
Feeding Experiences and Growth Status in a Rett Syndrome Population


I
INFORME FINAL DEL CONVENIO DE DONACIÓN Nº 70 ‐2009‐APECO‐CI
Evaluación del estado poblacional del huaucui y de las
comunidades vegetales que integra:
Bases para una estrategia de conservación en la
vertiente occidental peruana
Blanca León, Kenneth R. Young & José Roque G.
Resumen
Entre los endemismos más llamativos, por incorporar rareza en distribución con ambientes secos de la
vertiente occidental, está Orthopterygium huaucui (A. Gray) Hemsl. Este informe reseña las
observaciones y datos resaltantes del proyecto sobre las poblaciones y comunidades del huaucui que
permiten proponer temas para su conservación.
El informe recuenta la información de diez localidades visitadas en ocho cuencas en enero, febrero,
marzo, junio y diciembre 2009, setiembre 2010 y abril 2012. La exploración y verificación de las cuencas
permite señalar que el límite septentrional de esta especie sigue siendo la cuenca del Chillón. Las
poblaciones de esta especie varían en densidad a lo largo de la gradiente altitudinal, siendo ralas en los
límites inferiores y más abundantes y dominantes en altitudes intermedias. Se confirma que la especie
es una planta rara, pero localmente abundante. El límite altitudinal inferior de 800 m no se confirmó y
más bien la menor altitud corresponde a los 1200 m. La biología de Orthopterygium como planta dioica
hace difícil la observación ocasional de las poblaciones, excepto por los individuos femeninos que
retienen las infrutescencias por más de un año. La presencia de poblaciones de Orthopterygium
dominados por los extremos de tamaño, ya sea menores de 1m o mayores de 3m, la dominancia de
individuos con más de un tallo y el hábitat dinámico que ocupan en las laderas rocosas permite
proponer la hipótesis que Orthopterygium forma poblaciones clonales. Las semillas de Orthopterygium
son de rápida germinación, pero con alta mortalidad de las plántulas. Durante este estudio no se
registró la germinación de estas, excepto por una observación ocasional. Las respuestas fenológicas de
Orthopterygium ante asincronías ambientales podrían estar afectando su éxito reproductivo, por lo que
se sugiere ampliar las observaciones de estas condiciones. Este proyecto también ha servido para iniciar
la evaluación de la monofilia de Orthopterygium con el objeto de coadyuvar a entender los orígenes de
la flora de la vertiente occidental y responder si a través de su composición genética se puede incluir
paleoendemismos en la discusión de conservación de la flora. Una estrategia de conservación para esta
especie y la vertiente occidental requiere de múltiples conexiones y en particular para el estudio de las
características biológicas de especies en ambientes estacionales un apoyo de mediano y largo plazo.

León, Young & Roque – Orthopterygium bases para su conservación
1
INFORME FINAL DEL CONVENIO DE DONACIÓN Nº 70 ‐2009‐APECO‐CI
Antecedentes
Las laderas andinas del Pacifico central son reconocidas por la presencia de un complejo de
ambientes áridos y semiáridos, pero con escasa representación en el sistema oficial de áreas
protegidas. La flora de estos ambientes es un tema de interés para interpretar el origen de la flora
peruana; así Weberbauer (1945:627, 628) postuló que sus integrantes representaban vínculos
históricos a la del hemisferio norte o a México. Esta flora, además es interesante por incluir una
representación alta de endemismos (21% de la flora de las vertientes occidentales) y linajes de
probable evolución reciente (e.g. Malesherbia, Solanum sección Regmandra)
La especie, Orthopterygium huaucui (A. Gray) Hemsl. “huacui” o “huancoy”, objeto de este
proyecto fue seleccionada por tener rangos de distribución latitudinal y altitudinal limitados al
centro de la vertiente occidental del Perú. Su estudio en este proyecto formó parte de la iniciativa
que estudia las características de la flora y vegetación actual de la vertiente occidental, con el
objeto de reconstruir y entender los procesos que han modelado la composición, estructura y
dinámica de las poblaciones silvestres de plantas. Con estas bases consideramos se coadyuvará a
los esfuerzos de conservación de la biota peruana y se sustentará el integrar estos ambientes y su
flora a dichos esfuerzos.
Orthopterygium huaucui por ser un endemismo peruano fue categorizada por León &
Monsalve (2007) como “En Peligro”, sobre la base de sus poblaciones fragmentadas ubicadas en
un estrecho rango altitudinal y los riesgos a sus poblaciones provenientes de la expansión urbana y
varias actividades extractivas. El “huaucui” ocupa principalmente laderas rocosas, el mismo tipo de
hábitat de otras especies consideradas amenazadas, con las cuales forma en muchos casos
comunidades mixtas. Este proyecto propuso conocer: ¿es la estructura poblacional diferente en el
centro del rango de distribución que en los límites?, ¿cómo se distribuyen espacialmente los sexos
y cómo afecta este patrón al suceso reproductivo?, ¿es el huaucui una especie dominante? y si no
cómo se caracterizan las especies amenazadas en términos de dominancia.
Aspectos taxonómicos y biológicos del huaucui
Orthopterygium huaucui es una planta leñosa, ya sea árbol bajo o arbusto, dioico, restringido en
distribución a la vertiente occidental andina del Perú y miembro de la familia Anacardiaceae.
La historia taxonómica de Orthopterygium huaucui se inicia con su descripción en el
género Juliania por Asa Gray, sobre la base de una colección realizada en la cuenca del río Chillón
por la expedición Wilkes en 1854; el ejemplar depositado en el herbario de la Smithsonian consiste
de una rama sin hojas, la cual lleva solamente estructuras péndulas que fueron interpretadas
como frutos samaroides. Al parecer, Gray no tuvo acceso a una colección más temprana y
completa realizada por Andrew Mathews en abril de 1831 y depositada en los herbarios de Paris y
Kew (Hemsley 1907)

León, Young & Roque – Orthopterygium bases para su conservación
2
La casi ausencia de información morfológica contribuyó a las dificultades en establecer sus
afinidades evolutivas. Hemsley (1907) revisó mayor cantidad de material botánico permitiéndole
reconocer y proponer Orthopterygium para referirse al “huaucui”. Hemsley (op. cit) presentó
también una descripción detallada de las características de las plantas femeninas y masculinas, así
como una ilustración de una rama con hojas (Fig. 1). Tanto Orthopterygium como el género
mexicano‐guatemalteco Amphipterygium fueron ubicados por Hemsley (op. cit) en la familia
Julianiaceae sugiriendo vínculos evolutivos ya sea con las Juglandaceae o Anacardiaceae. Gracias a
trabajos morfo‐anatómicos y fitoquímicos posteriores (Young 1976) se resolvió que los vínculos de
las Julianiaceae se hallan con las Anacardiaceae, propuesta posteriormente apoyada por datos
moleculares (Pell 2004).
Figura 1. Ilustración de Orthopterygium huaucui (A. Gray) Hemsl.

León, Young & Roque – Orthopterygium bases para su conservación
3
La mayoría de los ejemplares de herbario de Orthopterygium huaucui están sin hojas, son
estériles o presentan plantas femeninas con los denominados frutos (Figura 2 A‐C). Sobre la base
de estos ejemplares se sabe entonces de la estacionalidad en la formación tanto de hojas como de
las inflorescencias.
Figura 2. Ejemplares de herbario (USM), tres de ellos (A‐C) femeninos y con “frutos”. La muestra D
es estéril.
Las hojas se inician al inicio de la estación de lluvias a fines de diciembre o enero, aunque
estos meses varían dependiendo de la disponibilidad de las lluvias. Las hojas son caducas y se
pierden en marzo, raras veces en abril. Weberbauer (1945) comentó que el desarrollo de las
inflorescencias ocurre al inicio de la estación seca cuando las hojas ya son caducas; sin embargo

León, Young & Roque – Orthopterygium bases para su conservación
4
nuestras observaciones no apoyan completamente tal aseveración, pues las inflorescencias
desarrollan antes o durante el desarrollo foliar, similar a los síndromes observados por Bullock
(1994).
Los tallos de Orthopterygium son por lo general erguidos, con corteza de color gris a
negruzco (Figura 3 A), similar en color a la de otros miembros de la flora acompañante como
Jatropha macrantha Müll. Arg. y Vasconcellea candicans A.DC. La profundidad de los tallos y raíces
depende de las condiciones del suelo y ladera (Figuras 3 B‐E). Durante este trabajo observamos
que los tallos pueden, en algunos casos, ramificarse (Figura 3 E)
Figura 3. A. Tronco de Orthopterygium huaucui. B‐D. Raíces expuestas en ladera de aprox. 90° de
inclinación. E. Tallo rastrero con dos ramas opuestas que llevan hojas y raíces adventicias. F. Vista
interior mostrando un corte transversal de tallo.

León, Young & Roque – Orthopterygium bases para su conservación
5
Las inflorescencias masculinas de Orthopterygium fueron consideradas como estructuras
que desarrollan luego de la caída de las hojas (Weberbauer 1945:157). Hemsley (1908) las
describió como pequeñas y erectas. Sin embargo, estas inflorescencias desarrollan en cimas
racemosas y péndulas que alcanzan hasta 15 cm de largo (Fig. 4 D). Jiménez et al. (en prep)
compararon las dimensiones de las flores de Orthopterygium y Amphipterygium reconociendo
pocas diferencias para ellas. Las flores masculinas en Orthopterygium se agrupan de 4—5 en cimas
con una bráctea lateral; cada flor es actinomorfa, sin pétalos, ni nectarios, con 3—7 sépalos
alternos a los estambres, los estambres, de hasta 1,45 mm, sobrepasan en longitud a los sépalos
(Hemsley 1907 figura incluida aqui; Jiménez et al, en prep).
Las estructuras florales femeninas en Amphipterygium fueron examinadas por Bachelier &
Endress (2007) confirmando que la estructura considerada una flor en realidad corresponde a una
inflorescencia, la cual para ese género es una cima de cuatro flores envueltas por un involucro a
manera de cúpula, en la cual solamente una flor se desarrolla hasta producir una semilla; la
inflorescencia se sostiene en un pedúnculo que se expande y aplana conforme avanza el desarrollo
del fruto. Las inflorescencias femeninas de Orthopterygium son estructuras cuyo desarrollo es
poco conocido y que en este estudio se examinaron en mayor detalle (Figura 4 A‐C). Estas
estructuras pasan inicialmente desapercibidas como indicó Hemsley (1907), si bien tienen una
posición apical debido a su desarrollo que precede al de las hojas quedando luego axilares a estas.
Los pedúnculos aplanados son la parte vistosa por su color rojizo (Figura 4 B y C), con superficie
cubierta de pelos blanquecinos que se aplana y ancha con la madurez floral. Estos pedúnculos son
inicialmente erguidos y probablemente luego de la fertilización continúan elongándose y
aplanando, al mismo tiempo que se encorvan (Fig. 4 C) hasta terminar péndulos. En las
inflorescencias jóvenes, los tres estigmas blanco‐amarillentos son protuberantes desde un estilo
grueso y rojizo, bilobados y que sobresalen del involucro, este a manera de “cúpula” (Fig 4C). La
secuencia de desarrollo de los ejes florales (León et al., en prep.) semeja al descrito para
Amphipterygium.
Weberbauer (1945:157) comentó la probable anemofilia de estas flores. Bullock (1994)
caracterizó la morfología de algunas plantas cuya polinización ocurre predominantemente por
viento, entre estas características incluyó la poca vistosidad floral, ausencia de pétalos, disposición
de los estambres, ausencia de nectarios. Es posible que anemofilia esté presente en
Orthopterygium; sin embargo, dado el color rojizo de los sépalos y de los ejes femeninos de

León, Young & Roque – Orthopterygium bases para su conservación
6
Orthopterygium deja abierta la posibilidad de otro tipo de síndrome, requiriéndose mayores
observaciones.
Figura 4. A. Árbol femenino en la cuenca del rio Chillón (foto K.R.Young). B. Rama sin hojas con
inflorescencias femeninas en la cuenca del Rímac (foto K.R.Young). C. Rama con hojas e
inflorescencias femeninas axilares a las hojas. Obsérvese los ejes péndulos que llevan las
infrutescencias (foto J. Roque) D. Ramas con hojas e inflorescencias masculinas (foto K.R.Young).

León, Young & Roque – Orthopterygium bases para su conservación
7
El pariente más cercano de Orthopterygium es Amphipterygium, un género mayormente
mejicano con el cual comparte un fruto sincárpico sobre un pedúnculo alado que le da apariencia
de una sámara (Figura 5). El estudio de Pell (2004) mostró que el linaje que incluye a estas plantas
americanas está conformado por géneros que provienen del hemisferio norte (Toxicodendron,
Pistacia y Comocladia). Ella también mostró que las afinidades del grupo
Orthopterygium/Amphipterygium son a géneros de bosques secos como Loxopterygium y
Cardenasiodendron. Loxopterygium está presente en la flora peruana en los bosques del noroeste
y en los fragmentos en la vertiente occidental, mientras que Cardenasiodendron es un endemismo
boliviano. Estos hallazgos parecerían apoyar la idea mencionada por Weberbauer (1945:628)
sobre la presencia en los ambientes xéricos de elementos de procedencia “mejicana o andina”. Por
tanto, hay necesidad por conocer el desarrollo histórico‐evolutivo de Orthopterygium para
entender cuán antigua es la historia misma de los ambientes que ocupa. Los trabajos de Linares
(2006), Pennington et al. (2010) o los de Sarkinen et al. (2012) muestran que para los ambientes
xéricos interandinos su flora data desde el Mioceno y que los mismos han diversificado en núcleos
con endemismos restringidos. Los resultados iniciales del trabajo de tesis de Víctor Jiménez (en
prep.) parecen apoyar un patrón similar para Orthopterygium: antigüedad y probable vínculo al
hemisferio norte. Si estos datos se confirman entonces se tendrá un paleoendemismo verificado.
Figura 5. A la izquierda frutos sincárpicos de Orthopterygium huaucui, a la derecha de
Amphipterygium adstringens (Schltdl.) Schiede ex Standl., material procedente de Guerrero,
México
Rangos de distribución y poblaciones
El conocimiento de la distribución de muchas especies en el Perú, como en el caso de
Orthopterygium huaucui, está fundamentado en el trabajo pionero de Weberbauer (1945).
Weberbauer (op. cit.) citó el rango de distribución latitudinal entre los 11°y 15°30’ S, los que
corresponden a 16 cuencas en el Pacífico central. Los sitios con poblaciones de Orthopterygium,
sin embargo están en seis cuencas. Estos sitios están ubicados principalmente en los
León, Young & Roque – Orthopterygium bases para su conservación
8
departamentos de Lima e Ica (Tabla 1 y Figura 6) con poblaciones que se extienden a Huancavelica
o Ayacucho y al extremo norte de Arequipa (Figura 6). El rango altitudinal de Orthopterygium va
de los 800 a 2800 m, siendo su amplitud aparentemente mayor en el sur (Weberbauer 1945:158).
Sin embargo, las altitudes más bajas observadas en este proyecto están en los 1200 m. En los
sitios históricos del valle del Chillón el límite inferior está ocupado por paisajes transformados por
el ser humano.
Información de colecciones históricas junto a los mapas de caminos disponibles por el
Ministerio de Transporte y Comunicaciones
(http://www.proviasnac.gob.pe/red_vial/mapas/departamental.html), fueron empleadas para
reubicar las poblaciones. Se examinó asimismo vistas de los valles en GoogleEarth, medio que
provee con cierta resolución imágenes que en algunos casos permitió reconocer las poblaciones
en sitios ya conocidos y en otros reconocer áreas potenciales.
La localidad original del tipo, citada como Yangas (1030 m) se halla en la cuenca del Chillón.
Mientras que la localidad del ejemplar más antiguo corresponde a la de “Cuesta de Purruchuca”
también en la misma cuenca. Otra localidad histórica es la de Chosica, la cual fue herborizada por
Weberbauer en 1910. La Tabla 1 reseña las cuencas principales con la presencia de poblaciones de
Orthopterygium. La cuenca del Chillón cuenta con mayor número de sitios, en parte por su
accesibilidad, al mismo tiempo que estos sitios parecen representar una misma población.
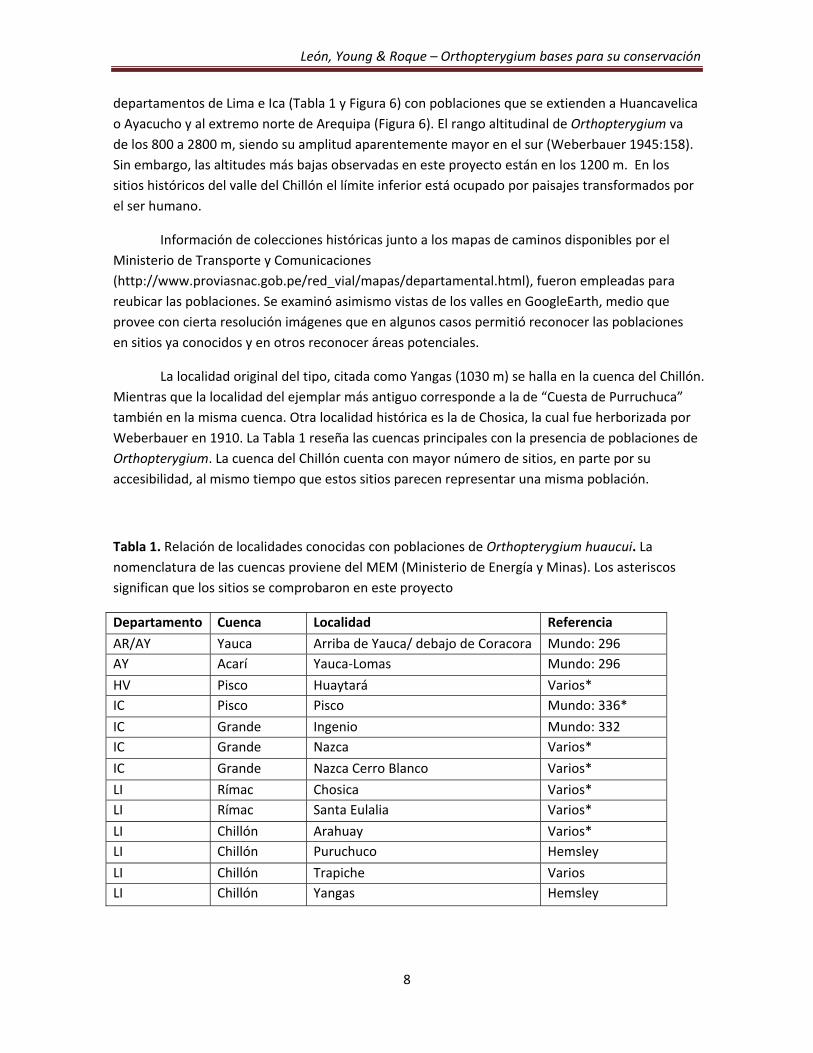
Tabla 1. Relación de localidades conocidas con poblaciones de Orthopterygium huaucui. La
nomenclatura de las cuencas proviene del MEM (Ministerio de Energía y Minas). Los asteriscos
significan que los sitios se comprobaron en este proyecto
Departamento Cuenca Localidad Referencia
AR/AY Yauca Arriba de Yauca/ debajo de Coracora Mundo: 296
AY Acarí Yauca‐Lomas Mundo: 296
HV Pisco Huaytará Varios*
IC Pisco Pisco Mundo: 336*
IC Grande Ingenio Mundo: 332
IC Grande Nazca Varios*
IC Grande Nazca Cerro Blanco Varios*
LI Rímac Chosica Varios*
LI Rímac Santa Eulalia Varios*
LI Chillón Arahuay Varios*
LI Chillón Puruchuco Hemsley
LI Chillón Trapiche Varios
LI Chillón Yangas Hemsley

León, Young & Roque – Orthopterygium bases para su conservación
9
Figura 6. Localidades conocidas de Orthopterygium huaucui sobre la base de colecciones botánicas
de este proyecto (Res. Direct. No. 012‐2009‐AG‐DGFF‐DGEFFS) y de otros (puntos morados) y
franjas azules representando los ocho valles visitados durante este proyecto, obsérvese los
rectángulos sin puntos que corresponden a valles sin aparente presencia del “huaucui”
Durante este proyecto se visitó diez localidades, de norte a sur: Huaral, Chillón, Santa
Eulalia, Jicamarca, Rímac, Lurín, Mala, Omas, Pisco y Nazca; estas dos últimas a cargo de Víctor
Jiménez para su trabajo de tesis. Estos sitios están en ocho cuencas (Figura 6), de los cuales cuatro
cuencas no habían sido previamente examinadas. No se hallaron poblaciones en Huaral, parte baja

León, Young & Roque – Orthopterygium bases para su conservación
10
(1200 m) de Jicamarca, ni en Lurín, Mala, tampoco en Omas. La ausencia en algunos casos
corresponde a la inaccesibilidad de los sitios, por tanto se requiere combinar mejores imágenes
con otras formas de acceso para verificación. En sitios como la vertiente sur del Huaral las
imágenes satelitales parecían mostrar poblaciones del Orthopterygium, pero la verificación de
campo reveló en cambio poblaciones dominadas por Vasconcellea y Armatocereus. Esa parta del
valle del Chancay‐Huaral al no incluir poblaciones del “huaucui” deja como límite septentrional de
esta especie a la cuenca del Chillón (ver Figura 7).
Se puede emplear estas observaciones para plantear los sitios a examinar a futuro que
permitan conocer los límites latitudinales y las condiciones ambientales que puedan contribuir al
patrón conocido. Además, queda sin explicación la razón para las distribuciones disyuntas.
Figura 7. Mapa de las cuencas del Pacífico central (basado en el Mapa de Cuencas Hidrográficas
del Ministerio de Energía y Minas) relevantes para la distribución del Orthopterygium huaucui: en
amarillo entero cuencas con presencia de esta especie (1025 Chillón; 1026 Rímac; 1034 Pisco;
1036 Grande; 1037 Acarí y 1038 Yauca); cuencas con rayas azules oblicuas indican sin presencia de
huaucui (1021 Pativilca; 1022 Supe; 1023 Huaura; 1024 Chancay‐Huaral; 1027 Lurín; 1028 Chilca,

León, Young & Roque – Orthopterygium bases para su conservación
11
1029 Mala, 1030 Omas; 1031 Cañete; 1032 Topara; 1033 San Juan; 1035 Ica; 1039 Chala; 1040
Chaparra; 1041 Atico)
Observaciones de las poblaciones del “huaucui”
Para conocer el tamaño y características de las poblaciones se establecieron al azar seis parcelas
temporales de 50X20 m (Figura 8) con la línea mayor sobre la pendiente. En las parcelas se
registró tanto la flora acompañante como cada individuo de Orthopterygium. Para cada planta de
“huaucui” se midió el alto, diámetro basal y a la altura del pecho, número de ramas y estado
fenológico.
Figura 8. Parcela en el camino a Arahuay, a 1250 m. Vista superior izquierda muestra la base de la
parcela sobre un tronco de Orthopterygium.

León, Young & Roque – Orthopterygium bases para su conservación
12
Weberbauer (1945:158) comentó sobre Orthopterygium que “esta planta es muy
frecuente por lo general y aún puede formar pequeños montes”. Estas observaciones son vigentes,
pudiéndose definirla como una especie rara localmente abundante y en la que sus poblaciones
marginales en altitud tienen individuos dispersos (Figura 9A), mientras que en el centro de su
distribución altitudinal son densos hasta constituir los montes a los que se refirió Weberbauer
(Figura 9 B). Lesica et al. (2006), examinando especies cercanamente relacionadas, propusieron
que especies con rangos de distribución reducidos pero localmente abundantes son una expresión
de especiación reciente. Para el caso de Orthopterygium este no parece ser el caso, dada su
probable emergencia Miocénica y su probable estatus de paleoendémica.
En 1000m2 se registró entre 3 y 76 individuos, contándose un total de 256 individuos en
seis diferentes parcelas a distintas altitudes. Las diferencias encontradas pueden asociarse a la
altitud, pero se requiere mayor información para confirmarlas. Las dos localidades más
depauperadas (debajo de Arahuay, cuenca del Chillón) corresponden al límite altitudinal inferior
de esta especie, ambas con condiciones ambientales más secas. En ambas se halló solo individuos
altos, dispersos y nula presencia de juveniles (Tabla 2), aunque sí presencia de infrutescencias en
el suelo y entre rocas (Figura 10). Sin embargo, en la población de Santa Eulalia, de la misma
cuenca y en una altitud intermedia del rango conocido, muestra dos picos de tamaños, uno para
individuos bajos igual o menores a 1,5 m y el otro para árboles con tamaños superiores a los 3 m
(Figura 9 B, Tabla 2), lo que podría indicar reproducción de juvenil a adulto.
Figura 9. Comunidades de Orthopterygium. A. Individuo aislado en comunidad dispersa en Pisco,
las flechas indican presencia de otros individuos en la ladera. B. Monte en Santa Eulalia con
individuos de hasta 5 m
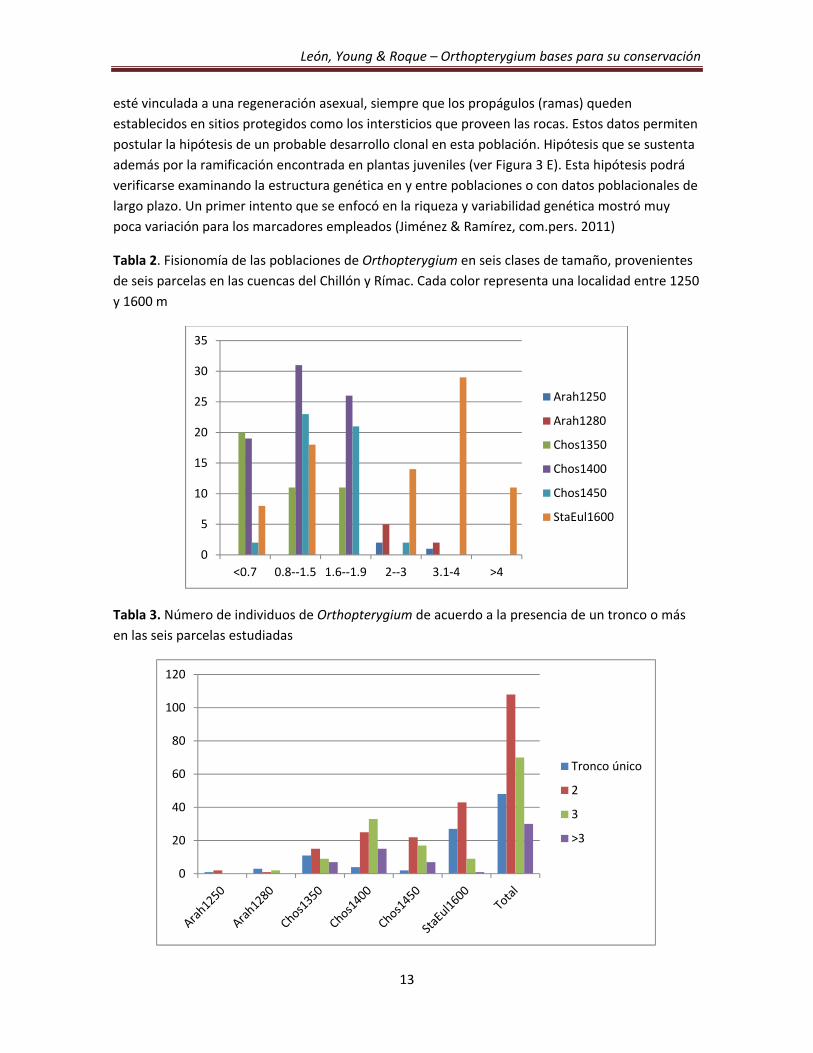
Las plantas de Orthopterygium crecen formando uno o más troncos, pero un tronco único
se halló solamente en 18,75% de todas las plantas observadas en este proyecto (Tabla 3), mientras
que la mayoría de los individuos son ramificados y con un 42% del total con dos tallos basales. Al
comparar las características en la Tabla 2 con los datos de la Tabla 3 se encuentra que la parcela
con el mayor número de individuos con más de un tronco es la población de Santa Eulalia. En
lugares tan dinámicos como las laderas andinas, la probable respuesta de las plantas más grandes
León, Young & Roque – Orthopterygium bases para su conservación
13
esté vinculada a una regeneración asexual, siempre que los propágulos (ramas) queden
establecidos en sitios protegidos como los intersticios que proveen las rocas. Estos datos permiten
postular la hipótesis de un probable desarrollo clonal en esta población. Hipótesis que se sustenta
además por la ramificación encontrada en plantas juveniles (ver Figura 3 E). Esta hipótesis podrá
verificarse examinando la estructura genética en y entre poblaciones o con datos poblacionales de
largo plazo. Un primer intento que se enfocó en la riqueza y variabilidad genética mostró muy
poca variación para los marcadores empleados (Jiménez & Ramírez, com.pers. 2011)
Tabla 2. Fisionomía de las poblaciones de Orthopterygium en seis clases de tamaño, provenientes
de seis parcelas en las cuencas del Chillón y Rímac. Cada color representa una localidad entre 1250
y 1600 m
Tabla 3. Número de individuos de Orthopterygium de acuerdo a la presencia de un tronco o más
en las seis parcelas estudiadas
0
5
10
15
20
25
30
35
<0.7 0.8‐‐1.5 1.6‐‐1.9 2‐‐3 3.1‐4 >4
Arah1250
Arah1280
Chos1350
Chos1400
Chos1450
StaEul1600
0
20
40
60
80
100
120
Tronco único
2
3
>3

León, Young & Roque – Orthopterygium bases para su conservación
14
La estacionalidad en la floración de Orthopterygium crea dificultades en reconocer los
sexos de esta planta dioica, especialmente de las plantas juveniles y de las masculinas. El
desarrollo floral y vegetativo en Orthopterygium huaucui responde a la precipitación, en forma
similar a la de la flora acompañante. Eventos “irregulares” de precipitación generaron floración de
plantas masculinas al menos en una de las poblaciones observadas por nosotros en el 2012. Esta
asincronía en la floración debido a condiciones ambientales requiere estudiarse para entender si
existe un estrés fisiológico a las plantas y cómo queda afectado el éxito reproductivo. Trabajos
recientes por Bertin (2008) y Mazer et al. (2013) identifican la influencia tanto ambiental como
genética sobre los aspectos fenológicos. El trabajo de Mazer et al. (2013) para el hemisferio norte
podría servir como referencia base para plantear estudios más detallados y continuos sobre
fenología en ambientes estacionales, secos y tropicales, en los cuales precipitación parece jugar un
papel importante en la fenología, mientras que temperatura juega un rol en el éxito de
establecimiento de las plántulas.
El reconocimiento de plantas femeninas en Tabla 4 incluye a aquellos individuos con frutos
colgantes. En Orthopterygium las infrutescencias quedan adheridas a las ramas por más de un año,
cuando maduran tornan los ejes de rojo a pajizo (ver Figura 10). Plantas dioicas frecuentemente se
presentan en relación 1:1 (por ejem. Morellato 2004) y en este proyecto observaciones en las
parcelas de Chosica muestran similar tendencia. El número alto de individuos con estadio
desconocido apoya sugerir observaciones continuas que aclaren la fenología de estas poblaciones.
Tabla 4. Número de individuos de Orthopterygium de acuerdo a su estadio floral (femenino,
masculino y desconocido) en cinco parcelas
Las comunidades conocidas de Orthopterygium ocupan ya sea suelos arenosos o en
laderas rocosas (León et al. 1997), en los primeros los individuos se presentan en forma dispersa
(Fig. 9 A), mientras que en laderas rocosas tiende a incrementarse el número de individuos. Estas
condiciones deben incorporarse para examinar el éxito de establecimiento de nuevas plantas. Las
infrutescencias de Orthopterygium por lo general llevan un solo embrión viable y estas
0
10
20
30
40
50
60
70
80
90
100
Arah1250 Arah1280 Chos1350 Chos1400 Chos1450 Total
Masc Fem desc

León, Young & Roque – Orthopterygium bases para su conservación
15
aparentemente no se dispersan distancias grandes, especialmente en casos donde la planta madre
ocupa laderas rocosas poco pronunciadas (menos de 20°).
Las semillas a su madurez son pajizas a café claro, con la cúpula gruesa y leñosa (Fig. 10 A‐
D). Como señalaron León et al. (1997) la germinación es iniciada cuando el agua alcanza a saturar
el tejido de la cúpula y llega al embrión. Las observaciones de León et al. (1997) en cuanto al
tiempo breve de germinación luego del tratamiento con agua fueron confirmadas por otros (S.
Palacios com.pers. 2011 y Jiménez com. pers. 2012). Los cotiledones emergen del ápice de la
cúpula, por donde persisten los restos de la flor (Fig. 10 D). En ninguna de las parcelas estudiadas
en este proyecto observamos plántulas o semillas germinando. La vista de la Figura 10 D
corresponde a un registro ocasional en una población aislada de la cuenca del Pisco.
Figura 10. A‐C. Infrutescencias sin germinar. A y C muestran semillas cerca de la planta madre
entre restos de la flora acompañante. B. Semillas mostrando variabilidad en tamaño y desarrollo
de la cúpula; la punta del lapicero muestra una semilla con agujero en base de la cúpula por
insecto; la barra representa 1 cm D. Germinación in situ, semilla observada en Pisco

León, Young & Roque – Orthopterygium bases para su conservación
16
La viabilidad de las semillas es un tema de interés asociado a la dinámica de las
comunidades de Orthopterygium. Palacios (com. pers.2011) notó que en 100 semillas tratadas con
escarificación, la germinación superó el 80%, con casos aislados de germinación de dos a tres
embriones por semilla. Como ya observó León et al. (1997) el crecimiento de estas plantas es muy
rápido, aunque las tasas de supervivencia son muy bajas. Poco se sabe el porqué de esta condición.
Durante este estudio se evaluaron seis semillas colectadas en 1996 en Nazca, de las cuales
solamente una desarrolló el embrión sin lograr emerger de la cúpula. Para el caso de
Amphipterygium adstringens en Mexico, la especie hermana del huaucui, las semillas incrementan
su éxito de germinación luego de un año de su cosecha, pero luego “el vigor de la semilla
disminuye paulatinamente” (Cervantes Sanchez & Sotelo Boyas 2002).
Comunidades vegetales asociadas
Las comunidades vegetales con presencia del huaucui incluyen una flora vascular rica en cactus y
arbustos deciduos, junto a otras plantas bulbíferas y herbáceas efímeras (Figura 11 A‐I). Junto a
Orthopterygium se encuentran otros endemismos: Armatocereus matucanensis Back. ex A.W. Hill,
Cnidoscolus peruvianus (Fig. 11 B), Cleistocactus spp., Haageocereus spp., Neoraimondia
arequipensis s.lat. , entre otras. A estas plantas se adicionan las presentes en la flora ecuatoriana o
boliviana en ambientes secos como Astrolepis sinuata (Fig. 11 D), Solanum cheesemannii (Fig. 11 F),
Vasconcellea candicans.
En las localidades que corresponden al límite altitudinal inferior de Orthopterygium, las
especies dominantes son Neoraimondia, Haageocereus o Cnidoscolus. Por encima de los 1400 m
hasta los 1800 m las poblaciones de Orthopterygium son extensas y es en este rango que esta
especie es dominante. Las comunidades vegetales con Orthopterygium pueden describirse como
ambientes de cactáceas columnares, a matorrales dominados por huaucui.
León et al. (1997) citó 19 especies asociadas o vecinas a las poblaciones de
Orthopterygium en Cerro Blanco ubicado a 1250 m, sitio que representa el límite altitudinal
inferior de esta especie. Se requiere completar la composición de la flora de la vertiente occidental,
pues como se mencionó antes muchas de las especies son caducifolias o efímeras y por tanto será
importante un registro de mediano plazo a lo largo de por lo menos dos estaciones para capturar
la información necesaria.
El rol de plantas nodrizas es un tema importante en estos ambientes que requiere
examinarse. La presencia de plantas ocupando el mismo sitio que plantas de Orthopterygium es
evidente principalmente en los individuos más desarrollados de estos últimos (ejemplo ver Figura
11C). Estas asociaciones son variables, así en el límite altitudinal inferior del huaucui, el probable
rol de cactáceas sea de facilitador para su establecimiento, especialmente cuando plantas como
Armatocereus y Neoraimondia parecen superar los 75 años de edad. Pero, para el caso de
comunidades dominadas por Orthopterygium el rol de nodriza podría ser de este para arbustos y
hierbas (Fig. 11H).

León, Young & Roque – Orthopterygium bases para su conservación
17
Figura 11. A. Espostoa melanostele (Vaup.) Borg y Haageocereus acranthus (Vaupel) Backeb. B.
Cnidoscolus cf. peruvianus (Müll. Arg.) J.F. Macbr. C. Neoraimondia arequipensis entre ramas de
Orthopterygium D. Astrolepis sinuata (Lag. ex Sw.) D.M. Benham & Windham subsp. sinuata al pie
de Espostoa E. Ladera con Orthopterygium y Furcraea cf. occidentalis Trel. F. Solanum cf.
cheesemanni (L. Riley) Fosberg G. Melocactus peruvianus Vaupel H. Ladera dominada por arbustos
de Orthopterygium, arbusto a la derecha es Ophryosporus cf. peruvianus (J.F. Gmel.) R.M. King & H.
Rob. I. Clinanthus sp.

León, Young & Roque – Orthopterygium bases para su conservación
18
Observaciones sobre amenazas y estado de conservación
En localidades con poco impacto humano y grandes poblaciones de Orthopterygium huaucui no
generan, por ahora, preocupaciones para la especie. Las poblaciones parecen estar muy bien
adaptadas a las condiciones ambientales, y es incluso abundante en algunos valles. Pero como se
mencionó antes queda por resolver el impacto de las asincronías ambientales con el éxito de
reclutamiento y establecimiento de plántulas. Además, observamos poblaciones más tenues en los
límites altitudinales, y que con agricultura o líneas de transmisión eléctrica o asentamientos
humanos simplemente ya no hay presencia de esta especie. No hay ninguna área protegida que la
incluye, así que su conservación activa es inexistente.
Mostramos que Orthopterygium huaucui es probablemente una especie paleoendémica,
que se mantiene frecuentemente como clon, con poca reproducción sexual y poca variación
genética. Confirmamos una distribución limitada, disyunta y dividida en tres partes. Expansión del
impacto humano en estos valles sería un riesgo grande para la especie. Además, los cambios
globales por un lado podrían afectar el reclutamiento de nuevos individuos (germinación,
crecimiento y establecimiento), y por otro podrían estar influyendo la necesidad de la especie en
subir en altitud, lo cual se dificulta por su poca reproducción y la intensidad de uso de suelo para
agricultura con riego en estos valles.
Otros estudios han documentado preocupaciones asociados con la línea de gas en Ica
(Perú LNG & Domus 2007), lo cual ha motivado esfuerzos para monitorear las poblaciones antes y
después de los cambios causados por la construcción de infraestructura energética. Nosotros
sugerimos acciones más activas que podrían incluir la reforestación de ciertas áreas con
Orthopterygium huaucui. Compartimos el deseo del proyecto de Kew en Ica (Whalen et al 2010)
de hacer restauración ecológica en los paisaje de las vertientes occidentales en el Perú.
Conclusiones y bases para una estrategia de conservación en la vertiente occidental
peruana
Los planes de conservación en la cuenca del Pacífico requieren con urgencia de datos verificables
sobre cambios en paisajes y riesgos ambientales. Las comunidades vegetales visitadas durante
este proyecto se ubican en áreas con alta tasa de cambio en uso del suelo ya sea por expansión
urbana o agrícola o de infraestructura, así como por extremos ambientales en temperatura e
irregularidades en la precipitación.
Weberbauer (1945) comentó la dificultad en distinguir los “elementos boreales de los
mejicanos” para los niveles medios del lado occidental de los Andes. La interpretación del origen y
patrones de distribución de los integrantes de una flora es importante para discutir objetivos de
investigación asociados a conservación (ver Sosa & De Nova 2012). Por ello, este proyecto incluyó
iniciar el examen del aparente vínculo que existe entre este género peruano y los taxones del
hemisferio norte empleando técnicas moleculares para conocer si Orthopterygium representa un
linaje monofilético o no, y que permita esclarecer la historia de migraciones y los vínculos
florísticos de la vertiente del Pacífico. Al mismo tiempo y en términos de conservación de la flora,

León, Young & Roque – Orthopterygium bases para su conservación
19
se planteó impulsar el estudio de los niveles intra e inter poblacionales de este endemismo, tanto
con estudios de biología reproductiva como los de análisis genético.
Las bases de una estrategia de conservación para Orthopterygium y en la vertiente
occidental central requieren del impulso y colaboración institucional para establecer vínculos
entre los estudios y monitoreo de plazos largos con los esfuerzos locales o regionales de
restauración o que involucren a las comunidades humanas. El aclarar las tasas de suceso
reproductivo coadyuvará a los esfuerzos de quienes consideran planes de propagación de
plántulas en viveros y su introducción a las localidades degradadas. La investigación de la ecología
y biología de Orthopterygium huaucui, combinada con programas activos de restauración es
nuestra sugerencia para los próximos pasos a seguir.
Agradecimientos
Nuestro reconocimiento a Asunción Cano, Miguel Chocce, Aniceto Daza, Paul Gonzales, Víctor
Jiménez, Sonia Palacios, Rina Ramírez, Diana Silva, Nanette Vega y a todos los que nos apoyaron
de diversa manera en este proyecto.

León, Young & Roque – Orthopterygium bases para su conservación
20
Referencias citadas
Bachelier, J. B. & P. K. Endress. 2007. Development of inflorescences, cupules, and flowers of
Amphipterygium and comparisons with Pistacia (Anacardiaceae). Int. J. Plant Sci. 168: 1237—
1253.
Bertin, R.I. 2008. Plant phenology and distribution in relation to recent climate change. J. Torrey
Bot. Club 135:126—146.
Bullock, S. H. 1994. Wind pollination of Neotropical dioecious trees. Biotropica 26:172—179.
Cervantes Sánchez, M.A. & Sotelo Boyas, M.E. 2002. Guias tecnicas para la propagacion sexual de
10 especies latifoliadas de selva baja caducifolia en el Estado de Morelos.31 pp. Secretaria de
Agricultura, Ganadería y Desarrollo Rural, Pesca y Alimentación. Instituto Nacional de
Investigaciones Forestales, Agricolas y Pecuarias. Centro de Investigación Regional del Centro.
Campo Experimental Zacatepec, Zacatepec, Morelos, México.
Perú LNG S.R.L. & DOMUS, 2007. Ecological Field Survey for the Peru LNG natural gas pipeline
project. 375 pp. Lima. Perú.
Hemsley, W. B. 1908. On the Julianiaceae: a new natural order of plants. Phil. Trans. Roy. Soc. Lond,
Ser. B. 199:169‐197.
Jiménez, V., B. León & R. Ramirez. Morfología de las flores estaminadas de Orthopterygium
huaucui (A. Gray) Hemsley, Anacardiaceae. Sometido a Rev. Peruana Biol.
León, B. & C. Monsalve 2007. Anacardiaceae endémicas del Perú. En B. León, J. Roque, C. Ulloa
Ulloa, P.M. Jørgensen, N. Pitman & A. Cano (Eds.). Libro Rojo de las Plantas endémicas del
Perú. Revista Peruana de Biología, Edición Especial 13(2): 971 pp. [diciembre 2006]
León, B. , K.R. Young, A. Cano, M.I. La Torre, M. Arakaki & J. Roque 1997.Botanical exploration and
conservation in Peru: the plants of Cerro Blanco, Nazca. BioLlania, Ed. Especial 6:431—448.
Lesica, P., R. Yurkewyckz & E. E. Crone 2006. Rare plants are common where find them. Amer. J.
Bot. 93:454‐‐459.
Mazer , S.J., S.E. Travers, B.I. Cook, T.J. Davie, K. Bolmgren, N.J.B. Kraft, N. Salamin & D.W. Inouye.
2013. Flowering date of taxonomic families predicts phonological sensistivity to temperature:
Implications for forecasting the effects of climate change on unstudied taxa. Amer. J. Bot. 100:
10.3732/ajb.1200455.
Morellato, L.P.C. 2004. Phenology, sex ratio, and spatial distribution among dioecious species of
Trichilia (Meliaceae). Pl. Biol. 6:491—497.
Pell, S. K. 2004. Molecular systematics of the cashew family (Anacardiaceae). Ph.D. dissertation.
Baton Rouge: Louisiana State University.

León, Young & Roque – Orthopterygium bases para su conservación
21
Sosa, V. & J.A. De Nova 2012. Endemic angiosperm lineages in Mexico: Hotspots for Conservation.
Acta Bot. Mexicana 100:293—315.
Weberbauer, A. 1945. El Mundo Vegetal de los Andes peruanos. Estudio fitogeográfico. 776pp.
Estación Experimental de La Molina, Dirección de Agricultura. Ministerio de Agricultura. Lima,
Peru.
Whalen, O. Q., D. Beresford‐Jones, W. Milliken, A. Orellana, A. Smyk & J. Leguia. 2010. An
ecosystem approach to restoration and sustainable management on dry forest in southern
Peru. Kew Bull. 65:613—641.
Young, D. A. 1976. Flavonoid chemistry and the phylogenetic relationships of the Julianiaceae. Syst.
Bot. 1:149—162.
Copyright © 2022 FDOKUMEN




















