
Escherichia coli YqhD Exhibits Aldehyde Reductase Activity and Protects from the Harmful Effect of...
8
Escherichia coli YqhD Exhibits Aldehyde Reductase Activity and Protects from the Harmful Effect of Lipid Peroxidation-derived Aldehydes * Received for publication, October 26, 2007, and in revised form, January 16, 2008 Published, JBC Papers in Press, January 22, 2008, DOI 10.1074/jbc.M708846200 Jose ´ Manuel Pe ´rez 1,2 , Felipe A. Arenas 2 , Gonzalo A. Pradenas 1 , Juan M. Sandoval 1 , and Claudio C. Va ´ squez 3 From the Laboratorio de Microbiologı ´a Molecular, Departamento de Biologı ´a, Facultad de Quı ´mica y Biologı ´a, Universidad de Santiago de Chile, Santiago 9170019, Chile Evidence that Escherichia coli YqhD is involved in bacterial response to compounds that generate membrane lipid peroxi- dation is presented. Overexpression of yqhD results in increased resistance to the reactive oxygen species-generating compounds hydrogen peroxide, paraquat, chromate, and potassium tellu- rite. Increased tolerance was also observed for the lipid peroxi- dation-derived aldehydes butanaldehyde, propanaldehyde, acrolein, and malondialdehyde and the membrane-peroxidizing compound tert-butylhydroperoxide. Expression of yqhD was also associated with changes in the concentration of intracellu- lar peroxides and cytoplasmic protein carbonyl content and with a reduction in intracellular acrolein levels. When com- pared with the wild type strain, an yqhD mutant exhibited a sensitive phenotype to all these compounds and also augmented levels of thiobarbituric acid-reactive substances, which may indicate an increased level of lipid peroxidation. Purified YqhD catalyzes the in vitro reduction of acetaldehyde, malondial- dehyde, propanaldehyde, butanaldehyde, and acrolein in a NADPH-dependent reaction. Finally, yqhD transcription was induced in cells that had been exposed to conditions favoring lipid peroxidation. Taken together these results indicate that this enzyme may have a physiological function by protecting the cell against the toxic effect of aldehydes derived from lipid oxi- dation. We speculate that in Escherichia coli YqhD is part of a glutathione-independent, NADPH-dependent response mech- anism to lipid peroxidation. Oxidative stress is detrimental to a number of cellular mac- romolecules including proteins, nucleic acids, and membrane lipids. In eukaryotic organisms oxidative modification of cellu- lar constituents has been implicated in the etiology of different pathological conditions such as diabetes, cataracts, pulmonary emphysema, arthritis, cancer, and aging (1, 2). The electron-transport chain provides a constant supply of reactive oxygen species (ROS). 4 Hydrogen peroxide (H 2 O 2 ), superoxide (O 2 . ), and hydroxyl radical (OH ) are typical side products of the aerobic metabolism. ROS compounds are also formed during cell exposure to redox-cycling chemicals like paraquat or metals and metalloids like chromate and potassium tellurite (K 2 TeO 3 ) (3–5). To alleviate ROS-generated oxidative damage, Escherichia coli cells induce the synthesis of a variety of antioxidant enzymes including catalases and superoxide dis- mutases (3, 6). Except for mechanisms involved in membrane peroxidation, prokaryotic and eukaryotic cells use similar mechanisms to respond to oxidative stress. Lipid oxidation is a common con- sequence of the activity of free radicals on cell membrane com- ponents. Oxidation of polyunsaturated fatty acids by HO 2 , the protonated form of superoxide anion, or OH leads to the for- mation of lipid peroxides (7). Enzymatic or chemical degrada- tion of lipid peroxides results in the generation of toxic break- down products like short-chain (C 3 -C 9 ) aldehydes such as 2-alkenals (,-unsaturated aldehydes), 2-propenal (acrolein), 4-hydroxynonenal, and malondialdehyde (MDA) (7, 8). The toxicity of these “reactive aldehydes” lies in their ability to form Michael adducts with thiol and amino groups of proteins, thus affecting several cellular processes (9). For example, inactiva- tion of glucose-6-phosphate dehydrogenase, glyceraldehyde-3- phosphate dehydrogenase, and cytochrome c oxidase by reac- tive aldehydes that modify lysine and cysteine residues located in enzyme active sites has been reported (10 –12). Reactive aldehydes can also generate DNA adducts causing an important increase of the rate of mutagenesis in E. coli and Salmonella (13, 14). Mammalian cells use three distinct systems to cope with the harmful effect of reactive aldehydes. The glutathione S-trans- ferase pathway fueled by the GSH or thioredoxin redox cycle conjugates aldehydes with glutathione (GSH), the aldo-keto reductase system reduces aldehydes to alcohols, and alcohol and aldehyde dehydrogenases oxidize aldehydes to carboxyl groups. In animals, the main detoxification pathway is GSH-de- pendent, and it has been shown that the alcohol/aldehyde dehy- * This work received financial support from Fondo Nacional de Ciencia y Tec- nologia Grant 1060022 and by Dicyt (Universidad de Santiago de Chile) grants (to C. C. V.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 Recipient of a doctoral fellowship from Mejoramiento de la Calidad y la Equidad de la Educacion Superior (MECESUP). 2 Recipient of a doctoral fellowship from Consejo Nacional de Ciencia y Tecnologia. 3 To whom correspondence should be addressed: Laboratorio de Microbio- logı ´a Molecular, Departamento de Biologı ´a, Facultad de Quı ´mica y Biolo- gı ´a, Universidad de Santiago de Chile, Casilla 40, Correo 33, Santiago, Chile. Tel.: 56-2-681-0357; Fax: 56-2-681-2108; E-mail: [email protected]. 4 The abbreviations used are: ROS, reactive oxygen species; MDA, malondial- dehyde; t-BOOH, tert-butylhydroperoxide; TBAR, thiobarbituric acid-reac- tive substance; qRT, quantitative real time; Cp, crossing point; wt, wild type. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 12, pp. 7346 –7353, March 21, 2008 © 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 7346 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 12 • MARCH 21, 2008 at Univ Western Ontario-OCUL on March 14, 2008 www.jbc.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Escherichia coli YqhD Exhibits Aldehyde Reductase Activity and Protects from the Harmful Effect of...

Escherichia coli YqhD Exhibits Aldehyde Reductase Activityand Protects from the Harmful Effect of LipidPeroxidation-derived Aldehydes*
Received for publication, October 26, 2007, and in revised form, January 16, 2008 Published, JBC Papers in Press, January 22, 2008, DOI 10.1074/jbc.M708846200
Jose Manuel Perez1,2, Felipe A. Arenas2, Gonzalo A. Pradenas1, Juan M. Sandoval1, and Claudio C. Vasquez3
From the Laboratorio de Microbiologıa Molecular, Departamento de Biologıa, Facultad de Quımica y Biologıa,Universidad de Santiago de Chile, Santiago 9170019, Chile
Evidence that Escherichia coli YqhD is involved in bacterialresponse to compounds that generate membrane lipid peroxi-dation is presented.Overexpression of yqhD results in increasedresistance to the reactive oxygen species-generating compoundshydrogen peroxide, paraquat, chromate, and potassium tellu-rite. Increased tolerance was also observed for the lipid peroxi-dation-derived aldehydes butanaldehyde, propanaldehyde,acrolein, andmalondialdehyde and themembrane-peroxidizingcompound tert-butylhydroperoxide. Expression of yqhD wasalso associated with changes in the concentration of intracellu-lar peroxides and cytoplasmic protein carbonyl content andwith a reduction in intracellular acrolein levels. When com-pared with the wild type strain, an yqhD mutant exhibited asensitive phenotype to all these compounds and also augmentedlevels of thiobarbituric acid-reactive substances, which mayindicate an increased level of lipid peroxidation. Purified YqhDcatalyzes the in vitro reduction of acetaldehyde, malondial-dehyde, propanaldehyde, butanaldehyde, and acrolein in aNADPH-dependent reaction. Finally, yqhD transcription wasinduced in cells that had been exposed to conditions favoringlipid peroxidation. Taken together these results indicate thatthis enzymemay have a physiological function by protecting thecell against the toxic effect of aldehydes derived from lipid oxi-dation. We speculate that in Escherichia coli YqhD is part of aglutathione-independent, NADPH-dependent response mech-anism to lipid peroxidation.
Oxidative stress is detrimental to a number of cellular mac-romolecules including proteins, nucleic acids, and membranelipids. In eukaryotic organisms oxidative modification of cellu-lar constituents has been implicated in the etiology of different
pathological conditions such as diabetes, cataracts, pulmonaryemphysema, arthritis, cancer, and aging (1, 2).The electron-transport chain provides a constant supply of
reactive oxygen species (ROS).4 Hydrogen peroxide (H2O2),superoxide (O2
. ), and hydroxyl radical (OH�) are typical sideproducts of the aerobic metabolism. ROS compounds are alsoformed during cell exposure to redox-cycling chemicals likeparaquat ormetals andmetalloids like chromate and potassiumtellurite (K2TeO3) (3–5). To alleviate ROS-generated oxidativedamage,Escherichia coli cells induce the synthesis of a variety ofantioxidant enzymes including catalases and superoxide dis-mutases (3, 6).Except for mechanisms involved in membrane peroxidation,
prokaryotic and eukaryotic cells use similar mechanisms torespond to oxidative stress. Lipid oxidation is a common con-sequence of the activity of free radicals on cell membrane com-ponents. Oxidation of polyunsaturated fatty acids by HO2, theprotonated form of superoxide anion, or OH� leads to the for-mation of lipid peroxides (7). Enzymatic or chemical degrada-tion of lipid peroxides results in the generation of toxic break-down products like short-chain (C3-C9) aldehydes such as2-alkenals (�,�-unsaturated aldehydes), 2-propenal (acrolein),4-hydroxynonenal, and malondialdehyde (MDA) (7, 8). Thetoxicity of these “reactive aldehydes” lies in their ability to formMichael adducts with thiol and amino groups of proteins, thusaffecting several cellular processes (9). For example, inactiva-tion of glucose-6-phosphate dehydrogenase, glyceraldehyde-3-phosphate dehydrogenase, and cytochrome c oxidase by reac-tive aldehydes that modify lysine and cysteine residueslocated in enzyme active sites has been reported (10–12).Reactive aldehydes can also generate DNA adducts causingan important increase of the rate of mutagenesis in E. coliand Salmonella (13, 14).Mammalian cells use three distinct systems to cope with the
harmful effect of reactive aldehydes. The glutathione S-trans-ferase pathway fueled by the GSH or thioredoxin redox cycleconjugates aldehydes with glutathione (GSH), the aldo-ketoreductase system reduces aldehydes to alcohols, and alcoholand aldehyde dehydrogenases oxidize aldehydes to carboxylgroups. In animals, themain detoxification pathway isGSH-de-pendent, and it has been shown that the alcohol/aldehyde dehy-
* This work received financial support from Fondo Nacional de Ciencia y Tec-nologia Grant 1060022 and by Dicyt (Universidad de Santiago de Chile)grants (to C. C. V.). The costs of publication of this article were defrayed inpart by the payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
1 Recipient of a doctoral fellowship from Mejoramiento de la Calidad y laEquidad de la Educacion Superior (MECESUP).
2 Recipient of a doctoral fellowship from Consejo Nacional de Ciencia yTecnologia.
3 To whom correspondence should be addressed: Laboratorio de Microbio-logıa Molecular, Departamento de Biologıa, Facultad de Quımica y Biolo-gıa, Universidad de Santiago de Chile, Casilla 40, Correo 33, Santiago, Chile.Tel.: 56-2-681-0357; Fax: 56-2-681-2108; E-mail: [email protected].
4 The abbreviations used are: ROS, reactive oxygen species; MDA, malondial-dehyde; t-BOOH, tert-butylhydroperoxide; TBAR, thiobarbituric acid-reac-tive substance; qRT, quantitative real time; Cp, crossing point; wt, wildtype.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 12, pp. 7346 –7353, March 21, 2008© 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
7346 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 12 • MARCH 21, 2008
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

drogenase and glutathione S-transferase pathways account for10 and 55% of aldehyde elimination by hepatocytes, respec-tively (15).Because the major peroxidation substrate, polyunsaturated
fatty acid, is found at very low levels in bacterial membranes,peroxidation of prokaryotic membranes has been a matter ofcontroversy. Increasing experimental evidence suggesting thatlipid peroxidation is physiologically important in microorga-nisms has been reported during the last few years. Bacterialaccumulation of reactive aldehydes has been demonstrated incells exposed to tert-butylhydroperoxide (t-BOOH), hydrogenperoxide (H2O2), potassium tellurite (K2TeO3), and titaniumoxide (TiO2) (4, 16–18).
It has been proposed that members of the ubiquitous thiolperoxidase thiol-dependent antioxidant/AhpC family partici-pate in protecting E. coli against alkyl and lipid hydroperoxides(19) such as alkyl hydroperoxidase reductase, bacterioferrincomigratory protein, and periplasmic thiol peroxidase. How-ever, direct experimental evidence for specific defense systemsagainst lipid peroxidation-derived aldehydes in bacteria havenot been reported to date.Sulzenbacher et al. (20) determined the crystal structure of a
new E. coli dehydrogenase, YqhD, whose physiological sub-strates could not be determined. Based mainly on the boundcoenzyme, NADP(OH)2, they proposed a putative relationshipbetween YqhD and oxidative stress. Genomic and proteomicstudies have shown that YqhD is expressed in E. coli after expo-sure to stress conditions such as cold shock (21), nutrient starv-ing (22), and oxidative stress generated by the Curvularia per-oxidase system (23). YqhD has several features similar to AdhE,an alcohol/aldehyde dehydrogenase involved in fermentative
metabolism in E. coli for which an antioxidant function underaerobic conditions has been proposed (24).We have found that YqhD is induced under tellurite expo-
sure, suggesting that it is involved in bacterial response to oxi-dative stress. In this work we show that E. coli YqhD is an alde-hyde reductase involved in resistance to ROS-generatingcompounds and reactive aldehydes. Evidence to confirm thatYqhD catalyzes the NADPH-dependent reduction of variousmembrane peroxidation-derived aldehydes, including acroleinand MDA, is provided. Increased levels of thiobarbituric acid-reactive substances (TBARs) and acrolein are found in �yqhDcells as compared with the parental, isogenic, wild type straincells. Expression of YqhD resulted in diminished concentra-tions of cytoplasmic peroxides and oxidized proteins. This anti-oxidant-like effect was only observed in the presence of oxygen,a condition required for lipid peroxidation. Finally, we deter-mined that yqhD transcription was induced upon exposure toH2O2, K2TeO3, paraquat, t-BOOH, and low temperatures, con-ditions known to promote membrane lipid peroxidation.Results suggest that YqhD is part of a specific defense mech-
anism against reactive aldehydes generated by membrane per-oxidation in E. coli. To our knowledge, this is the first commu-nication describing that such mechanism might be used bybacteria to cope with the toxicity of membrane lipid peroxida-tion-derived aldehydes.
EXPERIMENTAL PROCEDURES
Strains and Plasmids—E. coli strains and plasmids used inthis study are listed in Table 1.E. coliBW25113�yqhD, harbor-ing a KanR insertion in the chromosomal yqhD gene, was kindlyprovided by the NARA Institute, Japan.
TABLE 1Bacterial strains, plasmids, and primers used in this study
Strain Relevant genotype Source or referenceE. coli TOP10 F� mcrA �(mrr-hsdRMS-mcrBC) �80lacZ�M15 �lacX74 deoR nupG recA1
araD139 �(ara-leu)7697 galU galK rpsL(StrR) endA1 ��Invitrogen�
E. coli BW25113 laclq rmBT14 �lacZWJ16 hsdRS14 �araBADAH33 �rhaBADLD78 Datsenko and Wanner (46)E. coli pBAD E. coli BW25113 harboring pBAD This workE. coli pBAD yqhD E. coli BW25113 harboring pBAD yqhD This workE. coli �yqhD E. coli BW25113 �yqhD (yqhD::KanR) NARA Institute, JapanE. coli �yqhD pBAD E. coli BW25113 �yqhD harboring pBAD This workE. coli �yqhD pBAD yqhD E. coli BW25113 �yqhD harboring pBAD yqhD This workE. coli pDEST17 yqhD E. coli BL21 (DE3) harboring pDest17 yqhD plasmid Sulzenbacher et al. (20)
Plasmids Plasmid featurespBAD TOPO Expression vector, ApR Invitrogen�pBAD yqhD pBAD harboring E. coli yqhD gene This workpGEMT Cloning vector, ApR PROMEGA�pGEMT yqhDp pGEMT harboring E. coli yqhD gene plus 300-bp upstream of yqhD
translation start codonThis work
pDEST17 yqhD pDest17 harboring E. coli yqhD gene. Adds 6X His to YqhD and allows isopropyl 1-thio-�-D-galactopyranoside-induced over expression of YqhD
Sulzenbacher et al. (20)
Primers Forward (F) or Reverse (R), to amplify 5�-3� SequenceyqhD F F, yqhD CCGGAAAAGAGAATTATGATGCCAGGCTCyqhD R R, yqhD AAGCTTTTAGCGGGCGGCTTCGTATATACyqhD Ra R, yqhD ACGCTGCCGCCGCCGTAGGTAATsodAFa F, sodA CCTGCCATCCCTGCCGTATGCTTAsodARa R, sodA TCGATAGCCGCTTTCAGGTCACCCsoxSFa F, soxS TTACAGGCGGTGGCGATAATCGCTsoxSRa R, soxS CGAGCATATTGACCAGCCGCTTAA16sFa F, 16 S RNA ATGACCAGCCACACTGGAAC16sRa R, 16 S RNA TGACTTAACAAACCGCCTGCgapAFa F, gapA GTAAAGTTGGTATTAACGGTTTTGGgapARa R, gapA GGTGATGTGTTTACGAGCAG
a Primers used in qRT-PCR experiments. Primers were designed to generate a 300-bp product.
YqhD Is an Aldehyde Reductase
MARCH 21, 2008 • VOLUME 283 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 7347
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

Cloning and Expression of yqhD and Protein Purification—Plasmid pBAD yqhD was constructed by cloning the PCR-am-plified E. coli yqhD gene fragment into the expression vectorpBAD TOPO (Invitrogen). E. coli BW25113 chromosomalDNA and primers described in Table 1 was used to amplify theyqhD gene. The PCR product was ligated to the cloning vector,and the resulting plasmid pBAD yqhD was introduced intoE. coli Top10 by electroporation. Insert orientation was con-firmed byNcoI digestion, and gene integrity was determined bynucleotide sequencing. Plasmid pBAD yqhD was then mobi-lized into E. coli BW25113 to carry out the resistance studiesreported in this work.E. coli BL21(DE3) cells harboring plasmid pDEST17 yqhD (a
generous gift of Dr. C. Cambillau, CNRS, Marseille, France)were used to purify YqhD. After 5 h of induction with isopropyl1-thio-�-D-galactopyranoside (1 mM), cells were harvested,resuspended in buffer 50mMTris-HCl, pH7.0, and subjected tosonic disruption. Nucleic acids were precipitated with strepto-mycin sulfate (2%, w/v) on ice. After centrifuging at 13,000 � gfor 5 min, the supernatant was loaded onto a Ni2� column(nickel-nitrilotriacetic acid). Proteins were eluted with 0.5 Mimidazole and fractionated by SDS-PAGE to estimate proteinpurity.Assay for YqhD Dehydrogenase—The reaction mixture con-
tained 50 mM potassium phosphate buffer, pH 7.0, 2 mMNADPH, a defined aldehyde (acetaldehyde, propanal, orbutanal), or alcohol (butanol, propanol or isopropanol) as sub-strate and purified YqhD (10 �g/ml). The mixture was incu-bated for 2 min at 37 °C, and NADPH oxidation was deter-mined at 340 nm using a UV-Visible Agilent 8453spectrophotometer. One unit of enzyme activity was defined asthe amount of protein that oxidizes 1 pmol of NADPH/min at37 °C.Minimal Inhibitory Concentrations—Cells from overnight
cultures were diluted 100-fold with fresh LBmedium. Ten�l ofthis dilutionwere used to inoculate 1ml of LBmediumcontain-ing the appropriate antibiotics (Kan and/or Amp) and the tox-ics to be tested. Cells were grown for 48 h at 37 °C with shaking.Peroxide Intracellular Levels—The oxidant-sensitive probe
2�,7�-dichlorodihydrofluorescein diacetate was used to meas-ure peroxide levels in cells exposed to potassium tellurite (0.5�g/ml), hydrogen peroxide (1 mM), paraquat (50 �g/ml), orchromate (1 mM) (24). Cells were grown under aerobic condi-tions in LBmedium amended with the compounds to be testedto an A600 � 0.3. After washing with 10 mM potassium phos-phate buffer, pH 7.0, cells were incubated in the same buffercontaining 10 �M 2�,7�-dichlorodihydrofluorescein diacetate(dissolved in dimethyl sulfoxide) for 30min. Cells were washed,suspended in the same buffer, and disrupted by sonication. Cellextracts (100�l) weremixedwith 1ml of phosphate buffer, andfluorescence intensity was measured using an Applied Biosys-tems Cytofluor 4000 multi-well plate reader (excitation, 490nm; emission, 519 nm). Fluorescence emission valueswere nor-malized to protein concentration.Protein Carbonylation—Protein carbonyl content was deter-
mined according to Semchyshyn et al. (18). Nucleic acid-freeextracts (100 �l) prepared from E. coli BW25113 cells treatedand untreated for 30 min with K2TeO3 (0.5 �g/ml) or H2O2
(100 �M) were mixed with four volumes of 10 mM dinitrophe-nylhydrazine (dissolved in 2 M HCl) and incubated for 1 h atroom temperature with vortexing every 10–15 min. Proteinswere precipitated by adding 500 �l of 20% trichloroacetic acidand sedimented at 14,000 � g for 5 min. The pellet was washedat least 3 times with 1:1 ethanol:ethyl acetate to remove anyunreacted dinitrophenylhydrazine. Finally, the pellet was dis-solved at 37 °C in 450 �l of 50 mM dithiothreitol in 6 M guani-dine HCl. Carbonyl content was determined spectrophoto-metrically at 370 nm using a molar absorption coefficient of22,000 M�1cm�1 (18).TBARs—TBARs were determined in cell extracts as
described by Semchyshyn et al. (18). Cells were grown at 37 °Cin LB media to an A600 � 0.6 and exposed for 30 min to aparticular toxic compound. Cell suspensions (1.0ml) were thenprecipitated by the addition of 1.0ml of 20% (w/v) trichloroace-tic acid and centrifuged, and supernatants were mixed with 2.0ml of a saturated solution of thiobarbituric acid dissolved in 0.1M HCl and 10 mM butylated hydroxytoluene. Samples wereheated at 100 °C for 60 min, and 1.5-ml aliquots were removed,chilled on ice, and mixed with 1.5 ml of n-butanol. After cen-trifugation at 4,000 � g for 10 min, the organic fraction wasremoved, and the A535 was measured. TBARs content wasdetermined using a molar absorption coefficient of 156 mM�1
cm�1 (18, 25).RNA Purification and Quantitative Real Time (qRT)-PCR—
RNA purification was carried out using the Qiagen RNeasy kit(Qiagen). E. coli BW25113 cultures were diluted 1:100, inocu-lated in 200 ml of LB medium, and incubated at 37 °C withshaking to anA600 � 0.6. Cultures were then amended with 0.5�g/ml K2TeO3, 0.1 mM H2O2, 50 �g/ml paraquat, or 0.05%t-BOOH and incubated for 30 min. Cells were sedimented at13,000 � g for 3 min and used for RNA purification.Two �l of total RNA (�1 �g/�l) from control and treated
cells were used in qRT-PCR experiments. Reactions (20 �l)used the LightCycler� RNA Amplification kit SYBR Green I(Roche Applied Science) and contained Lightcycler� 0.4 �l ofRT-PCR Enzyme Mix, 4 �l of SYBR Green I Reaction Mix, 3.2�l of 25 mMMgCl2, 5.4 �l of H2O PCR grade, 4 �l of resolutionsolution, and 0.5 pmol of each specific primer. Amplificationproducts were detected by increase of fluorescence. Crossingpoints (Cp) are inversely proportional to the RNA content andwere determined using the software included in LightCycler�2.0. Primers used to amplify yqhD are shown in Table 1. Induc-tion of gene expressionwas expressed as the difference betweenthe crossing points of each RT-PCR determination (Cp �Cptoxic). Positive numbers reflect higher amounts of the partic-ular, specific mRNA in cells exposed to the toxic condition.Acrolein Concentration—Acrolein concentration was deter-
mined as described by Slininger and Bothast (26). Acroleinreacts with tryptophan yielding a purple complex absorbing at560 nm. Commercially available acrolein (Sigma) was used tostandardize the assay and to make a calibration curve. “Trypto-phan solution” consisted of tryptophan (2.05 g), 4.17ml of 8.9 MHCl, and 2.5 ml of toluene in 1 liter of bi-distilled water.Approximately 300 �l of crude extracts (or acrolein standard)were mixed with 600 �l of 8.9 M HCl and 1.5 ml of tryptophansolution. Duplicated samples were incubated for 20 min at
YqhD Is an Aldehyde Reductase
7348 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 12 • MARCH 21, 2008
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

40 °C until a purple color developed and became stable. A560was determined spectrophotometrically.
RESULTS
YqhD Mediates Resistance to ROS Elicitors in E. coli—Todetermine whether YqhD plays a role in resistance to oxidativedamage, minimal inhibitory concentrations for different ROS-generating compounds for E. coli strains lacking (�yqhD) andoverexpressing the yqhD gene (pBAD yqhD) were determined(Table 2). Although cells overexpressing yqhD showed increasedtolerance to compounds whose toxicity involves superoxide gen-eration like paraquat and potassium tellurite (4, 27), the hydroxylradical-generating chromate anion (5), and hydrogen peroxide,E. coli�yqhD showed increased sensitivity to all these compounds(Table 2). To assess whether YqhD-mediated resistance pheno-typewasalsorelated tocytoplasmthioldepletion (especiallyGSH),the effect of YqhDonE. coli resistance to oxidant compounds thatinvolve thiol oxidation like diamide and cadmium chloride (28)was determined. All E. coli strains tested exhibited similar resist-ance levels to these compounds, confirming the idea that YqhDprotects E. coli from ROS-generated damage.YqhD Is a NADPH-dependent Aldehyde Reductase—YqhD
exhibits 40–50% sequence similarity with a number of bacterialalcohol dehydrogenases. However, it did not show any detect-able enzymatic activity when tested with short-chain alcoholsas substrates (20). In our hands, purified YqhD did not showNADP�-dependent dehydrogenase activity on methanol, eth-anol, propanol, butanol, or isopropanol (not shown). YqhDshowed enzymatic activity with short-chain aldehydes includ-ing a few with unsaturations or hydroxylations. Km valuesranged from 0.67 mM for butanaldehyde to 28.5 mM for acetal-dehyde. The kcat value was similar for all substrates (Table 3).yqhD Elimination Increases TBARs Levels in E. coli—TBARs
are routinely used to estimate malondialdehyde content. Theycan also indicate the presence of a series of reactive aldehydesthat aremainly generated during lipid peroxidation (17, 18, 25).We have previously shown that E. coli cells exposed to H2O2 orK2TeO3 have increased levels of TBARs (4). E. coli �yqhD
exhibited increased content of TBARs even when H2O2 orK2TeO3 were not added to the culture, suggesting that YqhDmay function in controlling the amount of membrane peroxi-dation products that are generated during the normal basalmetabolism. E. coli �yqhD showed 2- (H2O2) and 4-fold(K2TeO3) TBARs increases when compared with the wild typestrain, indicating that YqhD affects the content of reactive alde-hydes within the cell (Fig. 1).Elimination of yqhD Alters the Intracellular Level of
Peroxides—The oxidation-sensitive probe 2�,7�-dichlorodihy-drofluorescein diacetate was used to estimate the level of cyto-plasmic peroxides in E. coli pBAD yqhD and E. coli �yqhD.Probe activation was monitored in the presence or absence ofthe ROS-generating compounds K2TeO3, H2O2, paraquat, andchromate. E. coli cells lacking the yqhD gene showed an impor-tant increase in probe activation as comparedwith thewild typecounterpart even in the absence of any compound.Overexpres-sion of yqhD resulted in decreased peroxide levels in untreatedcells. Peroxide levels were similar in wild type and pBAD yqhDcells (Fig. 2).YqhD Protects E. coli from Protein Oxidation—Formation of
carbonyl groups in proteins, a consequence of ROS and reactivealdehydes, is a commonly used marker to estimate oxidativestress. Anti-dinitrophenylhydrazineWestern blotting was usedto estimate the extent of protein oxidation. Analysis of cyto-plasmic protein extracts revealed that protein carbonylation inE. coli �yqhD was slightly higher than that of wild type andE. coli pBAD yqhD even in unstressed cells (Fig. 3A). Theseresults agree with those obtained spectrophotometrically withcells grown aerobically in LB medium and exposed to K2TeO3or H2O2 (Fig. 3B). E. coli �yqhD showed increased protein oxi-dation even when grown in the absence of these compounds(Fig. 3B). Genetic complementation of the deletion mutantrestored protein carbonyl content to wild type levels, whereasoverexpression of yqhD resulted in decreased protein oxidationregardless the tested toxic.YqhD Alters Acrolein Concentration in E. coli—Acrolein is a
highly reactive, toxic aldehyde that has been described as a
FIGURE 1. Elimination of yqhD increases the level of lipid peroxidationproducts in E. coli. TBARs were determined as described under “Experimen-tal Procedures” in wild type E. coli (wt) and a mutant lacking yqhD (�yqhD) inthe absence (control) or in the presence of the ROS elicitors H2O2, and K2TeO3.
TABLE 2YqhD mediates resistance to potassium tellurite and other ROSelicitors in E. coliMinimal inhibitory concentrations of K2TeO3, paraquat, diamide, and CdCl2 (�g/ml) and H2O2 and K2CrO7 (mM) were determined as described under “Experimen-tal Procedures.” Numbers are the mean of three independent determinations.
Strain K2TeO3 H2O2 Paraquat K2CrO7 CdCl2 DiamideE. coli pBAD 1.25 5 300 1.25 250 350E. coli pBAD yqhD 24 10 1000 5 250 350E. coli �yqhD 0.3 1.2 100 0.15 250 350
TABLE 3YqhD is a NADPH-dependent aldehyde reductaseEnzyme activity (�mol of NADPH/min mg prot) was determined as describedunder “Experimental Procedures.” Each determination corresponds at least to threeindependent trials with each substrate concentration analyzed.
Substrate Km kcat kcat/Km
mM s�1 s�1 M�1
Acetaldehyde 28.47 53.5 1880Propanaldehyde 3.31 45.06 13613Butanaldehyde 0.67 59.5 88018Acrolein 4.81 62.5 12994Malondialdehyde 1.78 60.1 33764
YqhD Is an Aldehyde Reductase
MARCH 21, 2008 • VOLUME 283 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 7349
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

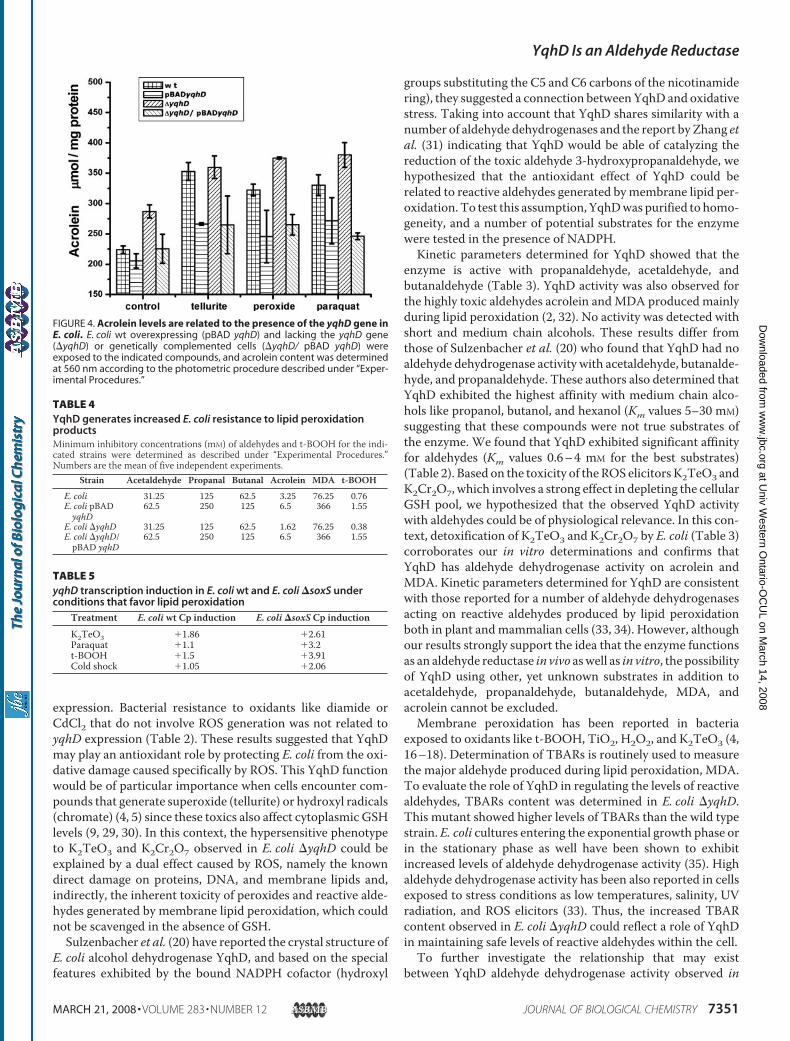
membrane peroxidation product in eukaryotic cells. Tellurite-,hydrogen peroxide-, or paraquat-mediated stress resulted inincreased concentrations of acrolein in E. coli (Fig. 4). Acroleincontent was proportional to the amount of K2TeO3, H2O2, ormethyl viologen used to stress the cells (not shown). E. coli�yqhD showed increased levels of acrolein even in the absenceof toxics, whereas yqhD overexpression prevented the ROS-mediated increase of acrolein concentration in E. coli. Geneticcomplementation of�yqhD cells restored thewild type acroleinphenotype (Fig. 4).Overexpression of yqhD Slightly Increases E. coli Resistance to
Aldehydes Generated during Lipid Peroxidation—To assess thephysiological relevance of YqhD in vivo, E. coli resistance toaldehydes generated duringmembrane peroxidationwas deter-mined. E. coli resistance to acetaldehyde, propanal, butanal,and acrolein doubled as result of yqhD overexpression. A
�4-fold increase in resistance to MDA, the main peroxidationproduct, was also observed. Although E. coli �yqhD and thewild type strain showed identical minimal inhibitory concen-trations for acetaldehyde, propanal, butanal, and MDA, E. coli�yqhD was slightly more sensitive to acrolein (Table 4).The effect of yqhD expression on E. coli resistance to the
classic membrane-peroxidizing compound t-BOOH was alsodetermined. The hypersensitive phenotype to t-BOOH exhib-ited byE. coli�yqhDwas reverted by genetic complementation.Overexpression of yqhD resulted in a significant increase int-BOOH resistance (Table 4), again suggesting that YqhD isinvolved in E. coli defense against membrane lipidperoxidation.Transcription of yqhD Is Induced in Response to Conditions
That Favor Lipid Peroxidation—Induction of yqhD transcrip-tion in response to the oxidative stress elicitors K2TeO3 andparaquat was evaluated by qRT-PCR. Cp were determined fortreated and untreated cells. A “Cp induction” value was definedas the difference between the Cp values obtained in each con-dition (see under “Experimental Procedures” for details).Increased amounts of yqhD mRNA were evidenced in cellsexposed to individual toxics when compared with unexposedwild type cells (Table 5). Cp values for yqhDmRNAwas �1–2-fold higher in treated cells than those observed in untreatedcells, reflecting an important difference in yqhDmRNAcontentbetween both conditions. The housekeeping gene gapA encod-ing glyceraldehyde-3-phosphate dehydrogenase showed aslight decrease in mRNA content in treated cells (not shown).The yqhD Promoter Contains a Putative SoxS Binding Site—
A putative SoxS binding site (CGTAATTTCTTAGGAATA)sharing 76% sequence identity with a SoxS binding sequencepreviously found inE. coliwas identified 172 bp upstreamof theyqhDATG start codon (Promoter prediction program). Signif-icant induction of yqhDmRNAsynthesiswas observed inE. colilacking SoxS (Table 5), suggesting that induction of yqhD tran-scription is rather SoxS-independent.
DISCUSSION
Our interests in bacterial resistance to potassium telluritelead us to investigate the synthesisof YqhD in E. coli grown in tellurite-amended media. We found thatYqhD was highly induced whencells were grown in the presence ofK2TeO3. This result and the factthat an important pathway of tellu-rite toxicity involves superoxidegeneration (4, 27) prompted us toevaluate the role that YqhD couldhave in E. coli resistance to telluriteand other oxidative stress elicitors.To assess whether YqhD exhibits
a general antioxidant function or arather specific effect on ROS-caused oxidation, resistance to sev-eral oxidant agents was evaluated.E. coli resistance to ROS elicitorswas directly related with yqhD
FIGURE 2. Elimination of yqhD results in increased intracellular peroxidelevels in E. coli. E. coli wt overexpressing (pBAD yqhD) and lacking the yqhDgene (�yqhD) were exposed to the indicated compounds, and intracellularperoxide levels were determined by the 2�,7�-dichlorodihydrofluoresceindiacetate procedure described under “Experimental Procedures.” AU, arbi-trary units.
FIGURE 3. Oxidized proteins in E. coli. A, SDS-PAGE showing cytoplasmic protein oxidation in the indicatedE. coli strains, as revealed by anti-dinitrophenylhydrazine antibodies. B, assessment of carbonyl protein con-tent in extracts of the indicated E. coli strains exposed to potassium tellurite or hydrogen peroxide, as deter-mined by the spectrophotometric procedure described under “Experimental Procedures.”
YqhD Is an Aldehyde Reductase
7350 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 12 • MARCH 21, 2008
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from
expression. Bacterial resistance to oxidants like diamide orCdCl2 that do not involve ROS generation was not related toyqhD expression (Table 2). These results suggested that YqhDmay play an antioxidant role by protecting E. coli from the oxi-dative damage caused specifically by ROS. This YqhD functionwould be of particular importance when cells encounter com-pounds that generate superoxide (tellurite) or hydroxyl radicals(chromate) (4, 5) since these toxics also affect cytoplasmic GSHlevels (9, 29, 30). In this context, the hypersensitive phenotypeto K2TeO3 and K2Cr2O7 observed in E. coli �yqhD could beexplained by a dual effect caused by ROS, namely the knowndirect damage on proteins, DNA, and membrane lipids and,indirectly, the inherent toxicity of peroxides and reactive alde-hydes generated by membrane lipid peroxidation, which couldnot be scavenged in the absence of GSH.Sulzenbacher et al. (20) have reported the crystal structure of
E. coli alcohol dehydrogenase YqhD, and based on the specialfeatures exhibited by the bound NADPH cofactor (hydroxyl
groups substituting the C5 and C6 carbons of the nicotinamidering), they suggested a connection betweenYqhD and oxidativestress. Taking into account that YqhD shares similarity with anumber of aldehyde dehydrogenases and the report by Zhang etal. (31) indicating that YqhD would be able of catalyzing thereduction of the toxic aldehyde 3-hydroxypropanaldehyde, wehypothesized that the antioxidant effect of YqhD could berelated to reactive aldehydes generated bymembrane lipid per-oxidation. To test this assumption, YqhDwas purified to homo-geneity, and a number of potential substrates for the enzymewere tested in the presence of NADPH.Kinetic parameters determined for YqhD showed that the
enzyme is active with propanaldehyde, acetaldehyde, andbutanaldehyde (Table 3). YqhD activity was also observed forthe highly toxic aldehydes acrolein andMDA produced mainlyduring lipid peroxidation (2, 32). No activity was detected withshort and medium chain alcohols. These results differ fromthose of Sulzenbacher et al. (20) who found that YqhD had noaldehyde dehydrogenase activity with acetaldehyde, butanalde-hyde, and propanaldehyde. These authors also determined thatYqhD exhibited the highest affinity with medium chain alco-hols like propanol, butanol, and hexanol (Km values 5–30 mM)suggesting that these compounds were not true substrates ofthe enzyme. We found that YqhD exhibited significant affinityfor aldehydes (Km values 0.6–4 mM for the best substrates)(Table 2). Based on the toxicity of the ROS elicitors K2TeO3 andK2Cr2O7, which involves a strong effect in depleting the cellularGSH pool, we hypothesized that the observed YqhD activitywith aldehydes could be of physiological relevance. In this con-text, detoxification of K2TeO3 and K2Cr2O7 by E. coli (Table 3)corroborates our in vitro determinations and confirms thatYqhD has aldehyde dehydrogenase activity on acrolein andMDA. Kinetic parameters determined for YqhD are consistentwith those reported for a number of aldehyde dehydrogenasesacting on reactive aldehydes produced by lipid peroxidationboth in plant andmammalian cells (33, 34). However, althoughour results strongly support the idea that the enzyme functionsas an aldehyde reductase in vivo aswell as in vitro, the possibilityof YqhD using other, yet unknown substrates in addition toacetaldehyde, propanaldehyde, butanaldehyde, MDA, andacrolein cannot be excluded.Membrane peroxidation has been reported in bacteria
exposed to oxidants like t-BOOH, TiO2, H2O2, and K2TeO3 (4,16–18). Determination of TBARs is routinely used to measurethe major aldehyde produced during lipid peroxidation, MDA.To evaluate the role of YqhD in regulating the levels of reactivealdehydes, TBARs content was determined in E. coli �yqhD.This mutant showed higher levels of TBARs than the wild typestrain. E. coli cultures entering the exponential growth phase orin the stationary phase as well have been shown to exhibitincreased levels of aldehyde dehydrogenase activity (35). Highaldehyde dehydrogenase activity has been also reported in cellsexposed to stress conditions as low temperatures, salinity, UVradiation, and ROS elicitors (33). Thus, the increased TBARcontent observed in E. coli �yqhD could reflect a role of YqhDin maintaining safe levels of reactive aldehydes within the cell.To further investigate the relationship that may exist
between YqhD aldehyde dehydrogenase activity observed in
FIGURE 4. Acrolein levels are related to the presence of the yqhD gene inE. coli. E. coli wt overexpressing (pBAD yqhD) and lacking the yqhD gene(�yqhD) or genetically complemented cells (�yqhD/ pBAD yqhD) wereexposed to the indicated compounds, and acrolein content was determinedat 560 nm according to the photometric procedure described under “Exper-imental Procedures.”
TABLE 4YqhD generates increased E. coli resistance to lipid peroxidationproductsMinimum inhibitory concentrations (mM) of aldehydes and t-BOOH for the indi-cated strains were determined as described under “Experimental Procedures.”Numbers are the mean of five independent experiments.
Strain Acetaldehyde Propanal Butanal Acrolein MDA t-BOOH
E. coli 31.25 125 62.5 3.25 76.25 0.76E. coli pBADyqhD
62.5 250 125 6.5 366 1.55
E. coli �yqhD 31.25 125 62.5 1.62 76.25 0.38E. coli �yqhD/pBAD yqhD
62.5 250 125 6.5 366 1.55
TABLE 5yqhD transcription induction in E. coli wt and E. coli �soxS underconditions that favor lipid peroxidation
Treatment E. coli wt Cp induction E. coli �soxS Cp inductionK2TeO3 �1.86 �2.61Paraquat �1.1 �3.2t-BOOH �1.5 �3.91Cold shock �1.05 �2.06
YqhD Is an Aldehyde Reductase
MARCH 21, 2008 • VOLUME 283 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 7351
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

vitro and the level of reactive aldehydes generated by lipid per-oxidation in vivo, we determined acrolein concentration in cellsexposed to various ROS elicitors. Acrolein, or 2-propenal, is ahighly toxic aldehyde generated as a byproduct during mem-brane peroxidation (32). Although acrolein generation has beenscantily described in bacteria to date, its lethal effect for pro-karyotes has been clearly demonstrated (13, 14).We found thatE. coli exposed to H2O2, paraquat, K2Cr2O7, and K2TeO3exhibited increased acrolein concentration (Fig. 4). Asexpected, cells that were exposed to low temperatures, a condi-tion that increases membrane polyunsaturated fatty acid con-tent (36), showed increased acrolein content (not shown) link-ing oxidative stress with acrolein production in E. coli. Theseresults confirm that the aldehyde reductase activity of YqhDobserved in vitro is probably related to the in vivo concentrationof YqhD substrates.Lipid hydroperoxides are intermediates of membrane lipid
peroxidation that can generate reactive aldehydes (7, 8). Lipidhydroperoxides exhibit increased polarity and long lifetimes ascomparedwith free radical intermediates, thus facilitating theirinteraction with intracellular targets (37). To cope with thishighly reactive molecules, cells produce catalases, peroxidases,and/or glutathione transferases that transform peroxides inmore stable molecules or unreactive adducts (19). We foundthat expression of yqhD directly affects intracellular peroxidelevels in E. coli exposed to H2O2, K2TeO3, K2Cr2O7, or para-quat. Cells incapable of producing yqhD showed an increasedconcentration of intracellular peroxides suggesting that YqhDplays an important role in detoxifying reactive aldehydes thatcould increase intracellular peroxide generation or affect cellu-lar defenses. This idea is consistent with the observation thatcells overexpressing yqhD exhibited decreased levels of intra-cellular peroxides.Another common marker used to assess oxidative stress is
carbonyl group formation in proteins. This results from aminoacid side chain modification preferentially caused by ROS andreactive aldehydes (18, 38). Because E. coli �yqhD containsincreased levels of reactive aldehydes, determined as TBARsand acrolein, we hypothesized that the protective role of YqhDwould be to prevent aldehyde-induced protein oxidation. Wefound that E. coli-defective in yqhD showed higher levels ofoxidized proteins than wild type cells, suggesting that YqhD issomehow involved in protecting the cell from cytoplasmic oxi-dation and from oxidants generated under stress and also dur-ing the basal metabolism.The respiratory electron transport chain is amajor (and con-
tinuous) source of ROS in bacteria (3, 19). The “naturally gen-erated” reactive oxygen species would be responsible for theincreased levels of TBARs and oxidized protein found in E. coli�yqhD. Both phenotypes are reverted to wild type levels upongenetic complementation with yqhD. In addition, overexpres-sion of yqhD resulted in decreased protein oxidation (Fig. 3B).This protective effect takes place in normally growing cells andis particularly high in cells subjected to conditions that favorlipid peroxidation. Activation of oxidative stress-inducible pro-moters by potassium tellurite supports this assumption. Recentresults from our laboratory have shown that tellurite-mediatedinduction of the ibpA gene promoter, a gene related with oxi-
dative response (39, 40), is blocked by yqhD overexpression.Similar findingswere not observedwith other stress-responsivepromoters not related to oxidative stress as cspA, p3RpoH, andsulA.5
Because in this work yqhD expression has been related toROS resistance, TBARs, acrolein, peroxide, and especially oxi-dized protein levels, we hypothesized that the role of YqhDaldehyde reductase is to protect cytoplasmic macromoleculesfrom aldehyde-mediated oxidation. E. coli resistance to lipidperoxidation-generated aldehydes revealed a correlationbetween yqhD expression and bacterial resistance, especially toacrolein (Table 4). These results are in agreement with theobserved effect of yqhD on acrolein concentration in vivo aswell as with enzyme activity exhibited by the purified protein.As proof that YqhD substrates come frommembrane lipid per-oxidation, we evaluated E. coli t-BOOH resistance. AlthoughyqhD overexpression resulted in increased resistance, cellslacking yqhD showed a hypersensitive phenotype to thisperoxide.Because the lipid peroxidation pathway includes an elonga-
tion step in which oxygen reacts with a carbonyl radical to gen-erate a lipoperoxide radical (41), we carried out K2TeO3 andH2O2 resistance analyses in E. coli under anaerobic conditions.No differences in resistance to K2TeO3 and H2O2 wereobserved between wt and yqhD-overexpressing cells in theabsence of oxygen. TBAR determination suggested that lipidperoxidation does not occur in this condition, indicating thattoxic YqhD substrates are not generated (unpublished results).5
Transcription activation of the yqhD gene has been observedin cells treatedwith the oxidantCurvularia haloperoxidase sys-tem (23) when exposed to cold shock (21) or when grown in alimited-nutrient medium (22). Here we demonstrate that yqhDis also induced in cells exposed to conditions favoring lipid per-oxidation, as are treatments with the ROS elicitors K2TeO3,H2O2, or paraquat or with the membrane lipid-peroxidatingcompound t-BOOH.A highly conserved SoxS binding motif was identified in the
promoter region of the yqhD gene (not shown). qRT-PCR stud-ies carried out using RNA from wt and �soxS E. coli strainssuggested that yqhD induction is not dependent on SoxS. How-ever, because gene activationmay result from the action of var-ious different transcriptional activators, this result does not ruleout the functionality of the identified SoxS binding site.In humans, a number of enzymes using cytotoxic reactive
aldehydes as substrates have been described. As YqhD, theseenzymes react with acrolein and MDA among others and arethought to form part of oxidative defense mechanisms able toneutralize the toxic effects of these lipid peroxidation-derivedreactive aldehydes in heart (42), kidney (43), brain (44), andarterial tissues (45).Somemechanistic insights have been communicated inAra-
bidopsis thaliana where a NADPH quinone oxidoreductase(P1-ZCR) has been involved in the detoxification of lipid per-oxidation-derived reactive aldehydes (33). The authors pro-posed that this proteinwas part of a defensemechanism against
5 J. Manuel Perez, F. A. Arenas, G. A. Pradenas, J. M. Sandoval, and C. C.Vasquez, unpublished results.
YqhD Is an Aldehyde Reductase
7352 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 12 • MARCH 21, 2008
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from

oxidative stress by scavenging highly toxic lipid peroxidation-derived unsaturated aldehydes as acrolein and 4-hydroxynon-enal. In addition, a NADPH-dependent AKR1C1 protein,which is induced byMichael acceptors and oxidative stress andthat exhibits aldehyde reductase activity involved in cellularprotection against reactive aldehydes, has been reported inhumans (34).To date no specific defense pathway dealing with lipid per-
oxidation-derived reactive aldehydes has been reported in bac-teria. The results of this work allow us to speculate that YqhD ispart of a response mechanism against lipid peroxidation-de-rived aldehydes. The role we propose for YqhD is especiallyrelevant for E. coli cells that are stressed by ROS elicitors and bycompounds that produceGSHoxidation. Asmentioned before,the first cell defense mechanism coping with lipid peroxidationproducts is GSH-dependent and consists in a set of glutathioneS-transferases, catalases, and peroxidases. In cells exposed toK2TeO3 or K2Cr2O7, two compounds that deplete the intra-cellular GSH pool and generate ROS-mediated lipid peroxi-dation, this first defense mechanism would be absentbecause of glutathione oxidation. In this context, the GSH-independent, NADPH-dependent YqhD putative mechanismdescribed here would be highly relevant in protecting the bac-terial cell from being damaged by MDA, acrolein, and otheraldehydes. In the absence of GSH, YqhD and probably other yetunknown aldehyde dehydrogenases or aldehyde reductaseswould act as cytosolic barriers against toxic products generatedduring lipid peroxidation in E. coli.
Acknowledgments—We are indebted to Dr. James A. Imlay and toDr. Sergio E. Pichuantes for helpful suggestions and critical read-ing of the manuscript.
REFERENCES1. Mates, J. M. (2000) Toxicology 153, 83–1042. Halliwell, B., and Gutteridge, J. (1999) Free Radicals in Biology and Med-
icine, 3rd Ed., Oxford University Press, NY3. Imlay, J. (2003) Annu. Rev. Microbiol. 57, 395–4184. Perez, J. M., Calderon, I. L., Arenas, F. A., Fuentes, D. E., Pradenas, G. A.,
Fuentes, E. L., Sandoval, J. M., Castro, M. E., Elıas, A. O., and Vasquez,C. C. (2007) Plos One 2, e211
5. Itoh, M., Nakamura, M., Suzuki, T., Hawai, K., Horitsu, H., and Taka-mizawa, K. (1995) J. Biochem. 117, 780–786
6. Fridovich, I. (1995) Annu. Rev. Biochem. 64, 97–1127. Esterbauer, H., Zollner, H., and Schaur, R. (1990) in Membrane Lipid
Oxidation (Vigo-Pelfrey, C., ed) Vol. 1, pp. 239–268, CRCPress, Inc., BocaRaton, FL
8. Esterbauer, H. (1982) in Free Radicals, Lipid Peroxidation, and Cancer(McBrien, D. C. H., and Slater, T. F., eds) pp. 101, Academia Press Ltd,London
9. Esterbauer, H., Zollner, H., and Scholz, N. (1975) Z. Naturforsch. 30,466–473
10. Szweda, L., Uchida, K., and Stadtman, R. (1993) J. Biol. Chem. 268,3342–3347
11. Uchida, K., and Stadtman, E. (1993) J. Biol. Chem. 268, 6288–639312. Chen, J., Schenker, S., Frosto, T., and Henderson, G. (1998) Biochim. Bio-
phys. Acta 1380, 336–34413. Yang, I., Hossain, M., Miller, H., Khullar, S., Johnson, F., Grollam, A., and
Moriya, M. (2001) J. Biol. Chem. 276, 9071–9076
14. Neudecker, T., Eder, E., Deininger, C., and Henschler, C. (1991) Mutat.Res. 264, 193–196
15. Hartley, D., Ruth, J., and Petersen, D. (1995) Arch. Biochem. Biophys. 316,197–205
16. Yoon, S., Park, J., Yang, J., and Park, J. (2002) J. Biochem. Mol. Biol. 35,297–301
17. Maness, P., Smolinski, S. Z., Blake, D., Huang, Z.,Wolfrum, E., and Jacoby,W. (1999) Appl. Environ. Microbiol. 65, 4094–4098
18. Semchyshyn, H., Bagnyukoba, T., Storey, K., and Lushchak, V. (2005)CellBiol. Int. 29, 898–902
19. Cha, M. K., Kim, H. K., and Kim, I. H. (2004) J. Bacteriol. 178, 5610–561420. Sulzenbacher, G., Alvarez, K., van den Heuvel, R., Versluis, C., Spinelli, S.,
Campanacci, V., Valencia, C., Cambillau, C., Eklund, H., and Tegoni, M.(2004) J. Mol. Biol. 342, 489–502
21. Phadtare, S., and Inouye, M. (2004) J. Bacteriol. 186, 7007–701422. Hua, Q., Yang, C., Oshima, T., Mori, H., and Shimizu, K. (2004) Appl.
Environ. Microbiol. 70, 2354–236623. Hansen, E., Schembri, M., Klemm, P., Schafer, T., Molin, S., and Gram, L.
(2004) Appl. Environ. Microbiol. 70, 1749–175724. Echave, P., Tamarit, J., Cabiscol, E., and Ros, J. (2003) J. Biol. Chem. 32,
30193–3019825. Rice-Evans, C., Diplock, A., and Symins, M. (1991) in Laboratory tech-
niques in Biochemistry andMolecular Biology (Buerdon, R., and van Knip-penberg, P.H., eds), Vol. 22, pp. 147–149, Elsevier Science Publishers B.V.,Amsterdam
26. Slininger, P. J., and Bothast, R. J. (1985) Appl. Environ. Microbiol. 50,1444–1450
27. Calderon, I. L., Arenas, F. A., Perez, J. M., Fuentes, D. E., Araya, M. A.,Saavedra, C. P., Tantalean, J. C., Pichuantes, S. E., Youderian, P. A., andVasquez, C. C. (2006) Plos ONE 1, e70
28. Wang, A., and Crowley, D. (2005) J. Bacteriol. 187, 3259–326629. Turner, R.,Weiner, J., andTaylor, D. (1999)Microbiology 145, 2549–255730. Ackerley, D., Barak, Y., Lynch, V., Curtin, J., and Matin, A. (2006) J. Bac-
teriol. 188, 3371–338131. Zhang, X., Tang, X., Zhuge, B., Shen, W., Rao, Z., Fang, H., and Zhuge, J.
(2005) Sheng Wu Gong Cheng Xue Bao 21, 743–74732. Uchida, K., Kanematsu,M., Sakai, K.,Matsuda, T., Hattori, N.,Mizuno, Y.,
Suzuki, D., Miyata, T., Noguch, N., Nikii, E., and Osawa, T. (1998) Proc.Natl. Acad. Sci. U. S. A. 95, 4882–4887
33. Mano, J., Torii, Y., Hayashi, S., Takimoto, K., Matsui, K., Nakamura, K.,Inze, D., Babiychuk, E., Kushnir, S., and Osada, K. (2002) Plant CellPhysiol. 43, 1445–1455
34. Burczynski, M., Sridhar, G., Palackal, N., and Pennings, M. (2001) J. Biol.Chem. 276, 2890–2897
35. Ho, K. K., and Weiner, H. (2005) J. Bacteriol. 187, 1067–107336. Denich, T., Beaudette, L., Lee, H., and Trevors, J. (2003) J. Microbiol.
Methods 52, 149–18237. Girotti, A. (1998) J. Lipid Res. 39, 1529–154238. Refsgaard, H., Tsai, l., and Stadtman, L. (2000) Proc. Natl. Acad. Sci.
U. S. A. 97, 611–61639. Shapiro, E., and Baneyx, F. (2002) Antimicrob. Agents Chemother. 46,
2490–249640. Kitagawa, M., Matsumura, Y., and Tsuchido, T. (2000) FEMS Microbiol.
Lett. 184, 165–17141. Porter, N. (1984)Methods Enzymol. 105, 273–28242. Srivastava, S., Chandra, A., Ansari, N. H., Srivastava, S. K., and Bhatnagar,
A. (1998) Biochem. J. 329, 469–47543. Zu, X., Yan, R., Robbins, S., Krishack, P. A., Liao, D. F., and Cao, D. (2007)
Toxicol. Sci. 97, 562–56844. Picklo, M. J., Olson, S. J., Markesbery, W. R., and Montine, T. J. (2001)
J. Neuropathol. Exp. Neurol. 60, 686–69545. Rittner, H. L., Hafner, V., Klimiuk, P. A., Szweda, L. I., Goronzy, J. J., and
Weyand, C. M. (1999) J. Clin. Investig. 103, 1007–1001346. Datsenko, K. A., andWanner, B. L. (2000) Proc. Natl. Acad. Sci. U. S. A. 97,
6640–6645
YqhD Is an Aldehyde Reductase
MARCH 21, 2008 • VOLUME 283 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 7353
at Univ W
estern Ontario-O
CU
L on March 14, 2008
ww
w.jbc.org
Dow
nloaded from




















