
The emotional Stroop task and posttraumatic stress disorder: A meta-analysis

Research ArticleEffects of L-Theanine on Posttraumatic Stress Disorder InducedChanges in Rat Brain Gene Expression
Tomaacutes Eduardo Ceremuga12 Stephanie Martinson2 Jason Washington2
Robert Revels2 Jessica Wojcicki2 Damali Crawford2 Robert Edwards2
Joshua Luke Kemper2 William Luke Townsend2 Geno M Herron2
George Allen Ceremuga3 Gina Padron3 and Michael Bentley2
1 US Army Graduate Program in Anesthesia Nursing AMEDDCampS Institute of Surgical Research Fort SamHouston TX 78234 USA2United States Army Graduate Program in Anesthesia Nursing Fort Sam Houston TX 78234 USA3The Geneva Foundation WA 98402 USA
Correspondence should be addressed to Tomas Eduardo Ceremuga teceremugagmailcom
Received 2 March 2014 Revised 11 April 2014 Accepted 13 May 2014 Published 3 August 2014
Academic Editor George Panagiotis Chrousos
Copyright copy 2014 Tomas Eduardo Ceremuga et al This is an open access article distributed under the Creative CommonsAttribution License which permits unrestricted use distribution and reproduction in any medium provided the original work isproperly cited
Posttraumatic stress disorder (PTSD) is characterized by the occurrence of a traumatic event that is beyond the normal rangeof human experience The future of PTSD treatment may specifically target the molecular mechanisms of PTSD In the USapproximately 20 of adults report taking herbal products to treat medical illnesses L-theanine is the amino acid in green teaprimarily responsible for relaxation effects No studies have evaluated the potential therapeutic properties of herbal medications ongene expression in PTSD We evaluated gene expression in PTSD-induced changes in the amygdala and hippocampus of Sprague-Dawley rats The rats were assigned to PTSD-stressed and nonstressed groups that received either saline midazolam L-theanineor L-theanine + midazolam Amygdala and hippocampus tissue samples were analyzed for changes in gene expression One-wayANOVAwas used to detect significant difference between groups in the amygdala and hippocampus Of 88 genes examined 17 hada large effect size greater than 0138 Of these 3 genes in the hippocampus and 5 genes in the amygdala were considered significant(119875 lt 005) between the groups RT-PCR analysis revealed significant changes between groups in several genes implicated in avariety of disorders ranging from PTSD anxiety mood disorders and substance dependence
1 Background
Posttraumatic stress disorder (PTSD) is characterized by theoccurrence of a traumatic event that is beyond the normalrange of human experience for example rape torturegenocide and war zone stress [1] The range of symptomsof PTSD necessitates many different therapeutic approachesto treating the disorder The most successful treatments arecognitive-behavioral therapy (CBT) andmedications [1]Theselective serotonin reuptake inhibitors (SSRIs) sertraline(Zoloft) and paroxetine (Paxil) are approved by the FDA fortreatment of PTSD [1] The future of PTSD treatment mayinclude medications specifically targeted for the molecularmechanisms responsible for PTSD symptoms Zhang and
colleagues used microarray technology to compare the geneexpression profile of stressed maladapted rats (PTSD) andstressed well adapted rats in the areas of the brain associatedwith PTSD the hippocampus and the amygdala [2] In thehippocampus they found about 110 genes were upregulatedby a two-fold change or greater and about 240 genes weredownregulated by less than a half-fold change [2]They foundthat multiple two signaling pathways were upregulated andmultiple one signaling pathways were downregulated in theamygdala [2]
In the US nearly 1 in 5 adults reports taking herbalproducts to treat a medical illness in fact as recently as 1980the US Pharmacopeia consisted of 59 herbal medicines [3]Green tea is an herbal product that has been consumed since
Hindawi Publishing Corporatione Scientific World JournalVolume 2014 Article ID 419032 10 pageshttpdxdoiorg1011552014419032
2 The Scientific World Journal
ancient times to promote relaxation L-theanine (marketedby the trade name Suntheanine) is the amino acid in greentea primarily responsible for its flavor and pharmacologiceffect [4] It was discovered in 1949 and is found almostsolely in green tea leaves [4] In a small-scale human studyby Junega and colleagues Suntheanine was found to promotethe generation of alpha brain waves indicative of a relaxedstate L-theanine is a derivative of L-glutamic acid which isone of themajor excitatory neurotransmitters in the brain [5]
Although little is known about the pharmacokinetics of L-theanine in humans animal studies show that it is absorbedin the small intestine via sodium-coupled active transportcrosses the blood brain barrier via a leucine-preferringtransport system and is metabolized by the kidneys [5] Inrats an intraperitoneal injection of L-theanine is taken up bythe brain within thirty minutes without metabolic changes[4]
In addition to the study by Junega and colleagues otherresearchers have investigated the ability of L-theanine toattenuate anxiety Lu and colleagues compared the acuteeffects of L-theanine alprazolam (a prototype anxiolytic) anda placebo on the behavioral measures of anxiety in healthyhuman subjects using an anticipatory anxiety model Luand colleagues found that L-theanine may have a relaxingeffect under resting conditions but neither L-theanine noralprazolam had anxiolytic effects in the anticipatory anxietymodel [6] However in a rat study by Heese et al L-theanineconcomitantly administered with midazolam was shown tohave an interactive anxiolytic effect and a decrease in fine andbasic motor movements in a rodent model [7]
No studies have evaluated the potential therapeutic prop-erties of herbal medications in PTSD or changes in geneexpression Because PTSD is categorized as an anxiety ornervous disorder it is important to study the potentialbenefits of L-theanine in the treatment of PTSD and themolecular pathways responsible The purpose of this studywas to investigate if L-theanine modulates gene expressionin the hippocampus and amygdala regions of the brain thathave been identified as critically important areas involved inthe pathogenesis of PTSD [8ndash10] This study was part of alarger study that also investigated neurobehavioral changesin this rodent model This current investigation provides abeginning framework to pursue targeted treatments of PTSDby identifying genes involved in the neurobiology of thisdevastating condition
2 Methods
Eighty male Sprague-Dawley rats (Harlan Sprague-DawleyLaboratories) weighing 300ndash350 grams were usedThey werehoused in groups of three in polycarbonate ldquoshoebox-sizedrdquocages lined with bedding The animals went through a 7-dayadaptation period in a temperature-controlled environment(22 plusmn 1∘C 60 humidity) with a reverse light-dark cyclewhere they received 12 hours of light (600 PM to 0600 AM)and 12 hours of darkness (600 AM to 0600 PM) They wereallowed free access to food and water The animals were onlyhandled for weighing drug administration and cleaning of
cages and were naıve to all instruments After acclimationthe investigators measured baseline body weight Statisticalanalysis demonstrated equivalency between the control andPTSD-stressed groups The use of laboratory rats in thisprotocol was in accordance with the NIH Guide for the Careand Use of Laboratory Animals and received approval fromthe Institutional Animal Care and Use Committee (IACUC)at the US Army Institute of Surgical Research San AntonioTexas
The animals were randomly assigned to one of eightgroups by use of a computer number generator Rats wereassigned to the nonstressed groups or the 3-day restraintshock PTSD rodent model groups There were four groupsin the nonstressed rats control vehicle (C-V) L-theanine(C-L) midazolam (C-M) and L-theanine + midazolam (C-M + L) There were also four groups in the PTSD-inducedrats control vehicle (P-V) L-theanine (P-L) midazolam (P-M) and L-theanine + midazolam (P-M + L) Each animalreceived a subcutaneous injection of the following (1) control(saline) (2) L-theanine 10mgkg [11] (3)midazolam (Versed)15mgkg or (4) L-theanine 10mgkg + midazolam (Versed)15mgkg (to evaluate interaction effects)The vehicle (saline)group served as a control All animals received equivalentvolumes of subcutaneous injection to total 2mLkg bodyweight Additionally all experimentation occurred on atimed schedule between 8 AM and 4 PM over 4 consecutivedays to control for the circadian rhythm of the animals
21 PTSD Stress Model Stress exposure consisted of 2-hour sessions per day of immobilization and tail-shocksfor 3 consecutive days The animals were stressed in themorning (between 0800 AM and 1200 PM) They wererestrained in a plexiglass tube and 40 electric shocks (2mA3-second duration) were applied at varying intervals (140ndash180 seconds) Commercial hardware and software controlledthe timing and amplitude of the shock (Precision AnimalShocker Coulbourn Instruments Columbus Ohio USA)Rats were stressed for 3 consecutive daysThe rationale for theapproach was because it had been previously demonstratedthat repeated stress sessions for 3 days were more effectivethan a single stress session in producing sustained physi-ological and behavioral abnormalities such as exaggeratedacoustic startle response and reduced body weight [12]
Several laboratory studies have established and tested theinescapable tail-shock model of stress in rats These testshave verified the neurobiological and behavioral alterationsinduced in the rat model and are similar to those found inhuman PTSD patients [12 13] Rats were stressed for threeconsecutive days based on previous literaturewhich indicatedrepeated stress sessions were more effective than a singlestress session in producing physiological and behavioralabnormalities including exaggerated ASR and reduced bodyweight (see Table 1) [12]
22 Gene Analysis After the completion of neurobehavioraltests the investigators anesthetized themale Sprague-Dawleyrats using isoflurane in a bell jar and the calvarium wasremoved exposing the whole brain The whole brain was
The Scientific World Journal 3
Table 1 Comparison of symptoms of PTSD in humans to dysfunction related to stress in rats (restraintshock) [12 13]
PTSD in humans Rat inescapable tail-shock model of stress (3 days)Weight loss Suppressed feeding and body weight lossDifficulty falling or staying asleep nightmares Altered sleep patterns
Psychomotor numbness Persistent behavioral abnormalities that is suppressed open-field activity longer hangingwire latencies
Poor concentration memory deficits Deficits in escapeavoidance learning and learning of an appetitive taskHyperarousalstartle response Exaggerated startle response
Brain cutting block
Razor blades
Sections
123456789
(a)
1 2 3 4
5 6 7 8
(b)
Figure 1 (a) Orientation of brain within brain cutting block (b) coronal brain sections from dissected regions (a) Illustration of rat brainin cutting block depicting specific areas where coronal slices were performed (b) numbers correspond to dissected sections from (b) FCfrontal cortexNA nucleus accumbensOT olfactory tubercle S septumCP caudate putamen RC remaining cortex GP globus pallidus aHanterior hypothalamus pH posterior hypothalamus A amygdala T thalamus SN substantia nigra VT ventral tegmentumH hippocampus[14]
removed intactly and placed immediately on a brain blockon ice [14] Coronal slices (200120583m thickness) in the vicinityof interaural and bregma coordinates 696mm and minus204respectively were dissected using a brain block (see redshaded region in Figure 1(a)) Circular punches (80ndash100 120583mdiameter)were retrieved from the left and right amygdala (seelower left red circle in area 6 of Figure 1(b)) Following thedissection of the amygdala the investigator grossly dissectedthe bilateral hippocampi (see upper medial red outlined areain area 6 of Figure 1(b)) Tissue was snap-frozen in liquidnitrogen placed in prelabeled eppendorf tubes and packedin dry ice for shipping to the QIAGEN Service Core forGenomics and Gene Expression in Frederick MD
RNA was isolated using the QIAGEN RNEasy MiniKit (Cat no 74104) following the manufacturerrsquos protocolfor tissue RNA quality was determined using the AgilentBioanalyzer (Agilent) with RNA 6000 Nano Kits (AgilentCat no 5067-1511) Total RNA yield 260280 and 260230ratios were measured using a NanoDrop spectrophotometer(Thermo) A reverse transcription reaction using 500 ng oftotal RNAwas completed using theQIAGENRT2 First StrandKit (QIAGEN Cat no 330401) cDNA samples were assayedusing a modified QIAGEN RT2 PCR array (Cat no PARN-60) according to manufacturerrsquos instructions [15 16] Thisarray containing 84 assays related to rat neurotransmitter
receptors and regulators was modified to include four addi-tional assays of genes from QIAGENThese genes are relatedto neurotransmission pathways that have been identified inprevious PTSD studies p11 (S100a10) (Cat no PPR06766)[17] 5HT
2A receptor (Htr2a) (Cat no PPR06850) [18] alpha1adrenergic receptor (Adra1a) (Cat no PPR43329) [19ndash21]andEgr1 (Zif268) (Cat no PPR44272) an effector immediateearly gene [15 16]
23 Rat Neurotransmitter Receptors and Regulators PCRArray The rat neurotransmitter receptors and regulatorsRT2 profiler PCR array profiles the expression of 84 genesinvolved in modulating the biological processes of neuro-transmitter biosynthesis uptake transport and signalingthrough neurotransmitter receptors This array containsreceptors for specific neurotransmitters such as acetyl-choline benzodiazepine dopamine gamma-aminobutyricacid (GABA) glutamate serotonin somatostatin and neu-ropeptides Genes involved in the regulation of neurotrans-mitter levels were included as well Analysis of the expressionof a focused panel of genes related to neuronal systems withthis array can be performed using real-time PCR
24 Statistical Analyses For this cross-sectional random-ized prospective study a one-way ANOVA was performed
4 The Scientific World Journal
to compare the 8 groups of which each unit was randomlyallocated All assumptions were examined (eg homogeneityof variance univariate normality etc) and in the event thatthe assumptions are not tenable remedial (eg transfor-mation) or alternative (eg nonparametric tests such asKruskal-Wallis) strategies were consideredMultiple compar-ison procedures (MCP) were performed in the event thatsignificance (120572 = 05) was obtained with the Tukey posthoc test A variance explained statistic eta-squared (1205782) wasused as the reported effect size and though ascertainingwhat constitutes a smallmediumlarge effect is primarily afunction of the contextdiscipline for this report 01059138was smallmediumlarge [22] All data were statisticallyanalyzed based on gene cycle thresholds (CTs) normalized tofive housekeeping genes per QIAGEN procedures [15 16]
3 Results
This study was designed to investigate the effects of L-theanine on PTSD-induced changes in rat brain gene expres-sion Eighty-eight genes were investigated in this study Dataanalysis showed a number of significant differences in geneexpression in both the hippocampus and amygdala Volcanoplots were developed to compare the gene expression of the40 nonstressed to the 40 PTSD-stressed rats for both thehippocampus and amygdala as demonstrated in Figure 2Thevolcano plots graphically display the relationship of ndashlog 10 119875value and the Log 2 fold change for the combined treatmentand control groups for all of the genes For the 8-group one-way ANOVA a large effect size (1205782 gt 138) was chosen toserve as a springboard for discussion and interpretation ofrelative between-group differences on gene expression
31 Hippocampus In the hippocampus the genes with aneffect size gt 0138 are displayed in Figure 3 In accordancewith Figure 3 Table 2 summarizes between-group changesin gene expression in the hippocampus The Egr1 gene wasfound to be significant and between-group differences wereobtained 119875 = 011 1205782 = 218 There was one significantpost hoc test with the P-M group having a significantly highermean (119872 = 558) than the P-V group (119872 = 457) Asignificant difference was found between groups for theMaoagene 119875 = 024 1205782 = 194 The post hoc test found onesignificant pairwise comparison (1) the C-M + L group hadthe highest mean (119872 = 332) and the P-V group had thelowest (119872 = 312) The Anxa9 gene was found to havesignificance between groups 119875 = 031 1205782 = 187 Multiplecomparison tests resulted in one significant pairwise resultthe P-M group had a significantly higher mean (119872 = 1073)than the P-V group (119872 = 1006)
The following genes in the hippocampus did not showsignificant differences between groups however due to thelarge effect size (variance explained) these genes are presentedin Table 2The S100a10 gene did not have significant between-group differences 119875 = 06 1205782 = 166 The P-M group had thehighest mean (119872 = 42) and the P-V group had the lowest(119872 = 382) Significant between-group differences were notobtained for the Gabrb2 gene 119875 = 123 1205782 = 142 The P-M
group had the highest mean (119872 = 332) and the P-V grouphad the lowest (119872 = 275) Finally the Chrm2 gene did notshow significance between groups A statistically significantdifference was not obtained 119875 = 138 1205782 = 138 Thoughnot significant the C-M + L group had the highest mean(119872 = 1056) and the P-V group had the lowest (119872 = 996)
32 Amygdala Genes in the amygdala with an effect sizegreater than 0138 are displayed in Figure 4 In accordancewith Figure 4 Table 3 summarizes between-groupdifferencesin gene expression within the amygdala The Gabra4 genewas 119875 = 001 1205782 = 271 Significant post hoc pairwisecomparisons were such that the C-L group (119872 = 352) had ahigher mean than each of the following (1) P-L (119872 = 304)(2) P-M (119872 = 311) and (3) C-M group (M = 309) Asignificant difference for the Slc5a7 gene was found119875 = 0241205782
= 195 The C-V group had the highest mean (119872 = 775)and the P-L group had the lowest (119872 = 667) howeveras per the post hoc test this pairwise comparison was notsignificant Significant between-group differences were alsofound for the Drd2 gene 119875 = 02 1205782 = 189 There was asignificant difference between P-V (119872 = 676) and the P-Mgroup (119872 = 448) The Drd1a gene was also significant 119875 =038 1205782 = 181 A significant difference was found betweenthe C-M + L (119872 = 641) and the P-M group (M = 483)Lastly the Glra2 showed significance 119875 = 04 1205782 = 179There was one significant post hoc test between the C-M + Lgroup with the higher mean (119872 = 562) and the P-M groupwith the lower mean (119872 = 489)
Again the following genes were not found to be signifi-cant however based on the large effect size (variability) usedin the study these genes are included in Table 3 No differencewas found for the Chrna6 gene 119875 = 075 1205782 = 159 The C-M group had the highest mean (119872 = 1651) and the C-M +L group had the lowest (119872 = 155) The Htr3a gene showedno significant difference between the groups 119875 = 081 1205782 =156 The C-L group had the highest mean (119872 = 674) andthe P-M + L group had the lowest (119872 = 635) The Chrnb4also showed no significant difference 119875 = 087 1205782 = 154The C-V group had the highest mean (119872 = 1159) and theC-M + L group had the lowest (119872 = 1043) No significantdifference was found for the Gabrd gene between the groups119875 = 089 1205782 = 153TheC-M+L group had the highestmean(119872 = 523) and the P-M group had the lowest (119872 = 456)The Prima1 gene did not have a significant difference betweenthe groups 119875 = 111 1205782 = 145 The P-V group had thehighest mean (119872 = 694) and the P-M group had the lowest(119872 = 625) No difference between the groups was found forthe Chrna3 gene 119875 = 127 1205782 = 141 The P-L group had thehighest mean (119872 = 1056) and the C-M + L group had thelowest (119872 = 967)
4 Discussion
L-theanine has previously been demonstrated to produceanxiolytic effects alone or in combination with concomitantadministered benzodiazepines [7] No studies have examined
The Scientific World Journal 5
2000
1500
1000
Hippocampal
Log 2 (fold difference)
minuslo
g Pva
lue
minus00750 minus00500 minus00250 00000 00250 00500
0500
0000
(a)
3000
2000
1000
Amygdalar
Log 2 (fold difference)
minuslo
g Pva
lue
0000minus0150 minus0100 minus0050 0000 0050 0100 0150
(b)
Figure 2 Differential expression of (a) hippocampal and (b) amygdalar mRNA between groups Volcano plot between Log 2 fold change onthe 119909-axis (for neurotransmitter receptors and regulators in L-theaninemidazolam treated groups 40 PTSD-stressed versus 40 nonstressed(control) groups) and minuslog of 119875 value on the 119910-axis Eighty male Sprague-Dawley rats were injected subcutaneously 30 minutes prior toevaluation of their performance on neurobehavioral tests The rats were then euthanized and cDNA prepared from the hippocampus (a) andamygdala (b) and were subjected to RT2 profiler PCR array for rat neurotransmitter receptor and regulator analysis as described inMaterialsand Methods PCR array profiles were performed for the expression of 88 genes potentially involved in PTSD andor rat neurotransmitterreceptors and neurotransmitter regulation
Hippocampal genes
Egr1 Maoa Anxa9 S100a10 Gabrb2 Chrm2
025
020
015
010
005
000
lowast
lowastlowast
Effec
t siz
e (1205782)
Large effect size (gt0138)
Figure 3 Effect size of gene changes in the hippocampus Genesdemonstrating a large effect size (1205782 gt 138) in the hippocampus asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
the administration of L-theaninersquos effects on PTSD-inducedchanges in brain gene expression We explored the modu-lation of gene expression in PTSD-induced changes in thehippocampus and amygdala and the effects of L-theanine andmidazolam on these changes
41 Hippocampus We found a downregulation of the Egr1gene in the PTSD-vehicle group compared to the PTSD-midazolam group Egr1 is an early growth response nuclear
Amygdalar genes
030
025
020
015
010
005
000
Effec
t siz
e (1205782)
Gab
ra4
Slc5
a7
Drd2
Drd1
a
Glra
2
Chrn
a6
Htr3
a
Chrn
b4
Gab
rd
Prim
a1
Chrn
a3
lowast
lowastlowast lowast lowast
Large effect sizes (gt0138)
Figure 4 Effect size of gene changes in the amygdala Genesdemonstrating a large effect size (1205782 gt 138) in the amygdala asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
transcription factor that plays a role in regulating the expres-sion of genes required for learning and memory [23 24] Inparticular previous studies have demonstrated a role for Egr1in hippocampal long-term potentiation a process critical tolearning and memory [25] Expression of the Maoa gene wasdecreased in the PTSD-vehicle group compared to the controlgroup that received L-theanine and midazolam
Decreased Maoa enzymatic activity leads to increasedconcentrations of its substrate biogenic amines including
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

2 The Scientific World Journal
ancient times to promote relaxation L-theanine (marketedby the trade name Suntheanine) is the amino acid in greentea primarily responsible for its flavor and pharmacologiceffect [4] It was discovered in 1949 and is found almostsolely in green tea leaves [4] In a small-scale human studyby Junega and colleagues Suntheanine was found to promotethe generation of alpha brain waves indicative of a relaxedstate L-theanine is a derivative of L-glutamic acid which isone of themajor excitatory neurotransmitters in the brain [5]
Although little is known about the pharmacokinetics of L-theanine in humans animal studies show that it is absorbedin the small intestine via sodium-coupled active transportcrosses the blood brain barrier via a leucine-preferringtransport system and is metabolized by the kidneys [5] Inrats an intraperitoneal injection of L-theanine is taken up bythe brain within thirty minutes without metabolic changes[4]
In addition to the study by Junega and colleagues otherresearchers have investigated the ability of L-theanine toattenuate anxiety Lu and colleagues compared the acuteeffects of L-theanine alprazolam (a prototype anxiolytic) anda placebo on the behavioral measures of anxiety in healthyhuman subjects using an anticipatory anxiety model Luand colleagues found that L-theanine may have a relaxingeffect under resting conditions but neither L-theanine noralprazolam had anxiolytic effects in the anticipatory anxietymodel [6] However in a rat study by Heese et al L-theanineconcomitantly administered with midazolam was shown tohave an interactive anxiolytic effect and a decrease in fine andbasic motor movements in a rodent model [7]
No studies have evaluated the potential therapeutic prop-erties of herbal medications in PTSD or changes in geneexpression Because PTSD is categorized as an anxiety ornervous disorder it is important to study the potentialbenefits of L-theanine in the treatment of PTSD and themolecular pathways responsible The purpose of this studywas to investigate if L-theanine modulates gene expressionin the hippocampus and amygdala regions of the brain thathave been identified as critically important areas involved inthe pathogenesis of PTSD [8ndash10] This study was part of alarger study that also investigated neurobehavioral changesin this rodent model This current investigation provides abeginning framework to pursue targeted treatments of PTSDby identifying genes involved in the neurobiology of thisdevastating condition
2 Methods
Eighty male Sprague-Dawley rats (Harlan Sprague-DawleyLaboratories) weighing 300ndash350 grams were usedThey werehoused in groups of three in polycarbonate ldquoshoebox-sizedrdquocages lined with bedding The animals went through a 7-dayadaptation period in a temperature-controlled environment(22 plusmn 1∘C 60 humidity) with a reverse light-dark cyclewhere they received 12 hours of light (600 PM to 0600 AM)and 12 hours of darkness (600 AM to 0600 PM) They wereallowed free access to food and water The animals were onlyhandled for weighing drug administration and cleaning of
cages and were naıve to all instruments After acclimationthe investigators measured baseline body weight Statisticalanalysis demonstrated equivalency between the control andPTSD-stressed groups The use of laboratory rats in thisprotocol was in accordance with the NIH Guide for the Careand Use of Laboratory Animals and received approval fromthe Institutional Animal Care and Use Committee (IACUC)at the US Army Institute of Surgical Research San AntonioTexas
The animals were randomly assigned to one of eightgroups by use of a computer number generator Rats wereassigned to the nonstressed groups or the 3-day restraintshock PTSD rodent model groups There were four groupsin the nonstressed rats control vehicle (C-V) L-theanine(C-L) midazolam (C-M) and L-theanine + midazolam (C-M + L) There were also four groups in the PTSD-inducedrats control vehicle (P-V) L-theanine (P-L) midazolam (P-M) and L-theanine + midazolam (P-M + L) Each animalreceived a subcutaneous injection of the following (1) control(saline) (2) L-theanine 10mgkg [11] (3)midazolam (Versed)15mgkg or (4) L-theanine 10mgkg + midazolam (Versed)15mgkg (to evaluate interaction effects)The vehicle (saline)group served as a control All animals received equivalentvolumes of subcutaneous injection to total 2mLkg bodyweight Additionally all experimentation occurred on atimed schedule between 8 AM and 4 PM over 4 consecutivedays to control for the circadian rhythm of the animals
21 PTSD Stress Model Stress exposure consisted of 2-hour sessions per day of immobilization and tail-shocksfor 3 consecutive days The animals were stressed in themorning (between 0800 AM and 1200 PM) They wererestrained in a plexiglass tube and 40 electric shocks (2mA3-second duration) were applied at varying intervals (140ndash180 seconds) Commercial hardware and software controlledthe timing and amplitude of the shock (Precision AnimalShocker Coulbourn Instruments Columbus Ohio USA)Rats were stressed for 3 consecutive daysThe rationale for theapproach was because it had been previously demonstratedthat repeated stress sessions for 3 days were more effectivethan a single stress session in producing sustained physi-ological and behavioral abnormalities such as exaggeratedacoustic startle response and reduced body weight [12]
Several laboratory studies have established and tested theinescapable tail-shock model of stress in rats These testshave verified the neurobiological and behavioral alterationsinduced in the rat model and are similar to those found inhuman PTSD patients [12 13] Rats were stressed for threeconsecutive days based on previous literaturewhich indicatedrepeated stress sessions were more effective than a singlestress session in producing physiological and behavioralabnormalities including exaggerated ASR and reduced bodyweight (see Table 1) [12]
22 Gene Analysis After the completion of neurobehavioraltests the investigators anesthetized themale Sprague-Dawleyrats using isoflurane in a bell jar and the calvarium wasremoved exposing the whole brain The whole brain was
The Scientific World Journal 3
Table 1 Comparison of symptoms of PTSD in humans to dysfunction related to stress in rats (restraintshock) [12 13]
PTSD in humans Rat inescapable tail-shock model of stress (3 days)Weight loss Suppressed feeding and body weight lossDifficulty falling or staying asleep nightmares Altered sleep patterns
Psychomotor numbness Persistent behavioral abnormalities that is suppressed open-field activity longer hangingwire latencies
Poor concentration memory deficits Deficits in escapeavoidance learning and learning of an appetitive taskHyperarousalstartle response Exaggerated startle response
Brain cutting block
Razor blades
Sections
123456789
(a)
1 2 3 4
5 6 7 8
(b)
Figure 1 (a) Orientation of brain within brain cutting block (b) coronal brain sections from dissected regions (a) Illustration of rat brainin cutting block depicting specific areas where coronal slices were performed (b) numbers correspond to dissected sections from (b) FCfrontal cortexNA nucleus accumbensOT olfactory tubercle S septumCP caudate putamen RC remaining cortex GP globus pallidus aHanterior hypothalamus pH posterior hypothalamus A amygdala T thalamus SN substantia nigra VT ventral tegmentumH hippocampus[14]
removed intactly and placed immediately on a brain blockon ice [14] Coronal slices (200120583m thickness) in the vicinityof interaural and bregma coordinates 696mm and minus204respectively were dissected using a brain block (see redshaded region in Figure 1(a)) Circular punches (80ndash100 120583mdiameter)were retrieved from the left and right amygdala (seelower left red circle in area 6 of Figure 1(b)) Following thedissection of the amygdala the investigator grossly dissectedthe bilateral hippocampi (see upper medial red outlined areain area 6 of Figure 1(b)) Tissue was snap-frozen in liquidnitrogen placed in prelabeled eppendorf tubes and packedin dry ice for shipping to the QIAGEN Service Core forGenomics and Gene Expression in Frederick MD
RNA was isolated using the QIAGEN RNEasy MiniKit (Cat no 74104) following the manufacturerrsquos protocolfor tissue RNA quality was determined using the AgilentBioanalyzer (Agilent) with RNA 6000 Nano Kits (AgilentCat no 5067-1511) Total RNA yield 260280 and 260230ratios were measured using a NanoDrop spectrophotometer(Thermo) A reverse transcription reaction using 500 ng oftotal RNAwas completed using theQIAGENRT2 First StrandKit (QIAGEN Cat no 330401) cDNA samples were assayedusing a modified QIAGEN RT2 PCR array (Cat no PARN-60) according to manufacturerrsquos instructions [15 16] Thisarray containing 84 assays related to rat neurotransmitter
receptors and regulators was modified to include four addi-tional assays of genes from QIAGENThese genes are relatedto neurotransmission pathways that have been identified inprevious PTSD studies p11 (S100a10) (Cat no PPR06766)[17] 5HT
2A receptor (Htr2a) (Cat no PPR06850) [18] alpha1adrenergic receptor (Adra1a) (Cat no PPR43329) [19ndash21]andEgr1 (Zif268) (Cat no PPR44272) an effector immediateearly gene [15 16]
23 Rat Neurotransmitter Receptors and Regulators PCRArray The rat neurotransmitter receptors and regulatorsRT2 profiler PCR array profiles the expression of 84 genesinvolved in modulating the biological processes of neuro-transmitter biosynthesis uptake transport and signalingthrough neurotransmitter receptors This array containsreceptors for specific neurotransmitters such as acetyl-choline benzodiazepine dopamine gamma-aminobutyricacid (GABA) glutamate serotonin somatostatin and neu-ropeptides Genes involved in the regulation of neurotrans-mitter levels were included as well Analysis of the expressionof a focused panel of genes related to neuronal systems withthis array can be performed using real-time PCR
24 Statistical Analyses For this cross-sectional random-ized prospective study a one-way ANOVA was performed
4 The Scientific World Journal
to compare the 8 groups of which each unit was randomlyallocated All assumptions were examined (eg homogeneityof variance univariate normality etc) and in the event thatthe assumptions are not tenable remedial (eg transfor-mation) or alternative (eg nonparametric tests such asKruskal-Wallis) strategies were consideredMultiple compar-ison procedures (MCP) were performed in the event thatsignificance (120572 = 05) was obtained with the Tukey posthoc test A variance explained statistic eta-squared (1205782) wasused as the reported effect size and though ascertainingwhat constitutes a smallmediumlarge effect is primarily afunction of the contextdiscipline for this report 01059138was smallmediumlarge [22] All data were statisticallyanalyzed based on gene cycle thresholds (CTs) normalized tofive housekeeping genes per QIAGEN procedures [15 16]
3 Results
This study was designed to investigate the effects of L-theanine on PTSD-induced changes in rat brain gene expres-sion Eighty-eight genes were investigated in this study Dataanalysis showed a number of significant differences in geneexpression in both the hippocampus and amygdala Volcanoplots were developed to compare the gene expression of the40 nonstressed to the 40 PTSD-stressed rats for both thehippocampus and amygdala as demonstrated in Figure 2Thevolcano plots graphically display the relationship of ndashlog 10 119875value and the Log 2 fold change for the combined treatmentand control groups for all of the genes For the 8-group one-way ANOVA a large effect size (1205782 gt 138) was chosen toserve as a springboard for discussion and interpretation ofrelative between-group differences on gene expression
31 Hippocampus In the hippocampus the genes with aneffect size gt 0138 are displayed in Figure 3 In accordancewith Figure 3 Table 2 summarizes between-group changesin gene expression in the hippocampus The Egr1 gene wasfound to be significant and between-group differences wereobtained 119875 = 011 1205782 = 218 There was one significantpost hoc test with the P-M group having a significantly highermean (119872 = 558) than the P-V group (119872 = 457) Asignificant difference was found between groups for theMaoagene 119875 = 024 1205782 = 194 The post hoc test found onesignificant pairwise comparison (1) the C-M + L group hadthe highest mean (119872 = 332) and the P-V group had thelowest (119872 = 312) The Anxa9 gene was found to havesignificance between groups 119875 = 031 1205782 = 187 Multiplecomparison tests resulted in one significant pairwise resultthe P-M group had a significantly higher mean (119872 = 1073)than the P-V group (119872 = 1006)
The following genes in the hippocampus did not showsignificant differences between groups however due to thelarge effect size (variance explained) these genes are presentedin Table 2The S100a10 gene did not have significant between-group differences 119875 = 06 1205782 = 166 The P-M group had thehighest mean (119872 = 42) and the P-V group had the lowest(119872 = 382) Significant between-group differences were notobtained for the Gabrb2 gene 119875 = 123 1205782 = 142 The P-M
group had the highest mean (119872 = 332) and the P-V grouphad the lowest (119872 = 275) Finally the Chrm2 gene did notshow significance between groups A statistically significantdifference was not obtained 119875 = 138 1205782 = 138 Thoughnot significant the C-M + L group had the highest mean(119872 = 1056) and the P-V group had the lowest (119872 = 996)
32 Amygdala Genes in the amygdala with an effect sizegreater than 0138 are displayed in Figure 4 In accordancewith Figure 4 Table 3 summarizes between-groupdifferencesin gene expression within the amygdala The Gabra4 genewas 119875 = 001 1205782 = 271 Significant post hoc pairwisecomparisons were such that the C-L group (119872 = 352) had ahigher mean than each of the following (1) P-L (119872 = 304)(2) P-M (119872 = 311) and (3) C-M group (M = 309) Asignificant difference for the Slc5a7 gene was found119875 = 0241205782
= 195 The C-V group had the highest mean (119872 = 775)and the P-L group had the lowest (119872 = 667) howeveras per the post hoc test this pairwise comparison was notsignificant Significant between-group differences were alsofound for the Drd2 gene 119875 = 02 1205782 = 189 There was asignificant difference between P-V (119872 = 676) and the P-Mgroup (119872 = 448) The Drd1a gene was also significant 119875 =038 1205782 = 181 A significant difference was found betweenthe C-M + L (119872 = 641) and the P-M group (M = 483)Lastly the Glra2 showed significance 119875 = 04 1205782 = 179There was one significant post hoc test between the C-M + Lgroup with the higher mean (119872 = 562) and the P-M groupwith the lower mean (119872 = 489)
Again the following genes were not found to be signifi-cant however based on the large effect size (variability) usedin the study these genes are included in Table 3 No differencewas found for the Chrna6 gene 119875 = 075 1205782 = 159 The C-M group had the highest mean (119872 = 1651) and the C-M +L group had the lowest (119872 = 155) The Htr3a gene showedno significant difference between the groups 119875 = 081 1205782 =156 The C-L group had the highest mean (119872 = 674) andthe P-M + L group had the lowest (119872 = 635) The Chrnb4also showed no significant difference 119875 = 087 1205782 = 154The C-V group had the highest mean (119872 = 1159) and theC-M + L group had the lowest (119872 = 1043) No significantdifference was found for the Gabrd gene between the groups119875 = 089 1205782 = 153TheC-M+L group had the highestmean(119872 = 523) and the P-M group had the lowest (119872 = 456)The Prima1 gene did not have a significant difference betweenthe groups 119875 = 111 1205782 = 145 The P-V group had thehighest mean (119872 = 694) and the P-M group had the lowest(119872 = 625) No difference between the groups was found forthe Chrna3 gene 119875 = 127 1205782 = 141 The P-L group had thehighest mean (119872 = 1056) and the C-M + L group had thelowest (119872 = 967)
4 Discussion
L-theanine has previously been demonstrated to produceanxiolytic effects alone or in combination with concomitantadministered benzodiazepines [7] No studies have examined
The Scientific World Journal 5
2000
1500
1000
Hippocampal
Log 2 (fold difference)
minuslo
g Pva
lue
minus00750 minus00500 minus00250 00000 00250 00500
0500
0000
(a)
3000
2000
1000
Amygdalar
Log 2 (fold difference)
minuslo
g Pva
lue
0000minus0150 minus0100 minus0050 0000 0050 0100 0150
(b)
Figure 2 Differential expression of (a) hippocampal and (b) amygdalar mRNA between groups Volcano plot between Log 2 fold change onthe 119909-axis (for neurotransmitter receptors and regulators in L-theaninemidazolam treated groups 40 PTSD-stressed versus 40 nonstressed(control) groups) and minuslog of 119875 value on the 119910-axis Eighty male Sprague-Dawley rats were injected subcutaneously 30 minutes prior toevaluation of their performance on neurobehavioral tests The rats were then euthanized and cDNA prepared from the hippocampus (a) andamygdala (b) and were subjected to RT2 profiler PCR array for rat neurotransmitter receptor and regulator analysis as described inMaterialsand Methods PCR array profiles were performed for the expression of 88 genes potentially involved in PTSD andor rat neurotransmitterreceptors and neurotransmitter regulation
Hippocampal genes
Egr1 Maoa Anxa9 S100a10 Gabrb2 Chrm2
025
020
015
010
005
000
lowast
lowastlowast
Effec
t siz
e (1205782)
Large effect size (gt0138)
Figure 3 Effect size of gene changes in the hippocampus Genesdemonstrating a large effect size (1205782 gt 138) in the hippocampus asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
the administration of L-theaninersquos effects on PTSD-inducedchanges in brain gene expression We explored the modu-lation of gene expression in PTSD-induced changes in thehippocampus and amygdala and the effects of L-theanine andmidazolam on these changes
41 Hippocampus We found a downregulation of the Egr1gene in the PTSD-vehicle group compared to the PTSD-midazolam group Egr1 is an early growth response nuclear
Amygdalar genes
030
025
020
015
010
005
000
Effec
t siz
e (1205782)
Gab
ra4
Slc5
a7
Drd2
Drd1
a
Glra
2
Chrn
a6
Htr3
a
Chrn
b4
Gab
rd
Prim
a1
Chrn
a3
lowast
lowastlowast lowast lowast
Large effect sizes (gt0138)
Figure 4 Effect size of gene changes in the amygdala Genesdemonstrating a large effect size (1205782 gt 138) in the amygdala asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
transcription factor that plays a role in regulating the expres-sion of genes required for learning and memory [23 24] Inparticular previous studies have demonstrated a role for Egr1in hippocampal long-term potentiation a process critical tolearning and memory [25] Expression of the Maoa gene wasdecreased in the PTSD-vehicle group compared to the controlgroup that received L-theanine and midazolam
Decreased Maoa enzymatic activity leads to increasedconcentrations of its substrate biogenic amines including
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

The Scientific World Journal 3
Table 1 Comparison of symptoms of PTSD in humans to dysfunction related to stress in rats (restraintshock) [12 13]
PTSD in humans Rat inescapable tail-shock model of stress (3 days)Weight loss Suppressed feeding and body weight lossDifficulty falling or staying asleep nightmares Altered sleep patterns
Psychomotor numbness Persistent behavioral abnormalities that is suppressed open-field activity longer hangingwire latencies
Poor concentration memory deficits Deficits in escapeavoidance learning and learning of an appetitive taskHyperarousalstartle response Exaggerated startle response
Brain cutting block
Razor blades
Sections
123456789
(a)
1 2 3 4
5 6 7 8
(b)
Figure 1 (a) Orientation of brain within brain cutting block (b) coronal brain sections from dissected regions (a) Illustration of rat brainin cutting block depicting specific areas where coronal slices were performed (b) numbers correspond to dissected sections from (b) FCfrontal cortexNA nucleus accumbensOT olfactory tubercle S septumCP caudate putamen RC remaining cortex GP globus pallidus aHanterior hypothalamus pH posterior hypothalamus A amygdala T thalamus SN substantia nigra VT ventral tegmentumH hippocampus[14]
removed intactly and placed immediately on a brain blockon ice [14] Coronal slices (200120583m thickness) in the vicinityof interaural and bregma coordinates 696mm and minus204respectively were dissected using a brain block (see redshaded region in Figure 1(a)) Circular punches (80ndash100 120583mdiameter)were retrieved from the left and right amygdala (seelower left red circle in area 6 of Figure 1(b)) Following thedissection of the amygdala the investigator grossly dissectedthe bilateral hippocampi (see upper medial red outlined areain area 6 of Figure 1(b)) Tissue was snap-frozen in liquidnitrogen placed in prelabeled eppendorf tubes and packedin dry ice for shipping to the QIAGEN Service Core forGenomics and Gene Expression in Frederick MD
RNA was isolated using the QIAGEN RNEasy MiniKit (Cat no 74104) following the manufacturerrsquos protocolfor tissue RNA quality was determined using the AgilentBioanalyzer (Agilent) with RNA 6000 Nano Kits (AgilentCat no 5067-1511) Total RNA yield 260280 and 260230ratios were measured using a NanoDrop spectrophotometer(Thermo) A reverse transcription reaction using 500 ng oftotal RNAwas completed using theQIAGENRT2 First StrandKit (QIAGEN Cat no 330401) cDNA samples were assayedusing a modified QIAGEN RT2 PCR array (Cat no PARN-60) according to manufacturerrsquos instructions [15 16] Thisarray containing 84 assays related to rat neurotransmitter
receptors and regulators was modified to include four addi-tional assays of genes from QIAGENThese genes are relatedto neurotransmission pathways that have been identified inprevious PTSD studies p11 (S100a10) (Cat no PPR06766)[17] 5HT
2A receptor (Htr2a) (Cat no PPR06850) [18] alpha1adrenergic receptor (Adra1a) (Cat no PPR43329) [19ndash21]andEgr1 (Zif268) (Cat no PPR44272) an effector immediateearly gene [15 16]
23 Rat Neurotransmitter Receptors and Regulators PCRArray The rat neurotransmitter receptors and regulatorsRT2 profiler PCR array profiles the expression of 84 genesinvolved in modulating the biological processes of neuro-transmitter biosynthesis uptake transport and signalingthrough neurotransmitter receptors This array containsreceptors for specific neurotransmitters such as acetyl-choline benzodiazepine dopamine gamma-aminobutyricacid (GABA) glutamate serotonin somatostatin and neu-ropeptides Genes involved in the regulation of neurotrans-mitter levels were included as well Analysis of the expressionof a focused panel of genes related to neuronal systems withthis array can be performed using real-time PCR
24 Statistical Analyses For this cross-sectional random-ized prospective study a one-way ANOVA was performed
4 The Scientific World Journal
to compare the 8 groups of which each unit was randomlyallocated All assumptions were examined (eg homogeneityof variance univariate normality etc) and in the event thatthe assumptions are not tenable remedial (eg transfor-mation) or alternative (eg nonparametric tests such asKruskal-Wallis) strategies were consideredMultiple compar-ison procedures (MCP) were performed in the event thatsignificance (120572 = 05) was obtained with the Tukey posthoc test A variance explained statistic eta-squared (1205782) wasused as the reported effect size and though ascertainingwhat constitutes a smallmediumlarge effect is primarily afunction of the contextdiscipline for this report 01059138was smallmediumlarge [22] All data were statisticallyanalyzed based on gene cycle thresholds (CTs) normalized tofive housekeeping genes per QIAGEN procedures [15 16]
3 Results
This study was designed to investigate the effects of L-theanine on PTSD-induced changes in rat brain gene expres-sion Eighty-eight genes were investigated in this study Dataanalysis showed a number of significant differences in geneexpression in both the hippocampus and amygdala Volcanoplots were developed to compare the gene expression of the40 nonstressed to the 40 PTSD-stressed rats for both thehippocampus and amygdala as demonstrated in Figure 2Thevolcano plots graphically display the relationship of ndashlog 10 119875value and the Log 2 fold change for the combined treatmentand control groups for all of the genes For the 8-group one-way ANOVA a large effect size (1205782 gt 138) was chosen toserve as a springboard for discussion and interpretation ofrelative between-group differences on gene expression
31 Hippocampus In the hippocampus the genes with aneffect size gt 0138 are displayed in Figure 3 In accordancewith Figure 3 Table 2 summarizes between-group changesin gene expression in the hippocampus The Egr1 gene wasfound to be significant and between-group differences wereobtained 119875 = 011 1205782 = 218 There was one significantpost hoc test with the P-M group having a significantly highermean (119872 = 558) than the P-V group (119872 = 457) Asignificant difference was found between groups for theMaoagene 119875 = 024 1205782 = 194 The post hoc test found onesignificant pairwise comparison (1) the C-M + L group hadthe highest mean (119872 = 332) and the P-V group had thelowest (119872 = 312) The Anxa9 gene was found to havesignificance between groups 119875 = 031 1205782 = 187 Multiplecomparison tests resulted in one significant pairwise resultthe P-M group had a significantly higher mean (119872 = 1073)than the P-V group (119872 = 1006)
The following genes in the hippocampus did not showsignificant differences between groups however due to thelarge effect size (variance explained) these genes are presentedin Table 2The S100a10 gene did not have significant between-group differences 119875 = 06 1205782 = 166 The P-M group had thehighest mean (119872 = 42) and the P-V group had the lowest(119872 = 382) Significant between-group differences were notobtained for the Gabrb2 gene 119875 = 123 1205782 = 142 The P-M
group had the highest mean (119872 = 332) and the P-V grouphad the lowest (119872 = 275) Finally the Chrm2 gene did notshow significance between groups A statistically significantdifference was not obtained 119875 = 138 1205782 = 138 Thoughnot significant the C-M + L group had the highest mean(119872 = 1056) and the P-V group had the lowest (119872 = 996)
32 Amygdala Genes in the amygdala with an effect sizegreater than 0138 are displayed in Figure 4 In accordancewith Figure 4 Table 3 summarizes between-groupdifferencesin gene expression within the amygdala The Gabra4 genewas 119875 = 001 1205782 = 271 Significant post hoc pairwisecomparisons were such that the C-L group (119872 = 352) had ahigher mean than each of the following (1) P-L (119872 = 304)(2) P-M (119872 = 311) and (3) C-M group (M = 309) Asignificant difference for the Slc5a7 gene was found119875 = 0241205782
= 195 The C-V group had the highest mean (119872 = 775)and the P-L group had the lowest (119872 = 667) howeveras per the post hoc test this pairwise comparison was notsignificant Significant between-group differences were alsofound for the Drd2 gene 119875 = 02 1205782 = 189 There was asignificant difference between P-V (119872 = 676) and the P-Mgroup (119872 = 448) The Drd1a gene was also significant 119875 =038 1205782 = 181 A significant difference was found betweenthe C-M + L (119872 = 641) and the P-M group (M = 483)Lastly the Glra2 showed significance 119875 = 04 1205782 = 179There was one significant post hoc test between the C-M + Lgroup with the higher mean (119872 = 562) and the P-M groupwith the lower mean (119872 = 489)
Again the following genes were not found to be signifi-cant however based on the large effect size (variability) usedin the study these genes are included in Table 3 No differencewas found for the Chrna6 gene 119875 = 075 1205782 = 159 The C-M group had the highest mean (119872 = 1651) and the C-M +L group had the lowest (119872 = 155) The Htr3a gene showedno significant difference between the groups 119875 = 081 1205782 =156 The C-L group had the highest mean (119872 = 674) andthe P-M + L group had the lowest (119872 = 635) The Chrnb4also showed no significant difference 119875 = 087 1205782 = 154The C-V group had the highest mean (119872 = 1159) and theC-M + L group had the lowest (119872 = 1043) No significantdifference was found for the Gabrd gene between the groups119875 = 089 1205782 = 153TheC-M+L group had the highestmean(119872 = 523) and the P-M group had the lowest (119872 = 456)The Prima1 gene did not have a significant difference betweenthe groups 119875 = 111 1205782 = 145 The P-V group had thehighest mean (119872 = 694) and the P-M group had the lowest(119872 = 625) No difference between the groups was found forthe Chrna3 gene 119875 = 127 1205782 = 141 The P-L group had thehighest mean (119872 = 1056) and the C-M + L group had thelowest (119872 = 967)
4 Discussion
L-theanine has previously been demonstrated to produceanxiolytic effects alone or in combination with concomitantadministered benzodiazepines [7] No studies have examined
The Scientific World Journal 5
2000
1500
1000
Hippocampal
Log 2 (fold difference)
minuslo
g Pva
lue
minus00750 minus00500 minus00250 00000 00250 00500
0500
0000
(a)
3000
2000
1000
Amygdalar
Log 2 (fold difference)
minuslo
g Pva
lue
0000minus0150 minus0100 minus0050 0000 0050 0100 0150
(b)
Figure 2 Differential expression of (a) hippocampal and (b) amygdalar mRNA between groups Volcano plot between Log 2 fold change onthe 119909-axis (for neurotransmitter receptors and regulators in L-theaninemidazolam treated groups 40 PTSD-stressed versus 40 nonstressed(control) groups) and minuslog of 119875 value on the 119910-axis Eighty male Sprague-Dawley rats were injected subcutaneously 30 minutes prior toevaluation of their performance on neurobehavioral tests The rats were then euthanized and cDNA prepared from the hippocampus (a) andamygdala (b) and were subjected to RT2 profiler PCR array for rat neurotransmitter receptor and regulator analysis as described inMaterialsand Methods PCR array profiles were performed for the expression of 88 genes potentially involved in PTSD andor rat neurotransmitterreceptors and neurotransmitter regulation
Hippocampal genes
Egr1 Maoa Anxa9 S100a10 Gabrb2 Chrm2
025
020
015
010
005
000
lowast
lowastlowast
Effec
t siz
e (1205782)
Large effect size (gt0138)
Figure 3 Effect size of gene changes in the hippocampus Genesdemonstrating a large effect size (1205782 gt 138) in the hippocampus asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
the administration of L-theaninersquos effects on PTSD-inducedchanges in brain gene expression We explored the modu-lation of gene expression in PTSD-induced changes in thehippocampus and amygdala and the effects of L-theanine andmidazolam on these changes
41 Hippocampus We found a downregulation of the Egr1gene in the PTSD-vehicle group compared to the PTSD-midazolam group Egr1 is an early growth response nuclear
Amygdalar genes
030
025
020
015
010
005
000
Effec
t siz
e (1205782)
Gab
ra4
Slc5
a7
Drd2
Drd1
a
Glra
2
Chrn
a6
Htr3
a
Chrn
b4
Gab
rd
Prim
a1
Chrn
a3
lowast
lowastlowast lowast lowast
Large effect sizes (gt0138)
Figure 4 Effect size of gene changes in the amygdala Genesdemonstrating a large effect size (1205782 gt 138) in the amygdala asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
transcription factor that plays a role in regulating the expres-sion of genes required for learning and memory [23 24] Inparticular previous studies have demonstrated a role for Egr1in hippocampal long-term potentiation a process critical tolearning and memory [25] Expression of the Maoa gene wasdecreased in the PTSD-vehicle group compared to the controlgroup that received L-theanine and midazolam
Decreased Maoa enzymatic activity leads to increasedconcentrations of its substrate biogenic amines including
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

4 The Scientific World Journal
to compare the 8 groups of which each unit was randomlyallocated All assumptions were examined (eg homogeneityof variance univariate normality etc) and in the event thatthe assumptions are not tenable remedial (eg transfor-mation) or alternative (eg nonparametric tests such asKruskal-Wallis) strategies were consideredMultiple compar-ison procedures (MCP) were performed in the event thatsignificance (120572 = 05) was obtained with the Tukey posthoc test A variance explained statistic eta-squared (1205782) wasused as the reported effect size and though ascertainingwhat constitutes a smallmediumlarge effect is primarily afunction of the contextdiscipline for this report 01059138was smallmediumlarge [22] All data were statisticallyanalyzed based on gene cycle thresholds (CTs) normalized tofive housekeeping genes per QIAGEN procedures [15 16]
3 Results
This study was designed to investigate the effects of L-theanine on PTSD-induced changes in rat brain gene expres-sion Eighty-eight genes were investigated in this study Dataanalysis showed a number of significant differences in geneexpression in both the hippocampus and amygdala Volcanoplots were developed to compare the gene expression of the40 nonstressed to the 40 PTSD-stressed rats for both thehippocampus and amygdala as demonstrated in Figure 2Thevolcano plots graphically display the relationship of ndashlog 10 119875value and the Log 2 fold change for the combined treatmentand control groups for all of the genes For the 8-group one-way ANOVA a large effect size (1205782 gt 138) was chosen toserve as a springboard for discussion and interpretation ofrelative between-group differences on gene expression
31 Hippocampus In the hippocampus the genes with aneffect size gt 0138 are displayed in Figure 3 In accordancewith Figure 3 Table 2 summarizes between-group changesin gene expression in the hippocampus The Egr1 gene wasfound to be significant and between-group differences wereobtained 119875 = 011 1205782 = 218 There was one significantpost hoc test with the P-M group having a significantly highermean (119872 = 558) than the P-V group (119872 = 457) Asignificant difference was found between groups for theMaoagene 119875 = 024 1205782 = 194 The post hoc test found onesignificant pairwise comparison (1) the C-M + L group hadthe highest mean (119872 = 332) and the P-V group had thelowest (119872 = 312) The Anxa9 gene was found to havesignificance between groups 119875 = 031 1205782 = 187 Multiplecomparison tests resulted in one significant pairwise resultthe P-M group had a significantly higher mean (119872 = 1073)than the P-V group (119872 = 1006)
The following genes in the hippocampus did not showsignificant differences between groups however due to thelarge effect size (variance explained) these genes are presentedin Table 2The S100a10 gene did not have significant between-group differences 119875 = 06 1205782 = 166 The P-M group had thehighest mean (119872 = 42) and the P-V group had the lowest(119872 = 382) Significant between-group differences were notobtained for the Gabrb2 gene 119875 = 123 1205782 = 142 The P-M
group had the highest mean (119872 = 332) and the P-V grouphad the lowest (119872 = 275) Finally the Chrm2 gene did notshow significance between groups A statistically significantdifference was not obtained 119875 = 138 1205782 = 138 Thoughnot significant the C-M + L group had the highest mean(119872 = 1056) and the P-V group had the lowest (119872 = 996)
32 Amygdala Genes in the amygdala with an effect sizegreater than 0138 are displayed in Figure 4 In accordancewith Figure 4 Table 3 summarizes between-groupdifferencesin gene expression within the amygdala The Gabra4 genewas 119875 = 001 1205782 = 271 Significant post hoc pairwisecomparisons were such that the C-L group (119872 = 352) had ahigher mean than each of the following (1) P-L (119872 = 304)(2) P-M (119872 = 311) and (3) C-M group (M = 309) Asignificant difference for the Slc5a7 gene was found119875 = 0241205782
= 195 The C-V group had the highest mean (119872 = 775)and the P-L group had the lowest (119872 = 667) howeveras per the post hoc test this pairwise comparison was notsignificant Significant between-group differences were alsofound for the Drd2 gene 119875 = 02 1205782 = 189 There was asignificant difference between P-V (119872 = 676) and the P-Mgroup (119872 = 448) The Drd1a gene was also significant 119875 =038 1205782 = 181 A significant difference was found betweenthe C-M + L (119872 = 641) and the P-M group (M = 483)Lastly the Glra2 showed significance 119875 = 04 1205782 = 179There was one significant post hoc test between the C-M + Lgroup with the higher mean (119872 = 562) and the P-M groupwith the lower mean (119872 = 489)
Again the following genes were not found to be signifi-cant however based on the large effect size (variability) usedin the study these genes are included in Table 3 No differencewas found for the Chrna6 gene 119875 = 075 1205782 = 159 The C-M group had the highest mean (119872 = 1651) and the C-M +L group had the lowest (119872 = 155) The Htr3a gene showedno significant difference between the groups 119875 = 081 1205782 =156 The C-L group had the highest mean (119872 = 674) andthe P-M + L group had the lowest (119872 = 635) The Chrnb4also showed no significant difference 119875 = 087 1205782 = 154The C-V group had the highest mean (119872 = 1159) and theC-M + L group had the lowest (119872 = 1043) No significantdifference was found for the Gabrd gene between the groups119875 = 089 1205782 = 153TheC-M+L group had the highestmean(119872 = 523) and the P-M group had the lowest (119872 = 456)The Prima1 gene did not have a significant difference betweenthe groups 119875 = 111 1205782 = 145 The P-V group had thehighest mean (119872 = 694) and the P-M group had the lowest(119872 = 625) No difference between the groups was found forthe Chrna3 gene 119875 = 127 1205782 = 141 The P-L group had thehighest mean (119872 = 1056) and the C-M + L group had thelowest (119872 = 967)
4 Discussion
L-theanine has previously been demonstrated to produceanxiolytic effects alone or in combination with concomitantadministered benzodiazepines [7] No studies have examined
The Scientific World Journal 5
2000
1500
1000
Hippocampal
Log 2 (fold difference)
minuslo
g Pva
lue
minus00750 minus00500 minus00250 00000 00250 00500
0500
0000
(a)
3000
2000
1000
Amygdalar
Log 2 (fold difference)
minuslo
g Pva
lue
0000minus0150 minus0100 minus0050 0000 0050 0100 0150
(b)
Figure 2 Differential expression of (a) hippocampal and (b) amygdalar mRNA between groups Volcano plot between Log 2 fold change onthe 119909-axis (for neurotransmitter receptors and regulators in L-theaninemidazolam treated groups 40 PTSD-stressed versus 40 nonstressed(control) groups) and minuslog of 119875 value on the 119910-axis Eighty male Sprague-Dawley rats were injected subcutaneously 30 minutes prior toevaluation of their performance on neurobehavioral tests The rats were then euthanized and cDNA prepared from the hippocampus (a) andamygdala (b) and were subjected to RT2 profiler PCR array for rat neurotransmitter receptor and regulator analysis as described inMaterialsand Methods PCR array profiles were performed for the expression of 88 genes potentially involved in PTSD andor rat neurotransmitterreceptors and neurotransmitter regulation
Hippocampal genes
Egr1 Maoa Anxa9 S100a10 Gabrb2 Chrm2
025
020
015
010
005
000
lowast
lowastlowast
Effec
t siz
e (1205782)
Large effect size (gt0138)
Figure 3 Effect size of gene changes in the hippocampus Genesdemonstrating a large effect size (1205782 gt 138) in the hippocampus asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
the administration of L-theaninersquos effects on PTSD-inducedchanges in brain gene expression We explored the modu-lation of gene expression in PTSD-induced changes in thehippocampus and amygdala and the effects of L-theanine andmidazolam on these changes
41 Hippocampus We found a downregulation of the Egr1gene in the PTSD-vehicle group compared to the PTSD-midazolam group Egr1 is an early growth response nuclear
Amygdalar genes
030
025
020
015
010
005
000
Effec
t siz
e (1205782)
Gab
ra4
Slc5
a7
Drd2
Drd1
a
Glra
2
Chrn
a6
Htr3
a
Chrn
b4
Gab
rd
Prim
a1
Chrn
a3
lowast
lowastlowast lowast lowast
Large effect sizes (gt0138)
Figure 4 Effect size of gene changes in the amygdala Genesdemonstrating a large effect size (1205782 gt 138) in the amygdala asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
transcription factor that plays a role in regulating the expres-sion of genes required for learning and memory [23 24] Inparticular previous studies have demonstrated a role for Egr1in hippocampal long-term potentiation a process critical tolearning and memory [25] Expression of the Maoa gene wasdecreased in the PTSD-vehicle group compared to the controlgroup that received L-theanine and midazolam
Decreased Maoa enzymatic activity leads to increasedconcentrations of its substrate biogenic amines including
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
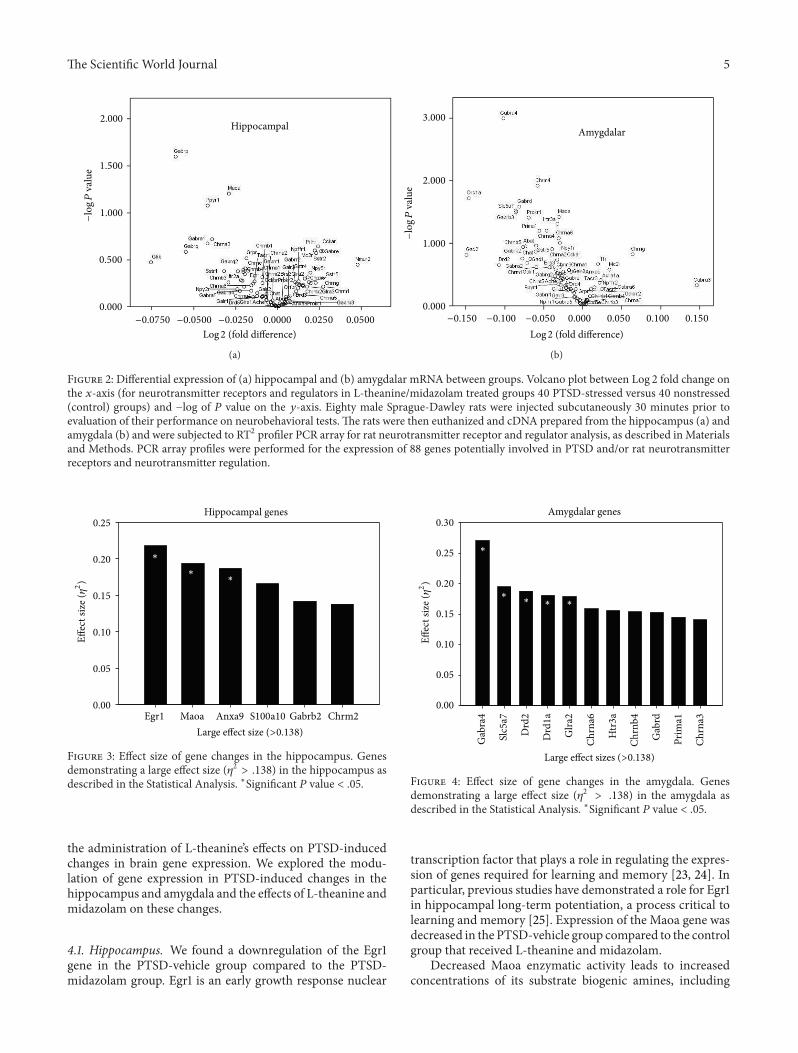
The Scientific World Journal 5
2000
1500
1000
Hippocampal
Log 2 (fold difference)
minuslo
g Pva
lue
minus00750 minus00500 minus00250 00000 00250 00500
0500
0000
(a)
3000
2000
1000
Amygdalar
Log 2 (fold difference)
minuslo
g Pva
lue
0000minus0150 minus0100 minus0050 0000 0050 0100 0150
(b)
Figure 2 Differential expression of (a) hippocampal and (b) amygdalar mRNA between groups Volcano plot between Log 2 fold change onthe 119909-axis (for neurotransmitter receptors and regulators in L-theaninemidazolam treated groups 40 PTSD-stressed versus 40 nonstressed(control) groups) and minuslog of 119875 value on the 119910-axis Eighty male Sprague-Dawley rats were injected subcutaneously 30 minutes prior toevaluation of their performance on neurobehavioral tests The rats were then euthanized and cDNA prepared from the hippocampus (a) andamygdala (b) and were subjected to RT2 profiler PCR array for rat neurotransmitter receptor and regulator analysis as described inMaterialsand Methods PCR array profiles were performed for the expression of 88 genes potentially involved in PTSD andor rat neurotransmitterreceptors and neurotransmitter regulation
Hippocampal genes
Egr1 Maoa Anxa9 S100a10 Gabrb2 Chrm2
025
020
015
010
005
000
lowast
lowastlowast
Effec
t siz
e (1205782)
Large effect size (gt0138)
Figure 3 Effect size of gene changes in the hippocampus Genesdemonstrating a large effect size (1205782 gt 138) in the hippocampus asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
the administration of L-theaninersquos effects on PTSD-inducedchanges in brain gene expression We explored the modu-lation of gene expression in PTSD-induced changes in thehippocampus and amygdala and the effects of L-theanine andmidazolam on these changes
41 Hippocampus We found a downregulation of the Egr1gene in the PTSD-vehicle group compared to the PTSD-midazolam group Egr1 is an early growth response nuclear
Amygdalar genes
030
025
020
015
010
005
000
Effec
t siz
e (1205782)
Gab
ra4
Slc5
a7
Drd2
Drd1
a
Glra
2
Chrn
a6
Htr3
a
Chrn
b4
Gab
rd
Prim
a1
Chrn
a3
lowast
lowastlowast lowast lowast
Large effect sizes (gt0138)
Figure 4 Effect size of gene changes in the amygdala Genesdemonstrating a large effect size (1205782 gt 138) in the amygdala asdescribed in the Statistical Analysis lowastSignificant 119875 value lt 05
transcription factor that plays a role in regulating the expres-sion of genes required for learning and memory [23 24] Inparticular previous studies have demonstrated a role for Egr1in hippocampal long-term potentiation a process critical tolearning and memory [25] Expression of the Maoa gene wasdecreased in the PTSD-vehicle group compared to the controlgroup that received L-theanine and midazolam
Decreased Maoa enzymatic activity leads to increasedconcentrations of its substrate biogenic amines including
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
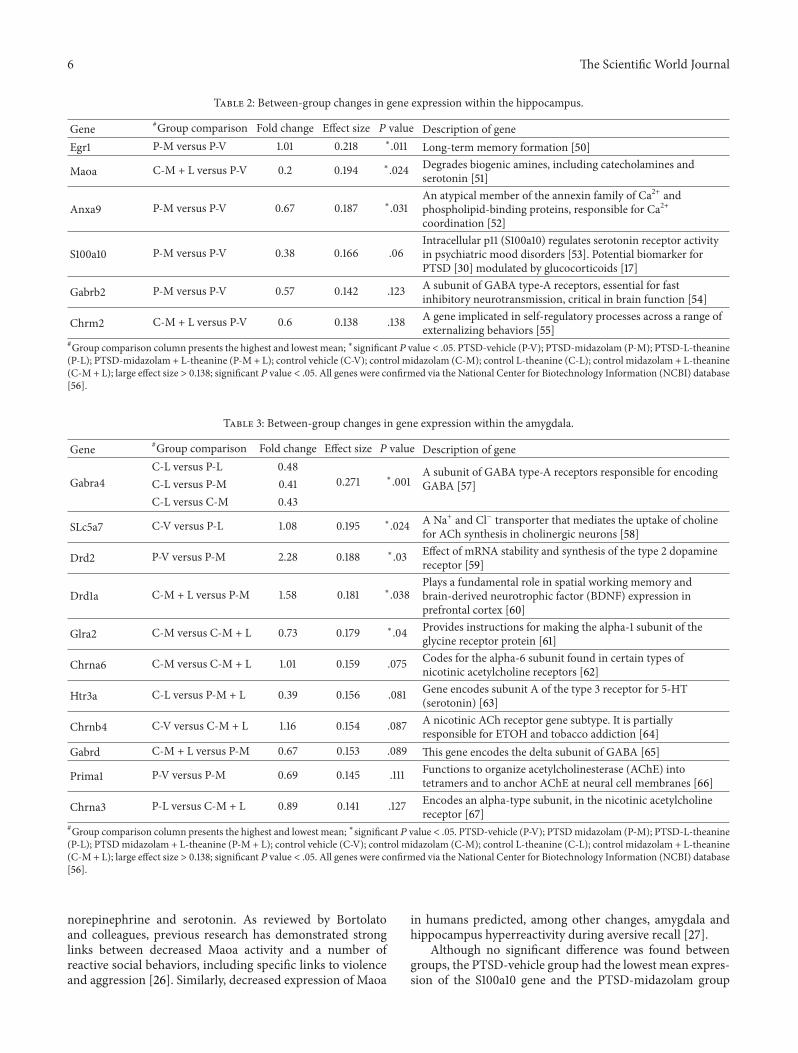
6 The Scientific World Journal
Table 2 Between-group changes in gene expression within the hippocampus
Gene Group comparison Fold change Effect size 119875 value Description of geneEgr1 P-M versus P-V 101 0218 lowast011 Long-term memory formation [50]
Maoa C-M + L versus P-V 02 0194 lowast024 Degrades biogenic amines including catecholamines andserotonin [51]
Anxa9 P-M versus P-V 067 0187 lowast031An atypical member of the annexin family of Ca2+ andphospholipid-binding proteins responsible for Ca2+coordination [52]
S100a10 P-M versus P-V 038 0166 06Intracellular p11 (S100a10) regulates serotonin receptor activityin psychiatric mood disorders [53] Potential biomarker forPTSD [30] modulated by glucocorticoids [17]
Gabrb2 P-M versus P-V 057 0142 123 A subunit of GABA type-A receptors essential for fastinhibitory neurotransmission critical in brain function [54]
Chrm2 C-M + L versus P-V 06 0138 138 A gene implicated in self-regulatory processes across a range ofexternalizing behaviors [55]
Group comparison column presents the highest and lowest mean lowastsignificant119875 value lt 05 PTSD-vehicle (P-V) PTSD-midazolam (P-M) PTSD-L-theanine(P-L) PTSD-midazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
Table 3 Between-group changes in gene expression within the amygdala
Gene Group comparison Fold change Effect size 119875 value Description of gene
Gabra4C-L versus P-L 048
0271 lowast001A subunit of GABA type-A receptors responsible for encodingGABA [57]C-L versus P-M 041
C-L versus C-M 043
SLc5a7 C-V versus P-L 108 0195 lowast024 A Na+ and Clminus transporter that mediates the uptake of cholinefor ACh synthesis in cholinergic neurons [58]
Drd2 P-V versus P-M 228 0188 lowast03 Effect of mRNA stability and synthesis of the type 2 dopaminereceptor [59]
Drd1a C-M + L versus P-M 158 0181 lowast038Plays a fundamental role in spatial working memory andbrain-derived neurotrophic factor (BDNF) expression inprefrontal cortex [60]
Glra2 C-M versus C-M + L 073 0179 lowast04 Provides instructions for making the alpha-1 subunit of theglycine receptor protein [61]
Chrna6 C-M versus C-M + L 101 0159 075 Codes for the alpha-6 subunit found in certain types ofnicotinic acetylcholine receptors [62]
Htr3a C-L versus P-M + L 039 0156 081 Gene encodes subunit A of the type 3 receptor for 5-HT(serotonin) [63]
Chrnb4 C-V versus C-M + L 116 0154 087 A nicotinic ACh receptor gene subtype It is partiallyresponsible for ETOH and tobacco addiction [64]
Gabrd C-M + L versus P-M 067 0153 089 This gene encodes the delta subunit of GABA [65]
Prima1 P-V versus P-M 069 0145 111 Functions to organize acetylcholinesterase (AChE) intotetramers and to anchor AChE at neural cell membranes [66]
Chrna3 P-L versus C-M + L 089 0141 127 Encodes an alpha-type subunit in the nicotinic acetylcholinereceptor [67]
Group comparison column presents the highest and lowest mean lowastsignificant 119875 value lt 05 PTSD-vehicle (P-V) PTSDmidazolam (P-M) PTSD-L-theanine(P-L) PTSDmidazolam + L-theanine (P-M + L) control vehicle (C-V) control midazolam (C-M) control L-theanine (C-L) control midazolam + L-theanine(C-M + L) large effect size gt 0138 significant 119875 value lt 05 All genes were confirmed via the National Center for Biotechnology Information (NCBI) database[56]
norepinephrine and serotonin As reviewed by Bortolatoand colleagues previous research has demonstrated stronglinks between decreased Maoa activity and a number ofreactive social behaviors including specific links to violenceand aggression [26] Similarly decreased expression of Maoa
in humans predicted among other changes amygdala andhippocampus hyperreactivity during aversive recall [27]
Although no significant difference was found betweengroups the PTSD-vehicle group had the lowest mean expres-sion of the S100a10 gene and the PTSD-midazolam group
The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

The Scientific World Journal 7
the highest The S100a10 gene and its associated protein p11are important in the regulation of serotonin receptors [28]and decreases in S100a10 gene expression have been linkedto PTSD major depressive disorder and bipolar disorder andproposed as a likely biomarker for suicidality [29 30]
A downregulation of the Gabrb2 gene in the PTSD-vehicle group compared to the PTSD-midazolam group wassimilar to Zhao and colleagues findings of a significantlydecreased expression of the Gabrb2 gene in the anteriorcingulate cortex in subjects suffering from major depressivedisorder or bipolar disorder [31]
A downregulation of the Anxa9 gene was found inthe PTSD-vehicle group compared to the PTSD-midazolamgroup The Anxa9 gene part of the calcium-dependentphospholipid-binding annexin family may play a role insuppression of apoptosis and is currently under investigationas a therapeutic target for the treatment of breast cancer[32] In a study using the Flinders rat model Chrm2 geneexpression levels were found to be significantly lower in theFlinders depression sensitive line [33]
42 Amygdala According to Ponomarev and colleaguesdownregulation of the Gabra4 gene can be correlated withstress enhanced fear learning (SEFL) in rats These datasuggest a significant role of GABA signaling in SEFL Themethod of inducing SEFL was similar to the method ofinducing PTSD in our study [34] Interestingly our findingssuggest an upregulation of this gene in the nonstressed-L-theanine group compared to PTSD-L-theanine PTSD-midazolam and nonstressed-midazolam groups
PCR analysis of tissue samples from the amygdala showeda 228-fold increase in Drd2 gene expression in the PTSD-vehicle group compared to the PTSD-midazolam groupwhich is inconsistent with the findings of a study thatinvestigated the effects of maternal deprivation and chronicmild stress in rats [35] Zhu and colleagues showed thatmRNA levels of the Drd2 gene were significantly decreased inall stressed groups compared to controls which enabled theinvestigators to correlate a downregulation of the Drd2 genewith depressive phenotypes [35]
Drd1a genes were significantly downregulated in thePTSD-midazolam group compared to the nonstressed-midazolam + L-theanine group concurrent with the resultsof another study exploring the effects of stress on the mouseprefrontal cortex transcriptome demonstrating a deregula-tion of the Drd1a gene in chronicmild stress exposed subjects[36]
The SLc5a7 gene was significantly downregulated inthe PTSD-L-theanine group compared to the nonstressed-vehicle group In comparison the SLc5a7 genewas found to beupregulated in tasks that required higher levels of attentionand it was found that decreased levels of cholinergic activityare involved in ADHD [37]
Prima1 DNAmethylation increased by 10ndash15 in subjectssuffering frommajor depressive disorder [38] which interest-ingly shows some correlation with our finding of the highestlevel of Prima1 gene expression in the PTSD-vehicle groupwhich is a 069-fold increase in expression compared with thePTSD-midazolam group with the lowest level of expression
Although no studies have investigated gene expressionchanges of Chrnb4 in PTSD or anxiety variations in thisgene were shown to increase sensitivity to nicotinersquos effectson attention mood and vulnerability to tobacco dependence[39] The Htr3a gene and serotonin have been linked to anxi-ety andmoods aswell as psychoses [40] and interactionswith5HT3 receptors are also involved with aggressive behaviordepression and the stress response [41] The Gabrd geneis a potential susceptibility gene for childhood-onset mooddisorders [42] and variations in this gene are associatedwith a range of genetic epilepsy syndromes [43] includingfebrile seizures [44] Chrna6 and Chrna3 genes are involvedin dopaminergic neurotransmission specifically noted inmesodiencephalic dopamine releasing neurons [45] At thistime there is no sufficient research linking the Glra2 genewith PTSD anxiety or mood disorders
5 Conclusions
RT-PCR analysis revealed significant changes betweengroups in several genes implicated in a variety of disordersranging from PTSD anxiety mood disorders and substancedependence These data further elucidate the transcriptomicfootprint of PTSD in the rodent amygdala and hippocampusas well as transcriptome changes effected by L-theanine andmidazolam interventions
These different neurotransmitter genes are significantbiologically and clinically as they are associated with var-ious central nervous system pathologies The hippocampalgenes related to long-term potentiation (Egr1) violenceand aggression (Maoa) and major depressive and bipolardisorder (S100a10 serotonin receptors) were shown to bedownregulated in PTSD animals and expression levels sig-nificantly changed Other neurotransmitter genes Gabrb2and Chrm2 were also modulated across treatment groupsand warrant further exploration as they are associated withdepression related clinical conditions Amygdalar genes thatwere changed across treatment groups also were related topsychological conditions such as depression (Drd2) stressenhanced fear learning (Gabra4) chronicmild stress (Drd1a)and anxiety mood disorders and psychoses (Htr3a) Thesefindings taken together demonstrate a relationship of possiblegene expression changes in important and known neuro-transmitter system genes and associated clinical pathologiesThis study lays the foundation for future studies to interrogateand investigate the translation of these genes into proteinsand the potential for these proteins to act as sites forpharmacologic therapeutic intervention
This understanding of the effect of PTSD at the levelof mRNA transcription contributes to a more completeunderstanding of the overall metabonomics of PTSD andmay allow for more targeted pharmacologic intervention inthe future
The next step to consider in clarifying the molecularmechanism of PTSD may be to characterize PTSD-inducedchanges to the rat proteome Changes in mRNA expressionare often linked to changes in protein expression but not in apredictable manner Small changes in mRNA expression can
8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

8 The Scientific World Journal
lead to variable changes in corresponding protein expressionWe propose repeating the current experiment using proteinassay methods to correlate changes in mRNA expressionwith changes in protein expression induced by PTSD andthe effects of L-theanine and midazolam on these changesMultiple methods and computational approaches to proteinassays are available [46]
We anticipate further investigation into the networkof gene relationships between control and PTSD-inducedrats using bioinformatic methods such as gene enrichmentanalysis and network enrichment analysis Characterizationof a comprehensive proteomic network in both control andPTSD-induced rats will allow for comparative proteomicanalysis that may lead to the identification of biomarkersfor PTSD susceptibility as well as potential targets for phar-macologic interventions [47ndash49] Additionally we anticipateinvestigation into the chemical components of other herbalsor pharmaceuticals that may have similar effects on geneexpression at the level of mRNA transcription with a partic-ular emphasis on those with demonstrated efficacy in treatingPTSD
Disclaimer
Theviews expressed in this paper are those of the authors anddo not reflect the official policy or position of theDepartmentof the Army Department of Defense or the US Government
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The researchers would like to acknowledge their families andthe support they provide as well as the US Army GraduateProgram in Anesthesia Nursing Faculty for their expertiseand guidance In addition the researchers are very gratefulto Jerry Merrill PhD Ms Kathleen McKay VSB FacilityManager at the US Army Institute of Surgical Research theUS Army Institute of Surgical Research Dr Dale Glaser andthe Department of Clinical Investigations at Brooke ArmyMedical Center for the use of their facilities and supportThisstudy was funded by TriService Nursing Research Program
References
[1] M Friedman PTSDHistory andOverviewTheNational Centerfor PTSD Department of Veterans Affairs 2007
[2] L Zhang R Zhou G Xing C J Hough X Li and H LildquoIdentification of genemarkers based onwell validated and sub-categorized stressed animals for potential clinical applicationsin PTSDrdquoMedical Hypotheses vol 66 no 2 pp 309ndash314 2006
[3] S Bent ldquoHerbal medicine in the United States review ofefficacy safety and regulation grand Rounds at University ofCalifornia San Francisco Medical Centerrdquo Journal of GeneralInternal Medicine vol 23 no 6 pp 854ndash859 2008
[4] D Chu T Okubo Y Nagato and H Yokogoshi ldquoL-theaninemdasha unique amino acid of green tea and its relaxation effect inhumansrdquo Trends in Food Science and Technology vol 10 no 6-7pp 199ndash204 1999
[5] G Eschenauer and B V Sweet ldquoPharmacology and therapeuticuses of theaninerdquo The American Journal of Health-SystemPharmacy vol 63 no 1 pp 26ndash30 2006
[6] K Lu M A Gray C Oliver et al ldquoThe acute effects ofL-theanine in comparison with alprazolam on anticipatoryanxiety in humansrdquo Human Psychopharmacology vol 19 no 7pp 457ndash465 2004
[7] T Heese J Jenkinson C Love et al ldquoAnxiolytic effects ofL-theaninemdasha component of green tea-when combined withmidazolam in the male Sprague-Dawley ratrdquo AANA Journalvol 77 no 6 pp 445ndash449 2009
[8] I Liberzon and B Martis ldquoNeuroimaging studies of emotionalresponses in PTSDrdquoAnnals of theNewYorkAcademy of Sciencesvol 1071 pp 87ndash109 2006
[9] R K Pitman L M Shin and S L Rauch ldquoInvestigating thepathogenesis of posttraumatic stress disorder with neuroimag-ingrdquo Journal of Clinical Psychiatry vol 62 supplement 17 pp47ndash54 2001
[10] R J Ursano L Zhang H Li et al ldquoPTSD and traumatic stressfrom gene to community and bench to bedsiderdquo Brain Researchvol 1293 pp 2ndash12 2009
[11] T Sugiyama and Y Sadzuka ldquoCombination of theanine withdoxorubicin inhibits hepatic metastasis of M5076 ovariansarcomardquo Clinical Cancer Research vol 5 no 2 pp 413ndash4161999
[12] R J Servatius J E Ottenweller and B H Natelson ldquoDelayedstartle sensitization distinguishes rats exposed to one or threestress sessions further evidence toward an animal model ofPTSDrdquo Biological Psychiatry vol 38 no 8 pp 539ndash546 1995
[13] T Garrick N Morrow A Y Shalev and S Eth ldquoStress-induced enhancement of auditory startle an animal model ofposttraumatic stress disorderrdquo Psychiatry vol 64 no 4 pp346ndash354 2001
[14] T G Heffner J A Hartman and L S Seiden ldquoA rapid methodfor the regional dissection of the rat brainrdquo PharmacologyBiochemistry and Behavior vol 13 no 3 pp 453ndash456 1980
[15] N Kozlovsky M A Matar Z Kaplan J Zohar and H CohenldquoA distinct pattern of intracellular glucocorticoid-relatedresponses is associated with extreme behavioral response tostress in an animal model of post-traumatic stress disorderrdquoEuropean Neuropsychopharmacology vol 19 no 11 pp 759ndash7712009
[16] Rat Neurotransmitter Receptors and Regulators PCR Arrayhttpwwwsabiosciencescomrt pcr productHTMLPARN-060Ahtml
[17] L Zhang H Li T P Su et al ldquop11 is up-regulated in theforebrain of stressed rats by glucocorticoid acting via twospecific glucocorticoid response elements in the p11 promoterrdquoNeuroscience vol 153 no 4 pp 1126ndash1134 2008
[18] X JiangGXing C YangAVerma L Zhang andH Li ldquoStressimpairs 5-HT
2119860
receptor-mediated serotonergic facilitation ofGABA release in juvenile rat basolateral amygdalardquo Neuropsy-chopharmacology vol 34 no 2 pp 410ndash423 2009
[19] S T Manion E H Gamble and H Li ldquoPrazosin administeredprior to inescapable stressor blocks subsequent exaggeration ofacoustic startle response in ratsrdquo Pharmacology Biochemistryand Behavior vol 86 no 3 pp 559ndash565 2007
The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

The Scientific World Journal 9
[20] L J Miller ldquoPrazosin for the treatment of posttraumatic stressdisorder sleep disturbancesrdquo Pharmacotherapy vol 28 no 5pp 656ndash666 2008
[21] M A Raskind E R Peskind D J Hoff et al ldquoA parallel groupplacebo controlled study of prazosin for trauma nightmares andsleep disturbance in combat veterans with post-traumatic stressdisorderrdquo Biological Psychiatry vol 61 no 8 pp 928ndash934 2007
[22] J Cohen Statistical Power Analysis for the Behavioral SciencesL Erlbaum Associates Hillsdale NJ USA 2nd edition 1988
[23] S Davis B Bozon and S Laroche ldquoHow necessary is theactivation of the immediate early gene zif268 in synapticplasticity and learningrdquo Behavioural Brain Research vol 142no 1-2 pp 17ndash30 2003
[24] E Knapska and L Kaczmarek ldquoA gene for neuronal plas-ticity in the mammalian brain Zif268Egr-1NGFI-A Krox-24TIS8ZENKrdquo Progress in Neurobiology vol 74 no 4 pp183ndash211 2004
[25] Z Penke C Chagneau and S Laroche ldquoContribution ofEgr1Zif268 to activity-dependent ArcArg31 transcription inthe dentate gyrus and area CA1 of the hippocampusrdquo Frontiersin Behavioral Neuroscience vol 5 article 48 2011
[26] M Bortolato K Chen S C Godar et al ldquoSocial deficits andperseverative behaviors but not overt aggression in MAO-Ahypomorphic micerdquo Neuropsychopharmacology vol 36 no 13pp 2674ndash2688 2011
[27] A Meyer-Lindenberg J W Buckholtz B Kolachana et alldquoNeuralmechanisms of genetic risk for impulsivity and violencein humansrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 103 no 16 pp 6269ndash62742006
[28] P Svenningsson K Chergui I Rachleff et al ldquoAlterationsin 5-HT1B receptor function by p11 in depression-like statesrdquoScience vol 311 no 5757 pp 77ndash80 2006
[29] T-P Su L Zhang M-Y Chung et al ldquoLevels of the potentialbiomarker p11 in peripheral blood cells distinguish patientswith PTSD from those with other major psychiatric disordersrdquoJournal of Psychiatric Research vol 43 no 13 pp 1078ndash10852009
[30] L Zhang T Su K Choi et al ldquoP11 (S100A10) as a potentialbiomarker of psychiatric patients at risk of suiciderdquo Journal ofPsychiatric Research vol 45 no 4 pp 435ndash441 2011
[31] J Zhao A-M Bao X-R Qi et al ldquoGene expression of GABAand glutamate pathway markers in the prefrontal cortex ofnon-suicidal elderly depressed patientsrdquo Journal of AffectiveDisorders vol 138 no 3 pp 494ndash502 2012
[32] Z Hu R Neve Y Guan and J Gray ldquoIdentification of newtherapeutic targets of breast cancer using siRNArdquo inProceedingsof the AACR Meeting Abstracts Online pp 49ndash56 2007
[33] E Blaveri F Kelly A Mallei et al ldquoExpression profiling of agenetic animal model of depression reveals novel molecularpathways underlying depressive-like behavioursrdquo PLoS ONEvol 5 no 9 Article ID e12596 pp 1ndash10 2010
[34] I PonomarevV Rau E I Eger RAHarris andM S FanselowldquoAmygdala transcriptome and cellular mechanisms underlyingstress-enhanced fear learning in a rat model of posttraumaticstress disorderrdquo Neuropsychopharmacology vol 35 no 6 pp1402ndash1411 2010
[35] X Zhu S Peng S Zhang and X Zhang ldquoStress-induceddepressive behaviors are correlated with Par-4 and DRD2expression in rat striatumrdquoBehavioural Brain Research vol 223no 2 pp 329ndash335 2011
[36] P Lisowski M Wieczorek J Goscik et al ldquoEffects of chronicstress on prefrontal cortex transcriptome inmice displaying dif-ferent genetic backgroundsrdquo Journal of Molecular Neurosciencevol 50 no 1 pp 33ndash57 2013
[37] B A EnglishMKHahn I R Gizer et al ldquoCholine transportergene variation is associated with attention-deficit hyperactivitydisorderrdquo Journal of Neurodevelopmental Disorders vol 1 no 4pp 252ndash263 2009
[38] S Sabunciyan M J Aryee R A Irizarry et al ldquoGenome-wideDNAmethylation scan inmajor depressive disorderrdquoPloSONEvol 7 no 4 Article ID e34451 2012
[39] M R Picciotto and P J Kenny ldquoMolecular mechanismsunderlying behaviors related to nicotine addictionrdquoCold SpringHarbor Perspectives in Medicine vol 3 no 1 Article ID a0121122013
[40] T Iidaka N Ozaki A Matsumoto et al ldquoA variant C178T inthe regulatory region of the serotonin receptor gene HTR3Amodulates neural activation in the human amygdalardquo Journalof Neuroscience vol 25 no 27 pp 6460ndash6466 2005
[41] N K Popova and V S Naumenko ldquo5-HT1A receptor as a keyplayer in the brain 5-HT systemrdquo Reviews in the Neurosciencesvol 24 no 2 pp 191ndash204 2013
[42] Y Feng K Kapornai E Kiss et al ldquoAssociation of the GABRDgene and childhood-onset mood disordersrdquo Genes Brain andBehavior vol 9 no 6 pp 668ndash672 2010
[43] R L Macdonald J-Q Kang and M J Gallagher ldquoMutationsin GABA
119860
receptor subunits associated with genetic epilepsiesrdquoThe Journal of Physiology vol 588 no 11 pp 1861ndash1869 2010
[44] J Nakayama ldquoProgress in searching for the febrile seizuresusceptibility genesrdquo Brain and Development vol 31 no 5 pp359ndash365 2009
[45] K Chakrabarty L von Oerthel A Hellemons et al ldquoGenomewide expression profiling of the mesodiencephalic region iden-tifies novel factors involved in early and late dopaminergicdevelopmentrdquo Biology Open vol 1 no 8 pp 693ndash704 2012
[46] M Vaudel A Sickmann and L Martens ldquoCurrent methodsfor global proteome identificationrdquoExpert Review of Proteomicsvol 9 no 5 pp 519ndash532 2012
[47] L Abatangelo R Maglietta A Distaso et al ldquoComparativestudy of gene set enrichment methodsrdquo BMC Bioinformaticsvol 10 article 275 2009
[48] A Shojaie and G Michailidis ldquoNetwork enrichment analysis incomplex experimentsrdquo Statistical Applications in Genetics andMolecular Biology vol 9 no 1 article 22 2010
[49] A Alexeyenko W Lee M Pernemalm et al ldquoNetwork enrich-ment analysis extension of gene-set enrichment analysis togene networksrdquo BMC Bioinformatics vol 13 no 1 article 2262012
[50] C Katche A Goldin C Gonzalez P Bekinschtein and JH Medina ldquoMaintenance of long-term memory storage isdependent on late posttraining Egr-1 expressionrdquo Neurobiologyof Learning and Memory vol 98 no 3 pp 220ndash227 2012
[51] Z-Y Chen G S Hotamisligil J-K Huang et al ldquoStructure ofthe human gene for monoamine oxidase type Ardquo Nucleic AcidsResearch vol 19 no 16 pp 4537ndash4541 1991
[52] M Chlystun A Markoff and V Gerke ldquoStructural and func-tional characterisation of the mouse annexin A9 promoterrdquoBiochimica et Biophysica ActamdashMolecular Cell Research vol1742 no 1ndash3 pp 141ndash149 2004
[53] N Hedhli D J Falcone B Huang et al ldquoThe annexinA2S100A10 system in health and disease emerging paradigmsrdquo
10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

10 The Scientific World Journal
Journal of Biomedicine and Biotechnology vol 2012 Article ID406273 13 pages 2012
[54] M K Mulligan X Wang A L Adler K Mozhui L Luand R W Williams ldquoComplex control of GABA(a) receptorsubunit mRNA expression variation covariation and geneticregulationrdquo PLoS ONE vol 7 no 4 Article ID e34586 2012
[55] S J Latendresse J E Bates J A Goodnight et al ldquoDifferentialsusceptibility to adolescent externalizing trajectories examin-ing the interplay between chrm2 and peer group antisocialbehaviorrdquo Child Development vol 82 no 6 pp 1797ndash1814 2011
[56] ldquoGene Databaserdquo httpwwwncbinlmnihgovgene[57] P J Mclean D H Farb and S J Russek ldquoMapping of the 120572
4
subunit gene (GABRA4) to human chromosome 4 defines an1205722
-1205724
-1205731
-1205741
gene cluster further evidence that modernGABA119860
receptor gene clusters are derived from an ancestral clusterrdquoGenomics vol 26 no 3 pp 580ndash586 1995
[58] S Apparsundaram S M Ferguson A L George Jr and RD Blakely ldquoMolecular cloning of a human hemicholinium-3-sensitive choline transporterrdquo Biochemical and BiophysicalResearch Communications vol 276 no 3 pp 862ndash867 2000
[59] J Duan M S Wainwright J M Comeron et al ldquoSynonymousmutations in the human dopamine receptor D2 (DRD2) affectmRNA stability and synthesis of the receptorrdquo Human Molecu-lar Genetics vol 12 no 3 pp 205ndash216 2003
[60] B Xing J Guo X Meng S Wei and S Li ldquoThe dopamine D1but not D3 receptor plays a fundamental role in spatial workingmemory and BDNF expression in prefrontal cortex of micerdquoBehavioural Brain Research vol 235 no 1 pp 36ndash41 2012
[61] J Feng N Craddock I R Jones et al ldquoSystematic screening formutations in the glycine receptor 1205722 subunit gene (GLRA2)inpatients with schizophrenia and other psychiatric diseasesrdquoPsychiatric Genetics vol 11 no 1 pp 45ndash48 2001
[62] E L Meyer D Yoshikami and J M McIntosh ldquoThe neuronalnicotinic acetylcholine receptors 1205724lowast and 1205726lowast differentiallymodulate dopamine release in mouse striatal slicesrdquo Journal ofNeurochemistry vol 105 no 5 pp 1761ndash1769 2008
[63] B Weiss A Mertz E Schrok M Koenen and G RappoldldquoAssignment of a human homolog of the mouse Htr3 receptorgene to chromosome 11q231-q232rdquo Genomics vol 29 no 1 pp304ndash305 1995
[64] X Gallego R J Cox J R Laughlin J A Stitzel and M AEhringer ldquoAlternative CHRNB4 31015840-UTRs mediate the alleliceffects of SNP rs1948 on gene expressionrdquo PLoS ONE vol 8 no5 Article ID e63699 2013
[65] B Sommer A Poustka N K Spurr and P H SeeburgldquoThe murine GABA
119860
receptor 120575-subunit gene structure andassignment to human chromosome 1rdquo DNA and Cell Biologyvol 9 no 8 pp 561ndash568 1990
[66] A L Perrier J Massoulie and E Krejci ldquoPRiMA the mem-brane anchor of acetylcholinesterase in the brainrdquo Neuron vol33 no 2 pp 275ndash285 2002
[67] C M Eng C A Kozak A L Beaudet and H Y ZoghbildquoMapping of multiple subunits of the neuronal nicotinic acetyl-choline receptor to chromosome 15 in man and chromosome 9in mouserdquo Genomics vol 9 no 2 pp 278ndash282 1991
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Copyright © 2022 FDOKUMEN




















