
A new aglaspidid arthropod from the Lower Ordovician of Wales

261
Copyright � 2000, SEPM (Society for Sedimentary Geology) 0883-1351/00/0015-0261/$3.00
Epizoan-sponge Interactions in the Early Ordovician ofthe Argentine Precordillera
MARCELO G. CARRERACatedra de Estratigrafıa y Geologıa Historica, Facultad de Ciencias Exactas, Fısicas y Naturales,
Universidad Nacional de Cordoba, Av. Velez Sarsfield 299 (5000) Cordoba, Argentina
PALAIOS, 2000, V. 15, p. 261–272
Evidence for biotic interaction is not obtainable easily in thefossil record. Epizoans cemented to the skeletons of host or-ganisms provide an opportunity to study possible host–epi-zoan interactions and the ecological structure of hard sub-stratum communities. One such possibility is sponges andtheir associated encrusters, but few such occurrences havebeen documented from the fossil record. Early Ordovicianlimestones of the San Juan Formation in the Argentine Pre-cordillera bear an important sponge fauna consistingmainly of demosponges. This Early Llanvirn assemblagehas the highest diversity, and most sponges were subjectedto epizoan encrustations. A total of nineteen species includ-ed in eleven genera of sponges were examined to record epi-zoans encrustations. Type and relative abundance of epi-zoans were compiled for each sponge species. Five kinds ofepizoans were recorded: bryozoans, stemmed echinoderms,brachiopods, other sponges, and scars and borings of un-determined organisms. Crinoids and blastozoans are wide-spread common epizoans on sponges. Small brachiopodsare attached to sponges showing no location preference;rarely, juvenile sponges appear on conspecific adults. Sinu-ous scars and worm-like organisms are found in almost allthe sponges. Deep borings, like those found in Devonian oryounger sponges, are absent. The analysis of epizoan distri-bution on each sponge species shows a fairly homogeneousdistribution of echinoderms on sponges but host specificityof bryozoans. Here, surface texture seems to have been a sig-nificant controlling factor. Almost all the trepostome bry-ozoa adhere to Rhopalocoelia tenuis Carrera. This taxon isa cylindrical sponge whose smooth surface texture con-trasts sharply with the rough and open textured spiculationobserved among other species in the fauna. This featureseems to have been a key factor for the mutual relationship.Rhopalocoelia tenuis shows a reaction to epizoans, modi-fying the shape of growth and changing the direction of thespicular structure in the vicinity of the epibiont, providingevidence that encrustation occurred while the sponge wasalive.
Diversity, composition and encrusting strategies of theArgentine epizoan community reflect the early evolutionarystage of this biotic relationships in the Early Paleozoic. Thiscontrasts with the more varied and diversified relationshipsamong organisms that were able to encrust, bore, and chem-ically perforate hard parts after the Devonian. The Early toMiddle Ordovician was a time of rapid evolutionary diver-sification among hard substrate communities. The Argen-tinean epizoan community appears to be transitional be-tween the first hard substrate encrusters (Late Cambrian),
which were eocrinoid dominated and were not accompaniedby macroborers or bryozoans, and the later Paleozoic com-munities, in which bryozoans were the most prolific utiliz-ers of attachment and macroborers were common.
INTRODUCTION
Modern sponges have served as substrates and shelterhabitats for many organisms. Cited benefits include sup-port, baffling against wave and other current action, pre-dation, and enhanced food supply by the generation of wa-ter currents (Frith, 1976; Bergquist, 1978; Villamizar andLaughlin, 1992 and references therein). Such associationsalso can be documented for fossil sponges due to the factthat many epizoan-encrusted stony sponges have highpreservation potential. Because evidence for biotic inter-action is not obtainable easily in the fossil record (Teveszand McCall, 1983; Alvarez and Taylor, 1987), sponges pro-vide unusual opportunities for observing the autecology ofhosts and epizoans (e.g., Spjeldnaes, 1984; Alvarez andTaylor, 1987) and the ecological structure of hard sub-strate communities (e.g., Taylor 1984; Liddell and Brett,1982).
Although, extensive studies have been carried out onhard substrate communities (see references in: Palmer1982; Tevesz and Mc Call, 1983; Wilson and Palmer, 1992;Wilson et al.,1992), few examples of sponges and their epi-bionts have been documented from the fossil record.Clarke (1908) described calcareous worm tubes aroundthe osculum of Devonian sponges from New York. Ziegler(1964) and Aldinger (1968) reported the occurrence ofbryozoans, serpulids, and brachiopods attached to Juras-sic and Cretaceous sponges. Gundrum (1979) described ademosponge assemblage with a dense population of at-tached organisms from Carboniferous rocks of NorthAmerica, and Kershaw (1980) reported bryozoans, corals,crinoids, and Trypanites borings on a stromatoporoid as-semblage from the Silurian of Gotland. Palmer and Fur-sich (1981) described a varied encrusting fauna on largeJurassic sponges mainly composed by bivalves, serpulids,and borings made by worms. A diverse fauna of sponges,bryozoans, and algae attached one to each other formingsuccessive incrustations as cryptobionts in the PermianCapitan reef (Wood et al., 1994).
Early Ordovician limestones of the San Juan Formationin the Argentine Precordillera (Fig. 1) bear an importantsponge fauna composed mainly of demosponges (Beresiand Rigby, 1993; Carrera, 1994 a, b, 1996 a, b; Carrera andRigby, 1999). Paleoecological aspects of the sponges andassociated faunas have been studied by Sanchez et al.
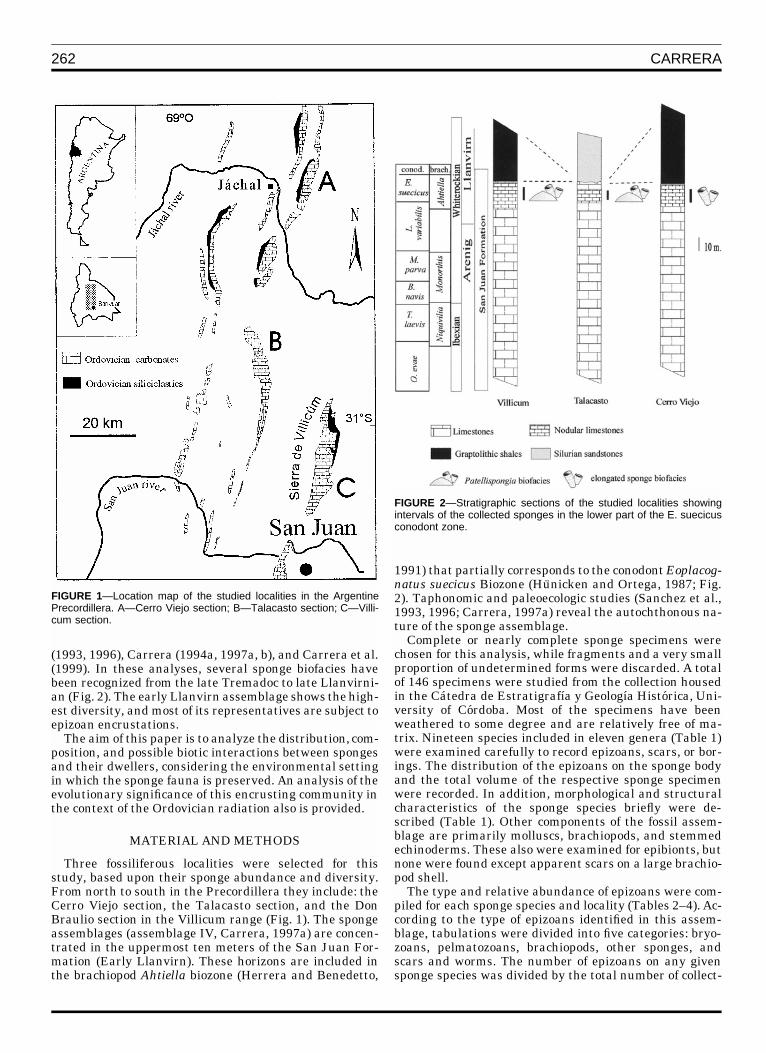
262 CARRERA
FIGURE 1—Location map of the studied localities in the ArgentinePrecordillera. A—Cerro Viejo section; B—Talacasto section; C—Villi-cum section.
FIGURE 2—Stratigraphic sections of the studied localities showingintervals of the collected sponges in the lower part of the E. suecicusconodont zone.
(1993, 1996), Carrera (1994a, 1997a, b), and Carrera et al.(1999). In these analyses, several sponge biofacies havebeen recognized from the late Tremadoc to late Llanvirni-an (Fig. 2). The early Llanvirn assemblage shows the high-est diversity, and most of its representatives are subject toepizoan encrustations.
The aim of this paper is to analyze the distribution, com-position, and possible biotic interactions between spongesand their dwellers, considering the environmental settingin which the sponge fauna is preserved. An analysis of theevolutionary significance of this encrusting community inthe context of the Ordovician radiation also is provided.
MATERIAL AND METHODS
Three fossiliferous localities were selected for thisstudy, based upon their sponge abundance and diversity.From north to south in the Precordillera they include: theCerro Viejo section, the Talacasto section, and the DonBraulio section in the Villicum range (Fig. 1). The spongeassemblages (assemblage IV, Carrera, 1997a) are concen-trated in the uppermost ten meters of the San Juan For-mation (Early Llanvirn). These horizons are included inthe brachiopod Ahtiella biozone (Herrera and Benedetto,
1991) that partially corresponds to the conodont Eoplacog-natus suecicus Biozone (Hunicken and Ortega, 1987; Fig.2). Taphonomic and paleoecologic studies (Sanchez et al.,1993, 1996; Carrera, 1997a) reveal the autochthonous na-ture of the sponge assemblage.
Complete or nearly complete sponge specimens werechosen for this analysis, while fragments and a very smallproportion of undetermined forms were discarded. A totalof 146 specimens were studied from the collection housedin the Catedra de Estratigrafıa y Geologıa Historica, Uni-versity of Cordoba. Most of the specimens have beenweathered to some degree and are relatively free of ma-trix. Nineteen species included in eleven genera (Table 1)were examined carefully to record epizoans, scars, or bor-ings. The distribution of the epizoans on the sponge bodyand the total volume of the respective sponge specimenwere recorded. In addition, morphological and structuralcharacteristics of the sponge species briefly were de-scribed (Table 1). Other components of the fossil assem-blage are primarily molluscs, brachiopods, and stemmedechinoderms. These also were examined for epibionts, butnone were found except apparent scars on a large brachio-pod shell.
The type and relative abundance of epizoans were com-piled for each sponge species and locality (Tables 2–4). Ac-cording to the type of epizoans identified in this assem-blage, tabulations were divided into five categories: bryo-zoans, pelmatozoans, brachiopods, other sponges, andscars and worms. The number of epizoans on any givensponge species was divided by the total number of collect-

EPIZOAN-SPONGE INTERACTIONS IN THE EARLY ORDOVICIAN 263
TABLE 1—Structural and morphological features of the Argentine Ordovician sponges.
Sponges Main morphologic features
Allosacus sp. Small discoidal sponge with reduced central cavity and large radial canals. Av-erage volume: 19 mm3
Archaeoscyphia minganensis Billings Long, erect, externally weakly annulate sponge with large and deep centralcavity and conpicuous radial canals. Average volume: 12 mm3
Archaeoscyphia pulchra Bassler Long, erect, strongly annulate sponge with large and deep central cavity andnumerous radial canals. Average volume: 16 mm3
Calicocoelia sp. Cup shaped, externally smooth sponge with deep central cavity and numerousradial canals. Average volume: 29 mm3
Hudsonospongia cyclostoma Raymond &Okulitch
Hudsonospongia irregularis Raymond &Okulitch
Cup shaped to piriform sponges with reduced and digitated central cavity. Av-erage volume: 32 mm3
Hudsonospongia porosa Raymond & Okulitch Globose to subglobose sponge without central cavity. Large openings all overthe surface. Average volume: 60 mm3
Patellispongia occulata BasslerPatellispongia robusta Beresi & RigbyPatellispongia argentina CarreraPatellispongia sp.
Large discoidal sponges with conspicuous radial canals and openings all overthe surface, without central cavity. Average volume: 453 mm3
Psarodictyum sp. Large discoidal sponge with conspicuos radial canals small openings in thesurface, without central cavity. Average volume: 140 mm3
Rhopalocoelia clarkii Raymond & Okulitch Long, erect and robust sponge with deep central cavity and dichotomous radialcanals. Average volume: 120 mm3
Rhopalocoelia regularis Raymond & Okulitch Small cylindrical sponge with smooth external surface open textured skeletonwith small spicules. Average volume: 14 mm3
Rhopalocoelia tenuis Carrera Small, slender, commonly digitate sponge with smooth external surface andfine espiculation, deep central cavity. Average volume: 8 mm3
Root-tufts Long root-tufts with giant monaxonic spicules. Average volume: 58 mm3
ed individuals of that species, to obtain relative abun-dance.
PALEOENVIRONMENTAL SETTING
The localities selected for this analysis have been sub-ject to sedimentological (Canas, 1995a, b, 1999; Carreraand Astini, 1997) and paleoecological studies (Sanchez etal., 1993, 1996; Carrera 1994a, 1997a). General sedimen-tological features of the sponge bearing limestones indi-cate a shallow subtidal environment in a middle-to-distalshelf ramp setting. A detailed study of biofacies of the SanJuan Formation was carried out by Carrera et al.(1999) in-cluding the sponge interval considered here, which wasplaced into the demosponge biofacies. Two of Carrera�s(1997a) different sponge biofacies are represented in theselected localities of the present study. The Patellispongiabiofacies occurs in the Talacasto and Villicum sections,and the ‘‘elongated sponge’’ biofacies is recorded in theCerro Viejo section. The former has been included in amiddle ramp setting sporadically affected by storm-wavebase. The huge discoidal forms of Patellispongia reflecttheir adaptation to fluctuating wave energy conditionsand imply the presence of soft or shifting substrates (San-chez et al., 1996; Carrera, 1997a). The Cerro Viejo assem-blage is represented by a high percentage of elongatedsponges indicative of low energy conditions. This locality isincluded in a middle-to-distal ramp setting rarely affectedby storm waves. In addition, Canas (1995 a, 1999) de-scribed these upper levels in the Cerro Viejo section asnodular wackestones and marlstones, having been depos-ited in more distal conditions.
ENCRUSTING EPIZOANS
Pelmatozoans
Crinoids and blastozoans are the most abundant andvariable epizoans on sponges. They are represented by dif-ferent kinds of holdfasts, from simple button-shapedstructures attached directly on small pits to large disc andrhizoid attachments (see Franzen, 1977 or Brett, 1981, forterminology). Holdfast attachments can be divided follow-ing the terminology of Brett (1981) into three main cate-gories:
(1) Simple discoidal holdfasts represent well over the80% of the epizoans, encrusting indistinctly erect or dis-coidal, small or large sponges. The general appearance isof a single solid disk, commonly with a mound or flatshape, that is circular in outline. Such attachment disksmay have entire, lobate or digitate outer margins, but theylack well-defined radicles (Figs. 3F, 4A, 4D). Both crinoidsand blastozoans have simple discoidal holdfasts.However,solid, mound-shaped holdfasts can be included in blasto-zoan attachments (T. Guensburg, pers. comm., 1999).
(2) Compound discoidal holdfasts are very similar ingeneral form to the simple discoidal holdfasts, but theyconsist of a cemented basal disc with septa-like ridgesoverlain by a mound-shape covering or integument. Thisintegument may be composed of discrete polygonal plates(Fig. 3B). They are considered to represent crinoid attach-ments.
(3) Radicular holdfast are very rare and occur mainly ondiscoidal sponges. They are terminal holdfast bearingwell-developed radicles (Fig. 4B, 4C).

264 CARRERA
TABLE 2—Epizoan distribution on sponges in the Villicum section. Numbers of epizoans per sponge species and, in brackets, average ofepizoan encrustation per sponge specimen.
Sponges (n� specimens) Bryozoans Pelmatozoans Brachiopods Sponges Scars—worms
Archaeoscyphia minganensis 5Archaeoscyphia pulchra 7Calicocoelia sp. 3Hudsonospongia cyclostoma 13Patellispongia robusta 4Patellispongia sp. 4
1 (0.2)—
1 (0.3)1 (00.7)
——
3 (0.6)7 (1)4 (1.3)
24 (1.8)—
10 (2.5)
1 (0.2)1 (0.1)—
2 (0.1)12 (3)
—
———
1 (00.7)—
3 (0.7)
8 (1.6)8 (1.1)4 (1.3)
20 (1.5)—
Psarodictyum sp. 11Rhopalocoelia clarkii 1Rhopalocoelia regularis 2Rhopalocoelia tenuis 1Root-tufts 4
—————
39 (3.5)1 91)3 (1.5)1 (1)6 (1.5)
4 (0.3)—
1 (0.5)—
1 (0.2)
—1 (1)
———
8 (0.7)
5 (2.5)—
1 (0.2)
‘ TABLE 3—Epizoan distribution on sponges in the Talacasto section. Numbers of epizoans per sponge species and, in brackets, average ofepizoan encrustation per sponge specimen.
Sponges (n� specimens) Bryozoans Pelmatozoans Brachiopods Sponges Scars—worms
Archaeoscyphia pulchra 13Hudsonospongia cyclostoma 5Patellispongia occulata 7Patellispongia robusta 1
——
2 (0.3)—
25 (2)12 (2.4)27 (3.8)
—
2 (0.1)2 (0.4)4 (0.5)6 (6)
—1 (0.2)
——
1 (0.07)1 (0.2)4 (0.5)
—Patellispongia argentina 6Rhopalocoelia clarkii 6Rhopalocoelia tenuis 6
6 (1)2 (0.3)
11 (1.8)
34 (5.6)7 (1.1)
17 (2.8)
1 (0.1)7 (1.1)2 (0.3)
——
1 (0.1)
——
1 (0.1)
Bryozoans
Two types of encrusting bryozoans were recognized. Themost abundant is a trepostome bryozoan that usuallyshows a laminar base and an incipient erect zoarium (Fig.3E). Some of these specimens resemble the immature partof the ramose trepostomes recorded in the matrix of thesame horizons. A very rare encrusting form also wasfound, mainly associated with the large discoidal spongePatellispongia (Fig. 3G). This encrusting form resemblesthe Berenicea-type bryozoan of Mergl (1983), Palmer andFursich (1974), and Wilson and Palmer (1992).
Brachiopods
Few brachiopods have been found on the sponge surfac-es. Some sponges bear the imprints of brachiopod shells ontheir skeletons (Fig. 3D). The only recognizable formsfound on sponges are: Platystrophia and Orthidium onroot-tufts, an indeterminate orthid on Archaeoscyphia andPetroria sp., and unidentified leptellinids on Psarodictyum(J.L.Benedetto, pers. comm., 1999).
Sponges
Although it is not a frequent occurrence, juvenile speci-mens can be seen on some conspecific sponges. In othercases, however, a sponge can overgrow different spongetaxa (root-tufts on Patellispongia).
Undetermined organisms
This category includes scars and worm-like organisms.Small scars and shallow pits probably are associated with
former crinoid attachments based upon remains found insome scars. Burrows or other kinds of deep perforationsare absent on the surface of the sponges, even in trans-verse thin sections of the sponge skeletons. The worm-likeorganisms are represented by small, and sinuous tubes. Inrelief, they appear as hollow cylinders that slowly taper toone end (Fig. 3A). They are very similar to serpulid tubesfound attached on Mesozoic sponges (Ziegler, 1974; Fur-sich and Werner, 1991) and also on hardgrounds (Wilsonand Palmer, 1992). Palmer (1982) recognised serpulid-type worms (‘‘spirorbid’’) recorded on hardground surfacesback to the Devonian, and Spjeldnaes (1984) reported thepresence of serpulid worms encrusting Silurian brachio-pods of Gotland.
EPIZOAN DISTRIBUTION
Virtually all the sponge specimens analysed bear somekind of epibiont. Epibionts are distributed all over thesponge body, suggesting that sponges were alive, or atleast in life position, during epibiont recruitment. In addi-tion, some sponge specimens show a change in their skel-eton growth as a consequence of epizoan encrustations, afeature that is discussed later. Epizoan numbers and dis-tribution on each sponge species (Tables 2–4, Fig. 5) showa fairly homogeneous encrustation of pelmatozoans and aclear selection by trepostome bryozoans for recruitment onRhopalocoelia tenuis. The latter is the only mutual rela-tionship with significant statistical value (Fig. 5). Rhopal-ocoelia tenuis occurs in the Talacasto and Cerro Viejo sec-tions, but has very few representatives in the Villicum sec-tion. The average number of bryozoan encrustations perspecimen is 2.7, with a maximum of eight bryozoans on a

EPIZOAN-SPONGE INTERACTIONS IN THE EARLY ORDOVICIAN 265
TABLE 4—Epizoan distribution on sponges in the Cerro Viejo section. Numbers of epizoans per sponge species and, in brackets, average ofepizoan encrustation per sponge specimen.
Sponges (n� specimens) Bryozoans Pelmatozoans Brachiopods Sponges Scars—worms
Allosacus sp. 4Aulocopium sp. 1Archaeoscyphia minganensis 4Archaeoscyphia pulchra 12Calicocoelia sp. 1
—————
3 (0.7)1 (1)
11 (2.7)18 (1.5)4 (4)
2 (0.5)—
1 (0.2)——
—————
———
6 (0.5)—
Hudsonospongia cyclostoma 10Hudsonospongia porosa 1Hudsonospongia irregularis 3Rhopalocoelia tenuis 7Psarodictyum sp. 1
———
8 (1.1)—
37 (3.7)4 (4)
10 (3.3)27 (3.8)4 (4)
1 (0.1)——
4 (0.5)—
—————
5 (0.5)————
Protachileum kayseri 1Rugospongia viejoensis 1Root-tufts 4Globose sponge indet. 6
————
3 (3)2 (2)
18 (4.5)13 (2.1)
—1 (1)
——
————
———
3 (0.5)
single sponge. In spite of the problems of preservation andsmall size, features that normally prevent taxonomic anal-ysis, these bryozoans all seem to be of the same species.
A high proportion of ramose trepostomes occurs in theTalacasto section. Fragments of these bryozoans appearedembedded in matrix, while small trepostome encrusta-tions occur on Rhopalocoelia tenuis in the same levels. Inthe Cerro Viejo section, ramose bryozoans in the matrixare very scarce, while R. tenuis, as in the Talacasto sec-tion, is crowded with trepostome epibionts. Two small andfragmentary specimens of R. tenuis were collected fromthe Villicum section, where they are devoid of bryozoanencrustations. Trepostomes are absent in the sediment ofthis section, but occur scarcely as small encrustations onArchaeoscyphia minganensis, Hudsonospongia cyclosto-ma, and Calicocoelia.
Bryozoan epibionts are restricted to four other sponges:Patellispongia argentina, Patellispongia cf. P. occulata,Rhopalocoelia clarkii, and Archaeoscyphia minganensis.Bryozoan occurrence on Patellispongia argentina is nearlyone epibiont per specimen on average. The other spongesshow average values on the order of 0.2 to 0.3 epibionts persponge. The relatively high percentage of bryozoans on P.argentina is due to the added occurrence of a different taxa(Fig 5), tentatively identified as a cyclostome (Fig. 3G; the‘‘Benericea type’’ of Palmer and Fursich, 1974, and Wilsonand Palmer, 1992).
Stemmed echinoderms appear on almost all sponges,and they constitute the most frequent epizoans. They alsoshow some preferences; for example, they appear mostcommonly on the different species of the discoidal spongesPatellispongia and Psarodyctium (see Tables 2–4 and Fig.5). However, they also encrust species of Archaeoscyphia,Hudsonospongia, and Rhopalocoelia. The only exception isthe highly porous Patellispongia robusta that apparentlyonly bears brachiopods in their surface (Fig. 5). In the Cer-ro Viejo section, where Patellispongia and Psarodictyumare absent, pelmatozoan encrustations increase on theother sponges. There is not a clear distribution of stemmedechinoderm holdfasts among sponges; but, in general, ra-dicular holdfasts are found commonly on flat discoidalsponges, while simple and compound holdfasts are foundindistinctly in all kind of sponges.
Epizoans on Patellispongia are distributed all over the
exterior surface but they are absent in the wide gastralsurfaces. Patellispongia shows different morphologies,from discoidal and laminar to wide conical forms. In thefield, all of these morphologies are found with the base ofthe cone down (Fig. 3C). Several epibionts, primarily echi-noderm attachments, were found on this external surface,suggesting that it was the real life position of this sponge.Wilson et al.(1992) found the similar encrusting mode oflife of Patellispongia on early Ordovician hardgrounds ofwestern North America.
SPONGE-EPIZOAN INTERACTIONS
Neontologists generally restrict usage of the term ‘‘epi-zoan’’ to associations in which the host was alive when en-crusted. The encrusting organisms attached to the Argen-tinean sponges show several indications of having beentrue epizoans. Sponges were subject to encrustationthroughout the sponge body, suggesting that at least somesponges were upright when the epizoans attached. How-ever, the most important feature shown by this associationis a reaction of the sponge against encrustation. Somesponges show halted skeletal production around a crinoidholdfast (Fig. 4E), and in the particular case of the bryo-zoans on Ropalocoelia tenuis, the sponge skeleton bendsoutward producing small domal elevations isolating theinitial bryozoan attachment (Figs.4B; 4E-G). These smallelevations clearly are distinguishable due to the otherwisesmooth surface of R. tenuis and their protruding skeletalnetwork. These features, and the fact that there are noother mounds or elevations along the rest of the spongewithout the bryozoan epibiont on top, suggest a reaction ofthe live sponge layer rather than the occasional settle-ment of the bryozoan on top of a previous mound on thesponge surface.
Reactions of sponges to their encrusters have not beenidentified previously with certainty in the fossil record(Gundrum, 1979), and very few examples have been de-scribed concerning other fossil-bearing epizoans. Chatter-ton (1975) recognized a response in the skeletal growth ofa brachiopod clearly related to the presence of an epizoan,as well as crinoids frequently reacted to the presence of en-crusters (Brett and Liddell, 1978). In modern sponges re-generation of damaged dermal layers is widespread

266 CARRERA
FIGURE 3—Different epibionts attachments on Argentine Ordovician sponges. (A) Worm-like tube attached to the surface of the discoidalPsarodictyum, and a compound crinoidal holdfast attached to the sponge surface (scale bar: 5 mm). (B) Multiplated crinoid holdfast attachedto the surface of Patellispongia. (scale bar: 10 mm). (C) The encrusting sponge Patellispongia covers important subtrate areas in the Talacastosection. These sponges commonly are found with epizoan attachments. (D) Mold and a fragment of a brachiopod shell cutting the spicular netof the sponge Psarodictyum (scale bar: 10 mm). (E) A large bryozoan attachment on Rhopalocoelia tenuis (scale bar: 5 mm). (F) Blastozoanholdfast attached to Psarodictyum. It overgrows the spicular mesh (scale bar: 5 mm). (G) Berenicea-type bryozoan on top of Patellispongiaargentina (scale bar: 5 mm).
(Storr, 1976; Jackson and Palumbi, 1979; Ayling, 1981;Woodley et al., 1981; Palumbi and Jackson, 1982). Somesponges are able to repair damaged areas at rates of 0.03–0.04 cm2 per day, depending on the diameter of the sponge
and the border of the damaged area (Ayling, 1981). Theyalso can begin rapid production of stiffer and strongerskeletons in high wave energy environments. Palumbi(1984) recorded a change in sponge skeletons within four
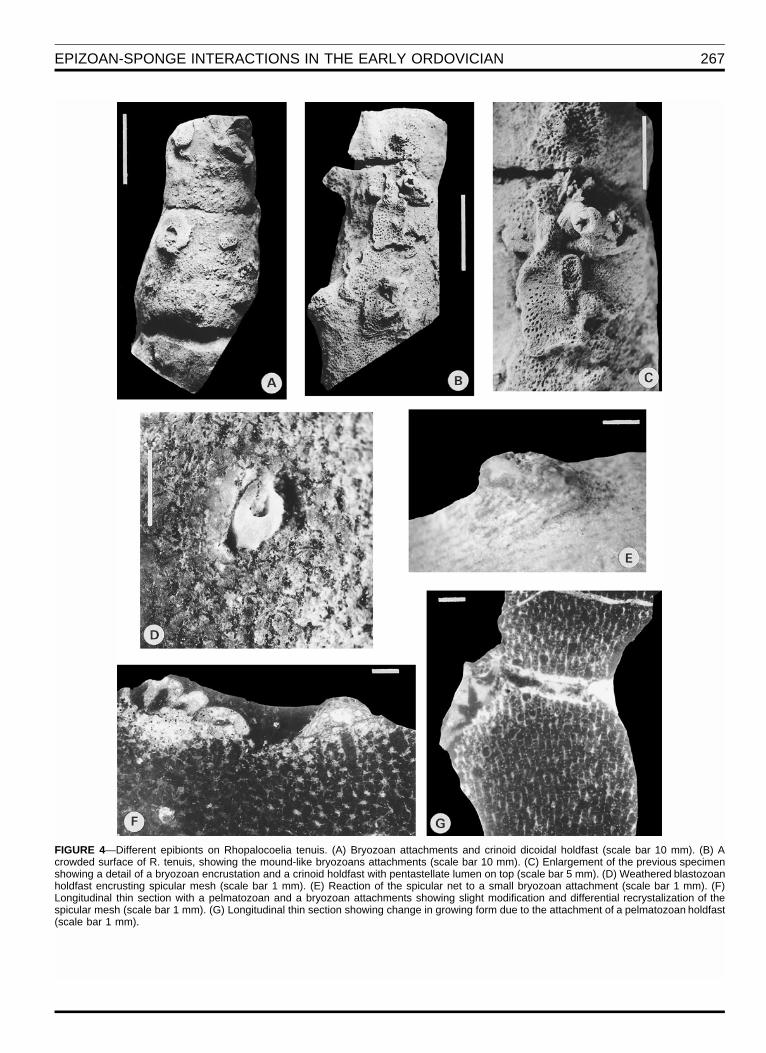
EPIZOAN-SPONGE INTERACTIONS IN THE EARLY ORDOVICIAN 267
FIGURE 4—Different epibionts on Rhopalocoelia tenuis. (A) Bryozoan attachments and crinoid dicoidal holdfast (scale bar 10 mm). (B) Acrowded surface of R. tenuis, showing the mound-like bryozoans attachments (scale bar 10 mm). (C) Enlargement of the previous specimenshowing a detail of a bryozoan encrustation and a crinoid holdfast with pentastellate lumen on top (scale bar 5 mm). (D) Weathered blastozoanholdfast encrusting spicular mesh (scale bar 1 mm). (E) Reaction of the spicular net to a small bryozoan attachment (scale bar 1 mm). (F)Longitudinal thin section with a pelmatozoan and a bryozoan attachments showing slight modification and differential recrystalization of thespicular mesh (scale bar 1 mm). (G) Longitudinal thin section showing change in growing form due to the attachment of a pelmatozoan holdfast(scale bar 1 mm).
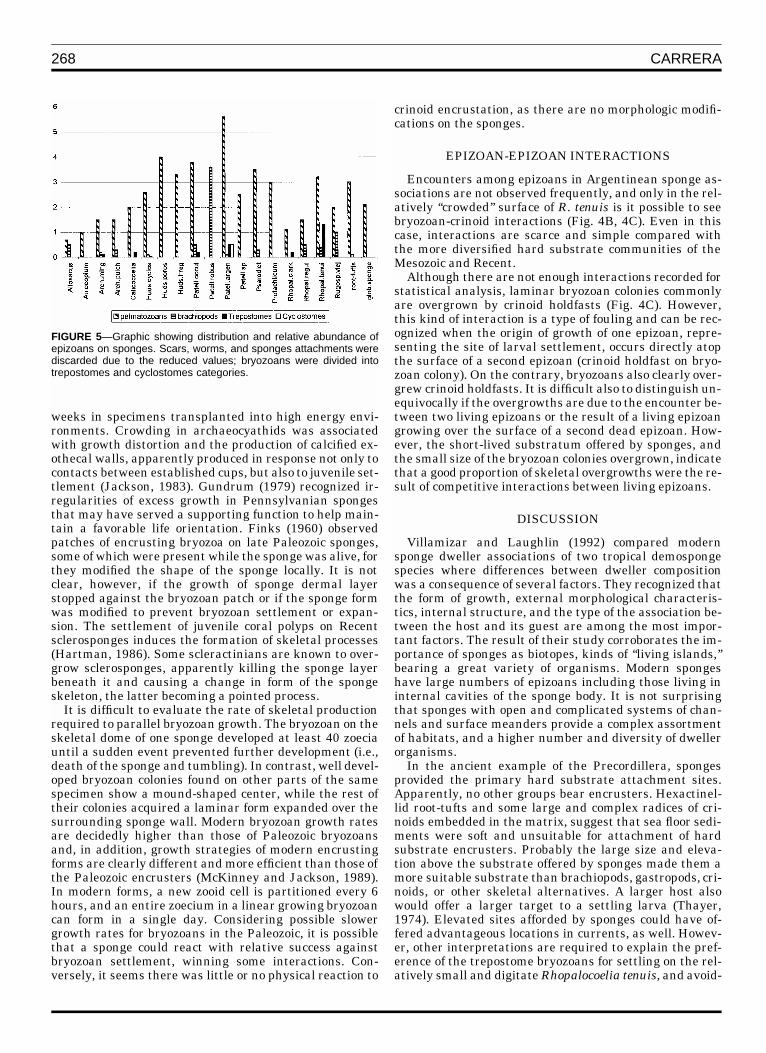
268 CARRERA
FIGURE 5—Graphic showing distribution and relative abundance ofepizoans on sponges. Scars, worms, and sponges attachments werediscarded due to the reduced values; bryozoans were divided intotrepostomes and cyclostomes categories.
weeks in specimens transplanted into high energy envi-ronments. Crowding in archaeocyathids was associatedwith growth distortion and the production of calcified ex-othecal walls, apparently produced in response not only tocontacts between established cups, but also to juvenile set-tlement (Jackson, 1983). Gundrum (1979) recognized ir-regularities of excess growth in Pennsylvanian spongesthat may have served a supporting function to help main-tain a favorable life orientation. Finks (1960) observedpatches of encrusting bryozoa on late Paleozoic sponges,some of which were present while the sponge was alive, forthey modified the shape of the sponge locally. It is notclear, however, if the growth of sponge dermal layerstopped against the bryozoan patch or if the sponge formwas modified to prevent bryozoan settlement or expan-sion. The settlement of juvenile coral polyps on Recentsclerosponges induces the formation of skeletal processes(Hartman, 1986). Some scleractinians are known to over-grow sclerosponges, apparently killing the sponge layerbeneath it and causing a change in form of the spongeskeleton, the latter becoming a pointed process.
It is difficult to evaluate the rate of skeletal productionrequired to parallel bryozoan growth. The bryozoan on theskeletal dome of one sponge developed at least 40 zoeciauntil a sudden event prevented further development (i.e.,death of the sponge and tumbling). In contrast, well devel-oped bryozoan colonies found on other parts of the samespecimen show a mound-shaped center, while the rest oftheir colonies acquired a laminar form expanded over thesurrounding sponge wall. Modern bryozoan growth ratesare decidedly higher than those of Paleozoic bryozoansand, in addition, growth strategies of modern encrustingforms are clearly different and more efficient than those ofthe Paleozoic encrusters (McKinney and Jackson, 1989).In modern forms, a new zooid cell is partitioned every 6hours, and an entire zoecium in a linear growing bryozoancan form in a single day. Considering possible slowergrowth rates for bryozoans in the Paleozoic, it is possiblethat a sponge could react with relative success againstbryozoan settlement, winning some interactions. Con-versely, it seems there was little or no physical reaction to
crinoid encrustation, as there are no morphologic modifi-cations on the sponges.
EPIZOAN-EPIZOAN INTERACTIONS
Encounters among epizoans in Argentinean sponge as-sociations are not observed frequently, and only in the rel-atively ‘‘crowded’’ surface of R. tenuis is it possible to seebryozoan-crinoid interactions (Fig. 4B, 4C). Even in thiscase, interactions are scarce and simple compared withthe more diversified hard substrate communities of theMesozoic and Recent.
Although there are not enough interactions recorded forstatistical analysis, laminar bryozoan colonies commonlyare overgrown by crinoid holdfasts (Fig. 4C). However,this kind of interaction is a type of fouling and can be rec-ognized when the origin of growth of one epizoan, repre-senting the site of larval settlement, occurs directly atopthe surface of a second epizoan (crinoid holdfast on bryo-zoan colony). On the contrary, bryozoans also clearly over-grew crinoid holdfasts. It is difficult also to distinguish un-equivocally if the overgrowths are due to the encounter be-tween two living epizoans or the result of a living epizoangrowing over the surface of a second dead epizoan. How-ever, the short-lived substratum offered by sponges, andthe small size of the bryozoan colonies overgrown, indicatethat a good proportion of skeletal overgrowths were the re-sult of competitive interactions between living epizoans.
DISCUSSION
Villamizar and Laughlin (1992) compared modernsponge dweller associations of two tropical demospongespecies where differences between dweller compositionwas a consequence of several factors. They recognized thatthe form of growth, external morphological characteris-tics, internal structure, and the type of the association be-tween the host and its guest are among the most impor-tant factors. The result of their study corroborates the im-portance of sponges as biotopes, kinds of ‘‘living islands,’’bearing a great variety of organisms. Modern spongeshave large numbers of epizoans including those living ininternal cavities of the sponge body. It is not surprisingthat sponges with open and complicated systems of chan-nels and surface meanders provide a complex assortmentof habitats, and a higher number and diversity of dwellerorganisms.
In the ancient example of the Precordillera, spongesprovided the primary hard substrate attachment sites.Apparently, no other groups bear encrusters. Hexactinel-lid root-tufts and some large and complex radices of cri-noids embedded in the matrix, suggest that sea floor sedi-ments were soft and unsuitable for attachment of hardsubstrate encrusters. Probably the large size and eleva-tion above the substrate offered by sponges made them amore suitable substrate than brachiopods, gastropods, cri-noids, or other skeletal alternatives. A larger host alsowould offer a larger target to a settling larva (Thayer,1974). Elevated sites afforded by sponges could have of-fered advantageous locations in currents, as well. Howev-er, other interpretations are required to explain the pref-erence of the trepostome bryozoans for settling on the rel-atively small and digitate Rhopalocoelia tenuis, and avoid-

EPIZOAN-SPONGE INTERACTIONS IN THE EARLY ORDOVICIAN 269
ing settlement on the large, discoidal Patellispongia or therobust and erect Archaeoscyphia, Hudsonospongia or Cal-icocoelia.
Substrate specificity or preference for a particular hostappears to be a fairly common phenomenon in the fossilrecord (Jackson, 1983; Conway Morris 1990). However,fossil epibionts restricted to a particular species or genusare not reported frequently. Dense monospecific associa-tions are strongly suggestive in themselves of preferentiallarval settlement on adults of the same species. Taxon spe-cific epibionts (see Jackson, 1983 and references therein)suggest a substrate preference by larval selection and set-tlement.
Bryozoan larvae are known to respond to direction andintensity of light, gravity, temperature, chemical sub-stances, physical characteristics of substrata, and waterflow (McKinney and McKinney, 1993). Larvae appear torespond to a physicochemical gradient whether expressedbiochemically or through surface texture (McKinney andMcKinney, 1993; Ryland, 1974). Some instances of gregar-iousness appear to be the result of biochemical stimuli; thesettling larvae tend to clump around previously settledlarvae.
Chemical characteristics of some potential substratesmay inhibit settlement of encrusting organisms (McKin-ney and McKinney, 1993). Modern sponges produce bio-chemical compounds, irritant mucus, and diffusible sur-face-active molecules that afford some protection againstovergrowth, either by killing the competitors or by makingthe sponge surface itself unsuitable for the settlement oflarvae of other organisms (Bergquist, 1978). Modernsponge-bryozoan interactions reported by Jackson andBuss (1975) showed that the effects of biochemical com-pounds utilized by the sponge appeared to be quite specificamong the bryozoan species. Microbial films on hard sub-strata are known commonly to stimulate settlement of lar-vae of sedentary benthic marine organisms, includingsome bryozoan species (McKinney and McKinney, 1993and references therein).
However, no evidence of either biochemical-microbialstimuli nor biochemical defenses can be identified in thefossil sponge-bryozoan association described here. Thephysical reaction of R. tenuis against bryozoans suggeststhat biochemical defenses against the bryozoan encrusta-tions were absent. Although, Girvanella coatings havebeen found on sponges from other levels of the same for-mation (Canas and Carrera, 1993), clear indications of itspresence are lacking in the sponges reported herein. Be-sides, there is no apparent reason why these microbialfilms would appear only on R. tenuis and not in the rest ofthe association.
The sponges found in the Argentine Precordillera showa great variety of form, size, and internal or external fea-tures. Sponge characteristics described in Table 1 are oftwo main morphotypes: discoidal and elongate. Represen-tatives of the discoidal morphotype range from the largePatellispongia or Psarodictyum to the small Allosacus,while the elongate morphotype includes the robust cup-shaped Calicocoelia, the cylindrical and annulate Ar-chaeoscyphia, or the small, digitate Rhopalocoelia tenuis.Globose or subglobose sponge forms could be included inthe discoidal morphotype or in a separate one. Clearly R.tenuis is one of the smallest sponges, if biovolume is con-
sidered and, in fact, it would be a very small target for lar-vae. Although it was more elevated above the substratethan the discoidal sponges, it was relatively short com-pared to other associated elongate sponge taxa.
Rhopalocoelia tenuis is texturally distinct from associ-ated sponges because of its fine spicules and smooth sur-face. Patellispongia argentina bears an important numberof bryozoan attachments, and also has a dense skeletonand small pores. Rhopalocoelia regularis also has smallspicules. However, the skeletal pattern is not dense, anddisplays more open pores than those of R. tenuis.
McKinney and McKinney (1993) have suggested thatphysical attributes of the substrate surface provoke a re-sponse in settling larvae. A general preference for asmooth surface was noted by Ryland (1974). In addition,McKinney (1996) stated that at least some of the differenc-es in substrate encrustation within an assemblage are dueto textural characteristics of the substrata. Thomsem(1977), in his study of bryozoan encrustations on erectbryozoans, recognized preferential settlement on bryozo-ans with relatively smooth surfaces. From these observa-tions and suggestions, and the characteristics of the Ar-gentinian sponges, it is possible to interpret that the spe-cific selection by trepostome bryozoan larvae for settlingonto R. tenuis could have been due to the smooth anddense external surface of this sponge.
Rhopalocoelia tenuis reacted against bryozoans but notagainst echinoderms, suggesting that the bryozoan infes-tation was unfavorable. Small individuals of modernsponges are subject to severe mortality as a result of bioticcompetition during the first year after settlement (Be-rgquist, 1978). A single small specimen of R. tenuis (8 cmlong) bears eight bryozoans and four echinoderms. Possi-bly bryozoans are potentially more harmful to the spongedue to their ability to encrust larger surface areas, whileechinoderm attachments represent a single small area offixation for the column that rarely increase in size.
As previously noted, the high proportion of epizoans re-corded on sponges is interpreted to reflect unfavorableconditions for settling on the sediment substrate in thethree localities analysed. Shifting bioclastic grainstonesand soft mudstones and wackestones occurred in thesponge-bearing limestones of the Precordillera. These un-stable conditions made the sponges a more suitable hardsubstrate, as ‘‘isolated islands,’’ for echinoderms, bryozo-ans, and some brachiopods (Fig. 6).
Modern, open textured and complex sponges bear amore diverse dweller association in their internal parts(Villamizar and Laughlin, 1992), but these apparently arelacking in the sponges studied here. Gundrum (1979) rec-ognized worm tubes protruding from a pore in one Penn-sylvanian sponge, recognizing a possible internal dweller.It is difficult to know if the fossil sponges once bore un-preservable soft organisms in their interiors. Surfaceworm tubes found in the Argentinian sponges are indica-tions of preference of these organisms for the exteriorparts.
PALEOBIOLOGICAL SIGNIFICANCE OF THEARGENTINE ENCRUSTING COMMUNITY
Some of the most studied hard substrate communitiesare those developed on early lithified carbonate sea floors

270 CARRERA
FIGURE 6—Reconstruction of the sponge-epizoan community of theArgentine Precordillera. 1- Patellispongia; 2- Archaeoscyphia; 3- Rho-palocoelia tenuis; 4- Rhopalocoelia clarkii; 5- Hudsonospongia; 6- Cri-noids (hipothetical forms); 7- Blastozoans (hypothetical forms); 8- Ber-enicea type bryozoan; 9- Trepostome bryozoan; 10- Pedunculate or-thid brachiopods; 11- Strophomenid brachiopods.
or hardgrounds. However, they provide only one of severaltypes of hard substrate that are studied by paleontologistswith an interest in the identity and paleoecology of thefaunas of such environments. By far the most abundanthard substrates in modern and ancient seas are those pro-vided by dead shells and other bioclastic debris and, insome cases, by the skeletons of the living biota as well(Wilson and Palmer, 1992). Epizoans cemented to theskeletons of host organisms offer clearer evidence of theirinterrelationship, and provide an opportunity to study theautecology of hosts and epizoans, the ecological structure,and evolutionary changes of hard substratum communi-ties.
Argentinean sponges represent an apparently exclusivehard substrate for encrusting epizoans in a carbonate softbottom environment. Although some differences are obvi-ous between the skeletons of a living biota and a hard-ground, it is possible to compare, in a broad sense, both en-crusting communities.
Argentinean sponge-epizoan communities are dominat-ed by crinoid holdfasts; blastozoans are a secondary com-ponent. Bryozoans comprise a low percentage of the epi-zoan community, and they exhibit low diversity. Brachio-pods are very rare as epibionts, and borings in sponges areabsent. Wilson et al.(1992) studied an Early Ordovicianhardground community, the Kanosh assemblages, that iscomparable in many ways to the Argentinean epizoancommunity. The low percentage of bryozoans, the lack ofborings, and the abundance of pelmatozoans make this as-semblage remarkably similar. However, it differs in theabundance of eocrinoid blastozoans, a dominant groupamong echinoderms in the early Middle Ordovician Ka-nosh assemblages and many of the Cambrian encrustingcommunities (Sprinkle and Guensburg, 1995).
Both the Argentinean and the Kanosh communities areclearly different from the Late Cambrian encrusting com-munity described by Brett et al. (1983), which is eocrinoiddominated, and lacking crinoids, bryozoans, or other skel-etal colonial encrusters. They also differ from the late Mid-dle Ordovician encrusting communities described by
Palmer and Palmer (1977) and Brett and Liddell (1978)which are bryozoan dominated.
The Early to Middle Ordovician was a time of rapid evo-lutionary diversification among hard substrate communi-ties (Palmer, 1982; Wilson and Palmer, 1992). Accordingto Wilson et al. (1992), the Kanosh hardground communi-ty appears to be transitional between the first hardgroundencrusting communities (Late Cambrian), which wereeocrinoid dominated and had no macroborers or bryozo-ans, and the later communities in which bryozoans werethe most prolific utilizers of hard substrate and macrobor-ers were common. The Argentinean assemblage is earlyLlanvirn in age, belonging to the E. suecicus conodontzones which are slightly younger than the upper ArenigKanosh assemblages. Possibly, this age difference makesthe Argentinean assemblage an evolutionary step further,when eocrinoids were displaced by crinoids and selectiveencrusting bryozoans. However, the community still re-mains dominated by solitary encrusters which is clearlydifferent from the Middle and Late Ordovician hard sub-strate assemblages dominated by colonial encrusters.
The presence of borers in hardground communities wasfirst recorded for the Middle Ordovician (Trypanites bor-ings of Palmer and Palmer, 1977). However, the oldest re-cord of borings in sponges dates from the Carboniferous(Gundrum, 1979), when specimens with borings of acro-thoracic barnacles occur. The absence of borings in the Or-dovician sponges of Argentina reflects the early evolution-ary state of development of boring organisms. However,Kershaw (1980) showed the presence of conspicuous Try-panites borings in Silurian stromatoporoids of Gotland.One possible explanation may be that the Trypanites ich-nogenus on hardgrounds and stromatoporoid skeletonswas a specialized organism that only bored chemically intocalcareous hard substrates and not in the siliceous skele-tons of the lithistid sponges.
Borers are a more significant component of the faunasof Mesozoic hardgrounds than those of earlier times. En-dolithic worms, sponges, and bivalves all started to be-come more important in both organic and inorganic Me-sozoic hard substrates (Palmer, 1982). However, the in-crease in the proportion of borers seems to start in the De-vonian and Carboniferous, with the appearance ofphoronids, acrothoracican barnacles, and recurved worms(Polydorites; Thayer, 1983; Signor and Brett, 1984; Ver-meij, 1987). Although Paleozoic borers are representedmainly by the simple and small Trypanites since the Mid-dle Ordovician (Wilson and Palmer, 1992), Devonian in-faunal assemblages are taxonomically and functionallymore diverse (Thayer, 1979; 1983). The first large scale di-versification of shell-breaking predators also took place inthe Devonian (Vermeij, 1983 Signor and Brett, 1984), andthis is possibly true for boring organisms that became im-portant dwellers in organic hard substrates. This first di-versification of borers and predators in the Late Paleozoicalso may have triggered the increase in the proportion ofencrusters with true exoskeletons, like cyclostome bryo-zoans, bivalves, and serpulid worms, that become commoncomponents later in the Mesozoic encrusting communities(Palmer, 1982, Wilson and Palmer, 1992).
ACKNOWLEDGEMENTSI wish to thank Dr. T. M. Sanchez, B.D. Chatterton, and
T. E. Guensburg for constructive suggestions on an early

EPIZOAN-SPONGE INTERACTIONS IN THE EARLY ORDOVICIAN 271
version of the manuscript. T. E. Guensburg provided help-ful comments on the determination of echinoderm hold-fasts. Appreciation is extended to the PALAIOS editorsand two anonymous reviewers for helpful suggestions thatsignificantly improved the manuscript. T. Benitez helpedwith drawings of community reconstruction. Financialsupport for this work was supplied by Consejo Nacional deInvestigaciones Cientıficas y Tecnicas (CONICET), Con-sejo de Investigaciones Cientıficas y tecnicas de la Prov-incia de Cordoba (CONICOR) and Agencia Nacional dePromocion Cientıfica y Tecnologica (ANPCYT). This paperis a contribution to the Project 410, IGCP-IUGS, TheGreat Ordovician Biodiversification event (GOBE).
REFERENCES
ALDINGER, H., 1968, Ecology of algal-sponge-reefs in the Upper Juras-sic of the Schwabische Alb, Germany: in MULLER, G., and FRIED-MAN, G., eds., Carbonate Sedimentology in Central Europe:Springer-Verlag, New York. p. 250–253.
ALVAREZ, F., and TAYLOR, P.D., 1987, Epizoan ecology and interac-tions in the Devonian of Spain: Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, v. 61, p. 17–31.
AYLING, A.M., 1981, The role of biological disturbance in temperatesubtidal encrusting communities: Ecology, v. 62, p. 830–847.
BERESI, S., and RIGBY, J.K., 1993, The Lower Ordovician sponges ofSan Juan, Argentina: Brigham Young University Geology Stud-ies, v. 39, p. 1–63.
BERGQUIST, P.R., 1978, Sponges: University of California Press,Berkeley, 268 p.
BRETT, C.E., 1981, Terminolgy and functional morphology of attach-ment structures in pelmatozoans echinoderms: Lethaia, v. 14, p.343–370.
BRETT, C.E., and LIDDELL, W.D., 1978, Preservation andPaleoecologyof a Middle Ordovician hardground community: Paleobiology, v. 4,p. 329–348.
BRETT, C.E., LIDDELL, W.D., and DERSTLER, K.L., 1983, Late Cambri-an hard substrate communities from Montana/Wyoming: The old-est known hardground encrusters: Lethaia, v. 16, p. 281–289.
CANAS, F.L., 1995a, Estratigrafıa y evolucion paleoambiental de lasrocas carbonaticas cambro-ordovıcicas de la Precordillera septen-trional: Unpublished Ph.D.Thesis, Universidad Nacional de Cor-doba, Cordoba, 300 p.
CANAS, F.L., 1995b, Early Ordovician Carbonate platform facies ofthe Argentine Precordillera: restricted shelf to open platform evo-lution: in COOPER, J.D., DROSER, M.L., and FINNEY, S.C., eds., Or-dovician Odyssey: Pacific Section Society for Sedimentary Geology(SEPM) Fullerton, Book 77, p. 221–224.
CANAS, F.L., 1999, Facies and sequences of the late Cambrian-earlyOrdovician carbonates of the Argentine Precordillera: A strati-graphic comparation with Laurentian platforms. In: KEPPIE, D.,and RAMOS, V., eds., Laurentia-Gondwana connections beforePangea: Geological Society of America, Special Paper 336: 43–62.
CANAS, F.L., and CARRERA, M.G, 1993, Early Ordovician microbial-sponge-receptaculitid bioherms of the Precordillera, Western Ar-gentina: Facies, v. 29, p.169–178.
CARRERA, M.G., 1994a, Taxonomıa, bioestratigrafıa y significado pa-leoambiental de las faunas de porıferos y briozoos del Ordovıcicode la Precordillera Argentina: Unpublished Ph.D.Thesis, Univer-sidad Nacional de Cordoba, Cordoba, 197 p.
CARRERA, M.G., 1994b, An Ordovician sponge fauna from San JuanFormation, Precordillera basin, western Argentina: Neues Jahr-buch fur Geologie und Palaontologie (Abhandlungen), v. 191, p.201–220.
CARRERA, M.G., 1996a, Ordovician Megamorinid Demosponges fromSan Juan Formation, Precordillera, Western Argentina: Geobios,v. 29, p. 643–650.
CARRERA, M.G., 1996b, Nuevos porıferos de la Formacion San Juan(Ordovıcico), Precordillera Argentina: Ameghiniana, v. 33, p. 335–342.
CARRERA, M.G., 1997a, Significado paleoambiental de los porıferos y
Briozoos de la Formacion San Juan (Ordovıcico), Precordillera Ar-gentina: Ameghiniana, v. 34, p. 179–199.
CARRERA, M.G., 1997b, Evolucion y recambio de la fauna de porıferosy Briozoos en el Ordovıcico de la Precordillera Argentina: Am-eghiniana, v. 34, p. 295–308.
CARRERA, M.G., and ASTINI, R., 1997, Valoracion de las restriccionesambientales durante la transicion Arenigiano-Llanvirniano, Or-dovıcico de la Precordillera Argentina: Revista Asociacion Geolo-gica Argentina, v. 52, p. 41–56.
CARRERA, M.G., and RIGBY, J.K., 1999, Biogeography of the Ordovi-cian sponges: Journal of Paleontology, v. 73, p. 26–37.
CARRERA, M. G., SANCHEZ, T. M., and BENEDETTO, J. L., 1999, Pa-leoenvironmental controls on biofacies in the Early Ordovicianlimestones of the Argentine Precordillera. Eight InternationalSymposium on the Ordovician System, Prague: Acta UniversitatisCarolinae-Geologica v. 43, p. 475–477.
CHATTERTON, B.D., 1975, A commensal relationship between a smallfilter feeding organism and Australian Devonian spiriferid bra-chiopods: Paleobiology, v. 1, p. 371–378.
CLARKE, J.M., 1908, The beginnings of dependent life: New YorkState Museum Bulletin, v. 121, p. 1–146.
CONWAY MORRIS, S., 1990, Parasitism: in BRIGGS, D.E.G., andCROWTHER, P.R., eds., Paleobiology: A synthesis: Blackwell Sci-entific Publications, Oxford, p. 376–381.
FINKS, R.M., 1960, Late Paleozoic faunas of the Texas region: Ameri-can Museum of natural History Bulletin, v. 120, p. 1–160.
FRANZEN, C., 1977, Crinoid holdfasts from the Silurian of Gotland:Lethaia, v. 10, p. 219–234.
FRITH, D.W., 1976, Animals associated with sponges at North Hay-ling, Hampshire: Journal of Linnean Society (Zoology), v. 58, p.353–362.
FURSICH, F.T., and WERNER, W., 1991, Palaeoecology of corallinesponge-coral meadows form the Upper Jurassic of Portugal: Pa-laeontogical Zentralblatt, v. 65, p. 35–69.
GUNDRUM, L.E., 1979, Demosponges as substrate: An example fromthe Pennsylvanian of North America: Lethaia, v. 12, p. 105–119.
HARTMAN, W.D., 1986, Astrorhizae, mamelons and symbionts of Re-cent Sclerosponges: Palaeontographica Americana, v. 56, p. 305–314.
HERRERA, Z.A., and BENEDETTO, J.L., 1991, Early Ordovician bra-chiopod faunas of the Precordillera basin, Western Argentina: bio-stratigraphy and paleobiogeographical affinities: in MAC KINNON,D.L., LEE, D.E., and CAMPBELL, J.D., eds., Brachiopods ThroughTime: Balkema, Rotterdam, p. 283–301.
HUNICKEN, M., and ORTEGA, G., 1987, Lower Llanvirn-Lower Cara-doc (Ord.) conodonts and graptolites from the Argentine Precor-dillera: IV European Conodont Symposium, ECOS IV, p. 136–145.
JACKSON, J. B., 1983, Biological Determinants of Present and Pastsessile animal distributions: in TEVESZ, M.J., and MCCALL, P.L.,eds, Biotic interactions in Recent and Fossil Benthic communities:Plenum Press, New York, p. 39–120.
JACKSON, J.B., and BUSS, L.W., 1975, Allelopathy and spatial compe-tition among coral reef invertebrates: Proceedings of the NaturalAcademy of Science, v. 72, p. 5160–5163.
JACKSON, J.B., and PALUMBI, S.R., 1979, Regeneration and partialpredation in cryptic coral reef environments: preliminary experi-ments on sponges and ectoprocts: in LEVY, C., and BOURY-ES-NOULT, N., eds., Biology des spongieres: Colloques Internationauxdu Centre National de la Recherche Scientifique (CNRS), v. 291,p. 303–308.
KERSHAW, S., 1980, Cavities and cryptic faunas beneath non-reefstromatoporoids: Lethaia, v. 13, p. 327–338.
LIDDELL, W.D., and BRETT, C.E., 1982, Skeletal overgrowths amongepizoans from the Silurian (Wenlockian) Waldron shale: Paleobi-ology, v. 8, p. 67–78.
MCKINNEY, F.K., 1996, Encrusting organisms on co-occurring disar-ticulated valves of two marine bivalves: Comparison of living as-semblages and skeletal residues: Paleobiology, v. 22, p. 543–567.
MCKINNEY, F.K., and JACKSON, J.B., 1989, Bryozoan Evolution: Uni-versity of Chicago Press, Chicago, 238 p.
MCKINNEY, F.K., and MCKINNEY, M.J., 1993, Larval behaviour andchoice of settlement site: correlation with environmental distri-bution pattern in an erect bryozoan: Facies, v. 29, p. 119–132.

272 CARRERA
MERGL, M., 1983, Rocky-bottom fauna of the Ordovician age in Bo-hemia (Arenigian; Prague basin, barrandian area): Vestnık Us-trednıho ustavu geologickeho, v. 58, p. 333–345.
PALMER, T.J., 1982, Cambrian to Cretaceous changes in hardgroundcommunities: Lethaia, v. 15, p. 309–323.
PALMER, T.J., and FURSICH, F.T., 1974, The ecology and a middle Ju-rassic Hardground and crevice fauna: Palaeontology, v. 17, p. 507–524.
PALMER, T.J., and PALMER, C.D., 1977, Faunal distribution and colo-nization strategy in a Middle Ordovician hardground community:Lethaia, v. 10, p. 179–199.
PALMER, T.J., and FURSICH, F.T., 1981, Ecology of sponge reefs fromthe Upper Bathonian of Normandy: Palaeontology, v. 24, p. 1–23.
PALUMBI, S.R., 1984, Tactics of acclimation: morphological changes ofsponges in an unpredictable environment: Science, v. 225, p.1478–1480.
PALUMBI, S.R., and JACKSON, J.B., 1982, Ecology of cryptic coral reefcommunities II. Recovery from small disturbance events by en-crusting bryozoa: The influence of ‘‘host’’ species and lesion size:Journal of Experimental Marine Biology and Ecology, v. 64, p.103–115.
RYLAND, J.S., 1974, Behaviour, settlement and metamorphosis ofbryozoan larvae: a review: Thalassia Jugoslavica, v. 10, p. 239–262.
SANCHEZ, T.M., WAISFELD, B.G., CARRERA, M.G., and TOFFOLO, S.,1993, Comunidades bentonicas en facies carbonaticas del Ordovı-cico Temprano de la Precordillera Argentina: Coloquios de Paleon-tologıa, v. 45, p. 139–162.
SANCHEZ, T.M., CARRERA, M.G., and BENEDETTO, J.L., 1996, Varia-ciones faunısiticas en el techo de la Formacion San Juan (Ordovı-cico temprano, Precordillera argentina): significado paleoambien-tal: Ameghiniana, v. 33, p. 185–200.
SIGNOR, P.W., and BRETT, C.E., 1984, The mid-Paleozoic precursor tothe Mesozoic marine revolution: Paleobiology, v. 10, p. 229–245.
SPJELDNAES, N., 1984, Epifauna as a tool in autoecological analysis ofSilurian brachiopods: Special Papers in Paleontology, v. 32, p.225–235.
SPRINKLE, J., and GUENSBURG, T.E., 1995, Origin of echinoderms inthe Paleozoic evolutionary fauna: The role of substrates: PA-LAIOS, v. 10, p. 437–453.
STORR, J.F., 1976, Field observations of sponge reactions as related totheir ecology: in HARRISON, F.W., and COWDEN, R.R., eds., Aspectsof Sponge Biology: Academic press, New York, p. 277–282.
TAYLOR, P.D., 1984, Adaptations for spatial competition and utiliza-tion in Silurian encrusting bryozoans: Special Papers in Paleon-tology, v. 32, p. 197–210.
TEVESZ, M.J., and MCCALL, P.L., 1983, Biotic Interactions in Recentand Fossil Benthic Communities: Plenum Press, New York, 837 p.THAYER, C.W., 1974, Substrate specificity of Devonian epizoa:Journal of Paleontology, v. 48, p. 881–894.
THAYER, C.W., 1979, Biological bulldozers and the evolution of ma-rine benthic communities: Science, v. 203, p. 458–461.
THAYER, C.W., 1983, Sediment-mediated biological disturbance andthe evolution of marine benthos: in TEVESZ, M.J., and MCCALL,P.L., eds., Biotic interactions in Recent and Fossil Benthic com-munities: Plenum Press, p. 480–626.
THOMSEM, E., 1977, Relations between encrusting bryozoans andsubstrate: an example from the Danian of Denmark: Bulletin ofthe Geological Society of Denmark, v. 26, p. 133–145.
VERMEIJ, G.J., 1983, Shell-breaking predation through time: inTEVESZ, M.J., and MCCALL, P.L., eds., Biotic interactions in Re-cent and Fossil Benthic communities: Plenum Press, p. 649–669.
VERMEIJ, G.J., 1987, Evolution and escalation: An ecological historyof life: Princenton University Press, 527 p.
VILLAMIZAR, E., and LAUGHLIN, R.A., 1992, Fauna associated with thesponges Aplysina archeri and Aplysina lacunosa in a coral reef ofthe Archipielago de los Roques, National Park, Venezuela: inREITNER, J., and KEUPP, H., eds., Fossil and Recent Sponges:Springer-Verlag, New York, p. 522–542.
WILSON, M.A., and PALMER, T.J., 1992, Hardgrounds andhardgroundfaunas: Institute of Earth Studies Publications, University ofWales, Aberystwyth, v. 9, p. 1–131.
WILSON, M.A., PALMER,T.J., GUENSBURG, T.E., FINTON, C. D., andKAUFMAN, L.E., 1992, The development of an Early Ordovicianhardground community in response to rapid sea-floor calcite pre-cipitation: Lethaia, v. 25, p. 19–34.
WOOD, R., DICKSON, J.A.D., and KIRKLAND-GEORGE, B., 1994, Turn-ing the Capitan reef upside down: a new appraisal of the ecology ofthe Permian Capitan reef, Guadalupe Mountains, Texas and NewMexico: PALAIOS, v. 9, p. 422–427.
WOODLEY, J.D., and NINETEEN AUTHORS, 1981, Hurricane Allen�s Im-pact on Jamaican coral reefs: Science, v. 214, p. 749–755.
ZIEGLER, B., 1964, Bewuchs auf Spongien: Palaeontological Zentral-blatt, Stuttgart, v. 38, p. 88–97.
ACCEPTED MAY 7, 2000
Copyright © 2022 FDOKUMEN




















