
Evapotranspiration (ET) and regulating mechanisms in two semiarid Artemisia-dominated shrub steppes...
10
Evapotranspiration (ET) and regulating mechanisms in two semiarid Artemisia-dominated shrub steppes at opposite sides of the globe Burkhard Wilske a, * , Hyojung Kwon b, 1 , Long Wei c , Shiping Chen c , Nan Lu a , Guanghui Lin c , Jing Xie a, d , Wenbin Guan d , Elise Pendall b , Brent E. Ewers b , Jiquan Chen a, c a Department of Environmental Sciences, University of Toledo, Toledo, OH 43606, USA b Department of Botany and Program in Ecology, University of Wyoming, Laramie, WY 82071, USA c State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, The Chinese Academy of Sciences, Xiangshan, Beijing 100093, PR China d College of Nature Conservation, Beijing Forestry University, Beijing 100083, PR China article info Article history: Received 31 July 2009 Received in revised form 3 March 2010 Accepted 18 May 2010 Keywords: Dryland ET PenmaneMonteith model Soil water potential Vapor pressure deficit Water vapor fluxes abstract Understanding the regulation of ecosystem ET is crucial to comprehend present and potential future vegetation and water availability in arid environments. We compared ET data from eddy-covariance flux measurements in two semiarid shrub-dominated ecosystems. Located in the Kubuqi Desert (KD, Inner Mongolia, China) and the Sierra Madre (SM, Wyoming, USA), the ecosystems shared similarities in latitude, climate and vegetation. We analyzed ET at KD and SM for the years 2006e2007 and 2004e2005, respectively, and tested regulatory functions of climate variables through modifications to the widely used FAO-PenmaneMonteith model. Important modifications included the insertion of soil water potential (Js) and the removal of vapor pressure deficit (VPD) and wind speed (u). The modifications improved daily estimates of growing season ET as they reduced deviations between model and measurement to 1e33%. According to model results, available energy (R n G) and Js were the main drivers of ET during growing seasons. VPD and u influenced ET mainly after precipitation events and at low Js (i.e., Js close to zero). Although our empirical model still requires validation through data from other sites, the results suggest that the regulatory power of Js and the conditional sensitivity to VPD are characteristic of ET in shrub steppes. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Evapotranspiration (ET) is one of the main components in the water cycle affecting the local-to-regional water balance and netting long-term feedbacks between vegetation and climate. A recent IPCC assessment projects the evaporative demand to increase almost everywhere because the water-holding capacity of the atmosphere increases with higher temperatures, but relative humidity is not projected to change markedly (Bates et al., 2008). Semiarid shrub-dominated ecosystems have so far drawn little attention regarding their ET, although open and closed shrublands cover 14% of the terrestrial surface (Loveland et al., 2000). From a climatic perspective, semiarid shrublands stand away from forests at the drier site of grasslands (Stephenson, 1990). Shifts between shrub and grass vegetation can have great impact on the hydrology, carbon and nutrient cycles in dryland areas (Frank and Karn, 2005; Scott et al., 2006). Measurements of ET in shrublands are rare because of their remoteness, but accurate ET predictions are required for assessment of runoff and groundwater recharge, primary productivity and rangeland management (Cleary et al., 2008). Recent installations of eddy-covariance flux measurements (EC) allowed us to investigate two shrub-dominated ecosystems at KD (Inner Mongolia) and SM (Wyoming). ET-relevant dissimilarity between the sites existed regarding a summer rain climate and deciduous shrubs at KD in contrast to winter precipitation and evergreen shrubs at SM. However, both sites shared also important similarities such as in the annual amount of precipitation, the mean air temperature, and the potential radiation because both sites are located at uplands of similar latitude. On top of these abiotic features, the dominant shrub vegetation consisted of congenous species (Artemisia sp.) of relatively similar height and cover. Hence, we considered a relaxed concept of vicariant species, which may imply further that similar parameters dominate the regulation of * Corresponding author. Tel.: þ1 419 530 4278. E-mail address: [email protected] (B. Wilske). 1 Present address: Department of Atmospheric Sciences, Yonsei University, Seoul, South Korea. Contents lists available at ScienceDirect Journal of Arid Environments journal homepage: www.elsevier.com/locate/jaridenv 0140-1963/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.jaridenv.2010.05.013 Journal of Arid Environments 74 (2010) 1461e1470
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Evapotranspiration (ET) and regulating mechanisms in two semiarid Artemisia-dominated shrub steppes...

lable at ScienceDirect
Journal of Arid Environments 74 (2010) 1461e1470
Contents lists avai
Journal of Arid Environments
journal homepage: www.elsevier .com/locate/ jar idenv
Evapotranspiration (ET) and regulating mechanisms in two semiaridArtemisia-dominated shrub steppes at opposite sides of the globe
Burkhard Wilske a,*, Hyojung Kwon b,1, Long Wei c, Shiping Chen c, Nan Lu a, Guanghui Lin c, Jing Xie a,d,Wenbin Guan d, Elise Pendall b, Brent E. Ewers b, Jiquan Chen a,c
aDepartment of Environmental Sciences, University of Toledo, Toledo, OH 43606, USAbDepartment of Botany and Program in Ecology, University of Wyoming, Laramie, WY 82071, USAc State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, The Chinese Academy of Sciences, Xiangshan, Beijing 100093, PR ChinadCollege of Nature Conservation, Beijing Forestry University, Beijing 100083, PR China
a r t i c l e i n f o
Article history:Received 31 July 2009Received in revised form3 March 2010Accepted 18 May 2010
Keywords:DrylandETPenmaneMonteith modelSoil water potentialVapor pressure deficitWater vapor fluxes
* Corresponding author. Tel.: þ1 419 530 4278.E-mail address: [email protected] (B.
1 Present address: Department of Atmospheric ScienSouth Korea.
0140-1963/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.jaridenv.2010.05.013
a b s t r a c t
Understanding the regulation of ecosystem ET is crucial to comprehend present and potential futurevegetation and water availability in arid environments. We compared ET data from eddy-covariance fluxmeasurements in two semiarid shrub-dominated ecosystems. Located in the Kubuqi Desert (KD, InnerMongolia, China) and the Sierra Madre (SM, Wyoming, USA), the ecosystems shared similarities inlatitude, climate and vegetation. We analyzed ET at KD and SM for the years 2006e2007 and 2004e2005,respectively, and tested regulatory functions of climate variables through modifications to the widelyused FAO-PenmaneMonteith model. Important modifications included the insertion of soil waterpotential (Js) and the removal of vapor pressure deficit (VPD) and wind speed (u). The modificationsimproved daily estimates of growing season ET as they reduced deviations between model andmeasurement to 1e33%. According to model results, available energy (Rn�G) and Js were the maindrivers of ET during growing seasons. VPD and u influenced ET mainly after precipitation events and atlow Js (i.e., Js close to zero). Although our empirical model still requires validation through data fromother sites, the results suggest that the regulatory power of Js and the conditional sensitivity to VPD arecharacteristic of ET in shrub steppes.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Evapotranspiration (ET) is one of the main components in thewater cycle affecting the local-to-regional water balance andnetting long-term feedbacks between vegetation and climate. Arecent IPCC assessment projects the evaporative demand toincrease almost everywhere because the water-holding capacity ofthe atmosphere increases with higher temperatures, but relativehumidity is not projected to change markedly (Bates et al., 2008).Semiarid shrub-dominated ecosystems have so far drawn littleattention regarding their ET, although open and closed shrublandscover 14% of the terrestrial surface (Loveland et al., 2000). Froma climatic perspective, semiarid shrublands stand away from forestsat the drier site of grasslands (Stephenson, 1990). Shifts between
Wilske).ces, Yonsei University, Seoul,
All rights reserved.
shrub and grass vegetation can have great impact on the hydrology,carbon and nutrient cycles in dryland areas (Frank and Karn, 2005;Scott et al., 2006). Measurements of ET in shrublands are rarebecause of their remoteness, but accurate ET predictions arerequired for assessment of runoff and groundwater recharge,primary productivity and rangeland management (Cleary et al.,2008).
Recent installations of eddy-covariance flux measurements (EC)allowed us to investigate two shrub-dominated ecosystems at KD(Inner Mongolia) and SM (Wyoming). ET-relevant dissimilaritybetween the sites existed regarding a summer rain climate anddeciduous shrubs at KD in contrast to winter precipitation andevergreen shrubs at SM. However, both sites shared also importantsimilarities such as in the annual amount of precipitation, the meanair temperature, and the potential radiation because both sites arelocated at uplands of similar latitude. On top of these abioticfeatures, the dominant shrub vegetation consisted of congenousspecies (Artemisia sp.) of relatively similar height and cover. Hence,we considered a relaxed concept of vicariant species, which mayimply further that similar parameters dominate the regulation of

Table 1Site characteristics of the shrub-dominated ecosystems in the Kubuqi Desert (KD,Inner Mongolia, China) and the Sierra Madre (SM, Wyoming, USA).
KD SMa
Latitude N 40�220510 0 N 41�19051.90 0 N 41Longitude E 108�320550 0 W 107�24002.40 0
Elevation (m) 1160 2260Annual mean T (�C) 8.2 [2006] 6.2e7.2Mean PPT (mmyr�1) 230e300 259e341b
Months incl.> 50% PPT yr�1 JulyeSeptember AprileJuneSoil type Sandy soil Sandy loamBulk soil density (g cm�3) 1.62� 0.08 (0e30 cm) 1.13 (0e40 cm)Soil water retentionc a¼ 377.53 a¼ 247.1926Js¼ a� exp(�b�VWC) b¼ 46.7055 b¼ 43.1260Dominant Artemisia spp. A. ordosica A. tridentata var. vaseyanaVegetation height (m) 0.55� 0.11 1.0Total vegetation cover (%) 17e23 27e39LAI 0.30� 0.13 1.2Stomatal resistance (sm�1) 45d 149e
Growing season dataET (days/season s/w)f 2006: 166/108 2004: 26/NA
2007: 120/77 2005: 43/NA
a From Ewers and Pendall (2007) and Kwon et al. (2008).b Measured at a site in 100 km distance (ibid.).c J(�MPa).d rs¼ 1/g, max. stomatal conductance from Xiao et al. (2005)z 1.0.e rs¼ 1/g, max. stomatal conductance from Loik (2007)¼ 0.3.f Days past quality control per growing (s) and non-growing season (w).
Nomenclature
ET evapotranspiration (measured) (mmd�1)ET0 reference ET of the FAO-PM for well-watered reference
crop (mmd�1)D slope of the saturation vapor pressure temperature
relationship (kPa �C�1)g psychrometric constant (kPa �C�1)G soil heat flux (Wm�2)Js soil water potential (MPa), note that positive values
were usedpa air pressure (kPa)ph standard pressure at elevation of sitePM PenmaneMonteith model, FAO-PM as adopted
by FAOFAO-PMs including shrub-adapted coefficients
FAO-PM_a and _b calculated using the radiation andtemperature term only
pm(i) modified FAO-PM, number (i¼ 1e4) indicating thestep of modification
pm(i)c and s denote use of crop and shrub-adapted coefficients,respectively
pm(i)s_a and _b if based on the radiation and temperature termonly
Rn net radiation (Wm�2)Rn�G available energy (Wm�2)Ta air temperature (�C)Ts soil temperature (�C)u wind speed (m s�1)u dir wind direction (degree)VPD vapor pressure deficit (kPa)VWC soil water content (cm3 cm�3)
B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e14701462
ecosystem ET. Vegetation can increase ET through transpiration andby using roots to exploit water from soil and groundwater resources(Scott et al., 2006; Wilske et al., 2009). Here, regulation of stomatalconductance becomes crucial as it adjusts the water use of plants tothe water availability in the ecosystem (Jarvis and McNaughton,1986; Fischer et al., 2008). Vapor pressure deficit (VPD) iscommonly accepted as being a key driver of ET in vegetatedecosystems (e.g., Mackay et al., 2007), because it integrates thecontrols on both transpiration and evaporation. In drylands inparticular, soil water availability has been identified as a key factorfor ruling the architecture of vegetation (Vezzoli et al., 2008). In thesame context, studies have shown that the drought resistance ofdesert shrubs (e.g., Artemisia tridentata) is controlled by the cavi-tation resistance of the root xylem rather than by the above-surfacepart of the plants (Kolb and Sperry, 1999; Sperry and Hacke, 2002).Soil water potential (Js, MPa) is commonly used in ecosystemscience to indicate soil water availability and to reflect simulta-neously soil properties that tend to retain rather than release water(Meidner, 1979; Campbell, 1988; Williams et al., 1993). In accor-dance with the considerations above, we set out from twohypotheses: (1) VPD is a major driver of ET in the shrub-dominatedecosystems and has a stronger control at the site with summer rainthan at the winter precipitation site; (2) Js is the second-mostimportant regulator of ET at both sites.
To test our hypotheses, we sought an available model to serve asa framework for ET simulations. ETmodels represent powerful toolsto examine land-to-atmosphere water vapor fluxes in correspon-dence to climate change, hydrology, land use, vegetation dynamics,crop production and other ecosystem functions (Rana and Katerji,1998; Rey, 1999; Xu et al., 2006; Fisher et al., 2008; Zhang et al.,2008). The suitability of ET models depends on the temporal andspatial scale of the desired simulations (Walker and Langridge,1996;Pereira and Pruitt, 2004; Fisher et al., 2005) and the available inputparameters, such as temperature, radiation and else (e.g.,Thornthwaite, 1948; Makkink, 1957; Priestley and Taylor, 1972;Hargreaves et al., 1985). The PenmaneMonteith model (Monteith,1965) has been employed to interpret ET in many terrestrialecosystems (e.g., forests, prairies, croplands and wetlands) due tothe consensus betweenmodel andmeasured parameters becomingavailablewithnetworks usingeddycovariance andother techniques(Rana et al., 1997; Fisher et al., 2005; Aires et al., 2008; Chen et al.,2008). The FAO adopted the PenmaneMonteith model to providereference ET (ET0) for crop vegetation (FAO-PM, Allen et al., 1998).The FAO-PM is probably the most widely used model regarding itsaccessibility and documentation (http://www.fao.org/docrep/X0490E/x0490e06.htm). Although like many other models it
performs less well under dry than moist conditions (Fisher et al.,2005), we found the FAO-PM most suitable for our purpose,because it contains separate terms for the radiation and thetemperature component and incorporates VPD and vegetationcharacteristics (e.g., height, leaf area index and stomatal resistance).
The objectives of our study were to: (1) examine the biophysicalregulation of ET in semiarid shrub-dominated ecosystems per-taining to parameters of major andminor influence, and (2) test ourtwo hypotheses regarding the influences of VPD and Js. Theseobjectives were achieved through analysis of measured ET andmodeling based on the framework of the FAO-PM.
2. Materials and methods
2.1. Study sites
ET and climate data were analyzed from two shrub-dominatedecosystems in uplands at similar latitude but at opposite sides ofthe globe (Table 1). One site was located in the Kubuqi Desert

B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e1470 1463
(KD), Inner Mongolia, China, and the other in the northwesternSierra Madre (SM), Wyoming, USA. The climate at both locationsis semiarid with long cold winters and dry warm summers. Theecosystems at KD and SM were dominated by Artemisia ordosicaKrasch. and A. tridentata Nutt. ssp. vaseyana (Rydb.) Beetle,respectively. A. ordosica is a minor deciduous shrub of 0.6e1 mheight (Xiao et al., 2003). A. tridentata Nutt. ssp. vaseyana(mountain big sagebrush) is evergreen and averages 1 m inheight (Cleary et al., 2008). Both species are moderately droughtresistant (Kolb and Sperry, 1999; Zheng et al., 2006). Co-domi-nant species included the shrub Hedysarum mongolicum Turcz. atKD, and the grasses Festuca idahoensis Elmer and Poa secunda J.Presl. at SM. The SM site was moderately grazed and containedmature sagebrush (z 40 years since fire) and burned sagebrush(z 5 years since fire) in equal proportions (Kwon et al., 2008).Vegetation height, cover and leaf area were higher in SM than KD(Table 1). The soils were characterized as sandy soil at KD (Zhang,1994) and as sandy loam (Calcic Pachic Argicryoll) with a 40-cmdeep, C-rich A-horizon at SM (Kwon et al., 2008).
2.2. Eddy-covariance (EC) instrumentation and micrometeorology
Water vapor fluxes and micrometeorological data at both siteswere obtained for parts of the periods 2006e2007 and 2004e2005,respectively. Net exchange of water vapor (i.e., latent heat, LE) wasmeasured by means of the EC technique using LI-7500 open-pathinfrared gas analyzers (IRGA; Li-Cor, Lincoln, NE, USA) for watervapor, CSAT3 3-dimensional sonic anemometers (Campbell Scien-tific Inc. (CSI) Logan, USA) for wind and temperature fluctuation,and CR5000 data loggers (CSI).
Net radiation (Rn, Wm�2) was measured with net radiometersmodel Q-7.1 (Radiation and Energy Balance Systems, REBS, Seattle,USA) and CNR 1 (Kipp & Zonen, Delft, the Netherlands), respec-tively. Air temperature (Ta, �C) and relative air humidity (RH, %)were recorded with HMP45AC probes (Vaisala, Helsinki, Finland).Soil temperatures (Ts, �C) were measured using T-type thermo-couples at four different levels (2, 5, 6, and 10 cm below thesurface). Soil heat flux (G, Wm�2) was measured with heat fluxplates HFT-3 (REBS) and HFP01SC (Hukseflux, Delft, theNetherlands) in KD and SM, respectively. Soil heat flux was calcu-lated as the overall average of three soil depths (max. depths 15 cm)and from close to and between shrubs. Precipitation (PPT) wasrecorded with tipping bucket rain gauges TE525 (CSI). Equipmentmalfunction limited availability of PPT data at SM (see Kwon et al.,2008).
Soil water content (VWC, cm3 cm�3) was measured at soildepths of 10, 20, 30, and 50 cm at KD, and 4, 15, 15e45, and45e75 cm at SM using horizontally and vertically installed soilmoisture probes (model CS616, Campbell Scientific). The relation-ship between VWC and soil water potential (Js) in SM wasexamined using a dewpoint potentiometer (modelWP4-T, DecagonDevices, Pullman, WA, USA). Data for the soil water retention curveof the KD site was obtained with a Pressure Membrane Extractor(Soilmoisture Equipment Corp., Santa Barbara, CA, USA) andgravimetrical measurements of VWC using the oven-dry method.To provide a comparable level for VWC and Js at both sites, weused the average of the 10- and 20-cm soil water data at KD to alignwith the 15-cm VWC at SM. We assumed that the differencebetween the 15-cm measurements and the average of the 10- and20-cm measurements will be absorbed, in reality, by the spatialvariability of soil moisture within the EC footprint area (e.g., Vivoniet al., 2010), and for the model application by the transformationfrom VWC to Js.
2.3. Data processing
Water vapor fluxes from both sites were processed to axis-rotated, WBL-corrected (Webb et al., 1980) 30-min ET data. Half-hourly data were removed, if: (1) the values for the Automatic GainControl (AGC) of the LI-7500 were >75, (2) the mean wind speedwas <1 m s�1, or (3) friction velocity (u*) was <0.1 m s�1 and<0.17 m s�1 at KD and SM, respectively. Daily ET was calculatedfrom 30-min ET data without gap-filling, if: (1) �45 30-min ETrecords were available, (2) PPT data indicated no precipitation forthe day, and (3) the auxiliary meteorological sensors deliveredreliable data for the same period. Past the quality measures, dailyET and climate data were available for 26, 43, 274, and 197 days forthe years 2004 (SM), 2005 (SM), 2006 (KD), and 2007 (KD),respectively. SM data comprised only of days within the growingseasons (i.e., data subsets SMs04 and SMs05). Therefore, KD datawere divided into four subsets including the growing season (s) ornon-growing season (w) based on a growing season threshold ofdaily mean Ta� 5 �C (Table 1).
2.4. Model tests
The FAO-PM refers to ET0 of a virtual homogenous grass cover of0.12 m height at any place and climate (http://www.fao.org/docrep/X0490E/x0490e06.htm). To assess the influence of individualparameters (e.g., VPD) on ET, we compared the correlation betweenmeasured and model ET with the model including either (1) thevariable, (2) the variable substituted by its seasonal mean, or (3) thevariable value set to 1 (i.e., exclusion of the parameter). Eq. (1)shows the FAO-PMwith the driving climate parameters substitutedby small letters (aee).
½FAO� PM� ET0 ¼ ððc1 � D� ðaÞÞ þ ðg� ðc2=bþ 273Þ � d
� eÞ=ðDþ g� ð1þ c3 � eÞÞ(1)
where D is the slope of the vapor pressure curve (kPa �C�1), (a) isthe available energy (Rn�G) in (Wm�2), g is the psychrometricconstant (kPa �C�1), b is daily mean Ta at 2 m height (�C), d is dailymean VPD at 2 m height (kPa), e is daily mean u at 2 m height(m s�1), c1 is the coefficient 0.035 converting Wm�2 into mmd�1,c2¼ 900, and c3¼ 0.34.
The first and second bracket in the numerator is called here-inafter the radiation term ((c1�D� (a))) and the temperatureterm (g� (c2/bþ 273)� d� e), respectively. The coefficients c2and c3 involve aerodynamic and surface resistances, which inturn reflect vegetation height and leaf area index (LAI), andthereby relate the driving variables to the vegetation. We testedthe FAO-PM and subsequent modifications with both (1) c2 and c3for the standard reference crop and (2) c2 and c3 to reflect theshrub vegetation (i.e., SM c2S¼ 2200, c3S¼ 2.94; KD c2S¼ 1480,c3S¼ 4.34; calculations according to http://www.fao.org/docrep/X0490E/x0490e06.htm#aerodynamicresistance(ra) and htm#(bulk) surface resistance (rs)). Shrub-adapted coefficients reflec-ted one value for vegetation height, LAI and stomatal resistanceper site for both years (Table 1).
The tests with the FAO-PM model at the two sites resulted inmodifications including exchange and elimination of variables. Themodifications turned the FAO-PM into an empirical model denotedas “pm”. The denotation is followed by a consecutive number,subscripts that mark the use of crop (c) or shrub-adapted coeffi-cients (s), and a final underscore _a or _b where only the radiation(a) or the temperature (b) term were used (see Nomenclature).

Table 2Precipitation (PPT, mm) relative to growing season data subset (s) at the SierraMadre (SM_yr) and the Kubuqi Desert (KD_yr).
PPT SM_04 SM_05 KD_06 KD_07
Before s N/A N/A 1.6 4.0Within s 5.8 18.5 202.3 151.2After s N/A N/A 25.5 47.1Annual 259e341a 229.4 202.3
N/A not applicable.a From Ewers and Pendall (2007).
B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e14701464
The first modification pm1c excluded u and substituted VPD (Eq.(2)). VPD was at initially substituted by VWC. VWC was inserted indenominator position, i.e., virtually replacing u, because VWCindicates water availability instead of water deprivation. VWC wasthen replaced by soil water potential Js. Along with the exclusionof u, the coefficient c3 became obsolete and was in the followingmodifications replaced by a simple multiplier to fit model tomeasured ET.
½pm1c� ET ¼ ððc1 � D� ðaÞÞ þ ðg� ðc2=bþ 273ÞÞ=ðDþ g
� ð4� gÞÞ (2)
where g represents (OJs) with Js in e(MPa); note that J (both insoil and leaf water potential) is usually a dimensionwith a negativesign, whereas we used positive values for the variable (i.e., (�1)�J).
We ran the model with both crop and shrub-adapted coeffi-cients. The radiation term essentially contributed linearity to themodel. Ta was linearly correlated with the quotient of standardpressure at altitude and actual pressure (R2: SMs04 0.89, SMs050.92, KDs06 0.57, KDs07 0.34). The inverse relationship was appliedto the radiation term in order to avoid overestimation and achievemaximum yield when the temperature influence is high and low,respectively (Eq. (3)).
hpm2s a
iET ¼ ðc4Þ2�ððc1 � D� ðaÞÞ=ðDþ g� ð4� gÞÞÞ (3)
where c4 is the pressure quotient of actual pressure to normalpressure at altitude (pa/ph) (kPa� kPa�1).
The same relationship was linked with a stark attenuatedtemperature term (Eq. (4)).
hpm3s
iET ¼ ðc4Þ2�
��ðc1 � D� ðaÞÞ þ 0:1� ðg� ðc2S=bþ 273ÞÞ=ðDþ g� ð4� gÞÞ� (4)
In accordance with the understanding that VPD has maximuminfluence on ET at high VWC (e.g., Mackay et al., 2007), the atten-uated temperature term alternatively was decoupled from thepressure regulation and instead subjected to a regulation by theproduct of VPD and VWC (Eq. (5)).
hpm4s
iET ¼
�ðc4Þ2�ððc1 � D� ðaÞÞ þ ð0:2� d� f Þ � ðg
� ðc2S=bþ 273ÞÞÞ=�Dþ g�
�4�g
��(5)
where d is VPD and f is VWC.
Table 3Pearson correlation coefficient (r) between daily ET and daily means of climateparameters at flux tower sites in the Sierra Madre (SM, yr 2004e2005) and theKubuqi Desert (KD, yr 2006e2007) for non-growing (w: Ta< 5 �C) and growingseasons (s: Ta> 5 �C).
ET SMs04 SMs05 KDw06 KDs06 KDw07 KDs07
Rn�G 0.7 0.7 0.5 0.6 0.5 0.5Rn 0.5 0.5 0.5 0.6 0.6 0.5VWC 0.8 0.6 �0.1 0.8 0.5 0.8u �0.1 0.5 0.0 �0.3 0.3 �0.4u dir 0.3 0.6 0.1 0.4 0.1 0.1G 0.4 0.4 0.2 0.0 0.3 0.3Ta �0.4 �0.3 0.1 0.6 0.5 0.6pa 0.3 0.3 0.0 �0.5 �0.4 �0.6VPD �0.4 �0.4 0.1 0.1 0.2 0.2Js 0.8 0.5 �0.1 0.8 0.4 0.8
2.5. Tests and statistics
The subsets of ET data available from SM and KD differed in sizeand year and encompassed periods with different amounts of PPT.Annual PPT was 229.4 mm at KD_06 and 202.3 mm at KD_07.However, PPT was 50 mm higher within the period encompassingthe growing season data subset KDs06 as compared with KDs07(Table 2). PPT was higher in SMs05 than in SMs04 (Table 2),although growing season data were restricted to a short period(Table 1).
We used standard T-test in S-Plus (S-Plus 6.1, Insightful Corp.,Seattle, WA, USA) to check site and seasonal differences in means ofET and climate parameters and the slope and intercept betweenmodel and measured ET. The differences not explicitly stated weresignificant (P� 0.05). We checked the Pearson correlation (r)between daily ET and individual climate parameters per site andseason (Table 3).
The plus operator in the FAO-PM numerator enables examina-tion of the individual contributions of the radiation and thetemperature terms. The difference between ET from the overallmodel and results from either the radiation or the temperatureterm indicated the contribution of either term. The performance ofthe FAO-PM and subsequent modifications was evaluated sepa-rately for each site and season by means of three measures: (1) thedeviation of modeled from measured seasonal total ET (mm), (2)the root mean square error (RMSE) between expected and esti-mated values, and (3) the slope and correlation (R2) of the linearregression between model and ET (Table 4).
3. Results
3.1. ET regulating parameters
Mean daily ET was significantly different between sites andseasons except for the two growing seasons at KD (P¼ 0.101,Fig. 1a). However, climate parameters showed many similaritiesbetween sites despite the difference in data size (Table 1, Fig.1beh).Although mean Rn was significantly different between all sites andseasons and themain range (i.e., mean� 2 SD) of growing season Rnwas generally higher at the more elevated site (SM) than the lowerelevated site (KD). The peak values of Rn were about similar atSMs04 (8699 Wm�2) and KDs06 (8517Wm�2, Fig. 1b). The mainrange of the climate parameters VWC, G, Ta, Ts, VPD and u wassimilar or not significantly shifted during growing seasons at thetwo sites (Fig. 1ceh). Furthermore, the means of data subsets weresimilar between seasons of different years and across sites. Mostimportantly, mean VWC was not significantly different betweenSMs04, KDs06 and KDs07 at similar soil depths (e.g., 15 cm,0.104e0.110 cm3 cm�3, P¼ 0.414e0.521, Fig. 1c) and Ta was notsignificantly different between the growing seasons at SM and KD(16.5e18.8 �C, P¼ 0.138e0.801, Fig. 1e). Mean Ts was not signifi-cantly different between SMs05 and KDs06 (P¼ 0.053) and KDs07(P¼ 0.165). VPD was significantly different between seasons andsites except for the two winter seasons at KD (P¼ 0.592) and

Table 4Performance of FAO-PM and pm modifications for the Sierra Madre and KubuqiDesert site per yr and season: (a) mm-deviation between total predicted andmeasured ET per period; slope (x� ET) and correlation (R2) based on the wholedataset; Eqs. (1)e(4) refer to the underlying equation, brackets indicate the use ofeither the shrub (s) or crop (c) coefficient, Eq. (3) (_a) is based on the radiation termonly; (b) the mean deviations from the predicted value (RMSE) within subsets.
(a) SMs04 SMs05 KDw06 KDs06 KDw07 KDs07 x� ET R2P
ET measured 41 118 9 148 14 90P(modeled ET�measured ET)
FAO-PM Eq. (1) 130 120 103 600 77 517 1.34 0.29FAO-PMs (Eq. (1)) 79 32 25 144 13 260 1.43 0.21pm1c Eq. (2) 1 �15 11 57 4 79 1.08 0.72pm1s (Eq. (2)) 16 20 22 94 13 108 1.34 0.75pm2s_a Eq. (3) 1 �2 �6 �8 �9 28 0.98 0.88pm3s Eq. (4) 5 7 �3 1 �7 35 1.04 0.88pm4s Eq. (5) 3 1 �6 �5 �9 30 1.00 0.88
(b) RMSE SMs04 SMs 05 KDw 06 KDs06 KDw07 KDs 07P
FAO-PM 1.77 1.58 1.77 1.90 1.75 2.14 1.82FAO-PMs 1.45 1.10 0.46 0.93 0.43 0.94 0.81pm1c 0.36 0.74 0.34 0.57 0.38 0.75 0.54pm1s 0.41 0.78 0.43 0.62 0.45 0.78 0.60pm2s_a 0.39 0.54 0.22 0.47 0.32 0.51 0.41pm3s 0.36 0.69 0.33 0.47 0.37 0.56 0.46pm4s 0.40 0.54 0.23 0.47 0.32 0.52 0.41
Fig. 1. Mean, main range (�2 SD, bar) and variance (line) of daily ET and climateparameters of the six data subset for the growing (s)/non-growing season (w) in theSierra Madre (2004, 2005) and the Kubuqi Desert (2006, 2007). Differences in meanswere not significant (P� 0.05) where indicated. *¼ between seasons at the same siteand **¼ between seasons across sites.
B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e1470 1465
between SMs05 (1.40 kPa) and KDs06 (1.41 kPa, P¼ 0.894, Fig. 1g).The variation in wind speed was larger on the high plateau (KD)than at themountainous site (SM). However, uwas not significantlydifferent between SMs04, SMs05 and KDw06 (P¼ 0.212e0.540)and not between the two winter (P¼ 0.693) and two summerseasons (P¼ 0.523) at KD. The overall changes in Rn and VWC dis-played pattern similar to daily ET (Fig. 1aec), whereas patterns inother climate parameters were less congruent. In particular, VPDwas higher at SMs04 than SMs05 whereas ET showed the oppositepattern (Fig. 1 a and g). VPD correlated inversely with VWC atSMs04 and SMs05 based on bins differing by 0.01 cm3 cm�3,however, statistics supported only two and three bins, respectively(data not shown). A similar correlation was not found for KD.
Climate parameters differed considerably regarding their indi-vidual correlation with daily ET (Table 3). Daily ET showed mostlypositive correlations (r� 0.6) with the parameters (Rn�G), Rn, VWCandJs,whereas itwas uncorrelated (�0.5< r< 0.5)withG,u, and udir. Correlations between daily ET and the parameters pa and Tashowed some distinction between sites and seasons. Ta showedpositive correlations and p showed negative correlations with dailyET at KDw06, KDs07, and KDw07, whereas they were uncorrelatedwith daily ET at SM or KDs06.
The lack of correlation between daily ET and VPD at both sitesappeared to reject our first hypothesis. The negative correlationbetween ET and VPD at SM (r¼�0.4) corresponded with theinverse relationship between VWC and VPD at SM. However, eventhe negative correlation disappeared when the data subsets wereanalyzed separately for day and night time correlations (i.e.,conditions Rn> 0 and Rn< 0, respectively). Overall, the correlationanalysis of ET and climate data confirmed our second hypothesisthat VWC was a main driver for ET, whereas it refuted our firsthypothesis because VPD was not correlated with daily ET.
3.2. Empirical modeling
The FAO-PMs with shrub-adapted coefficients overestimateddaily ET roughly 1.4-fold based on thewhole dataset from both sites(Appendix 1a, Appendices electronic version only). The tempera-ture term including VPD� u contributed 68e87% of the variation inthe daily fluxes (Appendix 1b). The model especially overestimated
ETof the subsets SMs04 and SMs05. The radiation term contributedless to model ET but the correlation with measured ET was muchhigher (R2¼ 0.61, Appendix 1c) as compared to the temperatureterm (R2< 0.1).
The FAO-PM with the reference crop coefficients overestimateddaily ET byw2.6 fold and correlated less well with ET based on thewhole dataset (R2¼ 0.29, Fig. 2a) and the individual subsets(R2< 0.1). Compared to FAO-PMs results, the temperature term inthe crop-specific FAO-PM contributed about 20% and <10% less tomodel ET at SM and KD, respectively.
To test our first hypothesis, VPD and u were alternatively elim-inated (i.e., variable value¼ 1) from the FAO-PM simulations.Elimination of VPD resulted in a loss of linear relationship betweenFAO-PM and ET, whereas eliminating u merely decreased the pre-dicted values (Fig. 2aec). Considering the correlation of parameterswith measured ET, we eliminated u and substituted VPD by VWC.With the insertion of VWC in denominator position the relationshipmodeled versus measured ET obtained a shape similar to curves ofsoil water retention (Fig. 2d). Replacement of VWC byJs recoveredand improved the linearity betweenmodel andmeasurements, andit practically aligned soil water conditions with respect to differentsoils (Fig. 2e). Reintroduction of VPD in its original position and inconcert with Js would have increased the scatter of model ET(Fig. 2f).
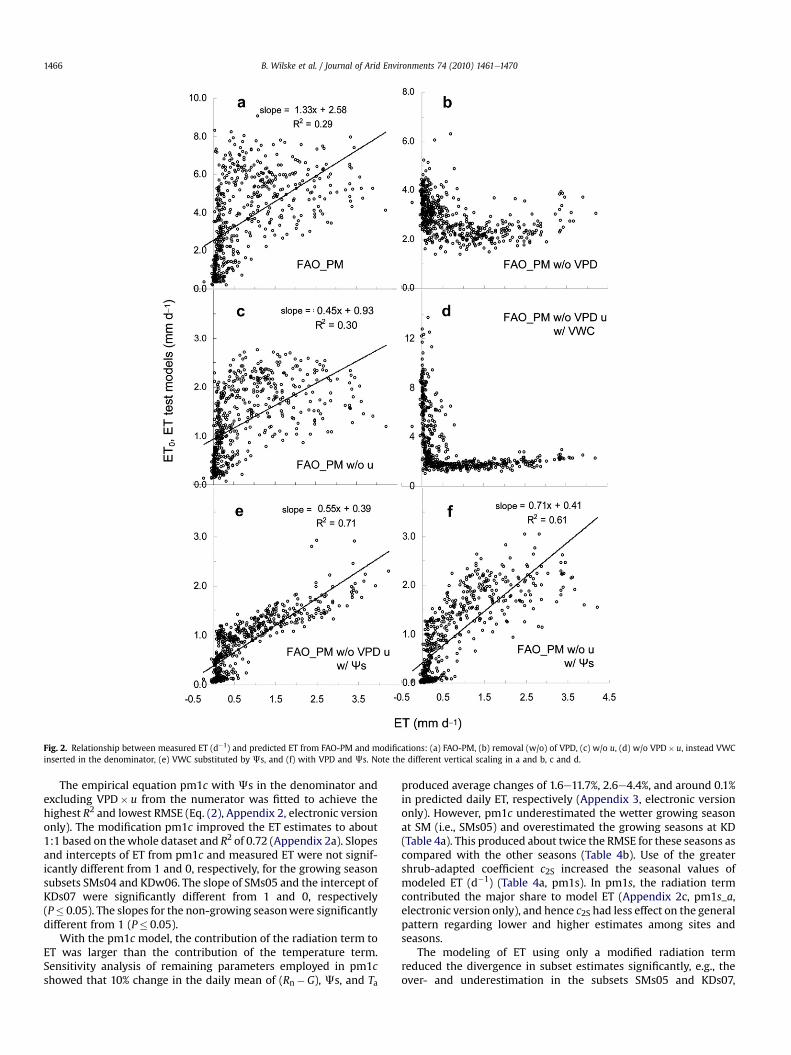
Fig. 2. Relationship between measured ET (d�1) and predicted ET from FAO-PM and modifications: (a) FAO-PM, (b) removal (w/o) of VPD, (c) w/o u, (d) w/o VPD� u, instead VWCinserted in the denominator, (e) VWC substituted by Js, and (f) with VPD and Js. Note the different vertical scaling in a and b, c and d.
B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e14701466
The empirical equation pm1c with Js in the denominator andexcluding VPD� u from the numerator was fitted to achieve thehighest R2 and lowest RMSE (Eq. (2), Appendix 2, electronic versiononly). The modification pm1c improved the ET estimates to about1:1 based on thewhole dataset and R2 of 0.72 (Appendix 2a). Slopesand intercepts of ET from pm1c and measured ET were not signif-icantly different from 1 and 0, respectively, for the growing seasonsubsets SMs04 and KDw06. The slope of SMs05 and the intercept ofKDs07 were significantly different from 1 and 0, respectively(P� 0.05). The slopes for the non-growing seasonwere significantlydifferent from 1 (P� 0.05).
With the pm1c model, the contribution of the radiation term toET was larger than the contribution of the temperature term.Sensitivity analysis of remaining parameters employed in pm1cshowed that 10% change in the daily mean of (Rn�G), Js, and Ta
produced average changes of 1.6e11.7%, 2.6e4.4%, and around 0.1%in predicted daily ET, respectively (Appendix 3, electronic versiononly). However, pm1c underestimated the wetter growing seasonat SM (i.e., SMs05) and overestimated the growing seasons at KD(Table 4a). This produced about twice the RMSE for these seasons ascompared with the other seasons (Table 4b). Use of the greatershrub-adapted coefficient c2S increased the seasonal values ofmodeled ET (d�1) (Table 4a, pm1s). In pm1s, the radiation termcontributed the major share to model ET (Appendix 2c, pm1s_a,electronic version only), and hence c2S had less effect on the generalpattern regarding lower and higher estimates among sites andseasons.
The modeling of ET using only a modified radiation termreduced the divergence in subset estimates significantly, e.g., theover- and underestimation in the subsets SMs05 and KDs07,

Fig. 4. Daily ET (>) and predictions with FAO-PM (—), pm3s (þ) and pm4s (A) duringgrowing seasons at the shrub-dominated sites in the (a, b) Sierra Madre and (c, d)Kubuqi Desert. Note that (1) pm3s and pm4s are often congruent, (2) the different scaleof ET0 in contrast to measured and modeled ET and (3) the pm3s and pm4s modifi-cations perform less well where the course of ET0 (demanding well-watered vegeta-tion) comes close to ET.
B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e1470 1467
respectively. The radiation term was multiplied with the square ofthe quotient of actual pressure to standard pressure at elevation ofsite (Eq. (3), pm2s_a, Appendix 4a, electronic version only). Thispressure correction in pm2s_a resulted in lower RMSE and growingseason estimates with less than 5% deviation from the seasonal ETfor SMs04, SMs05 and KDs06 (Table 4a). To achieve a similar resultwith the temperature term, it was attenuated to 0.1 (Eq. (4), Table4a, pm3s, Appendix 4b).
Along the pm modifications, Js showed a strong control onmodeled ET at high (negative) values but did not show similarregulating power at lower values. In pm3s, a major threshold was atabout (�) 10 MPa (Fig. 3). At values lower than (�) 10 MPa theresiduals between modeled and measured ET increased frommostly less than 0.2 up to 1.4. The lower control of Js on ETcoincided with periods of high VWC when VPD should have thestrongest influence on evaporation from soils and transpirationfrom vegetation. Hence, the attenuation of the temperature termwas combined with a reintroduction of VPD using the product ofVPD�VWC (Eq. (5), pm4s). The model pm4s achieved a 1.00 slopeat seasonal ET estimates and correlations with ET that were similarto pm2s_a and pm3s, respectively (Table 4a, Appendix 4c). TheRMSE in subsets was 0.40e0.54 and not significantly differentbetween sites (Table 4b). The mean deviation between the pm4smodel and measured ET was less than 0.3% per data point (i.e.,deviation (modeled�measured ET) in percent divided by thenumber of days per subset). The modified temperature term inpm4s including VPD�VWC helped in regulating some largeroverestimation at SMs04 and SMs05 (Fig. 4a and b). Conversely,pm4s did not eliminate some major underestimation at KDs06. Taand VPD were low or moderate during these days. Reviewing theoutliers that produced a difference in standard deviations> 2 SDbetween measured and modeled data (Fig. 4c: days 158, 210, 218,221, 226), we found that the preceding days were eliminated fromour analysis because of PPT (1.3e17.7 mm). The records showed noPPT for 3 days preceding d 235 at KDs07 (Fig. 4d, the highest pm4svalue) but 36.9 mm during d 230e231.
4. Discussion
We investigated the biophysical regulation of ET in two semiaridshrub-dominated ecosystems at upland sites and similar northernlatitudes in North America and East Asia.
Fig. 3. Varying control function of water potential in pm3s: 96% of simulated ET atJs� 10 included residuals< 0.2 (right of line a) and 43% at Js< 10 includedresiduals> 0.2 (left of line a and above line c).
Considering the variability that can exist among shrublandsregarding the climate regime and plant height and cover, we foundrather similar characteristics at the two sites. We hypothesized thatVPD and Js were the main drivers of ET at both sites. Correlationanalysis of ET and climate parameters supported our hypothesisrelative to Js but did not indicate that VPD had a major control onET in these ecosystems. Similarly, modifications to the FAO-PMmodel resulted in better ET predictions when VPD and u wereremoved and instead Js was inserted (i.e., models pm1cepm4s).Alternatively, the model pm4s included a stark attenuated VPD andtemperature term and predicted ET with a mean deviation of lessthan 0.3% per day data point at both sites. With both the FAO-PMand the pm modifications, the deviations between modeled andmeasured ET were larger between years than between sites.The agreement in model performance across sites suggests that thebiophysical regulation of ET was similar in the SierraMadre and theKubuqi Desert. Parameters of soil water status (VWC and Js)showed the best correlation with ET among all individual ET-regulating parameters during the growing seasons.Js theoreticallyequalized the soil water status relative to the different soil types(e.g., Passioura, 1980; Kramer and Boyer, 1995), Js in concert withavailable energy was the most efficient parameter in fitting themodeled to the measured daily ET. This suggests that these twoparameters were the key drivers of ET in both shrub-dominatedecosystems.
The importance of Js as driver of ET is in agreement withresults from previous studies showing that ET predictions

B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e14701468
improved significantly when existing models were modified toinclude soil water functions (Wegehenkel, 2000; Fisher et al.,2005). Evans and Trevisan (1995) pointed to the meaningful rela-tion between Js and ET. Rana et al. (1997) used the available soilwater (i.e., a 0e1 scale between field capacity and wilting point) tosubstitute predawn leaf water potential in their model simulatingET from crop under water stress. Recently, Chen et al. (2008)reported an improved linear relation between ET and soil mois-ture when soil moisture falls below a critical value in a Californianoak savanna. This study presented similar results, especially byshowing that Js> (�) 10 MPa had a strong control on ET, whereasJs did not show the same control below that threshold.Js obtainsfurther importance as driver of ET in shrub-dominated ecosystemsbecause the drought resistance of some species (e.g., A. tridentata)is controlled by the cavitation resistance of the root xylem (Kolband Sperry, 1999; Sperry and Hacke, 2002). However, A. ordosicaseems to depend on a shallower root system (Ohte et al., 2003;Cheng et al., 2006) as compared with A. tridentata, which report-edly has additional deep roots facilitating a significant hydraulic liftand water efflux from the upper root system (Caldwell andRichards, 1989). Hence, we have to consider that the depths ofrelevant Js may vary between ecosystems. We compared soilwater status at only one depth, because the remaining depths ofVWC and Js measurements were different at both sites.
The temperature term including VPD and u had the majorinfluence on ET modeled with the FAO-PM including both the cropand shrub-adapted coefficients. However, the temperature term inboth the FAO-PM and -PMs overestimated ET at both sites for thedry growing season (SMs04, KDs07) and underestimated ET at bothsites for the wet growing season (SMs05, KDs06). Furthermore, thecontrol of the temperature term on modeled ET showed no linearrelationship with the measured ET, which made its influencedisputable. Without enhancement by VPD and u, the temperatureterm contributed less than 40% to ET.
VPD was a dominant control variable on carbon fluxes at the SMsite when soil moisture was not limiting (Kwon et al., 2008).Analogous to the previous study, our investigation indicated thatVPD exerted a conditional control on ET when VWC was high andJs was low. Periods with precipitation and/or high VWC areusually short during the growing season in semiarid ecosystems,and our study did not corroborate that VPDwas a key driver of ET atSM and KD. However, days with PPT records were excluded fromour analysis due to the known failures of the EC method at thesetimes. Loik (2007) found effects of precipitation on the waterpotential in A. tridentata being maximal 2e3 days after pulseevents. Similarly in this study, some major outliers from a consis-tent modeled and measured ET course indicated that the influenceof rain events goes even beyond the day scale. We also have toassume that the control of VPD (or VPD� u) on ET is critical afterprecipitation when both soil- and canopy-intercepted waterscontribute to evaporation (e.g., Ziemer, 1979; Olchev et al., 2008).Similarly, frequent precipitation or snowmelt at the onset of thegrowing season can result in longer periods of high VWC. In someareas, both periods, shortly after precipitation events and aftersnowmelt, may account for a critical portion of ET with thepotential to change the monthly water balance.
Several previous studies have also suggested that the influenceof VPD and u on ETcan vary depending on the vegetation and/or theplant functional type of the dominant species (Pataki et al., 2000;Baldocchi et al., 2004; Fisher et al., 2008). The importance of VPDon ET is beyond doubt for grass and tree-dominated ecosystems,where the transpired water is acquired from recently accumulatedprecipitation or is pulled from far below surface. For example, VPDwas the most important atmospheric variable influencing canopytranspiration and ecosystem water use in both deciduous (Ewers
et al., 2002) and coniferous forests (Unsworth et al., 2004) innorthern USA when radiation was not limiting. Aires et al. (2008)found that VPD had a strong control on ET even during periodswith limiting soil moisture in a C3/C4 grassland in Spain. However,the influence of VPD on ET differed between various grain crops(Rana and Katerji, 1998), and the capacity of VPD to regulate ET canbe limited by the prevalence of water stress in dryland ecosystems(Mott and Parkhurst, 1991). Williams et al. (2004), for instance,found soil evaporation was positively correlated with daily VPD inan olive orchard in Morocco, whereas transpiration was not.
Wind speed reportedly caused overestimation of ET undercertain conditions across various ecosystems (Undersander, 1987;Fisher et al., 2005; Persaud et al., 2006). Conversely, Liu et al.(1998) reported surprisingly small influences of u on ecosystemET, as for example both upland slash pine flatwoods and Cypresswetlands in Florida were relatively insensitive to u. According toFAO, the PM model can require local calibration of the wind func-tion to achieve satisfactory results (http://www.fao.org/docrep/X0490E/x0490e06.htm). Wind speed was often high at KD, butour ET simulations performed better for both sites if uwas excludedfrom the model.
The altitude-dependent pressure quotient in pm2s_a, pm3s andpm4s improved the alignment of model outputs for the twoecosystems. Although the locations differed 1100 m in altitude andthe vegetation might adapt to the respective altitude, we do nothave a plausible explanation for a functional effect of the pressurequotient.
5. Conclusions
This study used stepwise modification of the FAO-PM toexamine the regulation of ET by individual biophysical parametersin two semiarid shrub-dominated ecosystems at opposite sides ofthe globe. Modifications to the FAO-PM based on climate parame-tereET correlations showed that soil water potential Js in concertwith available energy was the most important parameter to modelET in these ecosystems. While soil moisture provided the source forlandeatmosphere water fluxes, Js represented the competitor toatmospheric VPD. Over a large part of its range, Js controlled theextractable amount of water that VPD could draw from the semi-arid soils via evaporation and/or plants transpiration. In agreementwith common understanding, Js lost its control at low values(equivalent to high soil moisture). In semiarid environments, this isnot trivial because high (negative) Js is often ruling rather thanexceptional. This relationship was explained by an empirical pmmodel, which specifically describes ET in semiarid shrub-domi-nated ecosystems. The model is essentially driven by the FAO-PMradiation term and Js, whereas it allows much lesser influence ofVPD on ET. While the model is accurate for the two study sites,validation through EC data from other sites is needed to confirm itsgeneral use to predict ET in semiarid shrubs-dominated ecosys-tems. The pm4smodel can be useful to gap-fill missing ET data fromEC measurements. The difference between modeled and measuredET can also assist in identifying disturbance and/or climatic shiftthat have lasting impacts on the hydrology and water balance ina semiarid region.
Acknowledgements
This study was conducted as part of the Northern Eurasia EarthScience Program (NEESPI) and jointly supported by the NationalAeronautics and Space Administration (NASA) Grant (NEWS 2004NRA: NN-H-04-Z-YS-005-N) of the Land-Cover/Land-Use Programand the Natural Science Foundation of China (30928002). It wasfurther supported by the National Research Initiative of the USDA

B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e1470 1469
Cooperative State Research, Education and Extension Service, grantnumber #2003-35101-13652, the Wyoming Agricultural Experi-ment Station, and the USeChina Carbon Consortium (USCCC),which promotes collaborative research among institutions in theUS and China.
Appendix
Supplementary data associated with this article can be found inthe online version, at doi:10.1016/j.jaridenv.2010.05.013.
References
Aires, L.M., Pio, C.A., Pereira, J.S., 2008. The effect of drought on energy and watervapour exchange above a Mediterranean C3/C4 grassland in Southern Portugal.Agricultural and Forest Meteorology 148, 565e579.
Allen, R.G., Pereira, L.S., Raes, D., Smith, M., 1998. Crop Evapotranspiration e
Guidelines for Computing Crop Water Requirements e FAO Irrigation andDrainage Paper 56. FAO, Rome.
Baldocchi, D.D., Xu, L.K., Kiang, N., 2004. How plant functional-type, weather,seasonal drought, and soil physical properties alter water and energy fluxes ofan oakegrass savanna and an annual grassland. Agricultural and Forest Mete-orology 123, 13e39.
Bates, B.C., Kundzewicz, Z.W., Wu, S., Palutikof, J.P., 2008. Climate Change andWater. Technical Paper Intergovernmental Panel on Climate Change. IPCCSecretariat, Geneva.
Caldwell, M.M., Richards, J.H., 1989. Hydraulic lift: water efflux from upper rootsimproves effectiveness of water uptake by deep roots. Oecologia 79, 1e5.
Campbell, G.S., 1988. Soil water potential measurement: an overview. IrrigationScience 9, 265e273.
Chen, X., Rubin, Y., Ma, S., Baldocchi, D., 2008. Observations and stochastic modelingof soil moisture control on evapotranspiration in a Californian oak savanna.Water Resources Research 44, W08409. doi:10.1029/2007WR006646.
Cheng, X., An, S., Li, B., Chen, J., Lin, G., Liu, Y., Luo, Y., Liu, S., 2006. Summer rainpulse size and rainwater uptake by three dominant desert plants in a deserti-fied grassland ecosystem in northwestern China. Plant Ecology 184, 1e12.
Cleary, M.B., Pendall, E., Ewers, B.E., 2008. Testing sagebrush allometric relation-ships across three fire chronosequences in Wyoming, USA. Journal of AridEnvironments 72, 285e301.
Evans, S.P., Trevisan, M., 1995. A soil water-balance ‘bucket’ model for paleoclimaticpurposes. 1. Model structure and validation. Ecological Modelling 82, 109e129.
Ewers, B.E., Pendall, E., 2007. Spatial patterns in leaf area and plant functional typecover across chronosequences of sagebrushecosystems. Plant Ecology194, 67e83.
Ewers, B.E., Mackay, D.S., Gower, S.T., Ahl, D.E., Burrows, S.N., Samanta, S.S., 2002.Tree species effects on stand transpiration in northern Wisconsin. WaterResources Research 38, 1103, 11 pp. doi:10.1029/2001WR000830.
Fischer, B., Goldberg, V., Bernhofer, C., 2008. Effect of a coupled soil watereplant gasexchange on forest energy fluxes: simulations with the coupled vegeta-tioneboundary layer model HIRVAC. Ecological Modelling 214, 75e82.
Fisher, J.B., DeBiase, T.A., Qi, Y., Xu, M., Goldstein, A.H., 2005. Evapotranspirationmodels compared on a Sierra Nevada forest ecosystem. EnvironmentalModelling and Software 20, 783e796.
Fisher, J.B., Tu, K.P., Baldocchi, D.D., 2008. Global estimates of the landeatmospherewater flux based on monthly AVHRR and ISLSCP-II data, validated at 16 FLUX-NET sites. Remote Sensing of the Environment 112, 901e919.
Frank, A.B., Karn, J.F., 2005. Shrub effects on carbon dioxide and water vapor fluxesover grasslands. Rangeland Ecology and Management 58, 20e26.
Hargreaves, G.L., Hargreaves, G.H., Riley, J.P., 1985. Agricultural benefits for SenegalRiver basin. Journal of Irrigation and Drainage Engineering 111, 113e124.
Jarvis, P.G., McNaughton, K.G., 1986. Stomatal control of transpiration: scaling upfrom leaf to region. Advances in Ecological Research 15, 1e49.
Kolb, K.J., Sperry, J.S., 1999. Differences in drought adaptation between subspecies ofsagebrush (Artemisia tridentata). Ecology 80, 2373e2384.
Kramer, P.J., Boyer, J.S., 1995. Water Relations of Plants and Soil. Academic Press, SanDiego.
Kwon, H., Pendall, E., Ewers, B.E., Cleary, M., Naithani, K., 2008. Spring droughtregulates summer net ecosystem CO2 exchange in a sagebrush-steppeecosystem. Agricultural and Forest Meteorology 148, 381e391.
Liu, S., Riekerk, H., Gholz, H.L., 1998. Simulation of evapotranspiration from Floridapine flatwoods. Ecological Modelling 114, 19e34.
Loik, M.E., 2007. Sensitivity of water relations and photosynthesis to summerprecipitation pulses for Artemisia tridentata and Purshia tridentate. Plant Ecology191, 95e108.
Loveland, T.R., Reed, B.C., Brown, J.F., Ohlen, D.O., Zhu, Z., Yang, L., Merchant, J.W.,2000. Development of a global land cover characteristics database and IGBPDISCover from 1 km AVHRR data. International Journal of Remote Sensing 21,1303e1330.
Mackay, D.S., Ewers, B.E., Cook, B.D., Davis, K.J., 2007. Environmental drivers ofevapotranspiration in a shrub wetland and an upland forest in northern Wis-consin. Water Resources Research 43, W03442, 14 pp. doi:10.1029/2006WR005149.
Makkink, G.F., 1957. Testing the Penman formula by means of lysimeters. Journal ofthe Institute of Water Engineers 11, 277e288.
Meidner, H., 1979. Water uptake and translocation, stomatal movements andtranspiration. In: Goodall, D.W., Perry, R.A. (Eds.), Arid Land Ecosystems:Structure, Functioning, and Management, vol. 1. Cambridge University Press,Cambridge, pp. 491e507.
Monteith, J.L., 1965. Evaporation and environment. In: Fogg, G. (Ed.), The State andMovement of Water in Living Organisms. Proceedings of the XIXth Symposiumof the Society for Experimental Biology. Cambridge University Press,Cambridge, UK, pp. 205e234.
Mott, K.A., Parkhurst, D.F., 1991. Stomatal responses to humidity in air and helox.Plant Cell and Environment 14, 509e515.
Ohte, N., Koba, K., Yoshikawa, K., Sugimoto, A., Matsuo, N., Kabeya, N., Wang, L.H.,2003. Water utilization of natural and planted trees in the semiarid desert ofInner Mongolia, China. Ecological Applications 13, 337e351.
Olchev, A., Ibrom, A., Priess, J., Erasmi, S., Leemhuis, C., Twele, A., Radler, K.,Kreilein, H., Panferov, O., Gravenhorst, G., 2008. Effects of land-use changes onevapotranspiration of tropical rain forest margin area in Central Sulawesi(Indonesia): modelling study with a regional SVAT model. Ecological Modelling212, 131e137.
Passioura, J.B., 1980. The meaning of matrix potential. Journal of ExperimentalBotany 31, 1161e1169.
Pataki, D.E., Huxman, T.E., Jordan, D.N., Zitzer, S.F., Coleman, J.S., Smith, S.D.,Nowak, R.S., Seemann, J.R., 2000. Water use of two Mojave Desert shrubs underelevated CO2. Global Change Biology 6, 889e897.
Pereira, A.R., Pruitt, W.O., 2004. Adaptation of the Thornthwaite scheme for esti-mating daily reference evapotranspiration. Agricultural Water Management 66,251e257.
Persaud, N., Lesolle, D., Zhou, X., Wang, H., Joshua, W., 2006. Coefficients forPenman-based estimation of daily potential evapotranspiration in Botswana.Journal of Arid Environments 66, 764e772.
Priestley, C.H.B., Taylor, R.J., 1972. On the assessment of surface heat flux andevaporation using large-scale parameters. Monthly Weather Review 100,81e92.
Rana, G., Katerji, N., 1998. A measurement based sensitivity analysis of the Pen-maneMonteith actual evapotranspiration model for crops of different heightand in contrasting water status. Theoretical and Applied Climatology 60,141e149.
Rana, G., Katerji, N., Mastrorilli, M., 1997. Environmental and soileplant parametersfor modelling actual crop evapotranspiration under water stress conditions.Ecological Modelling 101, 363e371.
Rey, J.M., 1999. Modelling potential evapotranspiration of potential vegetation.Ecological Modelling 123, 141e159.
Scott, R.L., Huxman, T.E., Williams, D.G., Goodrich, D.C., 2006. Ecohydrologicalimpacts of woody-plant encroachment: seasonal patterns of water and carbondioxide exchange within a semiarid riparian environment. Global ChangeBiology 12, 311e324.
Sperry, J.S., Hacke, U.G., 2002. Desert shrub water relations with respect to soilcharacteristics and plant functional type. Functional Ecology 16, 367e378.
Stephenson, N.L., 1990. Climatic control of vegetation distribution: the role of thewater balance. American Naturalist 135, 649e670.
Thornthwaite, C.W., 1948. An approach toward a rational classification of climate.Geographical Review 38, 55e94.
Undersander, D.J., 1987. Alfalfa (Medicago sativa L.) growth response to water andtemperature. Irrigation Science 8, 23e33.
Unsworth, M.H., Phillips, N., Link, T., Bond, B.J., Falk, M., Harmon, M.E.,Hinckley, T.M., Marks, D., Paw, U.K.T., 2004. Components and controls ofwater flux in an old-growth Douglas-fireWestern Hemlock ecosystem.Ecosystems 7, 468e481.
Vezzoli, R., De Michele, C., Pavlopoulos, H., Scholes, R.J., 2008. Dryland ecosystems:the coupled stochastic dynamics of soil water and vegetation and the role ofrainfall seasonality. Physical Review E 77, 051908-1e051908-9.
Vivoni, E.R., Watts, C.J., Rodriguez, J.C., Garatuza-Payan, J., Mendez-Barroso, L.A.,Saiz-Hernandez, J.A., 2010. Improved landeatmosphere relations throughdistributed footprint sampling in a subtropical scrubland during the NorthAmerican monsoon. Journal of Arid Environments 74, 579e584.
Walker, B.H., Langridge, J.L., 1996. Modelling plant and soil water dynamics in semi-arid ecosystems with limited site data. Ecological Modelling 87, 153e167.
Webb, E.K., Pearman, G.I., Leuning, R., 1980. Correction of the flux measurements fordensity effects due to heat and water vapour transfer. Quarterly Journal of theRoyal Meteorological Society 106, 85e100.
Wegehenkel, M., 2000. Test of a modelling system for simulating water balancesand plant growth using various different complex approaches. EcologicalModelling 129, 39e64.
Williams, D.G., Cable, W., Hultine, K., Hoedjes, J.C.B., Yepez, E.A., Simonneaux, V.,Er-Raki, S., Boulet, G., de Bruin, H.A.R., Chehbouni, A., Hartogensis, O.K.,Timouk, F., 2004. Evapotranspiration components determined by stable isotope,sap flow and eddy covariance techniques. Agricultural and Forest Meteorology125, 241e258.
Williams, K., Caldwell, M.M., Richards, J.H., 1993. The influence of shade and cloudson soil water potential: the buffered behavior of hydraulic lift. Plant and Soil157, 83e95.
Wilske, B., Lu, N., Wei, L., Chen, S., Zha, T., Liu, C., Xu, W., Noormets, A., Huang, J.,Wei, Y., Zhang, Z., Ni, J., Sun, G., Guo, K., McNulty, S., John, R., Han, X., Lin, G.,Chen, J., 2009. Poplar plantation has the potential to alter the water balance in

B. Wilske et al. / Journal of Arid Environments 74 (2010) 1461e14701470
semiarid Inner Mongolia. Journal of Environmental Management 90,2762e2770.
Xiao, C.W., Zhou, G.S., Ceulemans, R., 2003. Effects of elevated temperature ongrowth and gas exchange in dominant plant species from Maowusu Sandland,China. Photosynthetica 41, 565e569.
Xiao, C.W., Zhou, G.S., Zhang, X.S., Zhao, J.Z., Wu, G., 2005. Responses of dominantdesert species Artemisia ordosica and Salix psammophila to water stress. Pho-tosynthetica 43, 467e471.
Xu, C.Y., Gong, L., Jiang, T., Chen, D., Singh, V.P., 2006. Analysis of spatial distributionand temporal trend of reference evapotranspiration and pan evaporation inChangjiang (Yangtze River) catchment. Journal of Hydrology 327, 81e93.
Zhang, X.S.,1994. The ecological background of theMaowusu sandland: principles andoptimal models for grass land management. Acta Phytoecologica Sinica 18, 1e6.
Zhang, Y., Chiew, F.H.S., Zhang, L., Leuning, R., Cleugh, H.A., 2008. Estimatingcatchment evaporation and runoff using MODIS leaf area index and the Pen-maneMonteith equation. Water Resources Research. 44, W10420, 15 pp.doi:10.1029/2007WR006563.
Zheng, Y., Xie, Z., Jiang, L., Shimizu, H., Rimmington, G.M., Zhou, G., 2006. Vegetationresponses along environmental gradients on the Ordos plateau, China.Ecological Research 21, 396e404.
Ziemer, R.R., 1979. Evaporation and transpiration. Reviews of Geophysics and SpacePhysics 17, 1175e1186.




















