
Expression and activity of cyclic and linear analogues of esculentin-1, an anti-microbial peptide...
7
Eur. J. Biochem. 263, 921–927 (1999) q FEBS 1999 Expression and activity of cyclic and linear analogues of esculentin-1, an anti-microbial peptide from amphibian skin Donatella Ponti 1 , Giuseppina Mignogna 1 , Maria L. Mangoni 2 , Daniela De Biase 1 , Maurizio Simmaco 2 and Donatella Barra 1 1 Dipartimento di Scienze Biochimiche ‘A. Rossi Fanelli’ and CNR, Centro di Biologia Molecolare, Universita ` La Sapienza, Roma, Italy; 2 Dipartimento di Scienze Biomediche, Universita ` ‘G. D’Annunzio’, Chieti, Italy Esculentin-1 is a potent anti-microbial peptide present in minute amounts in skin secretions of Rana esculenta. It contains 46 amino-acid residues and a C-terminal disulfide bridge. We have explored the possibility of producing analogues of this peptide by recombinant expression in Escherichia coli of a fusion protein which is sequestered in inclusion bodies. The peptide of interest has been inserted at the N-terminus of the protein, from which it can be released by cyanogen bromide cleavage. The anti-microbial activities of the recombinant peptide as well as that of a mutant linear form devoid of the disulfide bridge are presented. The recombinant analogues retain the biological activity of the natural peptide, as tested with an inhibition zone assay against a variety of microorganisms. However, experiments on the rate of bacterial killing show that gram-negative bacteria are more sensitive to the peptides than the gram-positive bacterium, the effect of the cyclic peptide being in all cases faster than that of the linear molecule. Moreover, the activity against gram-negative bacteria for both peptides is not affected by salts, whereas the activity against Staphylococcus aureus is lost at high salt concentration. Keywords: anti-microbial peptide; intramolecular disulfide; peptide expression; inclusion bodies; salt inactivation. Multicellular organisms are equipped with a variety of anti- microbial peptides representing important components of the innate immunity system, the first line of defense against invading microorganisms [1,2]. The numerous peptides described so far can be grouped in several structural families, totally unrelated from the evolutionary point of view. Each family is characterized by specific structural features such as size, occurrence of none, one or more disulfide bridges, predominance of some amino-acid residues (e.g. arginine or proline). However, despite such heterogeneity, all of them share common properties, such as an overall cationic character and the propensity to adopt a helical conformation often resulting in an amphipathic behaviour. These properties are believed to play important roles in the interaction of the peptides with the membrane of target cells and/or in the mechanism that eventually effects cell lysis [3,4]. A major goal in this field is represented by a more in-depth understanding of the mechanism of action of anti-microbial peptides, with special attention to the structural basis for their selective specificity (i.e. the ability to distinguish between prokaryotic versus eukaryotic cells, gram-positive versus gram- negative bacteria, the various bacterial strains, etc.). Advances of knowledge along these lines may have important con- sequences on suitability of natural anti-microbial peptides or their analogues for a number of practical applications, such as treatment of infections resistant to classical antibiotics [5]. In this context, it should be pointed out that in the various organisms and/or tissues, anti-microbial peptides are only rarely present as a unique molecule, but more frequently as a variety of forms, differing from each other by subtle structural features. This microheterogeneity is often underestimated because of the difficulties in achieving complete separation of the different components: some of them are present in minute amounts, others are only predicted from indirect genetic information. Obviously, availability of sufficient amounts of each component of a certain family of anti- microbial peptides in absolutely pure form would be highly desirable. It should also be pointed out that these peptides are synthesized on ribosomes, in contrast with other classical antibiotics from bacterial sources which are assembled by enzymatic machineries. Thus, although small peptides are easily made by chemical synthesis, the production of larger molecules can be achieved through recombinant DNA proce- dures [6–8]. We decided to exploit this strategy for the production of peptides belonging to the esculentin-1 family, which comprises a 46 amino-acid residue anti-microbial peptides from the skin secretion of Rana esculenta [9]. Esculentins are particularly suited to this approach, as they are the largest anti-microbial peptides isolated so far from amphibian skin, are recovered from the secretion in minute amounts and have been uniquely found in this Rana species. Moreover, esculentins are highly potent anti-microbial mol- ecules, characterized by the most favorable spectrum of action with negligible effects on eukaryotic cell membranes. Further- more, the structure of one of the precursors has been described and the corresponding cDNA is available [9]. Here we describe the production of an esculentin analogue and of a mutant form devoid of the C-terminal disulfide bridge (Fig. 1), both obtained following cyanogen bromide cleavage of a fusion protein expressed in Escherichia coli. This protein is Correspondence to D. Barra, Dipartimento di Scienze Biochimiche, Universita ` La Sapienza, Piazzale Aldo Moro 5, 00185 Roma, Italy, Fax: + 39 06 4440062, E-mail: [email protected] Abbreviations: GABA, 4-aminobutyrate; GABA-T, GABA transaminase; Gal-ONp, 2-nitrophenyl b-d galactoside. (Received 30 March 1999, revised 20 May 1999, accepted 27 May 1999)
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Expression and activity of cyclic and linear analogues of esculentin-1, an anti-microbial peptide...

Eur. J. Biochem. 263, 921±927 (1999) q FEBS 1999
Expression and activity of cyclic and linear analogues of esculentin-1,an anti-microbial peptide from amphibian skin
Donatella Ponti1, Giuseppina Mignogna1, Maria L. Mangoni2, Daniela De Biase1, Maurizio Simmaco2 andDonatella Barra1
1Dipartimento di Scienze Biochimiche `A. Rossi Fanelli' and CNR, Centro di Biologia Molecolare, UniversitaÁ La Sapienza, Roma, Italy;2Dipartimento di Scienze Biomediche, UniversitaÁ `G. D'Annunzio', Chieti, Italy
Esculentin-1 is a potent anti-microbial peptide present in minute amounts in skin secretions of Rana esculenta. It
contains 46 amino-acid residues and a C-terminal disulfide bridge. We have explored the possibility of producing
analogues of this peptide by recombinant expression in Escherichia coli of a fusion protein which is sequestered
in inclusion bodies. The peptide of interest has been inserted at the N-terminus of the protein, from which it can
be released by cyanogen bromide cleavage. The anti-microbial activities of the recombinant peptide as well as
that of a mutant linear form devoid of the disulfide bridge are presented. The recombinant analogues retain the
biological activity of the natural peptide, as tested with an inhibition zone assay against a variety of
microorganisms. However, experiments on the rate of bacterial killing show that gram-negative bacteria are more
sensitive to the peptides than the gram-positive bacterium, the effect of the cyclic peptide being in all cases faster
than that of the linear molecule. Moreover, the activity against gram-negative bacteria for both peptides is not
affected by salts, whereas the activity against Staphylococcus aureus is lost at high salt concentration.
Keywords: anti-microbial peptide; intramolecular disulfide; peptide expression; inclusion bodies; salt
inactivation.
Multicellular organisms are equipped with a variety of anti-microbial peptides representing important components of theinnate immunity system, the first line of defense againstinvading microorganisms [1,2]. The numerous peptidesdescribed so far can be grouped in several structural families,totally unrelated from the evolutionary point of view. Eachfamily is characterized by specific structural features such assize, occurrence of none, one or more disulfide bridges,predominance of some amino-acid residues (e.g. arginine orproline). However, despite such heterogeneity, all of them sharecommon properties, such as an overall cationic character andthe propensity to adopt a helical conformation often resulting inan amphipathic behaviour. These properties are believed to playimportant roles in the interaction of the peptides with themembrane of target cells and/or in the mechanism thateventually effects cell lysis [3,4].
A major goal in this field is represented by a more in-depthunderstanding of the mechanism of action of anti-microbialpeptides, with special attention to the structural basis for theirselective specificity (i.e. the ability to distinguish betweenprokaryotic versus eukaryotic cells, gram-positive versus gram-negative bacteria, the various bacterial strains, etc.). Advancesof knowledge along these lines may have important con-sequences on suitability of natural anti-microbial peptides ortheir analogues for a number of practical applications, such astreatment of infections resistant to classical antibiotics [5].
In this context, it should be pointed out that in the variousorganisms and/or tissues, anti-microbial peptides are onlyrarely present as a unique molecule, but more frequently as avariety of forms, differing from each other by subtle structuralfeatures. This microheterogeneity is often underestimatedbecause of the difficulties in achieving complete separationof the different components: some of them are present inminute amounts, others are only predicted from indirectgenetic information. Obviously, availability of sufficientamounts of each component of a certain family of anti-microbial peptides in absolutely pure form would be highlydesirable. It should also be pointed out that these peptidesare synthesized on ribosomes, in contrast with other classicalantibiotics from bacterial sources which are assembled byenzymatic machineries. Thus, although small peptides areeasily made by chemical synthesis, the production of largermolecules can be achieved through recombinant DNA proce-dures [6±8]. We decided to exploit this strategy for theproduction of peptides belonging to the esculentin-1 family,which comprises a 46 amino-acid residue anti-microbialpeptides from the skin secretion of Rana esculenta [9].Esculentins are particularly suited to this approach, as theyare the largest anti-microbial peptides isolated so far fromamphibian skin, are recovered from the secretion in minuteamounts and have been uniquely found in this Rana species.Moreover, esculentins are highly potent anti-microbial mol-ecules, characterized by the most favorable spectrum of actionwith negligible effects on eukaryotic cell membranes. Further-more, the structure of one of the precursors has been describedand the corresponding cDNA is available [9].
Here we describe the production of an esculentin analogueand of a mutant form devoid of the C-terminal disulfide bridge(Fig. 1), both obtained following cyanogen bromide cleavage ofa fusion protein expressed in Escherichia coli. This protein is
Correspondence to D. Barra, Dipartimento di Scienze Biochimiche,
UniversitaÁ La Sapienza, Piazzale Aldo Moro 5, 00185 Roma, Italy,
Fax: + 39 06 4440062, E-mail: [email protected]
Abbreviations: GABA, 4-aminobutyrate; GABA-T, GABA transaminase;
Gal-ONp, 2-nitrophenyl b-d galactoside.
(Received 30 March 1999, revised 20 May 1999, accepted 27 May 1999)

922 D. Ponti et al. (Eur. J. Biochem. 263) q FEBS 1999
constituted by an N-terminal part of 47 amino-acid residues,which corresponds to esculentin-1b with a Leu substituting forthe Met in position 28 and an additional methionine in position47 (or a mutant form in which the cysteine residues aresubstituted by Ser), and a C-terminal one corresponding tohuman GABA transaminase (GABA-T) (Fig. 2). The cDNAcoding for this enzyme was cloned in our laboratory and thecorresponding protein was found to be sequestered in inclusionbodies, following its expression in E. coli. We intended to takeadvantage of this feature for a rapid purification of the fusionprotein before cyanogen bromide treatment. In addition, theanti-microbial activity of the esculentin analogues produced inthis way was studied in detail with respect to the natural form.
E X P E R I M E N T A L P R O C E D U R E S
Mutagenesis of esculentin-1b
The DNA encoding the esculentin-1b analogue was obtained byPCR from pBlueScript-KS-Esc1-17-1 [9] using two oligo-nucleotide pairs. The region 1±28 of esculentin was amplifiedwith forward primer RX3
(50-CAGAATTCATGGGAATCTTCTCCAAACTGG-30), whichintroduces an EcoRI site and a Met codon at the 5 0-end of thegene, and reverse primer RY7 (5 0-CAGACCAACTTCCT-TGCCTAC-3 0), introducing the mutation Met28Leu. Theregion 29±46 was amplified using forward primer RY13(5 0-GACGTGGTCAGAACTGGCATA-3 0) and reverse primerEsc-dw (5 0-GAGTCGACCATACATTCAACCTTT-3 0), whichintroduces a Met codon in position 47 and a SalI site. To obtainan esculentin mutant in which the two cysteine residues weresubstituted by Ser, a suitably modified Esc-dw oligonucleotidewas used
(5 0-GAGTCGACCATAGATTCACCTTTAATTTTAGAA-3 0).The PCR products were ligated into the EcoRI and SalIrestriction sites of pBlueScript-SK. The construct was furtheramplified using forward primer Esc-up (5 0-GACATATGG-GAATCTTCTCCAAACTG-3 0), which introduces a NdeI site,and Esc-dw (see above).
Subcloning into the expression vector
For the expression of the esculentin analogues, the pHGTconstruct was used. It consists of a 1422-bp PCR-generatedfragment encoding the mature human GABA-T [10], direc-tionally subcloned into the expression vector pET22b (Strata-gene) at its NdeI and HindIII sites. This construct allows aconsistent overexpression of human GABA-T, but in aninsoluble form (D. De Biase et al., unpublished results).The engineered coding sequence of esculentin-1b wasinserted into pHGT at the NdeI±SalI restriction sites, atthe N-terminal region of the human GABA-T (Fig. 2). Thefidelity of the pET-Esc-GABA-T construct was confirmedby DNA sequencing.
Protein expression and isolation of inclusion bodies
For the expression of the fusion protein, E. coli BL21(DE3)/pLys was transformed with pET-Esc-GABA-T, andtranscription was induced by 1 mm isopropyl thio-b-d-galacto-side at 37 8C for 4 h. Bacterial cells were thawed on ice andlysed by sonication. The inclusion bodies were separated bycentrifugation and washed essentially according to [11].
Purification of the esculentin analogues
The inclusion bodies were resuspended in 80% trifluoroaceticacid (0.1 g´mL21) and treated with cyanogen bromide(50 mg´mL21) overnight at room temperature in the dark.After dialysis and lyophilization, the material soluble in 10%acetic acid was fractionated on a Sephadex G-50 column(2.5 � 60 cm) equilibrated and eluted with the same solvent.Fractions with anti-bacterial activity were pooled and furtherpurified by reversed-phase HPLC as reported [9].
Sequence and mass spectral analyses
N-terminal sequence analysis of the fusion protein wasperformed with a Perkin Elmer AB476A sequencer on a
Fig. 1. Amino acid sequences of natural
esculentins and analogues. Esculentin-1a,
natural peptide [9]; esculentin-1b, putative
peptide predicted from cDNA [9]; esculentin-C,
cyclic peptide; esculentin-L, linear peptide.
Substitution positions are boldfaced.
Cysteines are linked in a disulfide bridge.
M*, homoserine/homoserine lactone.
Fig. 2. Scheme of the construct used for the expression of the fusion protein esculentin-GABA transaminase (pET-Esc-GABA-T). The cDNA coding
for esculentin-1b [9], suitably mutated, was cloned upstream of the GABA-T coding region into the NdeI and SalI restriction sites.

q FEBS 1999 Recombinant expression of esculentin-1 (Eur. J. Biochem. 263) 923
sample electrotransferred onto ProBlott membrane (PerkinElmer) after SDS/PAGE. Sequence analysis of purified peptideswas performed on samples spotted on ProBlott membranes andcoated with polybrene (2 mL, 0.1 mg´mL21 in 50% methanol).The molecular mass of the purified peptides was determined byelectrospray ionization ion-trap mass spectrometry, using aFinnigan LCQ instrument.
Circular dichroism measurements
CD spectra were obtained on a Jasco J710 spectropolarimeter,equipped with a DP 520 processor, at 20 8C, using a quartz cellof 2-mm pathlength. Spectra were the average of a series of 3±7scans. Ellipticity is reported as the mean molar residueellipticity (u) expressed in deg´cm2´dmol21. Peptide concentra-tions were determined by quantitative amino-acid analysis.
Biological activity assays
The microorganisms used were: Bacillus megaterium Bm11,E. coli D21, and Staphylococcus aureus CowanI; Enterobacteragglomerans Bo-1S isolated from Bombina orientalis [12],Pseudomonas syringae pv. tabaci, Yersinia pseudotuberculosisYPIII, Pseudomonas aeruginosa ATCC15692 and Candidaalbicans.
The anti-bacterial activity was evaluated using an inhibitionzone assay on Luria±Bertani (LB)-agarose plates [9,13]. ForC. albicans, Winge medium (0.3% yeast extract, 0.2% glucose,pH 6.2) broth was used. The hemolytic activity was measuredusing a modification of the anti-bacterial inhibition zone assay[14]. Hemolytic capacity was also evaluated in liquid medium(0.9% NaCl) using 100 mL of a human red cell suspensionwhich after complete lysis in distilled water gives D500 nm of 0.5.
To study the bactericidal effect of the esculentin analoguesand the rate of killing, the peptides were added to a bacterialsuspension and the number of surviving bacteria was followedat different incubation times. P. syringae, P. aeruginosa andS. aureus were grown at 30 8C in LB medium to approximatelyD590 nm of 1. P. syringae and P. aeruginosa were diluted toabout 1 � 105 cells´mL21 in NaCl/Pi; S. aureus was alsodiluted to 1 � 105 cells´mL21 in phosphate buffer at differentNaCl concentrations. Peptides to be assayed were added to afinal concentration of 0.6 or 4 mm. Ten-microliter aliquots werewithdrawn at different intervals and spread on agar plates. Afterovernight incubation at 30 8C, the surviving bacteria, expressedas colony-forming units, were counted.
Inner membrane permeability was determined by measuringthe b-galactosidase activity using 2-nitrophenyl b-d galactoside(Gal-ONp, Sigma) as substrate according to [15]. E. coli D21(1 � 107 cells) were incubated for 90 min at 30 8C with 20 mmof each peptide. The bacterial culture was then passed through a0.2-mm filter and the hydrolysis of Gal-ONp was recorded onthe filtrate at 420 nm.
R E S U LT S
Expression of a cyclic esculentin analogue
The cDNA clone Esc1-17-1 encoding the esculentin-1bprecursor has been isolated previously from a R. esculentaskin library [9]. This clone was used as template to produce anengineered esculentin gene. The 5 0 and 3 0 regions of this genewere amplified by PCR using two pairs of oligonucleotides,RX3/RY7, and RY13/Esc-dw, respectively. The DNA fragmentswere subcloned into pBlueScript-SK in the correct reading
frame. This cDNA was inserted into the NdeI and SalIrestriction sites of the pET-GABA-T vector, upstream of theGABA-T coding region (Fig. 2). The vector was transformedinto E. coli BL21(DE3)/pLys host and the expressed fusionprotein (58 kDa) was found to accumulate in insolubleinclusion bodies. A second band of about 53 kDa was identifiedby automated Edman degradation as GABA-T. The simul-taneous production of the two proteins was due to the presenceof a second Shine±Dalgarno sequence [16,17] in the nucleotidesequence of esculentin. However, the level of GABA-Texpression was lower than that of the fusion protein.
Inclusion bodies (300 mg´L21 of bacterial culture) weretreated with cyanogen bromide to cleave the esculentinanalogue (esculentin-C) from GABA-T. After gel-filtration,fractions active against E. coli D21 were pooled and purified byRP-HPLC (Fig. 3). Sequence analysis of the active peaks(Fig. 3, peaks a and b) confirmed the expected structure. Massspectral analysis gave molecular masses of 4884.5 and 4866.1,respectively, corresponding to the homoserine/homoserinelactone containing peptides. The latter form accounts for 90%of the total. The purification yield was about 0.5 mg per litreculture.
Expression of an esculentin linear mutant
In order to produce an esculentin mutant lacking the disulfidebridge (esculentin-L), a different construct was designed,coding for a 46-residue peptide with a serine residuesubstituting for the cysteine in position 40 and a C-terminalmethionine substituting for Cys46. After cyanogen bromidecleavage of the expressed fusion protein, a peptide containing ahomoserine in position 46 should be produced. Expression ofthis construct yielded only the fusion protein but no GABA-T.This was possibly due to the fact that the deletion of one codonaltered the second Shine±Dalgarno sequence. Surprisingly, inthis case the fusion protein was soluble in the cytoplasm and noinclusion bodies were detectable. Therefore, we decided toproduce a GABA-T fusion protein identical to that used for theexpression of the cyclic analogue, except that both cysteine
Fig. 3. Reversed-phase HPLC purification of esculentin-C obtained by
cyanogen bromide cleavage of the fusion protein. The pooled active
fractions from Sephadex G-50 column were purified using an Acquapore
RP-300 column (7.0 � 250 mm, Perkin Elmer), eluted with a 60-min linear
gradient of 5±75% acetonitrile/isopropyl alcohol (4 : 1) in 0.2% (v/v)
trifluoroacetic acid, at a flow rate of 2 mL´min21. Peak a, esculentin-C with
a C-terminal homoserine; peak b, esculentin-C with a C-terminal
homoserine lactone.

924 D. Ponti et al. (Eur. J. Biochem. 263) q FEBS 1999
residues in positions 40 and 46 were replaced by Ser. In thiscase, inclusion bodies were formed which contained both thefusion protein and the human enzyme. The same proceduredescribed above was applied for the cleavage and purificationof the mutant peptide, except that, prior to treatment withcyanogen bromide, inclusion bodies were incubated in thepresence of 0.1% 2-mercaptoethanol for 2 h at 45 8C (seeDiscussion). The HPLC profile was similar to that reportedin Fig. 3, esculentin-L being eluted 1 min earlier thanesculentin-C. Mass spectral analysis gave molecular massesof 4854.8/4834.1 for the homoserine/homoserine lactonecontaining peptides, respectively, with the same relativeabundance as above. The yield of the linear analogue wasabout 1 mg per litre culture.
Both cyclic and linear esculentins were stable to carboxy-peptidase Y digestion in the pH range 6±8, as observed for thenatural peptide, thus confirming the prevalence of the lactoneform.
Secondary structure of esculentin analogues
Conformational studies using CD spectroscopy showed that,upon increasing the trifluoroethanol concentration, bothesculentin anologues undergo a gradual conformational change
Fig. 4. Circular dichroism spectra of esculentin-C (panel A) and
esculentin-L (panel B) at different trifluoroethanol concentrations.
Peptide concentrations, 13 mm.
Table 1. Anti-microbial and lytic activity of esculentin analogues.
Lethal concentration (mm)
Strains Esculentin-1a Esculentin-C Esculentin-L
Bacillus megaterium Bm11 0�.1 0�.1 0�.1
Staphylococcus aureus CowanI 0�.4 0�.4 1�.5
Escherichia coli D21 0�.2 0�.2 0�.2
Yersinia pseudotuberculosis YPIII 0�.1 0�.1 0�.1
Enterobacter agglomerans Bo-1S 0�.4 0�.3 0�.3
Pseudomonas syringae pv. tabaci 0�.2 0�.3 0�.3
Pseudomonas aeruginosa ATCC15692 0�.7 1�.4 1�.5
Candida albicans (clinical isolate) 0�.5 0�.4 0�.8
Human red cells . 200 . 200 . 200
Fig. 5. Anti-bacterial activity of esculentin analogues. Panel A, Pseu-
domonas syringae pv. tabaci (peptide concentration, 0.6 mm); panel B,
Pseudomonas aeruginosa ATCC15692 (peptide concentration, 4 mm). X,
Esculentin-C; P, esculentin-L. Activity is expressed as colony-forming
units. �, control.
q FEBS 1999 Recombinant expression of esculentin-1 (Eur. J. Biochem. 263) 925
from a random coil to an a-helical conformation [18]. Theconformational transition is already observed at 40% trifluoro-ethanol for the linear peptide, whereas for the cyclic peptide acomparable modification occurs at higher trifluoroethanolconcentrations (Fig. 4).
Biological activity of recombinant esculentin analogues
The recombinant products were tested in vitro forinhibitory activity against a variety of microorganisms.The lethal concentration values range from 0.1 to 1.5 mm(0.5±7 mg´mL21), similar to those of the natural peptide(Table 1). Similarly, the recombinant peptides do not inducehemolysis of human red cells in agarose medium assay at theconcentration used for anti-bacterial activity, while in the liquid
medium assay, at a concentration of 15 mm, 24% and 6%hemolysis were detected for the cyclic and the linearesculentins, respectively (data not shown).
The bactericidal activity and the rate of bacterial killing ofthe two esculentin analogues was determined in NaCl/Pi againsttwo gram-negative bacterial strains, P. syringae pv tabaci(Fig. 5, panel A) and P. aeruginosa ATCC 15692 (Fig. 5, panelB) and the gram-positive S. aureus Cowan I at different NaClconcentrations (Fig. 6). The time-killing courses show thatgram-negative bacteria are more sensitive to both peptides thanthe gram-positive bacterium. In fact, against P. syringae at apeptide concentration of 1 mm (i.e, three times higher the LCvalue) no bacteria survive after the first minute (data notshown). At 0.6 mm, within 30 min, a 98% reduction in colony-forming units is observed, although the two peptides act withdifferent kinetics, the effect of the cyclic peptide being fasterthan that of the linear one. Against P. aeruginosa, at aconcentration three times higher than the LC value, the cyclicesculentin kills all bacteria in the first 15 min, while a longertime (up to 90 min) is necessary for the linear peptide.Moreover, the activity against the gram-negative bacteria wasnot affected by high salt concentrations (136 mm NaCl). Incontrast, S. aureus is killed significantly more slowly (within60 min in phosphate buffer containing 34 mm NaCl) at apeptide concentration 3±10 times higher than the correspond-ing LC values. As shown in Fig. 6, the activity of bothesculentin analogues is influenced by salt concentration, thedifference between them clearly evident at 51 mm NaCl.Similar results were also found in experiments performed in LBmedium (at 90 mm NaCl concentration), according to thedifferent LC values obtained by the inhibition zone assay(Table 1).
The effect of the two peptides on the permeabilization of thebacterial inner membrane was studied by the unmasking ofcytoplasmic b-galactosidase catalytic activity of E. coli D21for an extracellular chromogenic substrate. Both peptides, after90-min incubation with the bacterial suspension, induce therelease of the same amount of enzyme, as suggested by thesimilar optical density values of 2-nitrophenol, the product ofGal-ONp hydrolysis by b-galactosidase (Fig. 7).
D I S C U S S I O N
Protein expression
The need to express eukaryotic polypeptides as fusion proteinsresults from the discovery that low levels of protein expressionare obtained in E. coli, because a protein recognized as foreignis degraded [19,20]. In fact, the direct expression of the46-residue anti-microbial peptide esculentin-1b in E. coli wasunsuccessful: although Northern blot analyses revealed highlevels of the corresponding mRNA the peptide was never foundin the extract, presumably because it is rapidly degraded. Asecond approach was followed, consisting of the expression inE. coli of the maltose-binding receptor [21] with the esculentinfused at the C-terminus. Although a band of the expected sizewas evident in SDS/PAGE, and its N-terminal sequencecorresponded to that of the maltose-binding receptor, theprotein was very labile, being degraded at the C-terminus. Athird approach was finally attempted, in which the peptideof interest was fused at the extremity of human GABA-T[10], and the expressed protein was found to be sequesteredin the E. coli inclusion bodies. We used this expression strategyto protect the peptide against degradation by intracellularproteases, with the advantage of a high enrichment of the
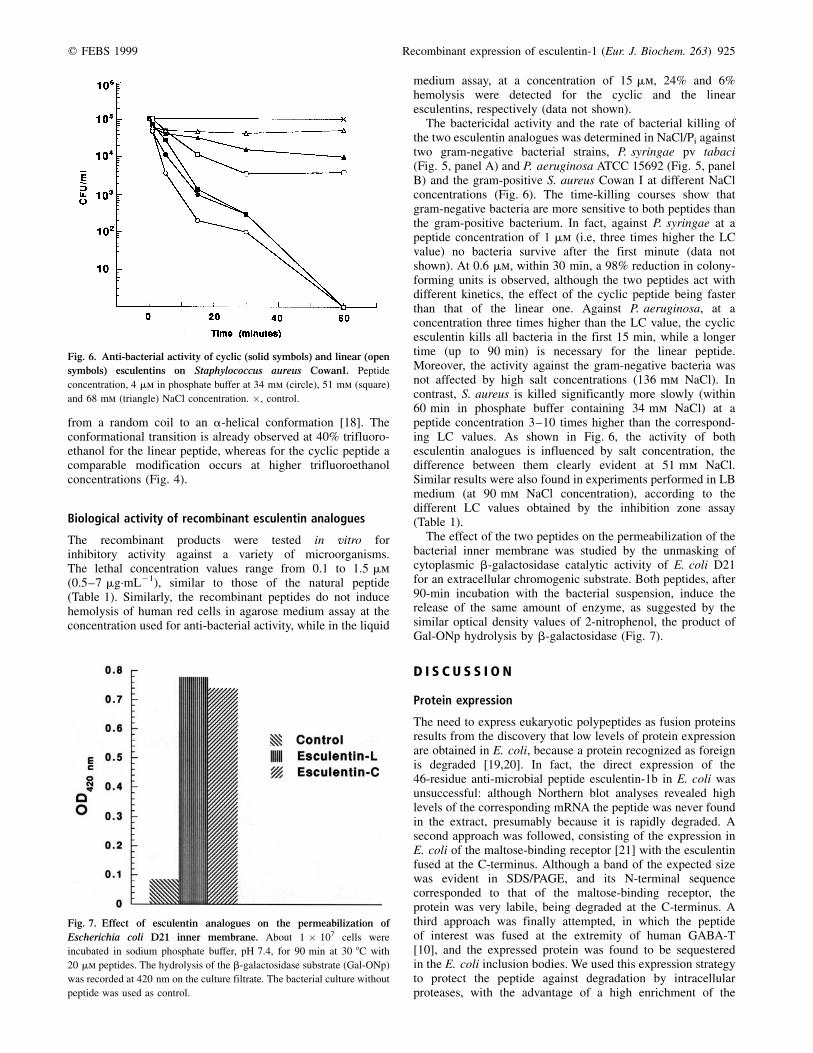
Fig. 6. Anti-bacterial activity of cyclic (solid symbols) and linear (open
symbols) esculentins on Staphylococcus aureus CowanI. Peptide
concentration, 4 mm in phosphate buffer at 34 mm (circle), 51 mm (square)
and 68 mm (triangle) NaCl concentration. �, control.
Fig. 7. Effect of esculentin analogues on the permeabilization of
Escherichia coli D21 inner membrane. About 1 � 107 cells were
incubated in sodium phosphate buffer, pH 7.4, for 90 min at 30 8C with
20 mm peptides. The hydrolysis of the b-galactosidase substrate (Gal-ONp)
was recorded at 420 nm on the culture filtrate. The bacterial culture without
peptide was used as control.

926 D. Ponti et al. (Eur. J. Biochem. 263) q FEBS 1999
desired protein. Also, the success of the expression procedurescould be evaluated in the early stages of purification, by directN-terminal sequencing of the appropriate molecular mass band.
The coexpression of the fusion protein and GABA-T wasfound to be essential for the formation of inclusion bodies. Asimilar behavior was observed in Bacillus thuringiensis, inwhich the simultaneous translation of two proteins from thesame transcript is required for the formation of inclusionbodies [22].
The formation of the esculentin intramolecular disulfidebond seems to occur spontaneously during the purificationprocedures. Moreover, it may function as scavenger, preventingmethionine oxidation. In initial experiments, the esculentinmutant lacking Cys residues could not be recovered aftercyanogen bromide cleavage. We suspected that the oxidation ofthe Met residue in position 47 has occurred, thus preventingcyanogen bromide action. When a preliminary incubation of theexpressed fusion protein with 2-mercaptoethanol was intro-duced in the purification protocol, the peptide was easilypurified using the same conditions as those followed forpurification of the Cys-containing analogue.
The pET-Esc-GABA-T vectors for the production of the tworecombinant peptides have been patented [23].
Biological activity
The availability of large amounts of esculentin analoguesallowed a more in-depth characterization of the biologicalactivities of this type of anti-microbial peptide. As shown inTable 1, the recombinant peptides behave in a manner similarto the natural esculentin-1a against most of the microorganismstested, without affecting mammalian cells. Moreover, bothrecombinant peptides exert the same permeabilization effect onthe bacterial inner membrane (Fig. 7). These results demon-strate that linearization of esculentin does not exhert a greateffect on conformation as well as on biological activity. This isin contrast with what is found in bactenecin, where lineariz-ation affects both conformation and anti-microbial activity[24]. However, these results are not surprising due to thedifferent length of the two peptides (12 versus 47 amino acidresidues).
A role for the disulfide bridge in esculentins can beenvisaged by analyzing the rate of bacteria killing. In fact,when the rate is measured against the gram-negative bacteriaP. syringae and P. aeruginosa, the cyclic and linear esculentinanalogues show a different behaviour: the cyclic peptide has amuch faster effect than the linear one (Fig. 5). Moreover, thisbehaviour is salt insensitive in the range 34±130 mm NaCl.
When the rate of killing is measured against the gram-positive S. aureus, the behavior is similar for both peptides.However, by increasing NaCl concentration (from 34 to51 mm), it is possible to discriminate the activities: the linearpeptide is inhibited to a greater extent when compared to thecyclic one (Fig. 6).
In peptides with more than one disulfide bridge, such asprotegrin-1, complete linearization causes a dramatic loss ofactivity at high salt concentrations against both gram-positiveand gram-negative bacteria [25]. However, this activity wasmeasured with an inhibition zone assay, condition in which theesculentin analogues are equally active against gram-negativebacteria, but display a different activity only against S. aureus(Table 1).
The results presented in this paper confirm the suitability ofexpressing esculentin-type peptides by recombinant DNAprocedures, allowing the design and production of several
different analogues, in order to exploit their potentialtherapeutic or biotechnological applications. Moreover, therecently described defensin subfamily from spinach, whichshares with esculentin a common N-terminal sequence [26],provides indication that esculentin can be used in plants toenhance the physiological response to infections.
A C K N O W L E D G E M E N T S
We thank Dr Maria Eugenia SchininaÁ for performing the mass spectrometry
analyses. This work was supported by grants from the Istituto Pasteur-
Fondazione Cenci Bolognetti, Consiglio Nazionale delle Ricerche (Target
project on Biotechnology) (D. B.), and Ministero dell'UniversitaÁ e della
Ricerca Scientifica e Tecnologica (D. B. and M. S.).
R E F E R E N C E S
1. Boman, H.G. (1995) Peptide antibiotics and their role in innate
immunity. Annu. Rev. Immun. 13, 61±92.
2. Lehrer, R.I. & Ganz, T. (1999) Antimicrobial peptides in mammalian
and insect host defence. Curr. Opin. Immunol. 11, 23±27.
3. Ojcius, D.M. & Young, J.D. (1991) Cytolytic pore-forming proteins
and peptides: is there a common structural motifs? Trends Biochem.
Sci. 16, 225±229.
4. Shai, Y. (1995) Molecular recognition between membrane-spanning
polypeptides. Trends Biochem. Sci. 20, 460±464.
5. Levy, S.B. (1998) The challenge of antibiotic resistance. Scientific
American, March issue, pp. 32±39.
6. Piers, K.L., Brown, M.H. & Hancock, R.E. (1993) Recombinant DNA
procedures for producing small antimicrobial cationic peptides in
bacteria. Gene 134, 7±13.
7. Lee, J.H., Minn, I., Park, C.B. & Kim, S.C. (1998) Acidic peptide-
mediated expression of the antimicrobial peptide buforin II as tandem
repeats in Escherichia coli. Protein Expr. Purif. 12, 53±60.
8. Zhang, L., Falla, T., Wu, M., Fidai, S., Burian, J., Kay, W. & Hancock,
R.E. (1998) Determinants of recombinant production of antimicrobial
cationic peptides and creation of peptide variants in bacteria.
Biochem. Biophys. Res. Commun. 247, 674±680.
9. Simmaco, M., Mignogna, G., Barra, D. & Bossa, F. (1994)
Antimicrobial peptides from skin secretion of Rana esculenta.
Molecular cloning of cDNAs encoding esculentin and brevinins
and isolation of new active peptides. J. Biol. Chem. 269,
11956±11961.
10. De Biase, D., Barra, D., Simmaco, M., John, R.A. & Bossa, F. (1995)
Primary structure and tissue distribution of human 4-aminobutyrate
aminotransferase. Eur. J. Biochem. 227, 476±480.
11. Marston, F.A. (1986) The purification of eukaryotic polypeptides
synthesized in Escherichia coli. Biochem. J. 240, 1±12.
12. Barra, D., Simmaco, M. & Boman, H.G. (1998) Gene-encoded peptide
antibiotics and innate immunity. Do `animalcules' have defence
budgets? FEBS Lett. 430, 130±134.
13. Hultmark, D., EngstroÈm, A., Andersson, K., Steiner, H., Bennich, H. &
Boman, H.G. (1983) Insect immunity. Attacins, a family of
antibacterial proteins from Hyalophora cecropia. EMBO J. 2,
571±576.
14. Mignogna, G., Simmaco, M., Kreil, G. & Barra, D. (1993)
Antibacterial and haemolytic peptides containing d-alloisoleucine
from the skin of Bombina variegata. EMBO J. 12, 4829±4832.
15. Lehrer, R.I., Barton, A. & Ganz, T. (1988) Concurrent assessment of
inner and outer membrane permeabilization and bacteriolysis in E.
coli by multiple-wavelength spectrophotometry. J. Immunol. Methods
108, 153±158.
16. Shine, J. & Dalgarno, L. (1974) The 3 0-terminal sequence of
Escherichia coli 16S ribosomal RNA: complementarity to nonsense
triplets and ribosome binding sites. Proc. Natl Acad. Sci. USA 71,
1342±1346.
17. Stormo, G.D., Schneider, T.D. & Gold, L.M. (1982) Characterization of

q FEBS 1999 Recombinant expression of esculentin-1 (Eur. J. Biochem. 263) 927
translational initiation sites in E. coli. Nucleic Acids Res. 10,
2971±2996.
18. Luo, P. & Baldwin, R.L. (1997) Mechanism of helix induction by
trifluoroethanol: a framework for extrapolating the helix-forming
properties of peptides from trifluoroethanol/water mixtures back to
water. Biochemistry 36, 8413±8421.
19. Goldschmidt, R. (1970) In vivo degradation of nonsense fragments in
E. coli. Nature 228, 1151±1154.
20. Lin, S. & Zabin, I. (1972) b-Galactosidase. Rates of synthesis and
degradation of incomplete chains. J. Biol. Chem. 247, 2206±2211.
21. Guan, C., Riggs, P.D. & Inouye, H. (1987) Vectors that facilitate the
expression and purification of foreign peptides in Escherichia coli by
fusion to maltose-binding protein. Gene 67, 21±30.
22. Rang, C., Bes, M., Moar, W.J. & Frutos, R. (1997) Simultaneous
production of the 34-kDa and 40-kDa proteins from Bacillus
thuringiensis subsp. thompsoni is required for the formation of
inclusion bodies. FEBS Lett. 412, 587±591.
23. Barra, D., Bossa, F., De Biase, D., Simmaco, M. & Ponti, D. (1998)
Plasmid for the production of anti-microbial peptides in recombinant
bacteria. Italian patent no. RM98A000393.
24. Wu, M. & Hancock, R.E.W. (1999) Interaction of the cyclic
antimicrobial cationic peptide bactenecin with the outer and
cytoplasmic membrane. J. Biol. Chem. 274, 29±35.
25. Harwig, S.L., Waring, A., Yang, H.J., Cho, Y., Tan, L. & Lehrer, R.
(1996) Intramolecular disulfide bonds enhance the antimicrobial and
lytic activities of protegrins at physiological sodium chloride
concentrations. Eur. J. Biochem. 240, 352±357.
26. Segura, A., Moreno, M., Molina, A. & Garcia-Olmedo, F. (1998) Novel
defensin subfamily from spinach (Spinacia oleracea). FEBS Lett.
435, 159±162.




















