
Effect of water temperature and dietary starch on growth and metabolic utilization of diets in...
6
Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles A. Couto a,b , P. Enes a,b , H. Peres b , A. Oliva-Teles a,b, ⁎ a Departamento de Zoologia e Antropologia, Faculdade de Ciências, Universidade do Porto, 4099-002 Porto, Portugal b CIMAR/CIIMAR-Centro Interdisciplinar de Investigação Marinha e Ambiental, Universidade do Porto, 4050-123 Porto, Portugal ABSTRACT ARTICLE INFO Article history: Received 8 January 2008 Received in revised form 23 May 2008 Accepted 24 May 2008 Available online 30 June 2008 Keywords: Carbohydrate Enzyme activity Feed utilization Gilthead sea bream Growth performance Water temperature Whole-body composition We evaluated the effect of dietary starch level on growth performance, feed utilization, whole-body composition and activity of selected key enzymes of intermediary metabolism in gilthead sea bream juveniles reared at 18 and 25 °C. A diet was formulated to contain 48% crude protein,12% lipids and 30% gelatinized maize starch (diet 30GS). Two other diets were formulated to include the same level of ingredients as diet 30GS except for the gelatinized starch, which was included at 20% (diet 20GS) or 10% (diet 10GS). No adjustment to diet composition was otherwise made. Each diet was fed to triplicate groups of gilthead sea bream (30 g initial mass) for 8 weeks, on a pair-feeding scheme. The higher temperature improved growth performance but the opposite was true for feed efficiency and protein efficiency ratio. Independently of temperature, growth performance, feed efficiency and protein efficiency ratio were lower in fish fed diet 30GS. No effect of temperature or dietary starch level on whole-body composition was noticed. Hepatosomatic index and liver glycogen were higher at 18 °C and, within each temperature, in fish fed diet 30GS. Glycemia was not affected by temperature, but was lower in fish fed diet 10GS. Data on enzyme activities showed that increasing water temperature enhances liver glucokinase (GK) and pyruvate kinase (PK) activities, suggesting that gilthead sea bream is more apt to use dietary starch at higher temperatures. No effect of temperature was noticed on hexokinase (HK), fructose-1,6- bisphosphatase (FBPase), glucose-6-phosphate dehydrogenase (G6PD) and glutamate dehydrogenase (GDH) activities. Dietary starch enhanced PK and FBPase activities while depressed GDH activity, suggesting a lack of significant regulation of hepatic glucose utilization and production in this species. HK, GK and G6PD activities were unaffected by dietary composition. Irrespectively of water temperature, gelatinized starch may be included up to 20% in diets for gilthead sea bream juveniles; at higher dietary levels, growth and efficiency of feed utilization are depressed. © 2008 Elsevier Inc. All rights reserved. 1. Introduction Gilthead sea bream (Sparus aurata) is one of the most important marine fish species reared in the Mediterranean region. Gilthead sea bream is a carnivorous species with high protein requirement (Oliva- Teles, 2000). The protein used in fish diets is mainly obtained from fish meal, an overexploited resource which is becoming scarce (Watanabe, 2002); thus, protein stands amongst the most expensive components of fish diets. Optimization of protein utilization for growth by reduc- tion of its use for energy purposes is therefore a research priority not only from an economical but also from an environmental perspective, since protein catabolism by-products account for the major source of nitrogen loading to the surrounding waters (Cowey, 1995; Watanabe, 2002). Thus, it is a priority to optimise the protein to energy ratio of the diets and improve the protein sparing by non-protein energy sources such as lipids and carbohydrates (Wilson, 1994; Hemre et al., 2002). Carbohydrate is the least expensive form of dietary energy; how- ever its utilization by fish depends of several factors, such as fish species, dietary carbohydrate level, origin, molecular complexity and physical state (Wilson, 1994; Stone, 2003). As fish are poikilothermic animals, environmental temperature also affects their feed intake, growth and metabolic responses (Peres and Oliva-Teles, 1999). Although few studies are available regarding the utilization of carbohydrates at different temperatures, such studies indicate that temperature affects carbohydrate utilization (Médale et al., 1991, 1999; Shikata et al., 1995; Papoutsoglou and Lyndon, 2005; Enes et al., 2006b, 2008b; Moreira et al., 2008). Technological processing including ap- plication of moisture and heat, which promotes gelatinization, improves starch utilization (Kaushik and Oliva-Teles, 1985; Krogdahl et al., 2005). Gelatinization breaks down the starch granules increasing its surface area, which renders starch more susceptible to enzymatic attack (Bergot and Brèque, 1983; Stone, 2003). The higher digestibility Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50 ⁎ Corresponding author. Departamento de Zoologia e Antropologia, Faculdade de Ciências, Universidade do Porto, 4099-002 Porto, Portugal. Tel.: +351 22 340 1507; fax: +351 22 340 1511. E-mail address: [email protected] (A. Oliva-Teles). 1095-6433/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpa.2008.05.013 Contents lists available at ScienceDirect Comparative Biochemistry and Physiology, Part A journal homepage: www.elsevier.com/locate/cbpa
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Effect of water temperature and dietary starch on growth and metabolic utilization of diets in...

Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part A
j ourna l homepage: www.e lsev ie r.com/ locate /cbpa
Effect of water temperature and dietary starch on growth and metabolic utilization ofdiets in gilthead sea bream (Sparus aurata) juveniles
A. Couto a,b, P. Enes a,b, H. Peres b, A. Oliva-Teles a,b,⁎a Departamento de Zoologia e Antropologia, Faculdade de Ciências, Universidade do Porto, 4099-002 Porto, Portugalb CIMAR/CIIMAR-Centro Interdisciplinar de Investigação Marinha e Ambiental, Universidade do Porto, 4050-123 Porto, Portugal
⁎ Corresponding author. Departamento de ZoologiaCiências, Universidade do Porto, 4099-002 Porto, Portfax: +351 22 340 1511.
E-mail address: [email protected] (A. Oliva-Teles).
1095-6433/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.cbpa.2008.05.013
A B S T R A C T
A R T I C L E I N F OArticle history:
We evaluated the effect o Received 8 January 2008Received in revised form 23 May 2008Accepted 24 May 2008Available online 30 June 2008Keywords:CarbohydrateEnzyme activityFeed utilizationGilthead sea breamGrowth performanceWater temperatureWhole-body composition
f dietary starch level on growth performance, feed utilization, whole-bodycomposition and activity of selected key enzymes of intermediary metabolism in gilthead sea bream juvenilesreared at 18 and 25 °C. A dietwas formulated to contain 48% crude protein,12% lipids and 30% gelatinizedmaizestarch (diet 30GS). Twoother dietswere formulated to include the same level of ingredients as diet 30GS exceptfor the gelatinized starch, which was included at 20% (diet 20GS) or 10% (diet 10GS). No adjustment to dietcompositionwas otherwisemade. Each dietwas fed to triplicate groups of gilthead sea bream (30 g initialmass)for 8weeks, on a pair-feeding scheme. The higher temperature improved growth performance but the oppositewas true for feed efficiency and protein efficiency ratio. Independently of temperature, growth performance,feed efficiency and protein efficiency ratio were lower in fish fed diet 30GS. No effect of temperature or dietarystarch level on whole-body composition was noticed. Hepatosomatic index and liver glycogen were higher at18 °C and, within each temperature, in fish fed diet 30GS. Glycemia was not affected by temperature, but waslower infish fed diet 10GS. Data on enzyme activities showed that increasingwater temperature enhances liverglucokinase (GK) and pyruvate kinase (PK) activities, suggesting that gilthead sea bream is more apt to usedietary starch at higher temperatures. No effect of temperature was noticed on hexokinase (HK), fructose-1,6-bisphosphatase (FBPase), glucose-6-phosphate dehydrogenase (G6PD) and glutamate dehydrogenase (GDH)activities. Dietary starch enhanced PK and FBPase activities while depressed GDH activity, suggesting a lack ofsignificant regulation of hepatic glucose utilization and production in this species. HK, GK and G6PD activitieswere unaffected by dietary composition. Irrespectively of water temperature, gelatinized starch may beincluded up to 20% in diets for gilthead sea bream juveniles; at higher dietary levels, growth and efficiency offeed utilization are depressed.
© 2008 Elsevier Inc. All rights reserved.
1. Introduction
Gilthead sea bream (Sparus aurata) is one of the most importantmarine fish species reared in the Mediterranean region. Gilthead seabream is a carnivorous species with high protein requirement (Oliva-Teles, 2000). The protein used in fish diets is mainly obtained from fishmeal, an overexploited resource which is becoming scarce (Watanabe,2002); thus, protein stands amongst the most expensive componentsof fish diets. Optimization of protein utilization for growth by reduc-tion of its use for energy purposes is therefore a research priority notonly from an economical but also from an environmental perspective,since protein catabolism by-products account for the major source ofnitrogen loading to the surrounding waters (Cowey, 1995; Watanabe,
e Antropologia, Faculdade deugal. Tel.: +351 22 340 1507;
l rights reserved.
2002). Thus, it is a priority to optimise the protein to energy ratio of thediets and improve the protein sparing by non-protein energy sourcessuch as lipids and carbohydrates (Wilson, 1994; Hemre et al., 2002).
Carbohydrate is the least expensive form of dietary energy; how-ever its utilization by fish depends of several factors, such as fishspecies, dietary carbohydrate level, origin, molecular complexity andphysical state (Wilson, 1994; Stone, 2003). As fish are poikilothermicanimals, environmental temperature also affects their feed intake,growth and metabolic responses (Peres and Oliva-Teles, 1999).Although few studies are available regarding the utilization ofcarbohydrates at different temperatures, such studies indicate thattemperature affects carbohydrate utilization (Médale et al., 1991,1999;Shikata et al.,1995; Papoutsoglou and Lyndon, 2005; Enes et al., 2006b,2008b; Moreira et al., 2008). Technological processing including ap-plication of moisture and heat, which promotes gelatinization,improves starch utilization (Kaushik and Oliva-Teles, 1985; Krogdahlet al., 2005). Gelatinization breaks down the starch granules increasingits surface area, which renders starch more susceptible to enzymaticattack (Bergot and Brèque, 1983; Stone, 2003). The higher digestibility

46 A. Couto et al. / Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
of gelatinized starch compared to rawstarchhas been shown in severalspecies (Bergot and Brèque,1983; Gouveia et al.,1995; Peres and Oliva-Teles, 2002; Stone et al., 2003), including gilthead sea bream (Venouet al., 2003). Treated starch was previously shown to improve per-formance and provide some protein sparing in gilthead sea bream,when used up to 20% (Fernandez et al., 2007; Enes et al., 2008a).
One difficulty in interpreting nutrient utilization is thatmodifying adietary nutrient inclusion level generally implies a concomitant mod-ification of other nutrient levels, therefore confounding the inter-pretation of effect of the nutrient under study. With the present studywe aimed, by pair-feeding the animals diets identical in all nutrientsbut carbohydrate, to gain further information on the effect of car-bohydrate level (gelatinized maize starch) on growth performance,body composition and activity of key enzymes of intermediary metab-olism in gilthead sea bream juveniles. This study also aims to evaluatethe effect of water temperature on carbohydrate utilization, this waycontributing to further understanding the potential use of carbohy-drates as energy sources in gilthead sea bream reared at differenttemperatures.
2. Materials and methods
The experiment was carried out at the Marine Zoology Stationof Faculty of Sciences, at Porto University, with gilthead sea bream(S. aurata) juveniles obtained from a commercial hatchery.
Afishmeal based dietwas formulated to contain 48% crude protein,12% crude lipids and30%gelatinized starch (diet 30GS); twoother dietswere formulated to include the same levels of all ingredients as diet30GS except for the gelatinized starch, whichwas included at 20% (diet20GS) or 10% (diet 10GS). No adjustment to diet composition wasotherwise made. All dietary ingredients were finely ground, wellmixed and dry pelleted in a laboratory pelletmill (CPM) through 3mmdye. Ingredients and proximate composition of the experimental dietsare presented in Table 1.
Table 1Ingredient composition and proximate analysis of the experimental diets
Diets
30GS 20GS 10GS
Ingredients (% dry weight)Fish meala 59.8 64.5 74.6Soluble fish protein concentrateb 1.0 1.1 1.3Cod liver oil 5.7 6.3 7.1Gelatinized starchc 30.0 22.2 12.5Vitamin premixd 1.0 1.1 1.3Mineral premixe 1.0 1.1 1.3Choline chloride (50%) 0.5 0.6 0.6Binderf 1.0 1.1 1.3
Proximate analysis (% dry weight)Dry matter (DM) 95.10 92.91 92.82Crude protein 47.17 52.76 58.34Crude fat 11.68 13.37 14.73Starch 25.5 18.5 8.0Ash 11.83 13.25 14.90Gross energy (kJ g−1 DM) 20.40 20.90 21.39
a Vacuum Dried LT. Pesquera Diamante, S. A. Peru.b G-Special. Soropêche, France.c C-Gel Instant—12016, Cerestar, Mechelen, Belgium.d Vitamins (mg kg−1 diet): retinol, 18,000 (IU kg−1 diet); calciferol, 2000 (IU kg−1
diet); alpha tocopherol, 35; menadion sodium bis., 10; thiamin, 15; riboflavin, 25; Capantothenate, 50; nicotinic acid, 200; pyridoxine, 5; folic acid, 10; cyanocobalamin,0.02; biotin, 1.5; ascorbyl monophosphate, 50; inositol, 400.
e Minerals (mg kg−1 diet): cobalt sulphate, 1.91; copper sulphate, 19.6; iron sulphate,200; sodium fluoride, 2.21; potassium iodide, 0.78; magnesium oxide, 830; manganeseoxide, 26; sodium selenite, 0.66; zinc oxide, 37.5; potassium chloride, 1.15 (g kg−1 diet);sodium chloride, 0.40 (g kg−1 diet); dibasic calcium phosphate, 5.9 (g kg−1 diet).
f Aquacube (Guar gum, polymethyl carbamide, Manioc starch blend, hydrate calciumsulphate). Agil, England.
The experimental facilities consisted of two thermo-regulated re-circulation water systems each one equipped with 9 fiberglasscylindrical tanks of 250 L water capacity, supplied with a continuousflow of filtered seawater. During the trial, water temperature wasmaintained at 25±0.5 °C in one system and at 18±0.5 °C in the othersystem, and salinity averaged 35±1‰. After transport to the experi-mental facilities, fish were kept in quarantine for 2 weeks and fed acommercial diet. Then they were transferred to the experimental sys-tems and allowed to adapt to the experimental conditions for 5 days.Thereafter, groups of 25 fish with an initial mean body mass of 30 gwere randomly distributed to each tank. Dietswere randomlyassignedto triplicate groups of these fish. During the trial, fishwere fed by handtwo times a day, 6 days a week, on a pair-feeding scheme as follows:the group receiving diet 30GS was fed to apparent visual satiation,while the other groups received 90% (diet 20GS) and 80% (diet 10GS) ofthe ration given to group 30GS, so that every group was fed the sameamount of all nutrients except carbohydrate. Utmost carewas taken toassure that all feed supplied was consumed. The trial lasted 8 weeks;during this period two fish from different tanks died fromunidentifiedreasons. During the trial, the fish were bulk weighed every 2 weeks,after 1 day of feed deprivation. Ten fish from the initial stock pop-ulation and six fish from each tank at the end of the trial were sampledand pooled for whole-body composition analysis. Whole fish, visceraand liver mass of these fish were recorded for determination ofhepatosomatic and visceral indices. To minimize manipulation stress,the remaining fish continued to be fed for threemore days, afterwhichblood from three fish and liver from six fish per tank were sampled 6 hafter the morningmeal. Blood was collected from the caudal veinwitha heparinised syringe, immediately centrifuged and the plasma frozenfor analysis. Livers were frozen in liquid nitrogen and stored at −80 °C.
Chemical composition of diets and whole fish were analyzed usingthe following procedures: dry matter after drying in an oven at 105 °Cuntil constant weight; ash by incineration in amuffle furnace at 450 °Cfor 16 h; protein content (N×6.25) by the Kjeldahl method after aciddigestion using Kjeltec digestion and distillation units; lipid bypetroleum ether extraction (SoxTec HT System); starch accordingto Thivend et al. (1972); gross energy by direct combustion in anadiabatic bomb calorimeter (PARR model 1261). Hepatic glycogenwasdetermined as described by Plummer (1987) and plasma glucose wasdetermined according to the enzymatic–colorimetric method fromSpinreact (glucose kit, cod. 1001191).
For measurement of hexoquinase (HK; EC 2.7.1.1)/glucokinase (GK;EC 2.7.1.2) and L-typepyruvate kinase (PK; EC 2.7.1.40) activities, a frozensample of liver (200 mg) was homogenized (dilution 1/10) in ice-coldbuffer (80mMTris; 5 mMEDTA; 2 mMDTT; 1 mMbenzamidine; 1 mM4-(2-aminoethyl) benzenesulfonyl fluoride, pH 7.6). After centrifugation(900 g for 10 min), the resultant supernatant was separated for HK/GKand PK activity measurements. The HK (low Km HKs) and GK (high KmHK or HK IV) activities were measured at 37 °C using 0.5 mM and100 mM of glucose, respectively, by coupling ribulose-5-phosphateformation from glucose-6-phosphate to the reduction of β-NADP usingpurified glucose-6-phosphate dehydrogenase (Sigma) and 6-phospho-gluconate dehydrogenase (Sigma) as coupling enzymes (Tranulis et al.,1996; Panserat et al., 2000). The method for measuring GK activity onfrozen samples needed correction bymeasuring glucose dehydrogenase(EC 1.1.1.47) activity as described by Tranulis et al. (1996). Tomeasure PKactivity the previously separated supernatant was centrifuged at10000 g for 20 min, for isolation of the cytosolic fraction. Theconsumption of β-NADH was monitored at 340 nm and 37 °C, usinglactate dehydrogenase (Sigma) in excess as coupling enzyme (Foster andMoon, 1985).
The activity of fructose-1,6-bisphosphatase (FBPase; EC 3.1.3.11)was measured on a frozen sample of liver (200 mg), homogenized(dilution 1/10) in ice-cold buffer (20 mM Tris; 5 mM EDTA; 2 mMDTT;0.24 M sucrose, pH 8). The homogenate was centrifuged at 900 gfor 10 min. Enzyme assay was performed on the cytosolic fraction by

Table 2Growth performance, feed, nitrogen and energy utilization of sea bream fed theexperimental diets
Temperature 25 °C 18 °C
Diets 30GS 20GS 10GS SEM 30GS 20GS 10GS SEM
Initial body mass (g) 30.66 30.72 30.64 0.03 30.68 30.72 30.73 0.01Final body mass (g) 64.44 67.16 69.49 0.83 46.71 50.65 51.21 0.81Mass gain(g kg ABW§−1 day−1)
10.10 10.42 10.86 0.13 6.40 7.46 7.59 0.21
Feed intake(g kg ABW−1 day−1)
19.45 16.69 14.77 0.69 12.17 10.26 9.25 0.45
Protein intake(g fish−1 day−1)
0.54 0.55 0.55 0.00 0.25 0.26 0.26 0.00
Starch intake(g fish−1 day−1)
0.29 0.19 0.07 0.03 0.14 0.09 0.03 0.01
Feed efficiencya 0.52 0.62 0.74 0.03 0.52 0.73 0.82 0.04Protein efficiency ratiob 1.10 1.18 1.26 0.03 1.11 1.38 1.41 0.05Nitrogen retention(% N intake) c
17.57 19.11 21.45 0.70 18.29 22.31 24.01 0.98
Energy Retention(% E intake) d
21.59 24.16 29.65 1.28 20.84 28.61 32.48 1.77
Two-way ANOVA
Variation source Temperature Diet Interaction Diete
30GS 20GS 10GS
Initial body mass (g) ns ns ns – – –
Final body mass (g) ⁎⁎⁎ ⁎⁎⁎ ns a b bMass gain(g kg ABW−1 day−1)
⁎⁎⁎ ⁎⁎⁎ ns a b b
Feed intake(g kg ABW−1 day−1)
⁎⁎⁎ ⁎⁎⁎ ⁎ a b c
Protein intake(g fish−1)
⁎⁎⁎ ns ns – – –
Starch intake(g fish−1)
⁎⁎⁎ ⁎⁎⁎ ⁎⁎⁎ a b c
Feed efficiency ⁎⁎ ⁎⁎⁎ ⁎ a b cProtein efficiencyratio
⁎⁎ ⁎⁎⁎ ⁎ a b b
N Retention(% N intake)
⁎ ⁎⁎ ns a b b
E Retention(% N intake)
⁎ ⁎⁎⁎ ns a b c
SEM: pooled standard error of the mean.Ns—non significant; *Pb0.05; **Pb0.01; ***Pb0.001.
§ Average body mass (initial body mass+final body mass)/2.a FE: wet mass gain/dry feed intake.b PER: wet mass gain/crude protein intake.c N retention (%): (N gain/N intake)×100.d Energy retention (%): (energy gain/energy intake)×100.e Different letters in the same row stand for statistical differences between diets
Pb0.05.
Table 3Whole-body composition (% fresh weight), visceral and hepatosomatic indices andplasmatic glucose of sea bream fed the experimental diets
Temperature 25 °C 18 °C
Diets 30GS 20GS 10GS SEM 30GS 20GS 10GS SEM
Dry matter 31.28 30.48 30.36 0.21 30.83 30.70 30.92 0.30Crude protein 15.65 15.77 16.26 0.15 15.65 15.67 16.02 0.16Crude lipids 12.10 11.55 11.40 0.18 12.27 11.78 12.10 0.20Ash 3.69 4.00 3.57 0.08 3.92 3.70 3.72 0.09Gross energy (kJg−1) 8.30 7.99 8.24 0.06 8.24 8.20 8.28 0.05VIa 6.47 5.96 5.95 0.11 8.06 7.76 7.35 0.16HSIb 1.54 1.13 1.16 0.04 3.06 2.60 2.16 0.10Liver glycogen(g 100 g−1 liver)
5.17 3.45 2.07 0.35 12.05 7.77 6.04 0.59
Plasma glucose (mmol L−1) 6.71 6.10 3.44 0.37 6.87 6.53 4.19 0.37
Two-way ANOVA
Variation source Temperature Diet Interaction Dietc
30GS 20GS 10GS
Dry matter ns ns ns – – –
Crude protein ns ns ns – – –
Crude lipids ns ns ns – – –
Ash ns ns ns – – –
Crude energy(kJ g−1)
ns ns ns – – –
VI ⁎⁎⁎ ⁎ ns a ab bHSI ⁎⁎⁎ ⁎⁎⁎ ⁎ a b bLiver glycogen(g 100 g−1 liver)
⁎⁎⁎ ⁎⁎⁎ ⁎ a b c
Plasma glucose(mmol L−1)
ns ⁎⁎⁎ ns a a b
SEM: pooled standard error of the mean; ns—non significant; *Pb0.05; **Pb0.01;***Pb0.001.
a Visceral index: (viscera mass/body mass)×100.b Hepatosomatic index: (liver mass/body mass)×100.c Different letters in the same row stand for statistical differences between diets
Pb0.05.
47A. Couto et al. / Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
monitoring β-NADPH formation, using purified glucose-6-phosphatedehydrogenase (Sigma) and 6-phosphogluconate dehydrogenase(Sigma) as coupling enzymes (Tranulis et al., 1996).
To measure glucose-6-phosphate dehydrogenase (G6PD; EC1.1.1.49) activity, a frozen sample of liver (200 mg) was homogenized(dilution 1/5) in ice-cold buffer (0.02 M Tris; 0.25 M sucrose; 2 mMEDTA; 0.1 M NaF; 0.5 mM PMSF; 0.01 M β-mercapto-ethanol, pH 7.4).Homogenates were centrifuged at 30000 g for 20 min, and theformation of β-NADPH was monitored at 37 °C and 340 nm (Bautistaet al., 1988).
For measurement of glutamate dehydrogenase (GDH; EC 1.4.1.2)activity, a frozen sample of liver (200 mg) was homogenized (dilution1/10) in ice-cold buffer (30mMHEPES, 0.25mMsucrose, 0.5mMEDTA,5 mM K2HPO4, 1 mM DTT, pH 7.4). After centrifugation (900 g for10 min), the resultant supernatant was sonicated for 1 min (pulse 1 s,amplitude 50) and centrifuged again at 15000 g for 20 min. The GDHactivity was measured using 10 mM of L-glutamate, at 37 °C andfollowed at 340 nm (Bergemeyer, 1974).
Protein concentration was determined according to Bradford(1976) using Sigma protein assay kit with bovine serum albumin as
a standard. Enzymatic activities were expressed as mU per mg ofhepatic soluble protein (specific activity). One unit of enzyme activity(U) was defined as the amount of enzyme catalysing the hydrolysis of1 μmol of substrate per min under standard conditions (37 °C).
Statistical analysis of datawas doneby two-wayanalysis of varianceusing the significance level of 0.05 for rejection of the null hypothesis.Whenever statistical differenceswere found, a Turkey'smultiple rangetest was performed to identify differences amongmeans. All statisticalanalysis was done using a SPSS 15.0 software package.
3. Results
Final bodymass andmass gainwere higher in fish reared at 25 thanat 18 °C (Table 2). Within each water temperature these parameterswere lower in fish fed diet 30GS than the other diets. Feed intake waslower at the lowest temperature and differences in feed intake amonggroups were due to the pair-feeding scheme applied. Differences werefound in FE and PER between rearing temperatures, with higher valuesat 18 than at 25 °C, except for fish fed diet 30GS (Table 2). FE increasedwith the decrease of dietary starch level, while PER was only lower inthe 30GS group comparatively to the 20GS and 10GS groups. Retentionof nitrogen and energy (in % of intake) were higher at 18 than at 25 °Cand increased with the reduction of dietary starch level (Table 2).
At the end of the trial, fish exhibited similar whole-body com-position (Table 3). Visceral (VI) and hepatosomatic (HSI) indices werehigher at 18 than at 25 °C. VI of fish fed diet 30GS was higher than thatof fish fed diet 10GS, while HSI of fish fed diet 30GS was higher thanthat of groups fed diets 20GS and 10GS, which were similar amongstthem. Liver glycogenwas also higher at 18 than at 25 °C and decreasedwith the decrease of dietary starch level (Table 3). Glycemia was
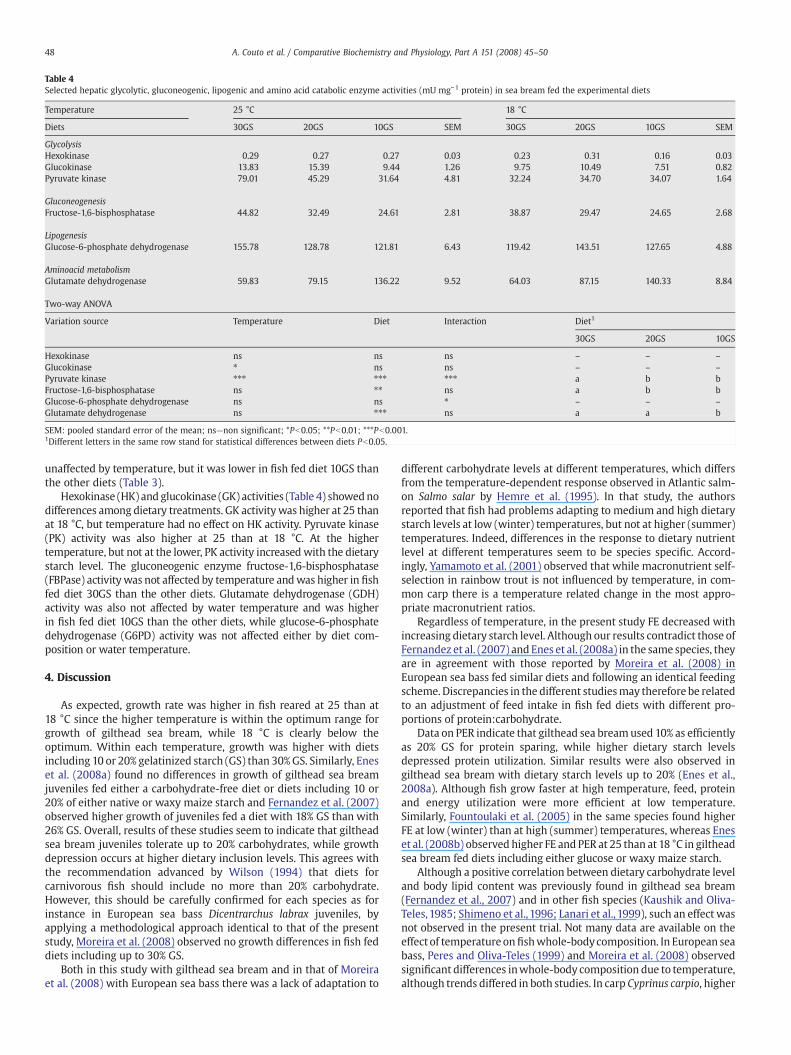
Table 4Selected hepatic glycolytic, gluconeogenic, lipogenic and amino acid catabolic enzyme activities (mU mg−1 protein) in sea bream fed the experimental diets
Temperature 25 °C 18 °C
Diets 30GS 20GS 10GS SEM 30GS 20GS 10GS SEM
GlycolysisHexokinase 0.29 0.27 0.27 0.03 0.23 0.31 0.16 0.03Glucokinase 13.83 15.39 9.44 1.26 9.75 10.49 7.51 0.82Pyruvate kinase 79.01 45.29 31.64 4.81 32.24 34.70 34.07 1.64
GluconeogenesisFructose-1,6-bisphosphatase 44.82 32.49 24.61 2.81 38.87 29.47 24.65 2.68
LipogenesisGlucose-6-phosphate dehydrogenase 155.78 128.78 121.81 6.43 119.42 143.51 127.65 4.88
Aminoacid metabolismGlutamate dehydrogenase 59.83 79.15 136.22 9.52 64.03 87.15 140.33 8.84
Two-way ANOVA
Variation source Temperature Diet Interaction Diet1
30GS 20GS 10GS
Hexokinase ns ns ns – – –
Glucokinase ⁎ ns ns – – –
Pyruvate kinase ⁎⁎⁎ ⁎⁎⁎ ⁎⁎⁎ a b bFructose-1,6-bisphosphatase ns ⁎⁎ ns a b bGlucose-6-phosphate dehydrogenase ns ns ⁎ – – –
Glutamate dehydrogenase ns ⁎⁎⁎ ns a a b
SEM: pooled standard error of the mean; ns—non significant; *Pb0.05; **Pb0.01; ***Pb0.001.1Different letters in the same row stand for statistical differences between diets Pb0.05.
48 A. Couto et al. / Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
unaffected by temperature, but it was lower in fish fed diet 10GS thanthe other diets (Table 3).
Hexokinase (HK) and glucokinase (GK) activities (Table 4) showednodifferences among dietary treatments. GK activitywas higher at 25 thanat 18 °C, but temperature had no effect on HK activity. Pyruvate kinase(PK) activity was also higher at 25 than at 18 °C. At the highertemperature, but not at the lower, PK activity increased with the dietarystarch level. The gluconeogenic enzyme fructose-1,6-bisphosphatase(FBPase) activity was not affected by temperature andwas higher in fishfed diet 30GS than the other diets. Glutamate dehydrogenase (GDH)activity was also not affected by water temperature and was higherin fish fed diet 10GS than the other diets, while glucose-6-phosphatedehydrogenase (G6PD) activity was not affected either by diet com-position or water temperature.
4. Discussion
As expected, growth rate was higher in fish reared at 25 than at18 °C since the higher temperature is within the optimum range forgrowth of gilthead sea bream, while 18 °C is clearly below theoptimum. Within each temperature, growth was higher with dietsincluding 10 or 20% gelatinized starch (GS) than 30%GS. Similarly, Eneset al. (2008a) found no differences in growth of gilthead sea breamjuveniles fed either a carbohydrate-free diet or diets including 10 or20% of either native or waxy maize starch and Fernandez et al. (2007)observed higher growth of juveniles fed a diet with 18% GS than with26% GS. Overall, results of these studies seem to indicate that giltheadsea bream juveniles tolerate up to 20% carbohydrates, while growthdepression occurs at higher dietary inclusion levels. This agrees withthe recommendation advanced by Wilson (1994) that diets forcarnivorous fish should include no more than 20% carbohydrate.However, this should be carefully confirmed for each species as forinstance in European sea bass Dicentrarchus labrax juveniles, byapplying a methodological approach identical to that of the presentstudy, Moreira et al. (2008) observed no growth differences in fish feddiets including up to 30% GS.
Both in this study with gilthead sea bream and in that of Moreiraet al. (2008) with European sea bass there was a lack of adaptation to
different carbohydrate levels at different temperatures, which differsfrom the temperature-dependent response observed in Atlantic salm-on Salmo salar by Hemre et al. (1995). In that study, the authorsreported that fish had problems adapting to medium and high dietarystarch levels at low (winter) temperatures, but not at higher (summer)temperatures. Indeed, differences in the response to dietary nutrientlevel at different temperatures seem to be species specific. Accord-ingly, Yamamoto et al. (2001) observed that while macronutrient self-selection in rainbow trout is not influenced by temperature, in com-mon carp there is a temperature related change in the most appro-priate macronutrient ratios.
Regardless of temperature, in the present study FE decreased withincreasing dietary starch level. Although our results contradict those ofFernandez et al. (2007) and Enes et al. (2008a) in the same species, theyare in agreement with those reported by Moreira et al. (2008) inEuropean sea bass fed similar diets and following an identical feedingscheme. Discrepancies in the different studiesmay therefore be relatedto an adjustment of feed intake in fish fed diets with different pro-portions of protein:carbohydrate.
Data on PER indicate that gilthead sea breamused 10% as efficientlyas 20% GS for protein sparing, while higher dietary starch levelsdepressed protein utilization. Similar results were also observed ingilthead sea bream with dietary starch levels up to 20% (Enes et al.,2008a). Although fish grow faster at high temperature, feed, proteinand energy utilization were more efficient at low temperature.Similarly, Fountoulaki et al. (2005) in the same species found higherFE at low (winter) than at high (summer) temperatures, whereas Eneset al. (2008b) observed higher FE and PER at 25 than at 18 °C in giltheadsea bream fed diets including either glucose or waxy maize starch.
Although a positive correlation between dietary carbohydrate leveland body lipid content was previously found in gilthead sea bream(Fernandez et al., 2007) and in other fish species (Kaushik and Oliva-Teles,1985; Shimeno et al., 1996; Lanari et al., 1999), such an effect wasnot observed in the present trial. Not many data are available on theeffect of temperature onfishwhole-bodycomposition. In European seabass, Peres and Oliva-Teles (1999) and Moreira et al. (2008) observedsignificant differences inwhole-body composition due to temperature,although trends differed in both studies. In carp Cyprinus carpio, higher

49A. Couto et al. / Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
body fat contentwas observed infish reared at 18 than at 25 °C (Médaleet al., 1999). Although no effect of water temperature on whole-bodycomposition was observed in our study, both lower temperature andhigh dietary starch level promoted higher visceral (VI) and hepatoso-matic (HSI) indices and liver glycogen deposition. Similar effects oftemperature (Enes et al., 2008b) and dietary carbohydrate level(Fernandez et al., 2007) were previously observed in this species. Theincrease in VI cannot be linked to a deposition of excess dietary energyas lipids in the viscera sincewhole-body lipid content does not supportthis argument. Instead, VI increase seems to be strongly correlated toliver glycogen deposition. The higher glycogen content and thus HSIobserved at lower water temperature may be a consequence of lowermetabolic rates at this temperature.
This study showed that glycemia was not affected by watertemperature even though feed intake was higher at the highest tem-perature. This agrees with the results of Enes et al. (2008b) also in thisspecies. Overall, these results suggest that temperature does not affectglucose regulation in gilthead sea bream juveniles. Fish generally havea limited ability to metabolize glucose, and high digestible carbohy-drate intake results in post-prandial hyperglycemia (Wilson, 1994;Peres et al., 1999). Accordingly, results of the present study showed ahigher glycemia in fish fed 30 and 20%GS than 10%GS. Indeed, 6 h afterfeeding, plasma glucose in the 10GS group was already near the basalglucose levels previously described for this species (Peres et al., 1999).
Our results cannot be explained by differences in starch digest-ibility. Although data are not presented, apparent digestibility of starchwas very high in all diets (N99%) and it was not affected by watertemperature. Results on GS digestibility are similar to those of Eneset al. (2008a) in the same species in fish fed up to 20% of raw or waxystarch. These results indicate that gilthead sea bream adapts, at thedigestibility level, to high levels of dietary starch, independently oftemperature. This lack of temperature effect on starchdigestibilitymaybe explained either by the lower feed intake, and hence lower starchintake at low temperatures, or by an adaptation of the carbohydraseactivity or expression at different temperatures; this needs to beelucidated in further studies.
At 25 °C, the enhanced PK activity levels due to the higher starchintake correlates to the glycemia levels observed in these groups. Suchinducible enzymatic response may also contribute to explain theincreased liver glycogen levels and HSI observed in fish fed the higherstarch level. Fernandez et al. (2007) in gilthead sea bream alsoreported higher PK activity in fish fed with high carbohydrate dietsand related these findings to a protein sparing effect by carbohydrates.However, this is not corroborated by the present data, as N retentiondid not improve with an increase of dietary carbohydrate level. On thecontrary, at high dietary starch level, there was even a decrease in Nretention. Contrary to PK, HK activity was unaffected by dietary starchlevel confirming that this enzyme is not under nutritional regulationas already observed in this species (Enes et al., 2008a,b) as well as inother species (Panserat et al., 2000; Enes et al., 2006a,b). Somehowunexpectedly, dietary starch level did not affect GK activity. Indeed,previous studies in fish, including gilthead sea bream (Panserat et al.,2000; Caseras et al., 2002; Enes et al., 2008a) showed an increase ofhepatic GK activity with the increase of dietary carbohydrates(Tranulis et al., 1996; Borrebaek and Christophersen, 2000; Panseratet al., 2000; Enes et al., 2006a).
Regarding temperature effect, GK and PK activities were higher inthewarm-acclimated fish, suggesting an enhanced glycolytic activityat a higher temperature. A reflection of these findings is the lowerglycogen deposition in liver of fish reared at the higher temperature.This is in line with the results of Hemre et al. (1995) in Atlanticsalmon who found a temperature-dependent glucose tolerance: fishtolerated and utilized higher levels of glucose at high temperatures.Results of the present study do not allow concluding if the higher GKand PK activities observed at 25 °C are related to a temperature effectper se or to the higher feed intake recorded at higher temperature.
However, very recently, Enes et al. (2008c) confirmed in Europeansea bass and gilthead sea bream that at least for GK such effect wasindeed do to temperature. In contrast to what was previously ob-served in gilthead sea bream (Enes et al., 2008b), present datashowed no effect of rearing temperature on HK activity. Thus, theeffect of temperature on the regulation of HK activity requires to befurther clarified.
Although Panserat et al. (2002) showed that in gilthead sea breamhepatic FBPase gene expression was depressed by feeding a 20%carbohydrate diet comparatively to a carbohydrate-free diet, our dataindicate that high levels of dietary starch (30%) promoted an increaseof FBPase activity. Furthermore, no differences in FBPase activity wereobserved in gilthead sea bream fed either a carbohydrate-free diet ordiets including 10 or 20% of either native or waxy maize starch (Eneset al., 2008a). Overall, these results sustain the hypothesis of a lack ofsignificant regulation of hepatic glucose utilization and production,which may contribute to explain the relatively low efficiency ofdietary glucose utilization by gilthead sea bream juveniles.
Excess hepatic dietary glucose is directed towards glycogensynthesis or lipogenesis. In the present study it seems that it wasmainly directed towards glycogen synthesis as the lipogenic enzymeG6PD activity was unaffected by dietary carbohydrate level. Moreover,data from whole-body composition showed no differences in whole-body lipid content among groups. On the contrary, other studiesperformed also with gilthead sea bream reported an increase of G6PDactivity as well as whole-body lipid content with increasing dietarycarbohydrate levels (Metón et al., 1999; Fernandez et al., 2007; Eneset al., 2008a). Irrespective of temperature, amino acid catabolismshowed a clear reduction with increasing dietary starch levels, asevidenced by GDH activities, suggesting a protein sparing by dietarycarbohydrates. However, these results conflict with those of whole-body N retention. Other studies performed on this very same speciesfurther observed a decrease of alanine aminotransferase (ALAT) ac-tivity with increasing dietary carbohydrate (Metón et al., 1999;Fernandez et al., 2007; Enes et al., 2008a).
In common carp, Shikata et al. (1995) observed that hepatopan-creatic enzyme activities were higher in cold-acclimated fish than inwarm-acclimated fish, suggesting a thermal compensation for lowacclimation temperature. The lack of differences due to temperature inactivities of FBPase, G6PD and GDH observed in the present study alsoseems to indicate an acclimation to low temperature in the activity ofthese enzymes, as also suggested by Ibarz et al. (2007). This howeverdid not occur with the glycolytic enzymes, GK and PK, which weremore active at the higher temperature.
Overall, results of the present study indicate that increasing watertemperature enhances liver glycolytic capacities, suggesting thatgilthead sea bream is more apt to use dietary starch at higher rearingtemperatures. Furthermore, dietary starch enhanced PK activitieswhile depressed protein catabolism. Gluconeogenesis was enhancedby high levels of dietary starch (30%) suggesting a lack of significantregulation of hepatic glucose synthesis in this species. Our datacorroborate previous results indicating that diets for gilthead seabream should not include more than 20% carbohydrate.
Acknowledgements
This workwas partially supported by the Fundação para a Ciência ea Tecnologia, Portugal (project POCI-CVT-57695-2004). The secondauthor was supported by a grant (BD/11414/2002) from Fundação paraa Ciência e a Tecnologia, Portugal. Wewould like to express our thanksto Mr. P. Correia for the assistance during the growth trial.
References
Bautista, J.M., Garrido-Pertierra, A., Soler, G., 1988. Glucose-6-phosphate dehydrogenasefrom Dicentrarchus labrax liver: kinetic mechanism and kinetics of NADPHinhibition. Biochim. Biophys. Acta 967, 354–363.

50 A. Couto et al. / Comparative Biochemistry and Physiology, Part A 151 (2008) 45–50
Bergemeyer, H.U., 1974. Methods of Enzymatic Analysis, vol. 4. Academic Press,New York, pp. 1704–1708.
Bergot, F., Brèque, J., 1983. Digestibility of starch by rainbow trout: effects of the physicalstate of starch and of the intake level. Aquaculture 34, 203–212.
Borrebaek, B., Christophersen, B., 2000. Hepatic glucose phosphorylating activities inperch (Perca fluviatilis) after different dietary treatments. Comp. Biochem. Physiol, B125, 387–393.
Bradford, M.M., 1976. A rapid and sensitive method for the quantification of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.72, 248–254.
Caseras, A., Metón, I., Vives, C., Egea, M., Fernandez, F., Baanante, I.V., 2002. Nutritionalregulation of glucose-6-phosphatase gene expression in liver of the gilthead seabream (Sparus aurata). Br. J. Nutr. 88, 607–614.
Cowey, C.B., 1995. Intermediary metabolism in fish with reference to output of endproducts of nitrogen and phosphorus. Water Sci. Technol. 31, 21–28.
Enes, P., Panserat, S., Kaushik, S., Oliva-Teles, A., 2006a. Effect of normal and waxy maizestarch on growth, food utilization and hepatic glucose metabolism in European seabass (Dicentrarchus labrax) juveniles. Comp. Biochem. Physiol. A 143, 89–96.
Enes, P., Panserat, S., Kaushik, S., Oliva-Teles, A., 2006b. Rapid metabolic adaptation inEuropean sea bass (Dicentrarchus labrax) juveniles fed different carbohydratesources after heat shock stress. Comp. Biochem. Physiol. A 145, 73–81.
Enes, P., Panserat, S., Kaushik, S., Oliva-Teles, A., 2008a. Growth performance andmetabolic utilization of diets with native and waxy maize starch by gilthead seabream (Sparus aurata) juveniles. Aquaculture 274, 101–108.
Enes, P., Panserat, S., Kaushik, S., Oliva-Teles, A., 2008b. Hepatic glucokinase andglucose-6-phosphatase responses to dietary glucose and starch in gilthead seabream (Sparus aurata) juveniles reared at two temperatures. Comp. Biochem.Physiol. A. 149, 80–86.
Enes, P., Panserat, S., Kaushik, S., Oliva-Teles, A., 2008c. Rearing temperature enhanceshepatic glucokinase but not glucose-6-phosphatase activities in European sea bass(Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) d with the same levelof glucose. Comp. Biochem. Physiol. A. doi:10.1016/j.cbpa.2008.04.596.
Fernandez, F., Miquel, A.G., Cordoba, M., Varas, M., Metón, I., Caseras, A., Baanante, I.V.,2007. Effects of diets with distinct protein-to-carbohydrate ratios on nutrientdigestibility, growth performance, body composition and liver intermediaryenzyme activities in gilthead sea bream (Sparus aurata, L.) fingerlings. J. Exp. Mar.Biol. Ecol. 343, 1–10.
Foster, G.D., Moon, T.W., 1985. Enzyme activities in Atlantic hagfish, Myxine glutinosa.Can. J. Zool. 64, 1080–1085.
Fountoulaki, E.E., Alexis, M.N., Nengas, I., 2005. Protein and energy requirements ofgilthead bream (Sparus aurata L.) fingerlings: preliminary results. Cah. OptionsMéditerr. 63, 19–26.
Gouveia, A., Oliva-Teles, A., Gomes, E., Peres, M.H., 1995. The effect of two dietary levelsof raw and gelatinized starch on growth and food utilization by the Europeanseabass. In: Castelló, I., Orvay, F., Calderer, I., Reig, A. (Eds.), Proc. of the Fifth NationalCongress on Aquaculture. Univ. Barcelona, Spain, pp. 516–521.
Hemre, G.I., Torrissen, O., Krogdahl, A., Lie, O.,1995. Glucose tolerance in Atlantic salmon,Salmo salar L., dependence on adaption to dietary starch and water temperature.Aquac. Nutr. 1, 69–75.
Hemre, G.I., Mommsen, T.P., Krogdahl, A., 2002. Carbohydrates in fish nutrition: effectson growth, glucose metabolism and hepatic enzymes. Aquac. Nutr. 8, 175–194.
Ibarz, A., Beltran, M., Fernandez-Borras, J., Gallardo, M.A., Sanchez, J., Blasco, J., 2007.Alterations in lipid metabolism and use of energy depots of gilthead sea bream(Sparus aurata) at low temperatures. Aquaculture 262, 470–480.
Kaushik, S.J., Oliva-Teles, A., 1985. Effect of digestible energy on nitrogen and energybalance in rainbow trout. Aquaculture 50, 89–101.
Krogdahl, A., Hemre, G.I., Mommsen, T.P., 2005. Carbohydrates in fish nutrition:digestion and absorption in postlarval stages. Aquac. Nutr. 11, 103–122.
Lanari, D., Poli, B.M., Ballestrazzi, R., Lupi, P., D'Agaro, E., Mecatti, M., 1999. The effects ofdietary fat and NFE levels on growing European sea bass (Dicentrarchus labrax L.).Growth rate, body and fillet composition, carcass traits and nutrient retentionefficiency. Aquaculture 179, 351–364.
Médale, F., Aguirre, P., Kaushik, S.J., 1991. Utilization of dietary carbohydrates by rainbowtrout at two water temperatures. In: Wenk, C., Boessinger, M. (Eds.), Proceedings ofthe XIIth symposium on energy metabolism of farm animals. Kartause Ittingen,Switzerland, pp. 392–395.
Médale, F., Poli, J.-M., Vallée, F., Blanc, D., 1999. Comparaison de l'utilisation digestive etmétabolique d'un régime riche en glucides par la carpe à 18 °C et 25 °C. Cybium 23,139–152.
Metón, I., Mediavilla, D., Caseras, A., Canto, E., Fernandez, F., Baanante, I.V.,1999. Effect ofdiet composition and ration size on key enzyme activities of glycolysis–gluconeogenesis, the pentose phosphate pathway and amino acid metabolism inliver of gilthead sea bream (Sparus aurata). Br. J. Nutr. 82, 223–232.
Moreira, I.S., Peres, H., Couto, A., Enes, P., Oliva-Teles, A., 2008. Temperature and dietarycarbohydrate level effects on performance and metabolic utilisation of diets inEuropean sea bass (Dicentrarchus labrax) juveniles. Aquaculture 274, 153–160.
Oliva-Teles, A., 2000. Recent advances in European sea bass and gilthead sea breamnutrition. Aquac. Int. 8, 477–492.
Panserat, S., Médale, F., Blin, C., Brèque, J., Vachot, C., Plagnes Juan, E., Gomes, E.,Krishnamoorthy, R., Kaushik, S., 2000. Hepatic glucokinase is induced by dietarycarbohydrates in rainbow trout, gilthead seabream, and common carp. Am. J. Physiol.278, R1164–R1170.
Panserat, S., Plagnes-Juan, E., Kaushik, S., 2002. Gluconeogenic enzyme gene expressionis decreased by dietary carbohydrates in common carp (Cyprinus carpio) andgilthead seabream (Sparus aurata). Biochim. Biophys. Acta 1579, 35–42.
Papoutsoglou, E.S., Lyndon, A.R., 2005. Effect of incubation temperature on carbohy-drate digestion in important teleosts for aquaculture. Aquac. Res. 36, 1252–1264.
Peres, H., Oliva-Teles, A., 1999. Influence of temperature on protein utilization injuvenile European seabass (Dicentrarchus labrax). Aquaculture 170, 337–348.
Peres, H., Oliva-Teles, A., 2002. Utilization of raw and gelatinized starch by European seabass (Dicentrarchus labrax) juveniles. Aquaculture 205, 287–299.
Peres, H., Goncalves, P., Oliva-Teles, A., 1999. Glucose tolerance in gilthead seabream(Sparus aurata) and European seabass (Dicentrarchus labrax). Aquaculture 179,415–423.
Plummer, P., 1987. Glycogen Determination in Animal Tissues. An Introduction toPractical Biochemistry, 3rd edn. McGraw Hill Book, Maidenhead. 332 pp.
Shikata, T., Iwanaga, S., Shimeno, S., 1995. Metabolic response to acclimation temperaturein carp. Fish. Sci. 61, 512–516.
Shimeno, S., Hosokawa, H., Takeda, M., 1996. Metabolic response of juvenile yellowtailto dietary carbohydrate to lipid ratios. Fish. Sci. 62, 945–949.
Stone, D.A.J., 2003. Dietary carbohydrate utilization by fish. Rev. Fish. Sci. 11, 337–369.Stone, D.A.J., Allan, G.L., Anderson, A.J., 2003. Carbohydrate utilization by juvenile silver
perch, Bidyanus bidyanus (Mitchell). III. The protein-sparing effect of wheat starch-based carbohydrates. Aquac. Res. 34, 123–134.
Thivend, P., Mercier, C., Guilbot, A., 1972. Determination of starch with glucoamylase. In:Whistler, R.L., Bemiller, J.N. (Eds.), Methods in Carbohydrate Chemistry. AcademicPress, New York, USA, pp. 100–105.
Tranulis, M.A., Dregni, O., Christophersen, B., Krogdahl, A., Borrebaek, B., 1996. Aglucokinase-like enzyme in the liver of Atlantic salmon (Salmo salar). Comp.Biochem. Physiol. B 114, 35–39.
Venou, B., Alexis, M.N., Fountoulaki, E., Nengas, I., Apostolopoulou, M., Castritsi-Cathariou, I., 2003. Effect of extrusion of wheat and corn on gilthead sea bream(Sparus aurata) growth, nutrient utilization efficiency, rates of gastric evacuationand digestive enzyme activities. Aquaculture 225, 207–223.
Watanabe, T., 2002. Strategies for further development of aquatic feeds. Fish. Sci. 68,242–252.
Wilson, R.P., 1994. Utilization of dietary carbohydrate by fish. Aquaculture 124, 67–80.Yamamoto, T., Shima, T., Furuita, H., Shiraishi, M., Sanchez Vazquez, F.J., Tabata, M., 2001.
Influence of decreasing water temperature and shortening of the light phase onmacronutrient self-selection by rainbow trout Oncorhynchus mykiss and commoncarl, Cyprinus carpio. Fish. Sci. 67, 420–429.




















