
Establishment, survival, and herbage production of novel, summer-active perennial pasture legumes in...
15
Establishment, survival, and herbage production of novel, summer-active perennial pasture legumes in the low-rainfall cropping zone of Western Australia as affected by plant density and cutting frequency Lalith D. B. Suriyagoda A,B,C , Daniel Real A,B,D , Michael Renton A,B,E , Hans Lambers A , and Megan H. Ryan A,B,F A School of Plant Biology and Institute of Agriculture, The University of Western Australia, 35 Stirling Highway, Crawley WA 6009, Australia. B Future Farm Industries Cooperative Research Centre, The University of Western Australia, Australia. C Present address: Faculty of Agriculture, University of Peradeniya, Peradeniya, 20400, Sri Lanka. D Department of Agriculture and Food, 3 Baron-Hay Court, South Perth, WA 6151, Australia. E CSIRO Ecosystem Sciences, Private Bag 5, Wembley, WA 6913, Australia. F Corresponding author. Email: [email protected] Abstract. Herbaceous perennial legumes that can provide forage in the summer–autumn dry period are urgently required in Mediterranean climates to complement annual pastures and the perennial legume lucerne (Medicago sativa). This study evaluated the establishment, survival, and herbage production of tedera (Bituminaria bituminosa var. albomarginata) and Cullen spp. native to Australia. Two experiments were replicated at Buntine (warmer site) and Newdegate (cooler site) in the low-rainfall cropping zone (<350 mm average annual rainfall) of Western Australia from June 2008 to September 2010. In the first experiment, established by transplanting seedlings, survival and herbage production of two accessions each of B. bituminosa and C. australasicum were studied under densities of 1, 2, 4, 8, and 16 plants/m 2 with 0, 1, 2, or 3 cuts in summer–autumn in addition to a winter–spring cut. In the second experiment, established from seed, emergence and survival of several accessions of B. bituminosa, C. australasicum, and M. sativa were studied, along with C. pallidum and C. cinereum. In the first experiment, B. bituminosa survived better than C. australasicum (70–80% v. 18–45%), especially at Buntine, but there was little impact of density or cutting frequency on survival. Plant death was highest during summer. Shoot dry weight (DW) accumulation varied greatly with site, year, and plant density. When rainfall was close to average, shoot DW was greater at Newdegate (B. bituminosa 7.4 t/ha, C. australasicum 4.5 t/ha) than at Buntine (2.3 t/ha), and both species produced much of their shoot DW in summer–autumn (e.g. 6 t/ha for B. bituminosa and 3 t/ha for C. australasicum at Newdegate). An early-summer cut reduced the DW that could be harvested later in summer–autumn. In the second experiment, emergence of B. bituminosa was either similar to, or higher than, emergence of the other species, being 43% at Buntine and 44% at Newdegate. Survival of B. bituminosa, compared with M. sativa, was similar at Buntine (13%) and slightly lower at Newdegate (14%). Emergence and survival of Cullen spp. varied among species and accessions, with survival of the best performing accession of C. australasicum (SA4966) similar to that of B. bituminosa and M. sativa at both sites. We conclude that B. bituminosa shows promise as a perennial summer forage for low-rainfall zones, with a density of 8–16 plants/m 2 and cutting frequency of 3 cuts/year (i.e. cut twice in summer–autumn), while C. australasicum and C. pallidum warrant further study. Additional keywords: Australian native legumes, Bituminaria bituminosa var. albomarginata, Cullen spp., drought- tolerant perennial pastures, summer–autumn feed gap, survival, tedera. Received 26 November 2012, accepted 15 April 2013, published online 3 May 2013 Introduction Perennial pasture legumes play an important role in the management of dryland salinity and can provide high-quality green forage, especially during the summer–autumn feed gap when annual forages have senesced (Cocks 2001; Bell et al. 2006b; Ward 2006). The only widely sown perennial legume in the low rainfall (<375 mm average annual rainfall) cropping zone of Western Australia is Medicago sativa L. (lucerne or Journal compilation ȑ CSIRO 2013 www.publish.csiro.au/journals/cp CSIRO PUBLISHING Crop & Pasture Science, 2013, 64, 71–85 http://dx.doi.org/10.1071/CP12398
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Establishment, survival, and herbage production of novel, summer-active perennial pasture legumes in...

Establishment, survival, and herbage production of novel,summer-active perennial pasture legumes in the low-rainfallcropping zone of Western Australia as affected by plantdensity and cutting frequency
Lalith D. B. SuriyagodaA,B,C, Daniel RealA,B,D, Michael RentonA,B,E, Hans LambersA,and Megan H. RyanA,B,F
ASchool of Plant Biology and Institute of Agriculture, The University of Western Australia,35 Stirling Highway, Crawley WA 6009, Australia.
BFuture Farm Industries Cooperative Research Centre, The University of Western Australia, Australia.CPresent address: Faculty of Agriculture, University of Peradeniya, Peradeniya, 20400, Sri Lanka.DDepartment of Agriculture and Food, 3 Baron-Hay Court, South Perth, WA 6151, Australia.ECSIRO Ecosystem Sciences, Private Bag 5, Wembley, WA 6913, Australia.FCorresponding author. Email: [email protected]
Abstract. Herbaceous perennial legumes that can provide forage in the summer–autumn dry period are urgently requiredin Mediterranean climates to complement annual pastures and the perennial legume lucerne (Medicago sativa). This studyevaluated the establishment, survival, and herbage production of tedera (Bituminaria bituminosa var. albomarginata)and Cullen spp. native to Australia. Two experiments were replicated at Buntine (warmer site) and Newdegate (cooler site)in the low-rainfall cropping zone (<350mm average annual rainfall) of Western Australia from June 2008 to September2010. In the first experiment, established by transplanting seedlings, survival and herbage production of two accessionseach of B. bituminosa andC. australasicumwere studied under densities of 1, 2, 4, 8, and 16 plants/m2 with 0, 1, 2, or 3 cutsin summer–autumn in addition to a winter–spring cut. In the second experiment, established from seed, emergence andsurvival of several accessions of B. bituminosa, C. australasicum, andM. sativa were studied, along with C. pallidum andC. cinereum.
In the first experiment, B. bituminosa survived better than C. australasicum (70–80% v. 18–45%), especially atBuntine, but there was little impact of density or cutting frequency on survival. Plant death was highest during summer.Shoot dry weight (DW) accumulation varied greatly with site, year, and plant density. When rainfall was close to average,shoot DW was greater at Newdegate (B. bituminosa �7.4 t/ha, C. australasicum �4.5 t/ha) than at Buntine (�2.3 t/ha),and both species produced much of their shoot DW in summer–autumn (e.g. 6 t/ha for B. bituminosa and 3 t/ha forC. australasicum at Newdegate). An early-summer cut reduced the DW that could be harvested later in summer–autumn.In the second experiment, emergence of B. bituminosa was either similar to, or higher than, emergence of the otherspecies, being 43% at Buntine and 44% at Newdegate. Survival of B. bituminosa, compared withM. sativa, was similar atBuntine (13%) and slightly lower at Newdegate (14%). Emergence and survival of Cullen spp. varied among species andaccessions, with survival of the best performing accession of C. australasicum (SA4966) similar to that of B. bituminosaand M. sativa at both sites. We conclude that B. bituminosa shows promise as a perennial summer forage for low-rainfallzones, with a density of 8–16 plants/m2 and cutting frequency of 3 cuts/year (i.e. cut twice in summer–autumn), whileC. australasicum and C. pallidum warrant further study.
Additional keywords: Australian native legumes, Bituminaria bituminosa var. albomarginata, Cullen spp., drought-tolerant perennial pastures, summer–autumn feed gap, survival, tedera.
Received 26 November 2012, accepted 15 April 2013, published online 3 May 2013
Introduction
Perennial pasture legumes play an important role in themanagement of dryland salinity and can provide high-qualitygreen forage, especially during the summer–autumn feed gap
when annual forages have senesced (Cocks 2001; Bell et al.2006b; Ward 2006). The only widely sown perennial legume inthe low rainfall (<375mm average annual rainfall) croppingzone of Western Australia is Medicago sativa L. (lucerne or
Journal compilation � CSIRO 2013 www.publish.csiro.au/journals/cp
CSIRO PUBLISHING
Crop & Pasture Science, 2013, 64, 71–85http://dx.doi.org/10.1071/CP12398

alfalfa), which may not persist well in areas with acid soils, lowsummer rainfall, or winter waterlogging (Humphries andAuricht2001; Ward et al. 2006). New perennial pasture legumes aretherefore sought for inclusion into this region and throughoutsouthern Australia (Bell et al. 2006a, 2006b; Dear et al. 2007; Liet al. 2008; Hayes et al. 2009; Bennett et al. 2011; Real et al.2011).
An evaluation of persistence and productivity of 103perennial forage legume and herb species over 3 years at arange of sites in southern Australia ranked 12 entries as mostpromising, including the Australian native Cullen australasicum(Schltdl.) J.W. Grimes and the exotic Bituminaria bituminosaC.H. Stirt. var. albomarginata (tedera) (Real et al. 2011). Cullenaustralasicum appears well adapted to both the alkaline soils ofthe South Australian cropping zone and the acidic soils of theWestern Australia cropping zone (Li et al. 2008; Bennett et al.2011, 2012). Bituminaria bituminosa var. albomarginata isnative to Lanzarote in the Canary Islands, Spain, where it isreported to be well adapted to low rainfall and heavy grazing(Gutman et al. 2000; Muñoz et al. 2000). For both of thesespecies, their value in current agricultural systems may accruefrom the provision of green feed during the summer–autumn feedgap (Finlayson et al. 2012). In this context, they are likely to begrown with annual plant-based pastures as companion speciesand to complement, at the farm level, lucerne pastures.
For all new pasture species, factors affecting seedlingemergence, survival, and herbage production are not wellunderstood (Li et al. 2008; Suriyagoda et al. 2012). Plantingdensity may affect survival rate, quantity, and quality (leaf tostem ratio, edible to non-edible ratio, branching pattern) ofpastures, and the degree of competition among plants of thesown species and with other species (Black 1960; Springer andGillen 2007; Dolling et al. 2011; Suriyagoda et al. 2011).
Cutting or grazing frequency also affects pasture quantityand quality. Cutting too frequently may cause low herbageproduction and plant death due, in part, to reductions in crownand root reserves (Kerridge and Skerman 1968; Gutteridge andWhiteman 1975; Humphries et al. 2006; Robinson et al. 2007;Striker et al. 2011). For instance, Kerridge and Skerman (1968)found that the yield of Psoralea eriantha Benth. [syn. Cullenpatens (Lindl.) J.W. Grimes] increased by 30% when cut every12 weeks instead of every 6 weeks. However, species andaccessions differ in their response to plant density and cuttingfrequency (Black 1960; Gutteridge and Whiteman 1975;Robinson et al. 2007). The importance of planting density andcutting frequency for survival and herbage production ofB. bituminosa and most Cullen species is not known.
A factor that could enhance adoption of a new perennialpasture species in the Mediterranean climate of south-westernAustralia is high herbage production in summer–autumn. Goodpasture herbage production has already been achieved inwinter–spring with annual pasture species, particularly withthe wide array of annual legumes now available (Nichols et al.2007). However, in summer–autumn (i.e. December–May),livestock must rely on the dry residues of annual species orcrops, or feed supplements either produced on-farm orpurchased. This summer–autumn feed gap also reduceslivestock production (e.g. net loss in fleece-free liveweight)(Moore et al. 2009). Thus, forage produced during this period
of low supply has high economic value (Bathgate and Pannell2002; Finlayson et al. 2012).
There were three objectives for the present study:
(i) To quantify the survival of C. australasicum andB. bituminosa in transplanted, spaced-plant experiments,under five plant densities (1, 2, 4, 8, and 16 plants/m2) andfour cutting frequencies (1, 2, 3, and 4 cuts/yearwith 0, 1, or2 cuts in summer–autumn) at two field experimental sitesin the low-rainfall cropping zone of Western Australia.The relatively low densities and cutting frequencieswere chosen to ensure plants remained productive in thesummer–autumn dry season and to allow companionannual pastures to be productive during the winter–spring growing season. Similar treatments have beenexamined in studies of other species with desirable out-of-season feed production (McGowan and Mathews 1992;Douglas et al. 1996).
(ii) To quantify shoot dry weight (DW) accumulation ofC. australasicum and B. bituminosa under five plantdensities and four cutting frequencies, particularly duringthe summer–autumn feed gap.
(iii) To quantify the survival of C. australasicum, C. cinereum(Lindl.) J.W. Grimes, C. pallidum (A. Lee), andB. bituminosa, along with M. sativa, in row experiments,established from seed, at two sites in the low-rainfallcropping zone of Western Australia.
We hypothesised that (i) at the relatively low plant densitiesexamined, higher plant density has little impact on survival ratebut increases DW accumulation; (ii) less frequent cuttingenhances survival and allows greater DW to be harvested inthe summer–autumn feed gap, but reduces annual productivity;(iii) more frequent cutting increases the leaf : stem ratio, andhence improves herbage quality; and (iv) M. sativa has betteremergence and survival than both B. bituminosa and the Cullenspecies at the site with the cooler weather, whereas the oppositeoccurs at the hotter site.
Materials and methodsSite description
Two experimental sites were established. One was at Buntine(30800ʹ30ʹʹS, 116820ʹ40ʹʹE; 20 km west of Buntine), a long-term research site of the Liebe Group (www.liebegroup.org.au), and the other was at the Department of Agricultureand Food, Western Australia (DAFWA) Research Station,Newdegate (33806ʹ16ʹʹS, 118849ʹ50ʹʹE; 18 km west ofNewdegate). These sites represented the low-rainfallcropping zone of Western Australia with long-term averageannual rainfall <375mm (see Supplementary Materials asavailable on journal’s website: Table 1S and Fig. 1S); rainfalllong-term averages showed drier summers and wetterautumn–winters at Buntine than at Newdegate, andtemperatures at Newdegate were consistently lower than atBuntine. Both sites had flat to undulating topography. Forthe previous 3 years, the Buntine site had been cultivatedwith wheat (Triticum aestivum L.) and the Newdegate sitewith narrow-leaf lupin (Lupinus angustifolius L.) and wheatin rotation. Both sites were low in mineral nitrogen (N), while
72 Crop & Pasture Science L. D. B. Suriyagoda et al.

available phosphorus (P) and potassium (K) were lower atNewdegate (Supplementary Material: Table 2S).
Plant material
For the density and cutting frequency experiments, hereafterreferred to as the ‘density experiments’, two accessions ofB. bituminosa (1, 6) and two accessions of C. australasicum(SA42566, SA44373) were used. For the experiments with seedsown in rows, hereafter referred to as the ‘row experiments’, fouraccessions of B. bituminosa (2, 3, 5, 9); three accessions ofC. australasicum (SA4966, SA42566, SA44373); oneaccession of C. cinereum (AusTRCF320100); one accessionof C. pallidum (SA44387); and four accessions (SA43158,SA43483, SA43484, SA44979), three breeding lines (PA4,TA22, A161), and one cultivar (SARDI Ten) of M. sativawere used. The accessions of each species used wereexperimental lines or cultivar controls, recommended bybreeders, for which adequate seed was available. Seeds ofC. australasicum, C. cinereum, and C. pallidum weremultiplied at The University of Western Australia ResearchStation at Shenton Park. Seeds of B. bituminosa were collectedfrom multiplication plots established at the DAFWA MedinaResearch Station, and M. sativa seeds were obtained from theGenetic Resource Centre at the South Australian Research andDevelopment Institute. Seeds were mechanically scarified andinoculated with peat cultures containing compatible rhizobiabefore seeding. Cullen australasicum was inoculated with RRI2761 and RRI 2781 sourced from the Rutherglen Centre,Victorian Department of Primary Industries. Bituminariabituminosa was inoculated with WSM 4083. The mixes ofroot nodule bacteria strains were based on the advice of theRutherglen Centre (Matt Denton and Bernadette Carmody, pers.comm.). At the end of the experiment, the presence of active, livenodules was observed, but numbers were not quantified. Beforesowing in the field, germination of each accession was tested inPetri dishes at 18–208C in dark conditions (four Petri dishes foreach accession with a minimum of 50 seeds each) before sowingin the field in order to adjust the sowing rate.
Field design, site preparation, and management
The density experiments were established as a split-split-plotdesign with four blocks and a 1-m buffer between blocks.Accessions were considered as the main plot factor. Densityand cutting frequency were considered as the subplot and split-subplot factors, respectively. There were five densities (1, 2, 4, 8,and 16 plants/m2, referred to as d01, d02, d04, d08, d16) andfour cutting frequencies (1, 2, 3 and 4 cuts/year, referred to as
Cut-1, Cut-2, Cut-3, Cut-4). All treatments were cut once inwinter–spring, while the number of cuts over summer–autumnvaried from 0 to 3. Each split subplot was 1 by 1m. Inoculatedseeds of all four accessions were raised for at least 6 weeks inseedling trays filled with a potting mixture before transplantingon 9 and 16 June 2008 at Newdegate and Buntine, respectively.Depending on the plant density, seedlings were transplanted at auniform spacing within each split subplot. Once plants had beenestablished, they were cut 2–4 cm from the ground at the timeintervals listed in Table 1. For C. australasicum at Newdegate,only Cut-2 and Cut-3 were made in the first and second year,respectively.
The row experiments (row nurseries) were established in arandomised complete block design with six blocks and with a1-m buffer between blocks. Each block consisted of 18 rows 1mlong and 1mapart, with each rowassigned to a randomly selectedaccession. Furrows 2 cm deep were prepared and seedsuniformly placed in them by hand and covered with a 1-cmlayer of fine topsoil. Seeds were sown at a rate calculated toresult in 50 germinable seeds per 1m row. Seeds were sown on9 and 16 June 2008 at Newdegate and Buntine, respectively. Aguard-row was established around each experiment usingM. sativa cv. SARDI Ten. There was no evidence of seed lossdue to disease or insect damage. Rows were cut 2–4 cm fromthe ground in January, May, and August 2009 and 2010. Slightvariation in cutting height occurred due to the unevenness inthe soil surface.
No fertilisers were added to the experiments, in line withgeneral farmer practice. Glyphosate (Roundup) was applied at arate of 540 g a.i./ha before site preparation. The grass-selectiveherbicide clethodim (Select) was applied at 60 g a.i./ha in August2008 to reduce competition from grasses, mainly annual ryegrass(Lolium rigidum L.), while broad-leaf weeds, mostly capeweed(Arctotheca calendula L. Levyns) and wireweed (Polygonumaviculare L.), were hand-weeded. Therefore, competition fromweeds was minimal. All sites were protected from wild animalsand livestock using meshed fences.
Soil analyses
At each site, in February 2010, ten soil samples were taken fromthe top 10 cm layer of soil from randomly selected locationswithin each block of the density experiments (four blocks) androw experiments (six blocks), in February 2010, using a 8-cm-diameter soil auger. Each soil sample was analysed separately atCSBP Ltd (Bibra Lake, W. Aust.). Ammonium-N and nitrate-Nconcentrations were calculated as described by Searle (1984),available P and Kwere measured using the Colwell test (Colwell
Table 1. Cutting schedule over the 2-year experimentAll treatments were cut once in winter–spring, and the number of cuts over summer–autumn varied from 0 to 3. Seedlings were transplanted into the field in
June 2008
Cutting frequency First year Second year(no. of cuts/year) Nov. 08 Jan. 09 May 09 Aug. 09 Dec. 09 Feb. 10 Apr. 10 Sept. 10
Early summer Summer Autumn Winter–spring Early summer Summer Autumn Winter–spring
1 � �2 � � � �3 � � � � � �4 � � � � � � � �
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 73

1965; Rayment and Higginson 1992), available sulfur (S)according to Blair et al. (1991), and organic carbon accordingto Walkley and Black (1934), with the remainder of analysesusing protocols outlined in Rayment and Higginson (1992).Cation exchange capacity was calculated from exchangeablesoil cations (Black 1968).
Data collection
Sites were visited at 1–2-month intervals and the numbers ofplants in density and row experiments counted. Final plantcounts were recorded on 9 and 30 September 2010 for Buntineand Newdegate, respectively. For the row experiments, theproportion of plant survival in each row was calculated as thenumber of plants present relative to the number of germinableseeds sown (50 seeds). As seedling counts tended to first increaseand then gradually decrease with time, the date at which thehighest number of seedlings was recorded was consideredpeak emergence. For the density experiments, the proportionof surviving plants in each plot was calculated as the number ofplants present relative to the density of seedlings transplanted.Towards the end of the first year of the density experiments,plants produced seeds. Therefore, naturally regeneratedseedlings were counted from each split-subplot using a 50 by50 cm quadrat on August 2009 (1 year after establishment). Asdetached seeds from plants couldmove on soil for short distancesdue to wind and water flow, naturally regenerated seedlingswere analysed only at the main-plot level (i.e. species), andthus the data from all cutting frequencies and all densitieswere pooled for each block. Daily mean, minimum, andmaximum temperatures and rainfall data for the period ofstudy were obtained from the Bureau of Meteorology website(www.bom.gov.au/climate/dwo/IDCJDW0600.shtml).
Biomass estimation from density experiments
The method of shoot cutting differed among density treatments.For d08 and d16, only the four central plants of each plot werecut, dried, and weighed. Remaining plants were cut anddiscarded. For d01, d02, and d04, where it was assumed thatcompetition among plants within a plot (and thus edge effects)was negligible, all plants within a plot were cut, dried, andweighed. In the first year (i.e. the initial four harvests), onerandomly selected plant from each plot was partitioned intoflowers, leaves, and edible and non-edible stems. Stems witha diameter >5mm were considered non-edible, as forC. australasicum these stems appeared woody. All sampleswere dried at 608C for 4 days, with DW expressed on a per m2
basis. Flowers, leaves, and edible and non-edible stem DWswere expressed as a percentage of whole plant DW.
Additional analysis was made of DW accumulation oversummer. For this analysis, DW accumulation between the firstand third cuts each year was considered (i.e. November 2008to May 2009 and December 2009 to April 2010) (Table 1). Theperiods from first to second cut, and from second to third cut,were considered as ‘early summer’ and ‘summer–autumn’periods, respectively. The growth rate (DW increase/m2.day)was estimated by dividing total DW produced per m2 by thenumber of growing days.
Volunteer annual pasture species were harvested at Buntineonly at the August 2009 sampling using a 50 by 50 cm quadrat in
each plot. Four randomly selected quadrats just outside theexperimental area were also harvested to study their growth innatural stand. Samples were dried at 608C for 1 week, with DWexpressed on a m2 basis.
Statistical analyses
Proportional survival data collected for 2 years were analysedusing binomial generalised linear mixed-effect models, usingPROC GLIMMIX with a logistic link function in SAS/STATsoftware Version 9.1 (SAS Institute Inc., Cary, NC, USA; SAS2003) considering site and time as random effects. Whenanalysing survival data of the density experiments, a fullmodel was used with time, site, block, species, density, cuttingfrequency, and their interactions considered as explanatoryvariables. For the row experiments, block, species, site, cuttingdate (i.e. time), and possible interactions were considered asexplanatory variables. During these analyses, the overallmodel was statistically significant (generalised chi-square/d.f. = 3.6, P < 0.0001 and 4.17, P < 0.0001 for density and rowexperiments, respectively), and significant effects for site, time,and numerous interactions between time and other explanatoryvariables were detected (Supplementary Material: Tables 3S,4S). Therefore, further analyses were performed for each siteand each time event separately. When comparing accessions ofa given species, a similar approach was used (SupplementaryMaterial: Tables 2S, 3S). As maximum emergence counts inthe row experiments were observed in September 2008 forBuntine and July 2008 for Newdegate, those seedling countswere used when comparing maximum emergence using thePROC GLIMMIX procedure, considering site, block, andspecies as explanatory variables (Supplementary Material:Table 3S). Results are presented as proportions with binomialstandard errors.
Dry weight from the density experiment was analysed usingthe procedureMIXED inSAS, considering cutting date (i.e. time)as a repeated-measure or within-subject factor. Explanatoryvariables in the model other than time were site, block,accession, density, cutting frequency, and all their possibleinteractions. The overall model was statistically significant(F1415, 6399 = 3.84� 103, P< 0.0001), and significant effectsfor site, time, and numerous interactions between time andother explanatory variables were detected (SupplementaryMaterial: Table 5S). Therefore, subsequent analyses wereperformed for each cutting date (time) and site (location). Ateach cutting date and site, the design was a split-split-plot withaccession, density, and cutting frequency as the main-plotfactor, subplot factor, and sub-subplot factor, respectively. Ateach site and cutting date, numerous significant interactionswere present among accessions, cutting frequency (cuts), anddensity (Supplementary Material: Table 6S). Accessions,densities, and cutting frequencies for each treatmentcombination were compared separately with the LSMEANSprocedure in SAS. A similar analysis was performed whenanalysing DW increase over summer, and DW of volunteerannual pasture species collected at Buntine (SupplementaryMaterial: Table 5S).
The proportion of edible DW (i.e. DW of flowers, leaves,and edible stems compared with total aboveground DW) wasanalysed using PROC CATMOD in SAS. The proportions were
74 Crop & Pasture Science L. D. B. Suriyagoda et al.

not affected by site, cutting frequency, or density for thefirst threecuts (November 2008, January 2009,May 2009) (SupplementaryMaterial: Table 5S). Therefore, when presenting results, datawere pooled across site, cutting frequency, and density. For thefourth cut of the first year (August 2009), site, cutting frequency,and accession all affected the proportion of flower, leaf, and stem(Supplementary Material: Table 5S). Counts of naturallyregenerated seedlings in the density experiment were subjectedto PROC CATMOD in SAS to test for species (accessions)and site effects. Soil characteristics were compared amongsites using PROC ANOVA in SAS. Comparisons betweenmeans were made using the LSMEANS procedure. Allsignificances were judged and expressed in the results sectionat a= 0.05
Results
Weather during the study period
At Buntine, total annual rainfall and summer and autumnrainfall totals were close to the long-term averages forboth years of the experiment (Supplementary Material:Fig. 1S, Table 1S). At Newdegate, a wet December (58mm)in the first year was followed by only 26% of the long-termaverage rainfall in autumn. The second summer was also verydry at this site, with only 5% of the long-term average summerrainfall received (5mm). Following average rainfall in Marchand April and 84mm in May, the site then received <10mm ofrainfall from June to September.
Plant survival in density experiments
Bituminaria bituminosa generally survived better thanC. australasicum throughout the study period, irrespective ofcutting frequency and density at both sites (Figs 1, 2). In addition,at the end of the experiment (September 2010), survival ofB. bituminosa was similar at Buntine and Newdegate, whereasC. australasicum SA44373 survived better at Newdegate thanBuntine.
At Buntine, irrespective of cutting frequency and density,survival of B. bituminosa was >90% after 12 months and ~80%by the end of the experiment. Survival of C. australasicum was~80% after 12 months and ~35% by the end of the experiment(Fig. 1). Survival of C. australasicum SA42566 was lower thanthat of SA44373 (21% v. 46%). For both species, plant deathoccurred primarily during the second summer.
At Newdegate, 12 months after establishment (July 2009),survival of both B. bituminosa accessions was >95%, whichdecreased to 73% by the end of the experiment, irrespectiveof cutting frequency and density (Fig. 2). There was little plantsdeath after establishment for C. australasicum at Newdegate.Therefore, the survival rate of C. australasicum was highlyvariable from the beginning of the experiment and lower thanthat of B. bituminosa. As a consequence, C. australasicum wasonly cut twice during the first year and three times during thesecond year. Plant death was not higher during summer.
Shoot dry weight
Shoot DW accumulation in the first year differed between sites,being up to 7.4 t/ha at Newdegate and up to 2.3 t/ha at Buntine(Figs 3, 4). Shoot DW also differed between years (P < 0.005),
with DW in the first year at least double that of thesecond year. There was also a three-way interaction amongcutting frequency, plant density, and species or accession ateach site (P < 0.001).
At Buntine in the first year, shoot DW production differedlittle among species and accessions (Fig. 3). However, in thesecond year, B. bituminosa production was much greater thanthat of C. australasicum, which produced very little DW. ShootDW tended to increase with plant density, irrespective of cuttingfrequency for all species or accessions, except C. australasicumin the second year (Fig. 3). Whereas B. bituminosa 1 generallyreached its highest shootDWat d16 (16plants/m2),B. bituminosa6 reached its greatest shootDWat d08–d16, andC. australasicumat d04–d08. Cutting frequency had little impact on shoot DW.For both species in thefirst year, >50%of shootDWaccumulatedin the first winter–spring growth period, i.e. between experimentestablishment in June 2008 and November 2008 (see Fig. 3,Cut-4, i.e. 4 cuts/year). The second most productive period wasMay–August 2009 for B. bituminosa. Cullen australasicum wasnot productive from May–August; shoot DW accumulationcontinued to be high from November 2008 to January 2009,before slowing in autumn and winter.
The DW produced at Newdegate was higher than that atBuntine, shoot DW accumulation in the first year reaching7.4 t/ha for B. bituminosa and 4.5 t/ha for C. australasicum(Fig. 4). In the second year, shoot DW accumulation at thehigher densities was generally ~50% less than in the first year,with the exception of B. bituminosa 6 in Cut-2, d16, which hadsimilar DWaccumulation in both years. Shoot DWaccumulationgenerally peaked at d08 or d16 forB. bituminosa andwas greatestat d16 for C. australasicum. Cutting frequency had little impacton shoot DW accumulation, with the exception of a reductionin shoot DW accumulation at Cut-4 for B. bituminosa 1 in thefirst year, which appeared to be due to a reduction in growthbetween establishment and January after a cut in November.Bituminaria bituminosa produced more DW thanC. australasicum over this period.
The cut in early summer (Figs 3, 4: bars with black filling inCut-4) reduced total annual DW production for B. bituminosa inthe first year at Newdegate and in the second year at Buntine.In addition, for both species, early summer cutting greatlyreduced accumulated DW that could be harvested later insummer–autumn (Figs 3, 4).
Shoot DW components
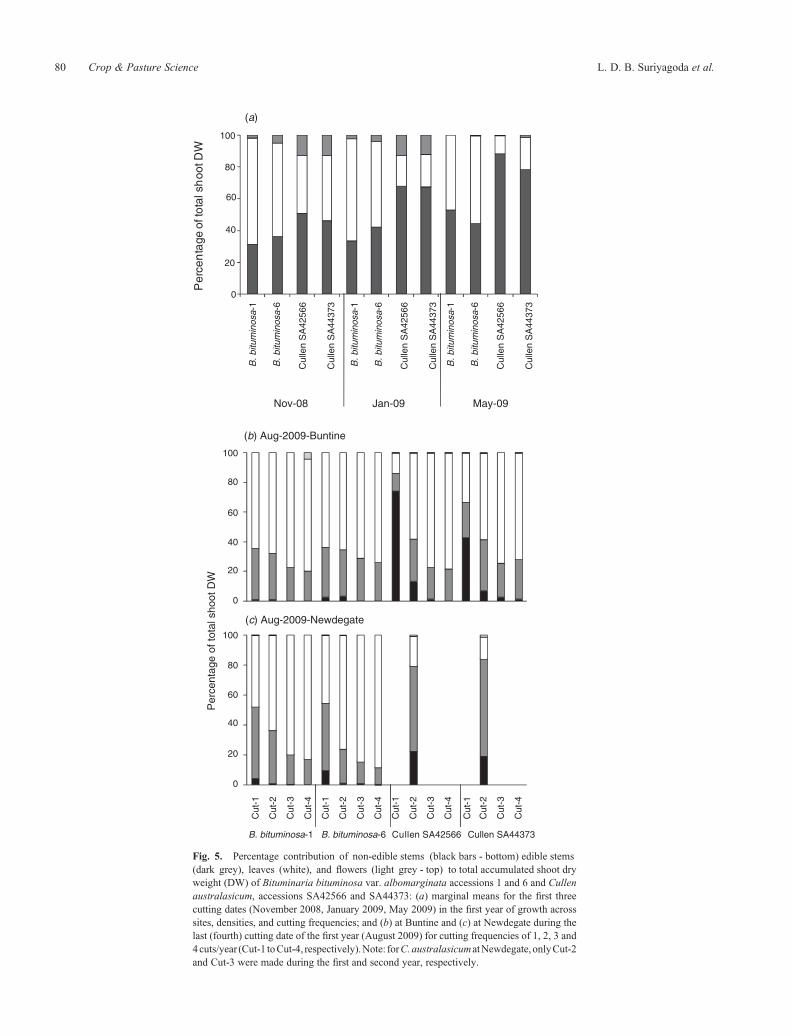
The contribution of leaves, stems, and flowers to total shoot DWat each harvest during the first year of growth is shown in Fig. 5.For the first three cutting times, DW differed among accessions(P< 0.001) (Fig. 5a). During this time, B. bituminosa had ahigher proportion of DW as leaves (and a lower proportion asedible stem or flowers) than C. australasicum; this differenceincreased (i.e. C. australasicum became stemmier) betweenNovember 2008 and May 2009.
At the fourth cut (i.e. August 2009, when Cut-1 was cut forthe first time), the interaction of accession� site� cuttingfrequency affected shoot DW distribution (P< 0.01; Fig. 5b,c). For B. bituminosa, the proportion of leaves increased withcutting frequency at each site, and only in Cut-1 and Cut-2 did
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 75

0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Age
(D
AT
)
Cut-3Cut-2 Cut-4
Proportion of survivalB
. bitu
min
osa
-1B
. bitu
min
osa
-6C
ulle
n S
A44
373
Cul
len
SA
4256
6Cut-1
Fig.1.
Proportionof
plantsurvival
ofBitu
minaria
bituminosavar.albo
margina
taaccessions
1and6andCullenau
stralasicum
accessions
SA42566andSA44373at
Buntin
ewith
time(daysafter
transplanting,DAT)atdensities
of1,2,4,8,and16
plants/m
2(¤
,&,~
,�,and
+,respectively)
andcutting
frequenciesof
1,2,3,and4cuts/year(Cut-1
toCut-4,respectively)
(i.e.0,1,2,or3cutsin
summer–autumnin
additio
nto
acutin
winter–spring).Binom
ialconfi
denceintervalfortreatm
entcom
parisons
isgivenin
thefirstsub-figu
re.
76 Crop & Pasture Science L. D. B. Suriyagoda et al.

Age
(D
AT
)
0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0 0
0.2
0.4
0.6
0.8
1.0
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Cut-2
Proportion of survival
Cut-1 Cut-3B
. bitu
min
osa
-1B
. bitu
min
osa
-6C
ulle
n S
A44
373
Cul
len
SA
4256
6Cut-4
Jul-08
Nov-08
Mar-09
Jul-09
Nov-09
Mar-10
Jul-10
Fig.2
.Propo
rtionof
plantsurvivalo
fBitu
minaria
bituminosavar.albo
marginata
accessions
1and6andCullenau
stralasicumaccessions
SA42
566andSA4437
3atNew
degatewith
time(daysafter
transplanting,
DAT)atdensities
of1,
2,4,
8and16
plants/m
2(¤
,&,~
,�,and
+,respectively)
andcutting
frequenciesof
1,2,
3,and4cuts/year(Cut-1
toCut-4,respectively)
(i.e.0
,1,2
,or3cutsin
summer–autumninadditio
ntoacutinwinter–spring).Binom
ialconfidenceintervalfortreatmentcom
parisons
isgiveninthefirstsub
-figu
re.N
ote,forC
.australasicum
,onlyCut-2andCut-3weremadedu
ring
thefirstand
second
year,respectively.
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 77
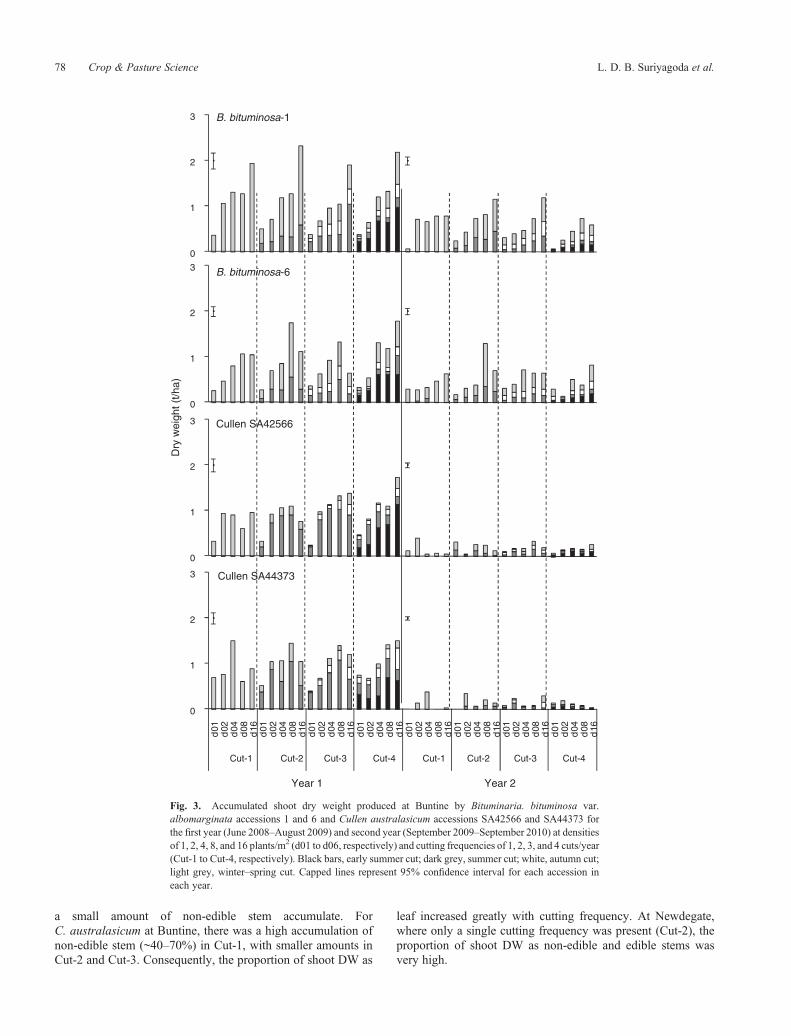
a small amount of non-edible stem accumulate. ForC. australasicum at Buntine, there was a high accumulation ofnon-edible stem (~40–70%) in Cut-1, with smaller amounts inCut-2 and Cut-3. Consequently, the proportion of shoot DW as
leaf increased greatly with cutting frequency. At Newdegate,where only a single cutting frequency was present (Cut-2), theproportion of shoot DW as non-edible and edible stems wasvery high.
0
1
2
3
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
Cut-4
0
1
2
3
0
1
2
3
0
1
2
3 B. bituminosa-1
B. bituminosa-6
Cullen SA44373
Dry
wei
ght (
t/ha)
Cullen SA42566
Year 1
Cut-1 Cut-2 Cut-3 Cut-4 Cut-1 Cut-2 Cut-3
Year 2
Fig. 3. Accumulated shoot dry weight produced at Buntine by Bituminaria. bituminosa var.albomarginata accessions 1 and 6 and Cullen australasicum accessions SA42566 and SA44373 forthe first year (June 2008–August 2009) and second year (September 2009–September 2010) at densitiesof 1, 2, 4, 8, and 16 plants/m2 (d01 to d06, respectively) and cutting frequencies of 1, 2, 3, and 4 cuts/year(Cut-1 to Cut-4, respectively). Black bars, early summer cut; dark grey, summer cut; white, autumn cut;light grey, winter–spring cut. Capped lines represent 95% confidence interval for each accession ineach year.
78 Crop & Pasture Science L. D. B. Suriyagoda et al.
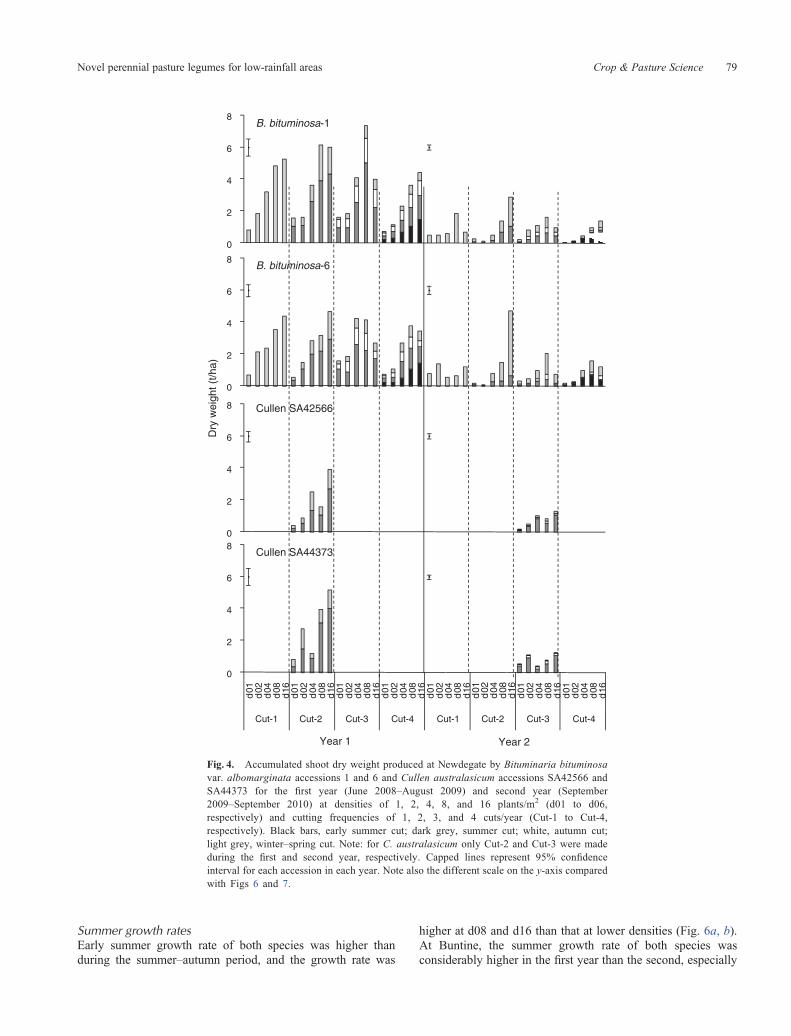
Summer growth ratesEarly summer growth rate of both species was higher thanduring the summer–autumn period, and the growth rate was
higher at d08 and d16 than that at lower densities (Fig. 6a, b).At Buntine, the summer growth rate of both species wasconsiderably higher in the first year than the second, especially
0
2
4
6
8
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
Cut-4
0
2
4
6
8
0
2
4
6
8
0
2
4
6
8B. bituminosa-1
B. bituminosa-6
Cullen SA44373
Cullen SA42566
Dry
wei
ght (
t/ha)
Year 1 Year 2
Cut-1 Cut-2 Cut-3 Cut-4 Cut-1 Cut-2 Cut-3
Fig. 4. Accumulated shoot dry weight produced at Newdegate by Bituminaria bituminosavar. albomarginata accessions 1 and 6 and Cullen australasicum accessions SA42566 andSA44373 for the first year (June 2008–August 2009) and second year (September2009–September 2010) at densities of 1, 2, 4, 8, and 16 plants/m2 (d01 to d06,respectively) and cutting frequencies of 1, 2, 3, and 4 cuts/year (Cut-1 to Cut-4,respectively). Black bars, early summer cut; dark grey, summer cut; white, autumn cut;light grey, winter–spring cut. Note: for C. australasicum only Cut-2 and Cut-3 were madeduring the first and second year, respectively. Capped lines represent 95% confidenceinterval for each accession in each year. Note also the different scale on the y-axis comparedwith Figs 6 and 7.
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 79
B. b
itum
inos
a-1
B. b
itum
inos
a-6
Cul
len
SA
4256
6
Cul
len
SA
4437
3
B. b
itum
inos
a-1
B. b
itum
inos
a-6
Cul
len
SA
4256
6
Cul
len
SA
4437
3
May-09
Per
cen
tage
of t
otal
sh
oot D
W
0
20
40
60
80
100
Cut
-1
Cut
-2
Cut
-3
Cut
-4
Cut
-1
Cut
-2
Cut
-3
Cut
-4
Cut
-1
Cut
-2
Cut
-3
Cut
-4
Cut
-1
Cut
-2
Cut
-3
Cut
-4
(c) Aug-2009-Newdegate
B. bituminosa-1 B. bituminosa-6 Cullen SA42566 Cullen SA44373
Per
cent
age
of to
tal s
hoot
DW
0
20
40
60
80
100
0
20
40
60
80
100B
. bitu
min
osa-
1
B. b
itum
inos
a-6
Cul
len
SA
4256
6
Cul
len
SA
4437
3
Nov-08 Jan-09
(a)
(b) Aug-2009-Buntine
Fig. 5. Percentage contribution of non-edible stems (black bars - bottom) edible stems(dark grey), leaves (white), and flowers (light grey - top) to total accumulated shoot dryweight (DW) of Bituminaria bituminosa var. albomarginata accessions 1 and 6 and Cullenaustralasicum, accessions SA42566 and SA44373: (a) marginal means for the first threecutting dates (November 2008, January 2009, May 2009) in the first year of growth acrosssites, densities, and cutting frequencies; and (b) at Buntine and (c) at Newdegate during thelast (fourth) cutting date of the first year (August 2009) for cutting frequencies of 1, 2, 3 and4cuts/year (Cut-1 toCut-4, respectively).Note: forC.australasicumatNewdegate, onlyCut-2and Cut-3 were made during the first and second year, respectively.
80 Crop & Pasture Science L. D. B. Suriyagoda et al.
0246810 0246810
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
0246810 0246810
B. b
itum
inos
a-1
B. b
itum
inos
a-6
Cul
len
SA
4256
6
Cul
len
SA
4437
3
Dry weight increase (kg/ha.day)
Yea
r 1
Yea
r 2
Yea
r 1
Yea
r 2
(a)
Bu
ntin
e
Cu
t 4C
ut 3
Cut
3C
ut 4
05
10
15
20
25
30051015202530
051015202530 051015202530
B. b
itum
inos
a-1
B. b
itum
inos
a-6
Cul
len
SA
4256
6
Cul
len
SA
4437
3
(b)
New
dega
te
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
d01
d02
d04
d08
d16
Cu
t 4C
ut 3
Cut
3C
ut 4
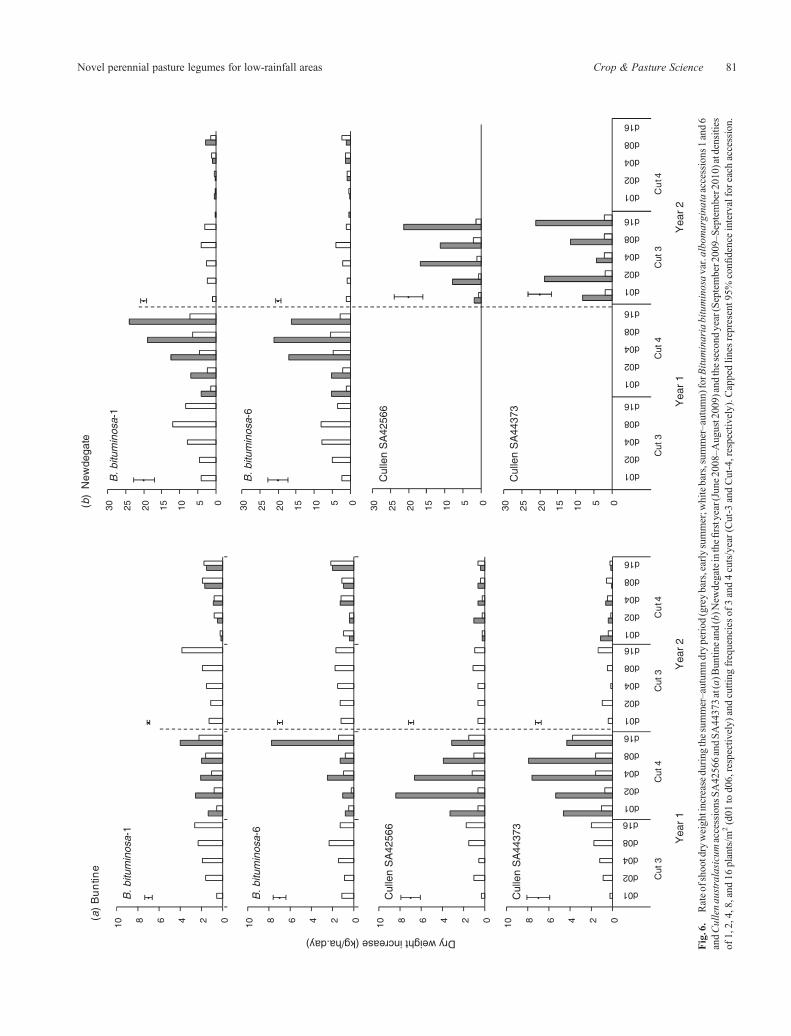
Fig.6.
Rateofshootdryweightincreasedu
ring
thesummer–autumndryperiod
(greybars,earlysummer;w
hitebars,sum
mer–autumn)forB
ituminariabituminosavar.albo
marginataaccessions
1and6
andCullenau
stralasicumaccessions
SA4256
6andSA4437
3at(a)B
untin
eand(b)N
ewdegateinthefirstyear(June
2008–August2009)andthesecond
year(September2009–September201
0)atdensities
of1,2,4,8,and16
plants/m
2(d01
tod0
6,respectiv
ely)
andcutting
frequenciesof
3and4cuts/year(Cut-3
andCut-4,respectively).C
appedlin
esrepresent95%
confi
denceintervalforeachaccession.
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 81
for C. australasicum in the early summer period (P< 0.01;Fig. 6a, grey bars). In the first year, C. australasicum had ahigher growth rate than B. bituminosa at the lower densities inthe early summer period (i.e. d01–d08), whereas growth ratesamong densities and accessions were similar in thesummer–autumn period. Summer–autumn growth rates wereslightly lower when there was a cut in early summer (Cut-4),especially for B. bituminosa. At Newdegate, summer growthrates followed similar trends to those at Buntine, but weregenerally 3–4 times higher (Fig. 6b).
Natural regeneration from self-sown seeds
The number of regenerated seedlings in the density experimentsdiffered among sites and species (c2 = 1852, P< 0.0001;Supplementary Material: Fig. 2S). Cullen australasicum hadmore regenerated seedlings than B. bituminosa at Buntine(26% and 7%, respectively) and B. bituminosa did notregenerate at Newdegate.
Volunteer annual pasture
Volunteer annual pasture DW at the end of the first year (August2009) decreased with plant density of B. bituminosa andC. australasicum (P < 0.01; Supplementary Material: Fig. 3S).
At d01, pasture DW from annual volunteers was similar to thatoutside the experimental plots.
Row experiments
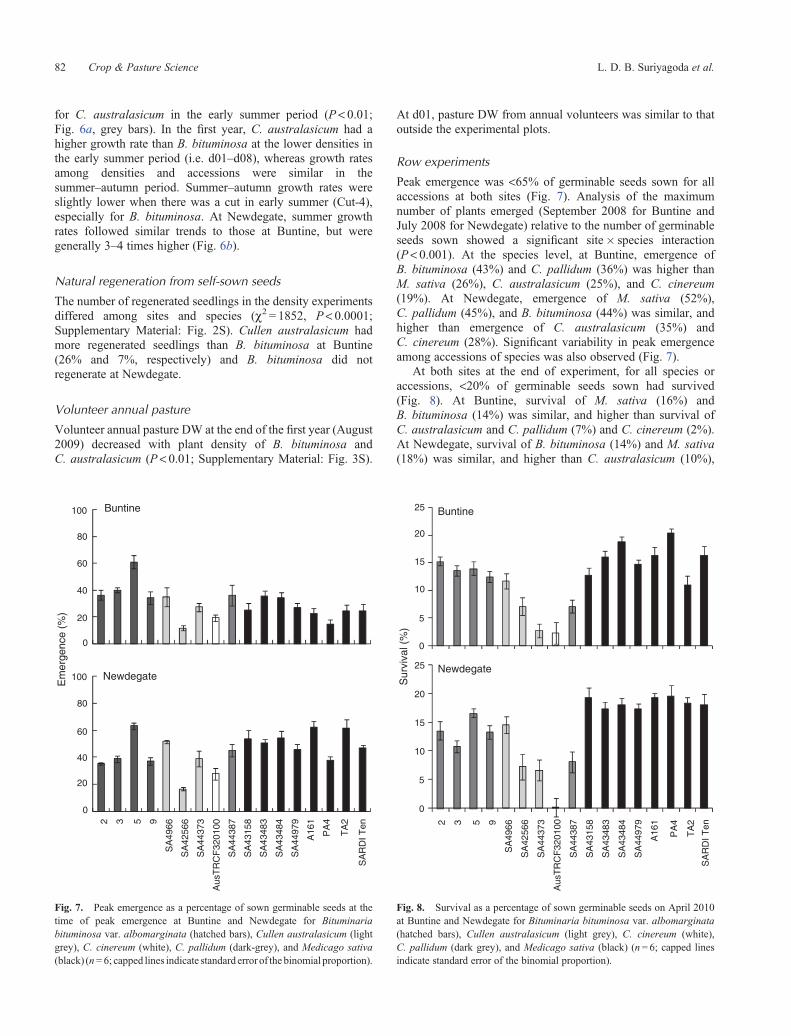
Peak emergence was <65% of germinable seeds sown for allaccessions at both sites (Fig. 7). Analysis of the maximumnumber of plants emerged (September 2008 for Buntine andJuly 2008 for Newdegate) relative to the number of germinableseeds sown showed a significant site� species interaction(P< 0.001). At the species level, at Buntine, emergence ofB. bituminosa (43%) and C. pallidum (36%) was higher thanM. sativa (26%), C. australasicum (25%), and C. cinereum(19%). At Newdegate, emergence of M. sativa (52%),C. pallidum (45%), and B. bituminosa (44%) was similar, andhigher than emergence of C. australasicum (35%) andC. cinereum (28%). Significant variability in peak emergenceamong accessions of species was also observed (Fig. 7).
At both sites at the end of experiment, for all species oraccessions, <20% of germinable seeds sown had survived(Fig. 8). At Buntine, survival of M. sativa (16%) andB. bituminosa (14%) was similar, and higher than survival ofC. australasicum and C. pallidum (7%) and C. cinereum (2%).At Newdegate, survival of B. bituminosa (14%) and M. sativa(18%) was similar, and higher than C. australasicum (10%),
0
20
40
60
80
100
0
20
40
60
80
100
Em
erge
nce
(%)
Buntine
Newdegate
2 3 5 9
SA
4966
SA
4256
6
SA
4437
3
SA
4438
7
Aus
TR
CF
3201
00
SA
4315
8
SA
4348
3
SA
4348
4
SA
4497
9
A16
1
PA
4
SA
RD
I Ten
TA
2
Fig. 7. Peak emergence as a percentage of sown germinable seeds at thetime of peak emergence at Buntine and Newdegate for Bituminariabituminosa var. albomarginata (hatched bars), Cullen australasicum (lightgrey), C. cinereum (white), C. pallidum (dark-grey), and Medicago sativa(black) (n= 6; capped lines indicate standard error of the binomial proportion).
0
5
10
15
20
25
0
5
10
15
20
25 Buntine
Newdegate
Sur
viva
l (%
)
2 3 5 9
SA
4966
SA
4256
6
SA
4437
3
SA
4438
7
Aus
TR
CF
3201
00
SA
4315
8
SA
4348
3
SA
4348
4
SA
4497
9
A16
1
PA
4
SA
RD
I Ten
TA
2
Fig. 8. Survival as a percentage of sown germinable seeds on April 2010at Buntine and Newdegate for Bituminaria bituminosa var. albomarginata(hatched bars), Cullen australasicum (light grey), C. cinereum (white),C. pallidum (dark grey), and Medicago sativa (black) (n = 6; capped linesindicate standard error of the binomial proportion).
82 Crop & Pasture Science L. D. B. Suriyagoda et al.

C. pallidum (8%), and C. cinereum (<1%). Survival variedamong accessions within species (Fig. 8, P < 0.05).
Discussion
Survival, dry weight production, and herbage quality
The first hypothesis—that at the relatively low plant densitiesexamined higher plant density has little impact on survival ratebut increases DW production—was partially supported. Densityhad little impact on survival rate for either species. This might bedue to the absence or extreme weakness of competition amongplants due to the deliberate use of quite low plant densities. ForDW production, annual shoot DW production increased withplant density in the first year at both sites, with the mostproductive accession of B. bituminosa (accession 1) producingthe most DW at d08 and d16, while accession 6 andC. australasicum had maximum DW between d04 and d16.However, the hypothesis was not so clearly supported in thesecond year, especially for the very poorly productiveC. australasicum at Buntine. There are several possibleexplanations. First, survival had declined by the second year,particularly for C. australasicum, with only 0–50% of plantsremaining. Second, as the experiments were unfertilised,perhaps P and K became limiting. Both B. bituminosaand C. australasicum respond strongly to addition of P(Suriyagoda et al. 2010; Pang et al. 2011). Finally, as bothsites had previously been sown to annual crops for at least3 years, herbage production in the first year may have beenenhanced by both species accessing, and then depleting, waterdeep in the profile (Bell et al. 2006a; Dolling et al. 2011).
As plant density had little effect on survival, we suggest thatboth species, when sown with the primary aim of providingfeed during summer and autumn, should be sown with a targetplant density of 8–16 plants/m2. Dolling et al. (2011)recommended >20 plants/m2 as the minimum density tooptimise M. sativa and total pasture herbage production for upto 4 years after establishment at sites that averaged 430mmannual rainfall. However, the sites in the current experimentreceived considerably less rainfall.
The second hypothesis was that lower cutting frequenciesenhance survival and allow greater DW to be harvested in thesummer–autumn feed gap, but reduce annual productivity.However, higher cutting frequency did not reduce plantsurvival, perhaps because the frequency of cutting (a minimumof 3 months) was always sufficient for plants to replenishdepleted carbohydrate reserves and enhance root growth(Gutteridge and Whiteman 1975; Humphries et al. 2006;Robinson et al. 2007). The increased plant death in thesummer periods for both species across sites presumablyresulted from moisture stress and was due to neither cuttingfrequency nor density.
The rate of DW increase in the summer–autumn feed-gapperiod showed little relationship to density and cuttingfrequency, but tended to increase with density in early summerwhen, presumably, water was more readily available. Contraryto the hypothesis, in most instances, more frequent cutting didnot increase annual shoot DW production. There was only oneinstance of a clear negative effect of cutting frequency. AtNewdegate in the first year, less shoot DW tended to
accumulate for B. bituminosa in Cut-4 (4 cuts/year, i.e. thehighest rate) than in Cut-3 (3 cuts/year). This reflected thecombined shoot DW from the first and second cuts (i.e.November and January) being less than that harvested whenthe first cut was in January. Perhaps the cut in Novemberreduced the ability of the plants to use the summer rainfall.Although the early summer cut did not reduce total annualDW accumulation, it reduced the accumulated DW later insummer–autumn. Therefore, early summer DW can beconserved for cutting in late summer–autumn when an earlysummer cut is not undertaken. Thus, we recommend that threecuts per year, with two in summer–autumn, is suitable forB. bituminosa, but for maximum summer herbage productionand conservation of DW for use in late summer–autumn, cuttingshould be avoided in early summer. Further work is required todetermine the cutting frequency effects on DW accumulationfor C. australasicum. Douglas et al. (1996) also found that a lowcutting frequency maximised yield of four shrub species grownin New Zealand, with yields higher from a single cut than thecombined yield from two or three cuts.
The third hypothesis—that more frequent cutting increasesthe leaf : stem ratio and hence improves herbage quality—wasgenerally supported, as more frequent cutting (4 cuts/year)mostly reduced the accumulation of edible stems and ensuredno accumulation of inedible stems. For C. australasicum atNewdegate, inedible stems accumulated to a very high level(>20% of shoot DW). This reflected the fact that only two cutswere made in the first year. Loss of mature leaves may also havecontributed.
Overall for these species, when considering DW producedper year and rate of DW produced during the summer–autumnfeed gap, a density of 8–16 plants/m2 and cutting frequency of3 cuts/year, twice during the summer–autumn feed-gap butnone in early summer, appears to be optimal, whereas acutting frequency of 4 cuts/year was the best for a higherherbage quality.
The fourth hypothesis—that M. sativa survives better thanthe Cullen species and B. bituminosa at the site with the coolerweather, while the opposite occurs at the hotter site—was mostlysupported. On average, M. sativa survival was the best atNewdegate, and was similar to B. bituminosa at Buntine.
When comparing accessions of B. bituminosa, accession 5had better emergence at both sites and better survival atNewdegate. Cullen australasicum SA4966, which was usedonly in the row experiment, had better survival than the othertwo accessions at both sites and its survival rate was similarto that of B. bituminosa. Inconsistent establishment forC. australasicum SA4966 was previously reported by Li et al.(2008), whereas Hayes et al. (2009) found that it establishedas well as M. sativa in a 3-year field study at Barmedman,New South Wales. In a spaced-plant experiment at Buntine,C. australasicum SA4966 had 100% survival over 18 months(Bennett et al. 2012). Lower survival in the current experiment(~12%) probably reflects dense seeding in a row (50 germinableseeds/m row), resulting in greater competition.
For the other Cullen species in the row experiment,C. pallidum had emergence and survival rates similar toC. australasicum. The survival rate of C. pallidum may alsohave been higher with superior germplasm, as survival of the
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 83

four accessions of C. pallidum investigated by Bennett et al.(2012) ranged from 22 to 89%, with the poorest performingaccession (SA44387) being the accession grown in thisexperiment. We agree with Bennett et al. (2012) that survivalof C. pallidum warrants further investigation. Survival ofC. cinereum was very low, even though the accession used inthe row experiment (AusTRCF320100) was one of the bestperforming of the 22 accessions of C. cinereum investigatedby Bennett et al. (2012), with 50% survival.
Volunteer pasture
At Buntine, at the end of the first year, there was a clear declinein the DW of volunteer annual pasture with increasing densityof perennials, particularly for B. bituminosa. Competition forwater, light, and nutrients was presumably responsible (Dearet al. 1998). Similar resultswere reported byDolling et al. (2011).An annual pasture component in low density pastures ofB. bituminosa and C. australasicum is desirable for severalreasons, including provision of a diverse feed supply forlivestock (Villalba et al. 2011) and maximising winter–springherbage production.
Natural regeneration of seedlings
Higher temperatures and higher autumn–winter rainfall atBuntine compared with Newdegate might explain why therewere more naturally regenerated seedlings at Buntine.However, this needs to be studied further. The management(e.g. cutting frequency) and environmental conditions (e.g.photoperiod, temperature, rainfall) conducive to flowering,seed set, and seedling establishment require further study forB. bituminosa and C. australasicum.
Concluding remarks
Survival (in both row and density experiments) and DWproduction (in the density experiment) of B. bituminosa andC. australasicum was influenced greatly by site, year, andplant density (for DW production). At Newdegate, the moreproductive site, summer–autumn herbage production wassubstantial, up to 3 t/ha for C. australasicum and 6 t/ha forB. bituminosa. Extremely dry conditions at Newdegate in thesecond year in summer and winter hindered assessment ofherbage production potential. A density of 8–16 plants/m2 islikely to maximise herbage production of the perennials andallow an understory of annual pasture. The results suggest thatthree cuts per year, with two in summer–autumn, is the mostdesirable for herbage accumulation and maintenance of a highleaf : stem ratio; avoidance of cutting in early summer isimportant to conserve herbage for use in late summer–autumn.Overall, B. bituminosa generally performed better in termsof survival and herbage production, although both specieshave potential for development as summer-active perennialpastures in the low-rainfall zone of the wheatbelt of WesternAustralia. Longer term experiments are now required todetermine whether adequate plant density can be maintainedover the longer term and to assess alternative germplasm ofB. bituminosa and C. australasicum.
Acknowledgments
We thank Steve Hughes, Ramakrishnan Nair, and Alan Humphries fromSARDI for providing theM. sativa seeds and Daniel Kidd for his assistancein data collection. Richard Bennett and Kevin Foster provided constructivecomments. This study was supported by the School of Plant Biology at TheUniversity of Western Australia, the Future Farm Industries CooperativeResearch Centre, and the Rural Industries Research and DevelopmentCorporation. LDB Suriyagoda also appreciates the SIRF/UIS Scholarshipawarded by The University of Western Australia and further scholarshipsupport from the late Frank Ford.
References
Bathgate A, Pannell D (2002) Economics of deep-rooted perennials inWestern Australia Agricultural Water Management 53, 117–132.doi:10.1016/S0378-3774(01)00160-3
Bell LW, Moore GA, Ewing MA, Ryan MH (2006a) Production, survivaland nutritive value of the perennial legumes Dorycnium hirsutum andD. rectum subjected to different cutting heights. Grass and ForageScience 61, 60–70. doi:10.1111/j.1365-2494.2006.00508.x
Bell LW, Ryan MH, Moore GA, Ewing MA (2006b) Comparative water useofDorycnium hirsutum-,M. sativa-, and annual legume-based pastures inthe Western Australian wheatbelt. Australian Journal of AgriculturalResearch 57, 857–865. doi:10.1071/AR05409
Bennett RG, Ryan MH, Colmer TD, Real D (2011) Prioritisation of novelpasture species for use in water-limited agriculture: a case study ofCullen in the Western Australian wheatbelt. Genetic Resources andCrop Evolution 58, 83–100. doi:10.1007/s10722-010-9567-3
Bennett RG, Colmer TD, Real D, Renton M, Ryan MH (2012) Phenotypicvariation for productivity and drought tolerance is widespread ingermplasm collections of Australian Cullen species. Crop & PastureScience 63, 656–671. doi:10.1071/CP11338
Black JN (1960) An assessment of the role of planting density in competitionbetween red clover (Trifolium pratense L.) and lucerne (Medicago sativaL.) in the early vegetative stage. Oikos 11, 26–42. doi:10.2307/3564882
Black CA (1968) ‘Soil–plant relationships.’ (Wiley Publishing & Sons: NewYork)
Blair GJ, Chinoim N, Lefroy RDB, Anderson GC, Crocker GJ (1991) A soilsulphur test forpastures andcrops.Australian Journalof SoilResearch29,619–626. doi:10.1071/SR9910619
Cocks PS (2001) Ecology of herbaceous perennial legumes: a review ofcharacteristics that may provide management options for the control ofsalinity and waterlogging in dryland cropping systems. AustralianJournal of Agricultural Research 52, 137–151. doi:10.1071/AR99170
Colwell JD (1965) An automatic procedure for the determination ofphosphorus in sodium hydrogen carbonate extract of soil. Chemistry &Industry 10, 893–895.
Dear BS, Cocks PS, Wolfe EC, Collins DP (1998) Established perennialgrasses reduce the growth of emerging subterranean clover seedlingsthrough competition for water, light, and nutrients. Australian Journalof Agricultural Research 49, 41–51. doi:10.1071/A97062
Dear BS, Li GD, Hayes RC, Hughes SJ, Charman N, Ballard RA (2007)Cullen australasicum (syn. Psoralea australasica): a review and somepreliminary studies related to its potential as a low rainfall perennialpasture legume. The Rangeland Journal 29, 121–132. doi:10.1071/RJ06039
Dolling PJ, Lyons AM, Latta RA (2011) Optimal plant densities of lucerne(Medicago sativa) for pasture production and soil water extraction inmixed pastures in south-western Australia. Plant and Soil 348, 315–327.doi:10.1007/s11104-011-0795-x
Douglas GB, Bulloch BT, Foote AG (1996) Cutting management ofwillows (Salix spp.) and leguminous shrubs for forage during summer.New Zealand Journal of Agricultural Research 39, 175–184.doi:10.1080/00288233.1996.9513176
84 Crop & Pasture Science L. D. B. Suriyagoda et al.

Finlayson J, Real D, Nordblom T, Revell C, Ewing M, Kingwell R (2012)A farm level assessment of a novel drought tolerant forage: Tedera(Bituminaria bituminosa C.H.Stirt var. albomarginata). AgriculturalSystems 112, 38–47. doi:10.1016/j.agsy.2012.06.001
Gutman M, Perevolotsky A, Sternberg M (2000) Grazing effects on aperennial legume, Bituminaria bituminosa (L.) Stirton, in aMediterranean rangeland. Cahiers Options Méditerranéennes 45,299–303.
Gutteridge RC, Whiteman PC (1975) Effect of defoliation frequency ongrowth and survival of four accessions of Psoralea erantha. AustralianJournal of Experimental Agriculture and Animal Husbandry 15,493–497. doi:10.1071/EA9750493
Hayes RC, Li GD, Dear BS, Humphries AW, Tidd JR (2009) Persistence,productivity, nutrient composition, and aphid tolerance of Cullen spp.Crop & Pasture Science 60, 1184–1192. doi:10.1071/CP09095
HumphriesAW,AurichtGC (2001)Breeding lucerne forAustralia’s southerndryland cropping environments. Australian Journal of AgriculturalResearch 52, 153–169. doi:10.1071/AR99171
Humphries AW, Kobelt ET, Bellotti WD, Auricht GC (2006) Tolerance ofAustralian lucerne (Medicago sativa) germplasm to grazing by sheep.Australian Journal of Experimental Agriculture 46, 1263–1270.doi:10.1071/EA04044
Kerridge PC, Skerman PJ (1968) The distribution and growth characteristicsof the native legume Psoralea eriantha in western Queensland. TropicalGrasslands 2, 41–50.
Li GD, Lodge GM, Moore GA, Craig AD, Dear BS, Boschma SP, AlbertsenTO,Miller SM, Harden S, Hayes RC, Hughes SJ, Snowball R, Smith AB,Cullis BC (2008) Evaluation of perennial pasture legumes and herbs toidentify species with high herbage mass and persistence in mixedfarming zones in southern Australia. Australian Journal ofExperimental Agriculture 48, 449–466. doi:10.1071/EA07108
McGowan AA, Mathews GL (1992) Forage production from hedges oftagasaste in a high rainfall temperate environment, and the effects ofplant spacing and frequency of harvesting. Australian Journalof Experimental Agriculture 32, 633–640. doi:10.1071/EA9920633
Moore AD, Bell LW, Revell DK (2009) Feed gaps in mixed-farmingsystems: insights from the grain and graze program. AnimalProduction Science 49, 736–748. doi:10.1071/AN09010
Muñoz A, Ortiz-Dorda AJ, Correal E (2000) Morphological and molecularcharacterization of Bituminaria bituminosa accessions from South-EastSpain and the Canary Islands. Cahiers Options Méditerranéennes 45,103–107.
Nichols PGH, Loi A, Nutt BJ, Evans PM, Craig AD, Pengelly BC, Dear BS,Lloyd DL, Revell CK, Nair RM, EwingMA, Howieson JG, Auricht GA,Howie JH, SandralGA,Carr SJ, deKoningCT,HackneyBF,CrockerGJ,SnowballR,HughesEJ,HallEJ, FosterKJ,SkinnerPW,BarbettiMJ,YouMP (2007) New annual and short-lived perennial pasture legumes forAustralian agriculture — 15 years of revolution. Field Crops Research104, 10–23. doi:10.1016/j.fcr.2007.03.016
Pang J, Yang J, Ward P, Siddique KHM, Lambers H, Tibbett M, Ryan M(2011) Contrasting responses to drought stress in herbaceous perenniallegumes. Plant and Soil 348, 299–314. doi:10.1007/s11104-011-0904-x
Rayment GE, Higginson FR (1992) ‘Australian laboratory handbook of soiland water chemical methods.’ (Inkata Press: Melbourne)
Real D, Li GD, Clark S, Albertsen TO, Hayes RC, Denton MD, D’AntuonoMF, Dear BS (2011) Evaluation of perennial forage legumes and herbsin six Mediterranean environments. Chilean Journal of AgriculturalResearch 71, 357–369. doi:10.4067/S0718-58392011000300003
Robinson K, Bell LW, Bennett RG, Henry DA, Tibbett M, Ryan MH (2007)Perennial legumes native to Australia: a preliminary investigation ofnutritive value and response to cutting. Australian Journal ofExperimental Agriculture 47, 170–176. doi:10.1071/EA06043
SAS (2003) ‘SAS/STAT User’s Guide, Version 9.1.’ (SAS Institute Inc.:Cary, NC)
Searle PL (1984) The berthelot or indophenol reaction and its use in theanalytical chemistry of nitrogen. A review. Analyst 109, 549–568.doi:10.1039/an9840900549
Springer TL, Gillen RL (2007) How does plant population density affectthe yield, quality, and canopy of native bluestem (Andropogon spp.)forage? Crop Science 47, 77–82. doi:10.2135/cropsci2005.12.0464
Striker GG, Manzur ME, Grimoldi AA (2011) Increasing defoliationfrequency constrains regrowth of the forage legume Lotus tenuis underflooding. The role of crown reserves. Plant and Soil 343, 261–272.doi:10.1007/s11104-011-0716-z
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2010) Multipleadaptive responses of Australian native perennial legumes with pasturepotential to grow in phosphorus- and moisture-limited environments.Annals of Botany 105, 755–767. doi:10.1093/aob/mcq040
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2011) Above- andbelow-ground interactions of grass and pasture legume species whengrown together under drought and low phosphorus availability. Plantand Soil 348, 281–297. doi:10.1007/s11104-011-0754-6
Suriyagoda LDB, Ryan MH, Lambers H, Renton M (2012) Comparison ofnovel and standard methods for analysing patterns of plant death indesigned field experiments. The Journal of Agricultural Science 150,319–334. doi:10.1017/S0021859611000566
Villalba JJ, Provenza FD, Clemensen AK, Larsen R, Juhnke J (2011)Preference for diverse pastures by sheep in response to intraruminaladministrations of tannins, saponins and alkaloids. Grass and ForageScience 66, 224–236. doi:10.1111/j.1365-2494.2010.00779.x
Walkley A, Black IA (1934) Estimation of soil organic carbon by thechromic acid titration method. Soil Science 37, 29–38. doi:10.1097/00010694-193401000-00003
Ward PR (2006) Predicting the impact of perennial phases on averageleakage from farming systems in south-western Australia. AustralianJournal of Agricultural Research 57, 269–280. doi:10.1071/AR04137
Ward PR, Micin SF, Dunin FX (2006) Using soil, climate, and agronomy topredict soil water use by lucerne compared with soil water use by annualcrops or pastures. Australian Journal of Agricultural Research 57,347–354. doi:10.1071/AR04132
Novel perennial pasture legumes for low-rainfall areas Crop & Pasture Science 85
www.publish.csiro.au/journals/cp




















