
Immature endodermal teratoma of the ovary: Embryologic correlations and immunohistochemistry

B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Effects of 5-hydroxytryptamine on substantia gelatinosaneurons of the trigeminal subnucleus caudalis inimmature mice
Hua Yin1, Seon Ah Park1, Seong Kyu Han, Soo Joung Park⁎
Department of Oral Physiology and Institute of Oral Bioscience, School of Dentistry, Chonbuk National University, Jeonju, Jeonbuk, 561-756,Republic of Korea
A R T I C L E I N F O
⁎ Corresponding author. School of Dentistry, Cof Korea. Fax: +82 63 270 4004.
E-mail address: [email protected] (S.J. PAbbreviations: ACSF, artificial cerebrospin
2,3-dione; EC50, effective concentration produ1-(m-chlorophenyl)-biguanide; RT-PCR, revecitrate; Vc, trigeminal subnucleus caudalis; Wboxamide maleate; 5-HT, 5-hydroxytryptam1 The first two authors contributed equally
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.10.050
A B S T R A C T
Article history:Accepted 14 October 2010Available online 26 November 2010
Serotonin (5-hydroxytryptamine, 5-HT) is involved in the descending modulation ofnociceptive transmission in the spinal dorsal horn. The trigeminal subnucleus caudalis(Vc; medullary dorsal horn) processes nociceptive input from the orofacial region, and 5-HT-containing axons are numerous in the superficial layers of the Vc. This study examined theactions of 5-HT on the substantia gelatinosa (SG) neurons of the Vc, using gramicidin-perforated patch-clamp recording in brainstem slice preparations from immature mice. Inorder to clarify the possible mechanisms underlying 5-HT actions in the SG of the Vc, thedirect membrane effects of 5-HT and effects of 5-HT receptor subtype agonists wereexamined. 5-HT induced a hyperpolarization in themajority (64/115, 56%) of the SG neuronstested. Thirty nine (34%) SG neurons showed no response, and 12 (10%) neurons respondedwith depolarization. The hyperpolarizing response to 5-HT was concentration-dependent(0.1–30 μM; n=7), not desensitized by repeated application (n=22), and significantlyattenuated by Ba2+ (K+ channel blocker; n=8). The 5-HT-induced hyperpolarization wasmaintained in the presence of TTX (Na+ channel blocker), CNQX (non-NMDA glutamatereceptor antagonist), AP5 (NMDA glutamate receptor antagonist), picrotoxin (GABAA
receptor antagonist), and strychnine (glycine receptor antagonist), indicating directpostsynaptic action of 5-HT on SG neurons (n=7). The 5-HT-induced hyperpolarizingeffects were mimicked by 8-OH-DPAT (5-HT1A receptor agonist) and α-methyl-5-HT (5-HT2
receptor agonist) and blocked by WAY-100635 (5-HT1A receptor antagonist) and ketanserin(5-HT2 receptor antagonist). Single-cell RT-PCR also revealed the presence of mRNA for 5-HT1A and 5-HT2C subtypes in the SG neurons. These results suggest that 5-HT acts directlyon SG neurons and 5-HT-induced hyperpolarization is mediated, in part, by 5-HT1A
receptors and 5-HT2 receptors, as well as by the activation of K+ channels, indicating an
Keywords:5-hydroxytryptamineTrigeminal subnucleus caudalisSubstantia gelatinosaGramicidin-perforated patch clampPostsynapticSingle-cell RT-PCR
honbuk National University, 664-14, 1 Ga, Deokjin-Dong, Jeonju, Jeonbuk, 561-756, Republic
ark).al fluid; AP5, D,L-2-amino-5-phosphonopentanoic acid; CNQX, 6-cyano-7-nitroquinoxaline-cing half-maximal response; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; mCPBG,rse transcriptase-polymerase chain reaction; SG, substantia gelatinosa; TTX, tetrodotoxinAY-100635, N-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-2-pyridinylcyclohexanecar-
ine; 8-OH DPAT, (±)-8-hydroxy-2-(di-n-propylamino) tetralinto this study.
er B.V. All rights reserved.
92 B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
important role for 5-HT in the modulation of orofacial nociceptive processing at the level ofthe SG of the Vc in mice.
© 2010 Elsevier B.V. All rights reserved.
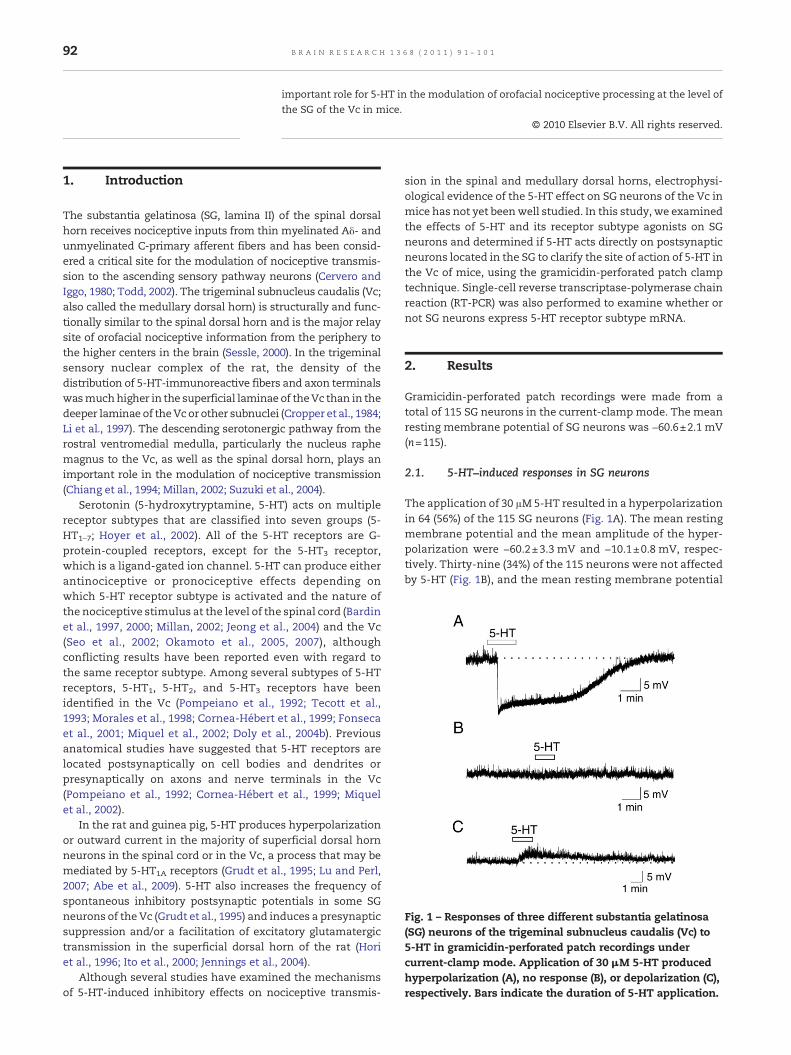
Fig. 1 – Responses of three different substantia gelatinosa(SG) neurons of the trigeminal subnucleus caudalis (Vc) to5-HT in gramicidin-perforated patch recordings undercurrent-clamp mode. Application of 30 μM 5-HT producedhyperpolarization (A), no response (B), or depolarization (C),respectively. Bars indicate the duration of 5-HT application.
1. Introduction
The substantia gelatinosa (SG, lamina II) of the spinal dorsalhorn receives nociceptive inputs from thin myelinated Aδ- andunmyelinated C-primary afferent fibers and has been consid-ered a critical site for the modulation of nociceptive transmis-sion to the ascending sensory pathway neurons (Cervero andIggo, 1980; Todd, 2002). The trigeminal subnucleus caudalis (Vc;also called the medullary dorsal horn) is structurally and func-tionally similar to the spinal dorsal horn and is the major relaysite of orofacial nociceptive information from the periphery tothe higher centers in the brain (Sessle, 2000). In the trigeminalsensory nuclear complex of the rat, the density of thedistribution of 5-HT-immunoreactive fibers and axon terminalswasmuchhigher in the superficial laminae of theVc than in thedeeper laminae of theVc or other subnuclei (Cropper et al., 1984;Li et al., 1997). The descending serotonergic pathway from therostral ventromedial medulla, particularly the nucleus raphemagnus to the Vc, as well as the spinal dorsal horn, plays animportant role in the modulation of nociceptive transmission(Chiang et al., 1994; Millan, 2002; Suzuki et al., 2004).
Serotonin (5-hydroxytryptamine, 5-HT) acts on multiplereceptor subtypes that are classified into seven groups (5-HT1–7; Hoyer et al., 2002). All of the 5-HT receptors are G-protein-coupled receptors, except for the 5-HT3 receptor,which is a ligand-gated ion channel. 5-HT can produce eitherantinociceptive or pronociceptive effects depending onwhich 5-HT receptor subtype is activated and the nature ofthe nociceptive stimulus at the level of the spinal cord (Bardinet al., 1997, 2000; Millan, 2002; Jeong et al., 2004) and the Vc(Seo et al., 2002; Okamoto et al., 2005, 2007), althoughconflicting results have been reported even with regard tothe same receptor subtype. Among several subtypes of 5-HTreceptors, 5-HT1, 5-HT2, and 5-HT3 receptors have beenidentified in the Vc (Pompeiano et al., 1992; Tecott et al.,1993; Morales et al., 1998; Cornea-Hébert et al., 1999; Fonsecaet al., 2001; Miquel et al., 2002; Doly et al., 2004b). Previousanatomical studies have suggested that 5-HT receptors arelocated postsynaptically on cell bodies and dendrites orpresynaptically on axons and nerve terminals in the Vc(Pompeiano et al., 1992; Cornea-Hébert et al., 1999; Miquelet al., 2002).
In the rat and guinea pig, 5-HT produces hyperpolarizationor outward current in the majority of superficial dorsal hornneurons in the spinal cord or in the Vc, a process that may bemediated by 5-HT1A receptors (Grudt et al., 1995; Lu and Perl,2007; Abe et al., 2009). 5-HT also increases the frequency ofspontaneous inhibitory postsynaptic potentials in some SGneurons of the Vc (Grudt et al., 1995) and induces a presynapticsuppression and/or a facilitation of excitatory glutamatergictransmission in the superficial dorsal horn of the rat (Horiet al., 1996; Ito et al., 2000; Jennings et al., 2004).
Although several studies have examined the mechanismsof 5-HT-induced inhibitory effects on nociceptive transmis-
sion in the spinal and medullary dorsal horns, electrophysi-ological evidence of the 5-HT effect on SG neurons of the Vc inmice has not yet beenwell studied. In this study, we examinedthe effects of 5-HT and its receptor subtype agonists on SGneurons and determined if 5-HT acts directly on postsynapticneurons located in the SG to clarify the site of action of 5-HT inthe Vc of mice, using the gramicidin-perforated patch clamptechnique. Single-cell reverse transcriptase-polymerase chainreaction (RT-PCR) was also performed to examine whether ornot SG neurons express 5-HT receptor subtype mRNA.
2. Results
Gramicidin-perforated patch recordings were made from atotal of 115 SG neurons in the current-clampmode. The meanresting membrane potential of SG neurons was −60.6±2.1 mV(n=115).
2.1. 5-HT–induced responses in SG neurons
The application of 30 μM5-HT resulted in a hyperpolarizationin 64 (56%) of the 115 SG neurons (Fig. 1A). The mean restingmembrane potential and the mean amplitude of the hyper-polarization were −60.2±3.3 mV and −10.1±0.8 mV, respec-tively. Thirty-nine (34%) of the 115 neurons were not affectedby 5-HT (Fig. 1B), and the mean resting membrane potential

93B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
of neurons showing no response was −60.6±3.2 mV. Howev-er, 5-HT induced a depolarization in 12 (10%) neurons (7.9±1.5 mV; Fig. 1C), and the mean resting membrane potentialof neurons excited by 5-HT was −60.9±2.8 mV. There was nosignificant difference in the resting membrane potentialsamong the various responding groups that were examined(P>0.05, one-way ANOVA).
The responses to 5-HT were reversible and reproducible.When 30 μM 5-HT was applied consecutively to the SG neu-rons showing hyperpolarizing responses to 5-HT, a secondapplication of 5-HT produced a hyperpolarization of similaramplitude to that produced by the first application in 22 SGneurons (Fig. 2A). There was no significant differencein mean amplitude of hyperpolarization between the first(−10.2±1.2 mV) and the second application (−9.5±1.3 mV) of5-HT (P>0.05, paired t-test; Fig. 2B). The 5-HT-inducedhyperpolarizations were concentration-dependent (0.1–30 μM; Fig. 3A), and an effective concentration producinghalf-maximal response (EC50) was 0.6 μM (Fig. 3B). The meanamplitude and the mean duration of hyperpolarizing re-sponse to 5-HT increased with increasing 5-HT concentra-tions (n=7; Fig. 3B and C).
2.2. 5-HT acts postsynaptically on SG neurons
To determine the site of the hyperpolarizing action of 5-HT onSG neurons, we examined 5-HT-induced hyperpolarization inthe presence of TTX (Na+ channel blocker) to block the actionpotential-dependent transmission, as well as AP5 (NMDAglutamate receptor antagonist), CNQX (non-NMDA glutamate
Fig. 2 – Responses to repeated application of 30 μM 5-HTin SG neurons. (A) The amplitude of 5-HT-inducedhyperpolarization by a second applicationwas similar to thatby the first application. Bars indicate the duration of 5-HTapplication. (B) No significant difference was found in themean amplitude of 5-HT-induced hyperpolarization betweenthe first and second applications of 5-HT (n=22, P>0.05). NS,not significant.
receptor antagonist), picrotoxin (GABAA receptor antagonist),and strychnine (glycine receptor antagonist) to rule outpresynaptic 5-HT actions mediated by glutamate, GABA, orglycine released from presynaptic nerve terminals. 5-HT-induced hyperpolarization persisted in the presence of0.5 μM TTX, 20 μM AP-5, 10 μM CNQX, 50 μM picrotoxin, and2 μM strychnine in 7 SG neurons (Fig. 4A), indicating a directpostsynaptic action of 5-HT on recorded SG neuron. Therewere no significant differences between the mean amplitudeaswell as themean duration of hyperpolarization produced by5-HT alone (−11.0±1.6 mV and 5.3±1.0 min) and those pro-duced by the 5-HT in the presence of the above-mentionedblockers (−9.7±1.5 mV and 5.1±1.1 min; n=7, both P>0.05,paired t-test; Fig. 4B and C).
2.3. Potassium ion channels, 5-HT1A and 5-HT2 receptorsare involved in 5-HT-induced hyperpolarization
To determine the ion channel involved in 5-HT-inducedhyperpolarization, we applied 0.5 mM BaCl2 (K+ channelblocker) with 5-HT in SG neurons, showing a hyperpolariz-ing response to 30 μM 5-HT. Immediately after the applica-tion of Ba2+, the action potentials were generated transientlywith various firing rates in 4 of 8 neurons but subsidedafterwards when we applied again 5-HT in the presence ofBa2+. BaCl2 completely or partially blocked the 5-HT-inducedhyperpolarization in 7 of 8 SG neurons (Fig. 5A). Theremaining neuron was unresponsive to BaCl2. The meanamplitude of the hyperpolarization was significantly re-duced from −13.3±2.4 mV by 5-HT alone to −5.6±1.7 mV by5-HT in the presence of BaCl2 (n=8, P<0.05, paired t-test;Fig. 5B).
To identify the receptor subtypes involved in 5-HT-induced hyperpolarization, we examined the effects of 5-HTreceptor subtype agonists in SG neurons that were hyperpo-larized by 5-HT (Fig. 6A). The major effects of each agonistin SG neurons hyperpolarized by 5-HT were as follows: the 5-HT1 receptor agonist, 8-OH-DPAT (30 μM), and the 5-HT2
receptor agonist, α-methyl-5-HT (30 μM), induced hyperpo-larization in 6 (75%) of 8 neurons and in 8 (73%) of 11 neurons,respectively. The 5-HT3 receptor agonist, mCPBG (100 μM),produced depolarization in 7 (64%) of 11 neurons. The 5-HT4
receptor agonist, RS 67506 (30 μM), induced hyperpolariza-tion and depolarization in 3 (38%) and 2 (25%) of 8 neurons,respectively. The effects of 5-HT receptor subtype agonists inSG neurons showing hyperpolarizing responses to 5-HT aresummarized in Fig. 6B. In addition, the 5-HT1 receptorantagonist, WAY-100635 (1 μM) and the 5-HT2 receptorantagonist, ketanserin (30 μM) attenuated the 5-HT-inducedhyperpolarization (Fig. 6C). The mean amplitude of hyperpo-larizing response to 30 μM 5-HT was significantly attenuatedby 1 μMWAY-100635 (from −9.3±0.5 mV to −5.4±0.5 mV; n=4,P<0.05, paired t-test) and by 30 μM ketanserin (from −8.7±0.7 mV to −4.3±0.5 mV; n=4, P<0.05, paired t-test).
On the other hand, in SG neurons showing a depolarizingresponse to 5-HT, the 5-HT2 receptor agonist, α-methyl-5-HT(30 μM), and the 5-HT3 receptor agonist, mCPBG, induceddepolarization in 2 of 2 neurons and 2 of 3 neurons,respectively. Some SG neurons (4/14, 29%) showing noresponse to 5-HT, responded with depolarization or
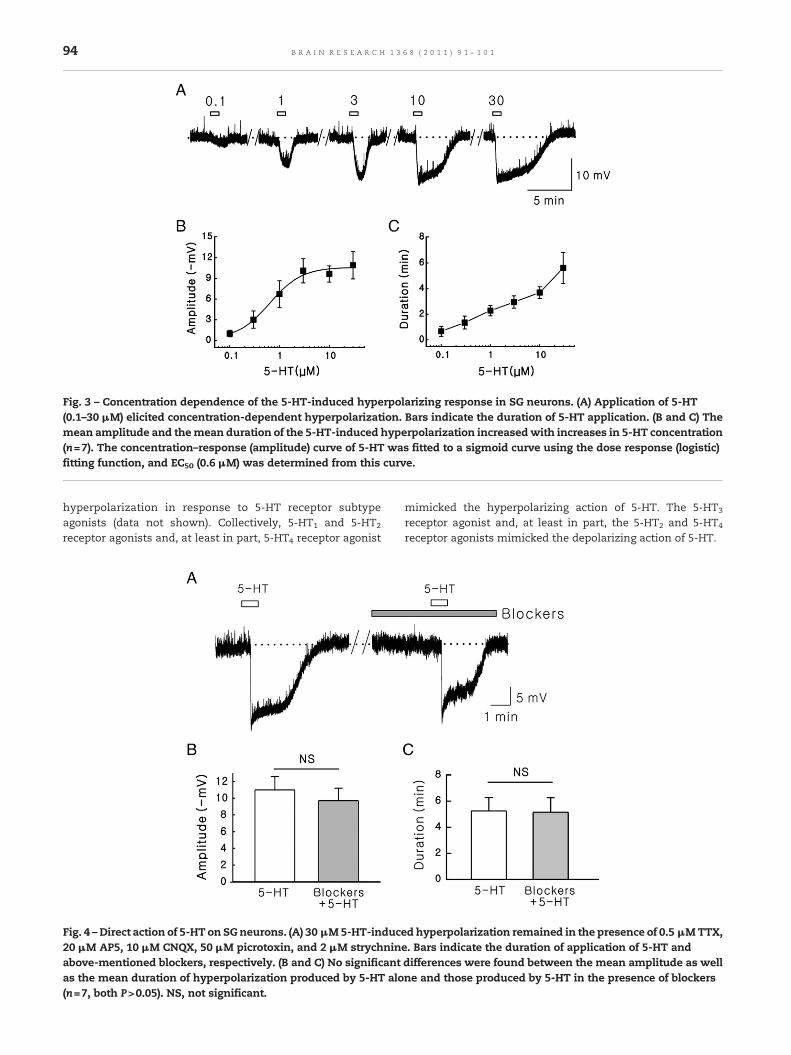
Fig. 3 – Concentration dependence of the 5-HT-induced hyperpolarizing response in SG neurons. (A) Application of 5-HT(0.1–30 μM) elicited concentration-dependent hyperpolarization. Bars indicate the duration of 5-HT application. (B and C) Themean amplitude and themean duration of the 5-HT-induced hyperpolarization increasedwith increases in 5-HT concentration(n=7). The concentration–response (amplitude) curve of 5-HT was fitted to a sigmoid curve using the dose response (logistic)fitting function, and EC50 (0.6 μM) was determined from this curve.
94 B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
hyperpolarization in response to 5-HT receptor subtypeagonists (data not shown). Collectively, 5-HT1 and 5-HT2
receptor agonists and, at least in part, 5-HT4 receptor agonist
Fig. 4 –Direct action of 5-HT on SGneurons. (A) 30 μM5-HT-induc20 μM AP5, 10 μM CNQX, 50 μM picrotoxin, and 2 μM strychninabove-mentioned blockers, respectively. (B and C) No significantas the mean duration of hyperpolarization produced by 5-HT alo(n=7, both P>0.05). NS, not significant.
mimicked the hyperpolarizing action of 5-HT. The 5-HT3
receptor agonist and, at least in part, the 5-HT2 and 5-HT4
receptor agonists mimicked the depolarizing action of 5-HT.
ed hyperpolarization remained in the presence of 0.5 μMTTX,e. Bars indicate the duration of application of 5-HT anddifferences were found between the mean amplitude as wellne and those produced by 5-HT in the presence of blockers

Fig. 5 – Effects of the K+ channel blocker Ba2+ on 5-HT-inducedhyperpolarization in SG neurons. (A) 0.5 mM BaCl2 partiallyblocked the hyperpolarization induced by 30 μM 5-HT. Barsindicate the duration of application of 5-HT and BaCl2,respectively. (B) Mean amplitudes of 5-HT-inducedhyperpolarization. BaCl2 significantly attenuates5-HT-induced hyperpolarization (n=8, *P<0.05).
95B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
In support of electrophysiological responses of SG neuronsto 5-HT receptor subtype agonists, single-cell RT-PCR revealedthe presence of mRNA for 5-HT receptor subtypes in the SGneurons (Fig. 7). We examined mRNA expression in SGneurons that showed the presence of the housekeeping geneglyceraldehyde 3-phosphate dehydrogenase (GAPDH). Ten(36%) of 28 SG neurons expressed 5-HT1A receptor mRNA, 16(48%) of 33 SG neurons expressed 5-HT2C receptor mRNA, 5(12%) of 43 SG neurons expressed 5-HT3A receptor subunitmRNA, and 1 (3%) of 35 SG neurons expressed 5-HT4 receptormRNA.
3. Discussion
Using gramicidin-perforated patch clamp recordings incurrent clamp mode, we examined the effects of 5-HT onSG neurons of the Vc of immature mice. Gramicidin formssmall pores in the cell membrane allowing only monovalentcations to permeate the membrane and preserving theintracellular second messenger system (Akaike, 1996). Themajority of SG neurons tested were hyperpolarized by 5-HTapplication. The 5-HT-induced hyperpolarizing effects wereconcentration-dependent, not desensitized by repeated ap-plication, and greatly diminished by Ba2+. The 5-HT-inducedhyperpolarization was maintained in the presence of TTX,CNQX, AP5, picrotoxin, and strychnine, indicating a directpostsynaptic action of 5-HT on SG neurons. The 5-HT-induced hyperpolarization was mimicked by 8-OH-DPATand α-methyl-5-HT and blocked by WAY-100635 and ketan-
serin. Single-cell RT-PCR revealed the presence of mRNA for5-HT1A, 5-HT2C, 5-HT3, and 5-HT4 subtypes in the SG neurons.
3.1. Themajority of SG neurons in the Vc are hyperpolarizedby 5-HT
The SG in the Vc and spinal dorsal horn is the importantrelay site for the transmission of nociceptive informationfrom primary afferent fibers (Cervero and Iggo, 1980).Previous electrophysiological studies using slice prepara-tions have shown that 5-HT produces hyperpolarization, oroutward current, in more than half of the SG neurons anddepolarization, or inward current, in a small population ofneurons both in the spinal dorsal horn (Lu and Perl, 2007; Abeet al., 2009) and the Vc (Grudt et al., 1995). In this study, wealso found that the majority (56%) of SG neurons tested werehyperpolarized, the 5-HT-induced hyperpolarizing effectswere concentration-dependent, and only 10% of the SGneurons were depolarized in mice. Both 5-HT and noradren-aline aremajor neurotransmitters of descendingmodulatorypathways originating from the brainstem (Millan, 2002) andexert similar effects on SG neurons (Grudt et al., 1995; Lu andPerl, 2007). Our previous study also has demonstrated thatthe majority (85%) of SG neurons are hyperpolarized byapplication of noradrenaline (Han et al., 2007). Yoshimuraand Furue (2006) has pointed out some quantitative differ-ences between the descending noradrenergic and seroto-nergic pathways in the spinal dorsal horn. G protein-coupledreceptors can be desensitized upon repeated application ofthe same agonist due to phosphorylation and uncoupling ofthe receptor from the G proteins (Kelly et al., 2008). Thisstudy revealed no noticeable decrease in 5-HT-inducedhyperpolarization in response to repeated 5-HT applications,indicating a lack of strong desensitization.
3.2. 5-HT has postsynaptic actions on SG neurons
Descending modulation of nociceptive transmission by 5-HTin the SG can occur presynaptically on the axon terminals ofthe first primary afferents or SG neurons, or postsynapticallyon the cell body or dendrites of SG neurons. The 5-HT-induced outward currents were not affected by the simulta-neous application of TTX (Abe et al., 2009). In this study, the5-HT-induced hyperpolarization was not significantly af-fected by pretreatment with TTX, AP5, CNQX, picrotoxin, andstrychnine. These results indicate that the 5-HT-inducedhyperpolarization in the SG of the Vc was mediated by directaction at postsynaptic 5-HT receptors, rather than byindirect action at presynaptic 5-HT receptors, i.e., actionpotential dependent- or action potential independent-neu-rotransmitter release from the axon terminals. Previousanatomical studies also indicate postsynaptic localizationof 5-HT1A receptors (Pompeiano et al., 1992), 5-HT2A recep-tors (Cornea-Hébert et al., 1999), and 5-HT3 receptors (Miquelet al., 2002) in the Vc. In the superficial layers of the spinaldorsal horn, immunoreactivities for 5-HT2A receptors (Dolyet al., 2004b) and 5-HT5A receptors (Doly et al., 2004a) weremainly found in postsynaptic cell bodies and dendrites, andat least some neurons show postsynaptic expression of 5-HT3 receptors (Miquel et al., 2002) and 5-HT7 receptors (Doly

96 B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
et al., 2005). These results suggest that a postsynapticmechanism is involved in the 5-HT-induced hyperpolariza-tion in the SG, although we cannot rule out some presynapticeffect of 5-HT (Hori et al., 1996; Ito et al., 2000; Jennings et al.,2004).
3.3. Potassium channels, 5-HT1A and 5-HT2 receptors areinvolved in 5-HT-induced hyperpolarization
Barium ions block a variety of K+ channels, and theapplication of 0.5 mM Ba2+ greatly reduced but did notcompletely inhibit 5-HT-induced hyperpolarization, a resultthat is consistent with a previous study in the SG of spinaldorsal horn (Abe et al., 2009). The hyperpolarizing effect of 5-HT was suggested to be mediated through the 5-HT1A
receptor and accompanying activation of the G-protein-coupled inwardly rectifying K+ channel in the lateralhypothalamic area (Muraki et al., 2004). G-protein-coupledinwardly rectifying K+ channel subunits are concentrated inthe SG of spinal dorsal horn and located almost exclusivelyat postsynaptic sites of putative excitatory synapses (Markeret al., 2005). However, our data should be interpreted withcaution because Ba2+ itself caused a burst of action poten-tials, in some neurons, which could influence on a manyvoltage-gated ion channels. Similar phenomena have beenfound in other whole-cell patch clamp experiments per-
Fig. 6 – Effects of 5-HT receptor subtype agonists and antagonistsresponses to 5-HT receptor subtype agonists in different neurons8-OH-DPAT (30 μM) andα-methyl-5-HT (30 μM). Bars indicate thof 5-HT receptor subtype agonists on SG neurons. 5-HT1 receptoα-methyl-5-HT (30 μM); 5-HT3 receptor agonist, mCPBG (100 μMhyperpolarization, depolarization or no response. The numbers(C) 5-HT1 receptor antagonist, WAY-100635 (1 μM) and 5-HT2 re5-HT-induced hyperpolarization, respectively.
formed on SG neurons of spinal dorsal horn in thatapplication of Ba2+ alone induced a small inward currentprobably due to a blockade of K+ channels (Jiang et al., 2003;Miyakawa et al., 2005; Tamae et al., 2005). We have chosencurrent clamp mode because the accompanying high accessresistance in perforated patch-clamp technique and thesignificant resistances offered by dendrites and axon mayintroduce errors in voltage clamp (Molleman, 2003). Despitedifficulties in interpretation, these results suggest that K+
channels are involved, at least partly, in the mechanism ofthe hyperpolarizing action of 5-HT.
Among various 5-HT receptor subtypes, the 5-HT1, 5-HT2,and 5-HT3 receptors are presumed to be involved in themodulation of nociceptive transmission in the Vc (Grudt et al.,1995; Seo et al., 2002; Jennings et al., 2004; Okamoto et al., 2005,2007). This study indicates that 5-HT-induced hyperpolariza-tion is mediated by the activation of 5-HT1A, 5-HT2, and some5-HT4 receptors and that the 5-HT-induced depolarization ismediated by the activation of 5-HT3 and some 5-HT2 and 5-HT4
receptors in the SG of the Vc. Detection of mRNA for 5-HT1A, 5-HT2C, 5-HT3, and 5-HT4 receptor subtypes by single-cell RT-PCR analysis in this study also supports the presence of 5-HTreceptor subtypes in SG neurons. The expressions of mRNAfor 5-HT3 and 5-HT4 receptors were detected in a relativelysmall number of SG neurons, which can be due to preferentialpresynaptic localization of their receptors (Miquel et al., 2002).
on SG neurons thatwere hyperpolarized by 5-HT. (A) Typical. The 5-HT-induced hyperpolarizing effect wasmimicked bye duration of agonist application. (B) Summary of the effectsr agonist, 8-OH-DPAT (30 μM); 5-HT2 receptor agonist,); and 5-HT4 receptor agonist, RS 67506 (30 μM) producedin the columns represent the number of neurons responded.ceptor antagonist, ketanserin (30 μM) attenuated the 30 μM

Fig. 7 – Examples of single-cell RT-PCR analysis of mRNA for5-HT receptor subtypes in SG neurons. (A) We examinedmRNA expression for 5-HT receptor subtype in SG neuronsthat showed the presence of the housekeeping gene GAPDH.Examples of GAPDH expression of samples used in (B).(B) 5-HT1A receptor mRNA was detected in 4 of the 10neurons. (C) 5-HT2C receptor mRNAwas detected in 6 of the 9neurons. (D) 5-HT3A receptor subunitmRNAwas detected in 1of the 9 neurons. (E) 5-HT4 receptor mRNA was detected in1 of the 9 neurons. L; 100 bp ladder, S1–S10; harvested singleSG neurons, T1 and T2; touching surface of the brainstemslice, N1 and N2; without reverse transcriptase for negativecontrol, PC; whole brain tissue for positive control.
97B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
Consistent with these findings, single-cell RT-PCR in thesuperficial layers of the mouse spinal dorsal horn alsoshowed mRNA expression of the 5-HT1A receptor in almosthalf of the neurons tested, and expression of the 5-HT2A
receptor and 5-HT3A receptor subunit in a smaller populationof the neurons tested (Huang et al., 2008; Wang et al., 2009).
Activation of the 5-HT1A receptor has been shown tomediate hyperpolarization or outward current in SG neuronsof the medullary or spinal dorsal horn (Grudt et al., 1995; Abeet al., 2009). 5-HT1A receptor activation leads to the inhibitionof adenylate cyclase and the opening of K+ channels in othercentral neurons (Lanfumey and Hamon, 2000). However,results concerning the roles of 5-HT2, 5-HT3, and 5-HT4
receptors in nociceptive processing are not consistent. A 5-HT2 receptor agonist depolarized a subpopulation of SGneurons of the Vc of guinea-pigs (Grudt et al., 1995) butelicited no response in SG neurons of the spinal dorsal hornin rats (Abe et al., 2009). A 5-HT3 agonist mimicked 5-HT-induced inward current, but a 5-HT4 agonist did not elicit any
outward or inward currents in the SG neurons of the spinaldorsal horn (Abe et al., 2009). A 5-HT3 agonist significantlyenhanced or suppressed jaw muscle activity evoked by MOinjection into the temporomandibular joint (Seo et al., 2002).Antinociceptive roles of 5-HT2A receptors and pronociceptiveroles of 5-HT3 receptors in orofacial nociceptive modulationwere suggested in that activation of 5-HT2A receptors orblocking of 5-HT3 receptors in the caudal portion of the Vcand upper cervical spinal cord reduced orofacial nocifensivebehaviors and nociceptive neural responses recorded fromthe superficial laminae of the above region (Okamoto et al.,2005, 2007).
The trigeminal brainstem sensory complex displays someproperties different from the spinal somatosensory system(Bereiter et al., 2000). For example, the extensive convergencepattern of the Vc, involving tooth pulp, periodontium, jawmuscles, temporomandibular joint, cornea, or dural vesselsin addition to cutaneous tissue, is a distinguishing featurethat differentiates the trigeminal brainstem sensory com-plex from the spinal cord (Sessle, 2000). The diverse effects of5-HT and its agonists on SG neurons might be due todifferences in animals (guinea pig, rat or mouse), centralnervous system regions (spinal or medullary dorsal horn),the location of receptors at the synapse (presynaptic orpostsynaptic), types of neurons (excitatory or inhibitory), 5-HT receptor subtypes (5-HT1–7 receptors) and associated Gproteins, and the patch clamp technique (whole-cell orgramicidin-perforated). Selective agonists and antagonistsfor some 5-HT receptor subtypes are not yet available, andfurther studies are required for better understanding of therole of each 5-HT receptor subtype in the descendingmodulation of orofacial nociception.
3.4. Physiological significance
Descending serotonergic pathways from the brainstem to thespinal cord have been reported to either inhibit or facilitatetransmission of nociceptive information (Millan, 2002). One ofthe major targets of descending serotonergic innervation isthe SG of the spinal dorsal horn (Marlier et al., 1991) and the Vc(Cropper et al., 1984; Li et al., 1997). The SG includesstructurally and functionally distinct types of neurons (Grudtand Perl, 2002; Davies and North, 2009), with some SG neuronstransmitting nociceptive information to neurons in lamina I orlaminae III-V (Gobel, 1978; Light and Kavookjian, 1988; Grudtand Perl, 2002).
According to anatomical studies, excitatory neurons com-prise a large proportion of neurons in the SG (Todd and Sullivan,1990; Landry et al., 2004). Recent electrophysiological studiesalso found that excitatory glutamatergic interneurons predom-inate within the SG neuronal circuitry (Lu and Perl, 2005; Santoset al., 2007). The 5-HT-induced outward currents are presumedto be evoked in the excitatory SG neurons through the 5-HT1A
receptor (Abe et al., 2009). Therefore, the descending serotoner-gic inhibition of orofacial nociceptive transmission can beattributed to a hyperpolarizing effect of 5-HT on the predomi-nant excitatory interneurons or projection neurons in the SG ofthe Vc. On the other hand, 5-HT3 receptors are present on axonterminalsof excitatoryneurons in the spinaldorsalhornandcancontribute to pronociceptive action of 5-HT (Conte et al., 2005).

98 B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
Inhibitory interneurons in the SG of the Vc are suggestedto be strategically located between primary afferents andprojection neurons in lamina I (Gobel, 1978). 5-HT producesinward currents in certain inhibitory interneurons in the ratSG (Lu and Perl, 2007). Immunocytochemical studies haverevealed that 5-HT axons contact cell bodies and dendrites ofenkephalin-containing neurons in the SG of the rat Vc (LópezCosta et al., 1994). The 5-HT3A receptor subunit is expressedin a subpopulation of GABAergic and enkephalinergicneurons in the mouse superficial dorsal horn (Huang et al.,2008). Antinociceptive effects of 5-HT in the spinal dorsalhorn may be induced by the excitation of inhibitory inter-neurons through 5-HT3 receptors, the release of GABA andenkephalin, and the subsequent inhibition of nociceptivetransmission (Kawamata et al., 2003). On the other hand,application of 5-HT generates outward currents in GABAergicgreen fluorescent protein neurons of the SG (Hantman andPerl, 2005). Some GABAergic and enkephalinergic neuronssimultaneously express 5-HT1A receptor mRNA in the super-ficial dorsal horn, and the hyperpolarizing effect of 5-HTon these inhibitory interneurons, which ismediated by the 5-HT1A receptor, can then induce disinhibition of nociceptiveprocessing (Zhang et al., 2002; Wang et al., 2009).
Descending serotonergic fibers terminate on excitatory orinhibitory interneurons in the SG and depolarizing andhyperpolarizing effects of 5-HT on these interneurons mightresult in antinociception or pronociception.
4. Experimental procedures
4.1. Animals and brainstem slice preparation
All experiments were approved by the Experimental AnimalCare and Ethics Committee of Chonbuk National University.Mice (Damul Science, Suwon, Korea) were housed under12:12 light:dark cycles with free access to food and water.Immature mice (postnatal 6–18 days) of either sex weredecapitated at between 10:00 and 12:00 h. The brains of intactICR mice were quickly removed and coronal brainstem slices(150–200 μm thickness), containing the Vc, were cut using avibratome (Microm, Walldorf, Germany) in ice-cold artificialcerebrospinal fluid (ACSF) containing (in mM) 126 NaCl, 2.5KCl, 2.4 CaCl2, 1.2 MgCl2, 11 D-glucose, 1.4 NaH2PO4 and 25NaHCO3 (pH 7.4, bubbled with 95% O2 and 5% CO2). Sliceswere allowed to recover in oxygenated ACSF for at least 1 h atroom temperature.
4.2. Electrophysiology
The brainstem slices were transferred to the recordingchamber, submerged, and superfused continuously withoxygenated ACSF at a rate of 4–5 ml/min. The slices wereviewed on the stage of an upright microscope (BX51WI,Olympus, Tokyo, Japan) using Nomarski differential interfer-ence contrast optics. The SG of the Vc was clearly visible as atranslucent band just medial to the spinal trigeminal tracttravelling along the lateral edge of the slice. Patch pipetteswere pulled from thin-walled borosilicate glass capillaries(PG52151-4, WPI, Sarasota, FL, USA) on a Flaming/Brown
micropipette puller (P-97; Sutter Instruments Co., Novato,CA, USA). The pipette solution was passed through adisposable 0.22 μm filter and contained (in mM): 130 KCl, 5NaCl, 0.4 CaCl2, 1 MgCl2, 10 HEPES and 1.1 EGTA (pH 7.3 withKOH). Gramicidin (Sigma, St. Louis, MO, USA) was firstdissolved in dimethylsulfoxide (Sigma) to a concentration of2.5–5 mg/ml and then diluted in the pipette solution, justbefore use, to a final concentration of 2.5–5 μg/ml andsonicated for 10 min. The gramicidin-perforated patch record-ings were made from SG neurons in current-clampmode. Thetip resistances of the electrodes were between 5 and 8 MΩ.Offset potentials were compensated directly before formationof a seal. After cell attachment, access resistance wasmonitored and experiments began when in the range of 40–100 MΩ. Recordings began 15–20 min after giga-seal formationwhen the resting membrane potential of the cell reached astable level below −45mV. The signals were acquired with apatch-clamp amplifier (Axopatch 200B, Axon Instruments, SanFrancisco, CA, USA), digitized with an A/D converter (Digidata1322A, Axon Instruments), and stored and analyzed with apersonal computer using Clampex 9 software (Axon Instru-ments). The signals were filtered (2 kHz, Bessel filter ofAxopatch 200B) prior to digitizing at a rate of 1 kHz. Allrecordings were made at room temperature.
4.3. Drugs
The drugs used were, as follows: 5-HT (0.1–30 μM); tetrodo-toxin citrate (TTX, 0.5 μM); 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX, 10 μM); D,L-2-amino-5-phosphonopentanoicacid (AP-5, 20 μM); picrotoxin (50 μM); strychnine hydrochlo-ride (2 μM); BaCl2 (0.5 mM); (±)-8-hydroxy-2-(di-n-propyla-mino) tetralin (8-OH DPAT, 30 μM); α-methyl-5-HT maleate(30 μM); 1-(m-chlorophenyl)-biguanide (mCPBG, 100 μM), RS67506 hydrochloride (30 μM), N-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-2-pyridinylcyclohexanecarboxamidemaleate (WAY-100635, 1 μM), and ketanserin tartrate (30 μM).TTX, mCPBG and RS 67506 were purchased from TocrisBioscience (Ellisville, MO, USA), and all other drugs werepurchased from Sigma.
Stock solutions of all drugs were made in distilled water,except picrotoxin and ketanserin, which were made indimethylsulfoxide. Stock solutions were diluted to workingconcentrations using ACSF immediately before use andapplied by superfusion. We applied 5-HT and its agonists for30 s–1 min and allowed awashout/recovery period of 5–10 minbetween subsequent applications.
4.4. Single cell RT-PCR
Patch electrodes used for harvesting were baked at 250 °C for6 h before being pulled. The cytoplasmic content was har-vested under visual control using a patch electrode filled with8 μl autoclaved solution containing (inmM) 140 KCl, 10 EGTA, 1MgCl2, 1 CaCl2, and 10 HEPES (pH 7.3). As a control for thedebris that might enter the pipette and be amplified, mockharvest was conducted, in which the patch electrode waslowered to the surface of the slice but cellular contents werenot aspirated. Immediately after harvesting, the individualcellular content was expelled from the patch electrode into a

Table 1 – Primers used for mRNA detection of 5-HT receptor subtypes.
Gene Accession number Sequence Product size (bp) Tm (°C)
5-HT1A NM_008308 5′GCATTTCTTTTTCCCTCCCTTCC3′ 368 bp 565′TGACCCAGAGTCCACTTGTTGAG3′
5-HT2C NM_008312 5′TCTCCCTTCCTTCCGTATTCCC3′ 452 bp 625′TGGCATCCTTCCACTTCTGTAGTC3′
5-HT3A NM_013561 5′CATGTATGCCATCCTCAACG3′ 310 bp 625′CTGCACATCAAAGGGGAAGT3′
5-HT4 NM_008313 5′TGAGTTCCAACGAGGGTTTCAG3′ 398 bp 605′TAATGCGATGCGTAGAGGGG3′
GAPDH M32599 5′TTGGCATTGTGGAAGGGCTC3′ 363 bp 625′TGCTGTTGAAGTCGCAGGAGAC3′
bp, base pair, Tm, melting temperature.
99B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
PCR tube containing RT pre-mixture consisting of 0.7 μl of 3 μg/μl random primers, 0.7 μl of 40 U/μl RNase OUT™ (Invitrogen,Carlsbad, CA, USA) and distilled water to a final volume of 5 μl.The cytoplasmic content and RT pre-mixture were mixed andthen collected after brief centrifugation. The sample wasincubated at 65 °C for 5 min, then on ice for at least 1 min.After the first reaction, RT reaction solution containing 4 μl of 5×RTbuffer, 2 μl of 0.1 MDTT, 1 μl of 10mMdNTP, 0.5 μl of 200U/μlSuperscriptase III (Invitrogen) and 0.5 μl of distilled water wasadded. The cDNA synthesis was performed at 25 °C for 5 min,then 50 °C for 1 h, and terminated by heating to 70 °C for 15min.The RT products were then kept in a freezer at −20 °C. Thesequences of RT-PCR primers for 5-HT receptor subtypes arepresented in Table 1. PCR was performed through 50 cycles ofamplification using the following conditions: denaturation at94 °C for 30 s, annealingat 56 °C, 60 °Cor 62 °C, dependingon theprimer pair, for 30 s, and elongation at 72 °C for 30 s. This wasfollowed by a final extension at 72 °C for 10min. The PCRproducts were electrophoresed on a 1.5% agarose gel (Promega,Madison, WI, USA) and then analyzed using Lab systemsanalyzer (Bio-Rad, Milano, Italy).
4.5. Statistical analysis
All values were expressed as the mean ± S.E.M. Differencesbetween baseline (pre-drug) values and post-drug values weretreated by a paired t-test, and differences among experimentalgroups were treated by a one-way ANOVA. The level ofsignificance was set at P<0.05.
Acknowledgments
This research was supported by Basic Science ResearchProgram through the National Research Foundation of Korea(NRF) funded by the Ministry of Education, Science andTechnology (2009-0070100).
R E F E R E N C E S
Abe, K., Kato, G., Katafuchi, T., Tamae, A., Furue, H., Yoshimura,M., 2009. Responses to 5-HT in morphologically identifiedneurons in the rat substantia gelatinosa in vitro. Neuroscience159, 316–324.
Akaike, N., 1996. Gramicidin perforated patch recording andintracellular chloride activity in excitable cells. Prog. Biophys.Mol. Biol. 65, 251–264.
Bardin, L., Bardin, M., Lavarenne, J., Eschalier, A., 1997. Effect ofintrathecal serotonin on nociception in rats: influence of thepain test used. Exp. Brain Res. 113, 81–87.
Bardin, L., Lavarenne, J., Eschalier, A., 2000. Serotonin receptorsubtypes involved in the spinal antinociceptive effect of 5-HTin rats. Pain 86, 11–18.
Bereiter, D.A., Hirata, H., Hu, J.W., 2000. Trigeminal subnucleuscaudalis: beyond homologies with the spinal dorsal horn. Pain88, 221–224.
Cervero, F., Iggo, A., 1980. The substantia gelatinosa of the spinalcord: a critical review. Brain 103, 717–772.
Chiang, C.Y., Hu, J.W., Sessle, B.J., 1994. Parabrachial areaand nucleus raphe magnus-induced modulation ofnociceptive and nonnociceptive trigeminal subnucleuscaudalis neurons activated by cutaneous or deep inputs.J. Neurophysiol. 71, 2430–2445.
Conte, D., Legg, E.D., McCourt, A.C., Silajdzic, E., Nagy, G.G.,Maxwell, D.J., 2005. Transmitter content, origins andconnections of axons in the spinal cord that possess theserotonin (5-hydroxytryptamine) 3 receptor. Neuroscience134, 165–173.
Cornea-Hébert, V., Riad, M.,Wu, C., Singh, S.K., Descarries, L., 1999.Cellular and subcellular distribution of the serotonin 5-HT2Areceptor in the central nervous system of adult rat. J. Comp.Neurol. 409, 187–209.
Cropper, E.C., Eisenman, J.S., Azmitia, E.C., 1984.5-HT-Immunoreactive fibers in the trigeminalnuclear complex of the rat. Exp. Brain Res. 55, 515–522.
Davies, A.J., North, R.A., 2009. Electrophysiological andmorphological properties of neurons in the substantiagelatinosa of the mouse trigeminal subnucleuscaudalis. Pain 146, 214–221.
Doly, S., Fischer, J., Brisorgueil, M.J., Vergé, D., Conrath, M., 2004a.5-HT5A receptor localization in the rat spinal cord suggests a rolein nociception and control of pelvic floor musculature. J. Comp.Neurol. 476, 316–329.
Doly, S., Madeira, A., Fischer, J., Brisorgueil, M.J., Daval, G.,Bernard, R., Vergé, D., Conrath, M., 2004b. The 5-HT2Areceptor is widely distributed in the rat spinal cordand mainly localized at the plasma membrane ofpostsynaptic neurons. J. Comp. Neurol. 472, 496–511.
Doly, S., Fischer, J., Brisorgueil, M.J., Vergé, D., Conrath, M., 2005.Pre- and postsynaptic localization of the 5-HT7 receptor in ratdorsal spinal cord: immunocytochemical evidence. J. Comp.Neurol. 490, 256–269.
Fonseca, M.I., Ni, Y.G., Dunning, D.D., Miledi, R., 2001. Distributionof serotonin 2A, 2C and 3 receptor mRNA in spinal cord andmedulla oblongata. Brain Res. Mol. Brain Res. 89, 11–19.

100 B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
Gobel, S., 1978. Golgi studies of the neurons in layer II of the dorsalhorn of the medulla (trigeminal nucleus caudalis). J. Comp.Neurol. 180, 395–413.
Grudt, T.J., Perl, E.R., 2002. Correlations between neuronalmorphology and electrophysiological features in the rodentsuperficial dorsal horn. J. Physiol. 540, 189–207.
Grudt, T.J., Williams, J.T., Travagli, R.A., 1995. Inhibition by5-hydroxytryptamine and noradrenaline in substantiagelatinosa of guinea-pig spinal trigeminal nucleus. J. Physiol.485, 113–120.
Han, S.K., Park, J.R., Park, S.A., Chun, S.W., Lee, J.C., Lee, S.Y., Ryu, P.D., Park, S.J., 2007. Noradrenaline inhibits substantia gelatinosaneurons in mice trigeminal subnucleus caudalis via α2 and βadrenoceptors. Neurosci. Lett. 411, 92–97.
Hantman, A.W., Perl, E.R., 2005. Molecular and geneticfeatures of a labeled class of spinal substantiagelatinosa neurons in a transgenic mouse. J. Comp. Neurol.492, 90–100.
Hori, Y., Endo, K., Takahashi, T., 1996. Long-lasting synapticfacilitation induced by serotonin in superficial dorsal hornneurones of the rat spinal cord. J. Physiol. 492, 867–876.
Hoyer,D.,Hannon, J.P.,Martin,G.R., 2002.Molecular, pharmacologicaland functional diversity of 5-HT receptors. Pharmacol. Biochem.Behav. 71, 533–554.
Huang, J., Wang, Y.Y., Wang, W., Li, Y.Q., Tamamaki, N., Wu, S.X.,2008. 5-HT3A receptor subunit is expressed in a subpopulationof GABAergic and enkephalinergic neurons in the mousedorsal spinal cord. Neurosci. Lett. 441, 1–6.
Ito, A., Kumamoto, E., Takeda, M., Shibata, K., Sagai, H.,Yoshimura, M., 2000. Mechanisms for ovariectomy-inducedhyperalgesia and its relief by calcitonin: participation of5-HT1A-like receptor on C-afferent terminals in substantiagelatinosa of the rat spinal cord. J. Neurosci. 20, 6302–6308.
Jennings, E.A., Ryan, R.M., Christie, M.J., 2004. Effects of sumatriptanon rat medullary dorsal horn neurons. Pain 111, 30–37.
Jeong, C.Y., Choi, J.I., Yoon, M.H., 2004. Roles of serotonin receptorsubtypes for the antinociception of 5-HT in the spinal cord ofrats. Eur. J. Pharmacol. 502, 205–211.
Jiang,N., Furue, H., Katafuchi, T., Yoshimura,M., 2003. Somatostatindirectly inhibits substantia gelatinosaneurons in adult rat spinaldorsal horn in vitro. Neurosci. Res. 47, 97–107.
Kawamata, T., Omote, K., Toriyabe, M., Yamamoto, H., Namiki, A.,2003. The activation of 5-HT3 receptors evokes GABA release inthe spinal cord. Brain Res. 978, 250–255.
Kelly, E., Bailey, C.P., Henderson, G., 2008. Agonist-selectivemechanisms of GPCR desensitization. Br. J. Pharmacol. 153(Suppl. 1), S379–S388.
Landry, M., Bouali-Benazzouz, R., El Mestikawy, S., Ravassard, P.,Nagy, F., 2004. Expression of vesicular glutamate transportersin rat lumbar spinal cord, with a note on dorsal root ganglia. J.Comp. Neurol. 468, 380–394.
Lanfumey, L., Hamon, M., 2000. Central 5-HT1A receptors: regionaldistribution and functional characteristics. Nucl. Med. Biol. 27,429–435.
Li, J.L., Kaneko, T., Nomura, S., Li, Y.Q., Mizuno, N., 1997.Association of serotonin-like immunoreactive axons withnociceptive projection neurons in the caudal spinal trigeminalnucleus of the rat. J. Comp. Neurol. 384, 127–141.
Light, A.R., Kavookjian, A.M., 1988. Morphology andultrastructure of physiologically identified substantiagelatinosa (lamina II) neurons with axons that terminate indeeper dorsal horn laminae (III-V). J. Comp. Neurol. 267,172–189.
López Costa, J.J., Averill, S., Saavedra, J.P., Priestley, J.V., 1994.Serotonin innervation of enkephalin containing neurones inthe rat spinal trigeminal nucleus. Neurosci. Lett. 168, 167–171.
Lu, Y., Perl, E.R., 2005. Modular organization of excitatory circuitsbetween neurons of the spinal superficial dorsal horn (laminaeI and II). J. Neurosci. 25, 3900–3907.
Lu, Y., Perl, E.R., 2007. Selective action of noradrenaline andserotonin on neurones of the spinal superficial dorsal horn inthe rat. J. Physiol. 582, 127–136.
Marker, C.L., Luján, R., Loh, H.H., Wickman, K., 2005. SpinalG-protein-gated potassium channels contribute in adose-dependent manner to the analgesic effect of μ- and δ- butnot κ-opioids. J. Neurosci. 25, 3551–3559.
Marlier, L., Sandillon, F., Poulat, P., Rajaofetra, N., Geffard, M.,Privat, A., 1991. Serotonergic innervation of the dorsal hornof rat spinal cord: light and electron microscopicimmunocytochemical study. J. Neurocytol. 20, 310–322.
Millan, M.J., 2002. Descending control of pain. Prog. Neurobiol. 66,355–474.
Miquel, M.C., Emerit, M.B., Nosjean, A., Simon, A., Rumajogee, P.,Brisorgueil,M.J.,Doucet, E.,Hamon,M.,Vergé,D., 2002.Differentialsubcellular localization of the 5-HT3-As receptor subunit in the ratcentral nervous system. Eur. J. Neurosci. 15, 449–457.
Miyakawa, A., Furue, H., Katafuchi, T., Jiang, N., Yasaka, T., Kato,G., Yoshimura, M., 2005. Action of neuropeptide Y onnociceptive transmission in substantia gelatinosa of the adultrat spinal dorsal horn. Neuroscience 134, 595–604.
Molleman, A., 2003. Patch Clamping: An Introductory Guide toPatch Clamp Electrophysiology. John Wiley and Sons, WestSussex.
Morales,M., Battenberg, E., Bloom, F.E., 1998. Distribution of neuronsexpressing immunoreactivity for the 5HT3 receptor subtype inthe rat brain and spinal cord. J. Comp. Neurol. 402, 385–401.
Muraki, Y., Yamanaka, A., Tsujino, N., Kilduff, T.S., Goto, K.,Sakurai, T., 2004. Serotonergic regulation of the orexin/hypocretin neurons through the 5-HT1A receptor. J. Neurosci.24, 7159–7166.
Okamoto, K., Kimura, A., Donishi, T., Imbe, H., Senba, E., Tamai, Y.,2005. Central serotonin 3 receptors play an important role inthe modulation of nociceptive neural activity of trigeminalsubnucleus caudalis and nocifensive orofacial behavior in ratswith persistent temporomandibular joint inflammation.Neuroscience 135, 569–581.
Okamoto, K., Imbe, H., Kimura, A., Donishi, T., Tamai, Y., Senba, E.,2007. Activation of central 5HT2A receptors reduces thecraniofacial nociception of rats. Neuroscience 147, 1090–1102.
Pompeiano, M., Palacios, J.M., Mengod, G., 1992. Distribution andcellular localization of mRNA coding for 5-HT1A receptor in therat brain: correlation with receptor binding. J. Neurosci. 12,440–453.
Santos, S.F., Rebelo, S., Derkach, V.A., Safronov, B.V., 2007.Excitatory interneurons dominate sensory processing in thespinal substantia gelatinosa of rat. J. Physiol. 581, 241–254.
Seo, K., Fujiwara, N., Hu, J.W., Cairns, B.E., Someya, G., 2002.Intrathecal administration of 5-HT3 receptor agonistmodulates jaw muscle activity evoked by injection of mustardoil into the temporomandibular joint in the rat. Brain Res. 934,157–161.
Sessle, B.J., 2000. Acute and chronic craniofacial pain: brainstemmechanisms of nociceptive transmission and neuroplasticity,and their clinical correlates. Crit. Rev. Oral Biol. Med. 11, 57–91.
Suzuki, R., Rygh, L.J., Dickenson, A.H., 2004. Bad news from thebrain: descending 5-HT pathways that control spinal painprocessing. Trends Pharmacol. Sci. 25, 613–617.
Tamae, A., Nakatsuka, T., Koga, K., Kato, G., Furue, H., Katafuchi,T., Yoshimura, M., 2005. Direct inhibition of substantiagelatinosa neurones in the rat spinal cord by activation ofdopamine D2-like receptors. J. Physiol. 568, 243–253.
Tecott, L.H., Maricq, A.V., Julius, D., 1993. Nervous systemdistribution of the serotonin 5-HT3 receptor mRNA. Proc. Natl.Acad. Sci. USA 90, 1430–1434.
Todd, A.J., 2002. Anatomy of primary afferents and projectionneurones in the rat spinal dorsal horn with particularemphasis on substance P and the neurokinin 1 receptor. Exp.Physiol. 87, 245–249.

101B R A I N R E S E A R C H 1 3 6 8 ( 2 0 1 1 ) 9 1 – 1 0 1
Todd, A.J., Sullivan, A.C., 1990. Light microscope study of thecoexistence of GABA-like and glycine-like immunoreactivitiesin the spinal cord of the rat. J. Comp. Neurol. 296, 496–505.
Wang, Y.Y., Wei, Y.Y., Huang, J., Wang, W., Tamamaki, N., Li, Y.Q.,Wu, S.X., 2009. Expression patterns of 5-HT receptor subtypes1A and 2A on GABAergic neurons within the spinal dorsal hornof GAD67-GFP knock-in mice. J. Chem. Neuroanat. 38, 75–81.
Yoshimura, M., Furue, H., 2006. Mechanisms for theanti-nociceptive actions of the descending noradrenergic andserotonergic systems in the spinal cord. J. Pharmacol. Sci. 101,107–117.
Zhang, Y.Q., Gao, X., Ji, G.C., Huang, Y.L., Wu, G.C., Zhao, Z.Q., 2002.Expression of 5-HT1A receptor mRNA in rat lumbar spinal dorsalhorn neurons after peripheral inflammation. Pain 98, 287–295.
Copyright © 2022 FDOKUMEN




















