
Effects of inorganic sediment and food concentrations on energetic processes of the zebra mussel,...
13
Effects of inorganic sediment and food concentrations on energetic processes of the zebra mussel, Dreissena polymorpha: implications for growth in turbid rivers Sharook P. Madon, Daniel W. Schneider, James A. Stoeckel, and Richard E. Sparks Abstract: We examined the effects of inorganic suspended sediment (0, 1, 10, and 100 mg⋅L –1 ) and food concentrations (0.1, 0.5, and 2.0 mg particulate organic matter (POM)⋅L –1 ) on clearance, ingestion, respiration, feces, and pseudofeces production rates and assimilation efficiencies of adult zebra mussels (Dreissena polymorpha) under laboratory conditions at 20 ± 1°C. Food quality indices (the proportion of inorganic to organic material in feeding suspensions = IO ratios) ranged from 0.304 to 63.04. Suspended inorganic sediment above 1 mg⋅L –1 greatly reduced clearance rates, ingestion rates, and assimilation efficiencies of zebra mussels, and ingestion of POM was maintained only at high food concentrations. Pseudofeces were produced copiously by zebra mussels in the 100 mg⋅L –1 sediment treatment, and there appeared to be an increased metabolic cost associated with pseudofeces production. Declining food quality caused exponential declines in clearance, ingestion, and absorption rates, water processing potential, and assimilation efficiencies, with the sharpest declines between IO ratios of 0.304 and 5.0. IO ratios in turbid rivers often exceed 1.71, our experimental threshold where scope for growth for zebra mussels was zero. We contend that zebra mussels in turbid rivers may exhibit low growth potential and may not stabilize at the high population densities seen in their lake counterparts. Résumé : Nous avons étudié les effets de la concentration de sédiments inorganiques (0, 1, 10 et 100 mg⋅L –1 ) et d’aliments (0,1, 0,5 et 2,0 mg de matière organique particulaire (MOP)⋅L –1 ) en suspension sur le filtrage, l’ingestion, la respiration, la production d’excréments et de pseudofèces et les taux d’assimilation chez la moule zébrée (Dreissena polymorpha) adulte en laboratoire à 20 ± 1°C. L’indice de qualité de nourriture (rapport matières inorganiques–organiques dans les suspensions d’alimentation = rapport IO) allait de 0,304 à 63,04. Lorsque la concentration de sédiments inorganiques dépassait 1 mg⋅L –1 , le filtrage, l’ingestion et les taux d’assimilation baissaient dans une grande mesure chez la moule zébrée; l’ingestion de MOP ne se maintenait qu’aux concentrations élevées de particules alimentaires. Les moules recevant une concentration de sédiments de 100 mg⋅L –1 ont produit des pseudofèces en quantité; il semble que la production de pseudofèces représente un surcroît pour le métabolisme. La baisse de la qualité de la nourriture s’est traduite par une baisse exponentielle du filtrage, de l’ingestion, du taux d’absorption, du potentiel de transformation de l’eau et des taux d’assimilation, les baisses les plus marquées correspondant aux rapports IO compris entre 0,304 et 5,0. Dans les rivières aux eaux troubles, les rapports IO dépassent souvent 1,71 : dans nos expériences cette valeur était le seuil au-delà duquel les possibilités de croissance de la moule zébrée étaient nulles. Selon nous, il se pourrait que le potentiel de croissance de la moule zébrée dans les cours d’eau troubles soit faible et que les populations ne se stabilisent pas aux densités élevées qu’on trouve dans les lacs. [Traduit par la Rédaction] Introduction Suspended sediments can have profound effects on energetic processes in filter-feeding bivalves. Bivalves reject excess fil- tered material in pseudofeces above certain threshold seston concentrations (Bayne and Newell 1983). Thus, inorganic sus- pended sediments may reduce the amount of food material ingested via “dilution” of organic particles (Widdows et al. 1979; Bricelj and Malouf 1984). Studies with marine bivalves indicate species-specific responses to suspended sediments loads. Species such as Spisula subtruncata and Mytilus edulis which control ingestion of nonfood particles by selecting out such particles via pseudofeces production (Widdows et al. 1979; Kiørboe et al. 1980; Møhlenberg and Kiørboe 1981) are Can. J. Fish. Aquat. Sci. 55: 401–413 (1998) Received December 18, 1996. Accepted June 12, 1997. J13806 S.P. Madon, 1 J.A. Stoeckel, and R.E. Sparks. Illinois Natural History Survey, River Research Laboratory, Long-Term Resource Monitoring Program, 704 N. Schrader, Havana, IL 62644, U.S.A. D.W. Schneider. Department of Urban and Regional Planning, University of Illinois at Urbana–Champaign, 611 E. Lorado Taft Drive, Champaign, IL 61820, U.S.A., and Illinois Natural History Survey, Champaign, IL 61820, U.S.A. 1 Author to whom all correspondence should be addressed. Mailing address: Department of Biological Sciences and Graduate Program in Environmental Science, Pace University, 861 Bedford Road, Pleasantville, NY 10570, U.S.A. e-mail: [email protected] 401 © 1998 NRC Canada
Transcript of Effects of inorganic sediment and food concentrations on energetic processes of the zebra mussel,...

Effects of inorganic sediment and food
concentrations on energetic processes of the
zebra mussel, Dreissena polymorpha:
implications for growth in turbid rivers
Sharook P. Madon, Daniel W. Schneider, James A. Stoeckel, andRichard E. Sparks
Abstract: We examined the effects of inorganic suspended sediment (0, 1, 10, and 100 mg⋅L–1) and food concentrations (0.1,
0.5, and 2.0 mg particulate organic matter (POM)⋅L–1) on clearance, ingestion, respiration, feces, and pseudofeces production
rates and assimilation efficiencies of adult zebra mussels (Dreissena polymorpha) under laboratory conditions at 20 ± 1°C.
Food quality indices (the proportion of inorganic to organic material in feeding suspensions = IO ratios) ranged from 0.304 to
63.04. Suspended inorganic sediment above 1 mg⋅L–1 greatly reduced clearance rates, ingestion rates, and assimilation
efficiencies of zebra mussels, and ingestion of POM was maintained only at high food concentrations. Pseudofeces were
produced copiously by zebra mussels in the 100 mg⋅L–1 sediment treatment, and there appeared to be an increased metabolic
cost associated with pseudofeces production. Declining food quality caused exponential declines in clearance, ingestion, and
absorption rates, water processing potential, and assimilation efficiencies, with the sharpest declines between IO ratios of
0.304 and 5.0. IO ratios in turbid rivers often exceed 1.71, our experimental threshold where scope for growth for zebra
mussels was zero. We contend that zebra mussels in turbid rivers may exhibit low growth potential and may not stabilize at
the high population densities seen in their lake counterparts.
Résumé: Nous avons étudié les effets de la concentration de sédiments inorganiques (0, 1, 10 et 100 mg⋅L–1) et d’aliments
(0,1, 0,5 et 2,0 mg de matière organique particulaire (MOP)⋅L–1) en suspension sur le filtrage, l’ingestion, la respiration, la
production d’excréments et de pseudofèces et les taux d’assimilation chez la moule zébrée (Dreissena polymorpha) adulte en
laboratoire à 20 ± 1°C. L’indice de qualité de nourriture (rapport matières inorganiques–organiques dans les suspensions
d’alimentation = rapport IO) allait de 0,304 à 63,04. Lorsque la concentration de sédiments inorganiques dépassait 1 mg⋅L–1,
le filtrage, l’ingestion et les taux d’assimilation baissaient dans une grande mesure chez la moule zébrée; l’ingestion de MOP
ne se maintenait qu’aux concentrations élevées de particules alimentaires. Les moules recevant une concentration de
sédiments de 100 mg⋅L–1 ont produit des pseudofèces en quantité; il semble que la production de pseudofèces représente un
surcroît pour le métabolisme. La baisse de la qualité de la nourriture s’est traduite par une baisse exponentielle du filtrage, de
l’ingestion, du taux d’absorption, du potentiel de transformation de l’eau et des taux d’assimilation, les baisses les plus
marquées correspondant aux rapports IO compris entre 0,304 et 5,0. Dans les rivières aux eaux troubles, les rapports IO
dépassent souvent 1,71 : dans nos expériences cette valeur était le seuil au-delà duquel les possibilités de croissance de la
moule zébrée étaient nulles. Selon nous, il se pourrait que le potentiel de croissance de la moule zébrée dans les cours d’eau
troubles soit faible et que les populations ne se stabilisent pas aux densités élevées qu’on trouve dans les lacs.
[Traduit par la Rédaction]
Introduction
Suspended sediments can have profound effects on energeticprocesses in filter-feeding bivalves. Bivalves reject excess fil-tered material in pseudofeces above certain threshold sestonconcentrations (Bayne and Newell 1983). Thus, inorganic sus-pended sediments may reduce the amount of food material
ingested via “dilution” of organic particles (Widdows et al.1979; Bricelj and Malouf 1984). Studies with marine bivalvesindicate species-specific responses to suspended sedimentsloads. Species such as Spisula subtruncata and Mytilus eduliswhich control ingestion of nonfood particles by selecting outsuch particles via pseudofeces production (Widdows et al.1979; Kiørboe et al. 1980; Møhlenberg and Kiørboe 1981) are
Can. J. Fish. Aquat. Sci. 55: 401–413 (1998)
Received December 18, 1996. Accepted June 12, 1997.J13806
S.P. Madon,1 J.A. Stoeckel, and R.E. Sparks.Illinois Natural History Survey, River Research Laboratory, Long-Term ResourceMonitoring Program, 704 N. Schrader, Havana, IL 62644, U.S.A.D.W. Schneider.Department of Urban and Regional Planning, University of Illinois at Urbana–Champaign, 611 E. Lorado Taft Drive,Champaign, IL 61820, U.S.A., and Illinois Natural History Survey, Champaign, IL 61820, U.S.A.
1 Author to whom all correspondence should be addressed. Mailing address: Department of Biological Sciences and Graduate Program inEnvironmental Science, Pace University, 861 Bedford Road, Pleasantville, NY 10570, U.S.A.e-mail: [email protected]
401
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:19 1998
Color profile: DisabledComposite Default screen

better adapted to turbid environments than those such as Mer-cenaria mercenaria which reduce clearance rates in responseto high inorganic sediment levels (Bricelj and Malouf 1984).Assimilation efficiencies may be positively or negatively cor-related with suspended sediments, depending on the sedimentconcentration and bivalve species (Kiørboe et al. 1980; Briceljand Malouf 1984; Bayne et al. 1987). Information on theeffects of suspended sediment on bivalve respiration rates in-dicates that bivalves either maintain relatively constant res-piration rates across a range of sediment concentrations(Widdows et al. 1979; Kiørboe et al. 1980; Møhlenberg andKiørboe 1981) or depress metabolic rates as sediment loadincreases (Aldridge et al. 1987; Alexander et al. 1994).
Already well established in the Great Lakes (Hebert et al.1989), the zebra mussel (Dreissena polymorpha) subsequentlycolonized major river systems in the United States such as theSt. Lawrence, Hudson, Illinois, Mississippi, and Ohio rivers(Alexander et al. 1994; Illinois Natural History Survey 1994;Strayer et al. 1996). Zebra mussels pose a major biofoulingproblem for raw-water intake systems and can potentially havesevere ecological impacts on native unionid mussels and otherbenthic and pelagic species (Mackie et al. 1989; Hebert et al.1991; Mackie 1991; MacIsaac et al. 1992; Griffiths 1993;Haag et al. 1993). Because of the severity of economic andecological impacts of zebra mussels on lake ecosystems, moststudies on the biology, physiological energetics, and ecologi-cal impacts of zebra mussel pertain to populations in lenticenvironments (Morton 1969; Walz 1978a, 1978b, 1978c,1978d, 1979; Hamburger et al. 1990; Garton and Haag 1991;Hebert et al. 1991; Schneider 1992; MacIsaac et al. 1992; Buntet al. 1993). Environmental conditions in lentic and lotic sys-tems, however, differ fundamentally, especially with respectto inorganic suspended sediment loads which may affectphysiological processes, and consequently growth rates, of ze-bra mussels. One way to predict growth potential for zebramussels in turbid riverine environments would be to evaluatefunctional responses of their energy intake and loss processesto suspended inorganic sediment.
The scope for growth, or the difference between energygained by an organism and all energy losses, is a useful indexof energy available for somatic growth and production of ga-metes (Bayne and Newell 1983). In large North Americanfloodplain rivers, suspended sediment levels range from 7 to592 mg⋅L–1 (U.S. Geological Survey 1983a, 1983b, 1991) andmay affect processes of energy supply and demand in the zebramussel, ultimately affecting its scope for growth. Alexanderet al. (1994) measured maintenance oxygen consumption inunfed zebra mussels over a range of water temperatures andturbidities and concluded that growth rates of zebra musselswould be depressed under typical ranges of temperature andturbidity in the Ohio River due to increased maintenance me-tabolism. However, the ability of zebra mussels to balanceenergy intake with metabolic costs will ultimately determinetheir growth potential in turbid rivers.
In this study, we examined the effects of inorganic sus-pended sediment and food concentrations on energetic proc-esses in adult zebra mussels. We also assessed the effects offood quality (inorganic to organic ratios in the food–sedimentsuspensions = IO ratios) on zebra mussel energetics and scopefor growth and discuss some implications for growth of theseorganisms in a large turbid rivers.
Materials and methods
We conducted experiments at the Illinois Natural History Survey’sRiver Research Laboratory between October 20, 1993, and February15, 1994. Mussels were collected as required by divers from a site atriver mile 162.3 in the Illinois River, near Peoria, Ill., between Octo-ber 1993 and February 1994. In the laboratory, we sorted mussels ofsimilar sizes (average dry weight ± SE = 20 ± 0.8 mg) and held themussels in filtered Quiver Creek water for at least 2 weeks prior to usein experiments. All mussels were maintained in similar conditions inthe laboratory prior to each experiment. Creek water was filteredthrough a pebble biofilter (2.1 × 0.8 × 0.4 m) several times daily andthen through a Triton TR-60 high-rate sand filter to remove all par-ticulate matter. We monitored the filtered creek water weekly for ni-trates, nitrites, and ammonia. Water temperature was controlled viaroom temperature and was maintained at 20 ± 1°C throughout theexperiments.
Food and sedimentThe food and sediment particle sizes that we selected for our studywere within the range of particle sizes selected by zebra mussels innatural systems (Ten Winkel and Davids 1982; Sprung and Rose1988). We maintained mussels on 4–6 mg⋅mussel–1⋅day–1 (2.7 × 103
to 4.0 × 103 cells⋅mussel–1⋅day–1) of a commercially available liquidmixture of preserved marine diatoms, Thallasiosira and Skeletonema(Diet B, Coast Sea Food Company, Bellevue, Wash.), which has beenused successfully in laboratory studies of zebra mussels elsewhere (A.Stoeckman, Ohio State University, Columbus, Ohio, personal com-munication). This mixture was also used as a food source in ourexperiments, providing uniform cell sizes (3–9 µm in diameter) andconsistent food quality of 76 mg particulate organic matter (POM)⋅g wet weight–1 (542.9 mg POM⋅g dry weight–1).
We collected bottom sediment from the Illinois River and sievedit to yield particles <40 µm in diameter. Sediment was concentratedby settling and decanting and then ashed for 7 h at 500–550°C in amuffle furnace to remove all associated organic matter; ashed sedi-ment was used in all experiments. We did not remove a significantfood source for zebra mussels by using ashed sediment, as initialanalysis revealed that only 6% of the sediment was organic matter.
Experimental apparatusThe experimental apparatus is shown in Fig. 1. A 120-L reservoircontained the stock food–sediment mixture which was maintained insuspension via air bubbles from air stones (A.S.) and two powerheads(P 1 and P 3). An additional powerhead (P 2) supplied the food–sediment mixture to a mixing chamber (M.C.), from where it was ledvia seven separate tubes to 300-mL experimental chambers (E.C.; ablowup of a typical experimental chamber is indicated by the brokenarrow). Flow to the chambers was controlled via flow clamps (F.C.)on the tubes leading into and out of the experimental chambers andwas constantly monitored via glass-bead flowmeters (F.M.). All ex-perimental chambers were placed on an immersible multistirrer plate(S.P.) in a water bath (W.B.) maintained at 20°C. Magnetic stir bars(S.B.) maintained homogenous particulate suspensions within eachchamber. Each experimental chamber had an inner chamber with adetachable 120-µm-mesh nitex sieve (S) on the bottom and threeslotted openings (O, only two are shown in the figure) to facilitatemixing of particulate matter. Zebra mussels were attached to plasticmounting plates (M.P.) and were separated from stir bars by the nitexsieve which allowed food and sediment particles (<40 µm in diame-ter) to pass through, but retained the feces and pseudofeces. Tubesattached to outlet ports below the sieve at the bottom of experimentalchambers led the food–sediment mixture to collecting buckets (C.B.).
ExperimentsExperiments were conducted to assess the effects of sediment andfood concentrations on clearance, ingestion, respiration, feces and
Can. J. Fish. Aquat. Sci. Vol. 55, 1998402
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:20 1998
Color profile: DisabledComposite Default screen

pseudofeces production rates, and assimilation efficiencies of adultzebra mussels. We used a complete factorial design with four levelsof sediment concentration (0, 1, 10, and 100 mg⋅L–1) and three levelsof food concentration (0.1, 0.5, and 2.0 mg POM⋅L–1). IO ratios wereused as an index of food quality and ranged from 0.304 to 63.04. Ourexperimental food and inorganic sediment levels and the correspond-ing IO ratios reflect levels commonly measured in the Illinois River(U.S. Geological Survey 1991; Illinois Natural History Survey, Long-Term Resource Monitoring Program (INHS-LTRMP), Havana, Ill.,unpublished data). We include data from two additional experimentsconducted without sediment at food levels of 1.0 and 5.0 mg POM⋅L–1
in our analysis of the effect of food quality on physiological responsesin zebra mussels.
Prior to each experiment, five batches of four mussels per batchwere glued at the outer edge of one valve with cyanoacrylate adhesiveto plastic platforms and habituated for 1 week to the appropriate ex-perimental mixtures of food and sediment. All mussels secreted bys-sal threads to the platform, indicating good condition. Duringhabituation and all experiments, food and sediment delivery to thechambers was regulated by a flow-through system to prevent reduc-tions of experimental particulate concentrations and to ensure steady-state conditions.
Clearance, ingestion, pseudofeces production rates, assimilationefficiencies, and fecal inorganic content were measured on day 1 un-der flow-through conditions to ensure that particulate levels in thechambers stayed constant during the experiments. For the experiment,each batch of mussels was randomly assigned to an experimentalchamber (five chambers = replicates) and allowed to adjust to condi-tions in the chambers under flows of 0.9 L⋅h–1 for 2 h. In addition, twocontrol chambers had no mussels. After allowing mussels to adjust tothe chambers, we ran experiments for 2–3 h. During this time, waterflowing out of each chamber was directed into collecting buckets. Atthe end of the experiment, flow to the chamber was shut off. A ho-mogenous 1-L subsample of water from each bucket was filtered onpreashed and preweighed 47-mm Whatman GF-C grade glass-fiberfilters. We dried filters at 105°C for 24 h, stored in the desiccator for15 min, weighed, ashed, desiccated again for 15 min, and reweighedto determine total dry mass and POM content. Total dry mass andPOM content of feces and pseudofeces picked individually from eachchamber were similarly determined. Respiration rate experimentswere conducted on day 2.
Clearance ratesClearance rates (litres of water cleared per mussel per hour) of totalsuspended matter (TSM = POM + inorganic sediment) were calcu-lated as
TSMc − TSMe
TSMc
⋅ FR
n
where TSMc and TSMe are the dry mass of total suspended matter inthe control and experimental chambers, respectively, FR is the flowrate (litres per hour) through the chamber, and n represents thenumber of mussels per chamber.
We also calculated clearance rates of POM as
POMc − POMe
POMc
⋅ FR
n
where POMc and POMe are the particulate organic matter contentmeasured in the control and experimental chambers, respectively.
Ingestion ratesIngestion rates (milligrams of POM per mussel per hour) were calcu-lated as
(POMc − POMe) ⋅ FR
n
Ingestion rates were adjusted for pseudofeces production by (1 – p)where p is the proportion of ingested material that was rejected aspseudofeces (Bayne et al. 1989).
Assimilation efficienciesAssimilation efficiencies (%) were based on the Conover (1966)method and were calculated as
RF − RE
(1 − RE)⋅RF⋅ 100
where RF and RE are ratios of organic matter to total dry matter in thefood and feces, respectively.
Absorption ratesAbsorption rates were estimated as the product of ingestion rate andassimilation efficiency and indicate the amount of POM absorbed bythe mussel.
Respiration ratesWe found that oxygen concentrations in experimental chambersdropped by 6–25% in a linear fashion over a 2-h period in initialexperiments in which oxygen consumption was monitored via po-larographic electrodes. Therefore, we used the Winkler method todetermine zebra mussel respiration rates. We allowed mussels to ad-just to the experimental chambers for 2 h under flows of 0.9 L⋅h–1.Flow was then shut off in the experimental chambers and controls.Prior to each experiment, we checked each chamber for the presenceof airspaces that could potentially introduce errors into our dissolvedoxygen measurements: no airspaces were found. Water was mixed inthe chambers with stir bars during the experiment. After 2 h, wesiphoned water from each chamber into 60-mL BOD bottles and
Fig. 1. Schematic diagram of the experimental apparatus. All labels
are explained in the text. Arrows indicate the flow of water
containing food and sediment suspensions through the system. The
apparatus consisted of seven outlets leading the food–sediment
mixture from the mixing chamber (M.C.) to seven separate
experimental chambers (E.C.). Only one such setup is shown here
to simplify illustration.
Madon et al. 403
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:25 1998
Color profile: DisabledComposite Default screen

determined dissolved oxygen in 50-mL samples. We calculated res-piration rates (milligrams of oxygen per individual per hour) by dif-ference between the control (no mussels) and experimental chambersand adjusted for the number of mussels per chamber (Madon andCulver 1993). Significant oxygen consumption in the control cham-bers (without zebra mussels) due to the presence of uneaten foodparticles and associated bacterial activity could potentially introduceerror into our respiration determinations, since food concentrations inthe experimental chamber and controls would differ. We thereforetested for oxygen consumption in the controls due to presence ofuneaten food particles and bacterial activity and found that it wasnegligible over a 2-h period.
Water processing potentialWe estimated the water processing potential as litres of water clearedof POM per millilitre of oxygen respired; this is a useful index of theability of mussels to balance their energy intake against their foodrequirement to meet energetic costs (Jørgensen 1952). Higher valuesof water processing potential indicate a greater ability of the organismto balance energy intake against energy costs (Bayne and Widdows1978). Scaling water processing potential against an environmentalparameter such as the proportion of inorganic to organic ratios insuspension provides insights into the organism’s ability to adapt tochanging food quality conditions. In general, organisms with higherwater processing potential will have lower requirements for minimumfood concentrations in their environment to adequately balance feed-ing gains against metabolic costs (Møhlenberg and Kiørboe 1981). Assuch, water processing potential provides a means of comparing habi-tat requirements of various species, where species with low waterprocessing potential will require an environment relatively rich inorganic particles compared with those with high water processingpotential (Jørgensen 1952; Bayne and Widdows 1978; Møhlenbergand Kiørboe 1981).
Pseudofeces production ratesAt experiment’s end on day 1, all pseudofeces collected on a sieveattached to the internal chamber (Fig. 1) were individually pickedfrom each chamber and dry mass and organic content were deter-mined. Mean mass-specific pseudofeces production rates (micro-grams of organic matter per milligram of tissue dry mass per hour)were calculated using mean dry mass of individuals from each cham-ber.
Fecal inorganic contentAll feces were individually picked from the sieves in each chamberand total dry mass and organic content of feces were determined.Mean mass-specific fecal inorganic content (micrograms of inor-ganics per milligram of tissue dry mass per hour) was calculated foreach chamber by subtracting organic mass from total dry mass andadjusting the result by the mean dry tissue weight of mussels fromeach chamber.
Routine metabolic rateThe gills of suspension-feeding bivalves are used for respiration aswell as for feeding. Therefore, respiration in bivalves is dependent onration (Bayne and Newell 1983). In the absence of food particles,pumping rates are low, and respiration rates represent the routinemetabolic rate. Respiration rates typically increase with increasingfood concentration and represent active respiration rates (Bayne andNewell 1983).
We measured respiration rates of zebra mussels at food levels of0.1, 0.5, 1.0, 2.0, and 5.0 mg POM⋅L–1. We converted these respira-tion rate measurements from milligrams of oxygen to calories byusing an oxycalorific value of 3.24 cal⋅mg O2
−1 (1 cal = 4.1868 J)(Elliot and Davidson 1975). A functional response of respiration ratesto food levels was described by a quadratic fit to the data, and routinemetabolic rate was calculated by extrapolation to zero ration. Because
extrapolation outside the domain of the data may compromise accu-racy, we also measured respiration rates at zero ration.
Scope for growthScope for growth is the amount of absorbed energy potentially avail-able for growth after accounting for metabolic and excretory losses.We estimated scope for growth (calories per hour) after convertingingestion rates and respiration rates (MR) to their respective caloricequivalents. Caloric density of POM (diatoms) was assumed to be5.31 cal⋅g dry mass–1 (Cummins and Wuycheck 1971). An oxycalo-rific value of 3.24 cal⋅mg O2
−1 was also assumed (Elliot and Davidson1975). We estimated scope for growth as
AR – MR – (0.064⋅AR)
where AR is the absorbed ration estimated as the product of ingestionrate and assimilation efficiency (proportion) and 0.064 represents theassumed proportion of absorbed ration that is lost via excretion(Schneider 1992).
Maintenance costsWe estimated maintenance costs of zebra mussels as
RMR/AR
where RMR is the routine metabolic rate. Maintenance costs reflectthe fraction of absorbed ration required for routine metabolism.
Standardization of ratesSimilar sized mussels of 20 ± 0.8 mg (±SE) average dry mass wereselected for our experiments. Because of the low variance in bodymass, we did not consider mussel mass to be an important covariateof physiological rates in our study. However, we standardized physi-ological rates to account for those variations in body mass that didoccur.
We standardized clearance, ingestion, and respiration rates to rep-resent rates in a mussel of 20 mg dry tissue weight, which was themean weight of all experimental mussels. We calculated the stand-ardized physiological rates (Ys) as suggested in Bayne et al. (1977):
Ys = (20/W)b⋅Ye
where b represents the slope of the allometric relationship and hasvalues of 0.61 for clearance and ingestion rates and 0.75 for respira-tion rates (Schneider 1992), W represents the measured mean drytissue weight of mussels from each chamber, and Ye is the measuredphysiological rate.
Statistical analysisWe conducted two separate sets of analyses (i) to determine the ef-fects of food and inorganic sediment on physiological responses ofzebra mussels and (ii) to evaluate physiological functional responsesof zebra mussels to food quality. In the first set of analyses, we useda two-way ANOVA with food and sediment as treatment categoriesto assess interactions of response variables between treatment catego-ries and to determine main effects of treatments on response variables.We also analyzed effects on response variables of food treatmentswithin each sediment treatment, and sediment treatments within eachfood treatment separately, via Tukey’s HSD multiple comparisons oftreatment effects generated with one-way ANOVA. All response vari-ables (x) were log10(x + 1) or arcsin(x)0.5 transformed to correct fornonnormality and nonhomogeneous variances. For the second set ofanalyses, we described functional responses of physiological vari-ables to food quality (IO ratios). High concentrations of inorganicsediment dilute the organic particles in suspension and potentiallyreduce the amount of food available to filter-feeders. Therefore, in-creasing IO ratios in suspension serve as an index of food quality,where food quality is superior at low IO ratios and inferior at high IOratios. We generated functional responses by fitting general linearmodels (Wilkinson 1990) to log-transformed or log10-transformedphysiological variables.
Can. J. Fish. Aquat. Sci. Vol. 55, 1998404
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:27 1998
Color profile: DisabledComposite Default screen
Results
Effects of food and sediment on zebra mussel energetics
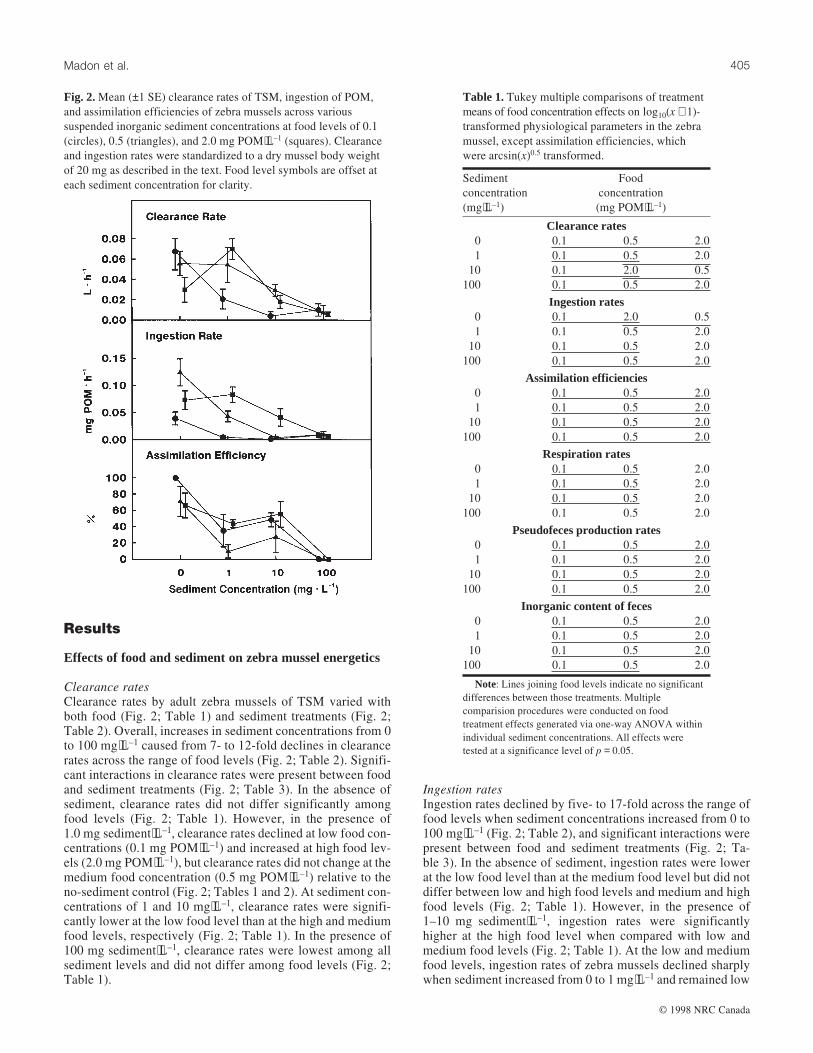
Clearance ratesClearance rates by adult zebra mussels of TSM varied withboth food (Fig. 2; Table 1) and sediment treatments (Fig. 2;Table 2). Overall, increases in sediment concentrations from 0to 100 mg⋅L–1 caused from 7- to 12-fold declines in clearancerates across the range of food levels (Fig. 2; Table 2). Signifi-cant interactions in clearance rates were present between foodand sediment treatments (Fig. 2; Table 3). In the absence ofsediment, clearance rates did not differ significantly amongfood levels (Fig. 2; Table 1). However, in the presence of1.0 mg sediment⋅L–1, clearance rates declined at low food con-centrations (0.1 mg POM⋅L–1) and increased at high food lev-els (2.0 mg POM⋅L–1), but clearance rates did not change at themedium food concentration (0.5 mg POM⋅L–1) relative to theno-sediment control (Fig. 2; Tables 1 and 2). At sediment con-centrations of 1 and 10 mg⋅L–1, clearance rates were signifi-cantly lower at the low food level than at the high and mediumfood levels, respectively (Fig. 2; Table 1). In the presence of100 mg sediment⋅L–1, clearance rates were lowest among allsediment levels and did not differ among food levels (Fig. 2;Table 1).
Ingestion ratesIngestion rates declined by five- to 17-fold across the range offood levels when sediment concentrations increased from 0 to100 mg⋅L–1 (Fig. 2; Table 2), and significant interactions werepresent between food and sediment treatments (Fig. 2; Ta-ble 3). In the absence of sediment, ingestion rates were lowerat the low food level than at the medium food level but did notdiffer between low and high food levels and medium and highfood levels (Fig. 2; Table 1). However, in the presence of1–10 mg sediment⋅L–1, ingestion rates were significantlyhigher at the high food level when compared with low andmedium food levels (Fig. 2; Table 1). At the low and mediumfood levels, ingestion rates of zebra mussels declined sharplywhen sediment increased from 0 to 1 mg⋅L–1 and remained low
Sediment
concentration
(mg⋅L–1)
Food
concentration
(mg POM⋅L–1)
Clearance rates0 0.1 0.5 2.0
1 0.1 0.5 2.0
10 0.1 2.0 0.5
100 0.1 0.5 2.0
Ingestion rates0 0.1 2.0 0.5
1 0.1 0.5 2.0
10 0.1 0.5 2.0
100 0.1 0.5 2.0
Assimilation efficiencies0 0.1 0.5 2.0
1 0.1 0.5 2.0
10 0.1 0.5 2.0
100 0.1 0.5 2.0
Respiration rates0 0.1 0.5 2.0
1 0.1 0.5 2.0
10 0.1 0.5 2.0
100 0.1 0.5 2.0
Pseudofeces production rates0 0.1 0.5 2.0
1 0.1 0.5 2.0
10 0.1 0.5 2.0
100 0.1 0.5 2.0
Inorganic content of feces0 0.1 0.5 2.0
1 0.1 0.5 2.0
10 0.1 0.5 2.0
100 0.1 0.5 2.0
Note: Lines joining food levels indicate no significant
differences between those treatments. Multiple
comparision procedures were conducted on food
treatment effects generated via one-way ANOVA within
individual sediment concentrations. All effects were
tested at a significance level of p = 0.05.
Table 1.Tukey multiple comparisons of treatment
means of food concentration effects on log10(x + 1)-
transformed physiological parameters in the zebra
mussel, except assimilation efficiencies, which
were arcsin(x)0.5 transformed.
Fig. 2. Mean (±1 SE) clearance rates of TSM, ingestion of POM,
and assimilation efficiencies of zebra mussels across various
suspended inorganic sediment concentrations at food levels of 0.1
(circles), 0.5 (triangles), and 2.0 mg POM⋅L–1 (squares). Clearance
and ingestion rates were standardized to a dry mussel body weight
of 20 mg as described in the text. Food level symbols are offset at
each sediment concentration for clarity.
Madon et al. 405
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:32 1998
Color profile: DisabledComposite Default screen
and did not differ significantly when sediment load was in-creased from 1 to 100 mg⋅L–1 (Fig. 2; Table 2). Conversely, atthe high food level, ingestion rates stayed relatively constant atsediment concentrations of 0–10 mg⋅L–1; a significant declinein ingestion rates occurred as sediment levels were increasedto 100 mg⋅L–1 (Fig. 2; Table 2).
Assimilation efficienciesAssimilation efficiencies in the zebra mussel did not varyamong food concentrations, and no significant interactionswere present between food and sediment treatments (Fig. 2;Tables 1 and 3). However, across the range of food levels,assimilation efficiencies declined from mean levels of 66–100% in the absence of sediment to 0% at 100 mg sediment⋅L–1 (Fig. 2). At the low food level, the decline in assimilationefficiency with increased sediment was characterized by asharp decrease between 0 and 1 mg sediment⋅L–1, followedby constant assimilation efficiencies between 1 and 10 mgsediment⋅L–1, and a decline again from 10 to 100 mg sediment⋅L–1 (Fig. 2; Table 2). This same pattern of decline was evident
at the medium food level, except that no significant differencein assimilation efficiencies was found between 10 and 100 mgsediment⋅L–1 (Fig. 2; Table 2). However, at the high foodlevel, assimilation efficiencies stayed relatively constant be-tween 0 and 10 mg sediment⋅L–1 and declined when sedimentincreased from 10 to 100 mg⋅L–1 (Fig. 2; Table 2).
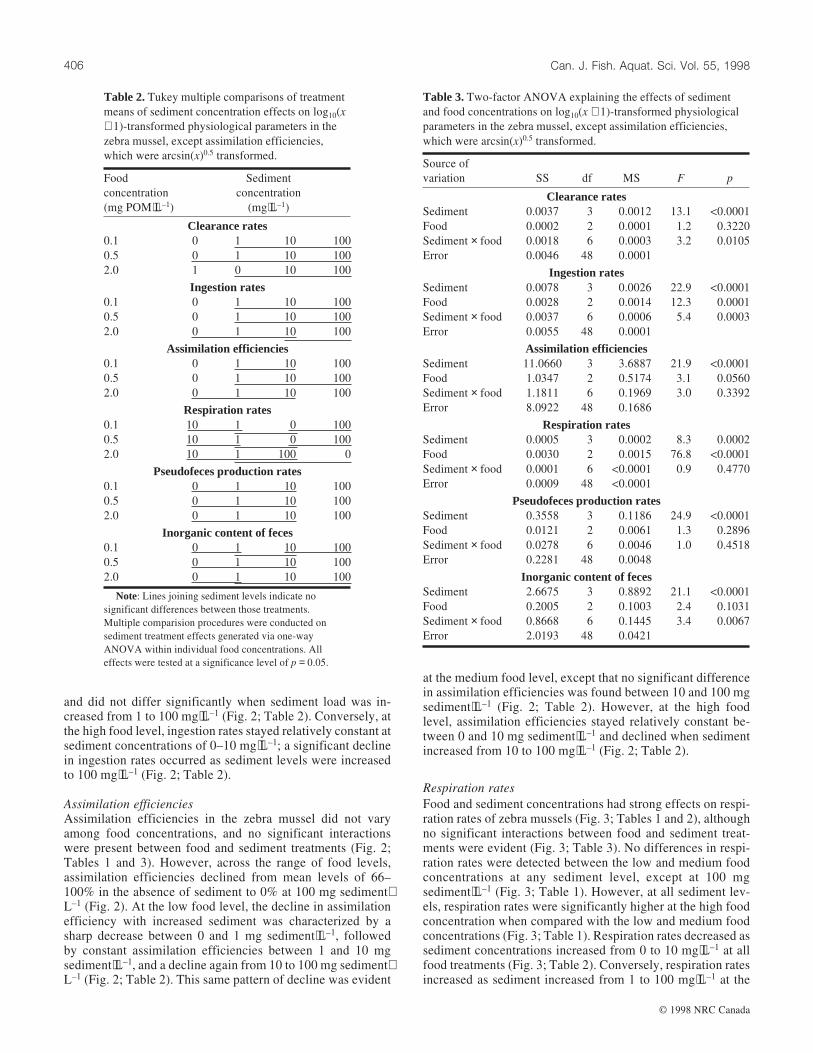
Respiration rates
Food and sediment concentrations had strong effects on respi-ration rates of zebra mussels (Fig. 3; Tables 1 and 2), althoughno significant interactions between food and sediment treat-ments were evident (Fig. 3; Table 3). No differences in respi-ration rates were detected between the low and medium foodconcentrations at any sediment level, except at 100 mgsediment⋅L–1 (Fig. 3; Table 1). However, at all sediment lev-els, respiration rates were significantly higher at the high foodconcentration when compared with the low and medium foodconcentrations (Fig. 3; Table 1). Respiration rates decreased assediment concentrations increased from 0 to 10 mg⋅L–1 at allfood treatments (Fig. 3; Table 2). Conversely, respiration ratesincreased as sediment increased from 1 to 100 mg⋅L–1 at the
Food
concentration
(mg POM⋅L–1)
Sediment
concentration
(mg⋅L–1)
Clearance rates0.1 0 1 10 100
0.5 0 1 10 100
2.0 1 0 10 100
Ingestion rates0.1 0 1 10 100
0.5 0 1 10 100
2.0 0 1 10 100
Assimilation efficiencies0.1 0 1 10 100
0.5 0 1 10 100
2.0 0 1 10 100
Respiration rates0.1 10 1 0 100
0.5 10 1 0 100
2.0 10 1 100 0
Pseudofeces production rates0.1 0 1 10 100
0.5 0 1 10 100
2.0 0 1 10 100
Inorganic content of feces0.1 0 1 10 100
0.5 0 1 10 100
2.0 0 1 10 100
Note: Lines joining sediment levels indicate no
significant differences between those treatments.
Multiple comparision procedures were conducted on
sediment treatment effects generated via one-way
ANOVA within individual food concentrations. All
effects were tested at a significance level of p = 0.05.
Table 2.Tukey multiple comparisons of treatment
means of sediment concentration effects on log10(x
+ 1)-transformed physiological parameters in the
zebra mussel, except assimilation efficiencies,
which were arcsin(x)0.5 transformed.Source of
variation SS df MS F p
Clearance ratesSediment 0.0037 3 0.0012 13.1 <0.0001
Food 0.0002 2 0.0001 1.2 0.3220
Sediment × food 0.0018 6 0.0003 3.2 0.0105
Error 0.0046 48 0.0001
Ingestion ratesSediment 0.0078 3 0.0026 22.9 <0.0001
Food 0.0028 2 0.0014 12.3 0.0001
Sediment × food 0.0037 6 0.0006 5.4 0.0003
Error 0.0055 48 0.0001
Assimilation efficienciesSediment 11.0660 3 3.6887 21.9 <0.0001
Food 1.0347 2 0.5174 3.1 0.0560
Sediment × food 1.1811 6 0.1969 3.0 0.3392
Error 8.0922 48 0.1686
Respiration ratesSediment 0.0005 3 0.0002 8.3 0.0002
Food 0.0030 2 0.0015 76.8 <0.0001
Sediment × food 0.0001 6 <0.0001 0.9 0.4770
Error 0.0009 48 <0.0001
Pseudofeces production ratesSediment 0.3558 3 0.1186 24.9 <0.0001
Food 0.0121 2 0.0061 1.3 0.2896
Sediment × food 0.0278 6 0.0046 1.0 0.4518
Error 0.2281 48 0.0048
Inorganic content of fecesSediment 2.6675 3 0.8892 21.1 <0.0001
Food 0.2005 2 0.1003 2.4 0.1031
Sediment × food 0.8668 6 0.1445 3.4 0.0067
Error 2.0193 48 0.0421
Table 3.Two-factor ANOVA explaining the effects of sediment
and food concentrations on log10(x + 1)-transformed physiological
parameters in the zebra mussel, except assimilation efficiencies,
which were arcsin(x)0.5 transformed.
Can. J. Fish. Aquat. Sci. Vol. 55, 1998406
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:33 1998
Color profile: DisabledComposite Default screen

low and medium food levels, but did not change significantlyat the high food level (Fig. 3; Table 2).
Pseudofeces production rates
Pseudofeces production rates were strongly affected by sedi-ment but not by food concentrations (Fig. 3; Tables 1 and 2),and no interactions were present between food and sedimenttreatments (Fig. 3; Table 3). Across the range of food treat-ments, pseudofeces production rates remained relatively con-stant as sediment increased from 0 to 10 mg⋅L–1, but increasedsharply as sediment increased from 10 to 100 mg⋅L–1 (Fig. 3;Table 2).
Fecal inorganic content
Inorganic content of feces was strongly affected by sedimentbut weakly by food concentration (Fig. 3; Tables 1 and 2),although there were significant interactions between food andsediment treatments (Fig. 3; Table 3). Across each sedimenttreatment, fecal inorganic content did not differ among foodlevels, except at 100 mg sediment⋅L–1 where fecal inorganiccontent was higher at the low and medium food levels than atthe high food level (Fig. 3; Table 1). At all food levels, therewas a general increase in fecal inorganic content as sedimentincreased from 0 to 100 mg⋅L–1 (Fig. 3; Table 2).
Effects of food quality on energetics
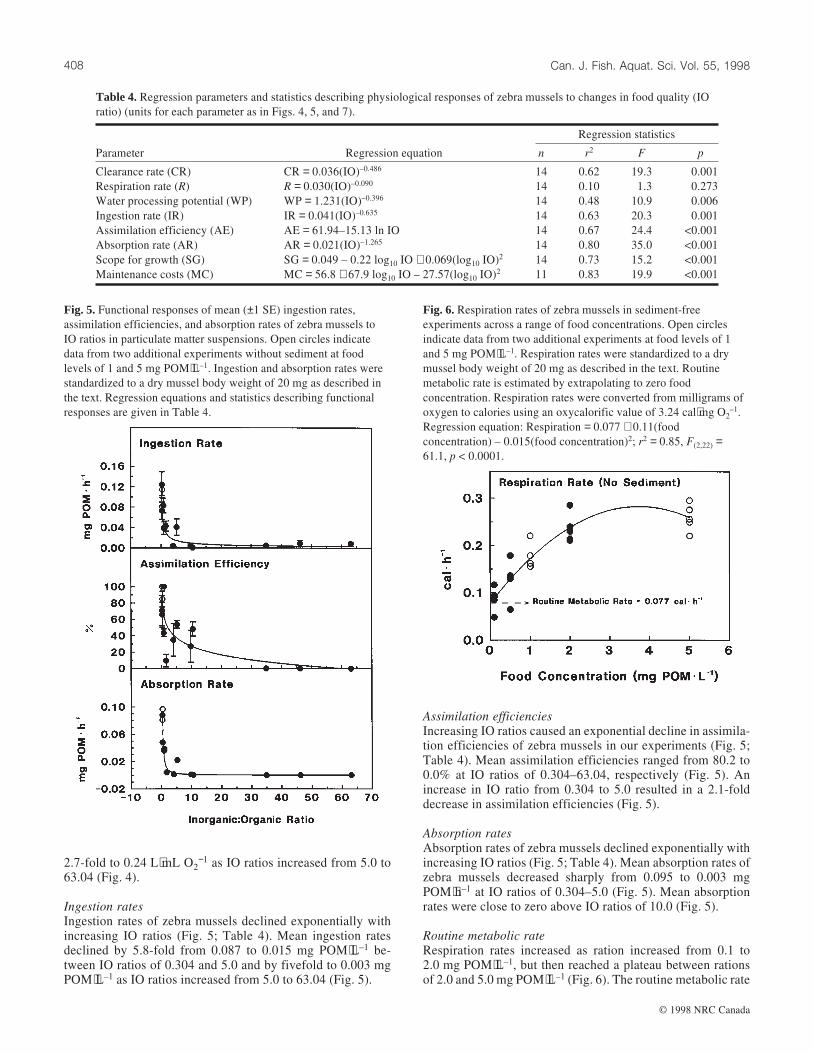
Clearance rates
Clearance rates of zebra mussels declined exponentially as IOratios increased (Fig. 4; Table 4). Mean clearance rates ofPOM showed a fourfold decrease from 0.064 to 0.016 L⋅h–1 asIO ratios increased from 0.304 to 5.0. Mean clearance ratesdeclined 3.3-fold to 0.0048 L⋅h–1 as IO ratios increased from5.0 to 63.04.
Respiration rates
Respiration rates of zebra mussels did not appear to changesignificantly with increasing IO ratios (Fig. 4; Table 4). Thismay have been an artifact of the high variation in respirationrates (Fig. 4) due to strong food level effects (Fig. 3; Table 1)and elevated respiration rates due to pseudofeces formation athigh sediment loads (Fig. 3; Table 2).
Water processing potential
Increasing IO ratios caused an exponential decline in waterprocessing potential of zebra mussels (Fig. 4; Table 4). Meanwater processing potential declined threefold from 1.97 to0.65 L⋅mL O2
−1 between IO ratios of 0.304 and 5.0 and
Fig. 3. Mean (±1 SE) respiration rates, weight-specific pseudofeces
production rates, and fecal inorganic content of zebra mussels
across various suspended inorganic sediment concentrations at food
levels of 0.1 (circles), 0.5 (triangles), and 2.0 mg POM⋅L–1
(squares). Respiration rates were standardized to a dry mussel body
weight of 20 mg as described in the text. Food level symbols are
offset at each sediment concentration for clarity.
Fig. 4. Functional responses of mean (±1 SE) clearance of POM,
respiration rates, and water processing potential of zebra mussels to
IO ratios in particulate matter suspensions. Open circles indicate
data from two additional experiments without sediment at food
levels of 1 and 5 mg POM⋅L–1. Clearance and respiration rates were
standardized to a dry mussel body weight of 20 mg as described in
the text. Regression equations and statistics describing functional
responses are given in Table 4.
Madon et al. 407
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:44 1998
Color profile: DisabledComposite Default screen
2.7-fold to 0.24 L⋅mL O2−1 as IO ratios increased from 5.0 to
63.04 (Fig. 4).
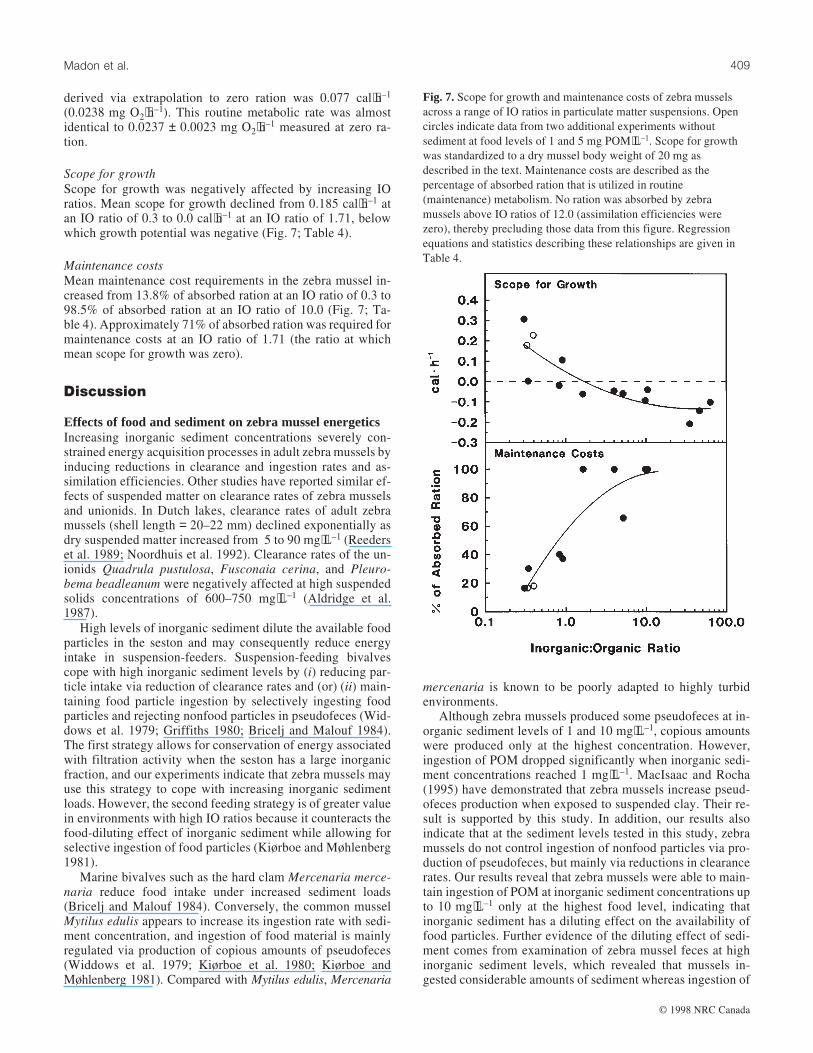
Ingestion ratesIngestion rates of zebra mussels declined exponentially withincreasing IO ratios (Fig. 5; Table 4). Mean ingestion ratesdeclined by 5.8-fold from 0.087 to 0.015 mg POM⋅L–1 be-tween IO ratios of 0.304 and 5.0 and by fivefold to 0.003 mgPOM⋅L–1 as IO ratios increased from 5.0 to 63.04 (Fig. 5).
Assimilation efficienciesIncreasing IO ratios caused an exponential decline in assimila-tion efficiencies of zebra mussels in our experiments (Fig. 5;Table 4). Mean assimilation efficiencies ranged from 80.2 to0.0% at IO ratios of 0.304–63.04, respectively (Fig. 5). Anincrease in IO ratio from 0.304 to 5.0 resulted in a 2.1-folddecrease in assimilation efficiencies (Fig. 5).
Absorption ratesAbsorption rates of zebra mussels declined exponentially withincreasing IO ratios (Fig. 5; Table 4). Mean absorption rates ofzebra mussels decreased sharply from 0.095 to 0.003 mgPOM⋅h–1 at IO ratios of 0.304–5.0 (Fig. 5). Mean absorptionrates were close to zero above IO ratios of 10.0 (Fig. 5).
Routine metabolic rateRespiration rates increased as ration increased from 0.1 to2.0 mg POM⋅L–1, but then reached a plateau between rationsof 2.0 and 5.0 mg POM⋅L–1 (Fig. 6). The routine metabolic rate
Regression statistics
Parameter Regression equation n r2 F p
Clearance rate (CR) CR = 0.036(IO)–0.486 14 0.62 19.3 0.001
Respiration rate (R) R = 0.030(IO)–0.090 14 0.10 1.3 0.273
Water processing potential (WP) WP = 1.231(IO)–0.396 14 0.48 10.9 0.006
Ingestion rate (IR) IR = 0.041(IO)–0.635 14 0.63 20.3 0.001
Assimilation efficiency (AE) AE = 61.94–15.13 ln IO 14 0.67 24.4 <0.001
Absorption rate (AR) AR = 0.021(IO)–1.265 14 0.80 35.0 <0.001
Scope for growth (SG) SG = 0.049 – 0.22 log10 IO + 0.069(log10 IO)2 14 0.73 15.2 <0.001
Maintenance costs (MC) MC = 56.8 + 67.9 log10 IO – 27.57(log10 IO)2 11 0.83 19.9 <0.001
Table 4.Regression parameters and statistics describing physiological responses of zebra mussels to changes in food quality (IO
ratio) (units for each parameter as in Figs. 4, 5, and 7).
Fig. 5. Functional responses of mean (±1 SE) ingestion rates,
assimilation efficiencies, and absorption rates of zebra mussels to
IO ratios in particulate matter suspensions. Open circles indicate
data from two additional experiments without sediment at food
levels of 1 and 5 mg POM⋅L–1. Ingestion and absorption rates were
standardized to a dry mussel body weight of 20 mg as described in
the text. Regression equations and statistics describing functional
responses are given in Table 4.
Fig. 6. Respiration rates of zebra mussels in sediment-free
experiments across a range of food concentrations. Open circles
indicate data from two additional experiments at food levels of 1
and 5 mg POM⋅L–1. Respiration rates were standardized to a dry
mussel body weight of 20 mg as described in the text. Routine
metabolic rate is estimated by extrapolating to zero food
concentration. Respiration rates were converted from milligrams of
oxygen to calories using an oxycalorific value of 3.24 cal⋅mg O2–1.
Regression equation: Respiration = 0.077 + 0.11(food
concentration) – 0.015(food concentration)2; r2 = 0.85, F(2,22) =61.1, p < 0.0001.
Can. J. Fish. Aquat. Sci. Vol. 55, 1998408
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:53 1998
Color profile: DisabledComposite Default screen
derived via extrapolation to zero ration was 0.077 cal⋅h–1
(0.0238 mg O2⋅h–1). This routine metabolic rate was almostidentical to 0.0237 ± 0.0023 mg O2⋅h–1 measured at zero ra-tion.
Scope for growth
Scope for growth was negatively affected by increasing IOratios. Mean scope for growth declined from 0.185 cal⋅h–1 atan IO ratio of 0.3 to 0.0 cal⋅h–1 at an IO ratio of 1.71, belowwhich growth potential was negative (Fig. 7; Table 4).
Maintenance costs
Mean maintenance cost requirements in the zebra mussel in-creased from 13.8% of absorbed ration at an IO ratio of 0.3 to98.5% of absorbed ration at an IO ratio of 10.0 (Fig. 7; Ta-ble 4). Approximately 71% of absorbed ration was required formaintenance costs at an IO ratio of 1.71 (the ratio at whichmean scope for growth was zero).
Discussion
Effects of food and sediment on zebra mussel energeticsIncreasing inorganic sediment concentrations severely con-strained energy acquisition processes in adult zebra mussels byinducing reductions in clearance and ingestion rates and as-similation efficiencies. Other studies have reported similar ef-fects of suspended matter on clearance rates of zebra musselsand unionids. In Dutch lakes, clearance rates of adult zebramussels (shell length = 20–22 mm) declined exponentially asdry suspended matter increased from 5 to 90 mg⋅L–1 (Reederset al. 1989; Noordhuis et al. 1992). Clearance rates of the un-ionids Quadrula pustulosa, Fusconaia cerina, and Pleuro-bema beadleanum were negatively affected at high suspendedsolids concentrations of 600–750 mg⋅L–1 (Aldridge et al.1987).
High levels of inorganic sediment dilute the available foodparticles in the seston and may consequently reduce energyintake in suspension-feeders. Suspension-feeding bivalvescope with high inorganic sediment levels by (i) reducing par-ticle intake via reduction of clearance rates and (or) (ii) main-taining food particle ingestion by selectively ingesting foodparticles and rejecting nonfood particles in pseudofeces (Wid-dows et al. 1979; Griffiths 1980; Bricelj and Malouf 1984).The first strategy allows for conservation of energy associatedwith filtration activity when the seston has a large inorganicfraction, and our experiments indicate that zebra mussels mayuse this strategy to cope with increasing inorganic sedimentloads. However, the second feeding strategy is of greater valuein environments with high IO ratios because it counteracts thefood-diluting effect of inorganic sediment while allowing forselective ingestion of food particles (Kiørboe and Møhlenberg1981).
Marine bivalves such as the hard clam Mercenaria merce-
naria reduce food intake under increased sediment loads(Bricelj and Malouf 1984). Conversely, the common musselMytilus edulis appears to increase its ingestion rate with sedi-ment concentration, and ingestion of food material is mainlyregulated via production of copious amounts of pseudofeces(Widdows et al. 1979; Kiørboe et al. 1980; Kiørboe andMøhlenberg 1981). Compared with Mytilus edulis, Mercenaria
mercenaria is known to be poorly adapted to highly turbidenvironments.
Although zebra mussels produced some pseudofeces at in-organic sediment levels of 1 and 10 mg⋅L–1, copious amountswere produced only at the highest concentration. However,ingestion of POM dropped significantly when inorganic sedi-ment concentrations reached 1 mg⋅L–1. MacIsaac and Rocha(1995) have demonstrated that zebra mussels increase pseud-ofeces production when exposed to suspended clay. Their re-sult is supported by this study. In addition, our results alsoindicate that at the sediment levels tested in this study, zebramussels do not control ingestion of nonfood particles via pro-duction of pseudofeces, but mainly via reductions in clearancerates. Our results reveal that zebra mussels were able to main-tain ingestion of POM at inorganic sediment concentrations upto 10 mg⋅L–1 only at the highest food level, indicating thatinorganic sediment has a diluting effect on the availability offood particles. Further evidence of the diluting effect of sedi-ment comes from examination of zebra mussel feces at highinorganic sediment levels, which revealed that mussels in-gested considerable amounts of sediment whereas ingestion of
Fig. 7. Scope for growth and maintenance costs of zebra mussels
across a range of IO ratios in particulate matter suspensions. Open
circles indicate data from two additional experiments without
sediment at food levels of 1 and 5 mg POM⋅L–1. Scope for growth
was standardized to a dry mussel body weight of 20 mg as
described in the text. Maintenance costs are described as the
percentage of absorbed ration that is utilized in routine
(maintenance) metabolism. No ration was absorbed by zebra
mussels above IO ratios of 12.0 (assimilation efficiencies were
zero), thereby precluding those data from this figure. Regression
equations and statistics describing these relationships are given in
Table 4.
Madon et al. 409
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:57:58 1998
Color profile: DisabledComposite Default screen

POM was minimal. In natural systems, zebra mussels may beable to derive nutrition from organic matter adsorbed on inor-ganic sediment particles. However, nutrition derived in thismanner is likely to be poor, considering that the organic frac-tion constituted only 6% of suspended sediment.
Inorganic suspended sediment greatly reduced the assimi-lation efficiencies of POM in zebra mussels, regardless of foodlevels. Ingestion of inorganic particles with increasing inor-ganic sediment concentrations could reduce gut residence timeof food particles, resulting in low assimilation efficiencies.Negative effects of suspended sediment on assimilation effi-ciencies of other bivalve species are demonstrated for Merce-naria mercenaria (Bricelj and Malouf 1984) and the Icelandscallop (Clamys iceladica) (Vahl 1980). Conversely, assimila-tion efficiencies in Mytilus edulis declined only slightly from65 to 52% as sediment concentrations increased from 0 to55 mg⋅L–1 (Kiørboe et al. 1980). Some bivalves such asSpisula subtruncata exhibit increased assimilation efficienciesin the presence of suspended sediment, which probably alsoserves as a source of supplemental organic material (Møhlen-berg and Kiørboe 1981). All organic matter was removed fromsediment used in our experiments, precluding it as a minorsource of organic material for zebra mussels in this study.However, subsequent feeding experiments on zebra musselswith natural river seston revealed that assimilation efficienciesrapidly declined from 80 to 10% as particulate inorganic mat-ter (PIM) increased from 0 to 10 mg⋅L–1 and remained at amean level of 5% between PIM levels of 12 and 55 mg⋅L–1
(D.W. Schneider, S.P. Madon, J.A. Stoeckel, and R.E. Sparks,unpublished data). These results are similar to those in thisstudy and reveal that adult zebra mussels in natural systemsmay be severely constrained in absorbing ingested organicmatter when PIM concentrations are above 10 mg⋅L–1.
Both food and inorganic sediment affected respiration ratesof zebra mussels in our experiments. Across all food levels,respiration rates declined as inorganic sediment concentrationsincreased from 0 to 10 mg⋅L–1. The gills are sites of both feed-ing and metabolism in filter-feeding bivalves, and respirationrates typically increase with filtration activity and ration(Bayne and Newell 1983). Clearance and ingestion rates ofzebra mussels in our experiments declined as inorganic sedi-ment levels increased to 10 mg⋅L–1, and consequently, respi-ration rates also decreased. Respiration rates of zebra musselsin our low and medium food experiments, however, increasedsignificantly as sediment levels increased from 10 to100 mg⋅L–1. High respiration rates observed at the 100 mg⋅L–1
sediment level cannot be adequately explained by clearancerates, which were very low. High levels of suspended sedimentcan dramatically reduce filtration efficiency of bivalves byclogging their gills (Bayne and Newell 1983). To counteractthis effect, bivalves reject excess particles in mucus secretionsfrom the gills (Foster-Smith 1975). These secretions trap ex-cess particles that, as in the case of zebra mussels, are thenrejected as pseudofeces via the inhalant siphon (Morton 1969).In our experiments, increases in respiration rates of zebra mus-sels coincide with, and may reflect costs associated with, in-creased pseudofeces production at the 100 mg⋅L–1 sedimentlevel. Respiration rates of zebra mussels at the 100 mg⋅L–1
sediment level did not increase at high food concentrations;mussels in this experiment also produced lower amounts ofpseudofeces than in the other two food treatments. Respiration
rates of bivalves also increase soon after ingestion of foodparticles due to specific dynamic action (SDA) costs associ-ated with processing and digestion of food material (Bayneet al. 1976; 1989). Below sediment levels of 100 mg⋅L–1, res-piration rates of zebra mussels were consistently higher at thehighest food level, and correlated with higher ingestion of foodmaterial, than at the low and medium food levels, thereby in-dicating metabolic costs due to the SDA component.
Effects of food quality on zebra mussel energeticsDeclining food quality had a negative effect on all functionalresponses, except respiration rates, of the zebra mussels testedin our experiments. Clearance of POM, ingestion, absorptionrates, and assimilation efficiencies of the zebra mussel all de-clined rapidly as food quality declined (IO ratios increased).The most rapid changes in functional responses occurred be-tween IO ratios of 0.304 and 5.0, indicating that growth ratesof zebra mussels would be adversely affected by food qualitychanges within this IO range.
In our experiments, the sharp decline in the water process-ing potential of zebra mussels with increasing IO ratios indi-cates that as food quality declines, zebra mussels have poorability to balance feeding gains against their energy costs.Water processing potential of zebra mussels never exceeded5.07 L⋅mL O2
−1 in our experiments and was consistently lowerthan 0.65 L⋅mL O2
−1 above IO ratios of 5.0. Conversely, waterprocessing potential values as high as 10 and 15 L⋅mL O2
−1
have been reported for Spisula subtruncata and Mytilus edulis,which typically inhabit turbid estuarine and marine environ-ments (Møhlenberg and Kiørboe 1981). We contend that zebramussels may be poorly adapted to turbid environments withhigh inorganic particle concentrations and generally require anenvironment relatively rich in organic particles.
The mean proportion of absorbed ration required to satisfycosts of routine metabolism (maintenance costs) in the zebramussel increased rapidly as IO ratios increased and was closeto 100% above IO ratios of 10.0. Mean scope for growth de-clined rapidly with increasing IO ratios and was negative at IOratios of 10.0. Scope for growth in the zebra mussel was zeroat IO ratios of 1.71, when 71% of the absorbed ration wasrequired for maintenance costs. The remaining proportion ofabsorbed ration was probably required for costs associatedwith feeding, digestion, and processing of food materials, leav-ing no surplus energy for growth. Our study reveals that withdeclining food quality, zebra mussels must expend increasingproportions of absorbed ration to meet maintenance costs, andconsequently have little or no energy available for growth orreproduction.
Only adult zebra mussels (20–25 mm) were used in ourstudy. Due to the allometry of energetic processes (Schneider1992), smaller zebra mussels (<20 mm) may show differentresponses to IO ratios than larger individuals. However, evi-dence from recent experiments on small zebra mussels fednatural river seston indicates that energy intake in 5- to 10-mmmussels decreased, while energy costs stayed relatively con-stant with increasing IO ratios (D.W. Schneider, S.P. Madon,J.A. Stoeckel, and R.E. Sparks, unpublished data). This patternis similar to that exhibited by adult mussels in this study.
Two additional factors that may affect energetic processesand growth of zebra mussels, but that were not considered inthis study, are temperature and organic matter adsorbed on
Can. J. Fish. Aquat. Sci. Vol. 55, 1998410
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:58:00 1998
Color profile: DisabledComposite Default screen

inorganic sediment particles. Our experiments were conductedat 20°C. Annual water temperatures in the Illinois River aretypically 20°C or above from May to September and below20°C from October to April. During the spring, lower tempera-tures may result in a more favorable scope for growth thanwould be predicted by this study. However, during this time,excess energy may be channeled into gamete production forthe coming spawn, at the expense of growth and (or) energystorage. During summer, when water temperatures are typi-cally above 20°C, increased maintenance metabolic costs maylead to lower growth rates (Walz 1978a; Alexander et al.1994). Our study indicates that the negative effects of hightemperature would be amplified by the high IO ratios typicalof the Illinois River during the summer months. If they survivethe summer, zebra mussels are likely to be in poor conditionin the fall, when they need to quickly replenish their stores tosurvive the winter. High IO ratios at tempreatures of 20°C orabove are likely to have a strong, negative impact on zebramussels in an environment such as the Illinois River.
Zebra mussels in our experiments ingested inorganic sedi-ment particles and may have been able to derive nutrition fromassociated organic material, if the sediment used had not beenpreashed. However, the fraction of organic material associatedwith inorganic particles in our river water was very small (6%)and would not have contributed substantially to the nutritionof zebra mussels. The nutritional value of sediment-associatedsubstrates in natural systems is typically low compared withthat of algal material (Arruda et al. 1983). Our feeding experi-ments with natural river seston revealed that although inges-tion rates were higher than those in this study for similar levelsof suspended matter, assimilation efficiencies rapidly declinedas natural seston loads increased, such that zebra mussels ab-sorbed only 5% of ingested material above seston loads of10 mg⋅L–1 (D.W. Schneider, S.P. Madon, J.A. Stoeckel, andR.E. Sparks, unpublished data). Therefore, whatever little nu-tritional benefit that could be derived by ingesting supplemen-tary organic material from natural seston may be partlynegated by highly reduced assimilation efficiencies as sestonconcentrations increase.
Implications for growth potential in turbid riversOur results demonstrate that inorganic sediments have detri-mental effects on energetic processes, and thus scope forgrowth, of zebra mussels. Reductions in surplus energy thatcan be used for growth and reproduction indicate that zebramussels populations in turbid rivers may not stabilize at veryhigh densities typical of lentic environments such as the GreatLakes. As such, zebra mussels are better adapted to lentic en-vironments with superior food quality (low IO ratios), and theirlong-term success in turbid rivers will depend on their abilityto adjust their energetic processes to high inorganic sedimentloads.
A large floodplain system such as the Illinois River presentsa mosaic of habitats ranging from low flow and turbidity inbackwater lake areas to high flow and turbidity in main chan-nel regions. Total suspended solids in the main channel areasof the Illinois River range from 15 to 550 mg⋅L–1 (U.S. Geo-logical Survey 1991). Recent measurements in backwater,side, and main channel areas of the Illinois River in 1994reveal that inorganic sediment levels range from 8 to180 mg⋅L–1 (R. Wright, INHS-LTRMP, Havana, Ill., personal
communication). In 1994, the Illinois River exhibited sharptemporal and spatial variations in IO ratios of seston, whichranged from 0.45 to 8.84, and were often above 1.71 (INHS-LTRMP, Havana, Ill., unpublished data), our experimentalthreshold for positive growth of adult zebra mussels. Zebramussels may be able to persist in high numbers in those areasof the Illinois River that exhibit low turbidity and low IO ra-tios.
Although zebra mussels colonized the Illinois River in largenumbers between 1992 and 1993, their long-term success re-mains questionable. From 1993 to 1995, populations in manystretches of the Illinois River have suffered high mortality,often resulting in greater than 99% population reduction atseveral monitoring sites (S.D. Whitney, INHS-LTRMP, Ha-vana, Ill., personal communication). At some sites, die-offsmay have been partially caused by factors such as fluctuatingwater levels, low dissolved oxygen (<3.0 mg⋅L–1), and watertemperatures exceeding 30°C (S.D. Whitney, INHS-LTRMP,Havana, Ill., personal communication), especially if musselswere already in poor condition after being exposed to high IOratios. However, at other sites, water levels, dissolved oxygen,and water temperatures were favorable and cannot reconcilemajor die-offs at these sites. It is likely that poor food qualityat these sites may have caused high mortality. Unfortunately,no IO ratio data were collected at these sites to explore thepossibility of food quality as a contributing mortality factor.We suggest that future studies routinely incorporate collectionof IO ratio data in their protocol.
Acknowledgements
This research was funded by grant No. CR 820574 awarded toD.W.S. and R.E.S. by the U.S. Environmental ProtectionAgency and administered through the U.S.Environmental Pro-tection Agency Environmental Research Laboratory, Duluth,Minn. Dr. B.L. Bayne provided helpful suggestions on thedesign of our experimental apparatus. We thank M.K.Stephens and A. Grese for their help in the field and laboratory.D. Blodgett, S.D. Whitney, and other staff members at theLong Term Resource Monitoring Program of the Illinois Natu-ral History Survey provided help with the collection of zebramussels from the Illinois River. Diana Padilla, Wendell Haag,Clifford Kraft, and two anonymous reviewers critically re-viewed earlier drafts of this manuscript.
References
Aldridge, D.W., Payne, B.S., and Miller, A.C. 1987. The effects ofintermittent exposure to suspended solids and turbulence on threespecies of freshwater mussels. Environ. Pollut. 45: 17–28.
Alexander, J.E., Jr., Thorp, J.H., and Fell, R.D. 1994. Turbidity andtemperature effects on oxygen consumption in the zebra mussel(Dreissena polymorpha). Can. J. Fish. Aquat. Sci. 51: 179–184.
Arruda, J.A., Marzolf, G.R., and Faulk, R.T. 1983. The role of sus-pended sediments in the nutrition of zooplankton in turbid reser-voirs. Ecology, 64: 1225–1235.
Bayne, B.L., and Newell, R.C. 1983. Physiological energetics of ma-rine mollusks. In The Mollusca. Vol. 4. Physiology, Part 1. Editedby K.M. Wilbur and A.S.M. Saleuddin. Academic Press, London,U.K. pp. 407–515.
Bayne, B.L., and Widdows, J. 1978. The physiological ecology oftwo populations of Mytilus edulis L. Oecologia (Heidelb.), 37:137–162.
Madon et al. 411
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:58:00 1998
Color profile: DisabledComposite Default screen

Bayne, B.L., Bayne, C.J., Carefoot, T.C., and Thompson, R.J. 1976.The physiological ecology of Mytilus californianus Conrad. Me-tabolism and energy balance. Vol. 1. Oecologia (Heidelb.), 22:211–228.
Bayne, B.L., Widdows, J., and Newell, R.I.E. 1977. Physiologicalmeasurements on estuarine bivalve mollusks in the field. In Biol-ogy of benthic organisms. Proceedings of the 11th EuropeanMarine Biology Symposium. Edited by B.F. Keegan, P.O.Ceidigh, and J.J.S. Boaden. Pergamon Press, Oxford, U.K.pp. 57–68.
Bayne, B.L., Hawkins, A.J.S., and Navarro, E. 1987. Feeding anddigestion by the mussel Mytilus edulis L. (Bivalvia:Mollusca) inmixtures of silt and algal cells at low concentrations. J. Exp. Mar.Biol. Ecol. 111: 1–22.
Bayne, B.L., Hawkins, A.J.S., Navarro, E., and Iglesias, I.P. 1989.Effects of seston concentration on feeding, digestion and growthin the mussel Mytilus edulis. Mar. Ecol. Prog. Ser. 55: 47–54.
Bricelj, V.M., and Malouf, R.E. 1984. Influence of algal and sus-pended sediment concentrations on the feeding physiology of thehard clam Mercenaria mercenaria. Mar. Biol. 84: 155–165.
Bunt, C.M., MacIsaac, H.J., and Sprules, W.G. 1993. Pumping ratesand projected filtering impacts of juvenile zebra mussels (Dreis-sena polymorpha) in western Lake Erie. Can. J. Fish. Aquat. Sci.50: 1017–1022.
Conover, R.J. 1966. Assimilation of organic matter by zooplankton.Limnol. Oceanogr. 11: 338–345.
Cummins, K.W., and Wuycheck, J.C. 1971. Caloric equivalents forinvestigations in ecological energetics. Verh. Int. Ver. Theor.Angew. Limnol. 18.
Elliot, J.M., and Davidson, W. 1975. Energy equivalents of oxygenconsumption in animal energetics. Oecologia (Heidelb.), 19:195–201.
Foster-Smith, R.L. 1975. The effect of concentration of suspension onthe filtration rates and pseudofecal production for Mytilus edulis
L., Cerastoderma edule (L.) and Venerupsis pullastra (Montagu).J. Exp. Mar. Biol. Ecol. 17: 1–22.
Garton, D.W., and Haag, W.R. 1991. Heterozygosity, shell length andmetabolism in the zebra mussel, Dreissena polymorpha, from arecently established population in Lake Erie. Comp. Biochem.Physiol. A Comp. Physiol. 99: 45–48.
Griffiths, R.J. 1980. Natural food availability and assimilation in thebivalve Choromytilus meridionalis. Mar. Ecol. Prog. Ser. 3:151–156.
Griffiths, R.W. 1993. Effects of zebra mussel (Dreissena polymor-pha) on the benthic fauna of Lake St. Clair. In Zebra mussels:biology, impacts, and control. Chap. 25. Edited by T.F. Nalepa andD.W. Schloesser. Lewis Publishers, Inc., Boca Raton, Fla.
Haag, W.R., Berg, D.J., Garton, D.W., and Farris, J.L. 1993. Reducedsurvival and fitness in native bivalves in response to fouling by theintroduced zebra mussel (Dreissena polymorpha) in western LakeErie. Can. J. Fish. Aquat. Sci. 50: 13–19.
Hamburger, K., Dall, P.C., and Jonasson, P.M. 1990. The role ofDreissena polymorpha Pallas (Mollusca) in the energy budget ofLake Esrom, Denmark. Verh. Int. Ver. Theor. Angew. Limnol. 24:621–625.
Hebert, P.D.N., Muncaster, B.W., and Mackie, G.L. 1989. Ecologicaland genetic studies on Dreissena polymorpha (Pallas): a new mol-lusc in the Great Lakes. Can. J. Fish. Aquat. Sci. 46: 1587–1591.
Hebert, P.D.N., Wilson, C.C., Murdoch, M.H., and Lazar, R. 1991.Demography and ecological impacts of the invading molluscDreissena polymorpha. Can. J. Zool. 69: 405–409.
Illinois Natural History Survey. 1994. Zebra mussels in the IllinoisRiver. Rep. No. 325, Illinois Natural History Survey, Champaign,Ill. pp. 3–4.
Jørgensen, C.B. 1952. On the relation between water transport andfood requirements in some marine filter feeding invertebrates.Biol. Bull. 3: 356–363.
Kiørboe, T., and Møhlenberg, F. 1981. Particle selection in suspen-sion feeding bivalves. Mar. Ecol. Prog. Ser. 5: 291–296.
Kiørboe, T., Møhlenberg, F., and Nøhr, O. 1980. Feeding, particleselection and carbon absorption in Mytilus edulis in different mix-tures of algae and resuspended bottom material. Ophelia, 19:193–205.
MacIsaac, H.J., and Rocha, R. 1995. Effects of suspended clay onzebra mussel (Dreissena polymorpha) faeces and pseudofecesproduction. Arch. Hydrobiol. 135: 53–64.
MacIsaac, H.J., Sprules, W.G., Johannsson, O.E., and Leach, J.H.1992. Filtering impacts of larval and sessile zebra mussels (Dreis-sena polymorpha) in western Lake Erie. Oecologia (Heidelb.), 92:30–39.
Mackie, G.L. 1991. Biology of the exotic zebra mussels, Dreissenapolymorpha, in relation to native bivalves and its potential impactin Lake St. Clair. Hydrobiologia, 219: 251–268.
Mackie, G.L., Gibbons, W.N., Muncaster, B.W., and Gray, I.M. 1989.The zebra mussel, Dreissena polymorpha: a synthesis of Europeanexperiences and a preview for North America. Ontario Ministry ofNatural Resources, Water Resources Branch, Great Lakes Section.Queen’s Printer, Toronto, Ont.
Madon, S.P., and Culver, D.A. 1993. Bioenergetics model for larvaland juvenile walleyes: an in-situ approach with experimentalponds. Trans. Am. Fish. Soc. 122: 797–813.
Møhlenberg, F., and Kiørboe, T. 1981. Growth and energetics inSpisula subtruncata (Da Costa) and the effect of suspended bot-tom material. Ophelia, 20: 79–90.
Morton, B.S. 1969. Studies on the biology of Dreissena polymorphaPallas. I. General anatomy and morphology. Proc. Malacol. Soc.Lond. 38: 301–321.
Noordhuis, R., Reeders, H., and Bij de Vaate, A. 1992. Filtration rateand pseudofeces production in zebra mussels and their applicationin water quality management. In The zebra mussel Dreissena po-lymorpha: ecology, biological monitoring and first applications inwater quality management. Edited by D. Nuemann and H.A. Jen-ner. Gustav Fischer Verlag, Stuttgart, Germany. pp. 101–114.
Reeders, H.H., Bij de Vaate, A., and Slim, F.J. 1989. The filtrationrate of Dreissena polymorpha (Bivalvia) in three Dutch lakes withreference to biological water quality management. FreshwaterBiol. 22: 133–141.
Schneider, D.W. 1992. A bioenergetics model of zebra mussel, Dreis-sena polymorpha, growth in the Great Lakes. Can. J. Fish. Aquat.Sci. 49: 1406–1416.
Sprung, M., and Rose, U. 1988. Influence of food size and foodquantity on the feeding of the mussel Dreissena polymorpha. Oe-cologia (Heidelb.), 77: 526–532.
Strayer, D.L., Powell, J., Ambrose, P., Smith, L.C., Pace, M.L., andFischer, D.T. 1996. Arrival, spread, and early dynamics of a zebramussel (Dreissena polymorpha) population in the Hudson Riverestuary. Can. J. Fish. Aquat. Sci. 53: 1143–1149.
Ten Winkel, E.H., and Davids, C. 1982. Food selection by Dreissenapolymorpha Pallas (Mollusca: Bivalvia). Freshwater Biol. 12:553–558.
U.S. Geological Survey. 1983a. Water resources data for Minnesota.MN-83-2, U.S. Department of the Interior, St. Paul, Minn.
U.S. Geological Survey. 1983b. Water resources data for Missouri.MO-83-1, U.S. Department of the Interior, Jefferson City, Mo.
U.S. Geological Survey. 1991. Water resources data for Illinois. IL-91-2, U.S. Department of the Interior, Urbana, Ill.
Walz, N. 1978a. The energy balance of the freshwater mussel Dreis-
sena polymorpha Pallas in laboratory experiments and in LakeConstance. I. Pattern of activity, feeding and assimilation effi-ciency. Arch. Hydrobiol. Suppl. 55(1): 83–105.
Walz, N. 1978b. The energy balance of the freshwater mussel Dreis-sena polymorpha Pallas in laboratory experiments and in LakeConstance. II. Reproduction. Arch. Hydrobiol. Suppl. 55(1):106–119.
Can. J. Fish. Aquat. Sci. Vol. 55, 1998412
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:58:02 1998
Color profile: DisabledComposite Default screen

Walz, N. 1978c. The energy balance of the freshwater mussel Dreis-sena polymorpha Pallas in laboratory experiments and in LakeConstance. III. Growth under standard conditions. Arch. Hydro-bio. Suppl. 55(2): 121–141.
Walz, N. 1978d. The energy balance of the freshwater mussel Dreis-
sena polymorpha Pallas in laboratory experiments and in LakeConstance. IV. Growth in Lake Constance. Arch. Hydrobiol. Suppl.55(2): 142–156.
Walz, N. 1979. The energy balance of the freshwater mussel Dreissena
polymorpha Pallas in laboratory experiments and in Lake Con-stance. V. Seasonal and nutritional changes in biochemical com-position. Arch. Hydrobiol. Suppl. 55(3/4): 235–254.
Widdows, J., Fieth, P., and Worrall, C.M. 1979. Relationships be-tween seston, available food and feeding activity in the commonmussel Mytilus edulis. Mar. Biol. 50: 195–207.
Wilkinson, L. 1990. SYSTAT: the system for statistics. Systat, Inc.,Evanston, Ill.
Madon et al. 413
© 1998 NRC Canada
F97-214.CHPMon Apr 20 09:58:02 1998
Color profile: DisabledComposite Default screen

















![Genotoxicity assessment and detoxification induction in Dreissena polymorpha exposed to benzo[a]pyrene](https://static.fdokumen.com/doc/165x107/6344d92703a48733920b14f7/genotoxicity-assessment-and-detoxification-induction-in-dreissena-polymorpha-exposed.jpg)


