DULAI, A. (1995): Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts.,...
20
ANNALES HISTORICO-NATURALES MUSEI NATIONALIS HUNGARICI Tomus 87. Budapest, 1995 p. 35-54 Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts., Hungary) by A. DULAI, Budapest DULAI, A . (1995): Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts., Hungary). - Annls hist.-nat. Mus. natu. hung. 87: 35-54 Abstract - Szob is very famous for its mollusc fauna, however, several other fossil groups can be found in the yellowish-grey sand, which are hardly known. The free-living, so-called lunulitiform Bryozoa are represented by two species at Szob: Cupuladria vindobonensis BALUK et RADWANSKI and Reussirella haidingeri (REUSS). Up to now, lunulitiform Bryozoa have only been mentioned in fauna-lists at numerous localities in Hungary. The way of life and the ecological characters of lunulitiform Bryozoa are well known from studies on the recent representatives of free-living Bryo- zoa. Thus this group is suitable for the reconstruction of the paleoenvironment of Szob. Lunulitiform Bryozoa choose small-particle sand bottom below the wave base and they live in normal or slightly reduced salt water in tropical or subtropical climate. Lunulitiform species adapted themselves to life on soft sediments, where they may be the dominant elements among Bryozoa. With 28 figures and 7 tables. INTRODUCTION The research of Bryozoa is a very neglected area of Hungarian paleontology. This fact holds especially true for the Miocene Bryozoa of Hungary, although important data have been lost because of the lack of such studies, as Bryozoa are excellent environment indica- tors. The aim of this paper is to show the free-living, so-called lunulitiform Bryozoa of the famous Szob locality. Bryozoa are generally fixed marine animals. The special feature of the examined group is its free-living habitude. "Lunulitiform" is an old-established term derived from the name of Lunulites. However, sometimes it had different meanings in the works of different authors. This paper follows BALUK & RADWANSKI'S (1984a) classification in which three families belong to lunulitiform Bryozoa (Cupuladriidae, Discoporellidae, Lunulitidae). Lunulitiform Bryozoa appeared in the Late Cretaceous and live up to now. The way of life and the ecological characters of lunulitiform Bryozoa are well known from studies on their recent representatives. The type species of genus Cupuladria, C. canadensis (BUSK), is one of the most minutely examined Bryozoa species. Lunulitiform Bryozoa were studied extensively in the last few decades by European speci- alists. The family Cupuladriidae was defined by LAGAAU (1952). The vertical distribution of lunulitiform Bryozoa in the Tertiary of the Netherlands was examined by LAGAAU (1953). A very accurate biological and ecological study was made by LAGAAU (1963) on recent Cupu- ladria canadensis (BUSK). ANNOSCIA (1963) mentioned three lunulitiform species from the Miocene deposits of Italy. The external morphological characters of Cupuladriidae have been discussed in detail by COOK (1965a, b). Lunulitiform Bryozoa species were divided into four groups by COOK (Cupuladria canadensis group, Cupuladria guineensis group, Cupuladria owenii group, Discoporella umbellata group). HAKANSSON (1973) examined the internal morphology of Cupuladria biporosa CANU et BASSLER by orientated thin sections and a very
Transcript of DULAI, A. (1995): Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts.,...

A N N A L E S H I S T O R I C O - N A T U R A L E S M U S E I N A T I O N A L I S H U N G A R I C I Tomus 87. Budapest , 1995 p . 3 5 - 5 4
Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts., Hungary)
by A. D U L A I , Budapest
D U L A I , A . (1995): Middle Miocene (Badenian) lunulitiform Bryozoa from Szob (Börzsöny Mts., Hungary). - Annls hist.-nat. Mus. natu. hung. 87: 35-54
Abs t rac t - Szob is very famous for its mollusc fauna, however, several other fossil groups can be found in the yellowish-grey sand, which are hardly known. The free-living, so-called lunulitiform Bryozoa are represented by two species at Szob: Cupuladria vindobonensis BALUK et RADWANSKI and Reussirella haidingeri (REUSS). Up to now, lunulitiform Bryozoa have only been mentioned in fauna-lists at numerous localities in Hungary. The way of life and the ecological characters of lunulitiform Bryozoa are well known from studies on the recent representatives of free-living Bryozoa. Thus this group is suitable for the reconstruction of the paleoenvironment of Szob. Lunulitiform Bryozoa choose small-particle sand bottom below the wave base and they live in normal or slightly reduced salt water in tropical or subtropical climate. Lunulitiform species adapted themselves to life on soft sediments, where they may be the dominant elements among Bryozoa. With 28 figures and 7 tables.
I N T R O D U C T I O N
The research of Bryozoa is a very neglected area o f Hungarian paleontology. This fact holds especially true for the Miocene Bryozoa of Hungary, although important data have been lost because of the lack of such studies, as Bryozoa are excellent environment indicators.
The aim of this paper is to show the free-living, so-called lunulitiform Bryozoa of the famous Szob locality. Bryozoa are generally fixed marine animals. The special feature o f the examined group is its free-living habitude. "Lunulitiform" is an old-established term derived from the name of Lunulites. However, sometimes it had different meanings in the works o f different authors. This paper follows B A L U K & R A D W A N S K I ' S (1984a) classification in which three families belong to lunulitiform Bryozoa (Cupuladriidae, Discoporellidae, Lunulitidae). Lunulitiform Bryozoa appeared in the Late Cretaceous and live up to now.
The way of life and the ecological characters of lunulitiform Bryozoa are well known from studies on their recent representatives. The type species o f genus Cupuladria, C. canadensis ( B U S K ) , is one of the most minutely examined Bryozoa species.
Lunulit iform Bryozoa were studied extensively in the last few decades by European specialists. The family Cupuladriidae was defined by L A G A A U (1952). The vertical distribution of lunulitiform Bryozoa in the Tertiary of the Netherlands was examined by L A G A A U (1953). A very accurate biological and ecological study was made by L A G A A U (1963) on recent Cupuladria canadensis ( B U S K ) . A N N O S C I A (1963) mentioned three lunulitiform species from the Miocene deposits of Italy. The external morphological characters of Cupuladriidae have been discussed in detail by C O O K (1965a, b). Lunulitiform Bryozoa species were divided into four groups by C O O K (Cupuladria canadensis group, Cupuladria guineensis group, Cupuladria owenii group, Discoporella umbellata group). H A K A N S S O N (1973) examined the internal morphology of Cupuladria biporosa C A N U et BASSLER by orientated thin sections and a very

regular internal structure was found with the repetition of a few basic elements. Miocene Bryozoa (of which 5 species belong to lunulitiform Bryozoa) of N W Germany were described by B U G E (1973). C A D É E (1975) described seven recent lunulitiform Bryozoa from the Guyana shelf, from which two were new. Three Miocene species were mentioned by C A D É E (1977) from West Germany. Miocene Bryozoa of Austria were summarized by V Á V R A (1977, 1979). Cupuladria canadensis was divided into three subspecies by C A D É E (1979, 1981). Badenian (Middle Miocene) lunulitiform Bryozoa of Korytnica Basin and Vienna Basin were examined by B A L U K & R A D W A N S K I (1977b, 1984a, b). In their later work a new family (Discoporellidae), a new genus (Reussirella) and a new species {Cupuladria vindobonensis) were described. A SEM atlas of type and figured material from L A G A A U (1952) was presented by B I S H O P & H A Y W A R D (1989). Five lunulitiform species were mentioned from the Pliocene deposits of SW Spain by R E G U A N T (1993).
L O C A L I T Y
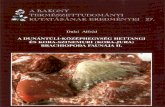
The locality is situated at the SW slopes of the Börzsöny Mountains, N Hungary. The old sandpit is N W of Szob on the left bank of Ipoly river, where the Damásd stream crosses the high road between the villages Szob and Ipolydamásd (Fig. 1). In the older literature this locality was mentioned as "Szob, Nagyfeltárás" (big outcrop).
The fossils of the yellowish-grey sand were already studied from the mid-nineteenth century owing to its rich mollusc fauna. After many smaller papers the mollusc fauna was examined monographically by C S E P R E G H Y - M E Z N E R I C S (1956). A short list is given in her work about the foraminifers of the locality. M Ü L L E R (1984) wrote about some decapods from Szob. M A N -
Z O N I (1877) mentioned Cupularia haidingeri Bryozoa species from the locality. In the same way as in the publications, mollusc centralisation is a characteristic feature of
the the collection of the Department of Geology and Paleontology of the Hungarian Natural History Museum, where molluscs form more than 90 per cent of the material of Szob.
MATERIAL AND METHODS
The examined material is derived from the coarse fraction (>2 mm) of 10-12 kg washed sand of Szob. The composition of the more than 10,000 specimens is the following (Fig. 2a): bivalves are the most frequent (46.57%), gastropods are significant (25.82%), but more than one-quarter of the fossils are composed of the other groups (27.6%).
Taking the other fossil groups into consideration (Fig. 2b) it can be seen that worm tubes are the most frequent (45.4%). Bryozoa (27.92%) and vertebrates (otoliths, fish-teeth and bones: 19.46%) are also significant, but decapods (4.41%) and scaphopods (2.75%) are less important.
Within Bryozoa (Fig. 2c) the number of lunulitiform Bryozoa is more than three times larger than that of all the other Bryozoa. Two lunulitiform species can be found in the Szob material: Cupuladria vindobonensis BALUK et RADWANSKI and Reussirella liaidingeri (REUSS).
The specimens are deposited in the Department of Geology and Paleontology of the Hungarian Natural History Museum in Budapest (inventory numbers: M.94.154.-M.94.175.).
Measurements of Bryozoa were taken according to COOK'S (1965a) methods. The length of zooecia was measured from the distal edge of the aperture to the distal edge of the next succeeding radial zooecial aperture, including the vibraculum. Mean, range and standard deviation of 50 measurements are given (5 zooecia of 10 specimens) for both species (Tables 1-2). One hundred measurements of the number of basal pores are given for Cupuladria vindobonensis BALUK et RADWANSKI (10 basal sectors of 10 specimens) (Table 3).
For Bryozoa terminology BASSLER (1953) and COOK (1965a, b) are followed.

SYSTEMATIC DESCRIPTIONS
Class: Bryozoa EHRENBERG, 1831 Subclass: Gymnolaemata A L L M A N , 1856
Order: Cheilostomata B U S K , 1852 Family: Cupuladriidae LAGAAD, 1952
Genus: Cupuladria C A N U et BASSLER, 1919
Cupuladria vindobonensis B A L U K et R A D W A N S K I , 1984 (Figs 4-10, 23, 27-28)
Cupularia canadensis BUSK: MANZONI 1877: 72, PI. 17, Figs 56a-c. Cupuladria canariensis (BUSK): LAGAAIJ 1952: 33, PI. 2, Figs la-b; LAGAAIJ 1953: 15, Pl. 1, Figs la-b; BUGE 1957:
139, Pl. 9, Fig. 5, Pl. 10, Fig. 3; LAGAAIJ 1963: 172, Pl. 25, Figs la-b, 3-5; ANNOSCIA 1963: 225, Pl. 9, Fig. 1, Pl. 10, Fig. 1, Pl. 11, Figs la-b, Pl. 12, Figs la-b; CHEETHAM & SANDBERG 1964: 1021, Figs 11, 13; COOK 1965b: 197, Figs la-f, Pl. 3, Fig. 4; BUGE 1973: 36, Pl. 6, Figs 1-2; CADÉE 1975: 323, Pl. 3d; BALUK & RADWANSKI 1977b: 143, Pl. 1, Figs 1-6, Pl. 2, Figs 1-5, Pl. 3, Figs. 1-2; CADÉE 1977: 45, Pl. 1, Fig. 3
Cupuladria canariensis canariensis (BUSK): CADÉE 1979: 446, Figs lc-d. Cupuladria canariensis cavernosa CADÉE 1979: 445, Figs le-g, Fig. 4; BISHOP & HAYWARD 1989: 8, Figs 18-21. Cupuladria vindobonensis BALUK & RADWANSKI 1984a: 22, Pis 2-3, PI. 9, Figs 1-2; BALUK & RADWANSKI 1984b:
243, Pis 1-4; REGUANT 1993: p.127.
M a t e r i a l : Numerous fragmentary specimens.
D i m e n s i o n s : length of zooecia 0.637±0.0324 mm (range: 0.59-0.70 mm), width of zooecia 0.276+0.013 mm (range: 0.22-0.31 mm), length of vibracular opesia 0.206+0.01 mm (range: 0.18-0.27 mm), number of pores per basal sectors 8.56± 2.99 (range: 2-17) (see details in Tables 1 and 3).
D e s c r i p t i o n : The discoidal, flattened cone-shaped zoaria are free in adult stages. Zooecia form a single layer, in which vibracula alternate with zooecia in the same radial series. The asymmetrical vibracula aie distal to each zooecia. The shape of zooecia is rounded rectangular. Central zooecia arc open, without closures by a calcareous lamina. Vestibular arch is not developed. The cryptocyst is simple, without any denticles. Ovicells are absent. Opesia are deprimed laterally. The number of basal kenozoidal chambers are between 1-5. The kenozoecial layer is about as thick as the zooecial layer. The basal surface of the colony is sectored by radial and tangential grooves. The majority of the basal sectors are long, with pores ranging usually between 5-13 and their maximum range observed 2 to 17 in a sector.
R e m a r k s : The family Cupuladriidae was described by L A G A A I J (1952) to include only the type genus Cupuladria. Later Discoporella also was classified to family Cupuladriidae by C H E E T H A M & SANDBERG (1964), C O O K (1965a) and B U G E (1973) but recently B A L U K & R A D W A N S K I (1984a) returned to the original interpretation and Discoporella was placed into a new family (Discoporellidae B A L U K et R A D W A N S K I ) .
According to C O O K (1965a) three species belong to her Cupuladria canariensis group, i.e. C. canariensis ( B U S K ) , C. monotreinata ( B U S K ) and C. biporosa C A N U et BASSLER) . This group was supplemented by C A D É E (1975) and B A L U K & R A D W A N S K I (1984a, b) with two later described species (C. surinamensis C A D É E , C. vindobonensis B A L U K et R A D W A N S K I ) .
A new subspecies was described by C A D É E (1979) for European Miocene-Pliocene Cupuladria canariensis (C. canariensis cavernosa C A D É E ) on the basis of the number o f kenozoecial chambers in the basal pad. Recent C. canariensis (BUSK) has only one kenozoecial chamber in a row but in the fossil forms there are more than one chambers in a row. In C A D É E ' S interpretation C. canariensis canariensis ( B U S K ) has 5.6-14.2 (1-28) and C. canadensis cavernosa ( C A D É E ) has 4.5-8.1 (1-28) pores at basal sectors.
However, according to B A L U K & R A D W A N S K I (1984a, b) the number of chambers depends on the thickness of the basal pad and is ascribable to ecological factors without any taxonomic value. Instead of this a new species was described (C. vindobonensis B A L U K et R A D W A N S K I )

Figs 4-6. Cupuladria vindobonensis BALUK et RADWANSKI: 4 = frontal view of M.94.154x240 (the pail marked with an arrow is magnified in Fig. 6), 5 = frontal view of M.94.155 x600 (note communication pores leading to
the basal kenozoidal chambers), 6 = magnified part of colony presented in Fig. 4

Figs 7-10. Cupuladria vindobonensis BALUK et RADWANSKI: 7 = basal view of M.94.156.x270 (the part marked with an arrow is magnified in Fig. 10), 8 = basal view of M.94.157.x240, 9 = basal view of M.94.158.x270,
10 = magnified part of colony presented in Fig. 7

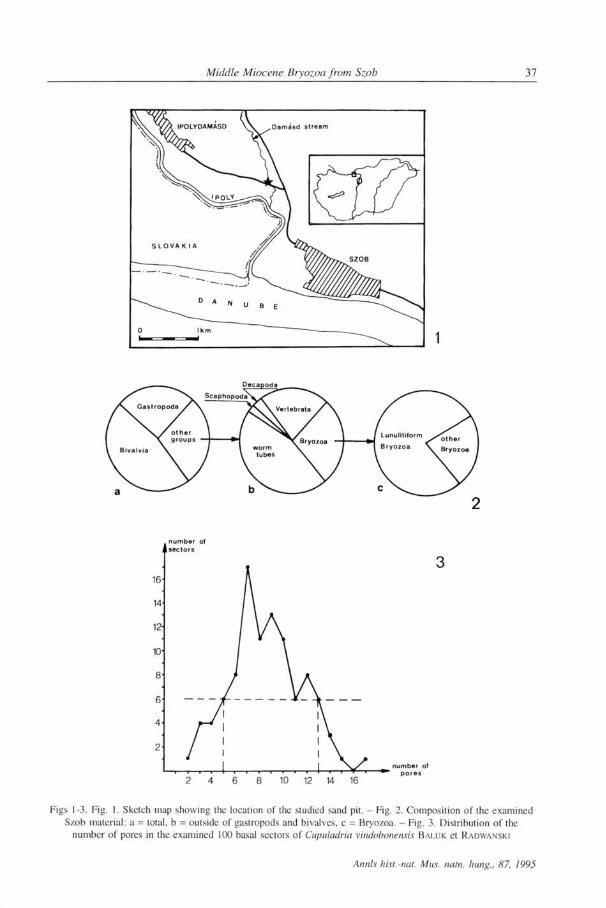
Figs 11-13. Reussirella haidingeri (REUSS): 11 = frontal view of M.94.159.x200 (the parts marked with an arrow are magnified in Figs 13 and 16), 12 = frontal view of M.94.160.X240 (the part marked with an arrow is
magnified in Fig. 25), 13 = magnified part of colony presented in Fig. 11

on the basis of the number of pores at basal sectors. This species contains all the European fossil specimens described as C. canariensis (BUSK). In their interpretation C. canariensis (BUSK) has 3-4 (1-6) and C. vindobonensis BALUK et RADWANSKI has 6-12 (1-28) pores at basal sectors.
The author identifies himself with this idea because the classification of this genus is mainly based on the number of pores at basal sectors. But it seems that the two interpretations have a very similar meaning.
The length of zooecia in the Szob specimens is very similar to C O O K ' S (1965b) data but slightly larger than given by CADÉE (1979, 1981) and R E G U A N T (1993) (Table 4). The width of zooecia is very similar to REGUANT's (1993) measurements but more or less smaller than reported by C O O K (1965b) and C A D É E (1979, 1981). The length of vibracular opesia slightly larger than indicated by COOK (1965b) and R E G U A N T (1993).
The number and shape of kenozoecial chambers in the Szob specimens can be seen on Figs 27-28. The number of kenozoecial chambers is maximum 8 in the Poland material ( B A L U K & R A D W A N S K I 1984b) and 3-4 in Vienna Basin ( B A L U K & R A D W A N S K I 1984a).
The number of pores per basal sector is 5-13 (2-17) in the Szob material (Fig. 3) which is very similar to the data of literature (Table 5). Regenerated specimens cannot be seen because of the very fragmentary preservation of the material. The diameter of the colonies is also unknown because of the incomplete colony preservation. The largest fragment is 5.2 mm long which refers to a size about 11-12 mm in diameter.
D i s t r i b u t i o n : This species (partly as C. canariensis (BUSK) was mentioned from the Transylvanian Basin (MANZONI 1877, COOK 1965b), Korytnica Basin (BALUK & RADWANSKI 1984b) and Vienna Basin (summarized by BALUK & RADWANSKI 1984a). Other occurrences outside the Central-Paratethys: Spain, the Netherlands, Germany, Denmark, Belgium (Miocene) and Spain, France, Great Britain, Belgium, the Netherlands, Italy (Pliocene).
Family Discoporellidae B A L U K et R A D W A N S K I , 1984
Genus Reussirella B A L U K et R A D W A N S K I , 1984
Reussirella haidingeri (REUSS, 1847) (Figs 11-22, 24-26)
Lunulites Haidingeri REUSS 1847: 58, PI. 7, Figs 26-27. Cupularia Haidingeri (REUSS): MANZONI 1877: 71, PI. 16, Fig. 54. Cupuladria haidingeri (REUSS): LAGAAIJ 1952: 35, PI. 2, Figs 2a-b; LAGAAIJ 1953: 16, Pl. 1, Figs 2a-b; BUGE 1957:
140, PI. 9, Figs 6-7; BUGE 1973: 36, Pl. 6, Figs 3-4; BALUK & RADWANSKI 1977b: 144, PI. 4, Figs 1-7; CADÉE 1977: 45, Pl. 1, Figs 2a-b.
Reussirella haidingeri (REUSS): BALUK & RADWANSKI 1984a: 25, Pis 4-6, PI. 10; BALUK & RADWANSKI 1984b: 246, Pis 5-6; BISHOP & HAYWARD 1989: 8, Figs 22-25; REGUANT 1993: 128, Pl. 1, Figs 5-6.
M a t e r i a 1: 38 more or less complete colonies and numerous fragmentary specimens.
D i m e n s i o n s: length of zooecia 0.545+0.0218 mm (range 0.47-0.61 mm), width of zooecia 0.267+0.0133 mm (range 0.21-0.30 mm), length of vibracular opesia 0.125+0.0083 mm (range 0.11-0.15 mm), diameter of colony 2.89+0.532 mm (range: 1.89-3.86 mm) (see details in Tables 2 and 6).
D e s c r i p t i o n : Zoaria are free in adult stages. Zooecia with asymmetrical vibracula situated distal to each zooecium in the same radial series. The shape of zooecia is rounded rhombic. Vicarious vibracula is not observed. Central zooecia are closed by a calcareous lamina. Zooecia are accompanied by well-developed vestibular arch. Zooecia are partly closed by a calcareous cryptocyst, with characteristic dentated opening. Ovicells cannot be observed. Kenozoidal chambers are absent within the basal pad. The zooecial layer is thicker than the basal pad. The concave side of the colony has radial grooves only. The surface between the grooves is covered by small tubercules.

R e m a r k s : B A L U K & R A D W A N S K I (1984a) described the new genus Reussirella for the members of C O O K ' S (1965a) Cupuladria owenii group on the basis of closed central zooecia. According to the describers this genus contains five species i.e. R. haidingeri (REUSS) , R. reussiana ( M A N Z O N I ) , R. muliispinata ( C A N U et BASSLER), R. doma (D ' ORBIGNY) and R. owenii ( G R A Y ) . The closed central zooecia refer to the relationship between Reussirella and Discoporella; for this reason, B A L U K & R A D W A N S K I (1984a) described the family Dis-coporellidae containing the two former genera. The calcareous lamina covering the central zooecia are interpreted as a passive excurrent chimney by B A L U K & R A D W A N S K I (1984b).
This species was described from the Vienna Basin by REUSS (1847) and it is known only from the European Neogene deposits, so it is an extinct endemic species. According to some authors, it may be only badly preserved Discoporella umbellata (DEFRANCE) but it is not a real assumption. This species can frequently be found together with Cupuladria vindobonensis B A L U K & R A D W A N S K I and/or Discoporella umbellata ( D E F R A N C E ) .
In sections it can be seen that kenozooecial chambers are absent within the basal pad (Fig. 26). Autozooecia sometimes descend near to the basal surface.
There are nearly 40 more or less complete specimens in the Szob material but the majority of the specimens are fragmentary. The regeneration o f the colonies is well visible in some specimens but the fragments of primary colonies are absent at the basal surfaces, in contrast to the Korytnica material ( B A L U K & R A D W A N S K I 1977b). The irregular or elongated shape of the colony and the arrangement of the zooecia refer to regeneration (Figs 17-18, 20-22, 24). Most of the complete specimens are rather small but the biggest fragment is 4.5 mm which refers to a size of about 9-10 mm in diameter.
Measurements on Reussirella haidingeri (REUSS) were given by R E G U A N T (1993). The length of zooecia in the Szob specimens is very similar to the Spanish material, however the width of zooecia and the length of vibracular opesia are smaller than in R E G U A N T ' S Pliocene specimens (Table 7).
D i s t r i b u t i o n : This species is known from the European Miocene and Pliocene sediments. In the area of Central Paratethys it was mentioned from the Vienna Basin (summarized by BALUK & RADWANSKI 1984a), Transyl-vanian Basin (MANZONI 1877), Carpathian Foredeep (MALECKI 1951), Korytnica Basin (BALUK & RADWANSKI 1977b, 1984b) and Pannonian Basin (MANZONI 1877). Over and above these Badenian localities this species was found in Austrian Eggenburgian sediments by VÁVRA (1977).
Other occurrences of Reussirella haidingeri outside the Central Paratethys: Mallorca, France, Italy, Germany (Miocene) and Mallorca, France, Belgium, the Netherlands, Great Britain, Portugal, Tunesia (Pliocene).
LIFE M O D E O F L U N U L I T I F O R M BRYOZOA
A t the end of the free-swimming larval stage a fine-grained, sandy bottom below the wave base is ideal for settlement. The larva settles on a small grain (foraminifers, quartz-grains). During a quick metamorphosis takes shape the ancestrular triplet, namely three zooecia form at the same time from the larval stage ( H A K A N S S O N 1973). From that moment the colony develops by radial budding ( L A G A A I J 1963). During growth the colony expands beyond the grain and becomes free-living. The developing colonies form a flattened cone, which shape hampers sinking of the colony in the soft sediment. Lunulitiform species well adapted themselves to life on soft sediments, where they may be the dominant elements among Bryozoa.
The investigations of recent species in aquarium and in natural marine environments confirm that the colonies lie on the bottom in apex-up position (GREELY 1967, 1970). Recently at the Guyana shelf the density of lunulitiform Bryozoa somewhere may be around 15,500 specimens/m ( C A D É E 1975). On the other hand, G R E E L Y (1967) mentioned 10 specimens/trT from Mississippi Sound.
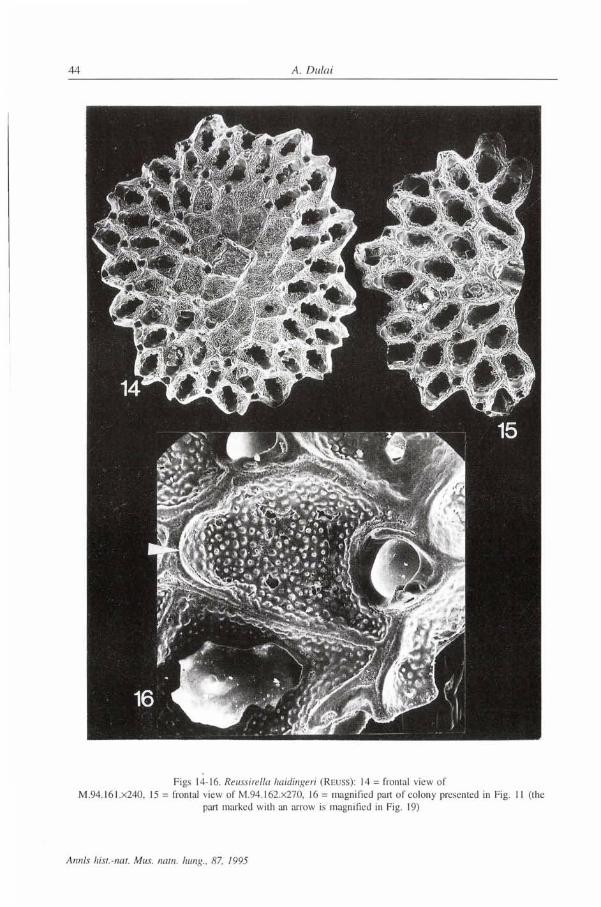
Figs 14-16. Reussirella haidingeri (REUSS): 14 = frontal view of M.94.161.x240, 15 - frontal view of M.94.162.x270, 16 = magnified part of colony presented in Fig. 11 (the
part marked with an arrow is magnified in Fig. 19)

Figs 17-19. Reussirella haidingeri (Rnuss): 17 = basal view of regenerated colony of M.94.163.x300, 18 = basal view of regenerated colony of M94.164.x390, 19 = magnified part of colony presented in Fig. 16
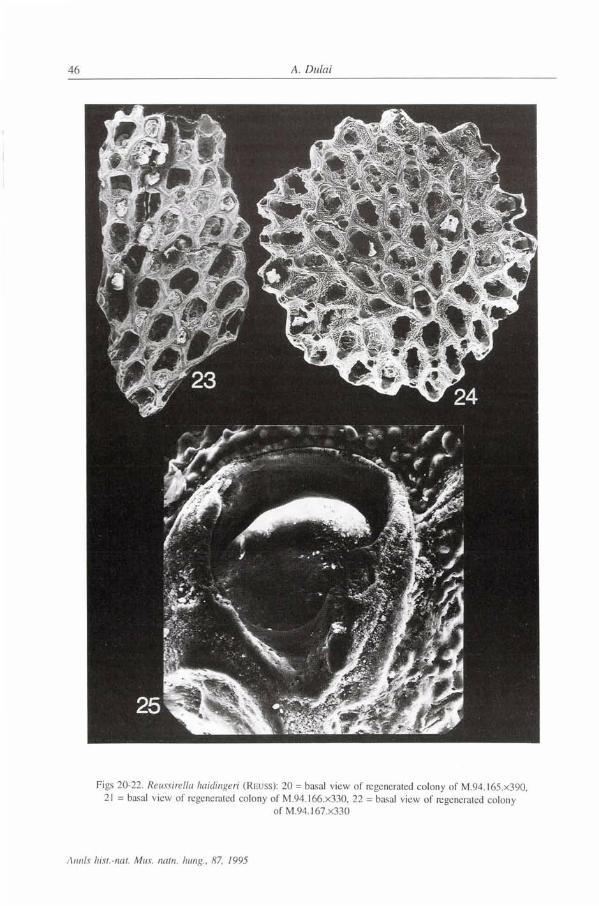
Figs 20-22. Reussirella haidingeri (REUSS): 20 = basal view of regenerated colony of M.94.165.x390, 21 = basal view of regenerated colony of M.94.166.X330, 22 = basal view of regenerated colony
of M.94.167.X330

Figs 23-25. Fig. 23. Cupuladria vindobonensis BALUK et RADWANSKI, frontal view of M.94.168.X240. - Figs 24-25. Reussirella haidingeri (REUSS): 24 = frontal view of
regenerated colony of M.94.169.x270, 25 - magnified part of colony presented in Fig. 12


The colonies of lunulitiform Bryozoa are very fragile, so unbroken specimens can hardly be found. I f the colony get hurt during the life of the animal, it can regenerate itself from a very small fragment. Injuries to colonies are caused by attacks of predators or different activities of benthic animals (holothurians, sea-urchins, hermit crabs) ( L A G A A I J 1963 , GREELY 1967) .
ECOLOGICAL CHARACTERS OF L U N U L I T I F O R M B R Y O Z O A
The fine-grained sandy bottom is ideal for lunulitiform Bryozoa. According to L A G A A I J (1963), recent species can never be found at a purely clayey bottom. In contrast, B A L U K & R A D W A N S K I (1977a, b, 1984b) desribed 3 lunulitiform Bryozoa from Miocene Korytnica Clay of Poland. So lunulitiform species can tolerate a small amount o f clay sedimentation.
The minimum tolerable water depth varies from place to place as a function of the intensity of water turbulence over the bottom (wave base, bottom currents). Lunulitiform Bryozoa cannot live in the littoral zone, they live only at the stable bottoms below wave base. For example, in recent seas they cannot be found between 0-10 m ( L A G A A I J 1963) or between 0-20 m ( C A D É E 1975). A bottom which is shallower but densely covered by sea grasses was mentioned for lunulitiform Bryozoa in Korytnica Basin ( B A L U K & R A D W A N S K I 1977a, b 1984b, and H O F F M A N 1977). The maximum water depth depends on the temperature of the bottom water.
According to L A G A A I J (1963), lunulitiform Bryozoa can be found at water temperatures between 12-30 ° C . The recent geographical distribution of Cupuladria canariensis ( B U S K ) is limited by the 14 ° C surface isocrymes, so the species refers to tropical-subtropical climate. The same holds true of Discoporella umbellata ( D E F R A N C E ) ( C O O K 1965a).
Lunulitiform Bryozoa mostly live in normal salt water but in shallow water they can tolerate a reduced salt content down to 28%o. However, higher than normal salinity is absolutely unacceptable for lunulitiform species.
PALEOBIOGEOGRAPHY
Lunulitiform Bryozoa were mentioned from some localities in the area of Central Paratethys. B A L U K & R A D W A N S K I (1984a) summarized the Badenian lunulitiform species o f the Vienna Basin and six species was described (Cupuladria canariensis ( B U S K ) , C. vindobonensis B A L U K et R A D W A N S K I , Reussirella haidingeri (REUSS), Reussirella reussiana ( M A N Z O N I ) , Discoporella umbellata (DEFRANCE), Lunulites androsaces M A N Z O N I ) . Three lunulitiform Bryozoa have been reported from Poland by M A L E C K I (1951) and B A L U K & R A D W A N S K I (1984b) ( C . vindobonensis B A L U K et R A D W A N S K I , R. haidingeri (REUSS), L. androsaces M A N Z O N I ) . Cupuladria canariensis ( B U S K ) was mentioned from the Transylvanian Basin by C O O K (1965b), however these specimens were regarded as C. vindobonensis by B A L U K & R A D W A N S K I (1984a, b).
Up to now lunulitiform Bryozoa have only been mentioned in fauna-lists in Hungary. In many cases the determination stopped at generic level. For this reason, researchers of the paleobiogeographic distribution of this group do not indicate the occurrence o f lunulitiform Bryozoa from Hungary. In some papers Eisenstadt in the Vienna Basin appeared as a part of Hungary based on C A N U & BASSLER's (1924) work where this recently Austrian town was mentioned as a Hungarian locality. One of the rare exceptions is constituted by the works of B A L U K & R A D W A N S K I (1984a, b), in which Reussirella haidingeri was mentioned from Szob, referring to M A N Z O N I (1877).
However, a short study of the literature wi l l show us that lunulitiform Bryozoa have been found in numerous Neogene localities of Hungary. Some examples, without a claim to completeness:

Cupularia Haidingeri, Szob ( M A N Z O N I 1877); Cupularia cfr. haidingeri, Rákospalota ( F R A N Z E N A U 1910); Lunulites sp., Nógrádszakái ( B O G S C H 1936); Lunulites sp., Balassagyarmat ( N O S Z K Y 1940); Cupularia (Lunulites) haidingeri, Acsa, Fót, Püspökhatvan ( N O S Z K Y 1940); Lunulites sp., Rétság ( B Á L D I et al. 1964); Lunulites sp., Fót, Csornád ( H Á M O R 1985).
These data indicate that lunulitiform Bryozoa are fairly common in the northern part of the Pannonian Basin. In the future the examination of the whole Pannonian Basin would be very instructive and interesting.
Lunulitiform Bryozoa are more common in the Pannonian Basin than it was reflected in literature. A more detailed examination of this group is desirable in the area of the Pannonian Basin.
Very useful data were given of the paleoenvironmental characteristics of Szob on the basis of the ecological features of lunulitiform Bryozoa before the examination of molluscs. Lunulitiform Bryozoa refer to soft sandy bottom below the wave base in normal or slightly reduced salty water in the tropical or subtropical climate.
A c k n o w l e d g m e n t s - This study was supported by the Hungarian National Scientific Research Fund (OTKA No. F 13975). The SEM photos were taken in the SEM Laboratory of the Hungarian Geological Survey by Mrs. TAKÁCS and Mrs. PELLÉRDY.
Table 1. Measurements of Cupuladria vindobonensis B A L U K et RADWANSKI (5 zooecia of 10 specimens). Key to the signs used: Lz: length of zooecia (mm);
lz: width of zooecia (mm); lo: length of vibracular opesia (mm); SD: standard deviation.
CONCLUSIONS
* * *
3 4 5 mean SD 0.044 0.011 0.019 0.029 0.029 0.008 0.030 0.013 0.016 0.021 0.007 0.010 0.097 0.018 0.005 0.013 0.013 0.008 0.015 0.007 0.011
1. Lz: lz: lo:
2. Lz: lz: lo:
3. Lz: lz: lo:
4. Lz: lz: lo:
5. Lz: lz: lo:
6. Lz: lz: lo:
7. Lz: lz: lo:
0.70 0.27 0.27 0.64 0.28 0.20 0.64 0.28 0.21 0.59 0.27 0.21 0.63 0.25 0.19 0.64 0.28 0.18 0.65 0.29 0.24
0.64 0.25 0.27 0.60 0.27 0.19 0.60 0.31 0.22 0.63 0.28 0.21 0.63 0.24 0.19 0.65 0.26 0.18 0.66 0.29 0.23
0.68 0.25 0.27 0.60 0.29 0.19 0.61 0.30 0.19 0.64 0.29 0.22 0.59 0.28 0.18 0.65 0.28 0.20 0.65 0.30 0.25
0.59 0.27 0.24 0.67 0.22 0.18 0.67 0.31 0.21 0.60 0.28 0.19 0.60 0.27 0.18 0.67 0.26 0.19 0.62 0.28 0.23
0.63 0.25 0.23 0.63 0.29 0.20 0.60 0.31 0.18 0.60 0.28 0.21 0.59 0.24 0.19 0.67 0.25 0.18 0.64 0.29 0.22
0.648 0.258 0.256 0.628 0.27 0.192 0.624 0.302 0.202 0.612 0.28 0.208 0.608 0.256 0.186 0.656 0.266 0.186 0.644 0.29 0.234

1 2 3 4 5 mean SD 8. Lz: 0.62 0.68 0.67 0.65 0.62 0.648 0.027
lz: 0.28 0.29 0.28 0.31 0.27 0.286 0.015 lo: 0.21 0.21 0.19 0.19 0.21 0.202 0.010
9. Lz: 0.68 0.63 0.61 0.65 0.64 0.642 0.025 lz: 0.27 0.27 0.27 0.28 0.25 0.268 0.010 lo: 0.19 0.21 0.21 0.19 0.19 0.198 0.010
10. Lz: 0.64 0.63 0.68 0.67 0.68 0.66 0.023 lz: 0.29 0.29 0.27 0.29 0.28 0.284 0.008 lo: 0.20 0.19 0.19 0.20 0.19 0.194 0.005
Total mean range
Lz: 0.637 0.59-0.70 (
0.276
0.206
0.22-0.31
0.18-0.27
SD
0.0324
0.013
0.010
Table 2. Measurements of Reussirella haidingeri (REUSS) (5 zooecia of 10 specimens). Key to the signs used: the same as in Table 1
1 2 3 4 5 mean SD 1. Lz: 0.50 0.48 0.52 0.55 0.54 0.514 0.028
lz: 0.24 0.21 0.23 0.27 0.24 0.238 0.021 lo: 0.14 0.12 0.12 0.12 0.13 0.126 0.008
2. Lz: 0.48 0.51 0.53 0.50 0.47 0.498 0.023 lz: 0.21 0.25 0.25 0.26 0.25 0.244 0.019 lo: 0.11 0.12 0.12 0.12 0.12 0.118 0.004
3. Lz: 0.55 0.58 0.56 0.53 0.53 0.55 0.021 lz: 0.30 0.27 0.27 0.28 0.28 0.28 0.012 lo: 0.15 0.14 0.14 0.12 0.12 0.134 0.013
4. Lz: 0.57 0.55 0.54 0.56 0.58 0.56 0.015 lz: 0.27 0.29 0.29 0.28 0.30 0.286 0.011 lo: 0.12 0.12 0.12 0.14 0.13 0.126 0.008
5. Lz: 0.57 0.56 0.58 0.56 0.55 0.564 0.011 lz: 0.30 0.29 0.29 0.28 0.28 0.288 0.008 lo: 0.14 0.13 0.14 0.14 0.13 0.136 0.005
6. Lz: 0.57 0.56 0.53 0.56 0.55 0.554 0.015 lz: 0.29 0.29 0.27 0.27 0.27 0.278 0.010 lo: 0.14 0.14 0.12 0.13 0.14 0.134 0.017
7. Lz: 0.53 0.58 0.61 0.57 0.59 0.576 0.029 lz: 0.28 0.27 0.28 0.27 0.25 0.27 0.012 lo: 0.13 0.13 0.14 0.12 0.12 0.128 0.008
8. Lz: 0.60 0.56 0.53 0.54 0.53 0.552 0.029 lz: 0.28 0.27 0.27 0.25 0.25 0.264 0.013 lo: 0.12 0.12 0.12 0.14 0.12 0.124 0.008
9. Lz: 0.56 0.54 0.55 0.58 0.61 0.568 0.027 lz: 0.25 0.27 0.26 0.26 0.24 0.256 0.011 lo: 0.12 0.11 0.11 0.13 0.12 0.118 0.008
10. Lz: 0.54 0.52 0.50 0.54 0.50 0.52 0.020 lz: 0.24 0.26 0.27 0.28 0.28 0.266 0.016 lo: 0.12 0.11 0.11 0.11 0.11 0.112 0.004
Total mean range SD
Lz: 0.545 0.47-0.61 0.0218
lz: 0.267 0.21-0.30 0.0133
lo: 0.125 0.11-0.15 0.0083

Table 3. Number of pores per basal sector in Cupuladria vindobonensis B A L U K et R A D W A N S K I (10 sectors of 10 specimens)
mean range SD 1. 13, 7, 5, 10, 9, 13, 12, 8, 11, 10 9.8 5-13 2.61 2. 8, 10, 11, 6, 8, 13, 12, 9, 3, 7 8.7 3-13 2.98 3. 14, 13, 4, 10, 7, 11, 8, 6, 12, 7 9.2 6-14 3.29 4. 10, 8, 7, 11, 9, 9, 3, 9, 6, 2 7.4 2-11 2.95 5. 7, 7, 7, 9, 13, 9, 10, 9, 7, 10 8.8 7-13 1.93 6. 7, 5, 4, 10, 6, 10, 8, 4, 7, 9 7.0 4-10 2.26 7. 12, 13, 11, 5, 7, 8, 7, 14, 5, 7 8.9 5-14 3.31 8. 12, 8, 6, 10, 9, 5, 8, 14, 9, 3 8.4 3-14 3.23 9. 15, 8, 12, 11, 6, 3, 6, 9, 4, 9 8.3 3-15 3.71
10. 7, 6, 10, 12, 17, 8, 7, 12, 7, 5 9.1 5-17 3.66
Total: 8.56 2-17 2.99
Table 4. Measurements of Cupuladria vindobonensis B A L U K et R A D W A N S K I according to the literature. Key to the signs used: the same as in Table 1
COOK 1965 b C. canariensis
CADÉE 1979 C. c. canariensis
CADÉE 1979 C. c. cavernosa
CADÉE 1981 C. c. cavernosa
REGUANT 1993 C. vindobonensis
This paper C. vindobonensis
Lz
mean
range
0.5-0.8
0.586-0.666 0.45-0.78
0.517-0.579 0.45-0.66
0.573+ 0.041
0.402 0.372-0.488
0.637 0.59-0.70
lz
mean
range
0.3-0.47
0.334-0.40 0.24-0.51
0.356-0.428 0.24-0.54
0.362+ 0.056
0.295 0.256-0.314
0.276 0.22-0.31
lo
mean
range
0.12-0.19
0.19 0.163-0.267
0.206 0.18-0.27
Table 5. Number of pores per basal sectors of Cupuladria vindobonensis B A L U K et R A D W A N S K I according to the literature
COOK (1965b)
CADÉE (1979)
BALUK & RADWANSKI (1984a)
BALUK & RADWANSKI (1984b)
This paper
C. canariensis
C. c. canariensis
C. c. cavernosa
C. vindobonensis
C. vindobonensis
C. vindobonensis
6-12 (4-20)
4- 14
4.5-8.1
6-12 (2-18)
6-12 (2-20)
5- 13 (2-17)

Table 6. Diameters of more or less complete colonies of Reussirella haidingeri (REUSS) (38 specimens, mm).
3.42 3.24 3.86 3.05 3.73 2.74 2.70 1.89 3.42 3.34 3.66 3.69
3.31 2.18 2.80 2.43 2.48 2.80 2.33 3.60 3.66 3.05 2.38 3.00
2.44 2.56 2.33 2.30 2.65 3.67 2.31 2.44 2.59 2.74 2.82 3.22
2.25 2.77
Total: mean: 2.89 mm; range: 1.89-3.86 mm; SD: 0.532
Table 7. Measurements of Reussirella haidingeri (REUSS) according to the literature. Key to the signs used: the same as in Table 1
Lz lz lo
mean mean mean
range range range
REGUANT 1993 0.523 0.334 0.204
R. haidingeri 0.492-0.585 0.292-0.385 0.185-0.215
This paper 0.545 0.267 0.215
R. haidingeri 0.47-0.61 0.21-0.30 0.11-0.15
REFERENCES
ANNOSCIA, E. (1963): Antozoi e briozoi nelle argille calabriane di Venosa (Potenza). - Geol. Romana 2: 215-237. BALUK, W. & RADWANSKI, A. (1977a): Organic communities and facies development of the Korytnica Basin (Middle
Miocene; Holy Cross Mountains, Central Poland). - Acta Geo!. Polonica 2 7 (2): 85-123. BALUK, W. & RADWANSKI, A. (1977b): The colony regeneration and life habitat of free-living bryozoans, Cupuladria
canariensis (BUSK) and C.haidingeri (REUSS) from the Korytnica Clays (Middle Miocene; Holy Cross Mountains, Poland). - Acta Geol. Polonica 27 (2): 143-156.
BALUK, W. & RADWANSKI, A. (1984a): Middle Miocene (Badenian) free-living bryozoans from the Vienna Basin. -Ann. Naturhist. Mus. Wien 8 6 A : 13-40.
BALUK, W. & RADWANSKI, A. (1984b): Free-living bryozoans from the Korytnica Clays (Middle Miocene; Holy Cross Mountains, Central Poland). - Acta Geol. Polonica 34 (3-4): 239-251.
BASSLER, R. G.(1953): Bryozoa. - In: Treatise on Invertebrate Paleontology G : GI-G253. BÁLDI, T., CSEPREGHY-MEZNERICS, I . & NYÍRÓ, R. (1964): La biostratigraphie des gisements oligocènes et miocènes
de l'Est de la Montagne Börzsöny. - Annls hist.-nat. Mus. natn. hung. 56 : 153-183. BISHOP, J. D. & HAYWARD, P. J. (1989): SEM Atlas of type and figured material from Robert Lagaaij's "The Pliocene
Bryozoa of the Low Counlries"(1952). - Meded. Rijks Geol. Dienst 43 (2): 1-64. BOGSCH, L. (1936): Tortonien fauna Nógrádszakálról. (Tortonische Fauna von Nógrádszakái.) - Ann. Inst. Hung.
Geol. 3 1 ( I ) : 1-112. BUGE, E. (1957): Les Bryozoaires du Néogene de l'ouest de la France. - Me'm. Mus. Nat. hist. Nat., n.s. C 6 : 1-436. BUGE, E. (1973): Les Bryozoaires Miocènes du Nord-Ouest de l'Allemagne. - Paläont. Z. 4 7 (1-2): 32-53. CADÉE, G. C. (1975): Lunulitiform Bryozoa from the Guyana shelf. - Neth. ./. Sea Res. 9 (3-4): 320-343. CADÉE, G. C. (1977): Bryozoa from the Late Miocene Mica Clay of Morsum Cliff, Sylt, Western Germany. - Meded.
Werkgr. Ten. Kwart. Geol. 1 4 (2): 43-50. CADÉE, G. C. (1979): The Cupuladria canariensis complex. - In: LARWOOD, G. P. & ABBOTT, M. B. (cds): Advances
in Bryozoology. Systematics Assoc. Spec. Volumes 13: 443-449. CADÉE, G. C. (1981): Notes on Bryozoa, 1. Cupuladria canariensis from the British Pliocene Coralline Crag. -
Meded. Werkgr. Tert. Kwart. Geol. 1 8 (1): 3-9. CANU, F. & BASSLER, R. S. (1924): Contribution a l'étude des Bryozoaires d'Autriche et de Hongrie. - Bull. Soc.
Géol. France (Se'r. 4) 24: 672-690.

CHEETHAM, A. H . & SANDBERG, P. A. (1964): Quaternary Bryozoa from Louisiana mudlumps. - J. Paleont. 38 (6): 1013-1046.
COOK, P. L. (1965a): Notes on Cupuladriidae (Polyzoa Anasca). - Bull. Brit. Mus. (Nat. Hist.), Zoology 1 3 (5): 153-187.
COOK, P. L. (1965b): Polyzoa from West Africa. The Cupuladriidae (Cheilostomata, Anasca). - Bull. Brit. Mus. (Nat. Hist.), Zoology 1 3 (6): 191-227.
CSEPREGHY-MEZNERICS, 1. (1956): A szobi és letkési puhatestű fauna.(Die Molluskenfauna von Szob und Letkés.) -Ann. Inst. Hung. Geol. 4 5 (2): 361-448.
FRANZENAU, Á . (1910): Középmiocén-rétegeknek új előfordulásáról Budapest környékén, Rákospalotán. (About the new occurrence of Middle Miocene deposits near Budapest, at Rákospalota.) - Földt. Közi 40 : 156-163.
GREELY, R. (1967): Natural orientation of lunulitiform bryozoans. - Geol. Soc. Amer. Bull. 7 8 (9): 1179-1182. GREELY, R. (1970): Life-orientation relationships of some Eocene bryozoan-coral associations. - / . Paleont. 44 (2):
343-345. HAKANSSON, E. (1973): Mode of growth of the Cupuladriidae (Bryozoa, Cheilostomata). - In: LARWOOD, G. P. (ed.):
Living and Fossil Bryozoa. Academic Press, New York, pp. 287-298. HÁMOR, G. (1985): A Nógrád-cserháti kutatási terület földtani viszonyai. (Geology of the Nógrád-Cserhát area.) -
Geol. Hung. Ser. Geol. 22: 1-307. HOFFMAN, A. (1977): Synecology of macrobenthic assemblages of the Korytnica Clays (Middle Miocene; Holy Cross
Mountains, Poland). - Acta Geol. Polonica 2 7 (2): 227-280. LAGAAIJ, R. (1952): The Pliocene Bryozoa of the Low Countries and their bearing on the marine stratigraphy of the
North Sea region. - Mededel. Geol. Stichting (Ser. C.) 5: 1-233. LAGAAIJ, R. (1953): The vertical distribution of the lunulitiform Bryozoa in the Tertiary of the Netherlands. -
Mededel. Geol. Stichting (n. ser.) 7: 13-19. LAGAAIJ, R.(1963): Cupuladria canariensis (BUSK) - portrait of a bryozoan. - Palaeontology 6 (1): 172-217. MALECKI, J. (1951): Contribution a la connaissance des Bryozoaires du Miocène de Benczyn. - Ann. Soc. Géol.
Pologne 1 9 (4): 487-491. MANZONI, A. (1877): I Briozoi fossili del Miocène d'Austria ed Ungheria. - Denkschr. Kais. Akad. Wiss. Math.-Na-
turwiss. Cl. 3 7 (2): 49-78. MÜLLER, P. (1984): A bádeni emelet tízlábú rákjai. (Decapod Crustacea of the Badenian.) - Geol. Hung. Ser. Paleont.
42: 1-317. NOSZKY, J. (1940): A Cserháthegység földtani viszonyai. (Das Cserhát-Gebirge.) - Geol. Besc.hr. Ung. Landsch. 3:
1-283. REGUANT, S. (1993): The Cheilostome Bryozoa from the Huelva Pliocene (SW-Spain) in the Western Meditenanean
context. - Mem. Sei. geol. 45 : 125-138. REUSS, A. E. (1847): Die fossilen Polyparien des Wiener Tertiär-beckens. - Naturwiss. Abh. 2: 1-109. VAVRA, N. (1977): Bryozoa tertiara. - In: ZAPFE, H. (ed.): Catalogus fossilium Austriae, Heft Vb/3. Springer, Österr.
Akad. Wiss., Wien, 210 pp. VÁVRA, N. (1979): Die Bryozoenfaunen des österreichischen Tertiärs. - N. Jb. Geol. Paläont. Abh. 157 (3): 366-392.
Author's address: ALFRÉD D U L A I
Department of Geology and Paleontology Hungarian Natural History Museum H-1088 Budapest, Múzeum körút 14-16 Hungary