
EFFECT OF INCORPORATED COMMON SUNFLOWER ( Helianthus annuus L.) LEAVES ON THE GROWTH OF SELECTED...
26
1 EFFECT OF INCORPORATED COMMON SUNFLOWER ( Helianthus annuus L.) LEAVES ON THE GROWTH OF SELECTED WEEDS AND CROPS GROWN SOLELY OR IN COMBINATION Mohammad Cholid Indonesian Tobacco and Fibre Crops Research Institute ABSTRACT The study was conducted with the following objectives: to determine the effect of incorporating common sunflower leaves in the soil on the growth and development of itchgrass [Rottboellia. cochinchinensis (Lour)W.D. Clayton], barnyardgrass (Echinochloa glabrescens Munro ex Hook.f.), water purslane [Ludwigia octovalvis (Jacq.) Raven], mungbean (Vigna radiata L.), and rice (Oryza sativa L.). Treatments were replicated 3 times and laid out using randomized complete block design. Treatments were species of crops/weeds: 1) rice; 2) barnyardgrass; 3) water purslane; 4) rice + barnyardgrass; 5) rice + water purslan; 6) mungbean; 7) itchgrass ; and 8) mungbean + itchgrass. The Result indicated that incorporation of common sunflower leaves significantly inhibited the emergence of itchgrass, barnyardgrass, and water purslane, slightly inhibited that in rice, while there was no effect in mungbean. Plant height, shoot and root dry weight and grain yield of itchgrass and water purslane decreased significantly with incorporation of common sunflower leaves over those of untreated. The incorporated leaves significantly delayed days to flowering in most of the test species, except barnyardgrass. Incorporated common sunflower leaves in the soil had inhibitory potential on water purslane and itchgrass, but less or no inhibition on rice and mungbean. In contrast, common sunflower leaves may be used for effective itchgrass and water purslane controls, without damaging mungbean and rice plants. Key word: Allelopathic, Helianthus annuus L, crops, weeds ABSTRAK Pengaruh lama pencampuran residu daun bunga matahari (Helianthus annuus L.) dalam tanah pada pertumbuhan beberapa tanaman dan gulma monokultur dan tumpangsari. Penelitian ini bertujuan untuk mengevaluasi pengaruh lama pencampuran residu dari daun tanaman bunga matahari dengan tanah terhadap pertumbuhan Rottboellia cochinchinensis (Lour) W.D. Clayton, Echinochloa glabrescens Munro ex Hook.f., Ludwigia octovalvis (Jacq.) Raven, kacang hijau (Vigna radiata L.), dan padi (Oryza sativa L.). Rancangan Percobaan yang digunakan adalah Rancangan Acak Kelompok Faktorial, dengan 3 ulangan. Perlakuan meliputi: 1) padi; 2) Echinochloa glabrescens; 3) Ludwigia octovalvis; 4) padi + Echinochloa glabrescens; 5) padi + Ludwigia octovalvis; 6) kacang hijau; 7) itchgrass ; and 8) kacang hijau + itchgrass. Hasil percobaan menunjukkan bahwa pencampuran residu daun tanaman bunga matahari dapat menghambat perkecambahan gulma, sedikit menghambat pada padi, dan tidak menghambat pada kacang hijau. Tinggi tanaman, berat kering batang dan akar, serta produksi biji dari Ludwigia octovalvis dan Rottboellia cochinchinensis menurun secara nyata pada perlakuan residu dibanding dengan kontrol. Residu daun bunga matahari menunda saat pembungaan pada semua tanaman yang diuji, kecuali pada Echinochloa glabrescens. Hal
Transcript of EFFECT OF INCORPORATED COMMON SUNFLOWER ( Helianthus annuus L.) LEAVES ON THE GROWTH OF SELECTED...

1
EFFECT OF INCORPORATED COMMON SUNFLOWER ( Helianthus annuus L.) LEAVES ON THE GROWTH OF SELECTED WEEDS AND CROPS
GROWN SOLELY OR IN COMBINATION Mohammad Cholid
Indonesian Tobacco and Fibre Crops Research Institute
ABSTRACT
The study was conducted with the following objectives: to determine the effect of
incorporating common sunflower leaves in the soil on the growth and development of
itchgrass [Rottboellia. cochinchinensis (Lour)W.D. Clayton], barnyardgrass (Echinochloa
glabrescens Munro ex Hook.f.), water purslane [Ludwigia octovalvis (Jacq.) Raven],
mungbean (Vigna radiata L.), and rice (Oryza sativa L.). Treatments were replicated 3
times and laid out using randomized complete block design. Treatments were species of crops/weeds: 1) rice; 2) barnyardgrass; 3) water purslane; 4) rice + barnyardgrass; 5) rice
+ water purslan; 6) mungbean; 7) itchgrass ; and 8) mungbean + itchgrass. The Result
indicated that incorporation of common sunflower leaves significantly inhibited the
emergence of itchgrass, barnyardgrass, and water purslane, slightly inhibited that in rice,
while there was no effect in mungbean. Plant height, shoot and root dry weight and grain
yield of itchgrass and water purslane decreased significantly with incorporation of
common sunflower leaves over those of untreated. The incorporated leaves significantly
delayed days to flowering in most of the test species, except barnyardgrass. Incorporated
common sunflower leaves in the soil had inhibitory potential on water purslane and
itchgrass, but less or no inhibition on rice and mungbean. In contrast, common sunflower
leaves may be used for effective itchgrass and water purslane controls, without damaging
mungbean and rice plants. Key word: Allelopathic, Helianthus annuus L, crops, weeds
ABSTRAK
Pengaruh lama pencampuran residu daun bunga matahari (Helianthus annuus L.) dalam tanah pada pertumbuhan beberapa tanaman dan gulma
monokultur dan tumpangsari.
Penelitian ini bertujuan untuk mengevaluasi pengaruh lama pencampuran residu dari daun tanaman bunga matahari dengan tanah terhadap pertumbuhan Rottboellia cochinchinensis (Lour) W.D. Clayton, Echinochloa glabrescens Munro ex Hook.f., Ludwigia octovalvis (Jacq.) Raven, kacang hijau (Vigna radiata L.), dan padi (Oryza sativa L.). Rancangan Percobaan yang digunakan adalah Rancangan Acak Kelompok Faktorial, dengan 3 ulangan. Perlakuan meliputi: 1) padi; 2) Echinochloa glabrescens; 3) Ludwigia octovalvis; 4) padi + Echinochloa glabrescens; 5) padi + Ludwigia octovalvis; 6) kacang hijau; 7) itchgrass ; and 8) kacang hijau + itchgrass. Hasil percobaan menunjukkan bahwa pencampuran residu daun tanaman bunga matahari dapat menghambat perkecambahan gulma, sedikit menghambat pada padi, dan tidak menghambat pada kacang hijau. Tinggi tanaman, berat kering batang dan akar, serta produksi biji dari Ludwigia octovalvis dan Rottboellia cochinchinensis menurun secara nyata pada perlakuan residu dibanding dengan kontrol. Residu daun bunga matahari menunda saat pembungaan pada semua tanaman yang diuji, kecuali pada Echinochloa glabrescens. Hal

2
ini menunjukkan bahwa residu daun dari tanaman bunga matahari efektif untuk mengendalikan gulma Ludwigia octovalvis, Rottboellia. Cochinchinensis, dengan tanpa menimbulkan kerusakan pada tanaman kacang hijau, serta sedikit penghambatan pada tanaman padi. Kata kunci : Alelopati, Helianthus annuus L, tanaman, gulma
Introduction
Most nations of the world face serious environmental problems, a portion of
which evolves from a heavy reliance on herbicides and pesticides for agricultural
production. Efforts toward a more sustainable form of agriculture must inevitably take
into account the adaptive mechanisms that have allowed success of certain plants, and
these include the allelochemicals they produce.
Allelopathy, particularly of crops, can play an important role in weed
management, if suitably managed and applied. A study conducted by Wilson and Rice
(1968) showed that common sunflower, Helianthus annuus L., possessed allelopathic
properties. Germinating sunflower seed reduced germination rate, shoot and radicle
length of weeds and seedling fresh weight of Trianthema portulacastrum L, Parthenium
hysterophorus L. and Amaranthus viridis L. weeds (Dharamraj, 1998). Dried sunflower
residues and leaf powder incorporated in the soil reduced the growth of sorghum,
soybean and sunflower itself (Irons and Burnside, 1982).
Natural phytotoxic chemicals from allelopathic plants can be exploited as novel
agrochemicals for controlling harmful weeds (Batish et al., 2001). Such chemicals not
only provide the desired results but also pose no threat to environment and have more
target sites in contrast to commercial herbicides. This study takes into consideration the
allelopathic effect of soil incorporated residues on the germination and further seedling
growth of selected major weeds and major staple grain crops
MATERIALS AND METHODS
Experiments were conducted from September 2003 to Februay 2004, at the
Central Experiment Station, Department of Agronomy, U.P. Los Baños, Philippines.
Mature seeds of itchgrass (R. cochinchinensis (Lour) W.D. Clayton), barnyardgrass
(Echinochloa glabrescens Munro ex Hook.f.), water purslane (Ludwigia octovalvis (Jacq.)
Raven) were collected from farms in Los Baños. Common sunflower (Helianthus annuus

3
L.) and rice (Oryza sativa L.) var. PSB PC-74 seeds were obtained from the UPLB Central
Experiment Station, and mungbean (Vigna radiata L.) var. MG 50-10A seeds were
obtained from the Bureau of Plant Industry. All seeds were sun-dried and stored in a
closed container at Seed Storage prior to use.
Treatments were replicated 3 times and laid out using randomized complete block
design. Treatments were species of crops/weeds: 1) rice; 2) barnyardgrass; 3) water
purslane; 4) rice + barnyardgrass; 5) rice + water purslan; 6) mungbean; 7) itchgrass ;
8) mungbean + itchgrass. Lowland and upland soils collected from 0-15 cm depth from
a uniform location, at the Central Experiment Station, Department of Agronomy, U.P.
Los Baños, and each soil type was mixed thoroughly, and separately placed in 30 cm
diameter clay pots.
Preparation of plant residues, common sunflower plants grown at the UPLB
Central Experimental Station were harvested at the start of flowering. Common
sunflower leaves were cut and chopped into 2 to 3 cm pieces and uniformly
incorporated in respective pots. Concentration 1:16 (w/v) and 0 day duration (at
seeding) of incorporation of common sunflower leaves residues into soil were selected in
this study. One gram foliage residues for every 16 grams soil (1:16 w/w) was equivalent
to 312.5 g residues per 5000 g soil.
Foliage residues were incorporated in clay pots filled with soil, and 10 seeds of
test crops (rice and mungbean) and 20 seeds test weeds (itchgrass, barnyardgrass and
water purslane) were sown at the same time. Water was added to the pot to maintain the
soil at field capacity. The plants were kept under field condition at the Central
Experiment Station, Department of Agronomy, U.P. Los Baños, Philippines.
Seedlings were thinned to two per pot, and plants were watered as required.
Fertilizer at recommended rate for each crop was used. Fertilizer at 90-30-30 kg/ha of
N, P, K was applied in rice, barnyardgrass and water purslane pots. One third of N and
all P and K were applied basally at seeding. The remaining N was top-dressed at equal
splits at maximum tillering and panicle initiation stage of rice plant. While mungbean
and itchgrass pots were applied 20-40-40 kg/ha of N, P, K. All of fertilizer (N, P and K)
was applied basally at seeding.
4
Azodrin (3 hydroxy-N-methyl crotonamide) 19.8 EC (2.0 kg ai/ha) insecticide
was applied to mungbean and rice plants at 14 DAS and 42 DAS to control leaf rollers
and leaf folders. No significant insect damage was observed during the experimental
period.
Three soil samples were taken and leaf residues were incorporated at 0 days
before sowing. Composite randomly collected samples were analyzed for pH, OM, total
N, available P, exchangeable K, and texture at the Department of Soil Science, University
of the Philippines Los Baños. Chemical and physical characteristics of mixed soil-foliage
residues incorporation are shown in Table 1.
Initial germination was observed three days after sowing to 14 days after sowing.
The germination was recorded when the radicle or hypocotyls were 2 cm in length, and
converted to percentage. Plant height was measured at two weeks intervals from
germination to maturity. Shoot and root length of the crops and weeds were measured
at the end of the experiment. Plant materials were dried at 800C for 48 hours in an oven
after which the dry weight was determined.
The F test of the RCBD was used to determine the significance among treatments
using analysis of variance. The F test showed significant results which was further
tested using LSD to evaluate the differences among treatment means.
Table 1. Chemical and physical characteristics of lowland and upland soil mixed with leaf of common sunflower used in Experiment.
CONSTITUENT LOWLAND UPLAND
Control 0 Control 0
pH 6.2 7.7 7.0 7.9 OM(%) 4.98 8.16 1.43 2.16 N(%) 0.24 0.32 0.08 0.06 P(ppm) 31.00 95.00 51.00 75.00 K(cmol/kgsoil) 0.83 12.7 0.74 8.4 Sand 13 24 Silt 33 36 Clay 55 39 Textural grade clay clay loam
RESULTS AND DISCUSSION
Seed Germination
Germination of rice, barnyardgrass, water purslane, and itchgrass was reduced
by common sunflower leaves incorporated in the soil, while germination of mungbean
5
was not affected. There was no significant difference in seed germination among mono
and mixed cultures of all test species. Effect of common sunflower leaves on seed
germination of rice, mungbean, barnyardgrass, water purslane, and itchgrass are shown
in Table 2.
Leaves incorporated in the soil reduced the germination of rice seeds in both
mono and mixed cultures (Table 2). Germination of mungbean was not significantly
different among the treatments. This implies that mungbean was less sensitive crop
species to common sunflower leaves than rice. Germination of both rice and mungbean
was not affected by the culture (mono culture and mixed culture).
Table 2. Effect of incorporated common sunflower leaves on seed germination of rice, mungbean, itchgrass, barnyardgrass and water purslane in clay pots.
TEST PLANT GERMINATION (%)
Untreated Leaf Incorporation %Inhibition
Rice Rice 75.46a 61.46b 18.55 s Rice+Barnyardgrass 80.04a 61.46b 23.21 s Rice+W.purslane 75.78a 62.92b 16.97 s Barnyardgrass Barnyardgrass 75.00a 36.67b 51.11 s Rice+Barnyardgrass 86.67a 28.33b 67.31 s Water purslane Water purslane 67.03a 25.30b 62.26 s Rice+W.purslane 65.19a 22.60b 65.33 s Mungbean Mungbean 84.61a 71.35a 15.67 ns Mungbean+Itchgrass 82.11a 75.84a 7.64 ns Itchgrass Itchgrass 75.00a 33.33b 55.56 s Mungbean+Itchgrass 73.33a 28.33b 61.37 s
s = significant, ns = not significant Numbers followed by the same letters within test species are not significantly different at 5% LSD
Germination of the weed species (itchgrass, barnyardgrass and water purslane)
was significantly reduced by leaves incorporated in soil. Among the weed species, water
purslane showed the most inhibition, followed by barnyardgrass and itchgrass. This
indicated that water purslane was most sensitive to inhibitory substances present in
sunflower. These results are in agreement with earlier studies reported by Iron and
Burnside (1982) that weed seed germination and seedling growth was extract-species-
concentration-temperature dependent. Tongma et al. (1997) also reported that the
degree of inhibition on plant growth varied depending on plant species. Aqueous
6
extracts of root and aboveground parts of sunflower significantly reduced germination
rate and percentage in tomatoes. The degree of reduction in germination was less in
maize. There were no significant effects on the germination and growth of soybean
(Beltran et al., 1997).
Plant Height
The height of rice, mungbean and barnyardgrass at 56 DAS were not
significantly reduced by common sunflower leaves incorporated in the soil (Table 3).
However, the heights of itchgrass and water purslane were significantly reduced.
Table 3. Effect of incorporated common sunflower leaves on plant height of rice, mungbean, itchgrass, barnyardgrass and water purslane at 56 DAS.
TEST PLANT PLANT HEIGHT (cm)
Untreated Leaf Incorporation % Inhibition (-)/ Stimulation (+)
Rice Rice 76.78a 74.77ab -2.62 ns Rice+Barnyardgrass 64.00b 64.93ab +1.45 ns Rice+W.purslane 74.63ab 74.80ab +0.23 ns Barnyardgrass Barnyardgrass 73.08a 66.75a -8.66 ns Rice+Barnyardgrass 69.32a 65.50a -5.51 ns Water purslane Water purslane 52.15a 13.47c -74.17 s Rice+W.purslane 34.50b 9.67c -71.97 s Mungbean Mungbean 36.48a 34.70a -4.88 ns Mungbean+Itchgrass 29.58a 32.17a +8.76 ns Itchgrass Itchgrass 119.30a 47.73b -59.99 s Mungbean+Itchgrass 100.95a 50.50b -49.98 s
s = significant, ns = not significant Numbers followed by the same letters within test species are not significantly different at 1% LSD
There was even slight stimulation in plant height of mixed culture rice (+1.45%)
+ barnyardgrass and rice (+0.23%) + water purslane when leaves were incorporated in
the soil as compared to the untreated. Conversely, slight inhibition in plant height
occurred in mixed culture barnyardgrass (-5.51%) when leaf residues were incorporated
in the soil. The magnitude of inhibition increased in mixed culture water purslane (-
71.97%) with leaves incorporated in the soil. The observed reduction of rice plant height
7
in mixed culture rice + barnyardgrass compared to monoculture rice, may be due to
competition between rice and barnyardgrass.
Plant height of mungbean was not affected by common sunflower leaves in
mono and mixed culture with itchgrass. There was slight stimulation in plant height of
mixed culture mungbean when leaves were incorporated in the soil. Conversely,
inhibition of plant height in mixed culture itchgrass (-49.98%) with leaves incorporated
in the soil was observed. The significant reduction in itchgrass plant height in soil
incorporated with leaves over the untreated may be due to inhibitory substances present
in common sunflower leaves. Curtis and Coltam (1950) reported that sunflower
influences succeeding generations through decomposition and released of
allelochemicals from underground plant parts. Kaur (1998) also found that phytotoxic
compounds produced during sunflower biomass decomposition inhibited the
germination of pearl millet and greengram.
Rice plants were taller in the untreated than that when common sunflower leaves
were incorporated from 14 to 56 days after sowing (Figure 1).
0.00
20.00
40.00
60.00
80.00
100.00
120.00
140.00
14 28 42 56 70 84 98
Days after seeding
Pla
nt
he
igh
t (c
m)
Rice (C) Barnyardgrass (C)Rice/R+B (C) Barnyardgrass/R+B (C)Rice (R) Barnyardgrass (R)Rice/R+B (R) Barnyardgrass/R+B (R)
C = Untreated, R = Leaf incorporation, R+B = Mixed culture rice + barnyardgrass
Figure 1. Plant height of rice and barnyardgrass in mono and mixed cultures
from 14 DAS until 98 DAS.
There was no significant difference in rice plant height at 70 DAS to 98 DAS. This can be
attributed to the fact that common sunflower leaves may have different effects,
depending on the stages of residue decomposition and stage of growth and

8
development of test plants. Inhibition effects of leaves of common sunflower on rice
growth were observed at the initial decomposition of leaves and decreased through
time.
From 84 to 98 DAS, rice plant height was constant. This may be due to the
transformation of the rice from vegetative to reproductive phase. Rice flowered at 72 to
84 days after seeding (Table 4). The height of rice plant was greater in the monoculture
than in mixed culture rice with barnyardgrass both in untreated plant and in the
treatment where leaves were incorporated. This may be attributed to the inter-specific
competition between rice and barnyardgrass. The same trend was shown in
barnyardgrass, where the height of barnyardgrass was greater in untreated compared to
the treated where leaves were incorporated in both mono and mixed culture.
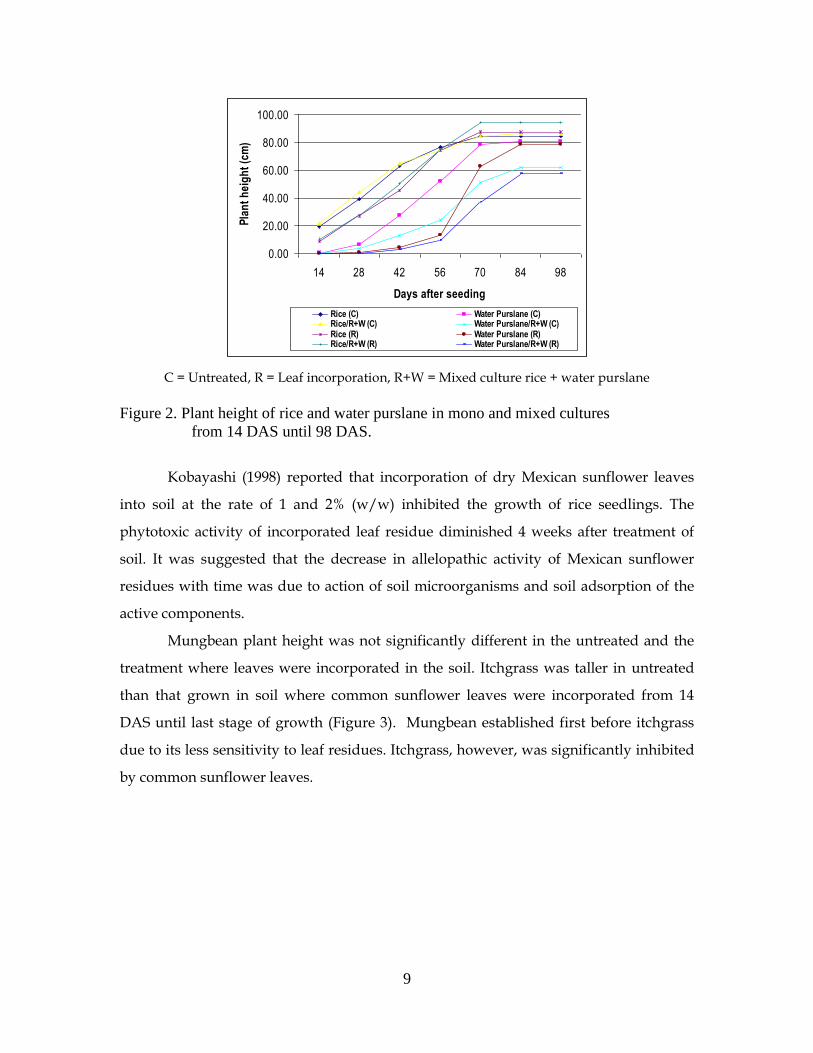
Water purslane plants were taller in the untreated than in those where leaves
were incorporated from 14 DAS to 70 DAS (Figure 2). The same trend was observed in
the mono and mixed cultures. From 84 to 98 DAS there were no significant differences in
height observed. This result collaborates the finding of Narwal et al. (1999b) who
reported that the phytotoxicity of sunflower biomass persisted in the soil up to 9 weeks
after soil incorporation. The harmful allelochemicals might have either decomposed by
soil microflora or adsorbed in soil complexes. The decomposition rate of toxic
substances was fastest in the beginning of incubation and decreased with time and
become constant at later stages. Sunflower residues incorporated in the soil were most
harmful to the following crop up to 30 DAS, moderate up to 60 DAS and least at 90 DAS.
Most of the allelochemicals might have been released during the fast rate of residue
decomposition in the early stage of incorporation.
Tongma et al. (1997) reported that water extract of Mexican sunflower [Tithonia
diversifolia (Hemsl.) A. Gray] applied in soil inhibited root growth of the plant tested but
this inhibitory activity declined as it stayed longer in the soil. Kitou and Yoshida (1998)
also observed that lettuce germination and radicle elongation were inhibited by soil
extracts amended with leaves of Solidago altissima during early period of incubation,
although the inhibitory effects disappeared with an increase in the duration of
incubation period.
9
0.00
20.00
40.00
60.00
80.00
100.00
14 28 42 56 70 84 98
Days after seeding
Pla
nt
he
igh
t (c
m)
Rice (C) Water Purslane (C)Rice/R+W (C) Water Purslane/R+W (C)Rice (R) Water Purslane (R)Rice/R+W (R) Water Purslane/R+W (R)
C = Untreated, R = Leaf incorporation, R+W = Mixed culture rice + water purslane
Figure 2. Plant height of rice and water purslane in mono and mixed cultures
from 14 DAS until 98 DAS.
Kobayashi (1998) reported that incorporation of dry Mexican sunflower leaves
into soil at the rate of 1 and 2% (w/w) inhibited the growth of rice seedlings. The
phytotoxic activity of incorporated leaf residue diminished 4 weeks after treatment of
soil. It was suggested that the decrease in allelopathic activity of Mexican sunflower
residues with time was due to action of soil microorganisms and soil adsorption of the
active components.
Mungbean plant height was not significantly different in the untreated and the
treatment where leaves were incorporated in the soil. Itchgrass was taller in untreated
than that grown in soil where common sunflower leaves were incorporated from 14
DAS until last stage of growth (Figure 3). Mungbean established first before itchgrass
due to its less sensitivity to leaf residues. Itchgrass, however, was significantly inhibited
by common sunflower leaves.

10
0.00
20.00
40.00
60.00
80.00
100.00
120.00
140.00
14 28 42 56 70 84 98
Days after seeding
Pla
nt
he
igh
t (c
m)
Mungbean (C) Itchgrass (C) Mungbean/M+I (C) Itchgrass/M+I (C)
Mungbean (R) Itchgrass (R) Mungbean/M+I (R) Itchgrass/M+I (R)
C = Untreated, R = Leaf incorporation, M+I = Mixed culture mungbean + itchgrass
Figure 3. Plant height of mungbean and itchgrass in mono and mixed cultures
from 14 DAS until 98 DAS.
The inhibitory effect of common sunflower leaves may be due to the presence of
allelochemicals. Several workers (Curtis and Coltam, 1950; Rice, 1983; Leather, 1983;
Narwal et al., 1999a) reported that sunflower influences succeeding crops through
decaying and exudates from above and underground plant parts. Rice (1983) and
Leather (1983) also reported that sunflower can have negative effects on successive crops
due to its strong allelopathic effect. Phenolics were thought to be responsible for such
behavior (Einhellig, 1986).
Days to Flowering
Days to flowering of most test plants were affected by incorporated leaves of
common sunflower. Table 4 presents the effect of the leaves on days to flowering of the
test species.
As indicated in Table 4, days to flowering of all test species except barnyardgrass
were delayed in treatment with leaves of common sunflower. Days to flowering of
barnyardgrass was not affected by leaves incorporated in the soil. Again here,
inhibitory substances in leaves of common sunflower exert some selectivity on
barnyardgrass.

11
Table 4. Effect of incorporated common sunflower leaves on days to flowering of rice, mungbean, itchgrass, barnyardgrass and water purslane.
TEST PLANT DAYS TO FLOWERING (DAS)
Untreated Leaf Incorporation
Rice Rice 73.67b 83.33a Rice+Barnyardgrass 72.67b 81.33a Rice+Water purslane 72.00b 82.00a Mungbean Mungbean 35.67b 41.67a Mungbean+Itchgrass 32.33b 41.33a Itchgrass Itchgrass 34.33b 43.33a Mungbean+Itchgrass 34.33b 42.33ab Barnyardgrass Barnyardgrass 42.67a 42.00a Rice+Barnyardgrass 40.67a 42.00a Water purslane Water purslane 34.00ab 40.67a Rice+Water purslane 32.00b 39.33a
Number followed by the same letters within test species are not significantly different at 1% LSD
This result is in agreement with that of Narwal et al. (1999a) reported that
common sunflower leaves through decomposition released allelochemicals that
inhibited seed germination, further growth and development rate of plant such as
extended days of flowering. Addition of 2 g residues to 80 g of soil significantly delayed
seed emergence, depression in growth, and reduced sorghum dry weight (Schon and
Einhellig, 1982). Akemo et al. (2000) found that rate of flowering and fruit set of tomato
plants varied directly with the proportion of rye to pea in the cover crop, and decreased
with increased rye to pea proportion. Kohli et al. (1998) also reported that
allelochemicals act via bringing certain changes in physiological functions, such as
respiration, photosynthesis and ion uptake. These, in turn, result in visible changes in
seed germination, and further reductions in growth and overall performance of the
target plants.
Shoot Dry Weight
The effect of incorporated common sunflower leaves on shoot dry weight of rice,
mungbean, itchgrass, barnyardgrass and water purslane at harvest is presented in
Table 5. As shown in Table 5, rice shoot dry weight in monoculture (+ 32.32%) was
12
significantly increased in the treatment with leaves compared to the untreated. Highest
shoot dry weight of rice was attained in monoculture rice (14.74 g/plant) with common
sunflower leaves incorporated in the soil, but was not significantly different from that in
the mixed culture with barnyardgrass.
Table 5. Effect of incorporated common sunflower leaves on shoot dry weight of rice, mungbean, itchgrass, barnyardgrass and water purslane at harvest.
TEST PLANT SHOOT DRY WEIGHT(g/plant)
Untreated Leaf Incorporation % Inhibition (-)/ Stimulation (+)
Rice Rice 11.14b 14.74a +32.32 s Rice+Barnyardgrass 4.05c 4.33c +6.91 ns Rice+W.purslane 12.21ab 13.94ab +14.17 ns Barnyardgrass Barnyardgrass 20.32a 18.82a -7.38 ns Rice+Barnyardgrass 18.49a 17.33a -6.27 ns Water purslane Water purslane 10.50a 8.29b -21.05 s Rice+W.purslane 4.73c 2.76d -41.65 s Mungbean Mungbean 3.28a 2.81ab -14.33 ns Mungbean+Itchgrass 1.87c 2.33bc +24.60 ns Itchgrass Itchgrass 5.41a 3.87b -28.47 s Mungbean+Itchgrass 2.38c 1.52c -36.13 ns
s = significant, ns = not significant Numbers followed by the same letters within test species are not significantly different at 5% LSD
Among the untreated (mono and mixed culture of rice and barnyardgrass), rice
shoot dry weight was lowest when rice was made to compete with barnyardgrass. Rice
shoot dry weight was not affected by the leaves incorporated in the mixed culture with
water purslane or barnyardgrass. However, leaves of common sunflower incorporated
in soil reduced shoot dry weights of water purslane in mono (-21.05) and mixed culture
(-41.65).
Stimulation in shoot dry weight of rice in mono but not in the mixed culture was
observed when leaves were incorporated in the soil compared to untreated. The
difference in stimulation in rice shoot dry weight in mixed cultures of barnyardgrass
and water purslane probably due to the greater competition ability of barnyardgrass
than water purslane against rice. Conversely, inhibition of shoot dry weight was

13
observed in mixed culture barnyardgrass (-6.27%) and mixed culture water purslane (-
41.65%) as companion crops of rice.
As in barnyardgrass and water purslane significant reduction in itchgrass shoot
dry weight was recorded when leaves were incorporated in the soil. The stimulation in
shoot dry weight of rice in monoculture could be due to the residues being source as
organic matter. Soil amendment of straw as a supply of organic matter had been
reported by Taja and Zaag (1986), and Schmidit (1982). Narwal et al. (1999) reported that
57.1% organic carbon from the sunflower shoot was mineralized/decomposed in 9
weeks.
The degree of inhibition on plant growth varied depending on test species. At 56
DAS and beyond, it was observed that leaves incorporated in the soil did not adversely
affect the growth of rice and barnyardgrass, but were still inhibitory to itchgrass and
water purslane. However, incorporated leaves did not affect rice when grown with
barnyardgrass or water purslane. Mungbean shoot dry weights were not affected by
incorporated leaves in both mono and mixed culture with itchgrass. While, leaf residues
of sunflower reduced shoot dry weights of itchgrass in monoculture. Shoot dry weight
of itchgrass was reduced in the monoculture but not in the mixed culture.
Root Dry Weight
Rice root dry weights with common sunflower leaves incorporated in the soil
were not significantly different from those of untreated (Table 6). Monoculture and
mixed culture rice with barnyardgrass or water purslane were not affected by
incorporated leaves. Likewise barnyardgrass root dry weight in mono or mixed culture
was not significantly changed by leaf incorporation. Only water purslane root dry
weight in mono or mixed culture, and itchgrass in monoculture were significantly
reduced by the incorporated leaves of common sunflower.

14
Table. 6. Effect of incorporated common sunflower leaves on root dry weight of rice, mungbean, itchgrass, barnyardgrass and water purslane at harvest.
TEST PLANT ROOT DRY WEIGHT(g/plant)
Untreated Leaf Incorporation % Inhibition (-)/ Stimulation (+)
Rice Rice 4.61a 5.51a +19.52 ns Rice+Barnyardgrass 0.99b 1.39b +40.40 ns Rice+W.purslane 3.87a 5.10a +31.78 ns Barnyardgrass Barnyardgrass 25.45a 23.23a -8.72 ns Rice+Barnyardgrass 24.28a 22.24a -8.40 ns Water purslane Water purslane 2.27a 2.02b -11.01 s Rice+W.purslane 0.73c 0.42d -42.47 s Mungbean Mungbean 0.53a 0.45a -15.09 ns Mungbean+Itchgrass 0.16c 0.27b +68.75 s Itchgrass Itchgrass 2.32a 1.79b -22.84 s Mungbean+Itchgrass 1.27c 0.90c -29.13 ns
s = significant, ns = not significant Numbers followed by the same letters within test species are not significantly different at 5% LSD
In contrast to the shoot dry weight, root dry weight of rice monoculture was not
stimulated by the incorporation of leaves were incorporated in the soil. Great inhibition
of root dry weight (-42.47%) occurred in mixed culture of rice and water purslane as
compared to barnyardgrass (-8.40%). This suggests that water purslane was more
sensitive to the incorporated leaves of common sunflower than barnyardgrass.
Mungbean root dry weight in mixed culture was higher than the untreated when leaves
were incorporated in the soil. Root dry weight of itchgrass in monoculture was
significantly reduced.
There was stimulation in root dry weight of mungbean mixed culture (+68.75%)
when leaf residues were incorporated in the soil as compared to the untreated. While,
itchgrass shoot dry weight on mixed culture was slightly inhibited. It may be inferred
that inhibitory effects of common sunflower leaves on weed species may eliminate
competition between mungbean and itchgrass, and may give favorable growth
condition to the crop.

15
Incorporated leaves of common sunflower reduced the germination, plant
height, and dry matter accumulation in weed species. This may be due to presence of
phytotoxic allelochemicals in the sunflower infested soil. These results collaborate the
findings of Wilson and Rice (1968), Nanjappa et al. (1998) and Narwal (1999). Park (1991)
also indicated that sunflower exudates reduced fresh weight of radish, and E. colona by
23% and 17% respectively, the fresh weight of rice was not affected.
Abeysekera (1992) also reported that residues of both wild sunflower [Tithonia
diversifolia (Herm SL.) Gray] and common sunflower [Helianthus annuus (L.)]
significantly inhibited germination, seedling growth, and reduced fresh weight, and dry
weight of E. colona, but no inhibitory effects on rice plants. Sandhu (1997) reported that
in the field the preceding sunflower crop decreased the plant height and forage or grain
yields of all crops, however, inhibition was maximum in greengram, maize and sesame
and minimum in rice and cowpea.
Grain Yield
Grain yield of rice and mungbean whether in mono or mixed cultures with the
weeds were not affected significantly by the incorporated leaves of common sunflower.
However, the seed yield of water purslane in both mono and mixed culture with rice
and itchgrass in mixed culture with mungbean was reduced (Table 7). Higher grain
yield was attained in monoculture rice (15.75 g/plant) with leaves incorporated in the
soil, but was not significantly different from that of the untreated. Seed yield of
barnyardgrass whether in mono or mixed culture with rice was not affected by the
incorporated leaves in mono and mixed culture with rice.
The magnitude of inhibition was greater in water purslane mixed culture with
rice than in the monoculture. The significant reduction in seed grain yield of water
purslane suggests inhibitory substances present in common sunflower leaves. The
observed reduction in water purslane seed yield due to the incorporated leaves of
common sunflower, reduced competition, increasing available growth factor such as,
water, soil nutrients, and light for the rice crop. The stimulation of grain yield of rice
though not significant could be due to the residues serving as source organic matter.
Soil amendment with straw as a supply of organic matter had been reported by Taja and
Zaag (1986), and Schmidit (1982).
16
Table 7. Effect of incorporated common sunflower leaves on grain seed yield of rice, mungbean, itchgrass, barnyardgrass and water purslane
TEST PLANT GRAIN SEED YIELD (g/plant)
Untreated Leaf Incorporation % Inhibition (-)/ Stimulation (+)
Rice Rice 12.90ab 15.75a +22.09 ns Rice+Barnyardgrass 4.40c 3.91c -11.14 ns Rice+W.purslane 11.42b 13.68ab +19.79 ns Barnyardgrass Barnyardgrass 6.19a 5.96a -12.44 ns Rice+Barnyardgrass 5.72a 6.17a +7.87 ns Water purslane Water purslane 7.03a 6.28b -10.67 s Rice+W.purslane 4.05c 1.79d -55.80 s Mungbean Mungbean 1.84a 1.81a -1.63 ns Mungbean+Itchgrass 1.40a 1.76a +25.71 ns Itchgrass Itchgrass 5.44a 4.05ab -25.55 ns Mungbean+Itchgrass 5.29a 3.06b -42.16 s
s = significant, ns = not significant Numbers followed by the same letters within test species are not significantly different at 5% LSD
Seed yield of rice in mixed culture rice + barnyardgrass (3.91 g/plant) was
significantly lower than that in mixed culture rice + water purslane (13.68 g/plant). This
suggests that barnyardgrass was more competitive to the rice as compared to water
purslane. Itchgrass seed yield in mixed culture with mungbean was highly reduced by
incorporated leaves of common sunflower. There was slight stimulation in grain yield of
mixed culture mungbean (+25.71%) but not significant when leaves were incorporated
in the soil as compared to the untreated.
Common sunflower leaves incorporated in the soil inhibited the germination of
most test plants except mungbean, but proved very slightly stimulatory to further
growth of rice. This indicates that comparatively, mungbean and rice were more tolerant
than the test weeds to the allelochemicals present in sunflower infested soil. These
results are in agreement with previous studies reported by Narwal et al. (1999a) that
preceding sunflower crop decreased the plant height, dry matter and yield of all
subsequent test crops as compared to those sown in fallow plots. Maximum reduction in
growth (plant height, dry matter), yield attributes and grain/fodder yield was observed

17
in cotton and sunflower, minimum in cereals (sorghum, pearlmillet, maize) and
moderate in legumes (clusterbean, cowpea, greengram) (Narwal et al.,1999a).
In soil incorporation of leaf extracts of common sunflower, abiotic (physical and
chemical) and biotic (microbial) soil barriers can limit the phytotoxicity of chemical in
terms of quality and quantity required to cause injury. Organic matter, reactive mineral
surfaces, ion exchange capacity, inorganic ions, and abiotic and biotic factors of soil
environment significantly influence allelochemical activity (Inderjit, 2000). Beltran (1997)
observed that allelopathic effects are difficult to ascertain in nature owing to the
difficulty of separating them from other factors (edaphic, climatic, and competition for
water, light and nutrients). Even in the absence of microbial metabolism, soil renders
compounds much less toxic than they are in laboratory solutions. This reduced toxicity
is due to slow diffusion rate and to formation of various complexes and sorptive
reactions in soil. Schmidt (1999) found that even extremely toxic compounds like 2,5-
dinitrophenol is much less toxic in soil than in laboratory bioassays.
Identification of chemicals in sunflower done by Dharamraj (1998) indicated that
phenolics are responsible for the observed interference in nature and are potent
allelochemicals. He suggested that the allelopathic effect of sunflower leaf litter is due to
presence of phenolic acids viz., p-coumaric, vanillic, syringic, p-hydroxybenzoic and
ferulic. Phenolic acids are released into soil by plants as decomposition metabolites and
in some plants as root exudates or leaf leachates (Cecchi et al., 1999). These chemicals
are involved in several soil processes, including the formation of humus, nutrient
availability, dissolution of minerals and allelopathy. Gogoi et al. (2000) reported that
phenols are common secondary plant metabolites involved in such phytotoxic activity,
as they occur in higher concentrations in plant tissues and soil. The phenolic acids affect
seed germination, radicle and plumule elongation and dry matter accumulation in many
crop species.
The stimulation of growth in the test plants could be due to the common
sunflower residues serving as source of organic matter. Soil amendment of straw as a
supply of organic matter had been reported by Taja and Zaag (1986), and Schmidit
(1982). These workers pointed out that upon decomposition, proteins, nucleic acids and
nitrogenous substances were converted to NO3- which would be utilized by the plant.

18
This phenomenon may explain the observation in monoculture rice with incorporated
leaves in the soil, that shoot growth of rice was stimulated. This result collaborates the
finding of Narwal et al. (1999b) who reported that the phytotoxicity of sunflower
biomass persisted in the soil up to 9 weeks after soil incorporation, then the harmful
effect decreased with passage of time. In 9 weeks, 57.1% organic carbon from the
sunflower shoot was mineralized/decomposed.
Debnath and Hajara (1972) found that soil treated sesbania also increased other
soil nutrients but it maintained the same pH as that of control. The present results are
agreement with the previous investigations as reported by Nagarajah and Nizar (1982).
One ton of fresh wild sunflower leaves and tender stem is capable of providing about 5
kg N, 1 kg P2O5, 10 kg k2O. Application of inorganic fertilizer can be appreciably
reduced when it is applied with green manure.
CONCLUSION
Incorporation of common sunflower leaves significantly inhibited the emergence
of itchgrass, barnyardgrass, and water purslane, slightly inhibited that in rice, while
there was no effect in mungbean. Plant height, shoot and root dry weights and grain
yield of itchgrass and water purslane decreased significantly with incorporation of
common sunflower leaves over those of the untreated.
Mungbean and barnyardgrass were less affected by leaf incorporation, while rice
was slightly affected. The slight enhancement of growth in rice may be due to the
organic matter coming from the incorporated leaves. Incorporated common sunflower
leaves in the soil had inhibitory potential on water purslane and itchgrass, but less or no
inhibition on rice and mungbean. It may be inferred that common sunflower leaves may
be used for effective itchgrass and water purslane controls, without damaging
mungbean and rice plants.

19
LITERATURE CITED
ABEYSEKERA, S.K. M.M.A. 1992. comparative study on agronomic characters and allelopathic activity of common sunflower (Helianthus annuus L.) and wild sunflower (Tithonia diversifolia (Herm SL.) Gray). MS Thesis . University of the Philippines at Los Baños. Philippines. 71p.
AKEMO, M.C., BENNETT and E.E. REGNIER. 2000. Tomato growth in spring-sown cover crops. Hortscience 35:843-848.
ALIOTTA, G. 1998. Localization of action site(s) of allelochemicals in seeds: Implications for weed control. In Abstracts. Allelopathy Journal 6(2).pp 281-282.
AN, M., I.R. JOHNSON and J.V. LOVETT. 1996. Mathematical modeling of allelopathy. I. Phytotoxicity caused by plant residues during decomposition. Allelopathy Journal 3 (1): 281-282.
______, J. PRATLEY and T. HAIG. 1998. Allelopathy: from concept to reality. Proceeding of the 9th Australian Agronomy Conference, Wagga wagga. pp. 1-5.
ANAYA, A.L. and H.R. PELAYO-BENAVIDES. 1997. Allelopathic potential of Mirabilis jalapa L. (Nyctaginaceae): Effects on germination, growth and cell division of some plants. Allelopathy Journal 4: 57-68.
BARANOV, V.I. 1980. Interaction of Phenolic compounds with metallic ions in regulation of growth. Sov. Plant Physiol. 27:805-811.
BATISH, D.R., H.P. SINGH and R.K. KOHLI. 2001. Allelopathy as a tool for sustainable weed management. The Proceeding of the 18th Asian-Pacific Weed Science Society Conference. May 28- June 2, 2001. Beijing, P.R. China. pp. 168-173.
___________, P. TUNG, H.P. SINGH and R.K. KOHLI. 2002. Phytotoxicity of sunflower residues against some summer season crops. J. Agronomy and Crop Science 188, 19-24.
BELL, D.T. and KOEPPE. 1972. Non competitive effects of giant foxtail on growth of corn . Agron. J. 64:321-325.
BELTRAN, L., LEYVA, A. and L. CAPARICON. 1997. A preliminary study of the allelopathic effect of sunflower (Helianthus annuus L.) on several economic crops. Cultivos Tropicales 18(1): 40-42.
BERES, I., SARDI, K. and S. KAMAN. 1998. Allelopathic effect of Ambrosia artemisifolia L. on germination and seedling growth of field crops.In Abstracts. Allelopathy Journal 6(2).pp 288.
_________ and G. KAZINCZI. 2000. Allelopathic effects of shoot extracts and residues of weeds on field crops. Allelopathy Journal 7(1): 93-98.

20
CHEEMA, Z.A. and KHALIQ. 2000. Use of sorghum allelopathic properties to control weeds in irrigated wheat in a semi arid region of Punjab. Agriculture, Ecosystem and Environment 79:105-112.
CECCHI, A.M., KOSKINEN, W.C., HAIDER, K. and H.H. CHENG. 1999. Effect of soil properties on sorption of phenolic acids to soil. In Abstracts. Allelopathy Journal 7(1).pp. 164.
COCHRAN, V.L., L.F. ELLIOTT and R.I. PAPENDICK. 1977. The production of phytotoxins from surface crop residues. Soil Sci. Soc. Am. J. 41:903-908.
COLTON, C.E. and F.A. EINHELLIG. 1980. Allelopathic mechanisms of velvetleaf (Abutilon theoprasti Medic., Malvaceae) on soybean. Am. J. Bot. 67: 1407-1413.
CONNOR, D.J. and A.J. HALL. 1997. Sunflower physiology. In : SCHNEITER, A.A. (ed.), Sunflower technology and production, Monograph No. 35, pp. 113-182. CSSA, Madison.
CRONJE, J.I., RETHMAN, N.F.G. and C.F. REINHARDT. 1999. Towards understanding, crop reactions to mulching and relative importance of allelopathy. In Abstracts. Allelopathy Journal 7(1).pp 118.
CRUZ-ORTEGA, R., A.L. ANAYA, B.E. HERNANDEZ-BAUTISTA and G. LAGUNA-HERNANDEZ. 1998. Effects of allelochemicals stress produced by Sicyos deppei on seedling root ultrastrucutre of Phaseolus vulgaris and Cucurbita ficifolia. Journal of Chemical Ecology 24:2039-2057.
CURTIS, J.T. and G. COTTAM. 1950. Antibiotic and autotoxic effects in prairie sunflower. Bull. Torr. Bot. Club 77, 137-191.
DAKSHININI, K.M.M. and INDERJIT. 1999. Multifaceted approach in the study of allelopathy. In Abstracts, Allelopathy journal 7(1). pp116, 2000.
DHARMARAJ, G. 1998a. Allelopathic influence of sunflower on field crops. In abstracts. Allelopathy Journal 6(1).pp.103.
____________. 1998b. Influence of germinating seeds and seedlings of sunflower on weed spp. In abstracts. Allelopathy Journal 6(1).pp.112.
____________. 1998c. Identification of allelochemicals in sunflower. In abstracts. Allelopathy Journal 6(1).pp.114.
DAYAN, F.E., J.G. ROMAGNI, M.R. TELLEZ, A.M. RIMARDO and S.O. DUKE. 1999. Managing weeds with natural products. Pestic. Outlook. 5, 185-188.
DUKE, S.O., F.E. DAYAN, A. HERNANDEZ, M.V. DUKE and H.K. ABBAS. 1997. Natural products as leads for new herbicide mode of action. Proc. Brighton Crop Prot. Conf., Weeds. 2, 579-586.
EINHELLIG, F.A. and L. KUAN. 1971. Effects of scopoletin and chlorogenic acid on stomata aperture in tobacco and sunflower. Bull. Torrey Bot. Club 98 : 155-162.

21
___________ and M. L. STILLE. 1979. Effects of ferulic and p-coumaric acids on plant water status. Abstr. Bot. Soc. Am., Misc. Ser. Publ. 157:40-41
___________ and G.R. LEATHER. 1988. Potential for exploiting allelopathy to enhance crop production. Journal of Chemical Ecology 14: 1829-1844.
___________. 1996. Interactions involving allelopathy in cropping systems. Agron. J. 88, 886-893.
ESPINO, R.R.C. and C. S. ATIENZA. 2000. Crop diversification in the Philippines. Department of Horticulture, College of Agriculture, UP Los Baños, Laguna, Philippines.15p.
GOGOI, B., KAUSHIK DAS and K.K. BARUAH. 2000. Effcet of allelochemicals on germination and seedling growth of rice (Oryza zativa L.) cultivars. Allelopathy Journal 7 (2): 279-284.
GLASS, A.D.M. 1973. 1973. Influence of phenolic acids on ion uptake. Plant Physiology 51: 1037-1041.
GRUMMER, G. 1955. Die gegenseitige bonere pflanzen-allelopathie. In: THOMPSON, A.C. (ed). 1985. The Chemistry of allelopathy. ACS Symp. Series 268. Washington, D.C. 4, 34-54.
GUENZI, W.D. and Mc CALLA. 1966. Phenolic acids in oats, wheat, sorghum and corn residues and their phytotoxicity. Agron. J. 58:303-304.
HALL, A.B., U. BLUM and R.C. FITES. 1982. Stress modification of allelopathy of Helianthus annuus L. debris on seed germination. Amer. J. Bot.: 776-783.
HAO, X. and E. DE JONG. 1988. Growth of wheat and barley seedlings at different matric and osmotic potentials. Agronomy Journal 80: 807-811.
HEGDE, R.S. and D.A. MILLER. 1990. Allelopathy and autotoxicity in alfafa: characterization and effects of preceding crops and residue incorporation. Crop. Sci. 30, 1255-1259.
HWANG, S.J., D.H. SHIN and K.U. KIM. 1997. Identification of biologically active substance from lilac (Syringa vulgaris L.). Korean Journal of Weed Science 17: 334-344.
INDERJIT and K.M.M. DAKSHINI. 1998. Allelopathic interference of Chickweed (Stellaria media) with seedling growth of wheat (Triticum aesticum). Cananadian Journal of Botany 76: 1317-1321.
________. 2000. Soil: environmental effects on allelochemical activity. Agronomy Journal 93: 79-84.
IRONS, M.S. and C.V. BURNSIDE. 1982. Competitive and allelopathic effects of sunflower. Weed Science 30:372-377.
JANIYA, J.D. 2002. Yield losses, major weed species and suggested management systems in selected major crops [Rice (Oryza sativa L.)]. In Weed management in major crops of the Philippines. WSSP, Inc. CPAP, Inc. pp.17-37.

22
JUNG QUAN, YU. 1999. Allelopathic problems and research in vegetable production. Advances in Horticulture 2: 336-343.
KALITA, D., H. CHOUDHARY and S.C. DEY. 1999. Assessment of allelopathic potential of some common upland rice weed species on morpho-physiological properties of rice (Oryza sativa L.) plant. Crop research 17: 41-45.
KIMBER, R.W.L. 1973. Phytotoxicity from plant residues. II. The effect of time of rotting of straw from some grasses and legumes on the growth of wheat seedlings. Plant Soil 38: 347-361.
KITOU, M. and S. YOSHIDA. 1998. Allelopathic effects of extracts of soil amended with some plant materials on germination and radicle elongation of lettuce. Journal of Weed Science and Technology 43: 1-9.
KOBAYASHI, K. 1998. Allelopathic activity of Mexican sunflower (Tithonia diversifolia) in soil. Weed Science 46: 432-437.
KOEPPE, D. E., L. M. ROHRBAUGH, E. L. RICE and S.H. WENDER. 1970. Tissue age and caffeoylquinic acid concentrations in sunflower, Phytochemistry 9:297-301.
KOHLI, R.K., D. BATISH and H.P. SINGH. 1998. Allelopathy and its implications in agroecosystems. Journal of Crop Production. 1: 169-202.
KOHLI, R.K. and PARIANA. 1992. Allelopathic impact of sunflower on crops. In Proceedings. National Symposium Allelopathy in Agroecosystems. TAURO, P. and S.S. NARWAL (eds). Hisar India : Indian Society of Allelopathy. pp. 29-30.
PARK, K.H. 1991. Allelopatic activity and potential natural herbicide from sunflower (Helianthus annuus L.). Ph.D Dissertation. University of the Philippines at Los Baños. Philippines. 132p.
LEATHER, G.R. 1983. Sunflowers (Helianthus annuus) are Allelopathic to weeds. Weed Science. 31:37-42.
LEE, C.W., Y.W. KIM and B.C. KOO. 1997. Influence of light, osmotic potential, pH and rice straw mulching on growth of barley (Hordeum vulgare L.) and water foxtail (Alopecurus aequalis var. amurensis (kom) Ohwi). Korean Journal of Weed Science 17:310-313.
LIN, W.X., K.U. KIM and D.H. SHIN. 2000. Rice allelopathic potential and its modes of action on barnyardgrass (Echinochloa crus-galli). Allelopathy journal 7(2): 215-224.
LOVETT, J.V. 1989. Phytochemical ecology: allelochemicals, mycotoxins and insect pheromones and allomones. CHOU, C.H. and G.R. WALLER (eds). Taipei. ROC. Pp. 49-67.
___________. 1990. Allelopathy and self defense in Barley. In BASSET, C., L.S. WHITEHOUSE and J.A. ZABKIEWIZZ (eds). Alternatives to chemical control of weeds. Rotorua, New Zealand. pp. 57-65.

23
MACIAS, F.A. 1995. Allelopathy in the search for natural herbicide model. In Inderjit, DAKSHINI, K.M.M. and F.A. EINHELLING (eds).. Allelopathy, organisms, processes, and applications. ACS Symp. Series 582.Washington, DC, 23, 310-329.
___________, A. TORRES, J.M.G. MOLINILLO, R.M. VARELLA, and D.
CASTELLANO. 1996. Potential allelopathic sesquiterpene lactones from sunflower leaves. Peredovick. Pergamon. Phytochemistry, 43 : 1205-1215
___________, ROSA M. VARELA, ASCENSION and JOSE MARIA G. MOLINILLO. 1993. Potential allelopathic guainolides from cultivar sunflower leaves, var. SH-222. Peredovick. Pergamon. Phytochemistry, 34 : 669-674.
___________, J.M.G. MOLINILLO, R.M. VARELA, A. TORRES and R.O. TRONCOSO. 1998. Allelocemicals in: OLOFDOTTER, M. (ed). 1998a. Allelopathy in Rice. International Rice Research Institute (IRRI). pp.69-79.
___________, R.M. OLIVA, A.M. SIMONET and J.C.G. GALINDO. 1998b. What are allelocemicals? in: implications for crop and soil management in agroecosystems. In Abstracts. Allelopathy Journal. 6(2).pp.301.
___________, OLIVA, R.M., VARELLA, R.M., TORRES, A. and MOLINILLO, M.G. 1999. Allelochemicals from sunflower leaves cv. Peredovick. Pergamon. Phytochemistry, 52 : 613-621.
MALLIK, M.A.B. 1999. Allelopathy and nitrogen fixation in legumes. In Abstracts. Allelopathy Journal 7(1).pp 120.
MANDAVA, N.B. 1985. Chemistry and biology of allelopathic agent. In : THOMPSON,
A.C. (ed).1985. The Chemistry of allelopathy. ACS Symp. Series 268. Washington, DC, 4: 34-54.
MOODY, K. 1986. A literature review of weeds in grain legumes in the Philippines. Philipp. J. Weed Sci. 13: 19-27.
MOLISH, H. 1937. Der Einflusslinear pflanze aufdie audere-Allelopathie; In:THOMPSON (ed). 1985. The Chemistry of allelopathy. ACS Symp. Series 268 Washington, D.C.4: 34-54.
MULLER, C.H. 1971. Biochemical interactions among plants. National Academy of Sciences, Washington, D.C. pp. 64-72.
MURPHY, D.J.B. and T.L. NOLAND. 1982. Temperature effects on seed inhibition and leakage mediated by viscosity and membranes. Plant Physiology 69:428-431.
NAGARAJAH, S. and B.M. NIZAR. 1982. Wild sunflower (Thithonia diversifolia) green manure for rice in the mid country wet zone. Tropical agriculturist 138:69-79.
NANJAPPA, H.V., R. POONGUZHALAN and B.K. RAMACHANDRAPPA. 1999. Influence of sunflower on subsequent crops. Allelopathy Journal 6: 69-74.

24
NARWAL, S.S., M.K. SARMAH and D.P.S. NANDAL. 1997. Allelopathic effects of wheat residues on growth and yield of fodder crops. Allelopathy Journal 4:111-120.
_____________, YADAVA, S. and S. GUPTA. 1999a. Allelopathic effects of sunflower on succeeding summer crops. 1. Field studies and bioassays. Allelopathy Journal 6:35-48.
_____________, T. SINGH, J.S. HOODA and M.K. KATHURIA. 1999b. Allelopathic effects of sunflower on succeeding summer crops. 2. Pot culture and biomass decomposition. Allelopathy Journal 6:209-226.
NAVAREZ, D.C. and R.P. ROBLES. 2002. Yield losses, major weed species and suggested management systems in selected major crops (Field legumes). In Weed management in major crops of the Philippines. WSSP, Inc. CPAP, Inc. pp. 42-44.
NYARKO, K.A. and S.K. DE DATTA. 1991. A handbook for weed control in rice. International Rice Research Institute (IRRI). Philippines. 113 p.
PALLER, E.C. 2002. Yield losses, major weed species and suggested management systems in selected major crops [Corn (Zea mays L.)]. In Weed management in major crops of the Philippines. WSSP, Inc. CPAP, Inc. pp. 38-41.
PANCHO, J.V. 1991. Grassy weeds in the Philippines. In Baker, F.M.G. and P.J. Terry: Tropical grassy weeds. CAB International for Casafa. Report Series 203,pp. 183-188.
PATTERSON, D.T. 1981. Effects of allelopathic chemicals on growth and physiological responses of soybean (Glycine max). Weed Sci. 29:53-58.
PAWAR, K.B. and P.D. CHAVAN. 1999. Influence of leaf leachates of plant species on mineral nutrition of Sorghum bicolor (L.) Moench. Allelopathy Journal 6: 87-92.
PURVIS, C.E. and G.P.D. JONES. 1990. Differential response of wheat to retain crop stubbles. II. Other factors influencing allelopathic potential; intraspecific variation, soil type and stubble quantity, Aust. J. Agric. Res. 41, 243-25 2.
PUTNAM, A.R. 1994. Phytotoxicity of plant residues. In: UNGER, P.W. (ed.), Managing Agricultural Residues, CRC Press, New York. pp. 285-314.
REHMAN, A. 1998. Allelopathic potential of Parthenium hysterophorus Linn. On germination, growth and dry matter production in Cassis sophera Linn. Bionature 18(1): 17-20.
REIGOSA, M.J., A.M. SANCHEZ-MOREIRAS and O. WEISS. 1999. Interacting factors in the manifestation of allelopathic effects. In abstracts Allelopathy Journal 7(1).pp 118.
_________, L. GONZALES, A. SANCHES-MOREIRAS, B. DURAN, D. PUIME, D. A. FERNANDEZ and J.C. BOLANO. 2002. Comparison of physiological effects of allelochemicals and commercial herbicides. Allelopathy journal 8(2): 211-220.

25
RICE, E.L. and S.K. PANCHOLY. 1973. Inhibition of nitrification by climax ecosystems II. Additional evidence and possible role of tannins. American Journal of Botany 60:691-720.
_________. 1974. Allelopathy. Academic Press, New York. 353 p.
_________, C.Y. LIN and C.Y. HUANG. 1981. Effects of decomposing rice straw on growth and nitrogen fixation by Rhizobium journal of chemical ecology 7: 333-344
_________. 1984. Allelopathy (second edition). Academic Press, Orlando. p. 29-30.
ROMEO, J.T. 2000. Raising the beam: moving beyond phytotoxicity. Journal of Chemical Ecology 26:2011-2014.
ROSS, M.A. and C.A. LEMBI. 1999 Applied weed science. 2nd edn. Prentice-Hall. Upper Saddle River, New Jersey. 452p.
SAJJAN, A.S. and B.D. BIRADAR. 1998. Allelopathic effect of Cassia sericea on seed and seedling parameters in rabi sorghum. World Weeds 5: 81-83.
SANDRAS, V.O. and N. TRAPANI. 1999. Leaf expansion and phonologic development: key determinants of sunflower plasticity, growth and yield. In: SMITH, D.L. and C. HAMEL (eds), Physiological control of growth and yield in filed crops. Springer-Verlag, Berlin. pp. 205-232.
SCHMIDT, S.K. 1999. Microbial competition limits the toxicity of allelochemicals in natural soils. In Abstracts. Allelopathy Journal 7(1).pp 164.
SCHMIDIT, E.L. 1982. Nitrification in the soils. In Agronomy Monograph. 22p.
SCHON, M.K. and F.A. EINHELLIG. 1982. Allelopathic effects of cultivated sunflower on grain sorghum. Botanical Gazette. 143: 505-510.
SPRING, O. and A. HAGER. 1982. Inhibition of elongation growth by two sesquiterpene lactones isolated from Helianthus annuus. Peredovick. Pergamon. Phytochemistry, 21 : 2551-2553.
___________, K. ALBERT and A. HAGER. 1983. Three biologically active heliangolides from Helianthus annuus. Peredovick. Pergamon. Phytochemistry, 21 : 2551-2553.
SPOLLEN, W. G. and R.E. SHARP. 1991. Spatial distribution of turgor and root growth at low water potentials. Plant Physiology 96: 438-443.
STEINSIEK, J.W., L.R. OLIVER and F.C. COLLINS. 1982. Allelopathic potential of wheat straw on selected weed species. Weed Science. 30: 495-497.
STERLING, T.M., R.L. HOUTZ and A.R. PUTNAM. 1987. Phytotoxic exudates from velvetleaf (Abutilon theophrasti) glandular trichomes. American Journal of Botany 74: 543-550.
TAJA, H. and P.V.D. ZAAG. 1986. Mulching and residue incorporation effects on chemical and physical properties of an isohyperthermis typic agrius. Toll paper. 27p.

26
TIWARI, J. P., S.P. KURCHANIA, C.S. BHALLA and N.R. PARADKAR. 1998. Studies on allelopathic effects of Cassia tora on Parthenium hysterophorus. In Abstracts. Allelopathy Journal 6(2).pp.292.
TONGMA, S., K. KOBAYASHI and K. USUI. 1997. Effect of water extracts from Mexican sunflower [Tithonia diversifolia (Hemsl.) A. Gray] on germination and growth of tested plants. Journal of Weed Science and Technology 42: 373-378.
TSAO, R. and Q. YU. 2000. Nematicidal activity of monoterpenoid compounds against economically important nematodes in agriculture. J. Essent. Oil Res. 12, 350-354.
TUNG. P., ASHIMA and R.K. KOHLI. 1996. Alleopathic. Impact of sunflower (Helianthus annus L.)on Phaseolus aureus Roxb. In : R.K. Kohli, N. Jerath, and D. Batish (eds). Some facets of biodiversity. Researchco Publications, New Delhi, India. pp.219-227.
VISALAKSHI, M. and K.K.V. SARMA. 1998. Allelopathic potential of five weedy species of Cassia on nodulation and dry matter production of Arachis hypogaea L. In Abstracts. Allelopathy Journal. 6(2).pp 289.
WARDLE, D. A., K.J. NICHOLSON and M. AHMED. 1992. Comparison of osmotic and allelopathy effects of grass leaf extracts on grass seed germination and radicle elongation. Plant and soil 140: 315-319.
_____________, M.C. NILSON, C. GALLET and O. ZACKRISSON. 1998. An ecosystem level perspective of allelopathy. In Abstracts. Allelopathy Journal. 6(2).pp 278.
WATANABE, K., N. OHNO, H. YOSHIOKA, J. GERSHENZON and TOM J. MABRY. 1982. Sesquiterpene lactones and diterpenoids from Helianthus argophyllus. Peredovick. Pergamon. Phytochemistry, 21 : 709-713.
WILSON, R.E. and E.L. RICE. 1968. Allelopathy as expressed by Helianthus annus and its role in old-field succession. Bulletin of the Torrey Botanical Club. 95: 432-448.
WORSHAM, A.D. 1989. Strategies capitalizing on allelopathy. In phytochemical ecology: allelochemicals, mycotoxins, and insect pheromones and allomones; Chou, C.H., Waller, G.R. Eds. Institute of Botany, Academia Sinica Monograph 9, Taipei, Taiwan. pp 275-291.
ZEEVAART, J.A.D. 1977. Sites of abscisic acid synthesis and metabolism in Ricinus communis L. Plant Physiol. 59: 788-791.
ZUBILLAGA, M.M., J.P. ARISTI and R.S. LAVADO. 2002. Effect of phosphorus and nitrogen fertilization on sunflower (Helianthus annus L.). J. Agronomy and Crop Science 188, 267-274.
ZWAIN, K.H. 1999. Allelopathic effects of wheat (Triticum aestivum L.) on nitrogen cycle. In Abstracts. Allelopathy Journal 6(1) .pp. 100.




















