
Charles E. Ives: Two Works Explored - Eastern Illinois University

ARTICLE IN PRESS
0306-4565/$ - se
doi:10.1016/j.jth
�CorrespondAcuaticos, Dep
San Diego, CA
E-mail addr
Journal of Thermal Biology 31 (2006) 506–513
www.elsevier.com/locate/jtherbio
Effect of salinity on the thermoregulatory behavior of juvenile blueshrimp Litopenaeus stylirostris Stimpson
Ana Denisse Rea,�, Fernando Dıaza, Gustavo Valdezb
aLaboratorio de Ecofisiologıa de Organismos Acuaticos, Departamento de Biotecnologıa Marina,
Centro de Investigacion Cientıfica y de Educacion Superior de Ensenada (CICESE), Km. 107 Carretera Tijuana-Ensenada, Ensenada Baja California,
Mexico, MexicobDepartamento de Acuicultura, Centro de Investigacion Cientıfica y de Educacion Superior de Ensenada (CICESE), Km. 107 Carretera Tijuana-Ensenada,
Ensenada Baja California, Mexico, Mexico
Abstract
Preferred temperature (PT) of juveniles of Litopenaeus stylirostris was not modified (P40.05) by salinity. The final preferendum of
juveniles was 27.8 1C.
The critical thermal maxima (CTMax) determined at 42 combinations (6 temperatures� 7 salinities) in blue shrimp was not affected
significantly by salinity (P40.05). We obtained a direct relationship between the CTMax and the acclimation temperature.
The end point of CTMax in L. stylirostris was defined as the loss of righting response (LRR).
The acclimation response ratio (ARR) for the juveniles of blue shrimp had an interval of 0.45–0.50, values that agreed with others
obtained for crustaceans from tropical and sub tropical climates.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Salinity; Temperature; Preferred temperature; CTMax; ARR; Litopenaeus stylirostris
1. Introduction
The salinity and temperature that modify the physiolo-gical responses of aquatic organisms are factors thatdetermine the distribution and survival of animals in thelagoon–estuarine ecosystems. Salinity is a factor that canmodify several physiological responses such as the toler-ance, metabolism, growth, life of cycle, nutrition and theintra- and inter-specific relationships (Kinne, 1971; Fry,1971; Venkataramiah et al., 1974).
Temperature is a direct factor that affects the metabo-lism of the organisms at all stages of the life cycle. It canenhance physiological responses within the biokineticrange or can be lethal when the organisms are stressedthus reducing the survival rate.
e front matter r 2006 Elsevier Ltd. All rights reserved.
erbio.2006.05.004
ing author. Laboratorio de Ecofisiologıa de Organismos
artamento de Biotecnologıa Marina, P.O. Box 434844,
92143-4844, USA. Fax: +52646 175 05 69.
ess: [email protected] (A. Denisse Re).
From the physiological viewpoint, thermoregulatorybehavior may be viewed as homeostatic mechanism, whichtends to maintain internal temperatures favorable forphysiological processes (Reynolds, 1979). The preferenceand avoidance responses are inseparable aspects ofthermoregulatory behavior of the organisms and theymay be modified for internal factors, anthropogenicactivity, age and the organism’s size and some can changeaccording to the time of day (Giattina and Garton, 1982;Nichelmann, 1983).The temperature intervals in which the organism
congregate or spend the time are defined operationally aspreferred temperature (PT) or thermal preferenda (Rey-nolds and Casterlin, 1979b). Fry (1947) proposed twomethods to enable a precise determination namely acuteand gravitational preferendum. The acute, thermal pre-ference was usually determined after 2 h of the immersionof organisms in the gradient from the acclimationtemperature. In the gravitational method the final prefer-endum is determined only after the organisms had asufficient period of time to gravitate towards a region of

ARTICLE IN PRESSA. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513 507
thermal preference (Reynolds and Casterlin, 1979a; Jo-bling, 1981).
Some studies have been carried out on the thermo-regulatory behavior of crustacean species in the marineenvironment. Among the most relevant are those onHomarus americanus (Reynolds and Casterlin, 1979b;Crossin et al., 1998; Jury and Watson III, 2000),Palaemonetes vulgaris (Casterlin and Reynolds, 1979),Panulirus argus (Reynolds and Casterlin, 1979c), Farfante-
penaeus duorarum (Reynolds and Casterlin, 1979d), F.
chinensis (Miao and Tu, 1995), Penaeus monodon (Chenand Chen, 1999) Hemigrapsus nudus (McGaw, 2003) andPanulirus interruptus (Dıaz-Iglesias et al., 2004).
Close to the tolerance zone we found the resistance zonewhich is characterized by the interaction of the organism withtemperature and exposure time (Paladino et al., 1980), theupper limit of this zone is represented by the critical thermalmaxima (CTMax) which was introduced by Cowles andBogert (1944) and modified by Lowe and Vance (1955) andstandardized by Hutchison (1961). Both are defined by Cox(1974) as the arithmetic media of the thermal points in whichlocomotory movements become disorganized and the shrimploses the ability to escape from the conditions which mayultimately lead to its death. These are determined by changingthe temperature at constant rate from the acclimationtemperature and observing this behavior.
The knowledge of the CTMax provides a relevant andecological index, when applied to the blue shrimp in itshabitat. The shrimp in nature experiences such spatialtemperatures temporally, such as acute fluctuations, out-side their limit of tolerance. Lutterschmidt and Hutchison(1997a, b) reported that CTMax may occur at differenttemperatures in different species, but the physiologicalresponses are the same across a diversity of taxa.
Paladino et al. (1980) mentioned that in aquaculturepractices it is crucially important to know CTMax andCTMin as both are good indicators of the thermalresistance. Both methods are non-destructive and theyidentify the temperatures at which the first stress symptomsoccur. For this reasons the CTMax is an excellent tool toevaluate the requirements in the thermal physiology.
According to Claussen (1977) acclimation ratio response(ARR) is an index of the magnitude of the thermalacclimation of the organisms. Dıaz Herrera et al. (1998);Dıaz et al. (2002, 2004a, b), Perez et al. (2003) and Re et al.(2005) suggested that species inhabiting cold and warmregions have low values of ARR in comparison to thosefrom subtropical and tropical which have high values,indicating that the values are dependent of the geographiczone where the organisms occur.
In Mexico, some blue shrimp farms are located inNorthwest region where fluctuations of temperature andsalinity are common. The management response in theseconditions is not efficient. This affects the productivitywhen the organisms are affected in their physiology duringthe dry or wet season, thus exposing them to thermal andsalinity changes (Lester and Pante, 1992). The aquacultural
potential that the blue shrimp, L. stylirostris has in Mexicomakes it necessary to know the effect of different combina-tions of salinity and temperature on the PT, upper thermaltolerance and their acclimation response ratio (ARR), todetermine the adequate environmental conditions to optimizeits cultivation in tropical Pacific Ocean region.
2. Material and methods
The experimental shrimps of L. stylirostris (n ¼ 1100)provided from Sinaloa and Sonora farms both Northweststates of Mexico. They were maintained in three oval tanks(2500 l) with constant aeration, seawater 35% and atemperature of 2871 1C. Photoperiod was maintained12 h light 12 h dark. They were fed daily with two rations8% of their wet body weight of commercial shrimp foodRangens. The specimens were randomly separated in togroups that were used to determine PT and CTMax.After that they were placed in four circular tanks (500 l)
for 15 days where they were acclimated at 28 1C at fourdifferent experimental salinities (10%, 20%, 30% and40%) which were made by dilutions of filtered sea water of35% and declorinated tap water, the salinity was diluted ata rate 2% daily, until the test salinities were reached. The40% was obtained by the addition of Instant Ocean saltsand the salinities variation range from 71%. Salinity ineach tank was measured with a refractometer YSI 30. Thefood remained; the feces and molts were extracted dailyfrom the tanks by siphoning.The PT of the shrimps was determined by the gravitation
method described by Reynolds and Casterlin (1979a). Foreach experimental condition thirty specimens with the sameweight range, were labeled with individual tags consisted ofsmall numbered plastic circles (15mm of diameter),attached with cyano-acrylate adhesive (Crazy Glue) andintroduced in groups of ten to the horizontal thermalgradient. The thermal gradient was established in a 200mmdiameter tube over a length of 4m that contained 20different segments (Dıaz et al., 2006). The specimens whereintroduced to the zone that corresponded to the originalacclimation temperature. Each experiment was repeatedthree times with a total of 120 shrimps (n ¼ 120). Theorganisms were not fed for 24 h prior to testing to avoidinterference due to the calorigenic effect of diet (Nelsonand Hooper, 1982; Beamish and Trippel, 1990).The horizontal thermal gradient was created by a
1000W heater controlled by a Medusa regulator, at theleft side of trough and Neslab (model HX 150), at the rightside to the water. The combinations of this equipmentgenerated a stable and linear 10.0–33.0 1C tempera-ture gradient (y ¼ 6.87+1.44xRsqr ¼ 0.97); where x ¼
segments of the gradient and y ¼ temperature on thesegments of gradient, to avoid the thermal stratificationinto the gradient, 20 air stones were placed in bottom ofeach segment producing 5–9mg l�1 dissolved oxygen inwater column. The water exchange rate was 180–200mlmin�1. Observations of number of shrimp in each

ARTICLE IN PRESSA. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513508
of the 20 segments were recorded hourly over a 24 h period.Simultaneously, temperatures were recorded with digitalthermometers along the 20 segments. The light exposurewas controlled to a photoperiod of 12 light and 12 darkwith a photophase and a scotophase intensity range from0.35� 1015–0.02� 1015 quanta s�1 cm�2.
PT data of the blue shrimp were processed with theExploratory Data Analysis (Tukey, 1977), and they wereplotted as parallel boxes; a Kruskall–Wallis test wasemployed for determining if (PT) differences (Po0.05)occurred in repetitions. After this was confirmed, ananalysis of variance of ranks was used (Sigma Stat Version3.1) to determine the effect of salinity.
The CTMax of 840 organisms acclimated gradually atexperimental salinities of 10%, 15%, 20%, 25%, 30%, 35%and 40%. The experimental temperatures were 20, 23, 25, 28,30 and 3371 1C, which were maintained by 1000W heatersconnected to temperature regulator provided a thermocouple,the increase and decreased rate of temperature was 2 1C perday. The organisms remained in these experimental condi-tions for 21 days to stabilize their internal medium and allowmetabolic compensation (Castille and Lawrence, 1981; Rosaset al., 2001). The acclimation phase took place in 13 circulartanks (500 l). The feeding was maintained at 8% ratio ofweight wet body specimens.
Twenty shrimp of each experimental condition weretagged (circular plastic tag 15mm) and in groups of fivewere introduced into 40 l aquarium provided with 1000Wimmersion heater and permanent aeration to maintain auniform temperature. The water was maintained atexperimental temperature for 30min to reduce the stressproduced by handling (Perez et al., 2003). Determinationsof CTMax were done between 9:00 and 14:00 h. Theheating rate used was 1 1Cmin�1 (Lutterschmidt andHutchison, 1997a, b). The stress event registered was lossof righting response (LRR) when the shrimp was on itsback and could not recover its upright posture, or remainsreclined 901 (Nelson and Hooper, 1982). When the shrimpreached this point they were returned to their acclimationsalinity and temperature. The organisms were used onceonly and the data for the animals that did not recover afterreturning them to their acclimation salinity and tempera-ture after LRR were discarded.
In the blue shrimp, we determined (ARR) defined byClaussen (1977) as DCTMax/DT or the change in the CTMaxper degree change in acclimation temperature. A two-wayanalysis of variance was used as previous determination ofthe normality and homocedasticity of the data (Sigma StatVersion 3.1) to determine the effect of salinity andtemperature on the thermal tolerance of the blue shrimp.
3. Results
In juveniles of L. stylirostris exposed to light/dark cycleof 24 h the interval of PT was 26.071.3–28.471.3 1C.Differences in the PT in juvenile of blue shrimp exposedbetween day and night were not observed (P40.05). PT for
the blue shrimp juveniles were found was 26.871.1,27.972.0, 28.071.6 and 27.571.02 1C (median7confi-dence interval at 95%) for acclimation salinities of 10%,20%, 30% and 40% salinity did not modify the PT ofshrimp (P40.05). The final preferendum determined forthe blue shrimp was 27.871.44 1C (Fig. 1).The CTMax of the blue shrimp were not modified when
salinity was increased from 10–40% as acclimationtemperature raised from 20 to 33 1C the thermal toleranceof shrimp was increased by 3.2–8.4 1C (Table 1). Ananalysis of variance indicated that the temperature had asignificant effect (Po0.05) on the CTMax, but the effect ofsalinity and the interaction temperature–salinity was notsignificant (P40.05).The ARR of the blue shrimp exposed to different
combinations of salinity and temperature was 0.49, 0.47,0.50 and 0.45 for the seven salinity groups (Table 2).
4. Discussion
In juvenile of L. stylirostris no significant statisticaldifferences were found between the determinations of PTduring the diurnal and nocturnal phases, in all experi-mental salinities. Thus, blue shrimp did not present aday–night cycle in relation at the PT, a response reported inother aquatic organisms (Reynolds and Casterlin, 1979c;Amador, 2001; Dıaz et al., 2000; Valdez, 2002; MedinaRomo, 2002; Perez et al., 2003; Luna-Figueroa et al., 2003).These results do not agree with the bioenergetic hypothesisproposed by Brett (1971) that food is not a limited sourcein natural environment.In this study, no differences were found in the final
preferendum of L. stylirostris, acclimated to differentsalinities. Valdez (2002) for a study on L. vannamei
juveniles exposed to 20%, 26% and 32% reported similarresults in the PT, also in agreement with Lester and Pante(1992) who reported that thermal preference was notinfluenced by salinity in most of shrimp species.This study demonstrated that L. stylirostris exposed to
different salinities had a PT of 26.8–28 1C. Robertson et al.(1991) reported the effect of manipulation temperature onreproduction L. stylirostris had optimal temperature rangeof 27–29. This showed that physiological process areoptimal within the range of PT. Thermal experimentsconducted on other warm-water crustaceans showedsimilar PTs range: the prawn Palaemonetes kadiakensis
PTs between 29 and 31 1C (Nelson and Hooper, 1982).Dıaz Herrera et al. (1993) compared the thermoregulatorybehavior of postlarvae and juveniles of Macrobrachium
rosenbergii using acute and gravitational methods; foundthat prawns were grouped between 29 and 31 1C. A finalpreferendum for M. tenellum was found to be 28 1C byHernandez et al. (1995). Dıaz et al. (2002) was determinedthe final preferendum of M. acanthurus, a mexican prawn,acclimated to five temperatures was 29.5 1C. Finally, Dıazet al. (2004a) obtained a PT of 28 1C for Cherax
quadricarinatus.
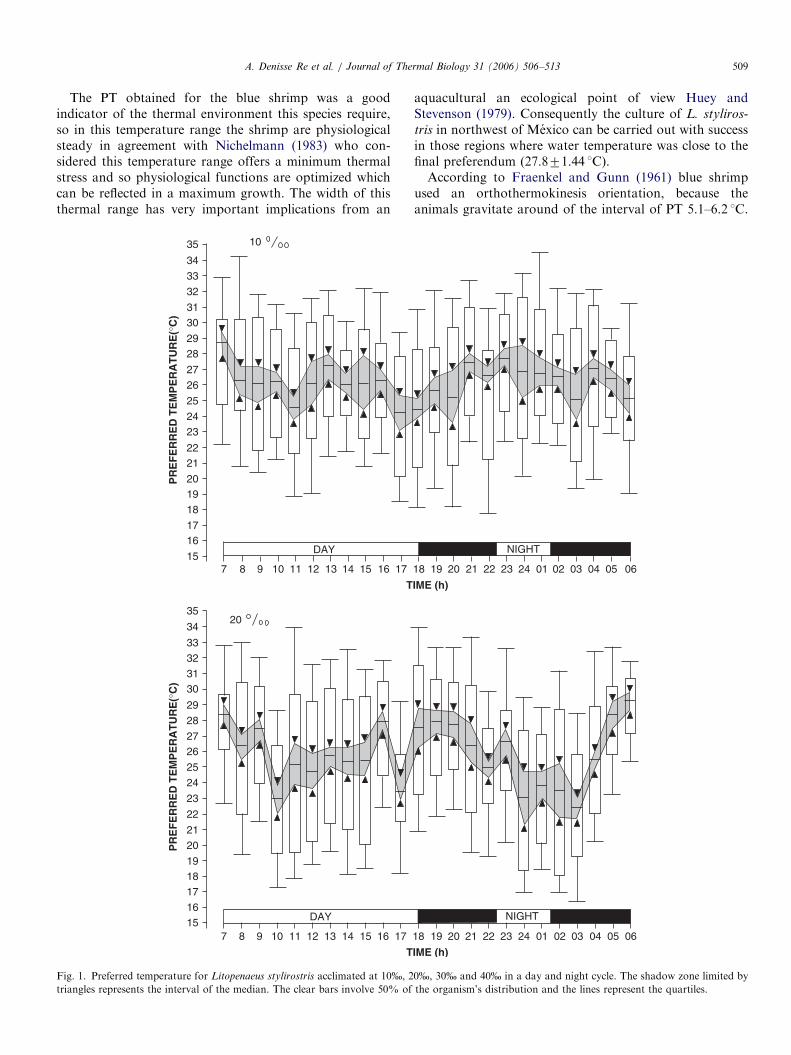
ARTICLE IN PRESSA. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513 509
The PT obtained for the blue shrimp was a goodindicator of the thermal environment this species require,so in this temperature range the shrimp are physiologicalsteady in agreement with Nichelmann (1983) who con-sidered this temperature range offers a minimum thermalstress and so physiological functions are optimized whichcan be reflected in a maximum growth. The width of thisthermal range has very important implications from an
7 8 9 10 11 12 13 14 15 16 1715
1716
18
1920
2122
2324
3031
32
33
34
25
26
29
27
28
35
T
DAY
PR
EF
ER
RE
D T
EM
PE
RA
TU
RE
(°C
)
7 8 9 10 11 12 13 14 15 16 1715
17
T
DAY
PR
EF
ER
RE
D T
EM
PE
RA
TU
RE
(°C
)
16
1819
20
21
22
23
24
3031
32
33
34
25
26
29
27
28
35
10
20
Fig. 1. Preferred temperature for Litopenaeus stylirostris acclimated at 10%, 2
triangles represents the interval of the median. The clear bars involve 50% of
aquacultural an ecological point of view Huey andStevenson (1979). Consequently the culture of L. styliros-
tris in northwest of Mexico can be carried out with successin those regions where water temperature was close to thefinal preferendum (27.871.44 1C).According to Fraenkel and Gunn (1961) blue shrimp
used an orthothermokinesis orientation, because theanimals gravitate around of the interval of PT 5.1–6.2 1C.
18 19 20 21 22 23 24 01 02 03 04 05 06
IME (h)
NIGHT
18 19 20 21 22 23 24 01 02 03 04 05 06
IME (h)
NIGHT
0%, 30% and 40% in a day and night cycle. The shadow zone limited by
the organism’s distribution and the lines represent the quartiles.
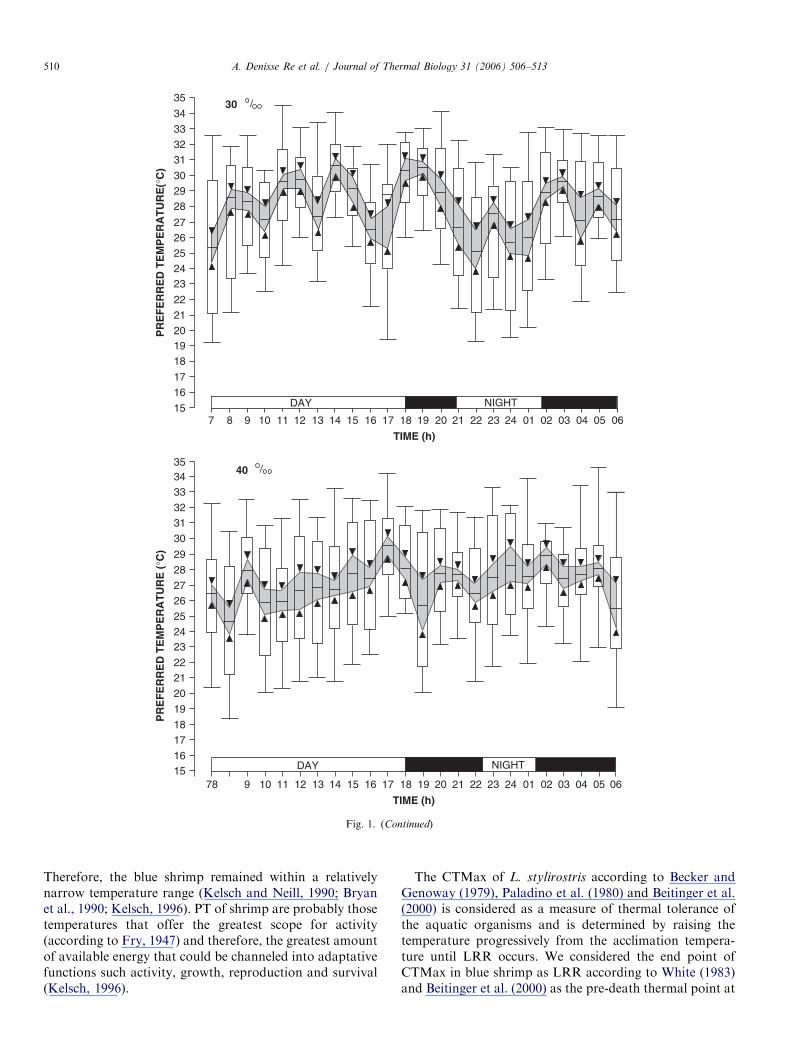
ARTICLE IN PRESS
15
17
20
24
27
29
31
33
7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 01 02 03 04 05 06
22
35
34
32
30
28
26
25
23
21
18
19
16
TIME (h)
DAY NIGHT
PR
EF
ER
RE
D T
EM
PE
RA
TU
RE
(°C
)
78 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 01 02 03 04 05 0615
17
16
18
19
20
21
22
2324
30
31
32
3334
25
26
29
27
28
35
TIME (h)
DAY NIGHT
PR
EF
ER
RE
D T
EM
PE
RA
TU
RE
(°C
)
/30
40 /
Fig. 1. (Continued)
A. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513510
Therefore, the blue shrimp remained within a relativelynarrow temperature range (Kelsch and Neill, 1990; Bryanet al., 1990; Kelsch, 1996). PT of shrimp are probably thosetemperatures that offer the greatest scope for activity(according to Fry, 1947) and therefore, the greatest amountof available energy that could be channeled into adaptativefunctions such activity, growth, reproduction and survival(Kelsch, 1996).
The CTMax of L. stylirostris according to Becker andGenoway (1979), Paladino et al. (1980) and Beitinger et al.(2000) is considered as a measure of thermal tolerance ofthe aquatic organisms and is determined by raising thetemperature progressively from the acclimation tempera-ture until LRR occurs. We considered the end point ofCTMax in blue shrimp as LRR according to White (1983)and Beitinger et al. (2000) as the pre-death thermal point at
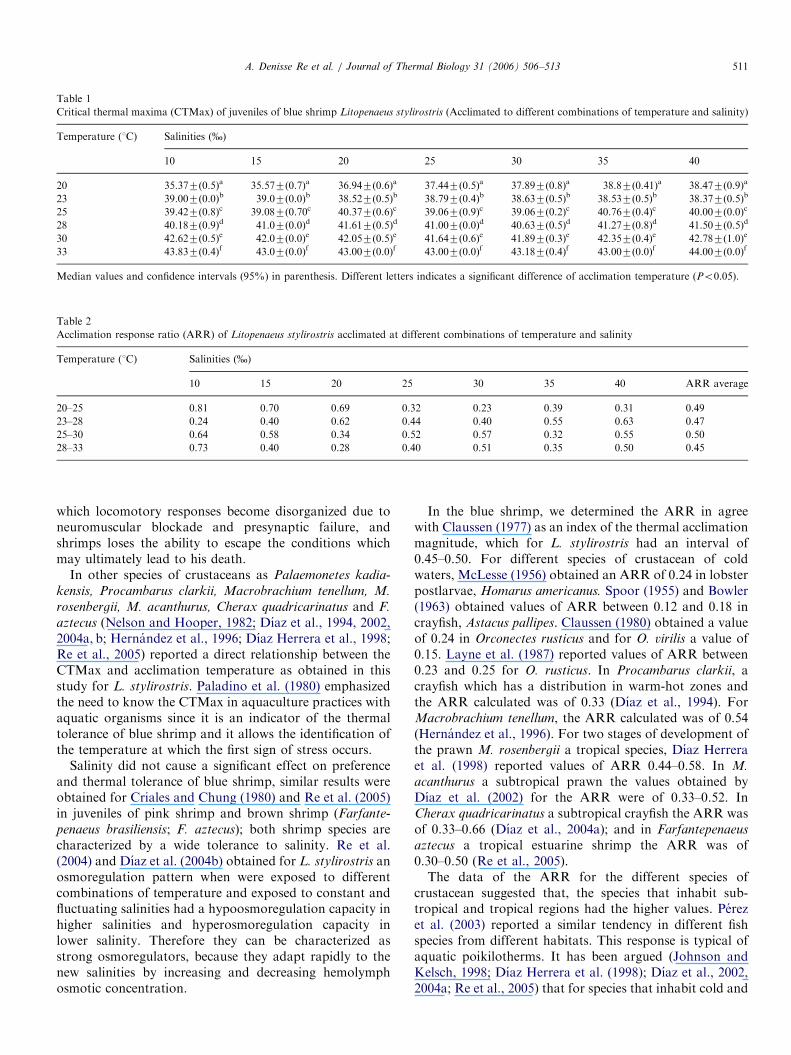
ARTICLE IN PRESS
Table 1
Critical thermal maxima (CTMax) of juveniles of blue shrimp Litopenaeus stylirostris (Acclimated to different combinations of temperature and salinity)
Temperature (1C) Salinities (%)
10 15 20 25 30 35 40
20 35.377(0.5)a 35.577(0.7)a 36.947(0.6)a 37.447(0.5)a 37.897(0.8)a 38.87(0.41)a 38.477(0.9)a
23 39.007(0.0)b 39.07(0.0)b 38.527(0.5)b 38.797(0.4)b 38.637(0.5)b 38.537(0.5)b 38.377(0.5)b
25 39.427(0.8)c 39.087(0.70c 40.377(0.6)c 39.067(0.9)c 39.067(0.2)c 40.767(0.4)c 40.007(0.0)c
28 40.187(0.9)d 41.07(0.0)d 41.617(0.5)d 41.007(0.0)d 40.637(0.5)d 41.277(0.8)d 41.507(0.5)d
30 42.627(0.5)e 42.07(0.0)e 42.057(0.5)e 41.647(0.6)e 41.897(0.3)e 42.357(0.4)e 42.787(1.0)e
33 43.837(0.4)f 43.07(0.0)f 43.007(0.0)f 43.007(0.0)f 43.187(0.4)f 43.007(0.0)f 44.007(0.0)f
Median values and confidence intervals (95%) in parenthesis. Different letters indicates a significant difference of acclimation temperature (Po0.05).
Table 2
Acclimation response ratio (ARR) of Litopenaeus stylirostris acclimated at different combinations of temperature and salinity
Temperature (1C) Salinities (%)
10 15 20 25 30 35 40 ARR average
20–25 0.81 0.70 0.69 0.32 0.23 0.39 0.31 0.49
23–28 0.24 0.40 0.62 0.44 0.40 0.55 0.63 0.47
25–30 0.64 0.58 0.34 0.52 0.57 0.32 0.55 0.50
28–33 0.73 0.40 0.28 0.40 0.51 0.35 0.50 0.45
A. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513 511
which locomotory responses become disorganized due toneuromuscular blockade and presynaptic failure, andshrimps loses the ability to escape the conditions whichmay ultimately lead to his death.
In other species of crustaceans as Palaemonetes kadia-
kensis, Procambarus clarkii, Macrobrachium tenellum, M.
rosenbergii, M. acanthurus, Cherax quadricarinatus and F.
aztecus (Nelson and Hooper, 1982; Dıaz et al., 1994, 2002,2004a, b; Hernandez et al., 1996; Dıaz Herrera et al., 1998;Re et al., 2005) reported a direct relationship between theCTMax and acclimation temperature as obtained in thisstudy for L. stylirostris. Paladino et al. (1980) emphasizedthe need to know the CTMax in aquaculture practices withaquatic organisms since it is an indicator of the thermaltolerance of blue shrimp and it allows the identification ofthe temperature at which the first sign of stress occurs.
Salinity did not cause a significant effect on preferenceand thermal tolerance of blue shrimp, similar results wereobtained for Criales and Chung (1980) and Re et al. (2005)in juveniles of pink shrimp and brown shrimp (Farfante-
penaeus brasiliensis; F. aztecus); both shrimp species arecharacterized by a wide tolerance to salinity. Re et al.(2004) and Dıaz et al. (2004b) obtained for L. stylirostris anosmoregulation pattern when were exposed to differentcombinations of temperature and exposed to constant andfluctuating salinities had a hypoosmoregulation capacity inhigher salinities and hyperosmoregulation capacity inlower salinity. Therefore they can be characterized asstrong osmoregulators, because they adapt rapidly to thenew salinities by increasing and decreasing hemolymphosmotic concentration.
In the blue shrimp, we determined the ARR in agreewith Claussen (1977) as an index of the thermal acclimationmagnitude, which for L. stylirostris had an interval of0.45–0.50. For different species of crustacean of coldwaters, McLesse (1956) obtained an ARR of 0.24 in lobsterpostlarvae, Homarus americanus. Spoor (1955) and Bowler(1963) obtained values of ARR between 0.12 and 0.18 incrayfish, Astacus pallipes. Claussen (1980) obtained a valueof 0.24 in Orconectes rusticus and for O. virilis a value of0.15. Layne et al. (1987) reported values of ARR between0.23 and 0.25 for O. rusticus. In Procambarus clarkii, acrayfish which has a distribution in warm-hot zones andthe ARR calculated was of 0.33 (Dıaz et al., 1994). ForMacrobrachium tenellum, the ARR calculated was of 0.54(Hernandez et al., 1996). For two stages of development ofthe prawn M. rosenbergii a tropical species, Dıaz Herreraet al. (1998) reported values of ARR 0.44–0.58. In M.
acanthurus a subtropical prawn the values obtained byDıaz et al. (2002) for the ARR were of 0.33–0.52. InCherax quadricarinatus a subtropical crayfish the ARR wasof 0.33–0.66 (Dıaz et al., 2004a); and in Farfantepenaeus
aztecus a tropical estuarine shrimp the ARR was of0.30–0.50 (Re et al., 2005).The data of the ARR for the different species of
crustacean suggested that, the species that inhabit sub-tropical and tropical regions had the higher values. Perezet al. (2003) reported a similar tendency in different fishspecies from different habitats. This response is typical ofaquatic poikilotherms. It has been argued (Johnson andKelsch, 1998; Dıaz Herrera et al. (1998); Dıaz et al., 2002,2004a; Re et al., 2005) that for species that inhabit cold and

ARTICLE IN PRESSA. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513512
temperate regions experiencing gradual long-term tem-perature fluctuations, would have the time necessary tomake metabolic adjustments, which result, in small in theirtolerance ranges. On the contrary, subtropical and tropicalspecies that experience the greatest fluctuations of tem-perature over short periods should have broad ranges oftolerance to survive the relatively rapid changes in watertemperature, without time for acclimation to adjust theirtolerance. Re et al. (2005) mentioned that higher ARRvalues might be found in species living in (or reproducingin lagoon–estuarine conditions).
Determination of thermal preference and tolerance in acommercial species such as blue shrimp is a good tool toenable the establishment of sites for culture based onregional temperatures that permits rapid the growth andlow mortality.
Acknowledgments
We thank SIMAC-CONACYT for financial support ofprojects 980111106004 and 200017509; and Jose M.Dominguez and Francisco Javier Ponce from the DrawingDepartment of CICESE for preparing the figures.
References
Amador, C.G., 2001. Termorregulacion de juveniles y adultos de la
langosta de quelas rojas Cherax quadricarinatus. M.Sc. Thesis,
CICESE.
Beamish, F.W.H., Trippel, E.A., 1990. Heat Increment: a static dynamic
dimension in bioenergetic models? Trans. Am. Fish. Soc. 119, 649–661.
Becker, C.D., Genoway, R.G., 1979. Evaluation of the critical thermal
maximum for determining thermal tolerance of freshwater fish.
Environ. Biol. Fish. 4, 245–256.
Beitinger, T.L., Bennett, W.A., McCauley, R.W., 2000. Temperature
tolerance of North American freshwater fishes exposed to dynamic
changes in temperature. Environ. Biol. Fish. 58, 237–275.
Bowler, K., 1963. A study of factors involved in acclimatization of
temperature and death at higher temperatures in Astacus pallipes, I:
experiments in intact animals. J. Cell. Comp. Physiol. 62, 119–132.
Brett, J.R., 1971. Energetic responses of salmon to temperature. A study
on some thermal relations in the physiology and freshwater ecology of
sockeye salmon Oncorhynchus nerka. Am. Zool. 11, 99–113.
Bryan, J.D., Kelsch, S.W., Neill, W.H., 1990. The maximum power
principle in behavioral thermoregulation by fishes. Trans. Am. Fish.
Soc. 119, 611–621.
Casterlin, M.E., Reynolds, W.W., 1979. Behavioral thermoregulation in
the grass shrimp Palaemonetes vulgaris (SAY). Rev. Can. Biol. 38,
45–46.
Castille Jr., F.J., Lawrence, A.L., 1981. The effect of salinity on the
osmotic, sodium and chloride concentrations in the hemolymph of
euryhaline shrimp of the genus Penaeus. Comp. Biochem. Physiol.
68A, 75–80.
Chen, J.C., Chen, J.N., 1999. Temperature preferendum of postlarval
black tiger shrimp Penaeus monodon. Mar. Freshwater Res. 50, 67–70.
Claussen, D.L., 1977. Thermal acclimation in ambystomatid salamanders.
Comp. Biochem. Physiol. 58A, 333–340.
Claussen, D.L., 1980. Thermal acclimation in the crayfish Orconectes
rusticus and O. virilis. Comp. Biochem. Physiol. 66A, 377–384.
Cowles, R.B., Bogert, C.M., 1944. A preliminary study of the thermal
requirements a desert reptile. Bull. Am. Mus. Nat. Hist. 83, 265–296.
Cox, D.K., 1974. Effect of three heating rates on the critical thermal
maximum of bluegill. In: Gibbons, J.W., Sharitz, R.R. (Eds.), Thermal
Ecology. AEC Symposium Series (Conf-73055), Springfield, VA, pp.
150–163.
Crossin, O.T., Al-Ayoub, S.A., Jury, S.H., Howell, W.H., Watson III,
W.H., 1998. Behavioral thermoregulation in the American lobster
Homarus americanus. J. Exp. Biol. 201, 365–374.
Criales, M.M., Chung, K.S., 1980. Tolerancia termica en postlarvas y
juveniles del camaron rosado Penaeus brasiliensis. Informes Museo del
Mar. 27, 1–15.
Dıaz, H.F., Espina, S., Buckle, R.L.F., 1994. Thermal stress responses of
Procambarus clarkii. Riv. Ital. Acquacol. 29, 149–154.
Dıaz, F., Del Rio-Portilla, M.A., Sierra, E., Aguilar, M., Re-Araujo,
A.D., 2000. Preferred temperature and critical thermal maxima of red
abalone Haliotis rufescens. J. Therm. Biol. 25, 257–261.
Dıaz, F., Sierra, E., Re, A.D., Rodrıguez, L., 2002. Behavioural
thermoregulation and critical thermal limits of Macrobrachium
acanthurus (Wiegman). J. Therm. Biol. 27, 423–428.
Dıaz, F., Re-Araujo, A., Sierra, E., Amador, G., 2004a. Behavioural
thermoregulation and critical limits applied to the culture of red claw
Cherax quadricarinatus (von Martens). Fresh Cray 14, 90–98.
Dıaz, F., Re-Araujo, A., Sierra, E., Dıaz Iglesias, E., 2004b. Effects of
temperature and salinity fluctuation on the oxygen consumption,
ammonium excretion and osmoregulation of the blue shrimp
Litopenaeus stylirostris (Stimpson). J. Shellfish Res. 23, 903–910.
Dıaz, F., Re Araujo, A., Medina, Z. Re, G., Valdez, G., Valenzuela, F.,
2006. Thermal preference and tolerance of green abalone Haliotis
fulgens (Philippi, 1845) and pink abalone Haliotis corrugata (Gray,
1828) Aqua. Res. 37, 877–884.
Dıaz Herrera, F., Gutierrez Morales, P., Garrido Mora, A., 1993.
Temperatura preferida y optima para el crecimiento de postlarvas y
juveniles de Macrobrachium rosenbergii (Crustacea:Palaemonidae).
Rev. Biol. Trop. 41, 153–155.
Dıaz Herrera, F., Sierra Uribe, E., Buckle Ramırez, L.F., Garrido Mora,
A., 1998. Critical thermal maxima and minima of Macrobrachium
rosenbergii (Decapoda:Palemonidae). J. Therm. Biol. 23, 381–385.
Dıaz-Iglesias, E., Dıaz, F., Re-Araujo, A.D., Baez Hidalgo, M., Lopez
Zenteno, M., Valdez Sanchez, G., Lopez Murillo, A.K., 2004.
Temperatura preferida y consumo de oxigeno circadiano de la
Langosta roja Panulirus interruptus (Randall, 1842). Cien. Mar. 30,
169–178.
Fraenkel, G.S., Gunn, D.L., 1961. The Orientation of Animals. Kineses,
Taxes and Compass Reactions. Dover Publications, New York,
376pp.
Fry, F.E.J., 1947. Effects of the environment on animal activity. Univ.
Toronto Stud. Biol. Ser. 55: Publ. Ontario Fish. Res. Lab. 68, 1–62.
Fry, F.E.J., 1971. The effect of environmental factors on the physiology of
fish. In: Hoar, W.S., Randall, D.J. (Eds.), Fish Physiology, vol. VII:
Environmental Relations and Behaviour. Academic Press, New York,
pp. 1–98.
Giattina, J.R., Garton, R.R., 1982. Graphical model of thermoregulatory
behavior by fishes with a new measure of eurythermality. Can. J. Fish.
Aquatic Sci. 39, 524–528.
Hernandez, R.M., Buckle, R.L.F., Dıaz, H.F., 1995. Preferred tempera-
ture of Macrobrachium tenellum (Crustacea, Palaemonidae). Riv. Ital.
Acquacol. 30, 93–96.
Hernandez, R.M., Buckle, R.L.F., Dıaz, F., 1996. Critical thermal
maximum of Macrobrachium tenellum. J. Therm. Biol. 21, 139–143.
Huey, R.B., Stevenson, R.D., 1979. Integrating thermal physiology and
ecology of ectotherms: a discussion of approaches. Am. Zool. 19,
357–366.
Hutchison, V.H., 1961. Critical thermal maxima in salamanders. Physiol.
Zool. 34, 92–125.
Jobling, M., 1981. Temperature tolerance and the final preferendum—
rapid methods for the assessment of optimum growth temperatures.
J. Fish. Biol. 19, 439–455.
Johnson, J.A., Kelsch, S.W., 1998. Effects of evolutionary thermal
environment on temperature preference relationship in fishes. Environ.
Biol. Fish. 53, 447–458.

ARTICLE IN PRESSA. Denisse Re et al. / Journal of Thermal Biology 31 (2006) 506–513 513
Jury, S.H., Watson III, W.H., 2000. Thermo sensitivity of the lobster,
Homarus americanus, as determined by cardiac assay. Biol. Bull. 199,
257–264.
Kelsch, S.W., 1996. Temperature selection and performance by bluegills:
evidence for selection in response in available power. Trans. Am. Fish.
Soc. 125, 948–955.
Kelsch, S.W., Neill, W.H., 1990. Temperature preference versus acclima-
tion in fishes: selection for changing metabolic optimum. Trans. Am.
Fish. Soc. 119, 601–610.
Kinne, O., 1971. Salinity: Animal Invertebrates. In: Kinne, O. (Ed.),
Marine Ecology, vol. I: Environmental Factors. Wiley Interscience,
London, pp. 821–995.
Layne, J.R., Claussen, D.L., Manis, M.L., 1987. Effects of acclimation
temperature, season and time of day on the critical thermal maxima
and minima of the crayfish Orconectes rusticus. J. Therm. Biol. 12,
183–187.
Lester, L.J., Pante, M.J.R., 1992. Penaeid temperature and salinity
responses. In: A. Fast, W., Lester, L.J. (Eds.), Marine Shrimp’s
Culture Principles and Practices. Elsevier, Amsterdam, pp. 515–534.
Lowe Jr., C.H., Vance, V.J., 1955. Acclimation of the critical thermal
maxima of the reptile Urosaurus ornatus. Science 122, 73–74.
Luna-Figueroa, J., Dıaz, F., Espina, S., 2003. Preferred temperature of the
mexican native cichlid Cichlasoma istlanum (Jordan and Snyder, 1899).
Hidrobiologıca 13, 271–275.
Lutterschmidt, W.I., Hutchison, V.M., 1997a. The critical thermal
maximum: data to support the onset of spasms the definitive end
point. Can. J. Zool. 75, 1553–1560.
Lutterschmidt, W.I., Hutchison, V.M., 1997b. The critical thermal
maximum: history and critique. Can. J. Zool. 75, 1561–1574.
McGaw, I.J., 2003. Behavioral thermoregulation in Hemigrapsus nudus,
the amphibious purple shore crab. Biol. Bull. 204, 38–49.
McLesse, D.W., 1956. Effects of temperature salinity and oxygen on
survival of the American lobster. J. Fish. Res. Bd. Canada 13, 247–272.
Medina Romo, Z., 2002. Temperatura optima de cultivo y temperatura
critica maxima del abulon azul Haliotis fulgens. B.Sc. Thesis, UABC.
Miao, S., Tu, S., 1995. Modeling thermal effect on growth of Chinese
shrimp, Penaeus chinensis (Osbeck). Ecol. Modell. 80, 187–196.
Nelson, D.H., Hooper, D.K., 1982. Thermal tolerance and preference of
the freshwater shrimp Palaemonetes kadiakensis. J. Therm. Biol. 7,
183–187.
Nichelmann, M., 1983. Some characteristics of the biological optimum
temperature. J. Therm. Biol. 39, 115–122.
Paladino, F.V., Spotila, J.R., Schubauer, J.P., Kowalski, K.T., 1980. The
critical thermal maximum: a technique used to elucidate physiological
stress and adaptation in fishes. Rev. Can. Biol. 39, 115–122.
Perez, E., Dıaz, F., Espina, S., 2003. Thermoregulatory behavior and
critical thermal limits of the angelfish Pterophyllum scalare (Lichten-
stein) (Pisces:Cichlidae). J. Therm. Biol. 28, 531–537.
Re, A.D., Dıaz, F., Sierra, E., Gomez-Jimenez, S., 2004. Consumo de
oxıgeno, excrecion de amonio y capacidad osmorreguladora de
Litopenaeus stylirostris (Stimpson) expuesto a diferentes combina-
ciones de temperatura y salinidad. Cien. Mar. 30, 443–453.
Re, A.D., Dıaz, F., Sierra, E., Rodriguez, J., 2005. Effect of salinity
and temperature on thermal tolerance of brown shrimp Farfantepe-
naeus aztecus (Ives) (Crustacea, Penaeidae). J. Therm. Biol. 30,
618–622.
Reynolds, W.W., 1979. Perspectives and introduction to the symposium:
thermoregulation in ectotherms. Am. Zool. 19, 193–194.
Reynolds, W.W., Casterlin, M.E., 1979a. Behavioral thermoregulation
and the ‘‘final preferendum’’ paradigm. Am. Zool. 19, 211–224.
Reynolds, W.W., Casterlin, M.E., 1979b. Behavioral thermoregulation
and activity in Homarus americanus. Comp. Biochem. Physiol. 64A,
25–28.
Reynolds, W.W., Casterlin, M.E., 1979c. Behavioral thermoregulation in
the spiny lobster Panulirus argus (Latreille). Hydrobiologia 66,
141–143.
Reynolds, W.W., Casterlin, M.E., 1979d. Thermoregulatory behavior of
the pink shrimp Penaeus duorarun Burkenroad. Hydrobiologia 67,
179–182.
Robertson, L., Bray, W., Lawrence, A., 1991. Reproductive response of
Penaeus stylirostris to temperature manipulation. J. World Aqua-
culture Soc. 22, 109–117.
Rosas, C., Lopez, N., Mercado, P., Martinez, E., 2001. Effect of salinity
acclimation on oxygen consumption of juvenile of the white shrimp
Litopenaeus vannamei. J. Crustacean Biol. 21, 912–922.
Spoor, W., 1995. Loss and gain of heat-tolerance by the crayfish. Biol.
Bull. 108, 77–87.
Tukey, J.W., 1977. Exploratory Data Analysis. Adisson-Wesley, Massa-
chusetts, 688pp.
Valdez-Sanchez, G., 2002. Determinacion de la preferencia termica y
fisiologıa energetica de juveniles de Litopenaeus vannamei aclimatados
a diferentes salinidades. M.Sc. Thesis, CICESE.
Venkataramiah, A., Lakshmi, G.J., Gunter, G., 1974. Studies on the
Effects of Salinity and Temperature on the Commercial Shrimp,
Penaeus aztecus Ives, with Special Regard to Survival Limits Growth,
Oxygen Consumption and Ionic Regulation. Gulf Coast Research
Laboratory, Ocean Springs, Mississippi, 134pp.
White, R., 1983. Effects of acute temperature change and acclimation
temperature on neuromuscular function and lethality in crayfish.
Physiol. Zool. 56, 174–194.
Copyright © 2022 FDOKUMEN
















