
Effect of chromium supplementation and copper status on glucose and lipid metabolism in Angus and...
13
Animal Feed Science and Technology 128 (2006) 253–265 Effect of chromium supplementation and copper status on glucose and lipid metabolism in Angus and Simmental beef cows H.S. Stahlhut, C.S. Whisnant, K.E. Lloyd, E.J. Baird, L.R. Legleiter, S.L. Hansen, J.W. Spears ∗ Department of Animal Science and Interdepartmental Nutrition Program, North Carolina State University, Raleigh, NC 27695-7621, United States Received 1 April 2005; received in revised form 11 October 2005; accepted 1 November 2005 Abstract Pregnant Angus (n = 83) and Simmental (n = 69) cows were blocked by age into three blocks and then randomly assigned by breed to one of two free choice mineral supplements to determine effects of dietary Cr and Cu status on glucose metabolism and blood metabolites in beef cows. Supplements consisted of: (1) control (no supplemental Cr) and (2) 40 mg Cr/kg of mineral (from Cr picolinate). Mineral supplements were formulated to contain all minerals typically supplemented to cattle diets with the exception of Cu. The study began approximately 75 days prepartum, at which time half of the cows in each treatment received a 25 g Cu oxide needle bolus. Blood was collected from 36 cows on days 28, 58, 97 (approximately 20-day postpartum), 155, 210, and 279 for plasma glucose and non-esterified fatty acid (NEFA) determination. Chromium supplementation reduced (P<0.05) plasma glucose concentrations. Plasma glucose concentrations were also affected by breed × Cu bolus (P<0.05). In non-Cu supplemented cows, plasma glucose levels were higher (P<0.05) in Angus versus Simmental cows. In cows receiving a Cu bolus, plasma glucose levels were similar between breeds. Plasma NEFA concentrations were affected by time (P<0.01), Cr × Cu bolus (P<0.05), Cr × time (P<0.01), and Cr × block (P<0.01). On days 97 and 155, plasma NEFA concentrations were lower Abbreviations: Cr, chromium; Cu, copper; GTT, glucose tolerance test; GnRH, gonadotrophin releasing hormone; NEFA, non-esterified fatty acids Use of trade names in this publication does not imply endorsement by the North Carolina ARS or criticism of similar products not mentioned. ∗ Corresponding author. Tel.: +1 919 515 4008; fax: +1 919 515 4463. E-mail address: Jerry [email protected] (J.W. Spears). 0377-8401/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.anifeedsci.2005.11.002
Transcript of Effect of chromium supplementation and copper status on glucose and lipid metabolism in Angus and...

Animal Feed Science and Technology128 (2006) 253–265
Effect of chromium supplementation and copperstatus on glucose and lipid metabolism in
Angus and Simmental beef cows�
H.S. Stahlhut, C.S. Whisnant, K.E. Lloyd, E.J. Baird,L.R. Legleiter, S.L. Hansen, J.W. Spears ∗
Department of Animal Science and Interdepartmental Nutrition Program, North CarolinaState University, Raleigh, NC 27695-7621, United States
Received 1 April 2005; received in revised form 11 October 2005; accepted 1 November 2005
Abstract
Pregnant Angus (n = 83) and Simmental (n = 69) cows were blocked by age into three blocks andthen randomly assigned by breed to one of two free choice mineral supplements to determine effectsof dietary Cr and Cu status on glucose metabolism and blood metabolites in beef cows. Supplementsconsisted of: (1) control (no supplemental Cr) and (2) 40 mg Cr/kg of mineral (from Cr picolinate).Mineral supplements were formulated to contain all minerals typically supplemented to cattle dietswith the exception of Cu. The study began approximately 75 days prepartum, at which time halfof the cows in each treatment received a 25 g Cu oxide needle bolus. Blood was collected from 36cows on days 28, 58, 97 (approximately 20-day postpartum), 155, 210, and 279 for plasma glucoseand non-esterified fatty acid (NEFA) determination. Chromium supplementation reduced (P<0.05)plasma glucose concentrations. Plasma glucose concentrations were also affected by breed × Cu bolus(P<0.05). In non-Cu supplemented cows, plasma glucose levels were higher (P<0.05) in Angus versusSimmental cows. In cows receiving a Cu bolus, plasma glucose levels were similar between breeds.Plasma NEFA concentrations were affected by time (P<0.01), Cr × Cu bolus (P<0.05), Cr × time(P<0.01), and Cr × block (P<0.01). On days 97 and 155, plasma NEFA concentrations were lower
Abbreviations: Cr, chromium; Cu, copper; GTT, glucose tolerance test; GnRH, gonadotrophin releasinghormone; NEFA, non-esterified fatty acids
� Use of trade names in this publication does not imply endorsement by the North Carolina ARS or criticism ofsimilar products not mentioned.
∗ Corresponding author. Tel.: +1 919 515 4008; fax: +1 919 515 4463.E-mail address: Jerry [email protected] (J.W. Spears).
0377-8401/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.anifeedsci.2005.11.002

254 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
(P<0.01) in cows receiving Cr relative to control cows. Chromium supplementation reduced (P<0.01)plasma NEFA concentrations in 2 and 3 and 4 and 5-year-old cows but not in older cows, relativeto control cows in those blocks. Chromium supplemented cows had lower (P<0.05) plasma NEFAconcentrations than control cows that did not receive a Cu bolus. No differences in plasma NEFAconcentrations occurred between treatments in cows that received a Cu bolus. At approximately 1-month prepartum and 1-month postpartum, 12 cows were cannulated in their jugular vein and glucosetolerance tests (GTT) were conducted. Cows used in GTT received 0.15 g of glucose/kg body weightas a 500 g/L dextrose solution. Chromium supplemented cows had lower plasma glucose (P<0.01),serum insulin (P<0.05) and NEFA (P<0.01) concentrations following the GTT conducted prepartumversus control cows. Clearance rates for glucose were not affected by treatment. In the postpartumGTT, plasma glucose was affected by an interaction between Cr supplementation and Cu status, asCr supplemented cows that received a Cu bolus had higher (P<0.001) plasma glucose after glucoseadministration than cows not supplemented with Cu. No differences in plasma glucose occurredbetween control cows regardless of Cu status. Chromium supplemented cows had lower (P<0.05)serum insulin concentrations 10–45 min after glucose administration versus control cows. Resultsindicate that plasma glucose is lower in cows receiving supplemental Cr, and that an interactionbetween Cr and Cu status may alter glucose metabolism.© 2005 Elsevier B.V. All rights reserved.
Keywords: Chromium; Cattle; Copper; Glucose tolerance
1. Introduction
Chromium (Cr) potentiates the action of insulin in animals (Davis and Vincent, 1997).Altered glucose metabolism has been reported following Cr supplementation to rats(Schwarz and Mertz, 1959), humans (Anderson et al., 1991), swine (Matthews et al., 2001)and ruminants (Bunting et al., 1994; Chang et al., 1995; Kegley et al., 1997; Hayirli et al.,2001).
Previous Cr research in ruminants has been conducted in pre-ruminant calves (Kegleyet al., 1997), growing and finishing steers (Bunting et al., 1994; Chang et al., 1995) anddairy cattle (Subiyatno et al., 1996; Hayirli et al., 2001). However, no research has beendone to determine the effect of dietary Cr on glucose metabolism in reproducing beef cows.Volatile fatty acids produced by microbes present in the rumen serve as the primary energysubstrates for ruminants, and because ruminants synthesize most glucose, their response toinsulin is different than that in non-ruminants (Fahey and Berger, 1988).
Currently, there is no established Cr requirement for ruminants. Improvement in glucosetolerance after Cr supplementation is the best means to determine if Cr is limiting insulinresponsiveness. Delayed insulin response, and general glucose intolerance, has also beenshown in copper-deficient rats (Choudry et al., 1981; Hassel et al., 1983); suggesting thatCu may play a role in glucose metabolism as well. Consequently, the objective of this studywas to determine effects of supplemental Cr, and Cu status, on glucose metabolism andblood metabolites in reproducing beef cows. This study was part of a larger study designedto determine effects of supplemental Cr, and Cu status, on performance and reproductionof beef cows (Stahlhut et al., 2005).

H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 255
Table 1Composition of free-choice mineral supplement
Ingredient g/kg
Mono-dicalcium phosphate (210 g P/kg) 38.0Calcium carbonate 13.5Magnesium oxide (580 g Mg/kg) 18.5Salt 16.0Cane molasses (liquid) 5.0Rice meal by-product 5.5Zinc sulfate 0.694Manganese sulfate 0.846EDDI 0.004Cobalt carbonate 0.008Sodium selenite premix (2 g Se/kg) 1.5Mineral oil 0.5Vitamin Aa (IU) 50000Vitamin Da (IU) 6818Vitamin Ea (IU) 57
a IU are on a per kg basis.
2. Materials and methods
2.1. Protocol
Care, handling, and sampling of cows in this study were approved by the North CarolinaState University Animal Care and Use Committee. One hundred and fifty-two pregnantAngus (n = 83) and Simmental (n = 69) cows were blocked by age into three blocks. Blockone contained cows 6 years of age or older (n = 59; initial body weight (BW), 584.4 ± 6.5 kg),block two contained cows of 4–5 years of age (n = 42; initial BW, 542.2 ± 7.9 kg), and blockthree contained 2- and 3-year-old females (n = 51; initial BW, 468.9 ± 6.9 kg). Within ablock, cows were randomly assigned by breed to one of two free choice fed mineral supple-ments. Supplements consisted of: (1) control (no supplemental Cr) and (2) 40 mg Cr/kg ofmineral (from Cr picolinate). Mineral supplements were formulated to contain all mineralstypically supplemented to cattle diets with the exception of Cu (Table 1). The study beganapproximately 75-day prepartum, at which time half of the cows in each treatment receiveda 25 g Cu oxide needle bolus (Copasure® Animax LTD, Bury St. Edmunds, Suffolk, UK).
Cows were rotated among tall fescue pastures at 14-d intervals to equalize pasture dif-ferences during the grazing season. During the winter, cows were fed grass hay free choice.In addition to hay, cows in all blocks were fed 5.5 kg (DM) of corn silage and 0.9 kg of corngluten feed (dehydrated) following calving, although on a DM/kg of BW, young cows (i.e.,block 3) received more energy and protein than older cows.
Calves were born between day 43 (16 October 2003) and day 104 (15 December2003). Calves born to Cu supplemented dams were given a 12.5 g Cu oxide needle bolus(Copasure®) on day 196 (17 March). Cows were synchronized using two doses of Lutal-yse (Pfizer Animal Health, New York, NY, USA) on days 141 and 152 and one dose ofgonadotrophin releasing hormone (GnRH, Intervet, Wilmington, DE, USA) on day 145.

256 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
Cows were observed for heat for 3 days and those showing signs of estrus were artificiallyinseminated. All cows not showing signs of estrus were given a dose of GnRH and insem-inated on day 155. Cows were then exposed to Angus sires for 28 days, beginning on day159. Cows were palpated to determine pregnancy and calves were weaned on day 284 (14June). Three cows died on days 5, 70 and 217 of the study from bovine leukemia. Four cowswere removed from the study on day 111, two of which had calves born dead, and two thatwere determined to be open.
Cows were weighed at 28-day intervals throughout the 284-day study. Thirty-six cowswere bled via jugular venipuncture on days 28, 58, 97, 155, 210, and 279 to determineplasma glucose and non-esterified fatty acid (NEFA) concentrations. The same cows werebled on all sampling dates. Samples for plasma glucose and NEFA were collected in tubescontaining potassium oxalate and sodium fluoride (Vacutainer 6383, Becton Dickinson,Franklin Lakes, NJ, USA).
Glucose challenges were conducted on day 65 (prepartum; n = 11) and day 126 (postpar-tum; n = 12). Cows averaged 21 days (range of 4–41 days) prepartum on days 65 and 49 days(range of 40–57 days) postpartum on day 126. Two cows per block per treatment were usedalthough only 3-year-old cows were chosen to represent block three. Cows were cannulatedin the jugular vein approximately 24 h prior to the glucose challenge to minimize effectsof stress. Cows were moved into pens approximately 3-day prior to each challenge andfed tall fescue hay (ad libitum) and corn silage top dressed with 0.11 kg of their treatmentmineral supplement. One cow calved in the pen before the prepartum glucose challengecould be conducted. Following the regular morning feeding, 0.15 g of glucose/kg of BWwas administered in a 500 g/L dextrose solution via the jugular catheter over a period of2 min. Blood samples were collected at −5, 0, 5, 10, 20, 30, 45, 60, 90, and 120 min relativeto infusion. Blood samples for serum insulin determination were put into glass tubes andkept on ice until centrifuged. Blood for plasma glucose and NEFA concentration determi-nation were put in tubes containing potassium oxalate and sodium fluoride and kept in iceuntil centrifuged.
2.2. Analytical procedures
Blood was transported in ice to the laboratory and centrifuged at 2500×g for 20 min.Plasma and serum were removed and frozen at −20 ◦C until analysis for glucose, insulinand NEFA concentrations. Plasma glucose was determined by electrochemical sensor cou-pled to a membrane-immobilized glucose oxidase enzyme (Industrial Analyzer, Model 27,Yellow Springs Instrument, Yellow Springs, OH, USA). Serum insulin was determinedby radioimmunoassay using antibody coated tubes (ImmuChemTM Insulin Coated Tube,ICN Pharmaceuticals, Inc., Costa Mesa, CA, USA). Plasma NEFA concentrations weredetermined by an enzymatic, colorimetric method (WAKO Pure Chemical Industries, Ltd.,Richmond, VA, USA).
2.3. Data analysis
Data were analyzed by repeated measures using the ‘Mixed’ procedure of SAS (2001).Individual animal was used as the experimental unit for blood data. The model for blood data

H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 257
included treatment, breed, bolus, time, block, treatment × breed, treatment × bolus, treat-ment × time, treatment × block, bolus × breed, breed × time, and bolus × time. The modelfor the prepartum glucose challenge included treatment, time, breed, treatment × time,breed × time, and the breed × treatment interaction. Bolus was not included in the modelfor the prepartum glucose challenge due to one cow calving prior to the glucose chal-lenge and lack of degrees of freedom. In the postpartum glucose challenge the modelincluded treatment, time, breed, bolus, treatment × time, breed × time, treatment × breed,treatment × bolus, and the bolus × time interaction. Interactions that were not significant(P<0.05) for parameters of interest were deleted from the models.
3. Results
3.1. Mineral consumption
Average mineral disappearance during the course of the experiment was 0.084 ± 0.008and 0.087 ± 0.007 kg/cow/d for the control and Cr treatments, respectively. Based on min-eral intake, Cr consumption in the Cr picolinate treatment averaged 3.5 mg/cow/d, whichapproached the target supplemental Cr intake of 4.0 mg/cow/d.
3.2. Blood metabolites
Plasma glucose concentrations were affected by time (days pre- or postpartum; P<0.01),treatment (P<0.01), breed (P<0.01), and breed × Cu bolus (P<0.02). Plasma glucose wasnot affected by block or a block × treatment interaction. Cows averaged 48 days (range of33–70 days) prepartum when the first blood sample was collected on day 28 of the study.Plasma glucose concentrations increased with time until day 79 postpartum in all treat-ment groups (Fig. 1). Cows receiving supplemental Cr had lower (P<0.01) plasma glucoseconcentrations versus control cows. Plasma glucose was not affected by a breed × time
Fig. 1. Effect of chromium on plasma glucose concentrations in reproducing beef cows: [pooled S.E.M. = 0.42;treatment (P<0.01); time (P<0.01)].

258 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
Table 2Effect of breed and copper supplementation on plasma glucose concentrations in beef cows
Breed S.E. P
Angus Simmental
Plasma glucose (mg/dL)a 55.6 53.7 0.004 0.01−Cu 56.3 52.9 0.006 0.01+Cu 54.9 54.4 0.006 0.58
a Means across sampling times; breed × copper bolus (P<0.05).
interaction. Angus cows had higher (P<0.01) plasma glucose than Simmental cows thatdid not receive supplemental Cu (Table 2). In those cows that received a Cu bolus, plasmaglucose concentrations did not differ between breeds.
Plasma NEFA concentrations were affected by treatment × time (P<0.01), treat-ment × Cu bolus (P<0.04; Table 3) and treatment × block × time (P<0.01). Chromiumsupplemented cows had lower (P<0.01) plasma NEFA concentrations on days 21 and 79postpartum. In cows that did not receive a Cu bolus, supplemental Cr reduced (P<0.01)plasma NEFA concentrations. In cows receiving Cu, supplemental Cr only numerially(P<0.16) reduced plasma NEFA concentrations compared to control animals. Chromiumsupplemented cows in blocks 2 and 3 had lower (P<0.01) NEFA concentrations than con-trols in those same blocks. In cows 6 years of age or older (block 1), Cr did not affect plasmaNEFA.
3.3. Glucose tolerance tests
In the glucose challenge conducted prepartum, plasma glucose concentrations wereaffected by time (P<0.01) and treatment (P<0.01; Fig. 2). Cows receiving supplementalCr had lower (P<0.01) plasma glucose concentrations following the glucose challenge ver-
Table 3Effect of dietary chromium and copper supplementation on plasma NEFA concentrations in beef cows
Treatment S.E. P
Control Chromium
Plasma NEFA (mequiv./dL)a,b 421 313 17.3 0.01−Cu 452 290 25.6 0.01+Cu 389 337 24.4 0.16
Time, days postpartum−48 297 220 42.0 0.20−18 451 377 42.0 0.21
21 569 389 42.6 0.0179 669 373 42.0 0.01
134 249 273 42.7 0.69203 289 248 42.7 0.49
a Treatment × Cu bolus (P<0.05).b Treatment × time (P<0.01).
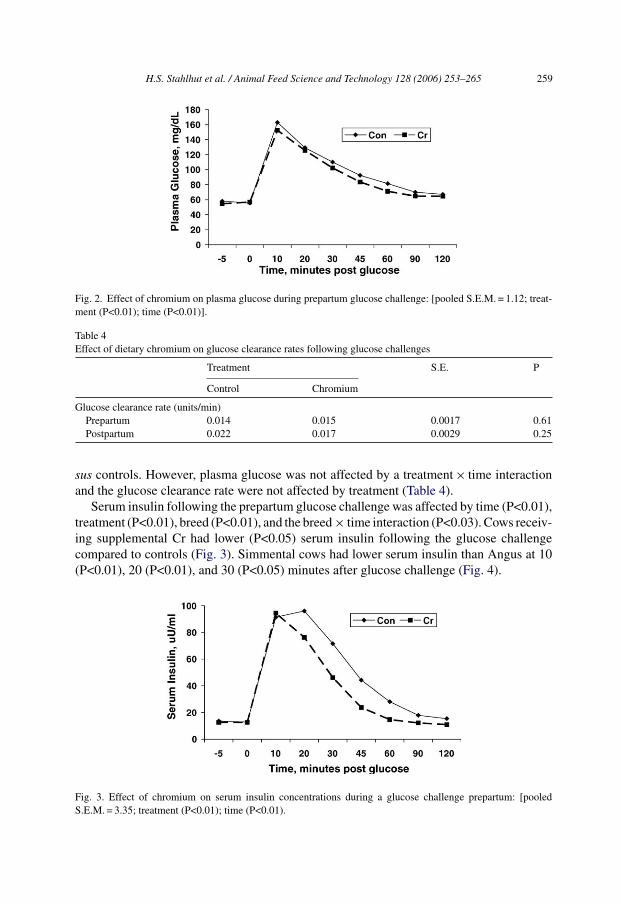
H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 259
Fig. 2. Effect of chromium on plasma glucose during prepartum glucose challenge: [pooled S.E.M. = 1.12; treat-ment (P<0.01); time (P<0.01)].
Table 4Effect of dietary chromium on glucose clearance rates following glucose challenges
Treatment S.E. P
Control Chromium
Glucose clearance rate (units/min)Prepartum 0.014 0.015 0.0017 0.61Postpartum 0.022 0.017 0.0029 0.25
sus controls. However, plasma glucose was not affected by a treatment × time interactionand the glucose clearance rate were not affected by treatment (Table 4).
Serum insulin following the prepartum glucose challenge was affected by time (P<0.01),treatment (P<0.01), breed (P<0.01), and the breed × time interaction (P<0.03). Cows receiv-ing supplemental Cr had lower (P<0.05) serum insulin following the glucose challengecompared to controls (Fig. 3). Simmental cows had lower serum insulin than Angus at 10(P<0.01), 20 (P<0.01), and 30 (P<0.05) minutes after glucose challenge (Fig. 4).
Fig. 3. Effect of chromium on serum insulin concentrations during a glucose challenge prepartum: [pooledS.E.M. = 3.35; treatment (P<0.01); time (P<0.01).

260 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
Fig. 4. Effect of breed on serum insulin concentrations following a glucose challenge prepartum: [pooledS.E.M. = 3.35; breed × time interaction (P<0.03); *P<0.05; **P<0.01)].
Plasma NEFA concentrations (data not shown) were affected by time (P<0.01), treat-ment (P<0.01), and a breed × treatment interaction (P<0.01). Angus cows receiving sup-plemental Cr had lower (P<0.01) plasma NEFA concentrations than Angus controls(174.69 ± 13.84 mequiv./mL versus 260.85 ± 11.30 mequiv./mL). No effect of treatmenton plasma NEFA concentrations was observed in Simmental cows.
In the glucose challenge conducted postpartum, plasma glucose concentrations wereaffected by time (P<0.01), treatment (P<0.01), and a treatment × bolus interaction (P<0.01;Fig. 5). In cows receiving a Cu bolus, Cr supplementation increased (P<0.01) plasma glucoseconcentrations following the postpartum glucose challenge. However, Cr did not affectplasma glucose concentrations in non-Cu supplemented cows. Glucose clearance rate wasnot affected by treatment (Table 4).
Serum insulin was affected by time (P<0.01), treatment (P<0.01), breed (P<0.01),breed × treatment (P<0.01), breed × time (P<0.01) and treatment × time (P<0.03) follow-ing the glucose challenge postpartum. Chromium supplementation resulted in lower seruminsulin at 10 (P<0.01) and 20 (P<0.05) minutes following glucose infusion compared to
Fig. 5. Effect of chromium and copper on plasma glucose concentrations during a glucose challenge postpartum:[pooled S.E.M. = 2.15; treatment × Cu bolus interaction (P<0.01); treatment (P<0.01); time (P<0.01)].
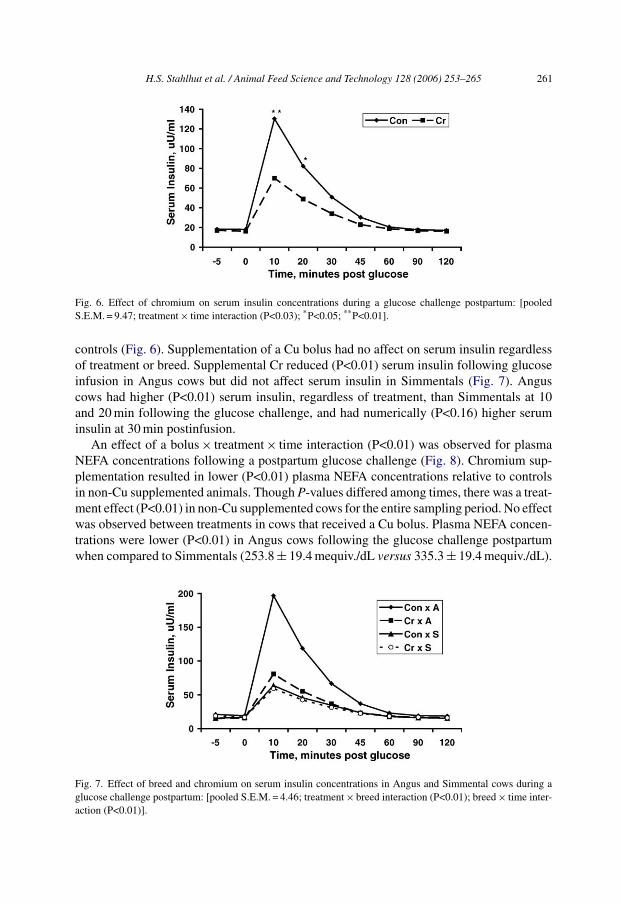
H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 261
Fig. 6. Effect of chromium on serum insulin concentrations during a glucose challenge postpartum: [pooledS.E.M. = 9.47; treatment × time interaction (P<0.03); *P<0.05; **P<0.01].
controls (Fig. 6). Supplementation of a Cu bolus had no affect on serum insulin regardlessof treatment or breed. Supplemental Cr reduced (P<0.01) serum insulin following glucoseinfusion in Angus cows but did not affect serum insulin in Simmentals (Fig. 7). Anguscows had higher (P<0.01) serum insulin, regardless of treatment, than Simmentals at 10and 20 min following the glucose challenge, and had numerically (P<0.16) higher seruminsulin at 30 min postinfusion.
An effect of a bolus × treatment × time interaction (P<0.01) was observed for plasmaNEFA concentrations following a postpartum glucose challenge (Fig. 8). Chromium sup-plementation resulted in lower (P<0.01) plasma NEFA concentrations relative to controlsin non-Cu supplemented animals. Though P-values differed among times, there was a treat-ment effect (P<0.01) in non-Cu supplemented cows for the entire sampling period. No effectwas observed between treatments in cows that received a Cu bolus. Plasma NEFA concen-trations were lower (P<0.01) in Angus cows following the glucose challenge postpartumwhen compared to Simmentals (253.8 ± 19.4 mequiv./dL versus 335.3 ± 19.4 mequiv./dL).
Fig. 7. Effect of breed and chromium on serum insulin concentrations in Angus and Simmental cows during aglucose challenge postpartum: [pooled S.E.M. = 4.46; treatment × breed interaction (P<0.01); breed × time inter-action (P<0.01)].

262 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
Fig. 8. Effect of chromium and copper bolus on plasma NEFA concentrations in postpartum glucose challenge:[pooled S.E.M. = 41.8; treatment × bolus × time interaction (P<0.01)].
4. Discussion
4.1. Blood metabolites
Supplementation with Cr lowered plasma glucose concentrations in our beef cows, bothprepartum and postpartum. Previous studies (Chang et al., 1995; Mowat et al., 1992) demon-strated that Cr supplementation lowered plasma glucose concentrations in growing andfinishing steers. However, other studies (Mowat et al., 1992; Kegley et al., 1997) found noeffect of Cr supplementation on plasma glucose concentrations in cattle.
Plasma NEFA concentrations were lower in cows receiving supplemental Cr shortly aftercalving (days 97 and 155), especially young cows. Our young cows (i.e., 2 and 3-year old)supplemented with Cr also lost less weight than controls (Stahlhut et al., 2005). The reducedweight loss in Cr supplemented cows is consistent with lower plasma NEFA concentrationsobserved in young cows shortly after calving, and suggests that Cr affected fat mobilizationfrom body stores to meet nutrient demands of lactation. Hayirli et al. (2001) reported that Crsupplementation did not affect BWs of dairy cows, but body condition score increased withincreasing supplementation of Cr methionine postpartum. Matthews et al. (2001) reportedthat swine supplemented with Cr picolinate had lower plasma NEFA concentrations versuscontrols. Similarly, studies conducted in sheep (Kitchalong et al., 1995) and dairy cattle(Hayirli et al., 2001) found that Cr supplementation lowered plasma NEFA concentrations.Conversely, supplemental Cr had no effect on plasma NEFA concentrations in growingsteers (Chang and Mowat, 1992; Bunting et al., 1994). Yang et al. (1996) also observedno effect on plasma NEFA concentrations in dairy cattle supplemented with chelated Cr.In the present study, responses in plasma NEFA concentrations to Cr were affected byCu status. Chromium supplementation lowered plasma NEFA concentrations in non-Cusupplemented cows, but only numerically lowered NEFA concentrations in those cows thatreceived a Cu bolus. Supplemental Cu alters metabolism of adipose tissue as well, as itsresponse to hormones responsible for lipolysis (Engle et al., 2000b), although in severalstudies supplemental Cu did not alter plasma NEFA concentrations in growing and finishingbeef steers (Engle et al., 2000a, 2000b).

H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 263
4.2. Glucose tolerance tests
Following a glucose challenge prepartum, plasma glucose concentrations were lowerin cows receiving supplemental Cr. Conversely, Kegley and Spears (1995) reported noeffect of supplementation with Cr nicotinic acid complex on plasma glucose concentrationsfollowing a glucose challenge in calves. In cows receiving a Cu bolus, Cr supplementationresulted in higher plasma glucose concentrations following a glucose challenge postpartum.However, no difference in plasma glucose concentrations was observed between treatmentsin cows that did not receive a Cu bolus. Although Cr supplementation reduced plasmaglucose concentrations regardless of time relative to glucose dosing, glucose clearancerates were not affected by Cr in either the prepartum or postpartum glucose challenges.Hayirli et al. (2001) reported no effect of supplemental Cr, from Cr methionine, on plasmaglucose concentrations (or area under the curve) in dairy cows following either prepartum orpostpartum glucose challenges. However, Cr picolinate supplementation increased glucoseclearance rate following a glucose challenge in growing calves (Bunting et al., 1994).
Serum insulin was lower in our cows receiving supplemental Cr following a glucosechallenge prepartum, and was lower at 10 and 20 min following glucose infusion in thepostpartum challenge. Subiyatno et al. (1996) observed that Cr supplementation decreasedserum insulin in primiparous dairy cows following a glucose challenge prepartum. Con-versely, Hayirli et al. (2001) reported higher serum insulin in dairy cows supplemented withCr methionine following a prepartum glucose challenge, However, following the postpar-tum challenge, serum insulin was lower in cows receiving Cr. Decreased levels of plasmaglucose and serum insulin following the prepartum glucose challenge in Cr supplementedcows suggests that less insulin was required to clear glucose from the blood, indicatinggreater tissue sensitivity to insulin. This is consistent with the role of Cr in potentiating theaction of insulin (Davis and Vincent, 1997). Bunting et al. (1994) observed increased tissuesensitivity to insulin in growing steers fed a corn grain-cottonseed hull diet supplementedwith Cr picolinate.
4.3. Breed differences in blood metabolites
Breed differences were observed in basal plasma glucose as well as plasma glucose,NEFA, and serum insulin following the glucose challenges. Angus cows had higher plasmaglucose concentrations than Simmentals throughout the study, as well as during the postpar-tum glucose challenge. Serum insulin was higher and plasma NEFA concentrations werelower in our Angus versus Simmental cows following glucose infusion during prepartum andpostpartum glucose challenges. Jones et al. (1991) reported that Angus heifers had higherserum insulin concentrations than Simmental heifers prior to puberty, but that glucose con-centrations were not affected by breed. Higher milk production in Simmental (Marston et al.,1992) may explain the lower plasma glucose, as glucose is utilized in synthesis of lactose.Mobilization of fat stores to meet energy demands of lactation may also explain the higherconcentration of plasma NEFA in Simmental cows. Differences in Cu requirements (Smartand Christensen, 1985; Ward et al., 1995; Mullis et al., 2003), average daily gain (Crouse etal., 1985; Gregory et al., 1994), fatty acid composition of carcass (Laborde et al., 2001) andmilk production (Marston et al., 1992) have been reported between Angus and Simmental

264 H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265
cattle, indicating a potential for differences in carbohydrate and lipid metabolism betweenthe breeds.
5. Conclusions
Supplementation with Cr resulted in lower plasma glucose concentrations in beef cowsthroughout the study. Plasma NEFA concentrations were reduced by Cr in early lactation butnot prepartum or in late lactation. Chromium altered glucose metabolism following glucosechallenges in reproducing beef cattle, however an unanticipated interaction between Crsupplementation and Cu status occurred postpartum. Further investigation is needed tounderstand the interaction between Cr supplementation and Cu status, as well as apparentdifferences in carbohydrate and lipid metabolism between Simmental and Angus cows.
Acknowledgement
Appreciation is extended to Prince Agri Products, Inc., Quincy, IL, USA for providingCr picolinate.
References
Anderson, R.A., Polansky, M.M., Bryden, N.A., Canary, J.J., 1991. Supplemental-chromium effects on glucose,insulin, glucagon, and urinary chromium losses in subjects consuming controlled low-chromium diets. Am. J.Clin. Nutr. 54, 909–919.
Bunting, L.D., Fernandez, J.M., Thompson Jr., D.L., Southern, L.L., 1994. Influence of chromium picolinate onglucose usage and metabolic criteria in growing Holstein calves. J. Anim. Sci. 72, 1591–1599.
Chang, X., Mowat, D.N., 1992. Supplemental chromium for stressed and growing feeder calves. J. Anim. Sci. 70,559–565.
Chang, X., Mowat, D.N., Mallard, B.A., 1995. Supplemental chromium and niacin for stressed feeder calves. Can.J. Anim. Sci. 75, 351–358.
Choudry, H., Srivastava, L., Murthy, L., Petering, H.G. 1981. Effects of dietary copper on insulin and thyroxinein male rats. Western Hemisphere Nutrition Congress VI, Los Angeles, CA, USA, August 1980. OfficialProgram, p. 84 (Abstact).
Crouse, J.D., Ferrell, C.L., Cundiff, L.V., 1985. Effects of sex condition, genotype and diet on bovine growth andcarcass characteristics. J. Anim. Sci. 60, 1219–1227.
Davis, C.M., Vincent, J.B., 1997. Chromium oligopeptide activates insulin receptor tyrosine kinase activity. Bio-chemistry 36, 4382–4385.
Engle, T.E., Spears, J.W., Armstrong, T.A., Wright, C.L., Odle, J., 2000a. Effects of dietary copper source andconcentration on carcass characteristics and lipid and cholesterol metabolism in growing and finishing steers.J. Anim. Sci. 78, 1053–1059.
Engle, T.E., Spears, J.W., Edens, F.W., 2000b. Dietary copper effects on lipid metabolism and circulating cate-cholamine concentrations in finishing steers. J. Anim. Sci. 78, 2737–2744.
Fahey, G.C., Berger, L.L., 1988. Carbohydrate nutrition of ruminants. The Ruminant Animal. Prentice-Hall,Englewood Cliffs, NJ, USA, pp. 269–297.
Gregory, K.E., Cundiff, L.V., Koch, R.M., Dikeman, M.E., Koohmaraie, M., 1994. Breed effects and retainedheterosis for growth, carcass, and meat traits in advanced generations of composite populations of beef cattle.J. Anim. Sci. 72, 833–850.

H.S. Stahlhut et al. / Animal Feed Science and Technology 128 (2006) 253–265 265
Hassel, C.A., Marchello, J.A., Lei, K.Y., 1983. Impaired glucose tolerance in copper-deficient rats. J. Nutr. 113,1081–1083.
Hayirli, A., Bremmer, D.R., Bertics, S.J., Socha, M.T., Grummer, R.R., 2001. Effect of chromium supplementationon production and metabolic parameters in periparturient dairy cows. J. Dairy Sci. 84, 1218–1230.
Jones, E.J., Armstrong, J.D., Harvey, R.W., 1991. Changes in metabolites, metabolic hormone and luteinizinghormone before puberty in Angus, Braford, Charolais, and Simmental heifers. J. Anim. Sci. 69, 1607–1615.
Kegley, E.B., Spears, J.W., 1995. Immune response, glucose metabolism, and performance of stressed feedercalves fed inorganic or organic chromium. J. Anim. Sci. 73, 2721–2726.
Kegley, E.B., Spears, J.W., Eisemann, J.H., 1997. Performance and glucose metabolism in calves fed a chromium-nicotinic acid complex or chromium chloride. J. Dairy Sci. 80, 1744–1750.
Kitchalong, L., Fernandez, J.M., Bunting, L.D., Southern, L.L., Bidner, T.D., 1995. Influence of chromium tripi-colinate on glucose metabolism and nutrient partitioning in growing lambs. J. Anim. Sci. 73, 2694–2705.
Laborde, F.L., Mandell, I.B., Tosh, J.J., Wilton, J.W., Buchanan-Smith, J.G., 2001. Breed effects on growthperformance, carcass characteristics, fatty acid composition, and palatability attributes in finishing steers. J.Anim. Sci. 79, 355–365.
Marston, T.T., Simms, D.D., Schalles, R.R., Zoellner, K.O., Martin, L.C., Fink, G.M., 1992. Relationship of milkproduction, milk expected progeny difference, and calf weaning weight in Angus and Simmental cow-calfpairs. J. Anim. Sci. 70, 3304–3310.
Matthews, J.O., Southern, L.L., Fernandez, J.M., Pontif, J.E., Bidner, T.D., Odgaard, R.L., 2001. Effect ofchromium picolinate and chromium propionate on glucose and insulin kinetics of growing barrows and ongrowth and carcass traits of growing-finishing barrows. J. Anim. Sci. 79, 2172–2178.
Mowat, D.N., Chang, X., Yang, W.Z., 1992. Chelated chromium for stressed feeder calves. Can. J. Anim. Sci. 73,49–55.
Mullis, L.A., Spears, J.W., McCraw, R.L., 2003. Estimated copper requirements of Angus and Simmental heifers.J. Anim. Sci. 81, 865–873.
SAS, 2001. SAS/STAT® User’s Guide (Release 8). SAS Institute Inc., Cary, NC, USA.Schwarz, K., Mertz, W., 1959. Chromium(III) and the glucose tolerance factor. Arch. Biochem. Biophys. 85,
292–295.Smart, M.E., Christensen, D.A., 1985. The effect of cow’s dietary copper intake, sire breed, and age on her copper
status and that of her fetus in the first ninety days of gestation. Can. J. Comp. Med. 49, 156–158.Stahlhut, H.S., Whisnant, C.S. Spears, J.W., 2005. Effect of chromium supplementation and copper status on
performance and reproduction of beef cows. Anim. Feed Sci. Technol., in press.Subiyatno, A., Mowat, D.N., Yang, W.Z., 1996. Metabolite and hormonal responses to glucose or proprionate
infusions in perparturient dairy cows supplemented with chromium. J. Dairy Sci. 79, 1437–1445.Ward, J.D., Spears, J.W., Gengelbach, G.P., 1995. Differences in copper status and copper metabolism among
Angus, Simmental, and Charolais cattle. J. Anim. Sci. 73, 571–577.Yang, W.Z., Mowat, D.N., Subiyanto, A., Liptrap, R.M., 1996. Effects of chromium supplementation on early-
lactation performance of Holstein cows. Can. J. Anim. Sci. 76, 221–230.




















