
Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales Productos bioactivos...
25
Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales) Carolina Villadóniga 1 , Sandra E. Vairo Cavalli 3* , Susana R. Morcelle del Valle 3* , Mª. Eugenia Errasti 3 , Mariela A. Bruno 3 , Miriam Barros 2 y Ana Mª. B. Cantera 1,2 1 Laboratorio de Enzimas Hidrolíticas, Facultad de Ciencias, Universidad de la República, Uruguay 2 Cátedra de Bioquímica, Facultad de Química, Universidad de la República, Uruguay. 3 Laboratorio de Investigación de Proteínas Vegetales (LIProVe), Universidad Nacional de La Plata, Argentina.*Carrera del Investigador Cientifico y Tecnológico, CONICET. Existe evidencia de que la dieta es un factor ambiental clave que afecta la incidencia de muchas enfermedades crónicas, por lo que se ha prestado mucho interés a la caracterización y explotación de los factores promotores de esta presente en los alimentos. La conciencia del efecto de la nutrición en la salud ha incentivado el desarrollo de productos promotores de la salud conocidos globalmente bajo el nombre de “alimentos funcionales”. Este concepto fue desarrollado primeramente en Japón en los años 1980s para certificar ciertos alimentos que presentaban beneficios documentados para la salud (Foods for Specified Health Use). De esta forma surgen los términos alimento funcional y nutracéutico. Un alimento funcional es un alimento convencional o similar en apariencia, que se consume como parte de la dieta normal, y que tiene efectos fisiológicos beneficiosos y/o reduce el riesgo de enfermedades crónicas más allá de las funciones nutricionales básicas. Estos alimentos se diseñan con un propósito concreto y debe existir evidencia científicamente controlada sobre su rol fisiológico (López, 2007). Un nutracéutico es un producto bioactivo aislado o purificado a partir de un alimento, que se suministra en una matriz no alimentaria (generalmente en píldoras, cápsulas o ampollas) y que se usa para mejorar la salud, en dosis que exceden aquellas que pueden ser obtenidas a partir del alimento natural. De esta manera surgen en el mercado alimentos con elevado contenido de diferentes componentes bioactivos como: ácidos grasos o esteroles, péptidos bioactivos, antioxidantes, carbohidratos prebióticos, vitaminas o minerales y bacterias prebióticas, entre otros. Los desarrollos tecnológicos en este campo han sido sorprendentes y estos productos serán probablemente una herramienta valiosa de la Ciencia de la Nutrición en el futuro (Juárez, 2007). Péptidos bioactivos Las proteínas, aparte de su función básica, son reservorio de péptidos que regulan los procesos vitales de los organismos (Karelin et al, 1998). Por lo tanto, actualmente son consideradas como fuente de péptidos biológicamente activos (bioactivos). Esto amplia el criterio usado por mucho tiempo para calificar el valor de las proteínas que se limitaba a su calidad nutricional: disponibilidad de nitrógeno 273
Transcript of Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales Productos bioactivos...

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
Carolina Villadóniga1, Sandra E. Vairo Cavalli3*, Susana R. Morcelle del Valle3*, Mª. Eugenia Errasti3, Mariela A. Bruno3, Miriam Barros2 y Ana Mª. B. Cantera1,2
1 Laboratorio de Enzimas Hidrolíticas, Facultad de Ciencias, Universidad de la República, Uruguay 2 Cátedra de Bioquímica, Facultad de Química, Universidad de la República, Uruguay. 3 Laboratorio de Investigación de Proteínas Vegetales (LIProVe), Universidad Nacional de La Plata,
Argentina.*Carrera del Investigador Cientifico y Tecnológico, CONICET.
Existe evidencia de que la dieta es un factor ambiental clave que afecta la incidencia de muchas enfermedades crónicas, por lo que se ha prestado mucho interés a la caracterización y explotación de los factores promotores de esta presente en los alimentos. La conciencia del efecto de la nutrición en la salud ha incentivado el desarrollo de productos promotores de la salud conocidos globalmente bajo el nombre de “alimentos funcionales”. Este concepto fue desarrollado primeramente en Japón en los años 1980s para certificar ciertos alimentos que presentaban beneficios documentados para la salud (Foods for Specified Health Use). De esta forma surgen los términos alimento funcional y nutracéutico. Un alimento funcional es un alimento convencional o similar en apariencia, que se consume como parte de la dieta normal, y que tiene efectos fisiológicos beneficiosos y/o reduce el riesgo de enfermedades crónicas más allá de las funciones nutricionales básicas. Estos alimentos se diseñan con un propósito concreto y debe existir evidencia científicamente controlada sobre su rol fisiológico (López, 2007). Un nutracéutico es un producto bioactivo aislado o purificado a partir de un alimento, que se suministra en una matriz no alimentaria (generalmente en píldoras, cápsulas o ampollas) y que se usa para mejorar la salud, en dosis que exceden aquellas que pueden ser obtenidas a partir del alimento natural.
De esta manera surgen en el mercado alimentos con elevado contenido de diferentes componentes bioactivos como: ácidos grasos o esteroles, péptidos bioactivos, antioxidantes, carbohidratos prebióticos, vitaminas o minerales y bacterias prebióticas, entre otros. Los desarrollos tecnológicos en este campo han sido sorprendentes y estos productos serán probablemente una herramienta valiosa de la Ciencia de la Nutrición en el futuro (Juárez, 2007).
Péptidos bioactivos
Las proteínas, aparte de su función básica, son reservorio de péptidos que regulan los procesos vitales de los organismos (Karelin et al, 1998). Por lo tanto, actualmente son consideradas como fuente de péptidos biológicamente activos (bioactivos). Esto amplia el criterio usado por mucho tiempo para calificar el valor de las proteínas que se limitaba a su calidad nutricional: disponibilidad de nitrógeno
273

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
derivado de aminoácidos esenciales y no esenciales, propiedades alergénicas y ausencia de anti-nutrientes (Dziuba & Darewicz, 2007).
Los péptidos bioactivos son productos de la degradación de proteínas por proteasas que poseen una función biológica específica solo luego de ser liberados de la proteína precursora. Han sido definidos, por lo tanto, como fragmentos específicos de una proteína que interactuando con los receptores específicos en las células blanco inducen respuestas fisiológicas que regulan las funciones del organismo. Muchos péptidos bioactivos han sido identificados, pero debe hacerse una distinción entre las proteínas bioactivas presentes naturalmente en los alimentos (por ejemplo: factores de crecimiento o inmunoglobulinas presentes en la leche) y los péptidos que se generan durante la digestión de fuentes proteicas intactas, o por el uso de hidrolizados de proteína como componentes de un sistema alimentario formulado.
Debe tenerse en cuenta que la actividad biológica de los péptidos puede tener una efecto positivo o negativo en el organismo. Los péptidos celiacos son un ejemplo de péptidos con una influencia negativa en el organismo (Kitts & Weiler, 2003).
Entre los efectos fisiológicos más conocidos de los péptidos bioactivos se encuentran su capacidad de actuar sobre los sistemas inmune, cardiovascular, endócrino y digestivo. Las actividades que han sido estudiadas comprenden aspectos muy variados, destacándose las siguientes: antitumoral, antimicrobiana (Martínez & Martínez, 2006), opioide, antihipertensiva, inmunomodulatoria, transportadora de calcio, antitrombótica, hipocolesterolémica y antioxidante (Haileselassie et al., 1999). Así actualmente, determinados péptidos son empleados en el tratamiento de enfermedades dentales, de mala absorción de minerales, diarrea, hipertensión, trombosis e inmunodeficiencias (Thomas et al., 2001)
Dentro de los péptidos bioactivos existe una gran diversidad. El tamaño es muy variado, conteniendo en promedio de 2 a 20 residuos de aminoácidos (Pihlanto-Lepälä, 2001). Se han identificado tanto di o tri péptidos así como polipéptidos complejos lineales o cíclicos que frecuentemente están modificados por glicosilación, fosforilación o acilación de los residuos de aminoácido (Dziuba & Darewicz, 2007).
Un péptido puede poseer más de una función biológica. También se han encontrado secuencias sobrepuestas de péptidos con diferentes funciones formando parte de la estructura de ciertas proteínas, como las caseínas (Meisel, 1998).
Los péptidos bioactivos pueden ejercer su acción tanto a nivel local (a través de la interacción con receptores en las células del tracto gastrointestinal) como sistémico, ya que pueden atravesar el epitelio intestinal (Meisel, 2004; Vermeirssen et al., 2004). Los péptidos mas pequeños (dos o tres residuos de aminoácidos) pueden ser absorbidos mas fácilmente que los más grandes. Los péptidos que ejercen su actividad localmente en el tracto intestinal, como los péptidos antimicrobianos, son más largos.
Si bien aún no ha sido plenamente establecida la relación estructura-actividad de muchos péptidos bioactivos se ha observado que los péptidos que manifiestan una determinada bioactividad muestran características estructurales comunes.
274

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
Activación proteolítica de los péptidos bioactivos
Las proteínas son precursores potenciales de péptidos bioactivos tanto como compuestos endógenos de los organismos vivos así como exógenos de origen alimenticio.
La liberación de péptidos bioactivos por proteólisis es un hecho que se ha demostrado tanto “in vivo” como “in vitro”. Se han reconocido y caracterizado un vasto número de péptidos bioactivos en fluidos corporales y tejidos de diferentes organismos (bacterias, hongos, plantas y animales), producidos por proteólisis endógena tanto de proteínas especializadas (precursores de hormonas, neurotransmisores) como de proteínas con otras funciones (enzimas, proteínas estructurales, de transporte, etc.). Los péptidos derivados de proteínas funcionales se cree que juegan un rol regulatorio a largo plazo que contribuye al balance homeostático del organismo (Ivanov et al, 1998, 2000 y 2006; Karelin et al, 1998). Por degradación enzimática in vitro se han obtenido péptidos bioactivos de preparaciones biológicas incluyendo tanto homogenatos de tejidos como proteínas individuales. Como ejemplos se encuentran los fragmentos bioactivos derivados de β-caseína (β-casomorfinas), hemoglobina (hemorfinas; Glämsta et al, 1991), proteína básica de mielina, gluten, seralbúmina bovina, citocromo c y lactoferrina (Karelin et al, 1998).
Las proteínas alimentarias representan una fuente muy importante de péptidos bioactivos que pueden ser liberados por degradación proteolítica tanto in vivo como in vitro (Rutherfurd-Markwick & Moughan, 2005).
Durante la digestión gastrointestinal, tanto a nivel de las enzimas gastrointestinales como de las enzimas de la flora intestinal, pueden liberarse péptidos bioactivos a partir de las proteínas de la dieta. Se ha reportado la presencia de péptidos opioides en los digeridos gastrointestinales tras el consumo de leche o caseína (Svedberg et al, 1985; Meisel & FitzGerald, 2000).
Los péptidos bioactivos pueden ser producidos a partir de proteínas precursoras por hidrólisis enzimática in vitro con enzimas digestivas o derivadas de microorganismos o plantas.
En el laboratorio se pueden simular las condiciones de hidrólisis en los distintos compartimentos del tracto gastrointestinal ajustando las variables de temperatura, pH, tiempo de digestión, así como concentración y tipo de proteasas para producir hidrolizados proteicos como los que surgirían en la digestión humana en adultos. La hidrólisis se puede llevar a cabo directamente en una celda de ultrafiltración con agitación a fin de remover, inmediatamente luego de la digestión, los péptidos de bajo peso molecular que se generan (Nass et al., 2008).
Las enzimas pancreáticas, principalmente tripsina, han sido usadas para liberar muchos péptidos bioactivos conocidos de las proteínas de la dieta. Otras enzimas y combinaciones diferentes de enzimas incluyendo alcalasa, quimotripsina, pancreatina y pepsina así como de fuentes bacterianas y fúngicas han sido aplicas para generar péptidos bioactivos a partir de varias fuentes proteicas (Korhonen & Pihlanto-Leppälä, 2003).
275

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
Además se pueden liberar péptidos bioactivos por hidrólisis durante la elaboración o el procesamiento de algunos alimentos (quesos y sueros de quesería) o por acción de enzimas proteolíticas nativas (leche) o enzimas bacterianas usadas en la preparación del alimento (leches fermentadas; Korhonen & Pihlanto-Leppälä, 2003).
También se han obtenido péptidos modificados por semisíntesis, diseñados a partir de péptidos naturales, con el fin de incrementar su actividad (Teschemacher et al., 1997).
Hasta ahora, la hidrólisis enzimática de las moléculas de proteína enteras ha sido la forma más común de producir péptidos bioactivos (Korhonen &. Pihlanto, 2006).
El creciente interés en los péptidos bioactivos ha estimulado la investigación. Los tópicos en los que se esta trabajando activamente incluyen: la obtención de péptidos bioactivos (evaluación de diferentes fuentes y enzimas proteolíticas, aislamiento de péptidos), caracterización funcional y estructural de los péptidos bioactivos (identificación de la bioactividad, determinación de la estructura, relación estructura-función), efectos in vivo (evidencia del rol fisiológico, efectos nocivos, tóxicos y alergénicos) y uso potencial para la promoción de la salud (alimentos funcionales y nutraceuticos).
Los desafíos tecnológicos principales están centrados en, primero, la manufactura de productos lácteos fermentados con alta concentración de péptidos bioactivos particulares o sus precursores, los cuales luego de la digestión en el tracto gastrointestinal darán lugar a péptidos bioactivos y, en segundo lugar, en la producción de hidrolizados de proteínas de leche con una alta concentración de péptidos que tengan una bioactividad específica y funcionalidad que los hagan adecuados como ingredientes en otros alimentos, incluidos los productos lácteos.
Fuentes naturales de péptidos bioactivos
Proteínas lácteas
La ruptura enzimática de las moléculas de proteína de la leche resulta en un número de péptidos bioactivos que ha sido caracterizado (Meisel et al., 1989; Séverin & Wenshui 2005; Clare & Swaisgood, 2000). Estos péptidos, inactivos dentro de la secuencia de las proteínas precursoras, una vez que son liberados, pueden modular diversos procesos regulatorios del cuerpo (Meisel, 2005; Möller et al., 2008).
Se han descrito numerosos efectos in vitro e in vivo, en ensayos con animales, de péptidos derivados de proteínas lácteas obtenidos en diversas condiciones de hidrólisis. Algunas de estos efectos incluyen la actividad antimicrobiana, antioxidante, antilipémica, opioide (hay péptidos que se ligan a receptores opioides con acción agonista o antagonista), inmunomoduladora (péptidos derivados de caseína que estimulan la proliferación de linfocitos humanos y la fagocitosis por macrófagos), transportadora de minerales (caseinofosfopéptidos). Asimismo, péptidos funcionales derivados de la caseína, presentes ya sea en la leche o en productos lácteos, han demostrado poseer efectos en el sistema cardiovascular debido principalmente a sus características antitrombótica o antihipertensiva. Adicionalmente, muchos derivados
276

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
peptídicos de la leche revelan una bioactividad multifuncional (Möller et al., 2008; Oleszczuk et al., 1997). En la tabla 20.1 se presentan algunos ejemplos de péptidos bioactivos derivados de proteínas de la leche.
Precursor Nombre Secuencia Bioactividad αS1-CN bovina (f 57-79)
Caseinofosfopéptido I*KQMEAESI*SSSEEI*V*
PNSV*EQK Transporte de minerales
αS1-CN bovina (f 1-23)
Isracidina RPKHPIKHQGLPEQVLNENLLRF
Antibacteriano
αS2-CN bovina (f 2-21)
caseinofosfopéptido NTM*EHV*SSSEESI*I*SQETY*K
Transporte de minerales
αS2-CN bovina (f 165-203)
Casocidina-I KTKLTEEEKNRLNFLKKISQRYQKFALPQYLKTVYQHQK
Antibacteriano
β-CN bovina (f 1-28)
Caseinofosfopéptido REL*EEL*NV*P*GEI*V*ESL*SSSEESI*TRI*NK
Transporte de minerales
β-CN bovina (f 60-66)
Β-casomorfina-7 YPFPGPI Agonista opioide, inhibidor de ACE
β-CN bovina (f 63-68)
Inmunopéptido PGPIPN Inmunomodulador
β-CN bovina (f 98-105)
Péptido antioxidante derivado de β-CN
VKEAMAPK Antioxidante
κ-CN bovina (f 25-34)
Casoxina C YIPIQYVLSR Antagonista opioide Inmunoestimulante Contractor del ileum Inhibidor de ACE
κ-CN bovina (f 106-116)
Casoplatelina MAIPPKKNQDK Antitrombótico
κ-CN bovina (f 96-106)
Péptido antioxidante derivado de κ-CN
ARHPHPHLSFM Antioxidante
κ-CN bovina (f 106-169)
Kapacina MAIPPKKNQDKTEIPTINTIAS*GEPTSTPTTEAVESTVATLEDS*PEVIESPPEINTVQVTSTAV
Antimicrobiano
α-La (f 50-53) α-Lactorfina YGLF Agonista opioide, Inhibidor de ACE
α-La (f 1-5) LTD 1 EQLTK Antibacteriano β-Lg (f 102-105) β-Lactorfina YLLF Agonista opioide
Inhibidor de ACE β-Lg (f 146-149) β- Lactokinina HIRL Inhibidor de ACE β-Lg (f 142-148) β- Lactokinina ALPMHIR Inhibidor de ACE β-Lg 25-40 No asignado AASDISLLDAQSAPLR Antibacteriano β-Lg 78-83 No asignado IPAVFK Antibacteriano β-Lg 92-100 No asignado VLVLDTDYK Antibacteriano Lactoferrina bovina (f 17-41)
Lactoferricina-B FKCRRWQWRMKKLGAPSITCVRRAF
Antibacteriano
Tabla 20.1. Péptidos bioactivos derivados de proteínas lácteas (Base de datos: BIOPEP, url: http://www.uwm.edu.pl/biochemia/index_en)
277

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
Además de la leche bovina y sus productos derivados se ha abordado la obtención e identificación de péptidos bioactivos de productos lácteos y leche de otros orígenes. Silva et al (2006) han aislado e identificado varios péptidos con actividad inhibitoria de ECA y/o antioxidante en sistemas semejantes a quesos ovinos y caprinos manufacturados con un cuajo vegetal. Hidrolizados de leche de yak producen los péptidos PPEIN (κ-CN; f156–160) y PLPLL (β-CN; f136–140) con actividad antihipertensiva (Mao et al., 2006 y 2007).
Los péptidos de origen lácteo se pueden producir en una escala industrial, y como consecuencia de ello, estos péptidos han sido considerados para su aplicación tanto como suplementos dietarios en "alimentos funcionales" o como drogas pudiendo ser utilizados como nutracéuticos en la formulación de alimentos y preparados farmacéuticos (Oleszczuk et al., 1997).
Otras proteínas alimentarias
Se han aislado y caracterizado péptidos bioactivos a partir de hidrolizados de otras fuentes de proteínas alimentarias incluyendo huevo, pescado, ostras, cereales (arroz, trigo), soja, y otras fuentes de proteínas animales como: sangre (hemoglobina, seralbumina) y músculo (actina, miosina). En la tabla 20.2 se muestra algunos péptidos bioactivos obtenidos por hidrólisis enzimática de proteínas no lácteas.
Se han encontrado un gran número de péptidos con actividad antioxidante en hidrolizados de fuentes de proteínas vegetales tales como: soja (Pena-Ramos & Xiong, 2002) y maiz (Kong & Xiong, 2006) La baja incidencia de hipercolesterolemia, enfermedades del corazón, y cáncer en algunas poblaciones de Asia, se ha asociado con un elevado consumo de proteína de soja. La estructura primaria de las proteínas de soja demuestra la presencia de secuencias con actividades: antihipertensiva, anticolesterolémica, antioxidante y anticancerígena.
También se han estudiado otras fuentes de proteínas animales como: pescado (Wu et al 2003), ostras (Qian et al., 2008), huevo (Dávalos et al., 2004) y gelatina (Kim et al., 2001; Mendis et al., 2005). Se han obtenido péptidos con actividad inhibitoria de ECA por hidrólisis de músculo de: atún WPEAAELMMEVDP (Qian et al., 2007) y de besugo GY, VY, GF y VIY (Fahmi et al., 2004).
Bases de datos
La predicción de la bioactividad de los péptidos puede ayudar a identificar las proteínas que contienen en su secuencia los péptidos con potencial para ser alimentos funcionales y las enzimas específicas para liberarlos. Sin embargo la bioactividad predicha debe ser medida para validar los hallazgos (Pripp et al, 2005). El modelado cuantitativo de relaciones estructura-función provee de una metodología para encontrar expresiones matemáticas que puedan ser útiles para estimar las actividades de cualquier compuesto relacionado y para la predicción de estructuras de alta actividad.
La base de datos de péptidos bioactivos (BIOPEP) desarrollada en el Departamento de Bioquímica de Alimentos, Universidad de Warmia y Mazury,
278
Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
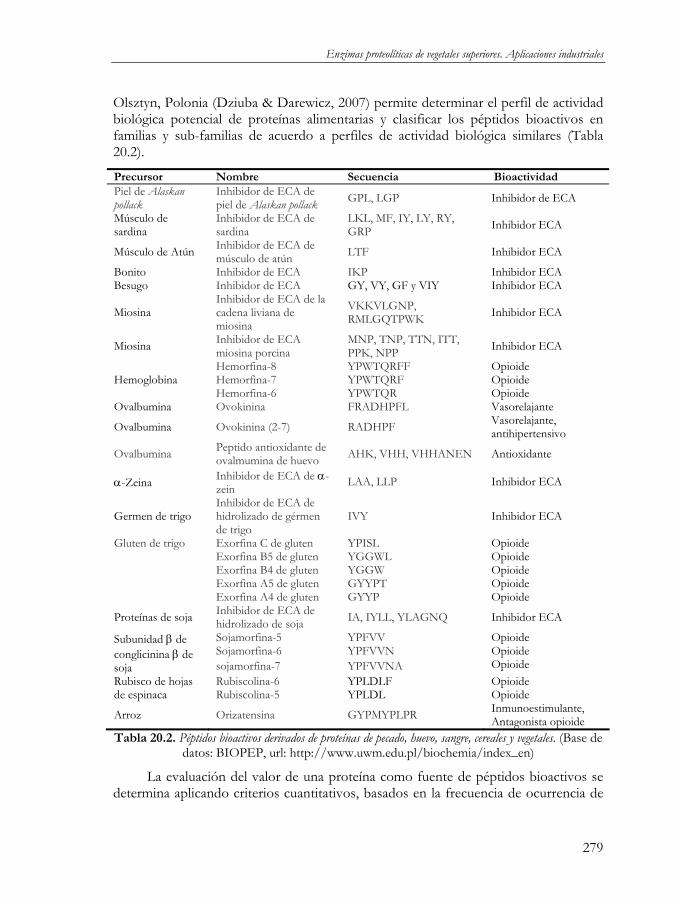
Olsztyn, Polonia (Dziuba & Darewicz, 2007) permite determinar el perfil de actividad biológica potencial de proteínas alimentarias y clasificar los péptidos bioactivos en familias y sub-familias de acuerdo a perfiles de actividad biológica similares (Tabla 20.2).
Precursor Nombre Secuencia Bioactividad Piel de Alaskan pollack
Inhibidor de ECA de piel de Alaskan pollack GPL, LGP Inhibidor de ECA
Músculo de sardina
Inhibidor de ECA de sardina
LKL, MF, IY, LY, RY, GRP Inhibidor ECA
Músculo de Atún Inhibidor de ECA de músculo de atún LTF Inhibidor ECA
Bonito Inhibidor de ECA IKP Inhibidor ECA Besugo Inhibidor de ECA GY, VY, GF y VIY Inhibidor ECA
Miosina Inhibidor de ECA de la cadena liviana de miosina
VKKVLGNP, RMLGQTPWK Inhibidor ECA
Miosina Inhibidor de ECA miosina porcina
MNP, TNP, TTN, ITT, PPK, NPP Inhibidor ECA
Hemorfina-8 YPWTQRFF Opioide Hemorfina-7 YPWTQRF Opioide Hemoglobina Hemorfina-6 YPWTQR Opioide
Ovalbumina Ovokinina FRADHPFL Vasorelajante
Ovalbumina Ovokinina (2-7) RADHPF Vasorelajante, antihipertensivo
Ovalbumina Peptido antioxidante de ovalmumina de huevo AHK, VHH, VHHANEN Antioxidante
α-Zeina Inhibidor de ECA de α-zein LAA, LLP Inhibidor ECA
Germen de trigo Inhibidor de ECA de hidrolizado de gérmen de trigo
IVY Inhibidor ECA
Gluten de trigo Exorfina C de gluten YPISL Opioide Exorfina B5 de gluten YGGWL Opioide Exorfina B4 de gluten YGGW Opioide Exorfina A5 de gluten GYYPT Opioide Exorfina A4 de gluten GYYP Opioide
Proteínas de soja Inhibidor de ECA de hidrolizado de soja IA, IYLL, YLAGNQ Inhibidor ECA
Sojamorfina-5 YPFVV Opioide Sojamorfina-6 YPFVVN Opioide
Subunidad β de conglicinina β de soja sojamorfina-7 YPFVVNA Opioide
Rubiscolina-6 YPLDLF Opioide Rubisco de hojas de espinaca Rubiscolina-5 YPLDL Opioide
Arroz Orizatensina GYPMYPLPR Inmunoestimulante, Antagonista opioide
Tabla 20.2. Péptidos bioactivos derivados de proteínas de pecado, huevo, sangre, cereales y vegetales. (Base de datos: BIOPEP, url: http://www.uwm.edu.pl/biochemia/index_en)
La evaluación del valor de una proteína como fuente de péptidos bioactivos se determina aplicando criterios cuantitativos, basados en la frecuencia de ocurrencia de
279

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
fragmentos bioactivos en una secuencia proteica (A) y la actividad potencial de los fragmentos de proteína (B). En la mencionada base de datos (url: http://www.uwm.edu.pl/biochemia/index_en) están documentadas 44 tipos diferentes de actividades biológicas que pueden ser ejercidas por péptidos de muy variado origen. La división en familias, es de primera importancia para el diseño de procesos de proteolisis y selección de las proteínas más ricas en péptidos con una determinada actividad.
Actividades biológicas de péptidos bioactivos obtenidos por hidrólisis
Péptidos antioxidantes
Como consecuencia de la respiración celular, en los organismos aeróbicos se forman radicales libres denominadas especies reactivas del oxígeno (EROs) que incluyen, entre otras, a los radicales hidroxílico (•OH) y anión superóxido (Ο2
• −). Estos debido a su inestabilidad química reaccionan rápidamente con diferentes moléculas (Martínez Sánchez, 2005). Si bien en condiciones normales las EROs son efectivamente eliminadas por sistemas celulares antioxidantes de defensa, un desbalance entre su generación y eliminación dará lugar a un stress oxidativo (Halliwell, 1996). La consecuencia inmediata es el daño al ADN, la membrana lipídica, las proteínas y los carbohidratos, alterando la función celular (Cooke et al., 2003; Mukul et al., 2005; Awasthi et al., 2003; Dröge W, 2002). Es conocido el papel de los radicales libres en el mecanismo fisiopatológico de los procesos de envejecimiento, cáncer, esclerosis múltiple, demencia senil, enfermedades cardiovasculares, artritis y ateroesclerosis, entre otras (Blake & Winyard, 1995; Halliwell & Gutteridge, 1990; Halliwell & Gutteridge, 1999).
Por su parte, la oxidación lipídica representa un problema en la industria alimenticia produciéndose olores rancios, colores oscuros y aromas indeseables así como productos de reacción potencialmente tóxicos alterando la calidad nutritiva y la vida media del alimento (Akoh & Min, 2002). Numerosas investigaciones han tenido como tema la relación entre la salud humana y nutrientes menores con actividad antioxidante (Chang et al., 2007).
Un antioxidante puede ser definido como aquella sustancia que en baja concentración respecto a un sustrato oxidable disminuye o inhibe la oxidación del mismo. Esta definición incluye antioxidantes no enzimáticos así como los enzimáticos (Sies, 1997).
Debido a los potenciales riesgos para la salud que tendría el uso de antioxidantes artificiales (Halliwell & Whiteman, 2004), en los últimos años se ha buscado reemplazarlos en los alimentos por antioxidantes naturales. Esto ha motivado la obtención de péptidos por síntesis o, degradación proteica por acción de proteasas. Las fuentes proteicas son muy variadas: derivadas de pescado (Wu et al., 2003; Je et al., 2005) de yema de huevo (Sakanaka et al., 2004); carne de cerdo (Saiga et al., 2003), soja (Gibbs et al., 2004), gluten de trigo y leche.
280

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
Los péptidos con actividad antioxidante serían de utilidad en la industria alimentaria, no solo como conservantes naturales, sino también en la formulación de alimentos funcionales destinados a la prevención de enfermedades en los individuos que los consuman.
Péptidos antihipertensivos
La hipertensión arterial (HA) constituye la principal causa de muerte. El riesgo de desarrollar enfermedades cardiovasculares esta directamente relacionado al nivel de presión arterial (FitzGerald et al., 2004). El mecanismo de control de la presión arterial (PA), que juega un rol principal y es conocido más detalladamente, es el ejercido por el sistema renina-angiotensina-aldosterona (RAAS). Dentro del RAAS la enzima convertidota de angiotensina-I (ECA-I; EC 3.4.15.1) cataliza la conversión de angiotensina I (DRVYIHPFHL) en angiotensina II (DRVYIHPF). El octapeptido angiotensinma II actúa directamente en las células de músculo liso del lecho vascular produciendo vasoconstricción y a su vez estimulando la liberación de aldosterona, hormona que induce la retención de sodio y agua y la excreción de potasio. La ECA-I actúa simultáneamente en el sistema quinina-calicreina, catalizando la degradación de las bradiquininas, compuestos de potente acción vasodilatadora. Al inhibirse ECA-I se produce simultáneamente la disminución de: la formación de angiotensina II y la degradación de bradiquinina, lo que globalmente conduce al descenso de la PA (Murray & FitzGerald, 2007).
Se han obtenido péptidos inhibidores potentes de ECA-I (IECA-I) a partir de hidrolizados de diversos alimentos incluyendo: leche, pescado, huevos, cereales y soja (Hong et al., 2008).
También se ha probado la liberación de péptidos IECA-I durante los procesos fermentativos de la leche: en el proceso proteolítico que tiene lugar durante la maduración de ciertas tipos de quesos y leches fermentadas (Korhonen & Pihlanto, 2006). Han sido aislados péptidos antihipertensivos a partir de queso Gorgonzola (Smacchi & Gobbetti, 1998) y queso Manchego producido con cultivos iniciadores diferentes (Gómez-Ruiz et al., 2002). En algunos casos maduraciones muy prolongadas hacen que se pierda la actividad antihipertensiva, esto ocurre en los quesos Parmesano y Gouda (Silva & Malcata, 2005).
Otras fuentes de péptidos antihipertensivos lo constituyen los hidrolizados de proteínas de clara de huevo en los que se ha probado su efecto hipotensor en ratas espontáneamente hipertensas (Manso et al., 2008).
La relación estructura-función de los péptidos antihipertensivos no ha sido completamente elucidada. Sin embargo, existen algunos rasgos comunes entre los péptidos ya identificados que han permitido sintetizar muy exitosamente péptidos con esta bioactividad. Estos péptidos son IECA-I competitivos. La bioactividad parece estar influenciada fuertemente por la secuencia del tripeptido C-terminal. En la mayoría de los casos, se ve que los dipéptidos son inhibidores más potentes que los tripéptidos que son resistentes a la acción de las endopeptidasas del tracto digestivo. Algunos IECA-I potentes derivados de alimentos contienen un residuo de prolina en
281

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
el extremo C-terminal mientras que otros contienen residuos de lisina o arginina. Esto sugiere la presencia de un residuo con carga positiva contribuye a la potencia del inhibidor. Muchos IECA-I tienen en común propiedades estructurales en las que están incluidas: tamaño pequeño, presencia de residuos hidrofóbicos (aromáticos o de cadenas laterales ramificada) además de residuos de prolina, arginina o lisina (Murray & FitzGerald, 2007).
La comprobación de actividad inhibitoria in vitro de un péptido no es suficiente para asegurar el efecto antihipertensivo in vivo. Luego de ser administrado oralmente el IECA debe ser absorbido de forma intacta en el intestino y ser resistentes a la degradación posterior por las peptidasas plasmáticas.
Ha sido demostrado el efecto antihipertensivo de ciertos péptidos, tanto aislados o como ingredientes de alimentos funcionales, en pacientes hipertensos. Esto ha impulsado el lanzamiento de alimentos funcionales o nutracéuticos capaces de prevenir y tratar la hipertensión, disminuyendo el riesgo de enfermedades cardiovasculares (Hong et al., 2008).
Existe evidencia de péptidos con efecto antihipertensivo, in vivo e in vitro, en productos lácteos fermentados con Lactobacillus helveticus y Saccharomyces cereviseae que contienen los tripéptidos IPP y VPP (Möller et al., 2008). Algunos ejemplos de preparaciones de origen lácteo que se encuentran en el mercado con propiedades antihipertensivas comprobadas en humano son los productos: CalpisTM (bebida láctea descremada fermentada con Lactobacillus helveticus y comercializada por Calpis Food Industry, Japón), Biozate® (hidrolizado de proteínas de suero producido por DAVICO Foods Internacional Inc., EEUU), Evolus® (leche agria fermentada con Lactobacillus helveticus y con concentrado de arándanos producida por Valio Ltd., Finlandia).
Péptidos opioides
Estos péptidos son ligandos de receptores opioides, que tienen efectos de tipo opioide, que pueden ser inhibidos por naloxone. Estos péptidos pueden interaccionar con diferentes receptores específicos, y sus efectos fisiológicos dependerán del tipo de receptor al que se unan. Los receptores opioides μ están vinculados al control de la motilidad intestinal y comportamiento emocional; los receptores δ, al control del comportamiento emocional, y los receptores κ están relacionados con la analgesia y la saciedad (Torres-Llanez et al., 2005).
Estudios recientes demostraron que existen péptidos en los productos lácteos que ejercen un rol principal en el sistema nervioso, y pueden tener actividades agonistas o antagonistas (Silva & Malcata, 2005). También se ha demostrado la presencia de péptidos opioides obtenidos a partir de proteínas de trigo, arroz, sangre (albúmina y hemoglobina), soja y espinaca (Teschemacher, 2003).
Los péptidos opioides provenientes de las proteínas alimentarias son conocidos como péptidos opioides exógenos atípicos. Son considerados exógenos debido a que provienen de una variedad de proteínas extrañas al organismo. A diferencia de los péptidos opioides endógenos, encefalinas, endorfinas y dinorfinas, que son liberados
282

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
exclusivamente a partir de tres proteínas precursoras endógenas (proencefalinas, proopiomelanocortina y prodinorfina). Los péptidos opioides exógenos se consideran atípicos, ya que presenta una secuencia aminoacídica N-terminal variable, a diferencia de los endógenos típicos que presentan la secuencia conservada: YGGF. La característica estructural común, entre los péptidos atípicos y típicos, es la presencia de un residuo de tirosina N-terminal (excepto para el caso de los provenientes de α-caseína) y de otro residuo aromático en la tercera o cuarta posición (Y-X-X-F o Y-X-F) que constituyen un motivo estructural importante para la unión al receptor opioide (Meisel & Bockelmann, 1998).
Los péptidos opioides derivados de proteínas lácteas que presentan actividad como agonistas son llamados exorfinas, en tanto que las casoxinas son los que ejercen actividad antagonista. Las β-casorfinas son opioides agonistas y son fragmentos provenientes de la hidrólisis enzimática de la β-caseína. La secuencia primaria de estos péptidos corresponde al fragmento 60-70 de dicha proteína, y han sido caracterizados como ligandos del receptor μ. Debido al alto contenido de residuos de Pro, estos péptidos son aparentemente resistentes al ataque proteolítico. Son absorbidos intactos durante la digestión, y son capaces de inhibir la motilidad gastrointestinal, así como la velocidad de vaciamiento del estómago por interacción directa con los receptores opioides (Haileselassie et al., 1999). Asimismo, se detectaron β-casorfinas en el plasma de mujeres encintas o en el período de lactancia (Shah, 2000). De los hidrolizados obtenidos por digestión con pepsina de la αS1 –caseína se aislaron las α-casorfinas, aunque el agonista opioide más activo es la morfoceptina, un tetrapéptido proveniente del grupo de las β-casorfinas bovinas. Más recientemente, Perpetuo y col. (2003) informaron un péptido nuevo obtenido de γ-caseína que posee actividad opioide in vitro y que fuera generado por hidrólisis tríptica de la caseína (Perpetuo et al., 2003). Otras secuencias de opioides agonistas también han sido identificados en las proteínas del suero de la leche, α-lactalbúmina (α-lactorfina) y β-lactoglobulina (Torrez-Llanez, 2005). La α-lactorfina demostró poseer una débil actividad opioide sobre la musculatura lisa, siendo capaz de provocar su contracción.
Contrariamente, los fragmentos provenientes de la digestión tríptica de la κ-caseína, conocidos como casoxinas, se comportan como antagonistas opioides (Shah, 2000). Dos casoxinas bovinas son ligandos del tipo μ con una potencia antagonista relativamente baja comparada con la naloxona (Meisel, 1998).
Las secuencias aminoacídicas correspondientes a la actividad opioide pueden encontrarse en otras fuentes de proteínas alimentarias, tales como las proteínas de la carne. Estos péptidos podrían generarse mediante el tratamiento proteolítico de dichas fuentes. La hemoglobina de la sangre bovina es un componente menor de la carne y de los productos cárnicos. Sin embargo, en algunos de ellos, la hemoglobina es un componente mayoritario. El estudio de hidrolizados pépticos de hemoglobina reveló la presencia de péptidos bioactivos con afinidad por los receptores opioides (Nyberg, 1997; Zhao et al., 1997). Han sido encontradas también tres secuencias opioides en la
283

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
β-conglicinina de la soja (“soymorfinas”), análogas a la β-casomorfina-4 humana pero con actividad más potente sobre el receptor μ. Asimismo, demostraron poseer actividad ansiolítica cuando fueron administradas por vía oral. También resultaron ser más potentes que otros péptidos opioides obtenidos de otras proteínas de origen vegetal como la rubiscolina-6, péptido δ-opioide proveniente de rubisco, o exorfinas del gluten, también activas en el receptor opioide δ (Ohinata, 2007). No obstante, aún está pendiente el hallazgo de estos péptidos como resultado de la hidrólisis ejercida por proteasas, ya sean propias del sistema gastrointestinal o exógenas.
Péptidos antimicrobianos
Debido al creciente problema de la resistencia a los antibióticos tradicionales presentada por los organismos patógenos, existe un creciente interés en la aplicación farmacológica de péptidos antimicrobianos para tratar infecciones. Sin embargo, el mecanismo de acción de estos nuevos agentes no está bien establecido. Muchos de ellos atacan la integridad de la bicapa lipídica de la membrana de los microorganismos. Además, la mayoría de los péptidos antimicrobianos son catiónicos y su interacción con la carga negativa de los fosfolípidos proveería una explicación para su especificidad hacia las membranas bacterianas. La interacción de las cargas positivas del péptido con las negativas de las membranas bacterianas es fundamental para la actividad antibacteriana (Epand & Vogel, 1999).
Fue puesto en evidencia que muchos péptidos antimicrobianos catiónicos pueden ser liberados por ruptura de proteínas intactas que pueden tener limitada actividad antimicrobiana o ninguna. Esto fue demostrado primeramente para la proteína láctea lactoferrina: la ruptura de la proteína bovina mediante hidrólisis con pepsina en condiciones ácidas libera un péptido de 25 residuos (lactoferricina B) que demuestra una potencia bacteriostática aumentada comparada con la proteína intacta. Esta región contiene aminoácidos básicos, y la actividad antimicrobiana reside en un hexapéptido compuesto por 3 residuos de Arg y 2 de Trp (Silva & Malcata, 2005; Epand & Vogel, 1999; Yamauchi et al., 1993). Distintos péptidos derivados de lactoferricina evidenciaron poder antibacteriano, antifúngico y antiviral, así como fueron útiles contra infecciones en plantas provocadas por hongos patógenos (Marcos et al., 2008)
En el suero de la leche obtenido luego de la acción de la quimosina se comprobó la existencia de péptidos de alto peso molecular de carácter básico, llamados casecidinas, que inhibían el crecimiento de bacterias patógenas y de Lactobacilos benignos. Estos péptidos derivan de αS1-caseína y κ-caseína. Uno de ellos, isracidina [(f1-23) proveniente de αS1-caseína] posee no sólo capacidad antibiótica, sino también la de estimular la resistencia del huésped contra la infección estafilocóccica (Lahov & Regelson, 1996). La digestión con pepsina durante 4 horas de αS2-caseína dio como resultado una fracción con gran actividad antibacteriana. El estudio estructural de los péptidos hallados en este hidrolizado demostró que no solamente eran ricos en aminoácidos básicos, sino que en su secuencia poseían
284

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
aminoácidos hidrofóbicos capaces de formar hélices α anfipáticas, lo que les concedería actividad antihemolítica. Estos fragmentos son los llamados (f 164-179) y (f 183-207), siendo este último el que presentó mayor actividad antibacteriana (Recio & Visser, 1999). El péptido inmunomodulatorio derivado de la β-caseína bovina, (f 193-209) demostró aumentar la actividad antimicrobiana de macrófagos de ratón (Sandre et al., 2001). Todos estos resultados respectivos a animales han sido obtenidos in vivo mediante la inyección de los péptidos; no hay estudios al presente que evidencien que tengan este efecto cuando dichos péptidos son digeridos (Silva & Malcata, 2005).
Péptidos hipocolesterolémicos
El riesgo de padecer enfermedades cardiovasculares está relacionado con una variedad de factores como la obesidad, el consumo de tabaco, la diabetes mellitus y un elevado nivel sanguíneo de lipoproteína de baja densidad (LDL), siendo este último factor un indicador del nivel de colesterol que se está depositando en el endotelio vascular. Por lo tanto, un incremento del colesterol en la sangre está directamente relacionado con la posibilidad de contraer una enfermedad cardiovascular (Wilson et al., 2008). Así pues, una disminución de la absorción intestinal de colesterol sería beneficiosa para el organismo. La absorción intestinal de colesterol se realiza a través de su solubilización en forma de micelas, constituidas por ácidos biliares y otros lípidos (Wang & Lee, 2008). Se ha observado que algunos péptidos pueden competir con el colesterol por un sitio en estas micelas, reduciendo de esta forma su solubilidad y absorción intestinal. Es decir, estos péptidos podrían considerarse hipocolesterolémicos ya que reducirían los niveles de colesterol plasmático.
Se han obtenido diversos hidrolizados proteicos de girasol utilizando pepsina, y de Brassica carinata con pepsina, pancreatina y carboxipeptidasa (Pedroche et al., 2006). En el caso del girasol se obtuvieron hidrolizados con alcalasa que resultaron de interés ya que mostraron una notable disminución en la incorporación de colesterol a la micela (Vioque et al., 2006).
Péptidos con actividad inmunomodulatoria
Los hidrolizados de proteínas lácteas pueden potenciar las funciones de la inmunidad celular, medida como la proliferación de linfocitos, la síntesis de anticuerpos y la regulación de citoquinas.
Existen péptidos presentes en la leche que afectan al sistema inmune y a la respuesta de proliferación de células. Los péptidos inmunoregulatorios obtenidos de β y αS1 caseína y α lactoalbúmina aumentan la fagocitosis y modulan la proliferación y diferenciación de linfocitos. La lactoferrina intacta o su péptido amino terminal también modulan la blastogénesis o diferenciación de linfocitos. (Torres-Llánez, 2005)
Durante la lactancia existen muchos factores que afectan el sistema inmune del neonato. En hidrolizados trípticos de leche humana se encontró actividad inmunoestimulante, debida a la presencia del hexapéptido VEPIPY de β-caseína (f54-59). También han sido identificados péptidos con actividad inmunoestimulante a
285

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
partir de caseínas bovinas: β-caseína (f63-68 y f191-193) y de αS1 caseína bovina (f194-199). Todos estos péptidos estimulan la actividad fagocítica de macrófagos humanos y murinos in vitro (Silva & Malcata, 2005).
En hidrolizados de caseína total realizados con quimosina, se encontró que la fracción soluble a pH 4,6 contiene los péptidos (f1-23) de la αS1-caseína y (f193-209) de la β-caseína, todos con actividad inmunomodulatoria (Minkiewicz et al., 2000). También se encontró un tripéptido de la κ-caseína que aumenta considerablemente la proliferación in vivo de linfocitos en sangre periférica. Mientras que la secuencia C-terminal de la β-caseína (f193-209) induce una respuesta similar en ratas (Coste et al., 1992).
También se han encontrado propiedades inmunomodulatorias en proteínas de lactosuero y sus hidrolizados. Luego de realizar digestiones con diversas enzimas proteolíticas se encontraron péptidos que incrementan significativamente la proliferación in vitro de linfocitos aislados a partir de bazo de ratón. (Mercier et al., 2004).
Péptidos transportadores de minerales
La leche y los productos lácteos son universalmente reconocidos como excelentes fuentes de calcio. Este elemento juega un importante rol en variadas etapas del metabolismo humano y se ha observado que el consumo de cantidades elevadas de calcio a temprana edad contribuye al desarrollo de una buena densidad ósea que previene la osteoporosis en la vida adulta. También se ha demostrado que existe una correlación entre la ingesta de calcio y la prevención de la hipertensión arterial (Mc Donagh & FitzGerald, 1998).
Las caseínas, proteínas mayoritarias de la leche, contienen en su secuencia numerosos residuos de serina, que unen fosfatos covalentemente por uniones monoéster. El grado de fosforilación depende de cada caseína en particular. Estos fosfatos serínicos actúan como quelantes del Ca+2 en la leche. Por ejemplo, la secuencia S(P)- S(P)- S(P)-E-E se encuentra presente en forma repetitiva en la αS1, αS2 y β caseínas (Torres-Llanez, 2005).
La elevada biodisponibilidad de Ca2+ de la leche y sus productos ha sido en parte atribuida a la producción de caseinofosfopeptidos (CPPs), péptidos fosforilados derivados de las caseínas que poseen la habilidad de unir y solubilizar minerales como el Ca2+. También pueden unir otros cationes bivalentes como Mg, Fe, Zn, Ba, Cr, Co Ni y Se. (Silva & Malcata, 2005). Poseen diferentes niveles de fosforilación y son producidos in vivo por la acción de las proteasas gastrointestinales sobre las caseínas. CPPs parecen ser resistentes a la degradación posterior, hecho debido probablemente a la elevada concentración de cargas negativas aportadas por los fosfatos. Los CCPs se acumulan en el extremo distal del intestino delgado, complejando catión y el limitando su precipitación, en donde juegan un rol en la absorción pasiva de Ca2+ y trazas de otros elementos. Por otra parte en la cavidad oral promueven la remineralización del esmalte de los dientes. Existen variados procedimientos para obtener hidrolizados enriquecidos en CPP involucrando generalmente etapas de formación de agregados
286

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
con Ca2+ seguidos de ultrafiltración (FitzGerald, 1998). Hidrolizados preparados con enzimas pancreáticas sobre caseinatos son empleados como suplemento dietario. Se hicieron hidrolizados in vitro de caseinatos sódico con muchas proteasas distintas, de origen bacteriano, animal, fúngico y vegetal, entre estas últimas la bromelaina de Ananas comosus. Se les agregó Ca2+ y se determinó que realmente los péptidos formados lo unen y solubilizan. Las diferentes proteasas ensayadas permitieron la obtención de hidrolizados con diferentes características, según la especificidad de corte (Mc Donagh & FitzGerald, 1998)
Péptidos con actividad antitrombótica
La actividad antitrombótica se origina por inhibición de la unión de fibrinógeno a una región específica del receptor en la superficie plaquetaria y por inhibición directa de la agregación de plaquetas. Las casoplatelinas son péptidos derivados del caseinoglicomacropéptido de la κ-caseína bovina (f106-116, f106-112, f112-116 y f113-116) con acción antitrombótica que actúan inhibiendo la agregación de plaquetas ADP-activadas así como también inhibiendo la unión de la cadena γ de fibrinógeno humano al sitio receptor específico en la superficie plaquetaria (Silva & Malcata, 2005).
Péptidos antitumorales
La evidencia epidemiológica sugiere que las dietas ricas en poroto de soja están asociadas con bajas tasas de mortalidad por cáncer, en particular cáncer de colon, mama y próstata, esto ha llevado a estudiar sus componentes bajo la suposición de que los mismos son responsables de prevenir la carcinogénesis (Jeong et al., 2003). Lunasin, de 43 aminoácidos, es un péptido particular y novedoso en la prevención del cáncer. Originalmente fue aislado y clonado a partir de porotos de soja pero también ha sido purificado a partir de semillas de cebada y trigo (Jeong et al., 2007). Este péptido ha demostrado ser un eficaz supresor de carcinogénesis en sistemas modelo in vitro e in vivo. Suprime la transformación de células de mamíferos causada por productos químicos carcinogénicos y cuando se aplica tópicamente inhibe la carcinogénesis de piel en ratones (Galvez et al., 2001). El extremo C-terminal de este péptido contiene 9 residuos de ácido aspártico, un motivo RGD de adhesión celular y una estructura de hélice obtenida por modelado homóloga a una región conservada de proteínas de unión a cromatina. Es de destacar que lunasin mata selectivamente células transformadas o en vías de transformación (Jeong et al., 2007).
Kim et al. (2000) realizaron hidrolizados de proteínas de soja con termoase y extrajeron los péptidos hidrofóbicos obtenidos con etanol, luego los fraccionaron por cromatografía y ensayaron la actividad anticancerígena in vitro de cada fracción midiendo su citotoxicicidad sobre una línea celular de monocitos macrófagos de ratón. De esta manera encontraron un péptido anticancerígeno de 1157 Da (X-MLPSYSPY).
Mediante hidrólisis in vitro de aislados de proteínas de soja emulando la digestión gastrointestinal con pepsina y pancreatina seguida de co-immunoprecipitación con topoisomerasa II humana fueron separados e identificados 3 péptidos inhibitorios de esta proteína (FEITPEKNPQ, IETWNPNNKP y VFDGEL). Su actividad inhibitoria
287

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
fue confirmada luego con péptidos sintéticos. Cabe señalar que las topoisomerasas son un blanco de varios agentes anticancerígenos porque su inhibición impide los procesos de proliferación celular y de diferenciación en la carcinogénesis (Wang et al., 2008).
Fraccionamiento y enriquecimiento de hidrolizados que contienen péptidos bioactivos
La explotación comercial de péptidos bioactivos, para la salud y nutrición humana, implica un desafío tecnológico y científico justificado por los beneficios potenciales que se pueden obtener de su comercialización. Para la producción de péptidos bioactivos a partir de proteínas alimentarias es necesaria la recuperación de estos péptidos, presentes en concentraciones en general bajas, a partir de hidrolizados muy complejos. La producción comercial de péptidos bioactivos ha estado limitada por la falta de tecnologías de escalado adecuadas. Se han utilizado diferentes técnicas de fraccionamiento como: separación por membrana (ultrafiltración), extracción bifásica con solventes orgánicos (Froidevaux et al., 2001) y extracción en sistema acuoso de dos fases (Murakami & Hirata, 2000) entre otros.
El proceso de hidrólisis realizado en forma convencional (en batch) presenta muchas desventajas tales como el alto costo relativo de las enzimas e ineficiencia. La introducción, en los años noventa, del reactor enzimático continuo, que integra la hidrólisis enzimática, la separación de los productos y la recuperación del catalizador en una única operación, superó algunos de los inconvenientes de los procesos en batch. Los reactores con membranas de ultrafiltración (UF) han mostrado mejor eficiencia de bioconversión enzimática y mejoras en el rendimiento y calidad de producto (uniformidad y masa molecular deseada), y pueden ser escalados fácilmente. La extracción continua de los péptidos en los reactores de membrana ha sido ampliamente empleada para proteínas lácteas. Entre otros, se han obtenido péptidos antitrombóticos (Bouhallab & Touzé, 1995), α-lactorfina (Bordenave et al., 1999). fosfopéptidos y precursores de casomorfinas (Righetti et al., 1997) utilizando reactores enzimáticos continuos.
Uno de los avances mas importantes en la separación de pequeñas biomoleculas cargadas, como péptidos, ha sido el desarrollo de membranas de nanofiltración (NF), debido a su alta selectividad en la separación de componentes en el rango de masa molecular de 300-1000 Da. Las membranas NF inorgánicas, en particular, poseen alta propiedades separativas debido a su comportamiento anfótero en función del pH. Se ha demostrado que las transmisiones de péptidos seleccionados a través de las membranas NF resultan de una combinación de los efectos de tamaño y carga y que está fuertemente afectada por las condiciones fisicoquímicas (pH y fuerza iónica). La filtración por electro-membrana (FEM) parece ser una tecnología promisoria para la producción de péptidos cargados. FEM combina la filtración por membrana tradicional con electroforesis. Se han aislado péptidos catiónicos con actividad
288

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
antimicrobiana utilizando filtración por electro membrana en batch (Bargeman et al., 2002).
Otra aproximación a la preparación de péptidos bioactivos en un proceso continuo es la proteólisis en un sistema heterogéneo en el cual la proteasa es inmovilizada en un soporte. En el contexto de la producción de péptidos bioactivos se conocen solo unos pocos escalados de procesos con enzimas inmovilizadas. Algunos ejemplos son: el uso de pepsina inmovilizada en un reactor en tanque con agitación continua para la obtención de un hidrolizado de hemoglobina enriquecido en tres péptidos bioactivos intermedios a una concentración estacionaria (Ticu et al, 2005) y la obtención de fosfopétidos que unen calcio preparados usando tripsina (Park & Allen, 1998a) o endopeptidasas específicas de ácido glutámico (Park & Allen, 1998b) inmovilizadas.
La cromatografía de intercambio en membrana ha emergido como una técnica promisoria para el enriquecimiento de fracciones peptídicas de hidrolizados de proteína. En este sistema, la proteína de interés es concentrada en un medio cromatográfico e hidrolizada in situ por una enzima apropiada. Los péptidos bioactivos resultantes son retenidos en el intercambiador iónico, mientras que los demás péptidos son lavados. Finalmente, la fracción que contiene los péptidos de interés es eluída del medio cromatográfico. Con éste método fue posible aislar y enriquecer péptidos antibacterianos catiónicos a partir de lactoferrina (Recio et al., 2000). La ventaja de estos métodos es que el aislamiento de la proteína precursora no es necesario y la enzima usada en el proceso puede ser recuperada. Esta técnica ofrece nuevas posibilidades para el enriquecimiento de péptidos con una masa molecular baja, y puede ser fácilmente escalado para dar rendimientos de gramos o aun kilogramos.
Perspectivas futuras
En vista de lo anteriormente expuesto queda evidenciada la importancia de descubrir nuevos péptidos beneficiosos para la salud a partir de hidrolizados y alimentos fermentados. Los péptidos bioactivos poseen aplicaciones muy interesantes como suplementos dietarios así como en preparaciones farmacéuticas. La identificación de nuevos compuestos bioactivos contribuiría al desarrollo de alimentos funcionales que puedan ser usados para mejorar la salud y la calidad de vida. No obstante, aún está pendiente de ser examinado con más detalle si estos péptidos bioactivos son beneficiosos y deseables como constituyentes de los alimentos o como fármacos en sí mismos. Los posibles efectos adversos asociados con los péptidos bioactivos deben ser considerados cuidadosamente, especialmente al ser incorporados en la formulación de un alimento funcional en concentraciones a las que no se llegan mediante la ingestión de los productos alimentarios originales. Sin embargo, las posibilidades de diseñar nuevos productos dietarios y fármacos naturales generan expectativas promisorias (Silva & Malcata, 2005; Korhonen & Pihlanto, 2003).
Es importante también lograr liberar en experimentos in vitro los mismos fragmentos peptídicos que se liberan in vivo durante la digestión de las proteínas de los
289

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
alimentos. La generación de péptidos bioactivos en las vías del tracto gastrointestinal, dado el daño que los péptidos puedan sufrir durante la digestión constituye una de las principales cuestiones a dilucidar. Algunos péptidos bioactivos conocidos no son absorbidos desde el tracto gastrointestinal hacia la circulación sanguínea y sus efectos probablemente sean mediados a través de receptores en la pared de la célula intestinal. En este aspecto, el conocimiento del mecanismo de acción de los péptidos es de fundamental importancia.
Por otro lado, los péptidos pueden interaccionar con otros componentes de los alimentos, especialmente si son sometidos a altas temperaturas. Es necesario conocer la formación potencial de sustancias tóxicas, alergénicas o carcinogénicas, para lo que se necesita el desarrollo de métodos analíticos modernos para estudiar la seguridad de los alimentos que contengan péptidos bioactivos, así como su biodisponibilidad en las formulaciones finales.
La producción de péptidos bioactivos en gran escala es todavía un desafío; en este aspecto, la obtención de los mismos por hidrólisis de las proteínas dietarias como alternativa a la síntesis química tradicional resulta una perspectiva interesante. Además de la hidrólisis enzimática, la fermentación microbiana representa una tecnología natural aplicable para la producción de péptidos bioactivos. La obtención de los mismos a partir de materiales crudos ricos en proteínas puede escalarse a nivel industrial empleando fermentación controlada en biorreactores. En el futuro, la producción comercial de secuencias peptídicas específicas podría emplear enzimas recombinantes y cepas microbianas seleccionadas, o aplicar alternativamente peptidasas aisladas de microorganismos, animales o vegetales. Esto también incluye el desarrollo de nuevos procesos de purificación de los productos deseados (Korhonen & Pihlanto, 2006).
Las peptidasas vegetales han sido poco exploradas como agentes capaces de desencriptar las secuencias peptídicas bioactivas presentes en proteínas alimentarias. Péptidos bioactivos se obtienen a partir de sustratos proteicos tanto de origen lácteo como de tejido muscular de porcinos, procesadas con proteasas vegetales. Se conoce la obtención de péptidos con actividad inhibitoria de la ECA por acción de enzimas de la planta Cynara cardunculus (Silva et al., 2006) y la fracciones peptídicas con esta bioactividad por la acción de las enzimas de la planta Bromelia antiacantha (Villadóniga et al., 2008) sobre proteínas lácteas. Componentes peptídicos antioxidantes se producen por la catálisis proteolítica de papaína (Saiga et al., 2003) sobre músculo porcino y por la acción de enzimas de la planta Cynara cardunculus (Silva et al., 2006) sobre proteínas lácteas. Extractos preparados a partir de látex de Ficus luschnathiana (higuerón) han sido utilizados para hidrolizar proteínas de diferentes lactosueros, obteniéndose fracciones peptídicas con actividad antimicrobiana sobre S aureus, siendo objeto de estudios tendientes a dilucidar aspectos referentes a estructuras, estabilidades y alcances de su acción (Barros, comunicación personal).
La aplicación de las mismas no sólo plantea la alternativa de obtener nuevos productos alimentarios enriquecidos en péptidos bioactivos, sino también la posibilidad de hallar nuevas secuencias peptídicas con actividades biológicas no
290

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
descubiertas aún. Una alternativa interesante que abre un amplio abanico de aplicaciones radica en el descubrimiento de nuevas secuencias aminoacídicas con funciones específicas que puedan incorporarse en otras proteínas mediante ingeniería de proteínas. Estos nuevos aspectos podrían ser aplicados no sólo en la formulación de alimentos funcionales y nutracéuticos.
Referencias
Akoh, C.C., & D.B. Min (2002) Food lipids: chemistry, nutrition, and biotechnology, 2nd edition. New York, Marcel Dekker Inc.
Awasthi Y.C., R. Sharma, J.Z. Cheng, Y. Yang, A. Sharma, S.S. Singhal & S. Awasthi (2003) Role of 4-hydroxynonenal in stress-mediated apoptosis signaling, Mol Aspects Med. 24: 219-30.
Blake, D., & P.G. Winyard (1995) Immunopharmacology of free radical species. San Diego, Academic Press.
Bordenave, S., F. Sannier, G. Ricart & J.M. Piot (1999) Continuous hydrolysis of goat whey in an ultrafiltration reactor: Generation of alpha-lactorphin, Prep. Biochem. Biotechnol. 29: 189-202.
Bouhallab S. & C. Touzé (1995) Continuous hydrolysis of caseinomacropeptide in a membrane reactor: Kinetic study and gram-scale production of antithrombotic peptides, Lait, 75: 251-8.
Chang, C.Y., K.C. Wu & S.H. Chiang (2007) Antioxidant properties and protein compositions of porcine haemoglobin hydrolysates, Food Chemistry 100: 1537-43.
Clare D.A. & H.E. Swaisgood (2000) Bioactive milk peptides: a prospectus, J. Dairy Sci., 83: 1187-95.
Cooke, M.S., M.D. Evans, M. Dizdaroglu & J. Lunec (2003) Oxidative DNA damage: mechanisms, mutation, and disease, FASEB J., 17: 1195-214.
Coste M., V. Rochet, J. Léonil, D. Mollé, S. Bouhallab & D. Tomé (1992) Identification of C-terminal peptides of bovine ß-casein that enhance proliferation of rat lymphocytes, Immunol. Lett. 33: 41-6.
Dávalos A., M. Miguel, B. Bartolomé & R. López-Fandiño (2004) Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis, J. Food Protect. 67: 1939-44.
Dröge, W. (2002) Free Radicals in the Physiological Control of Cell Function, Physiological Rev. 82: 47-95.
Dziuba J., P. Minkiewicz & D. Nalecz (1999) Biologically active peptides from plant and animal proteins, Pol. J. Food Nutr. Sci. 8: 3-16.
Dziuba, J. & M. Darewicz (2007) Food proteins as precursors of bioactive peptides - classification into families, Food Sci. Technol. Int.404.
Epand, R.M. & H.J. Vogel (1999) Diversity of antimicrobial peptides and their mechanisms of action, Biochim. Biophys. Acta, 1462: 11-28.
Fahmi, A., S. Morimura, H.C. Guo, T. Shigematsu, K. Kida & Y. Uemura (2004) Production of angiotensin I converting enzyme inhibitory peptides from sea bream
291

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
scales, Process Biochem. 39: 1195-200. FitzGerald R., B. Murray, D. Walsh (2004) Hypotensive peptides from milk proteins,
J. Nutr. 134: 980S-8S. FitzGerald, R.J. (1998) Potential uses of caseinophosphopeptides, Int. Dairy J. 8: 451-
7. Froidevaux, R., N. Nedjar-Arroume, L. Choisnard, M. Bigan, D. Guillochon. (2001)
Using an experimental design for the optimization of LVV-haemorphin-7 and VV-haemorphin-7 extraction by an organic solvent mixture in the course of bovine haemoglobin peptic hydrolysis, Biotechnol. Appl. Biochem. 33: 75-83.
Galvez, A.F., N. Chen, J. Macasieb & B.O. de Lumen (2001) Chemopreventive property of a soybean peptide (lunasin) that binds to deacetylated histones and inhibits acetylation, Cancer Res. 61: 7473-8.
Gibbs, B.F., A. Zougman, R. Masse & C. Mulligan (2004) Production and characterization of bioactive peptides from soy hydrolysate and soy-fermented food, Food Res. Int. 37: 123-31.
Glämsta E.L., A. Marklund, U. Hellman, C. Wernstedt, L. Terenius & F. Nyberg (1991) Isolation and characterization of a hemoglobin-derived opioid peptide from the human pituitary gland, Regul Pept. 34: 169-79.
Gómez-Ruiz, J.Á., M. Ramos, I. Recio (2002) Angiotensin-converting enzyme-inhibitory peptides in Manchego cheeses manufactured with different starter cultures, Int. Dairy J. 12: 697-706.
Haileselassie, S.S., B.H. Lee & B.F. Gibbs (1999) Purification and identification of potentially bioactive peptides from enzyme-modified cheese, J. Diary Sci., 82: 1612-7.
Halliwell B. & J.M.C. Gutteridge (1999) Free radicals, other reactive species and disease In: Free Radicals in Biology and Medicine (B. Halliwell & J.M.C. Gutteridge. eds.) 3rd edition, Oxford, Oxford University Press, pp. 617-783.
Halliwell, B. & J.M.C. Gutteridge (1990) Role of free radicals and catalytic metal ions in human disease: an overview, Meth. Enzymol. 186: 1-85.
Halliwell, B. & M. Whiteman (2004) Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean?, Brit. J. Pharmacol. 142: 231-55.
Halliwell, B. (1996) Antioxidants in human health and disease, Annu. Rev. Nutr. 16: 33-50.
Hong, F., L. Ming, S. Yi, L. Zhanxia, W. Yongquan & L. Chi (2008) The anti-hypertensive effect of peptides: A novel alternative to drugs, Peptides 29: 1062-71.
Ivanov V.T., A.A. Karelin, E.Y. Blischenko, M.M. Philippova & I.V. Nazimov (1998) Proteolytic degradation of hemoglobin in vivo. Role in formation of tissue specific peptide pool, Pure Appl. Chem. 70: 67-74.
Ivanov V.T., O.N. Yatskin, O.A. Kalinina, M.M. Philippova, A.A. Karelin & Y. Blishchenko (2000) Tissue-specific peptide pools, Generation and function, Pure Appl. Chem., 72: 355–63.
Ivanov, V.T., O.N. Yatskin, O.V. Sazonova, A.G. Tolmazova, K.V. Leontiev, M.M.
292

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
Philippova, A.A. Karelin & E.Y. Blishchenko (2006) Peptidomics: A modern approach to biodiversity of peptides, Pure Appl. Chem., 78: 963–75.
Je, J.Y., P.J. Park & S.K. Kimo (2005) Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate, Food Res. Int. 38: 45-50.
Jeong, H.J., J.B. Jeong, D.S. Kim, J.H. Park, J.B. Lee, D.H. Kweon, G.Y. Chung, E.W. Seo & B.O. de Lumen (2007) The cancer preventive peptide lunasin from wheat inhibits core histone acetylation, Cancer Lett. 255: 42–8.
Jeong, H.J., J.H. Park, Y. Lam & B.O. de Lumen (2003) Characterization of lunasin isolated from soybean, J. Agric. Food Chem. 51: 7901-6.
Juárez, M. (2007) Presente y futuro de los alimentos funcionales, IV JIPCA. CI-18: 32. Karelin A.A., E.Y. Blishehenko & V.T. Ivanov (1998) A novel system of peptidergic
regulation ,FEBS Lett. 428:.7-12. Kim S.E., H.H. Kim, J.Y. Kim, Y.I. Kang, H.J. Woo & H.J. Lee (2000) Anticancer
activity of hydrophobic peptides from soy proteins, Biofactors, 12: 151-5. Kim S.K., Y.T. Kim, H.G. Byun, K.S. Nam, D.S. Joo & F. Shahidi (2001) Isolation
and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin, J. Agric. Food Chem. 49: 1984-9.
Kitts, D.D. & K. Weiler (2003) Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery, Curr. Pharm.Design 9: 1309-23.
Kong B. & Y.L. Xiong (2006) Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action, J. Agric. Food Chem., 54: 6059-68.
Korhonen H. & A. Pihlanto-Leppälä (2003) Milk-Derived Bioactive Peptides: Formation and Prospects for Health Promotion En: Handbook of Functional Dairy Products. Functional Foods and Nutraceuticals (C. Shortt & J. O'Brien Eds.), Series 6.0, Florida, CRC Press.
Korhonen, H. & A. Pihlanto (2003) Food-derived bioactive peptides opportunities for designing future foods, Curr Pharm Des. 9: 1297-308.
Korhonen, H. & A. Pihlanto (2006) Bioactive peptides: production and functionality, Int. Dairy J. 16: 945-960.
Lahov, E. & W. Regelson (1996) Antibacterial and immunostimulating casein-derived substances from milk casecidin, isracidin peptides, Food Chem. Toxicol. 34: 131-45.
López, M.C. (2007) Alimentos funcionales y declaraciones de salud, desarrollo y legislación a nivel mundial y en MERCOSUR, IV JIPCA. CI: 19:33. Manso M.A., M. Miguel, J. Even, R. Hernández, A. Aleixandre & R. López-Fandiño
(2008) Effect of the long-term intake of an egg white hydrolysate on the oxidative status and blood lipid profile of spontaneously hypertensive rats, Food Chem. 109: 361-7.
Mao X.Y., J.R. Ni, W.L. Sun, P.P. Hao & L. Fan (2006) Value-added utilization of yak milk casein for the production of angiotensin-I-converting enzyme inhibitory peptides, Food Chem., 103: 1282-7.
Mao, X., F. Ren, Y. Li, Q. Nan & J. Song (2007) Comparison of the lymphoproliferation activity of yak milk casein hydrolysates hydrolyzed by
293

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
microbial-derived and animal-derived proteinase, J. Food Biochem,. 31: 289-99. Marcos, J.F., A. Muñoz, E. Pérez-Payá, S. Misra & B. López-García (2008)
Identification and rational design of novel antimicrobial peptides for plant protection, Annu. Rev. Phytopathol., 46: 273-301.
Martínez Agustin, O. & E. Martínez de Victoria Muñoz, (2006) Proteínas y péptidos en nutrición enteral, Nutr. Hosp. 21: 1-14.
Martínez Sánchez, G. (2005) Especies reactivas del oxígeno y balance redox, parte I: aspectos básicos y principales especies reactivas del oxígeno, Rev. Cub. Farm. 39: 3.
Mc Donagh, D. & R.J. FitzGerald (1998) Production of caseinophosphopeptides (CPPs) from sodium caseinate using a range of commercial protease preparations, Int. Dairy J., 8: 39-45.
Meisel H & R.J. FitzGerald (2000) Opioid peptides encrypted in intact milk protein sequences, Brit. J. Nutr. 84: S27-31.
Meisel H. (2004) Multifunctional peptides encrypted in milk proteins, Biofactors 21: 55-61.
Meisel, H. & W. Bockelmann (1999) Bioactive peptides encrypted in milk proteins: Proteolytic activation and thropho-functional properties, Antoine van Leeuwenhoek, 76: 207-15.
Meisel, H. (1998) Overview on milk-protein derived peptides, Int. Dairy J. 8: 363-73. Meisel, H. (2005) Biochemical properties of peptides encrypted in bovine milk
proteins, Curr. Med. Chem. 12: 1905-19. Meisel, H., H. Frister & E. Schlimme (1989) Biologically active peptides in milk
proteins, Z. Ernahrungswiss 28: 267-78. Mendis E, N. Rajapakse & S.K. Kim (2005) Antioxidant properties of a radical-
scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate, J Agric. Food Chem. 53: 581-7.
Mercier, A., S.F. Gauthier & I. Fliss (2004) Immunomodulating effects of whey proteins and their enzymatic digests, Int. Dairy J., 14: 175-83.
Millán, F. (2006) Peptídeos bioactivos em proteínas vegetáis de reserva, Braz. J. Food Technol. 99-102.
Minkiewicz, P., C.J. Slangen, J. Dziuba, S. Visser & H. Mioduszewska (2000) Identification of peptides obtained via hydrolysis of bovine casein by chimosin using HPLC and mass spectrometer, Milchwissenschaft 55:14-7.
Möller, N.P., K.E. Scholz-Ahrens, N. Roos & J. Schrezenmeir (2008) Bioactive peptides and proteins from foods: indication for health effects, Eur. J. Nutr. 47: 171-82.
Mukul, D., B. Kishore, P.R. Naveen & M.S. Lalit (2005) Oxidative damage of plasma proteins and lipids in epidemic dropsy patients: Alterations in antioxidant status, Biochim. Biophys. Acta 1722: 209-17.
Murakami, Y. & A. Hirata (2000) Novel process for enzymatic hydrolysis of proteins in an aqueous two-phase system for the production of peptide mixture, Prep. Biochem. Biotechnol. 30: 31-7.
Murray, B.A. & R.J. FitzGerald (2007) Angiotensin converting enzyme inhibitory
294

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
peptides derived from food proteins: biochemistry, bioactivity and production, Curr. Pharm. Design 13: 773-91.
Nass, N., R. Schoeps, R. Ulbrich-Hofmann, A. Simm, L. Hohndorf, C. Schmelzer, K. Raith, R.H.H. Neubert & K. Eder (2008) Screening for nutritive peptides that modify cholesterol 7 alpha-hydroxylase expression, J Agric Food Chem. 56: 4987-94.
Nyberg, F., K. Sanderson & E.L. Glämsta (1997) The hemorphins: a new class of opioid peptides derived from the blood protein haemoglobin, Biopolymers, 43: 147-56.
Ohinata, K., S. Agui & M. Yoshikawa (2007) Soymorphins, new μ opioid peptides from soy β-conglycinin β-subunit, have anxiolytic activities, Biosci. Biotech. Biochem., 71: 2618-21.
Oleszczuk, J., B. Wawrzycka, J.G. Maj & H. Meisel (1997) Biochemical properties of bioactive peptides derived from milk proteins: Potential nutraceuticals for food and pharmaceutical applications, Livest. Prod. Sci. 50: 125-38.
Park O. & J.C. Allen (1998a) Preparation of Phosphopeptides Derived from αs-Casein and ß-Casein Using Immobilized Glutamic Acid-Specific Endopeptidase and Characterization of Their Calcium Binding, J. Dairy Sci. 81: 2858–65.
Park O., H.E. Swaisgood & J.C. Allen (1998b) Calcium Binding of Phosphopeptides Derived from Hydrolysis of as-Casein or β-Casein Using Immobilized Trypsin, J. Dairy Sci, 81: 2850–7.
Pedroche J, M.M. Yust, H. Lqari, C. Megías, J Girón-Calle, M. Alaiz, J. Vioque & F. Millan (2006) Production of Brassica carinata protein hydrolyzates with a high Fischer's ratio using immobilized proteases, J. Agric. Food Chem. 54: 7621-7.
Pena-Ramos, E.A. & Y.L. Xiong (2002) Antioxidant Activity of Soy Protein Hydrolysates in a Liposomal System, J. Food Sci. 67: 2952-6.
Perpetuo, E.A., L. Juliano & I. Lebrun (2003) Biochemical and pharmacological aspects of two bradykinin-potentiating peptides obtained from tryptic hydrolysis of casein, J. Protein Chem. 22: 601–6.
Pihlanto-Lepälä, A. (2001) Bioactive peptides derived from bovine whey proteins: opioid an ACE-inhibitory peptides, Trends Food Sci. Technol. 11: 347-56.
Pripp A.H., T. Isaksson, L. Stepaniak, T. Sorhaug & Y. Ardö (2005) Quantitative structure activity relationship modelling of peptides and proteins as a tool in food science, Trends Food Sci. Technol. 16: 484-94.
Qian, Z.J., J.Y. Je & S.K. Kim (2007) Antihypertensive effect of angiotensin I converting enzyme-inhibitory peptide from hydrolysates of big eye tuna dark muscle, Thunnus obesus J. Agric. Food Chem, 55: 8398-403.
Qian, Z.J., W.K Jung, H.G. Byun, & S.K. Kim (2008) Protective effect of an antioxidative peptide purified from gastrointestinal digests of oyster, Crassostrea gigas against free radical induced DNA damage, Biores. Technol. 99: 3365-71.
Recio, I. & S. Visser (1999) Identification of two distinct antibacterial domains within the sequence of bovine αS2-casein, Biochim. Biophys. Acta-Gen. Subj. 1428: 314-26.
Recio I., C.J. Slangen & S. Visser (2000) Method for the production of antibacterial peptides from biological fluids at an ionic membrane. Application to the isolation of
295

Capítulo 20. Productos bioactivos obtenidos por proteólisis (nutracéuticos y alimentos funcionales)
nisin and caprine lactoferricin, Lait 80: 187–95. Righetti, P.G., F. Nembri, A. Bossi & M Mortarino (1997) Continuous enzymatic
hydrolysis of beta-casein and isoelectric collection of some of the biologically active peptides in an electric field, Biotechnol. Progr. 13: 58-64.
Rutherfurd-Markwick, K.J. & P.J. Moughan (2005) Bioactive peptides derived from food, J. AOAC Int. 88: 955-66.
Saiga, A., S. Tanabe, T. Nishimura (2003) Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment, J. Agric. Food Chem. 51: 3661-7.
Sakanaka, S., Y. Tachibana, N. Ishihara & L.R. Juneja (2004) Antioxidant activity of egg-yolk protein hydrolysates in a linoleic acid oxidation system, Food Chem. 86: 99-103.
Sandre, C., A. Gleizes, F. Forestier, R. Gorges-Kergot, S. Chilmonczyk, J. Leonil, M.C. Moreau & C. Labarre (2001) A peptide derived from bovine β-casein modulates functional properties of bone marrow-derived macrophages from germfree and human flora-associated mice, J. Nutr. 131: 2936-42.
Séverin S. & X. Wenshui (2005) Milk biologically active components as nutraceuticals: review, Crit. Rev. Food Sci. Nutr. 45: 645-56.
Shah, N.P. (2000) Effects of milk-derived bioactives: an overview Brit. J. Nutr. 84: S3-10.
Sies, H. (1997) Oxidative stress: oxidants and antioxidants, Exp. Physiol. 82: 291-5. Silva, S., A. Pihlanto & F.X. Malcata (2006) Bioactive peptides in ovine and caprine
cheeselike systems prepared with proteases from Cynara cardunculus, J. Dairy Sci. 89: 3336-44.
Silva, S.V. & F.X. Malcata (2005). Caseins as source of bioactive peptides, Int. Dairy J. 15: 1-15.
Smacchi, E. & M. Gobbetti (1998) Peptides from several Italian cheeses inhibitory to proteolytic enzymes of lactic acid bacteria, Pseudomonas fluorescens ATCC 948 and to the angiotensin I-converting enzyme, Enz. Microb. Technol. 22: 687-94.
Svedberg J., J. de Haas, G. Leimenstoll, F. Paul & H. Teschemacher (1985) Demonstration of beta-casomorphin immunoreactive materials in in vitro digests of bovine milk and in small intestine contents after bovine milk ingestion in adult humans, Peptides 6:825-30.
Teschemacher, H., G. Koch & V. Brantl (1997) Milk protein-derived opioid receptor ligands, Biopolymers 43: 99-117.
Teschemacher H. (2003) Opioid receptor ligands derived from food proteins, Curr. Pharm. Design 9: 1331-44.
Thomas, P.D., T.W. Nichols & A. Angstadt (2001) Dietary bioactive peptides in maintaining intestinal integrity and function, Am. J. Gastroenterol. 96: S311.
Torres-Llanez, M., B. Vallejo-Cordoba & A. González-Córdova (2005) Péptidos bioactivos derivados de las proteínas de la leche, Arch. Latinoamer. Nutr. 55: 111-7
Vermeirssen, V., J. Van Camp & W. Verstraete (2004) Bioavailability of angiotensin I converting enzyme inhibitory peptides, Brit. J Nutr. 92: 357-66.
296

Enzimas proteolíticas de vegetales superiores. Aplicaciones industriales
Villadóniga C., L. Macció, D. Vallés, M. Barros, A.M.B. Cantera (2008). Hidrólisis de proteínas de lactosuero con proteasas de Bromelia antiacantha. Evaluación de las propiedades antihipertensivas de los productos de hidrólisis. En: Actas del III Encuentro Regional de Biocatálisis y Biotransformaciones, octubre 2008, San Luis, Argentina, CO-8.
Vioque, J., J.Pedroche, M.M. Yust, H. Lqari, C. Megías, J. Girón-Calle, M Alaiz & Wang, D.Q. & S.P. Lee (2008) Physical chemistry of intestinal absorption of biliary
cholesterol in mice, Hepatology 48: 177-85. Wang, W., S.G. Rupasinghe, M.A. Schuler & E. Gonzalez de Mejia (2008)
Identification and characterization of topoisomerase II inhibitory peptides from soy protein hydrolysates, J. Agric. Food Chem., 56: 6267-77.
Wilson, P.W.F., S.R. Bozeman, T.M. Burton, D.C. Hoaglin, R. Ben-Joseph & C.L. Pashos (2008) Prediction of first events of coronary heart disease and stroke with consideration of adiposity, Circulation 118: 124-30.
Wu, H.C., H.M. Chen & C.Y. Shiau (2003) Free amino acids and peptides as related to antioxidant properties in protein hydrolyzates of mackerel (Scomber austriasicus), Food Res. Int. 36: 949-57.
Yamauchi, K., M. Tomita, T. Giehl & R.T. Ellison (1993) Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment, Infect. Immun. 61: 719-28.
Zhao, Q., I. Garreau, F. Sannier & J.M. Piot (1997) Opioid peptides derived from hemoglobin: hemorphins, Biopolymers 43: 75-98.
297




















