
Evaluation of reference genes for expression studies in leukocytes from term human pregnancy
6
Evaluation of reference genes for expression studies in leukocytes from term human pregnancy R. Vega-Sanchez a, * , M. Arenas-Hernandez b, c , J.A. Vazquez-Perez d , Y. Moreno-Valencia d , N. Gomez-Lopez b, e a Department of Nutrition and Bioprogramming, Instituto Nacional de Perinatología, Montes Urales 800, M exico D.F. 11000, Mexico b Department of Obstetrics & Gynecology, School of Medicine, Wayne State University, Detroit, MI, USA c National School of Biological Sciences, Instituto Polit ecnico Nacional, M exico D.F., Mexico d Department of Virology and Mycology Research, Instituto Nacional de Enfermedades Respiratorias, Calzada de Tlalpan 4502, M exico D.F. 14080, Mexico e Department of Obstetrics & Gynecology, Immunology & Microbiology, Perinatology Research Branch NICHD/NIH, School of Medicine, Wayne State University, C.S. Mott Center for Human Growth and Development, Room 305, 275 E. Hancock, Detroit, MI 48201, USA article info Article history: Accepted 14 November 2014 Keywords: Pregnancy Labor Leukocytes Housekeeping genes Stability geNorm abstract Introduction: Human labor is considered an inflammatory process modulated by systemic and local leukocytes that infiltrate into the maternalefetal interface. The putative roles of these leukocytes are currently being studied with gene expression assays. Such assays are normalized by the expression of housekeeping genes. However, expression of housekeeping genes may vary depending on the cell type and/or the experimental conditions. The aim of this study was to analyze the expression stability of several housekeeping genes in leukocytes from term human pregnancies, considering both anatomical origin and presence of labor. Methods: We analyzed the gene expression of ACTB, B2M, GAPDH, GUSB, PGK1, RN18S1 , TBP and UBC in leukocytes from maternal peripheral blood, placental blood and choriodecidua in women delivering at term with or without the presence of labor through real-time qPCR. Then we used geNorm to evaluate expression stability and pairwise variation. Results: The expression of all tested genes showed to be stable independent of the anatomical compartment and the absence or presence of labor. However, PGK1, GUSB and TBP showed to be the most stable and RN18S1 the least stable. Pairwise variation analyses showed that only two genes are needed for normalization yet the inclusion of a third improves its accuracy. Discussion: PGK1 , GUSB and TBP are the most adequate reference genes for gene expression normaliza- tion in leukocytes from term pregnancies, regardless of their anatomical origin (maternal peripheral blood, placental blood or choriodecidua) or the presence or absence of labor. Our study is the first report on housekeeping gene stability in leukocytes from healthy pregnant women. © 2014 Elsevier Ltd. All rights reserved. 1. Introduction Human parturition has been described as the result of an in- flammatory process that takes place within the intrauterine milieu when pregnancy comes to term [1], which seems to be modulated by specific leukocyte subsets that infiltrate into the choriodecidual tissues [2] and show an increased proinflammatory capacity when compared to peripheral blood leukocytes [3,4]. The characteriza- tion of these infiltrating leukocytes has been recently undertaken using cellular and molecular techniques, including gene expression assays. Housekeeping gene expression is used to normalize the expression of target genes in every expression assay. Over the last years, several studies have shown that housekeeping gene expression may vary depending on the cell type and/or the experimental conditions [5e8], stressing the importance of analyzing their expression in particular experimental conditions. * Corresponding author. Tel.: þ52 55 55209900x120. E-mail addresses: [email protected] (R. Vega-Sanchez), [email protected]. edu (M. Arenas-Hernandez), [email protected] (J.A. Vazquez-Perez), [email protected] (Y. Moreno-Valencia), [email protected] (N. Gomez-Lopez). Contents lists available at ScienceDirect Placenta journal homepage: www.elsevier.com/locate/placenta http://dx.doi.org/10.1016/j.placenta.2014.11.010 0143-4004/© 2014 Elsevier Ltd. All rights reserved. Placenta 36 (2015) 240e245
Transcript of Evaluation of reference genes for expression studies in leukocytes from term human pregnancy

lable at ScienceDirect
Placenta 36 (2015) 240e245
Contents lists avai
Placenta
journal homepage: www.elsevier .com/locate/placenta
Evaluation of reference genes for expression studies in leukocytesfrom term human pregnancy
R. Vega-Sanchez a, *, M. Arenas-Hernandez b, c, J.A. Vazquez-Perez d, Y. Moreno-Valencia d,N. Gomez-Lopez b, e
a Department of Nutrition and Bioprogramming, Instituto Nacional de Perinatología, Montes Urales 800, M�exico D.F. 11000, Mexicob Department of Obstetrics & Gynecology, School of Medicine, Wayne State University, Detroit, MI, USAc National School of Biological Sciences, Instituto Polit�ecnico Nacional, M�exico D.F., Mexicod Department of Virology and Mycology Research, Instituto Nacional de Enfermedades Respiratorias, Calzada de Tlalpan 4502, M�exico D.F. 14080, Mexicoe Department of Obstetrics & Gynecology, Immunology & Microbiology, Perinatology Research Branch NICHD/NIH, School of Medicine, Wayne StateUniversity, C.S. Mott Center for Human Growth and Development, Room 305, 275 E. Hancock, Detroit, MI 48201, USA
a r t i c l e i n f o
Article history:Accepted 14 November 2014
Keywords:PregnancyLaborLeukocytesHousekeeping genesStabilitygeNorm
* Corresponding author. Tel.: þ52 55 55209900x12E-mail addresses: [email protected] (R. Vega-Sanc
edu (M. Arenas-Hernandez), [email protected]@gmail.com (Y. Moreno-Valencia),(N. Gomez-Lopez).
http://dx.doi.org/10.1016/j.placenta.2014.11.0100143-4004/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Introduction: Human labor is considered an inflammatory process modulated by systemic and localleukocytes that infiltrate into the maternalefetal interface. The putative roles of these leukocytes arecurrently being studied with gene expression assays. Such assays are normalized by the expression ofhousekeeping genes. However, expression of housekeeping genes may vary depending on the cell typeand/or the experimental conditions. The aim of this study was to analyze the expression stability ofseveral housekeeping genes in leukocytes from term human pregnancies, considering both anatomicalorigin and presence of labor.Methods: We analyzed the gene expression of ACTB, B2M, GAPDH, GUSB, PGK1, RN18S1, TBP and UBC inleukocytes from maternal peripheral blood, placental blood and choriodecidua in women delivering atterm with or without the presence of labor through real-time qPCR. Then we used geNorm to evaluateexpression stability and pairwise variation.Results: The expression of all tested genes showed to be stable independent of the anatomicalcompartment and the absence or presence of labor. However, PGK1, GUSB and TBP showed to be the moststable and RN18S1 the least stable. Pairwise variation analyses showed that only two genes are neededfor normalization yet the inclusion of a third improves its accuracy.Discussion: PGK1, GUSB and TBP are the most adequate reference genes for gene expression normaliza-tion in leukocytes from term pregnancies, regardless of their anatomical origin (maternal peripheralblood, placental blood or choriodecidua) or the presence or absence of labor. Our study is the first reporton housekeeping gene stability in leukocytes from healthy pregnant women.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Human parturition has been described as the result of an in-flammatory process that takes place within the intrauterine milieuwhen pregnancy comes to term [1], which seems to be modulated
0.hez), [email protected] (J.A. Vazquez-Perez),[email protected]
by specific leukocyte subsets that infiltrate into the choriodecidualtissues [2] and show an increased proinflammatory capacity whencompared to peripheral blood leukocytes [3,4]. The characteriza-tion of these infiltrating leukocytes has been recently undertakenusing cellular and molecular techniques, including gene expressionassays.
Housekeeping gene expression is used to normalize theexpression of target genes in every expression assay. Over the lastyears, several studies have shown that housekeeping geneexpression may vary depending on the cell type and/or theexperimental conditions [5e8], stressing the importance ofanalyzing their expression in particular experimental conditions.

R. Vega-Sanchez et al. / Placenta 36 (2015) 240e245 241
Furthermore, it has been demonstrated that using a single refer-ence gene or an unstable gene rather than using two or morereference genes for normalizing the expression of a target gene, willyield different results [9e11].
Despite of these evidences, studies analyzing the stability ofhousekeeping genes in leukocytes from healthy individuals arescarce, even with contradictory results partly due to differentanalytical strategies [9,12,13], and none of the available studiesincluded pregnant women. Furthermore, there is not a single studywith leukocytes involved in pregnancy-related events addressingthe stability of these genes. In this study, we report for the first timethe expression stability of several housekeeping genes in leuko-cytes from healthy women with term pregnancies, analyzingwhether or not the anatomical compartment (peripheral blood,placental blood or choriodecidua) or the physiologic event (no la-bor or active labor) have any effects on said stability.
2. Methods
2.1. Participating women
This project was approved by the Internal Review Board of the Instituto Nacionalde Perinatología in Mexico City (register 212250-02191) and written informedconsent was obtained from all participating women. We included healthy womenwith term pregnancies (>37 weeks of gestation) who underwent elective cesareansection (n ¼ 25) or labor (n ¼ 19). Labor was considered to be present whenwomenpresented signs of uterine activity and cervical dilation was >2 cm. Women withdiabetes, autoimmune diseases, multiple pregnancy or clinical signs of infectionwere excluded.
2.2. Isolation of leukocytes
We isolated leukocytes from maternal peripheral blood (collected immediatelyafter delivery by venous puncture of the forearm) and from intervillous placentalblood (collected by manually compressing the cotyledons) by density gradient usingPolymorphprep (Axis-Shield, Oslo, Norway), recovering both the mononuclear andpolymorphonuclear bands. Leukocytes were washed with 1� PBS, incubated witherythrocyte lysis buffer (150 mM ammonium chloride, 10 mM potassium bicar-bonate, 0.1 mM EDTA) for 15 min and washed again with 1� PBS.
Additionally, to isolate choriodecidual leukocytes, the chorion of fetal mem-branes (cut from the placenta) was manually separated from the amnion, cut intosmall pieces (approximately 3� 3 cm) and incubated at 37 �C for 5e10minwith lowagitation in 100 ml of DMEM containing 0.1% trypsin for enzymatic digestion of thechoriodecidua. Media was recovered, diluted with 1� PBS and centrifuged. Pelletedcells were incubated in erythrocyte lysis buffer for 15 min and washed with 1� PBS.Choriodecidual leukocytes were then isolated from other cells by positive-selectionmagnetic cell sorting (MACS) using an anti-CD45 microbead-conjugated antibodyand MS columns, following the manufacturer's protocol (Miltenyi Biotec, Germany).All buffers were ice-cooled to minimize RNA degradation.
2.3. RNA isolation and cDNA synthesis
We isolated total RNA from leukocytes of the three anatomical compartmentsusing Trizol reagent following the manufacturer's protocol (Invitrogen, Carlsbad, CA,USA) and analyzed RNA integrity on half of the samples in an Agilent 2100 Bio-analyzer using RNA 6000 Nano kit (Agilent Technologies, Santa Clara, CA, USA). Allsamples showed RNA integrity numbers (RIN) over 7. Complementary DNA wassynthesized from approximately 500 ng of total RNA with the Transcriptor FirstStrand cDNA Synthesis Kit (Roche Applied Science, Mannheim), using randomhexamers, at 25 �C-10 min/55 �C-30 min/85 �C-5 min. cDNA was stored at �20 �Cuntil further used.
Table 1Primer sequences and amplification efficiencies.
Gene symbol RefSeq Forward primer
ACTB NM_001101.3 ccaaccgcgagaagatgacB2M NM_004048.2 tccgtggccttagctgtgGAPDH NM_002046.3 acaccatggggaaggtgaagGUSB NM_000181.3 catcgatgacatcaccgtcacPGK1 NM_000291.3 gtggaatggcttttaccttccRNA18S1 NR_003286.2 acgagactctggcatgctaaTBP NM_001172085.1 tgcccgaaacgccgaatataUBC NM_021009.5 tcggccttagaaccccagta
Amplification efficiencies were calculated from the slopes of serial dilution curves using
2.4. Quantitative real-time PCR
We analyzed the housekeeping genes ACTB, B2M, GAPDH, GUSB, PGK1, RNA18S1,TBP and UBC by high throughput gene expression analysis using DNA binding dye(Evagreen) for detection and highly specific primers designed for each gene (DEL-TAgene Assays, Fluidigm Corporation, San Francisco, CA, USA). We selected thesegenes over other housekeeping genes (like YWHAZ, HMBS, HPRT or SDHA) becausepredesigned, validated assays were already available from the manufacturer. Primersequences and amplification efficiencies are shown in Table 1.
First we performed a specific target preamplification reaction on each cDNAusing 2� TaqMan PreampMasterMix (Applied Biosystems, Foster City, CA, USA) and500 nM pooled primer mixture. Preamplification was carried at 95 �Ce10 min/14cycles of 95 �Ce15 s and 60 �Ce4 min. A cleanup step of preamplification productswas performed with 8 U of Exonuclease I (New England Biolabs, Ipswich, MA, USA)at 37 �Ce30 min/80 �C�15 min. The final clean-up products were diluted 1:5 withTE buffer.
For the qPCR, we followed the Fast Gene Expression Analysis Using EvaGreen onthe Biomark System Protocol from Fluidigm to prepare assay mixes (100 mM of eachpair of primers, 2X Assay Loading Reagent, 1� TE buffer) and sample mixes (dilutedpreamplified Exonuclease-treated cDNA, 2� SsoFast Master mix (BioRad, Hercules,CA, USA), 20� DNA Binding Dye Sample Loading Reagent). Assay and sample mixeswere loaded into the 48.48 Dynamic Array using the IFC Controller MX and thentransferred to the BioMark for qPCR (95 �C for 10 min; 40 cycles of 95 �C for 15 s and60 �C for 60 s). Each assay was run in duplicate and the results averaged. All samplesshowed Ct values <30 and inter-plate coefficient of variation was 9.83 (average ofthe ten samples with the lowest Ct values for every gene).
2.5. Analysis of housekeeping gene stability and variability
We used the geNorm VBA applet for Microsoft Excel (available at http://medgen.ugent.be/genorm/) to analyze expression stability and variability. It measures geneexpression stability (M) for a reference gene, as the average pairwise variation forthat gene against all other tested reference genes. Genes with the smallest M value(lower than 1.5) are considered the most stable.
In order to analyze whether or not the anatomical compartment (peripheralblood, placental blood or choriodecidua) or the physiological event (no labor orlabor) affects the stability of housekeeping genes, we performed two sets of ana-lyses. The first set tested the stability between compartments and was comprised oftwo different analyses, one with samples from women without labor and the otherwith samples fromwomen in labor. The second set tested the stability of genes in theabsence or presence of labor and was comprised of three different analyses, one foreach anatomical compartment. In order to confirm our results, we performed ananalysis including all samples.
Moreover, we also calculated the pairwise variation (Vn/n þ 1) in order toevaluate the minimum number of genes required for normalization in comparativegene expression analyses. According to Vandesompele et al., a pairwise variation of0.15 is considered to be the cut-off value below which the inclusion of an additionalreference gene is not required [9].
Since geNorm requires expression quantities (not Ct values), all Ct values weretransformed into said quantities using the comparative Ct method. Briefly, thehighest Ct value for each gene was set to 1 and values for the rest of the sampleswere calculated relative to this value. These expression quantities were the inputdata for geNorm.
3. Results
Fig. 1 shows the distribution of Ct values of all the testedgenes. All samples showed Ct values <30. From this graph, genescan be roughly grouped according to their median expressionlevels into three groups: high abundance genes with median Ctvalues �5, comprising RNA18S1 and B2M; medium abundancegenes with median Ct values between 5 and 15, comprising ACTB,
Reverse primer Amplification efficiency
tagcacagcctggatagcaa 1.075cccagacacatagcaattcagg 1.030gtgaccaggcgcccaata 1.043acaggttactgcccttgaca 0.985cttggctccctcttcatcaa 0.970aacgccacttgtccctctaa 1.053cgtggttcgtggctctctta 1.127gaaaaccagtgccctagagtca 1.067
the equation E ¼ 10(�1/m) e 1.
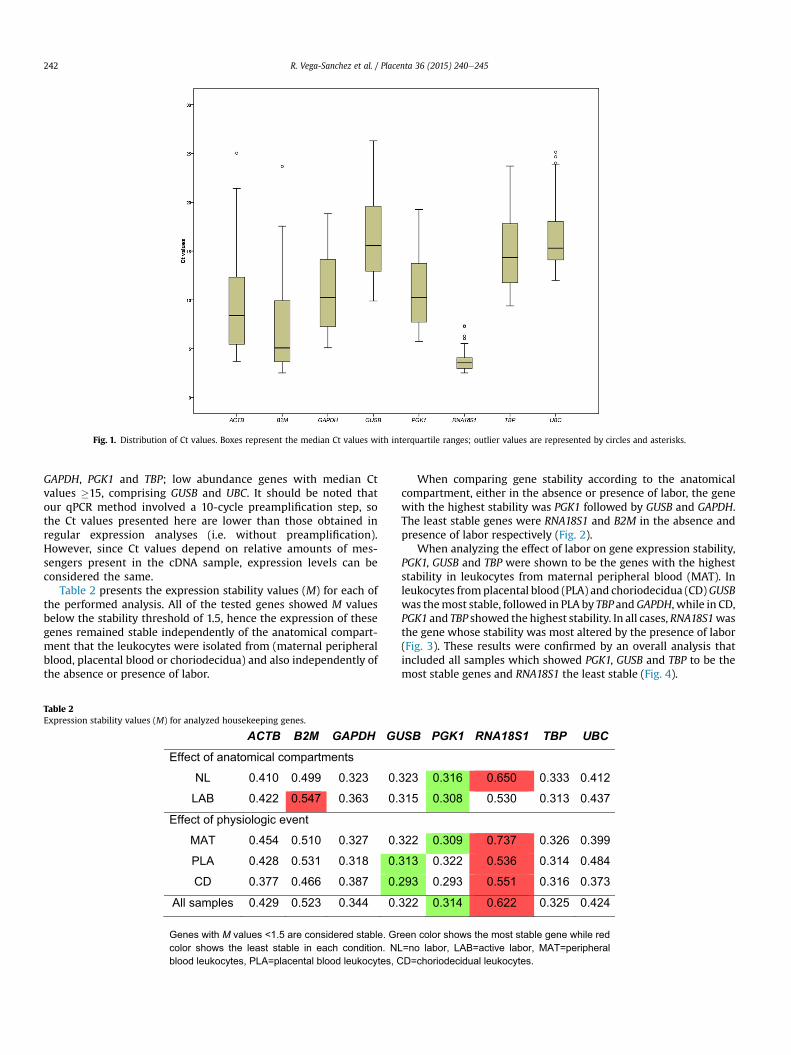
Fig. 1. Distribution of Ct values. Boxes represent the median Ct values with interquartile ranges; outlier values are represented by circles and asterisks.
R. Vega-Sanchez et al. / Placenta 36 (2015) 240e245242
GAPDH, PGK1 and TBP; low abundance genes with median Ctvalues �15, comprising GUSB and UBC. It should be noted thatour qPCR method involved a 10-cycle preamplification step, sothe Ct values presented here are lower than those obtained inregular expression analyses (i.e. without preamplification).However, since Ct values depend on relative amounts of mes-sengers present in the cDNA sample, expression levels can beconsidered the same.
Table 2 presents the expression stability values (M) for each ofthe performed analysis. All of the tested genes showed M valuesbelow the stability threshold of 1.5, hence the expression of thesegenes remained stable independently of the anatomical compart-ment that the leukocytes were isolated from (maternal peripheralblood, placental blood or choriodecidua) and also independently ofthe absence or presence of labor.
Table 2Expression stability values (M) for analyzed housekeeping genes.
ACTB B2M GAPDH GU
Effect of anatomical compartments
NL 0.410 0.499 0.323 0.
LAB 0.422 0.547 0.363 0.
Effect of physiologic event
MAT 0.454 0.510 0.327 0.
PLA 0.428 0.531 0.318 0.
CD 0.377 0.466 0.387 0.
All samples 0.429 0.523 0.344 0.
Genes with M values <1.5 are considered stable. Grcolor shows the least stable in each condition. NLblood leukocytes, PLA=placental blood leukocytes, C
When comparing gene stability according to the anatomicalcompartment, either in the absence or presence of labor, the genewith the highest stability was PGK1 followed by GUSB and GAPDH.The least stable genes were RNA18S1 and B2M in the absence andpresence of labor respectively (Fig. 2).
When analyzing the effect of labor on gene expression stability,PGK1, GUSB and TBP were shown to be the genes with the higheststability in leukocytes from maternal peripheral blood (MAT). Inleukocytes fromplacental blood (PLA) and choriodecidua (CD)GUSBwas themost stable, followed in PLA by TBP andGAPDH, while in CD,PGK1 and TBP showed the highest stability. In all cases, RNA18S1wasthe gene whose stability was most altered by the presence of labor(Fig. 3). These results were confirmed by an overall analysis thatincluded all samples which showed PGK1, GUSB and TBP to be themost stable genes and RNA18S1 the least stable (Fig. 4).
SB PGK1 RNA18S1 TBP UBC
323 0.316 0.650 0.333 0.412
315 0.308 0.530 0.313 0.437
322 0.309 0.737 0.326 0.399
313 0.322 0.536 0.314 0.484
293 0.293 0.551 0.316 0.373
322 0.314 0.622 0.325 0.424
een color shows the most stable gene while red =no labor, LAB=active labor, MAT=peripheral D=choriodecidual leukocytes.
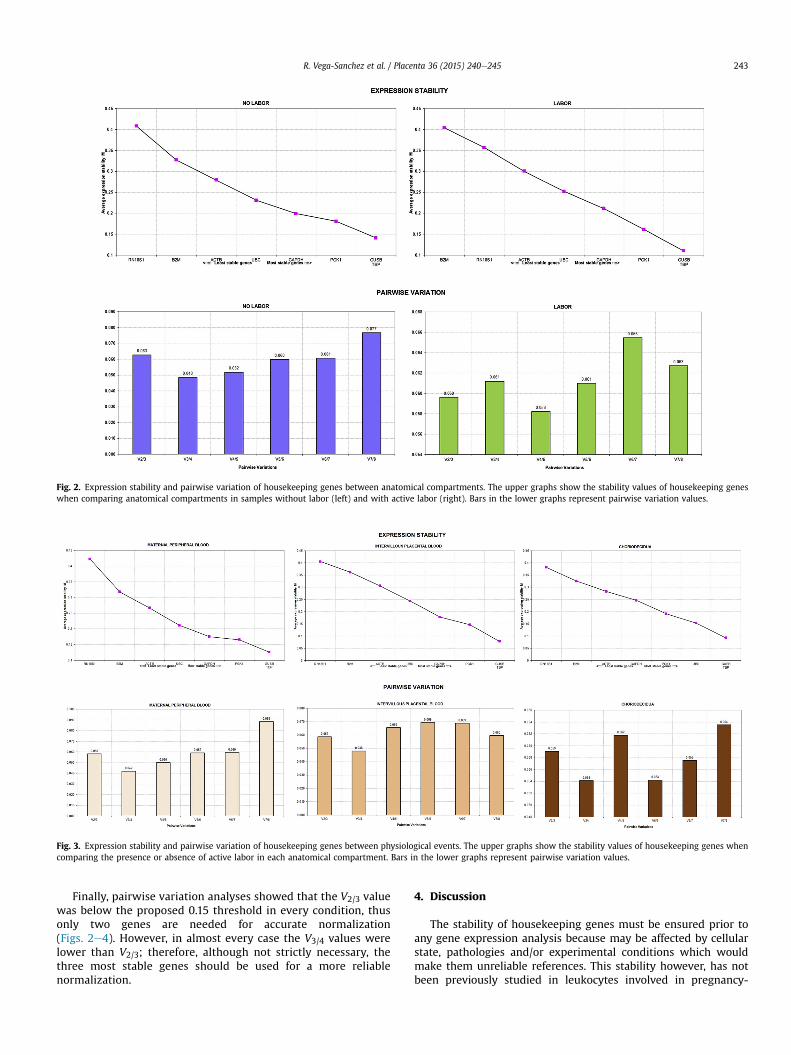
Fig. 2. Expression stability and pairwise variation of housekeeping genes between anatomical compartments. The upper graphs show the stability values of housekeeping geneswhen comparing anatomical compartments in samples without labor (left) and with active labor (right). Bars in the lower graphs represent pairwise variation values.
Fig. 3. Expression stability and pairwise variation of housekeeping genes between physiological events. The upper graphs show the stability values of housekeeping genes whencomparing the presence or absence of active labor in each anatomical compartment. Bars in the lower graphs represent pairwise variation values.
R. Vega-Sanchez et al. / Placenta 36 (2015) 240e245 243
Finally, pairwise variation analyses showed that the V2/3 valuewas below the proposed 0.15 threshold in every condition, thusonly two genes are needed for accurate normalization(Figs. 2e4). However, in almost every case the V3/4 values werelower than V2/3; therefore, although not strictly necessary, thethree most stable genes should be used for a more reliablenormalization.
4. Discussion
The stability of housekeeping genes must be ensured prior toany gene expression analysis because may be affected by cellularstate, pathologies and/or experimental conditions which wouldmake them unreliable references. This stability however, has notbeen previously studied in leukocytes involved in pregnancy-
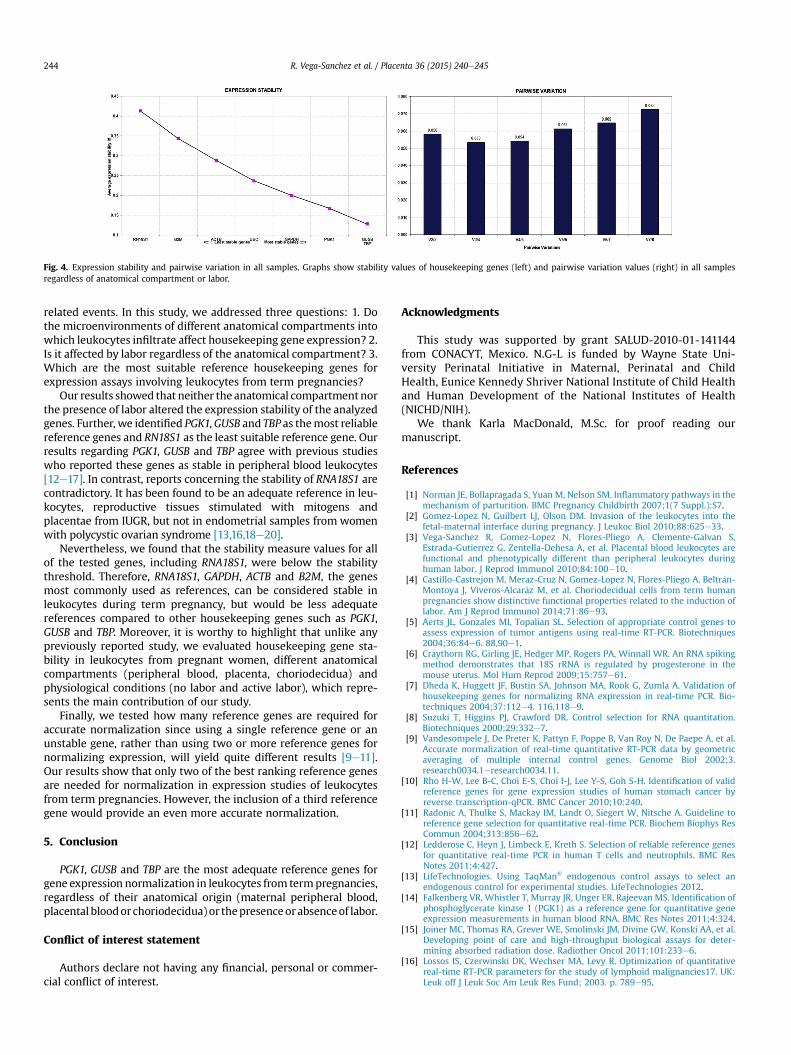
Fig. 4. Expression stability and pairwise variation in all samples. Graphs show stability values of housekeeping genes (left) and pairwise variation values (right) in all samplesregardless of anatomical compartment or labor.
R. Vega-Sanchez et al. / Placenta 36 (2015) 240e245244
related events. In this study, we addressed three questions: 1. Dothe microenvironments of different anatomical compartments intowhich leukocytes infiltrate affect housekeeping gene expression? 2.Is it affected by labor regardless of the anatomical compartment? 3.Which are the most suitable reference housekeeping genes forexpression assays involving leukocytes from term pregnancies?
Our results showed that neither the anatomical compartmentnorthe presence of labor altered the expression stability of the analyzedgenes. Further, we identified PGK1,GUSB and TBP as themost reliablereference genes and RN18S1 as the least suitable reference gene. Ourresults regarding PGK1, GUSB and TBP agree with previous studieswho reported these genes as stable in peripheral blood leukocytes[12e17]. In contrast, reports concerning the stability of RNA18S1 arecontradictory. It has been found to be an adequate reference in leu-kocytes, reproductive tissues stimulated with mitogens andplacentae from IUGR, but not in endometrial samples from womenwith polycystic ovarian syndrome [13,16,18e20].
Nevertheless, we found that the stability measure values for allof the tested genes, including RNA18S1, were below the stabilitythreshold. Therefore, RNA18S1, GAPDH, ACTB and B2M, the genesmost commonly used as references, can be considered stable inleukocytes during term pregnancy, but would be less adequatereferences compared to other housekeeping genes such as PGK1,GUSB and TBP. Moreover, it is worthy to highlight that unlike anypreviously reported study, we evaluated housekeeping gene sta-bility in leukocytes from pregnant women, different anatomicalcompartments (peripheral blood, placenta, choriodecidua) andphysiological conditions (no labor and active labor), which repre-sents the main contribution of our study.
Finally, we tested how many reference genes are required foraccurate normalization since using a single reference gene or anunstable gene, rather than using two or more reference genes fornormalizing expression, will yield quite different results [9e11].Our results show that only two of the best ranking reference genesare needed for normalization in expression studies of leukocytesfrom term pregnancies. However, the inclusion of a third referencegene would provide an even more accurate normalization.
5. Conclusion
PGK1, GUSB and TBP are the most adequate reference genes forgene expression normalization in leukocytes from termpregnancies,regardless of their anatomical origin (maternal peripheral blood,placental bloodorchoriodecidua) or thepresenceorabsenceof labor.
Conflict of interest statement
Authors declare not having any financial, personal or commer-cial conflict of interest.
Acknowledgments
This study was supported by grant SALUD-2010-01-141144from CONACYT, Mexico. N.G-L is funded by Wayne State Uni-versity Perinatal Initiative in Maternal, Perinatal and ChildHealth, Eunice Kennedy Shriver National Institute of Child Healthand Human Development of the National Institutes of Health(NICHD/NIH).
We thank Karla MacDonald, M.Sc. for proof reading ourmanuscript.
References
[1] Norman JE, Bollapragada S, Yuan M, Nelson SM. Inflammatory pathways in themechanism of parturition. BMC Pregnancy Childbirth 2007;1(7 Suppl.):S7.
[2] Gomez-Lopez N, Guilbert LJ, Olson DM. Invasion of the leukocytes into thefetal-maternal interface during pregnancy. J Leukoc Biol 2010;88:625e33.
[3] Vega-Sanchez R, Gomez-Lopez N, Flores-Pliego A, Clemente-Galvan S,Estrada-Gutierrez G, Zentella-Dehesa A, et al. Placental blood leukocytes arefunctional and phenotypically different than peripheral leukocytes duringhuman labor. J Reprod Immunol 2010;84:100e10.
[4] Castillo-Castrejon M, Meraz-Cruz N, Gomez-Lopez N, Flores-Pliego A, Beltr�an-Montoya J, Viveros-Alcar�az M, et al. Choriodecidual cells from term humanpregnancies show distinctive functional properties related to the induction oflabor. Am J Reprod Immunol 2014;71:86e93.
[5] Aerts JL, Gonzales MI, Topalian SL. Selection of appropriate control genes toassess expression of tumor antigens using real-time RT-PCR. Biotechniques2004;36:84e6. 88,90e1.
[6] Craythorn RG, Girling JE, Hedger MP, Rogers PA, Winnall WR. An RNA spikingmethod demonstrates that 18S rRNA is regulated by progesterone in themouse uterus. Mol Hum Reprod 2009;15:757e61.
[7] Dheda K, Huggett JF, Bustin SA, Johnson MA, Rook G, Zumla A. Validation ofhousekeeping genes for normalizing RNA expression in real-time PCR. Bio-techniques 2004;37:112e4. 116,118e9.
[8] Suzuki T, Higgins PJ, Crawford DR. Control selection for RNA quantitation.Biotechniques 2000;29:332e7.
[9] Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al.Accurate normalization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genes. Genome Biol 2002;3.research0034.1eresearch0034.11.
[10] Rho H-W, Lee B-C, Choi E-S, Choi I-J, Lee Y-S, Goh S-H. Identification of validreference genes for gene expression studies of human stomach cancer byreverse transcription-qPCR. BMC Cancer 2010;10:240.
[11] Radonic A, Thulke S, Mackay IM, Landt O, Siegert W, Nitsche A. Guideline toreference gene selection for quantitative real-time PCR. Biochem Biophys ResCommun 2004;313:856e62.
[12] Ledderose C, Heyn J, Limbeck E, Kreth S. Selection of reliable reference genesfor quantitative real-time PCR in human T cells and neutrophils. BMC ResNotes 2011;4:427.
[13] LifeTechnologies. Using TaqMan® endogenous control assays to select anendogenous control for experimental studies. LifeTechnologies 2012.
[14] Falkenberg VR, Whistler T, Murray JR, Unger ER, Rajeevan MS. Identification ofphosphoglycerate kinase 1 (PGK1) as a reference gene for quantitative geneexpression measurements in human blood RNA. BMC Res Notes 2011;4:324.
[15] Joiner MC, Thomas RA, Grever WE, Smolinski JM, Divine GW, Konski AA, et al.Developing point of care and high-throughput biological assays for deter-mining absorbed radiation dose. Radiother Oncol 2011;101:233e6.
[16] Lossos IS, Czerwinski DK, Wechser MA, Levy R. Optimization of quantitativereal-time RT-PCR parameters for the study of lymphoid malignancies17. UK:Leuk off J Leuk Soc Am Leuk Res Fund; 2003. p. 789e95.

R. Vega-Sanchez et al. / Placenta 36 (2015) 240e245 245
[17] Zampieri M, Ciccarone F, Guastafierro T, Bacalini MG, Calabrese R, Moreno-Villanueva M, et al. Validation of suitable internal control genes for expressionstudies in aging. Mech Ageing Dev 2010;131:89e95.
[18] Sadek KH, Cagampang FR, Bruce KD, Shreeve N, Macklon N, Cheong Y. Vari-ation in stability of housekeeping genes in endometrium of healthy andpolycystic ovarian syndrome women. Hum Reprod 2012;27:251e6.
[19] Arenas-HernandezM, Vega-Sanchez R. Housekeeping gene expression stabilityin reproductive tissues after mitogen stimulation. BMC Res Notes 2013;6:285.
[20] Murthi P, Fitzpatrick E, Borg AJ, Donath S, Brennecke SP, Kalionis BGAPDH. 18SrRNA and YWHAZ are suitable endogenous reference genes for relative geneexpression studies in placental tissues from human Idiopathic fetal growthrestriction. Placenta 2008;29:798e801.




















