
Effects of UV-B radiation in morpho-genetic characters of Gnaphalium luteo-album
6
Effects of UV-B radiation in morpho-genetic characters of Gnaphalium luteo-album Pedro Cuadra a, * , Daniela Vargas a , Víctor Fajardo a , Raúl Herrera b a Universidad de Magallanes, Facultad de Ciencias, P.O. Box 113-D, Punta Arenas, Chile b Laboratorio de Fisiología Vegetal y Genética Molecular, Instituto de Biología Vegetal y Biotecnología, Universidad de Talca, Casilla 747, Talca, Chile article info Article history: Received 28 January 2010 Received in revised form 22 June 2010 Accepted 29 June 2010 Available online 3 July 2010 Keywords: Plants’ response Gnaphalium DNA damage Ultraviolet-B radiation Polymorphism abstract Genetic analyses of Gnaphalium’s DNA showed a high degree of polymorphism in UV-B irradiated plants when compared to controls. Among the five tested primers the four ISSR primers selected for this analysis generated a total of 189 fragments. A high proportion of polymorphic bands ranging from 70% to 28% were found using these ISSR markers. Nei and Li similarity indexes [1] were used to evaluate genetic divergence among plants. The dendrograms obtained using these markers efficiently separate plants from different treatments. A linear relationship was observed between UV-B dose and percentage of dissimi- larity which may be related to DNA damage caused by the different UV-B treatments. Ó 2010 Elsevier B.V. All rights reserved. 1. Introduction In the Austral zone of South America the enhanced level of UV-B radiation reaching the ground has been affecting living organisms by producing several biochemical and physiological changes [2]. The most affected organisms are plants which have demonstrated a complete suite of responses to deal with high levels of this harm- ful radiation. Besides other reactions, the high absorption by pro- teins and nucleic acids has induced structural changes which may result in phenotype alterations [3–5]. Production of DNA photoproducts is another well-known effect which yields cyclobu- tane type pyrimidine dimmer [6–8]. Changes in the secondary metabolites content have been dem- onstrated in several studies [9–12] as one of the plants’ responses to UV-B radiation. For instance in Gnaphalium species, liphophilic flavonoids from the leaf surface have been involved in plant pro- tection against UV-B radiation [2]. Due to the particular structural and absorptive features of these compounds, it is considered that these molecules are participating in the chemical defense of plants. Flavonoids have functionally evolved with plants and one of their most important roles might be that of absorbing compounds, screening UV solar radiation as natural filters. Similarly, some enzymes from the phenylpropanoid biosyn- thetic pathway have also been involved in plant responses to UV-B radiation [13–15]. When these UV-B absorbing compounds accumulate on the leaf epidermal layers they act as UV filters and may play an important role in counteracting the harmful ef- fects of this radiation. This plant response seems to be particularly directed at avoiding CPD formation when they are under high levels of solar UV. The effectiveness of these compounds is demon- strated by reducing the CPD’s induction in exposed plants [6]. In general plants activate different mechanisms such as photoreacti- vation, excision repair of damaged bases or replicative bypass to reduce the amount of lesions on DNA strand. In another study, Kunz et al. [16] explained that plants counter- act cell damage by attenuating the UV dose received through accu- mulation of UV-absorbing secondary metabolites which neutralize reactive oxygen species (ROS) produced by UV radiation. Likewise, Qaderi et al. [17] reported that enhanced UV-B radiation increased the amount of UV-B absorbing compounds at epicuticular wax but lowered ethylene evolution more in the reproductive stage than in the vegetative stage. More evidence of the effects of UV radiation in the quantity and quality of leaf flavonoids is given by Hashiba et al. [12]. Two ten- dencies were revealed in this exclusion experiment: flavonoid accumulation decreases with an increase in the degree of exclu- sion. They concluded from the weak correlations between UV-B intensity and flavonoid accumulation under the natural habitat conditions that various microenvironmental factors may influence the production of flavonoids. Conversely, Albert et al. [18] found that plants from UV-B environments have a higher content of UV-B absorbing compounds compared with reduced UV-B treatment. On the other hand, the effects on DNA have been studied on sev- eral systems [19]. In fact, UV-B radiation produces multiple photo- products [20] and causes severe mutations during replication [16]. 1011-1344/$ - see front matter Ó 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.jphotobiol.2010.06.013 * Corresponding author. Tel.: +56 61 207053. E-mail address: [email protected] (P. Cuadra). Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75 Contents lists available at ScienceDirect Journal of Photochemistry and Photobiology B: Biology journal homepage: www.elsevier.com/locate/jphotobiol
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of UV-B radiation in morpho-genetic characters of Gnaphalium luteo-album

Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology B: Biology
journal homepage: www.elsevier .com/locate / jphotobiol
Effects of UV-B radiation in morpho-genetic characters of Gnaphalium luteo-album
Pedro Cuadra a,*, Daniela Vargas a, Víctor Fajardo a, Raúl Herrera b
a Universidad de Magallanes, Facultad de Ciencias, P.O. Box 113-D, Punta Arenas, Chileb Laboratorio de Fisiología Vegetal y Genética Molecular, Instituto de Biología Vegetal y Biotecnología, Universidad de Talca, Casilla 747, Talca, Chile
a r t i c l e i n f o a b s t r a c t
Article history:Received 28 January 2010Received in revised form 22 June 2010Accepted 29 June 2010Available online 3 July 2010
Keywords:Plants’ responseGnaphaliumDNA damageUltraviolet-B radiationPolymorphism
1011-1344/$ - see front matter � 2010 Elsevier B.V. Adoi:10.1016/j.jphotobiol.2010.06.013
* Corresponding author. Tel.: +56 61 207053.E-mail address: [email protected] (P. Cuadra)
Genetic analyses of Gnaphalium’s DNA showed a high degree of polymorphism in UV-B irradiated plantswhen compared to controls. Among the five tested primers the four ISSR primers selected for this analysisgenerated a total of 189 fragments. A high proportion of polymorphic bands ranging from 70% to 28%were found using these ISSR markers. Nei and Li similarity indexes [1] were used to evaluate geneticdivergence among plants. The dendrograms obtained using these markers efficiently separate plants fromdifferent treatments. A linear relationship was observed between UV-B dose and percentage of dissimi-larity which may be related to DNA damage caused by the different UV-B treatments.
� 2010 Elsevier B.V. All rights reserved.
1. Introduction
In the Austral zone of South America the enhanced level of UV-Bradiation reaching the ground has been affecting living organismsby producing several biochemical and physiological changes [2].The most affected organisms are plants which have demonstrateda complete suite of responses to deal with high levels of this harm-ful radiation. Besides other reactions, the high absorption by pro-teins and nucleic acids has induced structural changes whichmay result in phenotype alterations [3–5]. Production of DNAphotoproducts is another well-known effect which yields cyclobu-tane type pyrimidine dimmer [6–8].
Changes in the secondary metabolites content have been dem-onstrated in several studies [9–12] as one of the plants’ responsesto UV-B radiation. For instance in Gnaphalium species, liphophilicflavonoids from the leaf surface have been involved in plant pro-tection against UV-B radiation [2]. Due to the particular structuraland absorptive features of these compounds, it is considered thatthese molecules are participating in the chemical defense of plants.Flavonoids have functionally evolved with plants and one of theirmost important roles might be that of absorbing compounds,screening UV solar radiation as natural filters.
Similarly, some enzymes from the phenylpropanoid biosyn-thetic pathway have also been involved in plant responses toUV-B radiation [13–15]. When these UV-B absorbing compoundsaccumulate on the leaf epidermal layers they act as UV filters
ll rights reserved.
.
and may play an important role in counteracting the harmful ef-fects of this radiation. This plant response seems to be particularlydirected at avoiding CPD formation when they are under highlevels of solar UV. The effectiveness of these compounds is demon-strated by reducing the CPD’s induction in exposed plants [6]. Ingeneral plants activate different mechanisms such as photoreacti-vation, excision repair of damaged bases or replicative bypass toreduce the amount of lesions on DNA strand.
In another study, Kunz et al. [16] explained that plants counter-act cell damage by attenuating the UV dose received through accu-mulation of UV-absorbing secondary metabolites which neutralizereactive oxygen species (ROS) produced by UV radiation. Likewise,Qaderi et al. [17] reported that enhanced UV-B radiation increasedthe amount of UV-B absorbing compounds at epicuticular wax butlowered ethylene evolution more in the reproductive stage than inthe vegetative stage.
More evidence of the effects of UV radiation in the quantity andquality of leaf flavonoids is given by Hashiba et al. [12]. Two ten-dencies were revealed in this exclusion experiment: flavonoidaccumulation decreases with an increase in the degree of exclu-sion. They concluded from the weak correlations between UV-Bintensity and flavonoid accumulation under the natural habitatconditions that various microenvironmental factors may influencethe production of flavonoids. Conversely, Albert et al. [18] foundthat plants from UV-B environments have a higher content ofUV-B absorbing compounds compared with reduced UV-Btreatment.
On the other hand, the effects on DNA have been studied on sev-eral systems [19]. In fact, UV-B radiation produces multiple photo-products [20] and causes severe mutations during replication [16].

P. Cuadra et al. / Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75 71
The most common DNA damage is the dimerization of adjacentpyrimidine bases. UV-B induces the formation of two classes ofDNA photoproducts: the CPD and the pyrimidine 6–4 pyrimidonephotoproducts [7].
In this context, Matallana et al. [21] quantified the formation ofUV-induced photolesions in both isolated and cellular DNA. Theyshowed the formation of cytosine-containing photoproducts fromisolated DNA. This effect on the genome composition was furtherconfirmed in living cells [22], and due to the fact that cytosine-con-taining photoproducts are highly mutagenic, species with genomesexhibiting a high guanine–cytosine content are expected to bemore sensitive to UV-induced mutagenesis.
Additionally, Kang et al. [23] found that plants grown under vis-ible radiation with supplemental UV-B radiation were severaltimes taller than plants grown without UV-B. They also reporteda significant negative correlation between CPD levels induced byUV-B exposure and the content of UV-absorbing compounds. Like-wise, Hidema and Kumagai [9] reported that although UV-Babsorbing compounds are effective in reducing CPD induction inplants exposed to a challenge UV-B dose, certain levels of CPDare maintained constitutively in plants grown under light condi-tions with UV. They concluded that the systems for repairingDNA damage are essential to enable plants to cope with solar radi-ation containing UV-B. This CPD photolyase activity is a determin-ing factor in providing an explanation for the differences in UV-Bsensitivity among plants and cultivars.
They also suggested that any substitution of one or two bases inthe CPD photolyase gene can alter the activity of the enzyme andhence the associated resistance of the plant to UV-B radiation.According to Gerhart et al. [3] many UV-B effects are an indirectconsequence of UV-B radiation, dependent on increases in ROSmediated by UV-B.
Conversely, Hectors et al. [4] studied chronic UV-B effects inArabidopsis. They found several effects on morphology which weredescribed as redistribution rather than a cessation of growth. Theanalyses on gene expression indicated that plant morphogenesisinduced by UV-B is associated with subtle changes in phytohor-mone balance and is functionally uncoupled from stress responses.Similar effects, relating UV-B doses and the amount of CPDs in-duced, can be found in Zhou et al. [24]. In this study a DNA damagemodel as a result of UV-B was established. They demonstrated thata pre-treatment with baicalein reduced the amount of UV-B gener-ated CPDs, therefore baicalein can be used as a protective sub-stance against UV-B radiation.
In a previous work we analyzed for the first time the effects ofUV-B radiation on the leaf surface flavonoids in Gnaphalium vira-vira [2] and Gnaphalium luteo-album [25]. G. luteo-album plantsare cosmopolitan shrubs which also grow in Central Chile in se-verely degraded soils. Because of their location on the leaf surface,as well as in epidermal vacuoles, it seemed likely that they wereinteracting with the environment and, thus, irradiation using UV-B light could modify their content. Other important alterationson photosynthetic pigments, morphology and biomass wereobserved in the study of these plants. In order to find out moreabout these changes we set up experiments to analyze the effectsof UV-B radiation on Gnaphalium’s DNA and on UV-absorbingcompounds.
2. Materials and methods
2.1. Plant material
Seeds of G. luteo-album were obtained from our own seed bank.They were sown in a soil mixture of sand (40%) and compost (60%).After 90 days of growth they were individually potted using the
same soil mixture. They were then moved into the UV-B chamberthe following day. Plants were watered every 1–2 days, ambientdaytime temperatures inside the UV-B chamber were 20–32 �Cand relative humidity 38–42%. For all seven measurements leafsamples were taken from 140 leaves developed in an enhancedUV-B environment.
2.2. Light sources
In addition to greenhouse sunlight (PAR: 1200 lmol m�2 s�1),UV radiation was provided by four fluorescent tubes (Philips TL20 W/12; 1220 mm � 1000 mm). In order to keep the spectralquality of irradiance output relatively constant these tubes were70 h pre-burnt [26]. The UV-B radiation supplementation was ob-tained by shielding these tubes with a presolarized (7 h) celluloseacetate film (0.075 mm thickness) which cuts off all wavelengthsbelow 280 nm. To avoid UV photo degradation of acetate film,the filters were changed after 90 h exposure [26,27].
2.3. UV treatments
Plants were exposed to 8 h of UV-B irradiation daily and har-vested once a week after different times of exposure. Lamps wereplaced on a mobile rack and hung at 0.30 m above the plants. Wechose this distance because it produces a daily UV-B radiation flu-ence rate of 4.5 m Wm�2, which is within the average values thathave been measured in the Punta Arenas area during the last5 years (2.23–14.21 m Wm�2). After irradiation, the UV absorbingfilms were removed in order to give the same light conditions toall plants.
2.4. Radiation measurements
The spectral irradiance levels of plant height below the lampswere measured with a photometer/radiometer (Solar Light Co.PMA 2200 1.10). The spectral distribution of UV radiation providedby these UV-B tubes has been previously reported [2,25]. The radi-ometer was calibrated using a NIST traceable 1000 W tungsten fil-ament quartz halogen lamp.
2.5. Experimental design
The experimental pattern used in this study corresponded to arandomized complete block design with several variables. Fortyfour plants (90 days old) were distributed in two groups insidethe experimental chamber: control group (�UVB) and treatedgroup (+UVB). A UV absorbing plastic film was used to isolate bothgroups. Pot positions were randomized within each group every2 days to minimize position effects. The lamp rack height was ad-justed once a week to maintain the UV irradiance levels.
2.6. UV-B absorbing compounds
Leaves used in the absorbance measurements were taken fromthe top of plants (first stage). Internal UV-B absorbing phenolicswere extracted from leaves by grinding them using a pestle andmortar with 2 mL of MeOH:H2O:HCl = 79:20:1 (v/v) [28,29].Homogenates combined with further washing of the pestle andmortar with 1 mL of the same solvent mixture were centrifugedat 3000 rpm for 10 min. Supernatants were then filtered (What-man No. 1) and evaporated to dryness at 40 �C. Residues wereredissolved in MeOH and poured into a graduated cylinder to ad-just all samples at the same volume.

Table 2Similarity 413 indexes among plants after differentamounts of UV-B radiation.
Irradiation time (days) % of similarity
7 7014 4621 3628 28
72 P. Cuadra et al. / Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75
2.7. Chromatographic analysis
Thin layer chromatographic (TLC) analyses of MeOH extractswere carried out using cellulose plates (Merck, 0.1 mm) usingBAW = 4:1:5 (v/v) as eluent system. Compounds were detected asblue spots by developing the plates with Folin & Ciocalteu’sreagent.
2.8. Isolation of chromosomal DNA from leaves
Total plant DNA for PCR was isolated from 188 first stage leavesusing a CTAB protocol described by Doyle and Doyle [30] and mod-ified as follows. Frozen tissue (0.5–0.75 g) was ground in a pestleand mortar in liquid nitrogen and homogenized in 5 mL of pre-heated (60 �C) extraction buffer containing: 1 M Tris–HCl (pH8.0); 1.4 M NaCl; 20 mM EDTA; CTAB 3% (w/v). The mixture wasincubated for 20 min at 65 �C with occasional mixing by gentleswirling. DNA was extracted twice with an equal volume of chloro-form:isoamyl alcohol (24:1 v/v) and precipitated with one volumeof cold isopropanol. The precipitated DNA was extracted with aglass hook, washed in 70% (v/v) ethanol for 5 min, dried at roomtemperature and re-suspended in autoclaved water. Contaminat-ing RNA was removed by digestion with RNAse A. The DNA wasfurther purified using chloroform/isoamyl alcohol extraction andre-precipitated with ethanol and re-suspended in autoclavedwater.
2.9. RAPD and inter simple sequence repeat PCR
ISSR primers were obtained from The University of BritishColumbia (Vancouver, Canada) set # 9 and listed in Table 1. Theywere used for PCR amplification with the following protocol. PCRreaction mixture (20 lL) containing: one unit of Taq DNA polymer-ase, 1.5 mM MgCl2 buffer, 0.2 mM of each dNTP, with 0.3 lM of asingle primer and 10 ng of genomic DNA. PCR amplifications wereperformed with a MJR PTC-100 Thermocycler using the followingprogram: 30 cycles of 1 min at 94 �C, 2 min at 55 �C and 30 s at72 �C, with a final 5 min period at 72 �C. ISSR are complementaryto genomic microsatellites sequence and contain three bases‘‘anchor” that ensure the primers annealing to either the 50 or30 end [31].
2.10. Reproducibility and separation of PCR products
The amplification procedure was carried out in duplicate eachtime. Bands were only considered reproducible when the sameDNA pattern was obtained in at least two amplification runs usingDNA isolated on different occasions. ISSR-PCR products were sepa-rated by electrophoresis on 2% agarose gels, stained with ethidiumbromide and displayed using a UV transilluminator [32].
2.11. Data analysis
ISSR markers were treated throughout as ‘genetic’ phenotypes.The DNA profiles were recorded with the presence (1) or absence
Table 1List of primer sequences used for ISSR analysis.
ISSR primerscode
DNA sequence 50 ? 30
812 GAG AGA GAG AGA GAG AA820 GTG TGT GTG TGT GTG TC825 ACA CAC ACA CAC ACA CT834 AGA GAG AGA GAG AGA GYT855 ACA CAC ACA CAC ACA CYT
(0) of bands and the results compiled in a data matrix table. Adistance measurement was calculated from the binary ISSR datamatrix table using correlation coefficient (CORR). These distanceswere used to carry out principal components analysis (PCA) usingNTSYSpc [33].
2.12. Statistical analysis
Data of UV-B absorbing compounds were analyzed using proce-dures for a randomized complete block design. Statistical assess-ments (ANOVA, LSD test) were performed for all absorbancemeasurements using SAS 8.2 System Statistical Package.
3. Results
3.1. Internal UV-B absorbing compounds
The UV spectra of MeOH leaf extracts showed two majorabsorption peaks at 301, 304 nm and minor peaks at 395, 419and 448 nm. TLC analysis of these extracts revealed the presenceof 2–3 compounds with a light blue or green fluorescence underUV/NH3 (Rf = 0.79 and 0.62), typical of hydroxycinnamic deriva-tives. Because of their UV absorption spectra (270–310 nm) thesecompounds are likely to participate in plant protection againstUV-B radiation.
In order to compare the effects of UV-B radiation on the internalUV-B absorbing compounds (epidermal flavonoids and hydroxy-cinnamic esters) quantitation was carried out by measuring theMeOH extracts absorbance at 300 nm. Table 3 shows that treatedplants have a higher absorbance than non-irradiated plants andthat the differences between internal UV-B absorbing pigmentsin both plant groups are statistically significant after 21 days oftreatment, when the absorbance of treated plants is 58% higherthan in control plants.
Apparently plants cope with low levels of UV-B radiation butwhen these levels exceed their capacity to dissipate energy, plantsmust activate other mechanisms to deal with enhanced levels ofUV-B radiation.
3.2. Analysis of the polymorphisms
Extracted DNA from different leaves was bulked for PCR analy-sis and amplifications were performed in triplicates. These analy-ses were carried out using five different primer sequences (ISSR)on nine plants UV-B irradiated. In general primers are used in
Table 3Effect of UV-B radiation on internal UV-B absorbing compounds (A300)a,b.
Control Treatment F P
0.441 ± 0.020 0.762 ± 0.020 11.64 0.0001
a Mean concentration (UA; N = 10) and standard error of internal UV-B absorbingcompounds after 21 days of treatment.
b Degrees of freedom = 1; F and P (determined in ANOVA) are given for UV-Babsorbing compounds by treatment.
P. Cuadra et al. / Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75 73
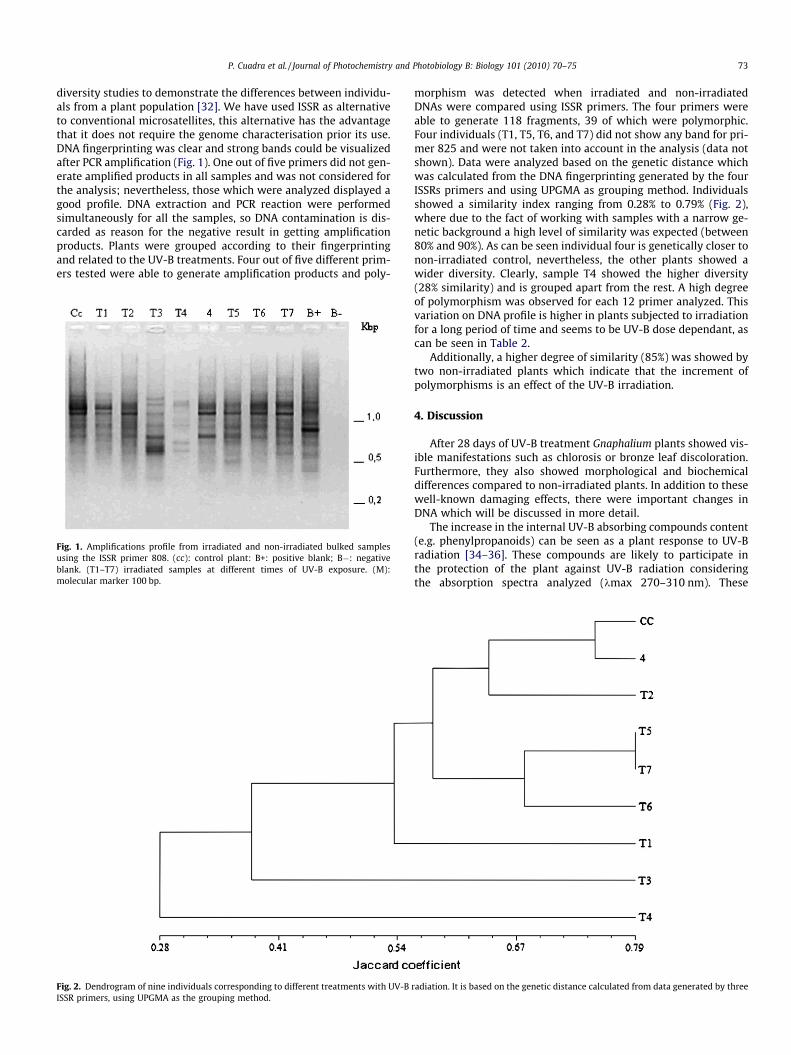
diversity studies to demonstrate the differences between individu-als from a plant population [32]. We have used ISSR as alternativeto conventional microsatellites, this alternative has the advantagethat it does not require the genome characterisation prior its use.DNA fingerprinting was clear and strong bands could be visualizedafter PCR amplification (Fig. 1). One out of five primers did not gen-erate amplified products in all samples and was not considered forthe analysis; nevertheless, those which were analyzed displayed agood profile. DNA extraction and PCR reaction were performedsimultaneously for all the samples, so DNA contamination is dis-carded as reason for the negative result in getting amplificationproducts. Plants were grouped according to their fingerprintingand related to the UV-B treatments. Four out of five different prim-ers tested were able to generate amplification products and poly-
Fig. 1. Amplifications profile from irradiated and non-irradiated bulked samplesusing the ISSR primer 808. (cc): control plant: B+: positive blank; B�: negativeblank. (T1–T7) irradiated samples at different times of UV-B exposure. (M):molecular marker 100 bp.
Fig. 2. Dendrogram of nine individuals corresponding to different treatments with UV-BISSR primers, using UPGMA as the grouping method.
morphism was detected when irradiated and non-irradiatedDNAs were compared using ISSR primers. The four primers wereable to generate 118 fragments, 39 of which were polymorphic.Four individuals (T1, T5, T6, and T7) did not show any band for pri-mer 825 and were not taken into account in the analysis (data notshown). Data were analyzed based on the genetic distance whichwas calculated from the DNA fingerprinting generated by the fourISSRs primers and using UPGMA as grouping method. Individualsshowed a similarity index ranging from 0.28% to 0.79% (Fig. 2),where due to the fact of working with samples with a narrow ge-netic background a high level of similarity was expected (between80% and 90%). As can be seen individual four is genetically closer tonon-irradiated control, nevertheless, the other plants showed awider diversity. Clearly, sample T4 showed the higher diversity(28% similarity) and is grouped apart from the rest. A high degreeof polymorphism was observed for each 12 primer analyzed. Thisvariation on DNA profile is higher in plants subjected to irradiationfor a long period of time and seems to be UV-B dose dependant, ascan be seen in Table 2.
Additionally, a higher degree of similarity (85%) was showed bytwo non-irradiated plants which indicate that the increment ofpolymorphisms is an effect of the UV-B irradiation.
4. Discussion
After 28 days of UV-B treatment Gnaphalium plants showed vis-ible manifestations such as chlorosis or bronze leaf discoloration.Furthermore, they also showed morphological and biochemicaldifferences compared to non-irradiated plants. In addition to thesewell-known damaging effects, there were important changes inDNA which will be discussed in more detail.
The increase in the internal UV-B absorbing compounds content(e.g. phenylpropanoids) can be seen as a plant response to UV-Bradiation [34–36]. These compounds are likely to participate inthe protection of the plant against UV-B radiation consideringthe absorption spectra analyzed (kmax 270–310 nm). These
radiation. It is based on the genetic distance calculated from data generated by three

74 P. Cuadra et al. / Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75
findings may be related to those reported in literature about UV-Beffects on plant flavonoid content. Many years ago, Wellmann [37]demonstrated that UV light stimulates flavonoid glycoside synthe-sis in Petroselinum hortense cell suspension cultures. More recentstudies show that plants accumulated unidentified UV-B absorbingcompounds pigments (reported as absorbance at 300 nm) afterirradiation with UV-B light. For instance, Kunz et al. [16] pointedout that plants counteract cell damage by attenuation of the UV-B dose received through accumulation of UV-absorbing secondarymetabolites. Another piece of experimental evidence relating tothe role of phenylpropanoids (e.g. hydroxycinnamic esters) in plantUV-B protection is reported by Burchard et al. [38]. In this study,authors analyzed the contribution of these metabolites to the epi-dermal shielding of rye primary leaves. Using chlorophyll fluores-cence measurements they investigated the relationship betweenepidermal UV shielding and the content of flavonoids and hydroxy-cinnamates. A good correlation was found between the epidermalUV-A and UV-B absorbance and the content of flavonoids. Theabaxial epidermis of young leaves showed a significantly higherabsorption as they are more exposed to UV. All these plant re-sponses to UV-B radiation are based on the absorption bands ofhydroxycinnamic esters being within the UV-B range. All these re-sponses can be explained on the basis of the UV absorption of thesemetabolites but also on biosynthetic grounds as they are precur-sors in the flavonoid biosynthetic pathway.
These increases in hydroxycinnamic derivatives are presumablylinked to increases in enzymatic activity and changes at genetic le-vel. In fact, several key enzymes of the phenylpropanoid biosyn-thetic pathway (e.g. PAL, CHS) have been shown to be specificallyinduced by UV-B radiation [39–41]. The role of UV-B light on geneactivation has also been reported [42,43].
The calculated indexes of treated plants showed at most 70% ofsimilarity between the controls and the irradiated plants afterbeing given the UV-B treatments (Table 2), indicating a gradual ef-fect at DNA level due to different UV-B radiation doses and changesat nucleotide sequences. A high degree of polymorphism was ob-served for each primer analyzed. This variation in DNA profile isgreater in more irradiated plants and seems to be UV-B dosedependant, as seen in Table 2. Plant individuals subjected to UV-B radiation increased the genetic variation, emphasizing particulardifferences. The intraspecific genetic variation on the genome ob-served in this study may be related to the morphological differ-ences (reduction in leaf area and leaf length, stem elongation,data not shown) observed in plants from different treatments.These findings tally well with those found by Usmanov et al.[44], where 19 different lines of Arabidopsis were used to studyintraspecific genetic differences. Their results showed that intra-specific responses to UV-B radiation were significantly different,exceeding both interspecific and intergeneric differences. Simi-larly, Ries et al. [45] studied the effect of elevated solar UV-B inArabidopsis and tobacco plants. They found that plants display awide variety of morphological and physiological responses.
These plant responses are mediated by UV-B specific pathwayswhich in turn mediate photomorphogenic responses to low levelsof UV-B and stimulate the expression of genes involved in UV-Bprotection. This meditative response was also suggested by Boccal-andro et al. [46]. Seemingly these morphological changes inducedby UV-B act as signal mechanisms which trigger the expressionof genes involved in plants’ protection against this radiation. Itseems that plants can use anatomical or mechanical differences -genetically originated- combined with other strategies to minimizethose external disturbances which exceed the usual dimensionsfound in their natural habitats.
Most studies of UV-B effects on plants show that plants try toattenuate the radiation received by using different strategies: accu-mulating UV-B absorbing secondary metabolites, neutralizing ROS
species, monomerizing pyrimidine dimmers or via photorepara-tion. However the cost of producing larger amounts of thesemetabolites and DNA repair mechanisms can divert photosynthesisat the expense of growth. In our study, this molecular responseseems to be designed to minimize the external disturbances towhich they are exposed in their natural habitat. Because these dis-ruptions exceed the natural levels to which they are exposed dur-ing the initial periods of growth, plants must use a more complexdefense mechanism by integrating different pathways to survive[47].
These changes at genetic level are more relevant, particularlythose involved in metabolic pathways, because they can changethe quality and quantity of final metabolites. This may have eco-logical consequences because some insects and other herbivoresrely on the production of these secondary metabolites to enablethem to recognize plants as the right place to oviposite so caterpil-lars can feed on them. Although UV-B effects are difficult to detectin natural conditions, they play a role in natural and managedecosystems.
The integration of molecular, chemical, physiological and bio-chemical information could provide some details about the actualresponses to this radiation and their consequences for the environ-ment. This provides clear evidence of the direct effect of UV-B radi-ation on DNA structure.
5. Abbreviations
ISSR, inter sequence simple repeat; CPD, cyclobutane pyrimi-dine dimmer; NIST, National Institute of Standards and Technol-ogy; TLC, thin layer chromatography; BAW, butanol, acetone,water; CTAB, hexadecyl trimethylammonium bromide; PCR,polymerization chain reaction; RAPD, random amplified poly-morphic DNA; PAL, phenylalanine ammonio lyase; CHS, chalconesynthase.
Acknowledgments
We would like to thank the Dirección de Investigación of theUniversidad de Magallanes (PR-F2-03RN-07-08) for financial sup-port and Mrs. Stephanie Venables for her kind revision of thismanuscript.
References
[1] M. Nei, W.H. Li, Mathematical model for studying genetic variation in termsof restriction endonucleases, Proc. Natl. Acad. Sci. USA 76 (1979) 5269–5273.
[2] P. Cuadra, J.B. Harborne, Changes in epicuticular flavonoids and photosyntheticpigments as a plant response to UV-B radiation, Z. Naturforsch. 51c (1996)671–680.
[3] K.E. Gerhardt, M.I. Wilson, B.M. Greenberg, Ultraviolet wavelength dependenceof photomorphological and photosynthetic responses in Brassica napus andArabidopsis thaliana, Photochem. Photobiol. 81 (2005) 1061–1068.
[4] K. Hectors, E. Prinzen, W. De Cohen, M.A.K. Jansen, Y. Guisez, Arabidopsisthaliana plants acclimated to low dose rates of ultraviolet-B radiation showspecific changes in morphology and gene expression in the absence of stresssymptoms, New Phytol. 175 (2007) 255–270.
[5] K.C. Smith, in: K.C. Smith (Ed.), The Science of Photobiology, UV RadiationEffects: DNA Repair and Mutagenesis, Plenum Press, New York, 1989, pp. 11–134.
[6] R. Yoshihara, T. Imaki, M. Hori, C. Watanabe, K. Yamamoto, K. Takimoto, CPDphotolyase gene from Spinacia oleracea: repair of UV-damaged DNA andexpression in plant organs, J. Radiat. Res. 46 (2005) 157–164.
[7] A. Yamamoto, N. Tanbir, T. Hirouch, M. Teramishi, J. Hidema, H. Morioka,Temperature-sensitive photoreactivation of cyclobutane thymine dimer insoybean, J. Radiat. Res. 49 (2008) 189–196.
[8] J. Cadet, C. Anselmino, T. Douki, L. Voituriez, Photochemistry of nucleic acid incells, J. Photochem. Photobiol. 15 (1992) 277–298.
[9] J. Hidema, T. Kumagai, Sensitivity of rice to ultraviolet-B radiation, Ann. Bot. 97(2006) 1377–1383.

P. Cuadra et al. / Journal of Photochemistry and Photobiology B: Biology 101 (2010) 70–75 75
[10] J.H. Sullivan, D.C. Gitz, L. Liu, C.P. Xu, W. Gao, J. Slusser, Coupling short termchanges in ambient UV-B levels with induction of UV-screening compounds,Photochem. Photobiol. 83 (2007) 863–870.
[11] N.J. Thines, L.A. Shipley, J.H. Bassman, J.R. Slusser, W. Gao, UV-B effects on thenutritional chemistry of plants and the responses of a mammalian herbivore,Oecologia 156 (2008) 125–135.
[12] K. Hashiba, T. Iwashina, S. Matsumoto, Variation in the quality and quantity offlavonoids in the leaves of coastal and inland Campanula punctata, Biochem.Syst. Ecol. 34 (2006) 854–861.
[13] K.C. Ravindran, A. Indrajith, V. Balakrishnan, K. Venkatesan, G. Kulandaivelu,Determination of defense mechanism in Phaseolus trilobus Ait. seedlingstreated under UV-B radiation, Afr. Crop Sci. J. 16 (2008) 111–118.
[14] D.C. Gitz, L. Gitz, J.W. Mc Clure, A.J. Huerta, Effects of a PAL inhibitor onphenolic accumulation and UV-B tolerance in Spirodela intermedia (Koch.), J.Exp. Bot. 55 (2004) 912–927.
[15] S.S. Singh, P. Kumar, A.K. Rai, in: A.K. Rai, T. Takabe (Eds.), AbioticStress Tolerance in Plants, Ultraviolet Radiation Stress: Molecular andPhysiological Adaptations in Trees, vol. 6, Springer, The Nederlands, 2006,pp. 91–110.
[16] B.A. Kunz, D.M. Cahill, P.G. Mohor, M.J. Osmond, E.J. Vonarx, InternationalReview of Cytology. A Survey of Cell Biology, vol. 255, Elsevier Academic Press,San Diego, USA, 2006. pp. 1–40.
[17] M.M. Qaderi, E.C. Yeung, D.M. Reid, Growth and physiological responses of aninvasive alien species, Silene noctiflora, during two developmental stages tofour levels of ultraviolet-B radiation, Ecoscience 15 (2008) 150–159.
[18] K.R. Albert, T.N. Mikkelsen, H. Ro-Poulsen, Ambient UV-B radiation decreasesphotosynthesis in high arctic Vaccinum uliginosum, Physiol. Plant. 133 (2008)199–210.
[19] L.O. Essen, T. Klar, Light-driven DNA repair by photolyases, Cell. Mol. Life Sci.63 (2006) 1266–1277.
[20] A. Sancar, G.B. Sancar, DNA repair enzymes, Ann. Rev. Biochem. 57 (1988) 29–67.
[21] S. Matallana, J.A. Meador, F. Joux, T. Douki, Effect of the GC content of DNA onthe distribution of UV-B induced bipyrimidine photoproducts, Photochem.Photobiol. Sci. 7 (2008) 794–801.
[22] S. Mouret, M. Charveron, A. Favier, J. Cadet, T. Douki, Differential repair UVBinduced cyclobutane pyrimidine dimers in cultivated human skin cells andwhole human skin, DNA Repair 7 (2008) 704–712.
[23] H.S. Kang, J. Hidema, T. Kumagai, Effects of light environment during cultureon UV-induced cyclobutyl pyrimidine dimmers and their photorepair in rice(Oryza sativa L.), Photochem. Photobiol. 68 (1998) 71–77.
[24] B.R. Zhou, D. Luo, F.D. Wei, X.E. Chen, J. Gao, Baicalein protects humanfibroblasts against ultraviolet-B induced cyclobutane pyrimidine dimmersformation, Arch. Derm. Res. 300 (2008) 331–334.
[25] P. Cuadra, J.B. Harborne, P.G. Waterman, Increases in surface flavonols andphotosynthetic pigments in Gnaphalium luteo-album in response to UV-Bradiation, Phytochemistry 45 (1997) 377–1383.
[26] P. Adamse, S. Britz, Spectral quality of two fluorescent UV sources during long-term use, Photochem. Photobiol. 56 (1992) 641–644.
[27] W.B. Sisson, M.M. Caldwell, Lamp/filter systems for simulation of solar UVirradiance under reduced atmospheric ozone, Photochem. Photobiol. 21 (1975)453–456.
[28] M.M. Caldwell, Solar ultraviolet radiation as an ecological factor for alpineplants, Ecol. Monogr. 38 (1968) 243–268.
[29] R. Mirecki, A.H. Teramura, Effects of ultraviolet-B irradiance on soybean, PlantPhysiol. 74 (1984) 475–480.
[30] J. Doyle, J. Doyle, Isolation of plant DNA from fresh tissue, Focus 12 (1990) 13–15.
[31] Y. Charters, A. Robertson, M. Wilkinson, G. Ramsay, PCR analysis of oilseedrape cultivars (Brassica napus L. ssp. oleifera) using 50-anchored simplesequence repeat (SSR) primers, Theor. Appl. Genet. 92 (1996) 442–447.
[32] R. Herrera, V. Cares, M. Wilkinson, P.D.S. Caligari, Characterization ofgenetic variation between Vitis vinifera cultivars from central Chile usingRAPD and Inter Simple Sequence Repeat markers, Euphytica 124 (2002) 139–145.
[33] F.J. Rohlf, NTSYS-pc Numerical Taxonomy and Multivariate Analysis System,Version 2.00. User Guide. Exeter Software, Applied Biostatistics Inc., Setauket,NY. USA, 2005.
[34] B. Berg, N.M. Schmidt, T.T. Hoye, P.J. Aastrup, D.K. Hendrichsen, M.C.Forchhammer, D.R. Klein, In: (FALTA AUTOR) Advances in EcologicalResearch High-Arctic Plant Herbivore Interactions Under Climate Influence,vol. 40, Academic Press, London, 2008. pp. 275–298.
[35] J. Rozema, L.O. Björn, J.F. Bornman, A. Gaberšcik, D.P. Häder, Trošt, M. Germ, M.Klisch, A. Gröniger, R.P. Sinha, M. Lebert, Y.Y. He, R. Buffoni-Hall, N.V.J. deBakker, J. van de Staaij, B.B. Meijkamp, The role of UV-B radiation in aquaticand terrestrial ecosystems – an experimental and functional analysis of theevolution of UV-absorbing compounds, J. Photochem. Photobiol. B: Biol. 66(2002) 2–12.
[36] J. Rozema, R.A. Broekman, P. Blokker, B.B. Meijkamp, N. de Bakker, J. van deStaaij, A. Van Beem, F. Ariese, S.M. Kars, UV-B absorbance and UV-B absorbingcompounds (para-coumaric acid) in pollen and sporopollen: the perspectiveto track historic UV-B levels, J. Photochem. Photobiol. B: Biol. 62 (2001) 108–117.
[37] E. Wellman, Phytochrome mediated flavone glycoside synthesis in cellsuspension cultures of Petroselinum hortense after preirradiation withultraviolet light, Planta 101 (1971) 283–286.
[38] P. Burchard, W. Bilger, G. Weissenböck, Contribution of hydroxycinnamatesand flavonoids to epidermal shielding of UV-A and UV-B radiation indeveloping rye primary leaves as assessed by ultraviolet-induced chlorophyllfluorescence measurements, Plant, Cell Environ. 23 (2000) 1373–1380.
[39] M. Tevini, U. Mark, G. Fieser, M. Saite, Effects of enhanced solar UV-B radiationon growth and function of crop plant seedlings, in: E. Riklis (Ed.), Proc. 10th Int.Congress Photobiology, Jerusalem, 1989.
[40] K. Bieza, R. Lois, An Arabidopsis mutant tolerant to lethal ultraviolet-B levelsshows constitutively elevated accumulation of flavonoids and other phenolics,Plant Physiol. 126 (2001) 1105–1115.
[41] T. Teklemariam, T. Blake, Phenylalanine ammonia-lyase-induced freezingtolerance in jack pine (Pinus banksiana) seedlings treated with low ambientlevels of ultraviolet-B radiation, Physiol. Plant 122 (2004) 244–253.
[42] T. Shimizu, S. Akada, M. Senda, R. Ishikawa, T. Harada, M. Niizeki, S. Dube,Enhanced expression and differential inducibility of soybean chalconesynthase genes by supplemental UV-B in dark-grown seedlings, Plant Mol.Biol. 39 (1999) 785–795.
[43] K. Ryan, E. Swinny, K. Markham, C. Winefield, Flavonoid gene expression andUV photoprotection in transgenic and mutant Petunia leaves, Phytochemistry59 (2002) 23–32.
[44] P.D. Usmanov, I.G. Mednik, B.I. Lipkind, Y. Giller, Genotypic characteristics ofthe response of plants to medium-wave ultraviolet radiation, Am. J. Bot. 79(1988) 863–871.
[45] G. Ries, W. Heller, H. Puchta, H. Sandermann, H.K. Seidlitz, B. Hohn, ElevatedUV-B radiation reduces genome stability in plants, Nature 406 (2000) 98–101.
[46] H.E. Boccalandro, C.A. Mazza, M.A. Mazzella, J.J. Casal, C.L. Ballaré, Ultraviolet Bradiation enhances a phytochrome-B-mediated photomorphogenic responsein Arabidopsis, Plant Physiol. 126 (2001) 280–288.
[47] J. Stratmann, Ultraviolet-B radiation co-opts defense signaling pathways,Trends Plant Sci. 8 (2003) 526–533.




















