GHS-R1a (C) in CL. Different su-perscripts denote statistically different values (P < 0.05). CL, corpus luteum;GHS-R1a, growth hormone secretagogue receptor; PF, preovulatory follicle.
Fig. 3. (A) Histologic findings in Bubalus bubalis CL. The representativesection was stained with hematoxylin (blue staining) and eosin (pink) tovisualize cellular nuclei and cytoplasm. Histologic evaluation showed thatluteal cells were round, with a large, central nucleus and were present in-side the capsule of the CL. Endothelial cells were small and flat and localizedaround the capillary inner layer. Pericytes were oval in shape and colo-calized with endothelial cells surrounding them in the outer layers of thecapillaries. (B) Immunohistochemical localization of ghrelin in Bubalusbubalis CL. Immunoreactivity of ghrelin in different stages of CL develop-ment was evaluated using CFL-488. Expression of ghrelin is merged with40 ,6-diamidino-2-phenylindole dihydrochloride counterstain (blue), indi-cating the nuclei of all cells in the sections. Representative pictures showimmunoreactivity against ghrelin (A: early luteal phase, B: mid-lutealphase, C: late luteal phase, D: regressed CL, and E: negative control). Con-trol sections are represented with isotype IgG and without primary anti-body labeling. Ghrelin was localized predominantly in cytoplasm of largeluteal cells and to some extent in small luteal. Scale bar ¼ 50 mm. (C)Immunohistochemical localization of GHS-R1a in Bubalus bubalis CL.Immunoreactivity of ghrelin in different stages of CL development wasevaluated using CFL-647. Expression of GHS-R1a is merged with 40 ,6-diamidino-2-phenylindole dihydrochloride counterstain (blue), indicatingthe nuclei of all cells in the sections. Representative pictures show immu-noreactivity against GHS-R1a (A: early luteal phase, B: mid luteal phase, C:late luteal phase, D: regressed CL, and E: negative control). Control sectionsare represented with isotype IgG and without primary antibody labeling.GHS-R1a was localized predominantly in cytoplasm of large luteal cells andsmall luteal cells. Scale bar ¼ 50 mm. BV, blood vessels; EC, endothelial cell;LLC, large luteal cell; Pe, pericytes; SLC, small luteal cell. (For interpretationof the references to color in this figure legend, the reader is referred to theweb version of this article.)
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3224
analysis of melting temperature, which is product-specificand a high-resolution gel electrophoresis to verify thattranscripts were of exact molecular size and furtherconfirmed by sequence analysis. Negative control PCRcontaining all components except template was includedfor each sample to check out the formation of primer dimer.
2.6. Antibodies and growth factor
Western blotting, immunohistochemistry, and im-munocytochemistry were performed using monoclonalanti-mouse b-actin (SC-81178; Santa Cruz Biotechnology,Inc., Dallas, TX), rabbit polyclonal ghrelin (SC-50297,Lot#H01111), goat polyclonal GHS-R1a (SC-10359, Lot#D1812), goat polyclonal 3b-HSD (sc-30820, Lot#G0512), goatpolyclonal CYP11A1 (SC-18043, Lot#D0412) antibody,mouseanti-goat IgG-HRP (sc-2354, Lot # G0910), goat anti-rabbitIgG-HRP (sc-2004, Lot #B1711), goat Anti Rabbit IgG-CFL-
488 (sc-362262, Lot#B0112), bovine anti-goat IgG-CFL-488(sc-362254, Lot#B0812), bovine anti-goat IgG-CFL-647 (sc-362284, Lot#B0312), and monoclonal anti-mouse b-actin(SC-81178). Human ghrelin (cat. no. 4990-100, Lot no. 40190)was procured from BioVision.
2.7. Western blot
To obtain total proteins, liquid nitrogen triturated lutealtissues of different stages were suspended in Tissue-PE LB (GBiosciences, St. Louis, MO) buffer and Halt protease inhibitorcocktail (Thermo Scientific), homogenized, and centrifugedat 12,000g. The supernatant containing mostly the totalsoluble protein was estimated using Bradford protein assay.
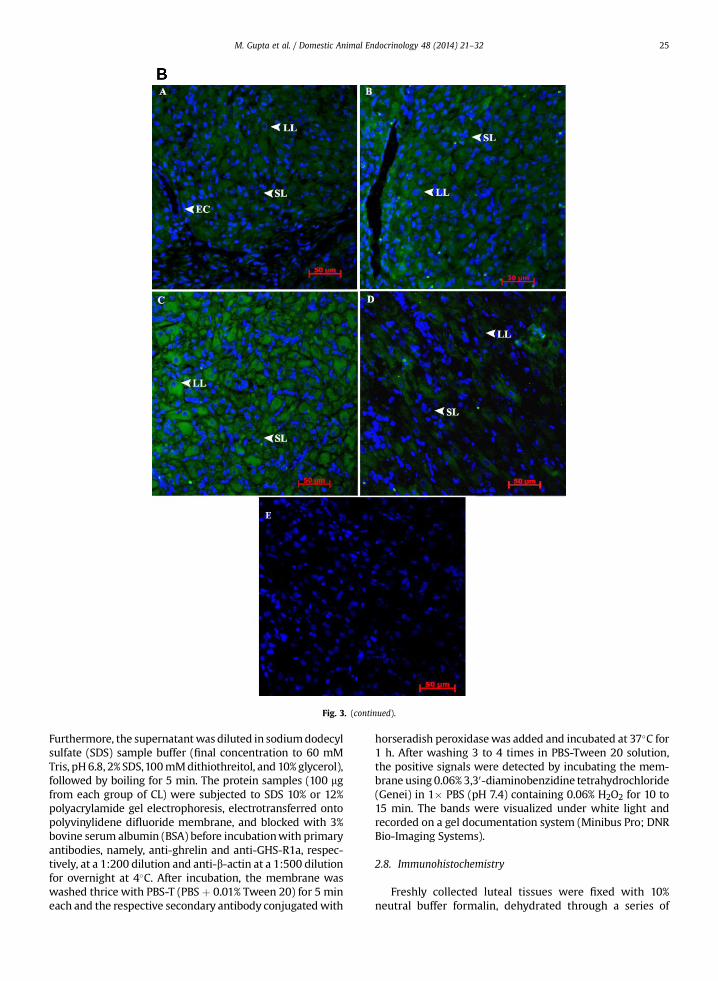
Fig. 3. (continued).
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–32 25
Furthermore, the supernatantwas diluted in sodiumdodecylsulfate (SDS) sample buffer (final concentration to 60 mMTris, pH6.8, 2% SDS,100mMdithiothreitol, and 10%glycerol),followed by boiling for 5 min. The protein samples (100 mgfrom each group of CL) were subjected to SDS 10% or 12%polyacrylamide gel electrophoresis, electrotransferred ontopolyvinylidene difluoride membrane, and blocked with 3%bovine serum albumin (BSA) before incubationwith primaryantibodies, namely, anti-ghrelin and anti-GHS-R1a, respec-tively, at a 1:200 dilution and anti-b-actin at a 1:500 dilutionfor overnight at 4�C. After incubation, the membrane waswashed thrice with PBS-T (PBS þ 0.01% Tween 20) for 5 mineach and the respective secondary antibody conjugatedwith
horseradish peroxidasewas added and incubated at 37�C for1 h. After washing 3 to 4 times in PBS-Tween 20 solution,the positive signals were detected by incubating the mem-brane using 0.06%3,30-diaminobenzidine tetrahydrochloride(Genei) in 1� PBS (pH 7.4) containing 0.06% H2O2 for 10 to15 min. The bands were visualized under white light andrecorded on a gel documentation system (Minibus Pro; DNRBio-Imaging Systems).
2.8. Immunohistochemistry
Freshly collected luteal tissues were fixed with 10%neutral buffer formalin, dehydrated through a series of
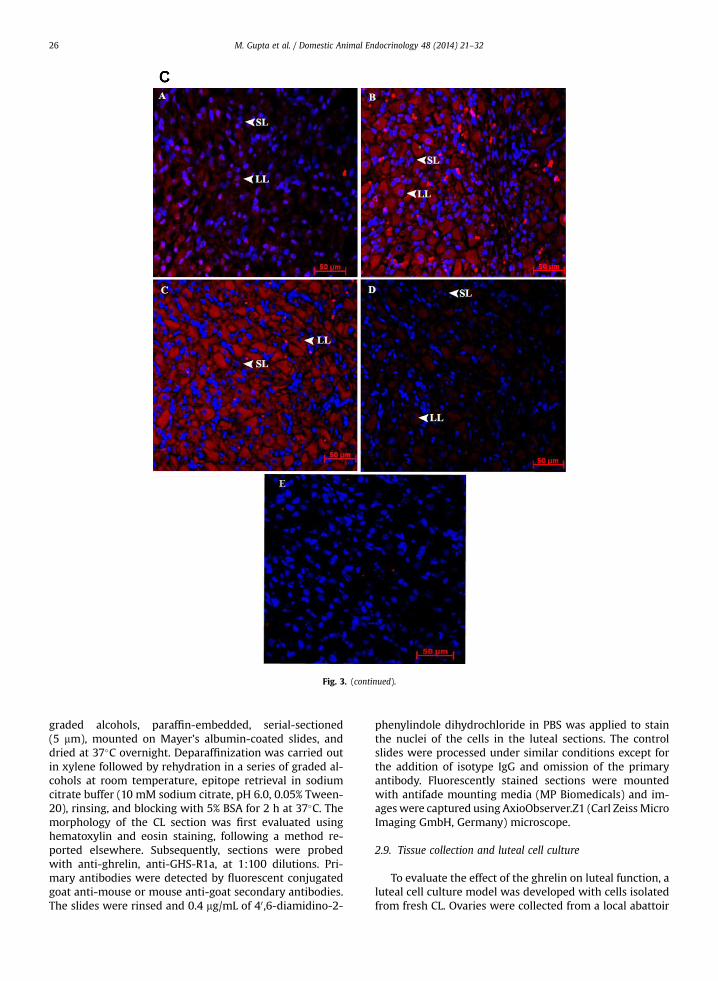
Fig. 3. (continued).
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3226
graded alcohols, paraffin-embedded, serial-sectioned(5 mm), mounted on Mayer’s albumin-coated slides, anddried at 37�C overnight. Deparaffinization was carried outin xylene followed by rehydration in a series of graded al-cohols at room temperature, epitope retrieval in sodiumcitrate buffer (10 mM sodium citrate, pH 6.0, 0.05% Tween-20), rinsing, and blocking with 5% BSA for 2 h at 37�C. Themorphology of the CL section was first evaluated usinghematoxylin and eosin staining, following a method re-ported elsewhere. Subsequently, sections were probedwith anti-ghrelin, anti-GHS-R1a, at 1:100 dilutions. Pri-mary antibodies were detected by fluorescent conjugatedgoat anti-mouse or mouse anti-goat secondary antibodies.The slides were rinsed and 0.4 mg/mL of 40,6-diamidino-2-
phenylindole dihydrochloride in PBS was applied to stainthe nuclei of the cells in the luteal sections. The controlslides were processed under similar conditions except forthe addition of isotype IgG and omission of the primaryantibody. Fluorescently stained sections were mountedwith antifade mounting media (MP Biomedicals) and im-ageswere captured using AxioObserver.Z1 (Carl ZeissMicroImaging GmbH, Germany) microscope.
2.9. Tissue collection and luteal cell culture
To evaluate the effect of the ghrelin on luteal function, aluteal cell culture model was developed with cells isolatedfrom fresh CL. Ovaries were collected from a local abattoir
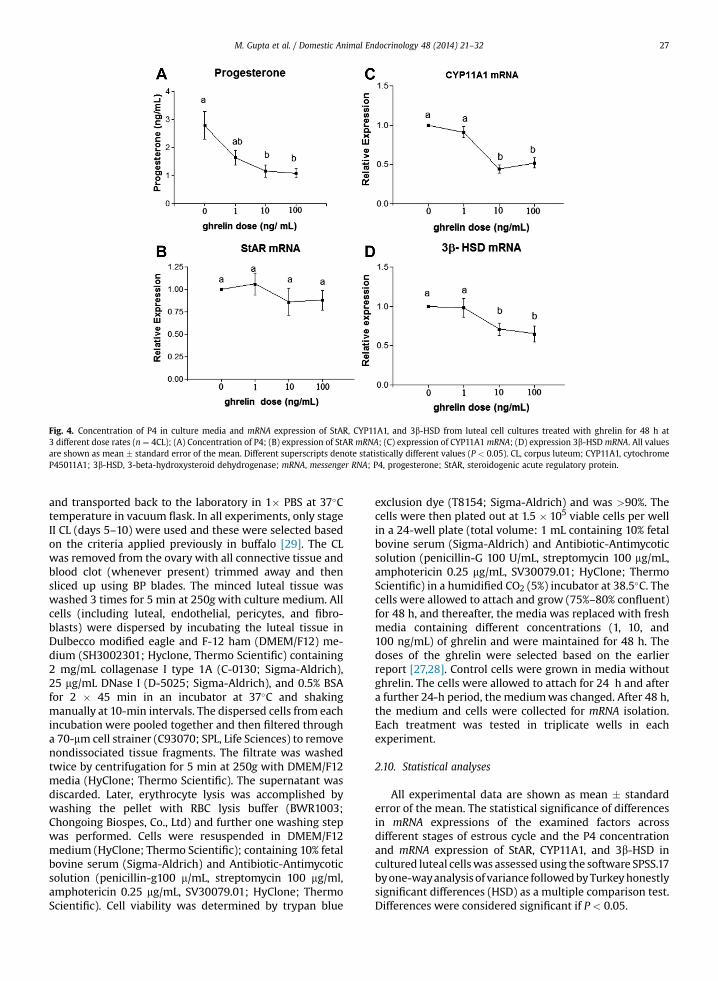
Fig. 4. Concentration of P4 in culture media and mRNA expression of StAR, CYP11A1, and 3b-HSD from luteal cell cultures treated with ghrelin for 48 h at3 different dose rates (n ¼ 4CL); (A) Concentration of P4; (B) expression of StAR mRNA; (C) expression of CYP11A1 mRNA; (D) expression 3b-HSDmRNA. All valuesare shown as mean � standard error of the mean. Different superscripts denote statistically different values (P < 0.05). CL, corpus luteum; CYP11A1, cytochromeP45011A1; 3b-HSD, 3-beta-hydroxysteroid dehydrogenase; mRNA, messenger RNA; P4, progesterone; StAR, steroidogenic acute regulatory protein.
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–32 27
and transported back to the laboratory in 1� PBS at 37�Ctemperature in vacuum flask. In all experiments, only stageII CL (days 5–10) were used and these were selected basedon the criteria applied previously in buffalo [29]. The CLwas removed from the ovary with all connective tissue andblood clot (whenever present) trimmed away and thensliced up using BP blades. The minced luteal tissue waswashed 3 times for 5 min at 250gwith culture medium. Allcells (including luteal, endothelial, pericytes, and fibro-blasts) were dispersed by incubating the luteal tissue inDulbecco modified eagle and F-12 ham (DMEM/F12) me-dium (SH3002301; Hyclone, Thermo Scientific) containing2 mg/mL collagenase I type 1A (C-0130; Sigma-Aldrich),25 mg/mL DNase I (D-5025; Sigma-Aldrich), and 0.5% BSAfor 2 � 45 min in an incubator at 37�C and shakingmanually at 10-min intervals. The dispersed cells from eachincubation were pooled together and then filtered througha 70-mm cell strainer (C93070; SPL, Life Sciences) to removenondissociated tissue fragments. The filtrate was washedtwice by centrifugation for 5 min at 250g with DMEM/F12media (HyClone; Thermo Scientific). The supernatant wasdiscarded. Later, erythrocyte lysis was accomplished bywashing the pellet with RBC lysis buffer (BWR1003;Chongoing Biospes, Co., Ltd) and further one washing stepwas performed. Cells were resuspended in DMEM/F12medium (HyClone; Thermo Scientific); containing 10% fetalbovine serum (Sigma-Aldrich) and Antibiotic-Antimycoticsolution (penicillin-g100 m/mL, streptomycin 100 mg/ml,amphotericin 0.25 mg/mL, SV30079.01; HyClone; ThermoScientific). Cell viability was determined by trypan blue
exclusion dye (T8154; Sigma-Aldrich) and was >90%. Thecells were then plated out at 1.5 � 105 viable cells per wellin a 24-well plate (total volume: 1 mL containing 10% fetalbovine serum (Sigma-Aldrich) and Antibiotic-Antimycoticsolution (penicillin-G 100 U/mL, streptomycin 100 mg/mL,amphotericin 0.25 mg/mL, SV30079.01; HyClone; ThermoScientific) in a humidified CO2 (5%) incubator at 38.5�C. Thecells were allowed to attach and grow (75%–80% confluent)for 48 h, and thereafter, the media was replaced with freshmedia containing different concentrations (1, 10, and100 ng/mL) of ghrelin and were maintained for 48 h. Thedoses of the ghrelin were selected based on the earlierreport [27,28]. Control cells were grown in media withoutghrelin. The cells were allowed to attach for 24 h and aftera further 24-h period, the mediumwas changed. After 48 h,the medium and cells were collected for mRNA isolation.Each treatment was tested in triplicate wells in eachexperiment.
2.10. Statistical analyses
All experimental data are shown as mean � standarderror of the mean. The statistical significance of differencesin mRNA expressions of the examined factors acrossdifferent stages of estrous cycle and the P4 concentrationand mRNA expression of StAR, CYP11A1, and 3b-HSD incultured luteal cellswas assessed using the software SPSS.17byone-wayanalysisof variance followedbyTurkeyhonestlysignificant differences (HSD) as a multiple comparison test.Differences were considered significant if P < 0.05.
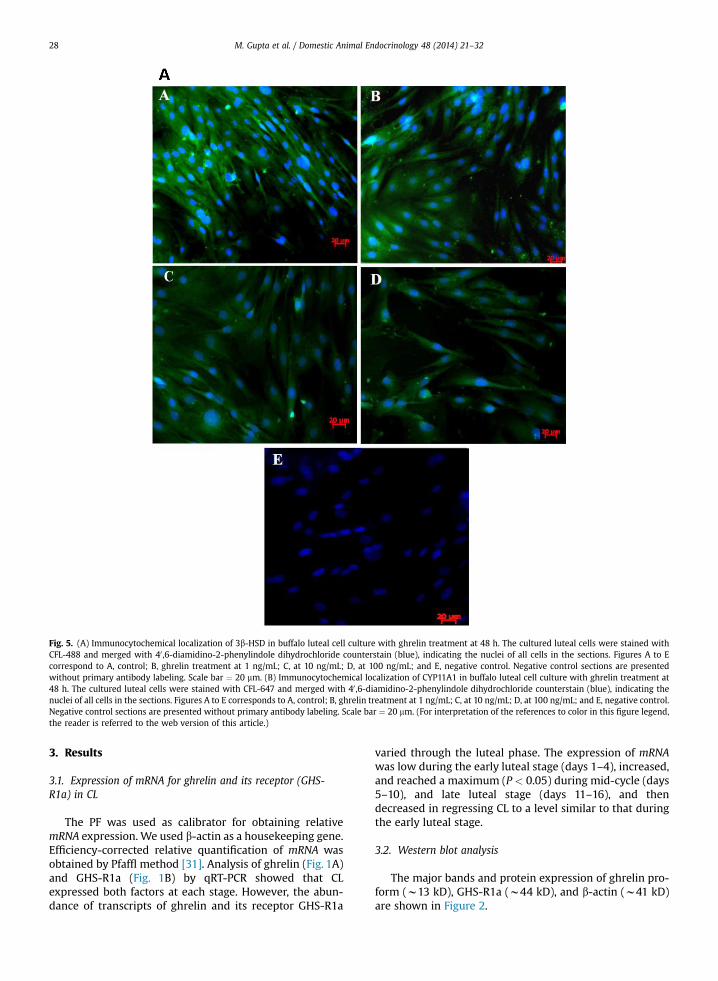
Fig. 5. (A) Immunocytochemical localization of 3b-HSD in buffalo luteal cell culture with ghrelin treatment at 48 h. The cultured luteal cells were stained withCFL-488 and merged with 40 ,6-diamidino-2-phenylindole dihydrochloride counterstain (blue), indicating the nuclei of all cells in the sections. Figures A to Ecorrespond to A, control; B, ghrelin treatment at 1 ng/mL; C, at 10 ng/mL; D, at 100 ng/mL; and E, negative control. Negative control sections are presentedwithout primary antibody labeling. Scale bar ¼ 20 mm. (B) Immunocytochemical localization of CYP11A1 in buffalo luteal cell culture with ghrelin treatment at48 h. The cultured luteal cells were stained with CFL-647 and merged with 40 ,6-diamidino-2-phenylindole dihydrochloride counterstain (blue), indicating thenuclei of all cells in the sections. Figures A to E corresponds to A, control; B, ghrelin treatment at 1 ng/mL; C, at 10 ng/mL; D, at 100 ng/mL; and E, negative control.Negative control sections are presented without primary antibody labeling. Scale bar ¼ 20 mm. (For interpretation of the references to color in this figure legend,the reader is referred to the web version of this article.)
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3228
3. Results
3.1. Expression of mRNA for ghrelin and its receptor (GHS-R1a) in CL
The PF was used as calibrator for obtaining relativemRNA expression. We used b-actin as a housekeeping gene.Efficiency-corrected relative quantification of mRNA wasobtained by Pfaffl method [31]. Analysis of ghrelin (Fig. 1A)and GHS-R1a (Fig. 1B) by qRT-PCR showed that CLexpressed both factors at each stage. However, the abun-dance of transcripts of ghrelin and its receptor GHS-R1a
varied through the luteal phase. The expression of mRNAwas low during the early luteal stage (days 1–4), increased,and reached a maximum (P < 0.05) during mid-cycle (days5–10), and late luteal stage (days 11–16), and thendecreased in regressing CL to a level similar to that duringthe early luteal stage.
3.2. Western blot analysis
The major bands and protein expression of ghrelin pro-form (w13 kD), GHS-R1a (w44 kD), and b-actin (w41 kD)are shown in Figure 2.

Fig. 5. (continued).
M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–32 29
3.3. Immunohistochemistry of CL
The morphology of each CL section was first evaluatedusing hematoxylin and eosin staining (Fig. 3A). As observedunder microscope, cells were identified according to shapeand localization. Luteal cells were round, with a large andcentral nucleus. Endothelial cells were small and flat andlocalized around the capillary inner layer. Pericytes wereoval in shape and colocalized with endothelial cells sur-rounding them in the outer layers of the capillaries. Theimmunoreactivities of ghrelin and GHS-R1a were observedusing immunohistochemistry with stage-specific differ-ence in reactivity. The localization of both the proteins wasconspicuous in various cell types in different stages of CLsections. The intensity of immunostaining varied fromphase to phase, and the number of positive cells wasinfluenced by the different stages of estrous cycle. Theimmunoreactivity was exclusively found in the cytoplasmof luteal cells and was greater during mid- and late phasesfor both factors. The negative controls, with isotype IgG and
without primary antibodies, showed only a weak back-ground staining (Fig. 3B, C).
3.4. Effect of ghrelin on P4 secretion and mRNA expression ofStAR, CYP11a1, and 3b-HSD
Luteal cells collected from CL during the mid-lutealphase were cultured for 48 h after attaining 75% to 80%confluence, without (control) or with increasing doses ofghrelin (1, 10, and 100 ng/mL). Concentrations of P4 incontrol culture averaged 2.78 � 0.45 ng/mL. P4 concen-trations decreased (P < 0.05) with ghrelin treatment in adose-dependent manner (Fig. 4A).
Expression of StAR, CYP11A1, and 3b-HSD (Fig. 4B–D)in cultured cells varied significantly according to the giventreatment. There was no difference in StAR mRNAexpression between control and treatment of all doses.CYP11A1 and 3b-HSD mRNA expression declined (P <
0.05) at 10 and 100 ng/mL dose of ghrelin as comparedwith control.

M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3230
3.5. Immunocytochemistry
The morphology and confluence of the cultured cellswas first observed under bright field illumination. Attachedluteal cells were spindle shaped, with a large and centralnucleus. Localization of CYP11A1 and 3b-HSD protein indifferent treatment was observed using fluorescent mi-croscopy. There was a decrease in number and intensity ofimmunoreactive cells in all doses of ghrelin-treated cells ascompared with control (Fig. 5A, B).
4. Discussion
The objective of the present study was to determineexpression and localization of ghrelin and its receptor,GHS-R1a, in the bubaline CL obtained from different stagesof the estrous cycle and to determine the modulatory effectof ghrelin on P4 production in buffalo luteal cell culture. Tothe best of our knowledge, our study is the first of its kindconducted in bubaline species to show the strong expres-sion of ghrelin and its receptor in buffalo CL and modula-tion of luteal function by it. The mRNA and proteinexpression of ghrelin and ghrelin receptor in buffalo CLwere consistent throughout the estrous cycle. Their relativeexpression, however, varied according to the stage ofestrous cycle; it was higher in mid- and late luteal phasesand lower in early and regressing phases. The presentfindings are in line with the earlier observation in ovine[16] and caprine [17] CL in which the greatest mRNAexpression of ghrelin and its receptor was observed in fullydeveloped CL. In porcine CL, the greatest ghrelin expressionwas observed in fully developed CL, but there was no dif-ference in GHS-R1a expression among different stages of CLas observed by Rak-Mardyla et al [18]. Similarly, Caminoset al [15] in rat and Zhang et al [32] in pig demonstratedthat ghrelin mRNA expression varied depending on thestage of the estrous cycle, with the greatest expressionduring diestrus. Ghrelin and GHS-R1a mRNA and proteinwere also detected in bovine CL [19]. Local ghrelin ovarianconcentration varied from 109 to 148 pg/mL during theestrous cycle as reported earlier in porcine species [20].
Results of mRNA and protein expression were also sup-ported by the localization of ghrelin and GHS-R1a proteinby immunohistochemistry. Ghrelin and GHS-R1a immu-noreactivity was observed in cyotoplasm of small and largeluteal cells. The intensity of immunostaining and number ofluteal cells observed containing ghrelin and GHS-R1a pro-tein in the present study increased during maturation anddecreased during regression of CL. These findings are inagreement with observations of Rak-Mardyla et al [18] inwhich increased immunoreactivity was observed with CLdevelopment. Similarly, in rat, Caminos et al [15] detectedspecific areas of intense ghrelin immunoreactivity in thecytoplasm of steroidogenic luteal cells during the diestrusphase which is in agreement with our findings. Also Gaytanet al [14] detected ghrelin protein in young (15–19 d of thecycle) and mature (20–24 d of the cycle) human lutealtissue, taking into consideration that day 28 of the men-strual cycle in the woman corresponds to late luteal phase.Localization of ghrelin and its receptor have also been re-ported in sheep [16,32], pig [33], and bovine ovary [19].
Expression and localization of ghrelin and its receptor inbubaline CL observed in the present study provides indirectevidence for autocrine and paracrine roles of ghrelin inluteal function. However, this evidencewas not sufficient todraw a conclusive inference about the role of ghrelin in theprocess. Therefore, in the second part of our study, anin vitro luteal cell culture model was used to evaluate theeffect of ghrelin on P4 secretion, mRNA expression of P4synthesis intermediates, and localization of CYP11A1 and3b-HSD proteins in luteal cell culture. At a ghrelin dose of1 ng/mL, there was no effect on concentrations of P4 or onthe expression of 3b-HSD and CYP11A1mRNA; however, 10and 100 ng/mL doses significantly inhibited P4 concentra-tion and 3b-HSD and CYP11A1 mRNA expression. Ourresults are in agreement with the findings obtained by Rak-Mardyla et al [18] in the pig that ghrelin inhibits P4secretion and 3b-HSD protein expression in luteal cells. Itwas proposed that ghrelin reduced P4 secretion by theinhibition of 3b-HSD enzyme activity and protein expres-sion [18]. Also, in human, it was observed that ghrelinreduced P4 secretion, decreased luteotropic PGE2 release,and increased luteolytic PGF2a release, suggesting that theimbalance between luteotropic and luteolytic factors couldbe a mechanism by which ghrelin negatively influencesluteal function [25].
Similarly, Viani et al [34] also found an inhibitory effectof ghrelin on granulosa luteal cells. In the present study, theinhibitory effect of ghrelin on P4 secretion could be due todecreased expression of 3b-HSD and CYP11A1, which areimportant mediators of the P4 biosynthetic pathway.Ghrelin also has other functions, including effects on cellproliferation, antiapoptotic actions, and modulation ofinsulin-like growth factor (IGF)-I [27,28]; therefore,increased ghrelin expression in fully developed CL ascompared with regressing CL, as observed in the in vivostudy, could be involved in these functions within the CL.Under in vivo conditions, a number of stimulatory andinhibitory factors work together for a particular functionand the resultant effect depends on the balance of them.However, under in vitro conditions, the effect of a singlefactor is demonstrated and, therefore, differences betweenin vitro and in vivo conditions are likely. A number oflocally produced growth factors such as IGF-I, IGF-II,vascular endothelial growth factor, basic fibroblast growthfactor, epidermal growth factor, tumor necrosis factoralpha, interferon gamma, Endothelin1, and leptin affect P4production by CL in addition to gonadotropins in animals[35–37]. Of the aforementioned factors, IGF-I [38], IGF-II[39], vascular endothelial growth factor [40], epidermalgrowth factor [41], and leptin [42] are stimulatory, whereasfibroblast growth factor [43], tumor necrosis factor alpha[44], and interferon gamma [45] have been reported toinhibit P4 production although their expression level wasalso reported to be greater in fully mature CL [18,46–48].Similarly, ghrelin, which exhibits greatest expression inmature buffalo CL, also suppresses P4 synthesis and pro-duction as evidenced from our in vitro study. Hence, theincreased expression of ghrelin and increased productionof P4 in fully functional CL may not be related.
At the systemic level, an inhibitory effect of ghrelin onthe hypothalamic-pituitary axis has been reported in the

M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–32 31
human, rat, and pig [22,36]. Ghrelin inhibits gonadotropin-releasing hormone release, luteinizing hormone re-sponsiveness to gonadotropin-releasing hormone, andconcentrations of circulating follicle-stimulating hormoneand luteinizing hormone in several species [49,50]. Ghrelinis also an important regulator of energy balance. Becausethe reproductive axis is highly dependent on body energystatus, ghrelin, by acting both at the central and the pe-ripheral level, appears to inhibit reproduction during timesof negative energy balance. The present findings in buffaloalso indicate an inhibitory effect of ghrelin at the local levelrelative to several aspects of luteal function.
Although the reproductive actions of ghrelin and theirmechanisms require further study, the present data showfor the first time that ghrelin has a role in the regulation offemale buffalo reproductive function. The present studydemonstrated the dynamics of expression and localizationof ghrelin and its functional receptor in buffalo CL duringthe luteal phase and an inhibitory effect on P4 productionand expression of P4 synthetic intermediates in luteal cellculture. These observations suggest a possible regulatoryrole for the ghrelin in luteal development, function, andregression.
Acknowledgments
Funding support received from Department of Biotech-nology and Department of Science and Technology, Gov-ernment of India and Director Indian Veterinary ResearchInstitute are gratefully acknowledged.
Conflicts of interest: The authors declare that there is noconflict of interest on publishing the article.
References
[1] Food and Agriculture Organization of the United Nations.FAOSTAT. <http://faostat.fao.org/site/573/DesktopDefault.aspx?PageID¼573#ancor>; 2011 [accessed 02.02.14].
[2] Prakash BS. Influence of climate on animal reproduction. Izatnagar,India National Workshop on animal-Climate Interaction, IVRI; 2002.
[3] Madan ML, Prakash BS. Reproductive endocrinology and biotech-nology applications among buffaloes. Soc Reprod Fertil Suppl 2007;64:261–81.
[4] Stocco C, Telleria C, Gibori G. The molecular control of corpusluteum formation, function, and regression. Endocr Rev 2007;28:117–49.
[5] Berisha B, Schams D. Ovarian function in ruminants. Domest AnimEndocrinol 2005;29:305–17.
[6] Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K.Ghrelin is a growth hormone-releasing acylated peptide fromstomach. Nature 1999;402:656–60.
[7] Howard AD, Feighner SD, Cully DF, Arena JP, Liberator PA,Rosenblum CI, et al. A receptor in pituitary and hypothalamus thatfunctions in growth hormone release. Science 1996;273:974–7.
[8] Van der Lely AJ, Tschop M, Heiman ML, Ghigo E. Biological, physi-ological, pathophysiological, and pharmacological aspects of ghrelin.Endocr Rev 2004;25:426–57.
[9] Gualillo O, Lago F, Omez-Reino JG, Casanueva FF, Dieguez C. Ghrelin,a widespread hormone: insights into molecular and cellular regu-lation of its expression and mechanism of action. FEBS Lett 2003;552:105–9.
[10] Fernandez-Fernandez R, Tena-Sempere M, Aguilar E, Pinilla L.Ghrelin effects on gonadotropin secretion in male and female rats.Neurosci Lett 2004;362:103–7.
[11] Fernandez-Fernandez R, Tena-Sempere M, Navarro VM,Barreiro ML, Castellano JM, Aguilar E, Pinilla L. Effects of ghrelinupon gonadotropin-releasing hormone and gonadotropin secretion
in adult female rats: in vivo and in vitro studies. Neuroendocri-nology 2005;82:245–55.
[12] Fernandez-Fernandez R, Martini AC, Navarro VM, Castellano JM,Dieguez C, Aguilar E, Pinilla L, Tena-Sempere M. Novel signals forthe integration of energy balance and reproduction. Mol CellEndocrinol 2006;254-255:127–32.
[13] Fernandez-Fernandez R, Tena-Sempere M, Roa J, Castellano JM,Navarro VM, Aguilar E, Pinilla L. Direct stimulatory effect of ghrelinon pituitary release of LH through a nitric oxide-dependent mech-anism that is modulated by estrogen. Reproduction 2007;133:1223–32.
[14] Gaytan F, Barreiro ML, Chopin LK, Herington AC, Morales C, Pinilla L,Casanueva FF, Aguilar E, Dieguez C, Tena-Sempere M. Immunoloc-alization of ghrelin and its functional receptor, the type 1a growthhormone secretagogue receptor, in the cyclic human ovary. J ClinEndocrinol Metab 2003;88:879–87.
[15] Caminos JE, Tena-Sempere M, Gaytan F, Sanchez-Criado JE,Barreiro ML, Nogueiras R, Casanueva FF, Aguilar E, Dieguez C.Expression of ghrelin in the cyclic and pregnant rat ovary. Endo-crinology 2003;144:1594–602.
[16] Du C, Xilingaowa Cao G, Wang C, Li H, Zhao Y, Siqingaowa, Cao J.Expression of orexigenic peptide ghrelin in sheep ovary. Dom AnimEndocrinol 2009;36:89–98.
[17] Chandra V, Ram H, Sharma AK. Expression profile of ghrelin andghrelin receptor in cyclic goat ovary. Indian J Anim Sci 2012;82:984–7.
[18] Rak-Mardyla A, Gregoraszczuk EL, Karpeta A, Dud M. Expression ofghrelin and the ghrelin receptor in different stages of porcinecorpus luteum development and the inhibitory effects of ghrelin onprogesterone secretion, 3b-hydroxysteroid dehydrogenase (3b-honestly significant difference (HSD)) activity and protein expres-sion. Theriogenology 2012;77:1505–12.
[19] Deaver SE, Hoyer PB, Dial SM, Field ME, Collier RJ, Rhoads ML.Localization of ghrelin and its receptor in the reproductive tract ofHolstein heifers. J Dairy Sci 2013;96:150–7.
[20] Rak-Mardyla A, Gregoraszczuk EL. Expression of ghrelin and itsreceptor in porcine ovarian follicles collected from prepubertal andestrous cycle animals. J Physiol Pharmacol 2012;63:195–9.
[21] Wertz-Lutz AE, Knight TJ, Pritchard RH, Daniel JA, Clapper JA,Smart AJ, Trenkle A, Beitz DC. Circulating ghrelin concentrationsfluctuate relative to nutritional status and influence feedingbehavior in cattle. J Anim Sci 2006;84:3285–300.
[22] Miller WL. Molecular biology of steroid hormone synthesis. EndocrRev 1988;9:295–318.
[23] Stucco DM, Clark BJ. Regulation of the acute production of steroidsin steroidogenic cells. Endocr Rev 1996;17:221–44.
[24] Cherradi N, Rossier MF, Vallotton MB, Timberg R, Friedberg I,Orly J, Wang XJ, Stocco DM, Capponi AM. Submitochondrial dis-tribution of three key steroidogenic proteins (steroidogenic acuteregulatory protein and cytochrome P450scc and 3b-hydroxyste-roid dehydrogenase isomerase enzymes) upon stimulation byintracellular calcium in adrenal glomerulosa cells. J Biol Chem1997;272:7899–907.
[25] Tropea A, Tiberi F, Minici F, Orlando M, Gangale MF, Romani F,Miceli F, Catino S, Mancuso S, Sanguinetti M, Lanzone A, Apa R.Ghrelin affects the release of luteolytic and luteotropic factors inhuman luteal cells. J Clin Endocrinol Metab 2007;92:3239–45.
[26] Sirotkin AV, Grossmann R. Effects of ghrelin and its analogues onchicken ovarian granulosa cells. Domest Anim Endocrinol 2008;34:125–34.
[27] Sirotkin AV, Meszarosova M. Comparison of effects of leptin andghrelin on porcine ovarian granulosa cells. Domest Anim Endocrinol2010;39:1–9.
[28] Sirotkin AV, Rafay J, Kotwica J, Darlak K, Valenzuela F. Role of ghrelinin regulating rabbit ovarian function and the response to LH andIGF-I. Domest Anim Endocrinol 2009;36:162–72.
[29] Sarkar M, Schilffarth S, Schams D, Meyer HH, Berisha B. Theexpression of leptin and its receptor during different physiologicalstages in the bovine ovary. Mol Reprod Dev 2010;77:174–81.
[30] Kumar L, Panda RP, Hyder I, Yadav VP, Sastry KVH, Sharma GT,Mahapatra RK, Bag S, Bhure SK, Das GK, Mitra A, Sarkar M.Expression of leptin and its receptor in corpus luteum duringestrous cycle in buffalo (Bubalus bubalis). Anim Reprod Sci 2012;135:8–17.
[31] Pfaffl M. A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res 2001;29:2002–7.
[32] Miller DW, Harrison JL, Brown YA, Doyle U, Lindsay A, Adam CL,Lea RG. Immuno histochemical evidence for an endocrine/paracrine

M. Gupta et al. / Domestic Animal Endocrinology 48 (2014) 21–3232
role for ghrelin in the reproductive tissues of sheep. Reprod BiolEndocrinol 2005;3:60.
[33] Zhang W, Lei Z, Su J, Chen S. Expression of ghrelin in the porcinehypothalamo–pituitary–ovary axis during the estrous cycle. AnimReprod Sci 2008;109:356–67.
[34] Viani I, Vottero A, Tassi F, Cremonini G, Sartori C, Bernasconi S,Ferrari B, Ghizzoni L. Ghrelin inhibits steroid biosynthesis bycultured granulosa-lutein cells. J Clin Endocrinol Metab 2008;93:1476–81.
[35] Muccioli G, Lorenzi T, Lorenzi M, Ghe C, Arnoletti E, Raso GM,Castellucci M, Gualillo O, Meli R. Beyond the metabolic role ofghrelin: a new player in the regulation of reproductive function.Peptides 2011;32:2514–21.
[36] Webb R, Woad KJ, Armstrong DG. Corpus luteum (CL) function: localcontrol mechanisms. Domest Anim Endocrinol 2002;23:277–85.
[37] Miyamoto A, Shirasuna K, Sasahara K. Local regulation of corpusluteum development and regression in the cow: Impact of angio-genic and vasoactive factors. Domest Anim Endocrinol 2009;37:159–69.
[38] McArdle CA, Holtorf A-P. Oxytocin and progesterone release frombovine corpus luteal cells in culture: effects of insulin-likegrowth factor I, insulin, and prostaglandins. Endocrinology 1989;124:1278–86.
[39] Sauerwein H, Miyamoto A, Gunther J, Meyer HHD, Schams D.Binding and action of insulin-like growth factors and insulin inbovine luteal tissue during the oestrous cycle. J Reprod Fertil Suppl1992;96:103–15.
[40] Kashida S, Sugino N, Takiguchi S, Karube A, Takayama H,Yamagata Y, Nakamura Y, Kato H. Regulation and role of vascularendothelial growth factor in the corpus luteum during mid-pregnancy in rats. Biol Reprod 2001;64:317–23.
[41] Serta RT, Seibel MM. The influence of epidermal growth factor onprogesterone production by human granulosa–luteal cells in cul-ture. Hum Reprod 1993;8:1005–10.
[42] Nicklin LT, Robinson RS, Marsters P, Campbell BK, Mann GE,Hunter MG. Leptin in the bovine corpus luteum: receptor expres-sion and effects on progesterone production. Mol Reprod Dev 2007;74:724–9.
[43] Spicer LJ, Stewart RE. Interactions among basic fibroblast growthfactor, epidermal growth factor, insulin, and insulin-like growthfactor-I (IGF-I) on cell numbers and steroidogenesis of bovine thecalcells: role of IGF-I receptors. Biol Reprod 1996;54:255–63.
[44] Benyo DF, Pate JL. Tumor necrosis factor-alpha alters bovine lutealcell synthetic capacity and viability. Endocrinology 1992;130:854–60.
[45] Chen XL, Gao HJ, Wei P, Song XX, Hu ZY, Liu YX. Regulatory effect ofIFN-g on expression of TGF-b1, TbR-II, and StAR in corpus luteum ofpregnant rhesus monkey. Acta Pharmacol Sin 2003;24:435–41.
[46] Schams D, Berisha B. Regulation of corpus luteum function in cat-tledan overview. Reprod Dom Anim 2004;29:241–51.
[47] Sakumoto R, Berisha B, Kawate N, Schams D, Okuda K. Tumor ne-crosis factor-aand its receptor in bovine corpus luteum throughoutthe estrous cycle. Biol Reprod 2000;62:192–9.
[48] Petroff MG, Petroff BK, Pate JL. Expression of cytokine messengerribonucleic acid in the bovine corpus luteum. Endocrinology 1999;140:1018–21.
[49] Garcia MC, Lopez M, Alvarez CV, Casanueva F, Tena-Sempere M,Dieguez C. Role of ghrelin in reproduction. Reproduction 2007;133:531–40.
[50] Lorenzi T, Meli R, Marzioni D, Morroni M, Baragli A, Castellucci M,et al. Ghrelin: a metabolic signal affecting the reproductive system.Cytokine Growth Factor Rev 2009;20:137–52.




















