
Doctorado en Ciencia Animal Tesis - ridaa unicen
86
I Doctorado en Ciencia Animal Tesis Reproducción experimental y aspectos patológicos e inmunohistoquímicos de bronconeumonías causadas por Rhodococcus equi en potrillos Por: Jorge Pablo García Facultad de Ciencias Veterinarias U.N.C.P.B.A 2020
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Doctorado en Ciencia Animal Tesis - ridaa unicen

I
Doctorado en Ciencia Animal
Tesis
Reproducción experimental y aspectos patológicos e inmunohistoquímicos de bronconeumonías causadas
por Rhodococcus equi en potrillos
Por: Jorge Pablo García
Facultad de Ciencias Veterinarias U.N.C.P.B.A
2020

II
Dedicada a Julia, Maura y Uziel

III
Agradecimientos
Al Dr. Paco Uzal, director de esta tesis, le agradezco profundamente no solo porque marcó un antes y un después en mi vida profesional, sino también por toda su ayuda en muchos momentos difíciles en mi paso por el laboratorio CAHFS de San Bernardino y por el apoyo incondicional en la preparación del examen del ACVP.
A la Dra. Élida Fumuso, co-directora de esta tesis, por brindarme la posibilidad de formar parte de su grupo de trabajo y por el tiempo que me brindó a lo largo de la realización de la misma.
Al Dr. Adolfo Casaro, quien fuera director de esta Tesis Doctoral, por ser una de las personas más influyentes que me motivó tanto en lo profesional como en lo personal.
A la comisión de Doctorado de la UNCPBA y especialmente a los evaluadores externos e internos, cuyos valiosos aportes mejoraron el diseño de los experimentos y la interpretación de los resultados obtenidos.
Y un profundo agradecimiento a todas aquellas personas que, de manera directa o indirecta, se involucraron y colaboraron para que esta tesis fuese posible.

IV
RESUMEN
Rhodococcus equi (R. equi) es un cocobacilo gram positivo, inmóvil e intracelular
facultativo que produce bronconeumonía en potrillos. La infección por Rhodococcus equi se
encuentra distribuida mundialmente y produce un impacto económico-productivo negativo
importante en la industria equina. Esta enfermedad se manifiesta principalmente en potrillos
menores de 6 meses, produciendo bronconeumonía piogranulomatosa asociada con alta morbilidad
como mortalidad. La virulencia de R. equi está determinada por la presencia de un plásmido de entre
85 y 90 kb, que le proporciona la característica de ser intracelular facultativo. La inhalación de
aerosoles que contienen cepas virulentas de R. equi desde el ambiente, y la replicación dentro de los
macrófagos alveolares, son componentes esenciales de su patogénesis. Debido a que solo el modelo
experimental equino reproduce las lesiones pulmonares típicas de esta enfermedad, el mismo ha
sido utilizado en estudios para evaluar la implementación de métodos profilácticos y terapéuticos.
Las lesiones macroscópicas producidas se caracterizan por consolidación multifocal de tamaño
variable con cavitaciones ocupadas con un material purulento. La lesión microscópica que produce
es una bronconeumonía piogranulomatosa caracterizada por abundante cantidad de neutrófilos,
macrófagos, células gigantes multinucleadas tipo Langhans y una menor cantidad de linfocitos y
células plasmáticas.
Los objetivos de la presente tesis fueron:
I: Demostrar que la dosis de 5x10³ UFC/mL de la cepa 103+ de R. equi es capaz de producir
bronconeumonía piogranulomatosa en potrillos menores de 6 meses.
II: Diagnosticar de forma precoz la infección pulmonar mediante la interpretación en conjunto de
los signos clínicos respiratorios, el aumento de temperatura rectal, la elevación de fibrinógeno
sanguíneo y la observación de lesiones pulmonares ultrasonográficas.
III: Caracterizar las lesiones pulmonares macroscópicas y microscópicas de los potrillos infectados
experimentalmente con una dosis baja de la cepa 103+ de R. equi (5x10³ UFC/mL) y confirmar la
respuesta inmune Th-1 característica de esta infección, a través de la utilización del estudio
histopatológico y la técnica de inmunohistoquímica para la detección de linfocitos T (CD3) y
linfocitos B (CD79).
La tesis está dividida en tres capítulos. El capítulo I se basa en el conocimiento reciente de
aspectos relevantes de esta enfermedad, tales como los factores de virulencia, epidemiología,
patogénesis, mecanismo inmune, patología, diagnóstico clínico, tratamiento y pronóstico. El
capítulo II describe la reproducción experimental y aspectos clínicos de la bronconeumonía causada

V
por la cepa 103+ de Rhodococcus equi en potrillos. El capítulo III comprende el estudio
anatomopatológico e inmunohistoquímico de lesiones pulmonares inducidas experimentalmente
por la cepa 103+ de Rhodococcus equi en potrillos.
Este estudio demostró que la inoculación de la cepa 103+ de R. equi con una dosis baja
(5x103 UFC/mL) es capaz de producir exitosamente bronconeumonía en potrillos menores de 6
meses. En cuanto a la detección clínica de la enfermedad, la evaluación conjunta de los signos
clínicos respiratorios, temperatura rectal, fibrinógeno sanguíneo y la observación de lesiones
piogranulomatosas mediante la ultrasonografía torácica, permitieron la detección de la enfermedad
a los 13,5 días -en promedio-, desde la inoculación.
La totalidad de los potrillos desafiados presentaron lesiones pulmonares macroscópicas
similares, caracterizadas por consolidación pulmonar y/o cavitación severa con áreas de
consolidación multifocales a coalescentes en la región cráneoventral y caudodorsal de los
pulmones. En cuanto a las lesiones pulmonares microscópicas, el 100% de los potrillos inoculados
presentó bronconeumonía piogranulomatosa, el 54% presentó proliferación de neumocitos tipo II
(epitelización), el 36% presentó trombosis pulmonar y el 27% presentó fibroplasia pulmonar.
Además, se observaron diversas lesiones extrapulmonares, tales como hipopión, hipema, necrosis
hepática centrolobulillar y miocarditis multifocal.
Este estudio demostró que la inoculación de una dosis baja de la cepa 103+ de R. equi es
capaz de producir lesiones pulmonares similares a las producidas por la infección natural. Sin
embargo, se debería investigar el efecto de una dosis por debajo de 5x103 UFC/mL, la cual
especulamos que podría producir lesiones severas, pero con un intervalo de tiempo mayor entre la
inoculación y los signos clínicos respiratorios, asemejándose aun más a una infección natural.

VI
SUMMARY
Rhodococcus equi is a grampositive, nonmotile, facultative intracellular coccobacillus that
produces bronchopneumonia in foals. Rhodococcosis has a worldwide distribution and causes
important productive and economic losses in the equine industry. The disease typically occurs in
foals less than 6 months of age, causing mainly pyogranulomatous bronchopneumonia
associatedwith high morbidity and mortality. The virulence of Rhodococcus equi is associated with
an 85-90 kb virulence plasmid that provides the organism with the ability to be afacultative
intracellular pathogen. The inhalation of aerosolized virulent R. equi strains from the environment
and replication within alveolar macrophages are essential components of the pathogenesis. Due to
its importance and to the factthat only the equine experimental model can reproduce the classic
pulmonary lesions of this disease, this model has been used in the evaluation of prophylactic and
therapeutic methods. The macroscopic lesions are characterized by multifocal consolidation of
variable size with cavitations filled withpurulent material. The microscopic lesions consist of
piogranulomatous bronchopneumonia, with abundant amount of neutrophils, macrophages,
multinucleatedLanghans typegiant cells, and less lymphocytes and plasma cells.
The aims of this thesis were:
I: To determine if an inoculation dose of 5x103 CFU/mL of the 103+ strain of R. equi is able to
produce piogranulomatous broncopneumonia in foals younger than 6 months of age.
II: To detect early pulmonary infection by evaluating respiratory clinical signs, rectal temperature,
blood fibrinogen and ultrasonographic pulmonary findings.
III: Tocharacterize thegross and microscopic pulmonary lesions of foals experimentally infected
with 5x10³ CFU of the 103+ strain of R. equi, and to evaluate the immunologic response to this
challenge by histopathology and immunohystochemistry.
The thesis is divided into three chapters. Chapter I reviews the recent knowledge on relevant
aspects of rhodococcosis including etiology, clinical aspects, epidemiology, pathogenesis, immune
response, pathology, diagnosis, treatment and prognosis. Chapter II describes the experimental
reproduction and clinical aspects of bronchopneumonia caused by Rhodococcus equi strain 103+ in
foals. Chapter III describes the gross, microscopic and immunohistochemical evaluation of
pulmonary lesions experimentally induced by R. equi train 103+ in foals.
This study shows that an inoculum of 5x103 CFU of R. equi strain 103+ is able to produce
bronchopneumoniain foals less than 6 months of age. The evaluation of the respiratory clinical signs
in conjunction with rectal temperature, blood fibrinogen and observation of pulmonary

VII
piogranulomatous lesions by means of thoracic ultrasonography, allowed for diagnosis to be made
as early as 13.5 days after inoculation.
All of the foals challenged in this study had similar gross pulmonary lesions, characterized
by severe lung consolidation with multifocal to coalescent pyogranulomas in the cranioventral and
caudodorsal regions of the lung. The pulmonary microscopic lesions in all of the inoculated foals
consisted of pyogranulomatous bronchopneumonia; 54% of the foals had type II pneumocyte
hyperplasia (epithelization), 36% showed thrombosis and 27% exhibited fibroplasia.
Extrapulmonary lesions included hypopion (27%), hyphema (9%), centrolobular hepatic necrosis
(27%) and multifocal myocarditis (9%).
This study shows that the inoculation of a low dose of Rhodococcus equi strain 103+ can
produce pulmonary lesions similar to the lesions observed in the natural infection. The effect of a
dose lower than 5x10³ CFU should be investigated; we speculate that lower doses of this
microorganism could produce severe lesions with a longer incubation period, which migh bear more
resemblance to natural infection.

VIII
ABREVIATURAS
Ac: Anticuerpos BAL: Lavaje broncoalveolar CGB: Conteo de glóbulos blancos CMH II: Complejo mayor de histocompatibilidad tipo II DO: Densidad óptica ELISA: Ensayo por inmunoadsorción FC: Frecuencia cardíaca FNT-a: Factor de necrosis tumoral alfa FR: Frecuencia respiratoria H&E: Tinción de hematoxilina-eosina IFN-g: Interferón gamma IHC: Inmunohistoquímica IL-12: Interleuquina 12 Mab 10G5: Anticuerpo monoclonal de ratón PAI: Isla de patogenicidad PCR: Reacción en cadena de la polimerasa PHI: Plasma hiperinmune equino PTAH: Tinción de hematoxilina ácida fosfotúngstica SFE: Solución fisiológica estéril SDRA: Síndrome de distrés respiratorio agudo RGB: Recuento de glóbulos blancos Th1: Linfocitos colaboradores tipo 1 Th2: Linfocitos colaboradores tipo 2 TLR: Receptores tipo TollG Vap-A: Proteína A asociada a la virulencia VirR: Regulador de transcripción VirS: Sensor de membrana histidina quinasa
PALABRAS CLAVES
Rhodococcus equi, potrillos, reproducción experimental, bronconeumonía, clínica, patología, inmunohistoquímica.

2
ÍNDICE GENERAL
AGRADECIMIENTOS ................................................................................................. III
RESUMEN… .................................................................................................................. IV
SUMMARY… ................................................................................................................. VI
ABREVIATURAS… .................................................................................................... VIII
PALABRAS CLAVES ................................................................................................. VIII
CAPÍTULO I .................................................................................................................... 5
CAPÍTULO II ................................................................................................................. 23
CAPÍTULO III ............................................................................................................... 50
CAPÍTULO I: Infección causada por Rhodococcus equi en potrillos: revisión bibliográfica
1. Introducción .......................................................................................................... 5
2. Virulencia… ......................................................................................................... 6
3. Epidemiología y patogenia… .............................................................................. 7
4. Respuesta inmunológica… ................................................................................. 10
5. Diagnóstico clínico… .......................................................................................... 11
6. Patología macroscópica… .................................................................................. 14
7. Patología microscópica… ................................................................................... 15
8. Prevención ............................................................................................................ 16
9. Tratamiento… ..................................................................................................... 16
10. Pronóstico… ...................................................................................................... 18
11. Bibliografía… ..................................................................................................... 18

3
CAPÍTULO II: Reproducción experimental y aspectos clínicos de bronconeumonías causadas por la cepa 103+ de Rhodococcus equi en potrillos
Introducción ...................................................................................................................... 23
Hipótesis y objetivos ........................................................................................................ 24
1. Materiales y métodos ........................................................................................... 25
1.1 Animales ......................................................................................................... 25
1.2 Inóculo ............................................................................................................ 25
1.3 Inoculación ..................................................................................................... 27
1.4 Criterio de enfermedad .................................................................................. 28
1.5 Estudio bacteriológico… ............................................................................... 28
1.6 Estudio clínico ................................................................................................. 28
1.7 Exámenes complementarios ......................................................................... 29
1.7.1 Medición de fibrinógeno sanguíneo ................................................. 29
1.7.2 Ultrasonografía torácica ................................................................... 30
1.7.3 Radiografía torácica ........................................................................... 32
2. Resultados ............................................................................................................. 32
2.1 Reproducción experimental .......................................................................... 32
2.2 Estudio bacteriológico… ................................................................................ 33
2.3 Estudio clínico .................................................................................................. 35
2.4 Medición de fibrinógeno sanguíneo. ............................................................. 37
2.5 Ultrasonografía torácica ................................................................................ 37
2.6 Radiografía torácica ........................................................................................ 42
3. Discusión y conclusión .......................................................................................... 45
4. Bibliografía ............................................................................................................ 48

4
CAPÍTULO III: Estudio anatomopatológico e inmunohistoquímico de lesiones pulmonares inducidas
experimentalmente por la cepa 103+ de Rhodococcus equi en potrillos
Introducción ....................................................................................................................... 50
Hipótesis y objetivo ........................................................................................................... 52
1. Materiales y métodos ............................................................................................ 52
1.1 Animales .......................................................................................................... 52
1.2 Inoculación experimental ............................................................................... 52
1.3 Estudio anatomopatológico ............................................................................ 53
1.4 Tinciones especiales ........................................................................................ 53
1.5 Evaluación inmunohistoquímica .................................................................... 53
1.6 Evaluación de la respuesta inmune ................................................................ 54
2. Resultados ............................................................................................................. 54
2.1 Patología macroscópica ................................................................................. 54
2.2 Patología microscópica y tinciones especiales .............................................. 60
2.3 Evaluación inmunohistoquímica y de la respuesta inmune ....................... 72
3. Discusión y conclusión ......................................................................................... 73
4. Bibliografía ........................................................................................................... 77

5
CAPÍTULO I
Infección causada por Rhodococcus equi en potrillos: revisión bibliográfica
1. Introducción
Rhodococcus equi (R. equi) se caracteriza por producir principalmente enfermedad
respiratoria en equinos jóvenes (pertenece al Philum XXVI Actinobacteria, clase I: Actinobacteria,
al orden V: Corynebacteriales, familia IV: Nocardiaceae, género IV: Rhodococcus: R. equi). Este
género comprende más de 20 especies, incluyendo patógenos de animales (R. equi), plantas (R.
fascians) y humanos (R. equi, R. rhodochrous y R. erythropolis) (Conville y Witebsky, 2007;
Prescott, 1991; VonBargen y Hass, 2009). R. equi es un cocobacilo gram positivo, inmóvil, no
flagelado e intracelular facultativo (Harrington et al., 2005; VonBargen y Hass, 2009). Su forma
varía de bacilo a coco de acuerdo a las condiciones de crecimiento: luego de 4 horas en medio
líquido presenta forma de bacilo, pero tras un día de cultivo en este medio o en agar sangre desarrolla
la forma de coco (VonBargen y Hass, 2009). Sus colonias son de 1 mm de diámetro, irregulares,
suaves, de aspecto mucoide y toman color rojizo claro luego de 1 semana de crecimiento (Conville y
Witebsky, 2007; VonBargen y Hass, 2009). Las características bioquímicas más importantes
incluyen: positividad a las pruebas de catalasa, ureasa, lipasa, nitrato y al fenómeno CAMP, y
negatividad a las pruebas de citocromo oxidasa, fermentación de hidratos de carbono, hidrólisis de
gelatina, indol, hipurato y esculina (Songer y Post, 2005; VonBargen y Hass, 2009).
La bronconeumonía producida por R. equi es considerada una de las principales
enfermedades respiratorias de los potrillos debido a la alta morbilidad y letalidad con la que se
presenta. Muchas veces producen epizootias (Cortez et al., 2008; Giguère et al., 2003; Noah et al.,
2002), y la mayoría de las bronconeumonías ocasionadas por R. equi afecta a potrillos de entre 2 y
4 meses de edad (Giguère et al., 2003; Pusterla et al., 2007). La morbilidad en establecimientos en
donde la enfermedad se presenta de forma enzoótica puede superar el 40% (Giguère et al., 2003).
El primer brote de bronconeumonía por R. equi en Argentina fue reportado en potrillos sangre pura
de carrera entre los meses de septiembre y diciembre de 1979, en un grupo de 60 animales. En un
período de 3 meses se observó una morbilidad del 65% y una letalidad del 55% (Campero et al.,
1981). R. equi produce un impacto económico-productivo negativo en la producción equina
nacional y mundial debido a la pérdida directa de animales, baja ganancia de peso y gastos de
tratamiento (Cohen et al., 2005; Giguère et al., 2003; Muscatello et al., 2009).
R. equi es también un patógeno oportunista emergente de personas inmunodeprimidas como
en el caso de pacientes infectados por el virus de la inmunodeficiencia adquirida, personas bajo

6
tratamiento prolongado con corticoides y pacientes con diabetes y leucemia, entre otros (Harrington
et al., 2005; VonBargen y Hass, 2009). En humanos, R. equi puede causar bronconeumonía severa
con desarrollo de fiebre, tos y dolor de pecho, y abscesos cerebrales, hepáticos, renales o de otros
órganos, con resultados pobres al tratamiento (Pei et al., 2007).
R. equi también es frecuentemente aislado en cerdos con linfadenitis de los linfonódulos
submaxilares y tonsilitis (Prescott, 1991; VonBargen y Hass, 2009). Esta bacteria causa lesiones
granulomatosas similares a la tuberculosis en los linfonódulos de bovinos, habiendo sido
observadas en el 0,008% a la inspección post mortem en mataderos (Flynn et al., 2001). En cabras,
R. equi produce abscesos hepáticos y pulmonares, y bronconeumonía (Tkachuk-Saad, 1998). Los
ovinos, camélidos, felinos y caninos son ocasionalmente afectados por R. equi (Muscatello et al.,
2009; VonBargen y Hass, 2009).
A pesar de que existe una extensa cantidad de literatura sobre R. equi, mucha de esta
información se encuentra dispersa. La siguiente revisión bibliográfica se enfocará en la información
reciente sobre los factores de virulencia, epidemiología, patogenia, respuesta inmune, patología,
diagnóstico, tratamiento y pronóstico de las infecciones por R. equi en equinos.
2. Virulencia
R. equi produce enzimas solubles (fosfolipasa y colesterol oxidasa) denominadas “factores
equi”. Estos cooperan sinérgicamente con la fosfolipasa D de Corynebacterium pseudotuberculosis,
la toxina b de Staphylococcus aureus y la hemolisina de Listeria monocytogenes para producir
hemólisis completa de glóbulos rojos ovinos; esta característica es también conocida como
fenómeno CAMP, el cual confirma la identidad de R. equi (Campero et al., 1981; VonBargen y
Hass, 2009). R. equi también posee cadenas largas de ácido micólico en la pared celular,
característica que comparte con miembros del género Mycobacterium y que actúan interfiriendo
con la habilidad de los leucocitos para fagocitar la bacteria (VonBargen y Hass, 2009).
Sin embargo, la virulencia de R. equi está asociada a la presencia de un plásmido de entre
85 kb y 90 kb, que es necesario (pero no suficiente) para causar enfermedad en los potrillos
(Harrington et al., 2005; Kuskie et al., 2007; Muscatello, 2012). Este plásmido posee una isla de
patogenicidad (denominada por su abreviatura en inglés: PAI) que porta los genes que codifican las
proteínas asociadas a la virulencia. Estas proteínas se conocen como Vap-A, Vap-C, Vap-D, Vap-
E, Vap-G y Vap-H, siendo la proteína de superficie Vap-A la más importante (MacArthur et al.,
2010; Muscatello, 2012). La expresión de la proteína Vap-A es termorregulada, ocurriendo de forma
óptima entre los 34 ºC y 41 ºC. Además, su producción se incrementa en respuesta a la

7
privación de hierro (Harrington et al., 2005; Jain et al., 2003). Aunque todas las proteínas Vap son
inmunogénicas, la proteína Vap-A y la proteína secretoria Vap-C son las que inducen una respuesta
inmune más importante, tanto en animales adultos como en potrillos (Dawson et al., 2010; Jaks et
al., 2007).
La respuesta de anticuerpos séricos contra la proteína Vap-A ocurre naturalmente en
potrillos y se correlaciona con la carga de R. equi en la materia fecal durante los primeros 3 meses
de vida; estos anticuerpos también se encuentran elevados en los potrillos con bronconeumonía.
Así, la determinación de anticuerpos en suero contra la proteína Vap-A se considera insuficiente
para el diagnóstico de esta infección. Ello es así debido a la dificultad de discernir entre potrillos
afectados de bronconeumonía y potrillos sanos con proliferación intestinal a causa de la exposición
natural a R. equi (Dawson et al., 2010).
Dentro de la PAI existen dos reguladores transcripcionales: VirR/VirS y los receptores
nucleares orphan (orf). El sistema regulador global VirR/VirS tiene dos componentes clásicos: un
regulador de transcripción (VirR) y un sensor de membrana histidina quinasa (VirS). Estos fueron
descubiertos hace 24 años cuando se demostró que regulaban la producción de toxinas codificadas
por plásmidos (toxina alfa y perfringolisina) y algunas otras enzimas extracelulares de Clostridium
perfringens tipo A (Lyristis et al., 1994; Shimizu et al., 1994). Los receptores nucleares orphan
pertenecen a una superfamilia que regula diversos procesos biológicos. Estas moléculas son factores
de transcripción activados por ligandos que actúan como sensores naturales de un amplio rango de
hormonas esteroides y xenobióticos. Los receptores nucleares orphan son definidos por la falta de
ligandos identificables que controlan sus funciones in vivo (Mukherjee y Mani, 2010). Las
mutaciones en cualquiera de estos dos reguladores transcripcionales, VirR/VirS u orf, producen la
atenuación completa de la virulencia de R. equi (Ren y Prescott, 2004). Las cepas carentes del
plásmido en las que se encuentra la PAI no poseen la habilidad de producir la proteína Vap-A. Esta
última es indispensable para la supervivencia y el crecimiento de R. equi dentro de los macrófagos,
debido a que detiene la maduración fagosomal (Giguère, 2001; MacArthur et al., 2010). Dichas
cepas son, por lo tanto, avirulentas.
3. Epidemiología y patogenia
El suelo es el principal reservorio de R. equi (Giguère et al., 2003; Harrington et al., 2005).
Esta bacteria tiene requerimientos mínimos de crecimiento, siendo los ácidos orgánicos simples
encontrados en la materia fecal de los herbívoros, una importante fuente de nutrientes para su
desarrollo (Dawson et al., 2010; Slovis et al., 2005). A pesar de la amplia dispersión de R. equi en
el ambiente, es característico hallar haras endémicos, haras con infección esporádica y haras sin la

8
presencia de enfermedad producida por este microorganismo (Giguère et al., 2003; Muscatello,
2012). Esto refleja el efecto de diferentes factores que influyen en el desarrollo de la infección,
incluyendo condiciones ambientales, de manejo y virulencia de cepas (Slovis et al., 2005). La alta
temperatura es el factor ambiental más importante que predispone al desarrollo de bronconeumonías
en potrillos (Dawson et al., 2010).
La eliminación fecal proveniente de yeguas y potrillos es una fuente importante de
contaminación progresiva del ambiente (Giguère, 2001; Muscatello, 2012). Las yeguas madres de
potrillos afectados no eliminan más R. equi en las heces que las yeguas madres de potrillos sanos,
indicando que la alta eliminación producida por ciertas yeguas no está necesariamente vinculada a
la infección de sus potrillos (Wilkins, 2009). La carga intestinal de R. Equi en herbívoros adultos
es principalmente pasiva debido a que esta bacteria no logra replicarse en las condiciones
anaeróbicas del intestino grueso de animales adultos. Sin embargo, esta bacteria puede multiplicarse
en el intestino de potrillos de hasta 3 meses de edad, pudiendo alcanzar concentraciones de hasta
105 UFC de R. equi por gramo de materia fecal (Wilkins, 2009). Por tal razón, los potrillos
infectados son considerados la mayor fuente de contaminación en los establecimientos de cría
equina (Dawson et al., 2010). También se ha demostrado la presencia de cepas virulentas de R. equi
en el aire exhalado por potrillos con bronconeumonía, lo que demuestra la importancia de los
aerosoles provenientes de animales enfermos como fuente de contagio, sugiriendo la posibilidad de
una vía alternativa de transmisión (Muscatello et al., 2009; Muscatello, 2012; Takai y Cohen, 2011).
Los potrillos adquieren la infección respiratoria a través de la inhalación de aerosoles, polvo
o materia fecal conteniendo R. equi (Muscatello, 2012; Wilkins, 2009; Zachary, 2012). La ingestión
de R. equi también es una importante vía de exposición y posiblemente de inmunización, pero
raramente se produce la diseminación hematógena de la bacteria a través de esta vía, aunque esto
puede ocurrir si existen múltiples exposiciones con grandes cantidades de R. equi (Wilkins, 2009).
El período de incubación varía de 6 a 18 días en potrillos inoculados experimentalmente con
1x104 UFC de cepas virulentas de R. equi, mientras que en condiciones de campo la incubación es
variable dependiendo de la puerta de entrada y la cantidad de bacterias virulentas. Basados en
estudios realizados en Argentina y Japón, el período de incubación teórico promedio, calculado
según el modelo epidemiológico Starwell, fue de 49 días (Horowitz et al., 2001; Muscatello, 2012;
Wilkins, 2009).
Cuando R. equi es inhalado, se deposita en la mucosa de las vías respiratorias (tráquea y
bronquios) debido a la fuerza centrífuga y la turbulencia del aire inspirado. Si este microorganismo
logra alcanzar la unión bronquiolo-alveolar, el mismo ya no puede ser eliminado por el barrido
mucociliar y es detectado por los macrófagos alveolares (célula presentadora de antígenos), los

9
cuales fagocitan la bacteria a través de la interacción antígeno (Ag)/receptor. Los macrófagos
presentan fragmentos de péptidos antigénicos (incluyendo antígenos de R. equi) asociados al
complejo mayor de histocompatibilidad tipo II (CMH II) a los linfocitos T colaboradores
sensibilizados. Sin embargo, para que R. equi se adhiera inicialmente a los macrófagos, esta bacteria
debe ser opsonizada con Ac o con fragmentos del complemento (Ishino et al., 1992; Zachary, 2012).
La habilidad fagocítica de los macrófagos y neutrófilos en los potrillos es comparable a la de los
adultos, pero la falta de capacidad de opsonización del plasma es un factor limitante hasta las 3 o 4
semanas de edad (Dawson et al., 2010).
Las células dendríticas captan la bacteria mediante endocitosis, a través de seudópodos que
penetran entre los estrechos espacios del epitelio respiratorio y presentan fragmentos de péptidos
antigénicos (incluyendo antígenos de R. equi) asociados al CMH II a los linfocitos T colaboradores
vírgenes (Zachary, 2012). Los potrillos menores de 3 meses podrían tener un aumento de la
susceptibilidad a la infección por R. equi, debido a una disminución de la función de las células
dendríticas, por una expresión reducida (edad dependiente) del CMH II, lo que produce una menor
presentación de antígenos a los linfocitos T colaboradores vírgenes (Flaminio et al., 2007; Pargass
et al., 2009; Dwason et al., 2010). A su vez, los potrillos parecen producir un rango limitado de
isotipos de Ac hasta aproximadamente los 2 meses de edad (Caston et al., 2006). Los principales
isotipos de inmunoglobulinas (Ig) equinas son la IgG, IgA, IgM, IgE e IgD. La IgG representa el
80% del total de las inmunoglobulinas del plasma de los potrillos (Dawson et al., 2010). Las IgGa
e IgGb son las subclases más abundantes de IgG en el suero y calostro del equino adulto. La síntesis
del isotipo IgGa ha sido detectada en las primeras 5 a 8 semanas de vida. La opsonización de los
componentes capsulares de R. equi con IgG, y fragmentos del complemento, parece promover la
fagocitosis de R. equi y aumentar la destrucción bacteriana a través de la fusión fagosoma-lisosoma
(Cauchard et al., 2004).
R. equi expresa moléculas de superficie como la proteína Vap-A, que se une a receptores de
membrana expresados en los macrófagos alveolares como los Toll Like Receptor 2 (TLR),
receptores de complemento, receptor de manosa y Mac-1 (integrinas β2) de los neutrófilos (Dawson
et al., 2010; Zachary, 2012). Además, las moléculas CD-1 de los macrófagos alveolares actúan
como receptores no clásicos del complejo mayor de histocompatibilidad capaz de presentar
componentes antigénicos lipídicos inusuales, como el ácido micólico y lipoarabinomanan presentes
en la pared celular de R. equi. Los macrófagos alveolares de los potrillos expresan niveles mínimos
de moléculas CD-1, pudiendo ser este otro mecanismo por el cual R. equi evade la respuesta inmune
y predispone a los potrillos jóvenes a la infección por R. equi (Dawson et al., 2010). Se ha
demostrado que, a diferencia de lo que sucede en el equino, la mayor parte de las cepas de R. equi

10
que afectan al ser humano no portan el plásmido de virulencia, desconociéndose el rol de la proteína
Vap-A en la infección en el ser humano (Nicholson y Prescott, 1997).
Los receptores mencionados anteriormente producen una señal que lleva a la activación de
varios factores de transcripción, como el factor nuclear kappa β y la proteína quinasa mitogénica
activada que regulan la producción de citoquinas y otras moléculas reguladoras que aumentan la
respuesta de la inmunidad innata (Dawson et al., 2010; Zachary, 2012).
Usualmente, durante la fagocitosis de R. equi por parte de los macrófagos, este
microorganismo es envuelto por una porción de la membrana plasmática y es internalizado,
formando una nueva organela llamada fagosoma que posteriormente madura por la interacción con
vesículas endocíticas tempranas, tardías y lisosomales, culminando con la formación del
fagolisosoma. Normalmente, en el fagolisosoma la bacteria es expuesta a varios factores
bactericidas, tales como el pH ácido (4,0-5,0), enzimas hidrolíticas (proteasas, lipasas, ADNasas,
ARNasas, entre otras), y metabolitos reactivos del oxígeno (radicales superóxidos, peróxido de
hidrógeno y óxido nítrico) que contribuyen a la destrucción y eliminación de la bacteria (Wilkins,
2009; VonBargen y Hass, 2009). Sin embargo, en los macrófagos que han fagocitado cepas
virulentas de R. equi no se produce la maduración del fagosoma; por lo tanto, ocurre la replicación
intracelular de R. equi (VonBargen y Hass, 2009). Debido a la rápida replicación de R. equi dentro
del fagosoma y a la producción de los factores equi, se produce la muerte prematura de los
macrófagos alveolares con la consecuente liberación de grandes cantidades de bacterias al
parénquima adyacente, donde son fagocitadas por nuevos macrófagos provenientes del sistema
circulatorio y linfonódulos regionales, perpetuándose de este modo el proceso (Zachary, 2012).
Los neutrófilos forman parte de la respuesta inflamatoria aguda y son capaces de eliminar
R. equi a través de una exitosa formación del fagolisosoma y el estallido respiratorio. Sin embargo,
este mecanismo no es efectivo para controlar la enfermedad; en su lugar, produce una extensa
destrucción pulmonar por la liberación de enzimas lisosomales y metabolitos reactivos del oxígeno
(Demmers et al., 2001; Zachary, 2012).
4. Respuesta inmunológica
Frecuentemente, los potrillos están expuestos a R. equi inmediatamente luego del
nacimiento. Sin embargo, mientras la mayoría de los potrillos desarrollan una respuesta inmune
protectora, algunos animales exhiben una susceptibilidad única a infecciones a causa de la
inmadurez de su sistema inmune. Debido a la placentación epiteliocorial del equino, la transferencia
de anticuerpos maternos al potrillo se realiza exclusivamente a través de la ingestión de calostro.
Esta transferencia pasiva juega un rol crítico en la resistencia contra una variedad de agentes

11
infecciosos durante los primeros meses de vida (Boyd et al., 2003). En potrillos, la mayor parte de
los anticuerpos desaparece a los 3 meses de vida, lo que coincide con la edad típica en la que son
diagnosticadas las bronconeumonías por R. equi. Este hallazgo es considerado evidencia de la
función protectora de los anticuerpos contra la infección producida por este microorganismo
(Dawson et al., 2010). Aunque la IgA es la primera línea de defensa en el tracto respiratorio, la
misma está ausente en la mucosa nasal durante los primeros 28 días de vida en los potrillos, lo que
puede incrementar la susceptibilidad de los mismos a la infección en la etapa temprana de vida
(Dawson et al., 2010). No obstante, se requiere tanto de una respuesta mucosa local (IgA) como de
una respuesta humoral sistémica (IgGa e IgGb) para proteger contra la infección pulmonar e
intestinal de R. equi (Hooper-McGrevy et al., 2005).
Es aceptado que la respuesta de los linfocitos T colaboradores tipo 1 (Th1) es necesaria para
eliminar la infección contra R. equi y que la respuesta de los linfocitos T colaboradores tipo 2 (Th2)
es perjudicial. La diferenciación hacia una respuesta Th1 o Th2 está determinada por la
concentración de diferentes citoquinas en el lugar de la infección. Por ejemplo, la interleuquina 12
(IL-12), producida por células dendríticas y macrófagos, estimula a los linfocitos indiferenciados a
diferenciarse en células Th1. Una vez que un subgrupo celular es diferenciado, la respuesta inmune
tiende a persistir en esa dirección debido a que las citoquinas producidas por un subgrupo celular
inhiben el desarrollo del otro subgrupo celular (en este caso la respuesta Th2). Este fenómeno es
conocido como regulación cruzada (Snyder, 2007).
El factor de necrosis tumoral alfa (FNT-α) y el interferón gamma (IFN-γ), producidos por
los linfocitos T colaboradores y citotóxicos, estimulan la capacidad fagocítica de los macrófagos
(Dawson et al., 2010). La expresión de estos factores se ha observado incrementada en desafíos con
cepas virulentas de R. equi en potrillos y equinos adultos (Dawson et al., 2010). Sin embargo, los
potrillos hasta los 3 meses de edad parecen tener una expresión reducida de IFN-γ cuando se la
compara con la de los equinos adultos; la razón de esta disminución permanece desconocida
(Dawson et al., 2010; Ryan et al., 2010).
5. Diagnóstico clínico
Actualmente no existe un método de diagnóstico único, con la suficiente especificidad y
sensibilidad, para el diagnóstico de bronconeumonías causadas por R. equi en animales vivos. Por
eso el diagnóstico está basado en la detección de R. equi en muestras de exudado broncoalveolar en
conjunto con los hallazgos clínicos, anormalidades hematológicas y la evidencia radiográfica y/o
ultrasonográfica de lesiones pulmonares (Muscatello, 2012).

12
Es importante la detección temprana de los potrillos infectados para que la aplicación de
medidas terapéuticas y preventivas resulten exitosas (Cortez et al., 2008; Giguère et al., 2003;
Harrington et al., 2005; Muscatello, 2012). A pesar de ello, el reconocimiento clínico precoz de las
bronconeumonías severas causadas por R. equi es difícil debido a la lenta diseminación respiratoria
de este microorganismo, a la extraordinaria habilidad de los potrillos para compensar la pérdida
progresiva funcional del volumen pulmonar y a que los signos clínicos y la auscultación
frecuentemente no se correlacionan con el grado de consolidación pulmonar (Harrington et al.,
2005; Slovis, 2007).
Los signos clínicos iniciales más importantes son la tos esporádica o intermitente, anorexia,
hipertermia, depresión, disnea y descarga nasal. A pesar de ser inespecíficos, estos ayudan a
establecer un diagnóstico presuntivo de neumonía por R. equi (Giguère, 2001; Muscatello, 2012).
Entre los signos clínicos observados en los potrillos con estadios más avanzados de enfermedad se
encuentran: ollares dilatados, esfuerzo abdominal o franca respiración abdominal, intolerancia al
ejercicio, hipopión, hipema, expresión ansiosa, decúbito esternal, cianosis, anemia,
trombocitopenia, diarrea y claudicación severa (Caston et al., 2006). Aun en la forma crónica de la
enfermedad, los signos clínicos respiratorios tienen un comienzo agudo, ya que la misma progresa
de forma insidiosa hasta que existe suficiente cantidad de parénquima pulmonar dañado como para
causar signos clínicos de falla respiratoria (Wilson, 1992).
El diagnóstico de la bronconeumonía por R. equi debe ser confirmado por medio del cultivo
bacteriológico o la amplificación del gen Vap-A mediante la reacción en cadena de la polimerasa
(PCR) en muestras de exudado broncoalveolar obtenido de potrillos con signos clínicos de
enfermedad pulmonar, evidencia citológica de infección de las vías aéreas o evidencia radiográfica
o ultrasonográfica de bronconeumonía (Harrington et al., 2005; Giguère et al., 2011; Muscatello,
2012; Pusterla et al., 2007).
Sin embargo, es necesario tener en cuenta que el cultivo bacteriológico no distingue entre
cepas virulentas y avirulentas y puede, además, producir resultados falsos negativos debido a la
previa administración de antibióticos (Muscatello, 2012). Así, en potrillos el cultivo bacteriológico
debe ser realizado en conjunto con la técnica de PCR del exudado broncoalveolar o de la cepa
aislada (Giguère et al., 2011; Pusterla et al., 2007). Se ha observado que potrillos sin signos
respiratorios provenientes de haras enzoóticos pueden ser positivos a la PCR del aspirado
broncoalveolar, haciendo que este análisis solo no pueda ser utilizado para confirmar el diagnóstico.
Por lo tanto, el cultivo bacteriológico y la PCR de aspirados broncoalveolares siempre deben ser
interpretados en el contexto de los hallazgos clínicos y nunca deben ser utilizados individualmente

13
como prueba inicial para detectar una infección por R. equi en animales clínicamente sanos (Giguère
et al., 2003).
Además de los métodos mencionados anteriormente, entre las técnicas de laboratorio a
utilizar para la detección temprana de la enfermedad se encuentra el conteo de glóbulos blancos
(CGB) y la medición de fibrinógeno, aunque el aumento de fibrinógeno siempre parece anteceder
al incremento de los glóbulos blancos (Webb, 2011). A pesar de que estos hallazgos no son
específicos, un conteo de glóbulos blancos por encima de 20 000 células/µL junto con una
concentración de fibrinógeno de más de 700 mg/dL y evidencia de bronconeumonía
piogranulomatosa es un hallazgo presuntivo de una neumonía causada por R. equi (Giguère et al.,
2003; Giguère et al., 2011). La neutrofilia con desviación a la izquierda es generalmente moderada
en el equino; por esta razón se utiliza el aumento de fibrinógeno como otro indicador de inflamación
(Webb, 2011).
También es necesario tener en cuenta que la concentración de fibrinógeno por sí solo no es
útil como método de diagnóstico temprano, dado que puede verse aumentado por infecciones
causadas por otros patógenos diferentes de R. equi (Giguère et al., 2011). Estudios recientes han
utilizado la medición de otras proteínas de fase aguda como el amiloide sérico A, aunque se observó
que su medición realizada semanalmente no representa un marcador precoz de confianza en el
diagnóstico clínico. A pesar de ello, su medición -una vez que los signos clínicos respiratorios están
presentes-, permite al veterinario clínico la evaluación del progreso de la infección y la eficacia del
tratamiento (Passamonti et al., 2015).
La ultrasonografía de tórax es un método de diagnóstico complementario, eficiente y preciso
para la detección precoz de bronconeumonías por R. equi y permite la evaluación de la severidad
de las lesiones neumónicas y su evolución en respuesta a la terapia antibiótica (Cortez et al., 2008;
Giguère, 2001). Las lesiones pulmonares aparecen como áreas encapsuladas hiperecoicas bien
definidas, de tamaño variable, con un centro hipoecoico (Muscatello, 2012; Ramirez et al., 2004).
Esta técnica se realiza en establecimientos endémicos en los potrillos entre las 3 y 6 semanas de
edad, lo que coincide con la revisación ultrasonográfica de las yeguas con fines reproductivos.
Las limitaciones de esta técnica radican en que no existen lesiones patognomónicas debido
a que otros agentes etiológicos que no son comunes en equinos, como Aspergillus fumigatus,
Histoplasma capsulatum y Coccidioides immitis, pueden producir lesiones similares. Otra
limitación es que solo detecta lesiones que se encuentran en la periferia pulmonar (Muscatello,
2012; Ramirez et al., 2004).

14
A pesar de que la radiografía torácica es útil para detectar inflamación intersticial,
linfoadenopatía hiliar y ocasionalmente cavitaciones, no se recomienda como único criterio de
pronóstico y de eutanasia, debido a que muchos animales infectados con R. equi, pero sin signos
clínicos, muestran lesiones radiográficas (Caston et al., 2006; Giguère, 2001). De acuerdo con el
conocimiento actual, las pruebas serológicas, como la inmunodifusión en gel de agar y la técnica
de ELISA, no han demostrado tener la sensibilidad y/o especificidad adecuadas para su utilización
como métodos de detección temprana de la enfermedad (Giguère et al., 2011). La presencia de
anticuerpos indica exposición, infección subclínica o transferencia de anticuerpos maternales, pero
no necesariamente indica la presencia de enfermedad clínica. Por esta razón, se excluye la
utilización de pruebas diagnósticas serológicas para detectar bronconeumonías causadas por R. equi
(Dawson et al., 2010; Giguère et al., 2011).
6. Patología macroscópica
La lesión macroscópica pulmonar característica de la infección por R. equi es una
bronconeumonía piogranulomatosa, caracterizada por áreas multifocales a coalescentes
consolidadas de color blanco amarillento, firmes, de 1 a 10 cm de diámetro (Zachary, 2012). Con
el tiempo, los centros de los piogranulomas se vuelven friables a causa del desarrollo de necrosis
piogranulomatosa licuefactiva (Caswell y Williams, 2016). Estas lesiones suelen ser erróneamente
interpretadas como abscesos, pero no poseen una cápsula fibrosa (Caswell y Williams, 2016).
Aunque generalmente la lesión pulmonar se observa en las áreas cráneoventrales, la totalidad del
parénquima pulmonar puede verse afectado (Brown et al., 2007). Los linfonódulos
traqueobronquiales se encuentran agrandados, de consistencia firme, y al corte presentan áreas
coalescentes de exudado de color blanco amarillento, caracterizados histológicamente por
linfadenitis piogranulomatosa necrotizante con edema difuso (Brown et al., 2007; Zachary, 2012).
El desarrollo de pleuritis no es común (Caswell y Williams, 2016; Valdes y Johnson, 2005).
Aproximadamente la mitad de los potrillos con bronconeumonía por R. equi desarrolla
enterocolitis. El intestino de los potrillos con bronconeumonía está expuesto a una continua
exposición bacteriana proveniente de la deglución de exudados respiratorios con gran cantidad de
R. equi. En general, el barrido mucociliar no es afectado directamente por R. equi; por lo tanto esta
bacteria es movilizada hacia la nasofaringe, desde donde, mediante la deglución, accede al tracto
digestivo (Zachary, 2012). En el intestino, las lesiones comienzan en las placas de Peyer, debido a
que R. equi utiliza a las células M para acceder al tejido linfoide intestinal; las mismas son afectadas
por la inflamación granulomatosa con necrosis, lo que da lugar a la aparición de úlceras irregulares
de entre 1 y 2 cm de diámetro, frecuentemente cubiertas por restos necróticos o purulentos (Brown

15
et al., 2007; Zachary, 2012). Posteriormente, la infección se traslada a los linfonódulos
mesentéricos, los cuales se observan aumentados de tamaño, de consistencia firme y color gris,
debido al edema y a la inflamación piogranulomatosa (Brown et al., 2007; Zachary, 2012).
Aproximadamente un tercio de los potrillos con bronconeumonía por R. equi desarrolla
polisinovitis supurativa, producida por la diseminación hematógena de la bacteria (Caswell y
Williams, 2016). Una cantidad menor de potrillos desarrolla poliartritis inmunomediada,
particularmente de las articulaciones tibiotarsianas y carpianas, caracterizada por sinovitis
linfoplasmacítica y depósitos de inmunoglobulinas en la membrana sinovial de la que no se aíslan
microorganismos (Caswell y Williams, 2007; Muscatello, 2012). Las infecciones diseminadas
pueden también ocasionar uveítis, panoftalmitis, empiema de las bolsas guturales, sinusitis,
abscedación cutánea, esplénica o hepática y osteomielitis de la pelvis, metáfisis de huesos largos y
cuerpos de las vértebras (Caswell y Williams, 2016; Clark-Price et al., 2003; Giguère, 2001).
7. Patología microscópica
La lesión pulmonar microscópica típica es una bronconeumonía piogranulomatosa
multifocal caracterizada inicialmente por la infiltración de las vías respiratorias y alvéolos con
macrófagos, neutrófilos y escasa cantidad de linfocitos y células plasmáticas.
Posteriormente, en las áreas de inflamación se desarrolla necrosis piogranulomatosa
licuefactiva con abundante cantidad de restos celulares, células inflamatorias degeneradas, fibrina
y edema, rodeada por una banda de neutrófilos y células gigantes multinucleadas tipo Langhans,
las que a la tinción de Gram, demuestran la presencia de cocobacilos gram positivos
intracitoplasmáticos (Caswell y Williams, 2016).
En el intestino, R. equi puede producir enterocolitis caracterizada por inflamación
piogranulomatosa multifocal de la lámina propia y tejido linfoide asociado a las mucosas
intestinales con gran cantidad de macrófagos, células gigantes multinucleadas, neutrófilos y menor
cantidad de linfocitos y células plasmáticas. En la mucosa, el infiltrado inflamatorio produce un
engrosamiento de las vellosidades y distorsiona las criptas intestinales (Zachary, 2012). Las áreas
de inflamación piogranulomatosa evolucionan posteriormente con la formación de úlceras, las
cuales se cubren con membranas fibrinonecróticas, neutrófilos y macrófagos con bacterias
intracitoplasmáticas (Caswell y Williams, 2016).
Desde la pared intestinal los microorganismos drenan hacia los linfonódulos intestinales
regionales produciendo linfadenitis piogranulomatosa con grandes cantidades de neutrófilos,
macrófagos y células gigantes multinucleadas tipo Langhans (Brown et al., 2007).

16
En cortes fijados en formol e incluidos en parafina, la presencia de R. equi puede ser puesta
en evidencia a través de la técnica de inmunohistoquímica (Retteg et al., 2009). Para la
inmunomarcación se utiliza el anticuerpo monoclonal de ratón (MAb 10G5) contra la proteína Vap-
A. Generalmente, la bacteria se detecta dentro de los macrófagos presentes en las lesiones
(Madarame et al., 1996).
8. Prevención
Actualmente no existe una vacuna efectiva que proteja a los potrillos contra la neumonía
producida por R. equi (Giguère et al., 2003; Giguère et al., 2011). Por esta razón la profilaxis se
basa en la inmunización pasiva mediante el uso de plasma hiperinmune equino (PHI), el cual reduce
significativamente las tasas de morbilidad y mortalidad (Caston et al., 2006; Muscatello 2012;
Slovis, 2007). El componente protector del PHI es desconocido (Muscatello, 2012). Se recomienda
la administración de 1 litro de PHI dentro de los primeros 2 días de vida a todos los potrillos, con
una segunda dosis de similar volumen entre la segunda y cuarta semana de edad (Caston et al.,
2006; Slovis, 2007). También se puede utilizar el manejo ambiental que tiene como objetivo
disminuir la carga de polvo en el ambiente mediante el riego abundante de los corrales, el uso de
potreros cubiertos de pasto y la desinfección de los establos (Muscatello, 2012).
La concentración de cepas virulentas de R. equi en el suelo no se correlaciona positivamente
con un incremento en la frecuencia de bronconeumonía por R. equi; por lo tanto, la remoción del
estiércol no es una práctica útil para el control de las bronconeumonías por R. equi (Dawson
et al., 2010).
9. Tratamiento
En los últimos años la aparición de macrólidos de nueva generación, como la claritromicina
y la azitromicina en combinación con rifampicina, reemplazaron a la eritromicina, la cual produce
la hipertermia inducida por anhidrosis debido a una falla en el mecanismo de sudoración (Stieler et
al., 2015). Estas combinaciones son más estables químicamente, tienen una gran biodisponibilidad
vía oral, adquieren altas concentraciones en tejidos y células fagocíticas y siguen permaneciendo
actualmente como el tratamiento de elección (Giguère et al., 2004; Reuss y Cohen, 2015). Sin
embargo, esta combinación comúnmente causa diarrea, generalmente pasajera, aunque algunos
potrillos pueden desarrollar diarrea severa con deshidratación (Giguère et al., 2011). La
administración de azitromicina se recomienda a razón de 10 mg/Kg vía oral cada 24 horas, más la
administración de rifampicina oral a razón de 5 mg/Kg cada 12 horas (Cohen, 2014). Se ha
demostrado que la combinación con rifampicina previene las mutaciones bacterianas que

17
intervienen en la resistencia antimicrobiana contra los macrólidos (Cohen, 2014). Aunque la
mayoría de los aislamientos de R. equi son altamente susceptibles a los macrólidos y la rifampicina,
se han encontrado cepas resistentes a cada una de estas drogas. La rifampicina no debe ser utilizada
sola, dado que incrementa la posibilidad de que se desarrollen cepas resistentes (Boyen et al., 2011;
Giguère et al., 2011; Muscatello, 2012). Tampoco se recomiendan los tratamientos
quimioprofilácticos porque pueden promover resistencia de cepas de R. equi o de otras bacterias a
los macrólidos (Giguère et al., 2011; Muscatello, 2012).
Recientemente, se ha demostrado que la administración intramuscular de gamitromicina
produce una concentración inhibitoria mínima (CIM) por encima de lo requerido para inhibir el
crecimiento del 90% de organismos de R. equi en el BAL durante aproximadamente nueve días;
esto indica que la administración semanal de gamitromicina podría ser factible (Cohen, 2014).
La tulatromicina, un nuevo antibiótico macrólido sintético inyectable, aprobado para su uso
en porcinos y bovinos, se concentra en células broncoalveolares de potrillos luego de la
administración por vía intramuscular (Giguère et al., 2011; Venner et al., 2007). Sin embargo, la
tulatromicina ha demostrado ser menos efectiva que el tratamiento estándar de
azitromicina/rifampicina (Giguère et al., 2011; Muscatello, 2012).
Los criterios utilizados para determinar la duración del tratamiento, que generalmente puede
variar de 3 a 12 semanas, son la resolución de los signos clínicos, la normalización de los valores
plasmáticos de fibrinógeno y la resolución de las lesiones pulmonares a la ultrasonografía y
radiografía (Giguère et al., 2011).
Los tratamientos de soporte consisten en una adecuada nutrición e hidratación y en mantener
al potrillo afectado en un ambiente fresco y bien ventilado. El uso de antinflamatorios no esteroideos
o dipirona oral, así como las duchas frías, ayudan a controlar la hipertermia y mejorar la actitud en
potrillos letárgicos y/o anoréxicos. En casos de polisinovitis, artritis séptica u osteomielitis se
requiere un tratamiento agresivo que involucra lavajes articulares, desbridamiento quirúrgico y
perfusión endovenosa o intraósea de agentes antimicrobianos (Giguère et al., 2011; Kelmer et al.,
2011).

18
10. Pronóstico
En establecimientos endémicos que usan ultrasonografía torácica como método de
identificación temprana de la enfermedad junto con el tratamiento de los potrillos afectados, la tasa
de sobrevivencia es cercana al 100% (Giguère et al., 2011; Venner et al., 2012). El pronóstico del
rendimiento atlético de potrillos luego de un tratamiento exitoso contra bronconeumonías
producidas por R. equi es excelente (Giguère et al., 2011).
11. Bibliografía
1. Boyen, F.; Pasmans, F.; Haesebrouck, F. (2011). Acquired antimicrobial resistance in equine
Rhodococcus equi isolates. Vet Rec. 168, 1-2.
2. Boyd, N.K.; Cohen, N.D.; Lim, W.S.; Martens, R.J.; Chaffin, M.K.; Ball, J.M. (2003). Temporal
changes in cytokine expression of foals during the first month of life. Vet Immunol and
Immunopathol. 92, 75-85.
3. Brown, C.C.; Baker, D.C.; Barker, L.K. (2007). Pathology of domestic animals, ed. Saunders
Elsevier, 5th ed., p. 226. Philadelphia, PA.
4. Cauchard, J.; Sevin, C.; Ballet, J.; Taouji, S. (2004). Foal IgG and opsonizing anti Rhodococcus
equi antibodies after immunization of pregnant mares with a protective Vap-A candidate vaccine.
Vet Microb. 104, 73-81.
5. Campero, C.M.; Camezzana, O.R.; Elbusto, C.A.; Igarza, L.M. (1981). Bronconeumonía a
Corynebacterium equi en potrillos pura sangre de carrera. Gac Vet. 43, 775-781.
6. Caston, S.S.; McClure, R.S.; Martens, J.R.; Chaffin, K.M.; Miles, G.K.; Griffith, W.R.; Cohen,
D.N. (2006). Effect of hyperimmune plasma on the severity of pneumonia caused by
Rhodococcus equi in experimentally infected foals. Vet Therap. 7, 361-376.
7. Caswell, J.L.; Williams, K.J. (2016). Pathology of domestic animals, ed. Elsevier, 6th ed., Vol. 2,
pp. 569-571. St. Louis, MO.
8. Cohen, N.D. (2014). Rhodococcus equi foal pneumonia. Vet Clin Equine. 30, 609-622.
9. Cohen, N.D.; O’Conor, S.O.; Chaffin, M.K.; Martens, R.J. (2005). Farm characteristics and
management practices associated with development of Rhodococcus equi pneumonia in foals.
JAVMA. 226, 404-413.
10. Conville, P.S.; Witebsky, F.G. (2007). Manual of clinical microbiology, ed. ASM press, 9th ed.,
pp. 515-519. Washington, DC.
11. Clark-Price, S.C.; Rush, B.R.; Gaugham, E.M.; Cox, J.H. (2003). Osteomyelitis of the pelvis
caused by Rhodococcus equi in a two-year-old horse. JAVMA. 222, 969-972.

19
12. Cortez, J.; Aguilar, J.J.; Lagiogia, M.; Fernández, R.; Losino, L. (2008). Use of ultrasonography
to detect pulmonary lesions in thoroughbred foals in Argentina. Eq Vet Ed. 20, 154-158.
13. Dawson, T.R.; Horohov, D.W.; Meijer, W.G.; Muscatello, G. (2010). Current understanding of
the equine immune response to Rhodococcus equi. An immunological review of R. equi
pneumonia. Vet immunol and immunopathol. 135, 1-11.
14. Demmers, S.; Johannisson, A.; Grondahl, G.; Jensen-Waern, M. (2001). Neutrophil functions and
serum IgG in growing foals. Equine Vet J. 33, 676-680.
15. Flaminio, M.J.; Borges, A.S.; Nydam, D.V.; Horohov, D.Q.; Hecker, R.; Matychak, M.B. (2007).
The effect of CpG-ODN on antigen presenting cells of the foal. J Immun Based Ther Vacc. 5, 1-
17.
16. Flynn, O.; Quigley, F.; O’Grady, D.; Gogarty, A.; McGuirk, J.; Takai, S. (2001). Virulence
associated protein characterization of Rhodococcus equi isolated from bovine lymph nodes. Vet
Microbiol. 78 (3), 221-228.
17. Giguère, S. (2001). Rhodococcus equi pneumonia. Proceedings of the Annual Convention of the
AAEP. 47, 456-465.
18. Giguère, S.; Cohen, N.D.; Chaffin, K.; Slovis, N.M.; Holandus, M.K.; Hines, S.A.; Prescott, J.F.
(2011). Diagnosis, treatment, control and prevention of infections caused by Rhodococcus equi
in foals. J of Vet Inter Med. 25, 1209-1220.
19. Giguère, S.; Hernandez, J.; Gaskin, J.; Miller, C.; Bowman, J.L. (2003). Evaluation of white blood
cell concentration, plasma fibrinogen concentration, and an agar gel immunodiffusion test for
early identification of foals with Rhodococcus equi pneumonia. JAVMA.222, 775-781.
20. Giguère, S.; Jacks, S.; Roberts, G.D.; Hernandez, J.; Long, M.T.; Ellis, C. (2004). Retrospective
comparison of azitromycin, claritromycin, and eritromycin for the treatment of foals with
Rhodococcus equi pneumonia. J Vet Medic. 18, 568-573.
21. Harrington, J.R.; Golding, M.C.; Martens, R.J.; Halbert, N.D.; Cohen, N.D. (2005). Evaluation of
a real-time quantitative polymerase chain reaction assay for detection and quantitation of virulent
Rhodococcus equi. AJVR. 66, 755-761.
22. Hooper-McGrevy, K.E.; Wilkie, B.N.; Prescott, J.F. (2005). Virulence associated protein specific
serum immunoglobulin G-isotype expression in young foals protected against Rhodococcus equi
pneumonia by oral immunization with virulent R. equi. Vaccine. 23, 5760-5767.
23. Horowitz, M.L.; Cohen, N.D.; Takai, S.; Becu, T.; Chaffin, K.M.; Chu, K.K.; Magdesian, K.G.;
Martens, R.J. (2001). Application of Sartwell’s model (log-normal distribution of incubation
periods) to age at onset and age at death of foals with Rhodococcus equi pneumonia as evidence
of perinatal infection. J Vet Inter Med. 15, 171-175.

20
24. Ishino, S.; Kumagai, K.; Kuniyoshi, S.; Nakazawa, M.; Matzuda, I.; Oka, M. (1992).
Immunohistochemical observation on pneumonic lesions caused by Rhodococcus equi in foals. J
Vet Med Scien. 54, 509-515.
25. Jacks, S.; Giguère, S.; Crawford, P.C.; Castleman, W.L. (2007). Experimental infection of
neonatal foals with Rhodococcus equi triggers adults-like gamma interferon induction. Clin
Immunol. 14, 669-677.
26. Jain, S.; Bloom, B.J.; Holandus, M.K. (2003). Deletion of Vap-A encoding virulence associated
protein A attenuates the intracellular actinomycete Rhodococcus equi. Mol Microbiol. 50, 115-
128.
27. Kelmer, G.; Hayes, M.E. (2011). Regional limb perfusion with erythromycin for treatment of
septic physitis and arthritis caused by Rhodococcus equi. Vet Rec. 165, 291-292.
28. Kuskie, K.R.; Smith, J.L.; Wang, N.; Carter, C.N.; Chaffin, M.K.; Slovis, N.M.; Stepusin, R.S.;
Cattoi, A.E.; Muscatello, G.; Leadon, D.; Ocampo-Sosa, A.; Lewis, D.; Fogarty, U.; Buckley, T.;
Gilkerson, J.; Meijer, W.; Vazquez-Boland, J. (2007). Rhodococcus equi infection in foals: the
science of rattles. Equine Vet J. 39, 470-478.
29. Lyristis, M.; Bryant, A.E.; Sloan, J.; Awad, M.M.; Nisbet, I.T.; Stevens, D.L.; Rood, J.I. (1994).
Identification and molecular analysis of a locus that regulates extracellular toxin production in
Clostridium perfringens. Mol Microbiol. 12, 761-777.
30. MacArthur, I.; Parreira, V.R.; Lepp, D.; Mutharia, L.M.; Vazquez-Boland, J.A.; Prescott, J.F.
(2010). The sensor kinase MprB is required for Rhodococcus equi virulence. Vet Microbiol. 147,
133-141.
31. Madarame, H.; Takai, S.; Morisawa, N.; Fuji, M.; Hidaka, D.; Tsubaki, S.; Hasegawa, Y. (1996).
Immunohistochemical detection of virulence associated antigens of Rhodococcus equi in
pulmonary lesions of foals. Vet Pathol, 33, 341-343.
32. Mukherjee, S.; Mani, S. (2010). Orphan nuclear receptors as targets for drug development. Pharm
Res. 27, 1439-1468.
33. Muscatello, G. (2012). Rhodococcus equi pneumonia in the foal – part 2: diagnosis, treatment and
disease management. Vet J. 192, 27-33.
34. Muscatello, G.; Gilkerson, J.R.; Browning, G.F. (2009). Detection of virulent Rhodococcus equi in exhalated air samples from naturally infected foals. J Clin Microbiol. 47, 734-737.
35. Nicholson, V.M.; Prescott, J.F. (1997). Restriction enzyme analysis of the virulence plasmids of
Vap-A positive Rhodococcus equi strains isolated from humans and horses. J Clin Microbiol. 35,
738-740.

21
36. Noah, D.C.; Chaffin, K.; Martens, R.J. (2002). How to prevent and control pneumonia caused by
Rhodococcus equi at affected farms. Proceedings of the Annual Convention of the AAEP. 48,
295-299.
37. Pargass, I.S.; Wills, T.B.; Davis, W.C.; Wardrop, K.J.; Alperin, D.C.; Hines, S.A. (2009). The
influence of age and Rhodococcus equi infection on CD1 expression by equine presenting cells.
Vet Immunol Immunopathol. 130, 197-209.
38. Passamonti, F.; Vardi, D.M.; Stefanetti, V.; Marenzoni, M.L.; Prato, S.; Cevese, P.; Coletti, M.;
Pepe, M.; Casagrande, P.; Olea-popelka, F. (2015). Rhodococcus equi pneumonia in foals: an
assessment of the early diagnostic value of serum amyloid A and plasma fibrinogen
concentrations in equine clinical practice. Vet J. 203, 211-218.
39. Pei, Y.; Nicholson, V.; Woods, K.; Prescott, J.F. (2007). Immunization by intra-bronquial
administration to one week old foals of an unmarked double gene disruption strain of
Rhodococcus equi strain 103+. Vet Microbiol. 125, 100-110.
40. Prescott, J.F. (1991). Rhodococcus equi: an animal and human pathogen. Clin Microbiol Rev. 4,
20-34.
41. Pusterla, N.; Wilson, W.D.; Mapes, S.; Leutenegger, C.M. (2007). Diagnostic evaluation of real-
time PCR in the detection of Rhodococcus equi in faeces and nasopharyngeal swabs from foals
with pneumonia. Vet Rec. 161, 272-275.
42. Ramirez, S.; Lester, G.; Roberts, G. (2004). Diagnostic contribution of thoracic ultrasonography
in 17 foals with Rhodococcus equi pneumonia. Vet Radiol Ultrasound. 45, 172-176.
43. Rayn, C.; Giguère, S.; Hagen, J.; Hartnett, C.; Kalyuzhny, A.E. (2010). Effect of age and mitogen
on the frequency of interleukin-4 and interferon gamma secreting cells in foals and adult horses
as assessed by an equine-specific ELISPOT assay. Vet Immunol and Immunopathol. 133, 66-71.
44. Ren, J.; Prescott, J.F. (2004). The effect of mutation on Rhodococcus equi virulence plasmigene
expression and mouse virulence. Vet Microbiol. 103, 219-230.
45. Retteg, P.S.; Jottini, S.; Takai, S.; Venner, M.; Wohlsein, P. (2009). Immunohistological
demonstration of Rhodococcus equi in a trotter foal. Dtsch Tierarztl Wochenschr. 116, 335-339.
46. Reuss, S.M.; Cohen, N.D. (2015). Update on bacterial pneumonia in the foal and weanling. Vet
Clin North Am Equine Pract. doi: 10.1016/j.cveq. 2014.11.004.
47. Shimizu, T.; Ba-Thein, W.; Tamaki, M.; Hayashi, H. (1994). The virR gene, a member of a class
of two-component response regulators, regulates the production of perfringolysin O, collagenase,
and hemagglutinin in Clostridium perfringens. J Bacteriol. 176, 1616-1623.
48. Slovis, N. (2007). Rhodococcus equi pneumonia: new perspectives on screening. Proceedings of
the NAVC. January 13-27. Orlando, USA.

22
49. Slovis, N.; McCracken, J.; Mundy, G. (2005). How to use thoracic ultrasound to screen foals for
Rhodococcus equi at affected farms. Proceedings of the Annual Convention of the AAEP. 51.
50. Snyder, P.W. (2007). Diseases of immunity. En: Base patológica de enfermedades veterinarias,
5th ed., pp 204. Mosby Elsevier, St. Louis, MO.
51. Songer, J.G.; Post, K.W. (2005). Veterinary microbiology. Bacterial and fungal agents of animal
disease, 1st ed. Elsevier Saunders, p. 110. St, Louis, MO.
52. Stieler, A.L.; Sanchez, L.C.; Mallicote, M.F.; Martabano, B.B.; Burrow, J.A.; MacKay, R.J.
(2015). Macrolide-induced hyperthermia in foals: Role of impaired sweat response. Equine Vet.
J. 10, 1111-1115.
53. Takai, S.; Cohen, N.D. (2011). Effects of location for collection of air samples on a farm and
time of day of sample collection on airborne concentrations of virulent Rhodococcus equi at two
horse breeding farms. AJVR. 72, 73-79.
54. Tkachuk-saad, O.; Lusis, P.; Welsh, R.D. (1998). Rhodococcus equi infections in goats. Vet
Rec. 143, 311-312.
55. Valdes, A.; Johnson, J.R. (2005). Septic pleuritis and abdominal abscess formation caused by Rhodococcus equi in a foal. JAVMA. 227, 960-963.
56. Venner, M.; Kerth, R.; Klug, E. (2007). Evaluation of tulathromycin in the treatment of
pulmonary abscesses in foals. The Vet J. 174, 418-421.
57. Venner, M.; Rodiger, A.; Laemmer, M.; Giguère, S. (2012). Failure of antimicrobial therapy to
accelerate spontaneous healing of subclinical pulmonary abscesses on a farm with endemic
infections caused by Rhodococcus equi. Vet J. 192, 293-298.
58. VonBargen, K.; Haas, A. (2009). Molecular and infection biology of the horse pathogen
Rhodococcus equi. FEMS Microbiol Lett. 33, 870-891.
59. Webb, J.L. (2011). Leukocytes in clinical pathology, ed. Wiley-Blackwell, 5th ed., pp 70.
Ames, IO.
60. Wilkins, P.A. (2009). Large animal internal medicine, ed. Mosby Elsevier, 4th ed., p. 512. St,
Louis, MO.
61. Wilson, D.W. (1992). Foal pneumonia. An overview. Proceedings of the Annual Convention of
the AAEP. 38, 203-214.
62. Zachary, J.F. (2012). Pathologic basis of veterinary disease, ed. Mosby Elsevier, 5th ed., p. 179.
St, Louis, MO.

23
CAPÍTULO II Reproducción experimental y aspectos clínicos de bronconeumonías causadas por la cepa
103+ de Rhodococcus equi en potrillos
Introducción
La bronconeumonía producida por Rhodococcus equi ocupa un lugar importante entre las
enfermedades respiratorias en potrillos debido a la alta morbilidad y letalidad con la que se presenta
(Campero et al., 1981; Cortez et al., 2008; Noah et al., 2002). Esta enfermedad produce un
considerable impacto negativo en la producción equina nacional y mundial a través de la pérdida
directa de animales, baja ganancia de peso y gastos de tratamiento (Ainsworth et al., 1998; Cohen,
2014; Horohov et al., 2011; Takai et al., 2004).
La infección por R. equi ha sido reproducida experimentalmente en varias especies animales.
Sin embargo, solo el modelo experimental equino reproduce las lesiones pulmonares
piogranulomatosas típicas de esta enfermedad en la especie equina (Giguère et al., 1999; Jacks et
al., 2007; Wada et al., 1997). Los lechones y cobayos desarrollan lesiones pulmonares pero sin la
formación de piogranulomas y finalmente la infección es eliminada (Prescott, 1991). Los ratones
eliminan infecciones inducidas de forma experimental por vía endovenosa o intranasal que
inducirían enfermedad pulmonar en potrillos, demostrando así su relativa insensibilidad a este
microorganismo (Giguère et al., 1999; Jacks et al., 2007; VonBargen y Hass, 2008). El modelo
experimental de bronconeumonías por R. equi en potrillos ha sido utilizado en estudios de uso
profiláctico de plasma hiperinmune (Caston et al., 2006; Hooper-McGrevy et al., 2001), en
modulación de la respuesta de citoquinas mediada por plásmidos de virulencia de R. equi (Giguère
et al., 1999), en la evaluación del incremento en la expresión pulmonar de citoquinas inflamatorias
asociadas al incremento pulmonar del factor nuclear-kB (Uhl et al., 2002), en el uso terapéutico de
estreptolisina como adyuvante en el tratamiento de bronconeumonía causada por R. equi (Horohov
et al., 2011) y en la evaluación de vacunas inactivadas contra R. equi.
Muchos de los modelos de infección por R. equi en potrillos publicados hasta ahora, han
utilizado desafíos con dosis relativamente altas (1,4x107 UFC y 2,5x109 UFC por animal) que
inducen lesiones pulmonares severas y signos clínicos fulminantes entre los 3 y 10 días post
inoculación (Giguère et al., 1999; Hooper-McGrevy et al., 2001). Sin embargo, la edad promedio
al momento del diagnóstico de la bronconeumonía causada por R. equi en establecimientos
endémicos es de aproximadamente 37 días (Jacks et al., 2007). Este hallazgo indica que el período
de incubación en condiciones naturales es más largo que el observado en la reproducción
experimental con las dosis utilizadas hasta el momento. Ello es así, posiblemente debido a una

24
menor dosis infectiva y diferente vía de entrada. El objetivo de este trabajo fue demostrar que la
inoculación intrabronquial de R. equi con una dosis baja (inóculo total 5x10³ UFC por animal) en
potrillos menores de 6 meses, es suficiente para provocar el desarrollo de bronconeumonía
piogranulomatosa similar a la que se presenta en infecciones naturales. Además, este estudio
propone como herramientas para la detección temprana de la infección producida por R. equi, la
evaluación conjunta de: los signos clínicos respiratorios, la medición de la temperatura rectal, el
nivel de fibrinógeno sanguíneo y la detección de piogranulomas pulmonares mediante la utilización
de ultrasonografía torácica en potrillos con sospecha de enfermedad.
Hipótesis
1. La inoculación intrabronquial con una dosis de 5x10³ UFC/mL de la cepa 103+ de R. equi, es
suficiente para el desarrollo de bronconeumonía severa en potrillos menores de 6 meses.
2. La evaluación e interpretación en forma conjunta de los signos clínicos de distrés respiratorio,
aumento de temperatura, aumento de fibrinógeno y lesiones torácicas ultrasonográficas es suficiente
para realizar el diagnóstico de la enfermedad de forma temprana.
Objetivos
1. Determinar si la dosis de 5x10³ UFC/mL de la cepa 103+ de R. equi, es capaz de producir
bronconeumonía piogranulomatosa en potrillos menores de 6 meses.
2. Diagnosticar la infección pulmonar de forma precoz interpretando en conjunto los signos clínicos
respiratorios, aumento de temperatura, aumento de fibrinógeno y lesiones ultrasonográficas
pulmonares luego de la reproducción experimental.

25
1. Materiales y métodos
Este estudio fue aprobado por la comisión de Bienestar Animal de la Facultad de Ciencias
Veterinarias U.N.C.P.B.A. (aval número: 9/2018).
1.1 Animales
Se utilizaron 11 potrillos criollos/cruza, de 42 días de edad, provenientes de un proyecto de
investigación destinado a probar una vacuna contra R. equi. Dentro de los 2 días del nacimiento de
los potrillos, se obtuvieron los valores de frecuencia cardíaca, frecuencia respiratoria, temperatura
rectal y concentración de fibrinógeno sanguíneo. Estos parámetros fueron utilizados como valores
normales para cada uno de los potrillos.
1.2 Inóculo
Se utilizó la cepa 103+ de R. equi. El plásmido de virulencia con la isla de patogenicidad
(PAI) que codifica para la proteína asociada a virulencia A (Vap-A) se detectó en la totalidad de los
inóculos utilizados (Figura 1), mediante la técnica de reacción en cadena de la polimerasa (PCR) en
el Laboratorio de Virología de la Facultad de Ciencias Veterinarias U.N.C.P.B.A. Para obtener del
ADN plasmídico se utilizaron dos métodos diferentes. En primera instancia se emulsificó el ansa
completa de colonias bacterianas en 200 μL de agua, se hirvió durante 10 minutos, se centrifugó
durante 3 minutos y se utilizaron 3 μL como plantilla para PCR. Posteriormente, para una mayor
rigurosidad en el ensayo, se purificó el ADN plasmídico utilizando un kit de purificación de DNA
(Qiagen, Valencia, CA, USA) para bacterias gram positivas, de acuerdo a las instrucciones del
fabricante para bacterias gram positivas. El ADN obtenido se utilizó como templado para la
reacción de PCR. Brevemente, se combinaron 10 µL de 2x HF mastermix, 0,5 µL Fw-Vap-A
(100pmol) 5´TAATGCGACCGTTCTTGATTCC3´, 0,5 µL de Bw-Vap-A (100 pmol)
5´TGTAGAGACGCTGAAGGTCATTTG3´ y 9 µL de ddH20 haciendo un volumen total de 20 µL.
Posteriormente, se realizó el protocolo de PCR en un equipo termociclador PTC-100 (MJ Research),
se sembró en gel de agarosa y se coloreó con Sybr safe.

26
Figura 1: La línea 1 muestra la banda de 239 pares de bases correspondiente al inóculo. La línea 2 muestra
el control negativo. Las líneas 3, 4, 5 y 6 corresponden a diferentes controles positivos y la línea 7
corresponde a los marcadores de 250 pb, 500 pb, 750 pb, 1000 pb y 1250 pb.
Los primeros utilizados fueron Vap-A F 5`TAATGCGACCGTTCTTGATTCC 3` y Vap-A R
5`TGTAGAGACGCTGAAGGTCATTTG3`.
Para estimar la concentración bacteriana, las colonias conservadas en nitrógeno líquido
fueron sembradas en agar tripteína soya (Oxoid, CM 0129) durante 48 horas a 35 °C, luego se tomó
una colonia y fue inoculada en un pico de flauta con agar tripteína soya (Oxoid, CM 0129) seguido
por la incubación durante 48 horas a 35 °C. Posteriormente se realizó un lavaje con 3 mL de solución
fisiológica estéril para obtener una suspensión bacteriana. Dicha suspensión fue leída con un
espectrofotómetro a DO540nm y se ajustó a una densidad óptica de 0,17 correspondiente con 5x107
UFC/ml. Para la determinación exacta del número de UFC del inóculo, luego de 48 horas se realizó
el conteo de UFC mediante la utilización de diluciones seriadas de entre 1/10 y 1/160 hasta obtener
una concentración de 5x103 UFC/mL, de la cual una alícuota fue sembrada en agar tripteína soya
durante 48 horas a 35 °C con el objetivo de realizar un recuento retrospectivo de viabilidad, ya que
la densidad óptica no se ve alterada por la presencia de bacterias inviables. Finalmente, se tomó 1
ml de la suspensión bacteriana y se diluyó en 24 mL de solución fisiológica estéril para obtener el
inóculo final (Figura 2).

27
Diluciones seriadas
Recuento
retrospectivo- Viabilidad
Cada potrillo recibió 25 mL de 5x103 UFC/mL. Luego de la inoculación, los potrillos fueron
ubicados en parcelas y en grupos separados junto con sus madres, para su monitoreo y revisación.
Una vez que los potrillos se consideraron enfermos, los mismos se aislaron en boxes individuales
junto con sus madres para su seguimiento clínico y tratamiento.
48 hs. – 35 ˚C
TSA
48 hs. – 35 ˚C
Placas de agar tripteína soya
Pico de flauta
Lavaje 3 mL SFE
Suspensión bacteriana
Espectrofotómetro LO = 540 nm
se ajustó 7
DO = 0,17 = 5x10 UFC/mL
2 3 4 5 6 5x10 5x10 5x10 5x10 5x10 CFU/mL
Figura 2: Esquema del procedimiento para estimar la concentración y preparación del inóculo.
1.3 Inoculación
Previo a la inoculación, los potrillos fueron anestesiados de forma general con 0,5 mg/Kg
de xilacina, 0,02 mg/Kg de butorfanol, 4 mg/Kg de ketamina y 0,2 mg/Kg de diazepan (Muir, 2008).
Para la inoculación se utilizó un fibroscopio óptico flexible (Olimpus GIF) y una sonda de biopsia,
la cual se introdujo a través de una fosa nasal, laringe y tráquea hasta arribar al bronquio principal
derecho, luego se acopló la jeringa a la sonda de biopsia y se descargó el inóculo.
+ 24 mL SFE - inóculo

28
1.4 Criterio de enfermedad
Los potrillos inoculados se consideraron enfermos cuando presentaron uno o más de los
siguientes signos clínicos respiratorios: tos, taquipnea, disnea y respiración abdominal durante un
mínimo de 48 horas y áreas ecogénicas en la ultrasonografía torácica (interpretadas como
piogranulomas pulmonares) junto con temperatura rectal por encima de 39,5 °C durante 24 horas.
1.5 Estudio bacteriológico
A los potrillos considerados enfermos, de acuerdo al criterio de enfermedad descripto
anteriormente, se les realizó un lavaje broncoalveolar (BAL) para confirmar la presencia de R. equi
in vivo mediante el cultivo bacteriológico. Además, en los potrillos muertos se tomaron muestras
de pulmón en forma aséptica, las que fueron luego sembradas en agar tripteína soya durante 48
horas a 35 °C para el aislamiento bacteriológico.
El BAL fue realizado inmediatamente luego de la radiografía torácica, con el animal bajo
anestesia y previo a la instauración del tratamiento antibiótico (ver en p. 34). Para esta técnica se
utilizó un catéter de lavaje broncoalveolar bibona de 10 mm de diámetro por 240 cm de largo
(Laboratorios Jorgensen, CO, USA). El catéter fue introducido en las fosas nasales y a través de la
laringe hasta la bifurcación de la tráquea en donde se insufló el balón terminal. Se inyectaron 50
mL de solución salina estéril, la cual fue aspirada inmediatamente luego de su inyección para
obtener la muestra de BAL.
1.6 Estudio clínico
Los signos clínicos (tos, disnea, depresión, adinamia, distensión articular, hipopión e
hipema) fueron evaluados de forma subjetiva y registrados periódicamente durante 102 días post
inoculación o hasta la muerte espontánea de los potrillos.
La temperatura rectal y la frecuencia respiratoria se evaluaron cada 3 días. La medición de
la temperatura rectal se realizó con un termómetro clínico, considerándose anormal cuando la
temperatura superó los 39,5 °C. La frecuencia respiratoria se evaluó mediante la observación de los
movimientos del flanco con el animal en reposo, y fue considerada anormal cuando superó los 60
movimientos por minuto.

29
Es importante aclarar que, a pesar de que la temperatura rectal forma parte del examen clínico
general, la misma no posee un componente subjetivo como sí lo tiene la evaluación de los signos
clínicos respiratorios; por este motivo la temperatura fue considerada separadamente del examen
clínico.
1.7 Exámenes complementarios
1.7.1 Medición de fibrinógeno sanguíneo
Se determinó cada 7 días de forma indirecta a través de un refractómetro -utilizando sangre
de la vena yugular, colocada en tubos con y sin anticoagulante- para obtener plasma y suero,
respectivamente.
La diferencia entre el nivel de proteína total de plasma y suero fue multiplicada por 1 000
para obtener un resultado de fibrinógeno en mg/mL. Se consideró un valor anormal de fibrinógeno
por encima de 400 mg/dL. Los protocolos de la revisación clínica y exámenes complementarios se
grafican en la Figura 3.
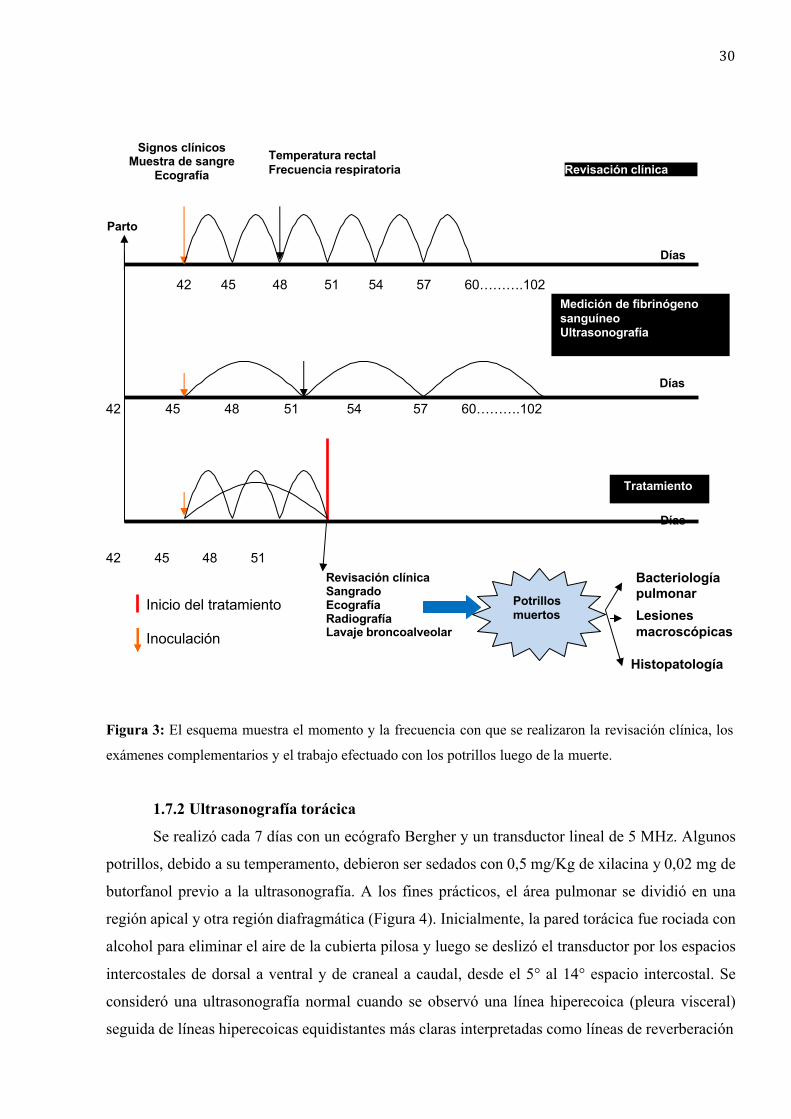
30
Signos clínicos Muestra de sangre
Ecografía
Temperatura rectal Frecuencia respiratoria Revisación clínica
Parto
Días
42 45 48 51 54 57 60……….102
42 45 48 51 54 57 60……….102
Días
Días
42 45 48 51
Inicio del tratamiento
Inoculación
Revisación clínica Sangrado Ecografía Radiografía Lavaje broncoalveolar
Potrillos muertos
Bacteriología pulmonar Lesiones macroscópicas
Histopatología
Figura 3: El esquema muestra el momento y la frecuencia con que se realizaron la revisación clínica, los
exámenes complementarios y el trabajo efectuado con los potrillos luego de la muerte.
1.7.2 Ultrasonografía torácica
Se realizó cada 7 días con un ecógrafo Bergher y un transductor lineal de 5 MHz. Algunos
potrillos, debido a su temperamento, debieron ser sedados con 0,5 mg/Kg de xilacina y 0,02 mg de
butorfanol previo a la ultrasonografía. A los fines prácticos, el área pulmonar se dividió en una
región apical y otra región diafragmática (Figura 4). Inicialmente, la pared torácica fue rociada con
alcohol para eliminar el aire de la cubierta pilosa y luego se deslizó el transductor por los espacios
intercostales de dorsal a ventral y de craneal a caudal, desde el 5° al 14° espacio intercostal. Se
consideró una ultrasonografía normal cuando se observó una línea hiperecoica (pleura visceral)
seguida de líneas hiperecoicas equidistantes más claras interpretadas como líneas de reverberación
Medición de fibrinógeno sanguíneo Ultrasonografía
Tratamiento

31
de la pleura visceral (Cortez, 2008). La observación de áreas hiperecoicas focales bien definidas de
tamaño variable, con un centro hipoecoico y generalmente sin la presencia de vías aéreas, fueron
interpretadas como piogranulomas pulmonares. Estas observaciones fueron registradas por
separado para los pulmones derecho e izquierdo.
Figura 4: Esquema de la región pulmonar apical (A) y de la región diafragmática (B) consideradas en este
estudio.
Para el registro del resultado ultrasonográfico se construyó un sistema de calificación de acuerdo al
tamaño de las lesiones (Tabla 1).
Tabla 1: Criterio para la interpretación ultrasonográfica.
Calificación Tamaño de las áreas ecogénicas (cm)
1 < 1
2 1 – 2
3 2,1 – 3
4 > 3
B A
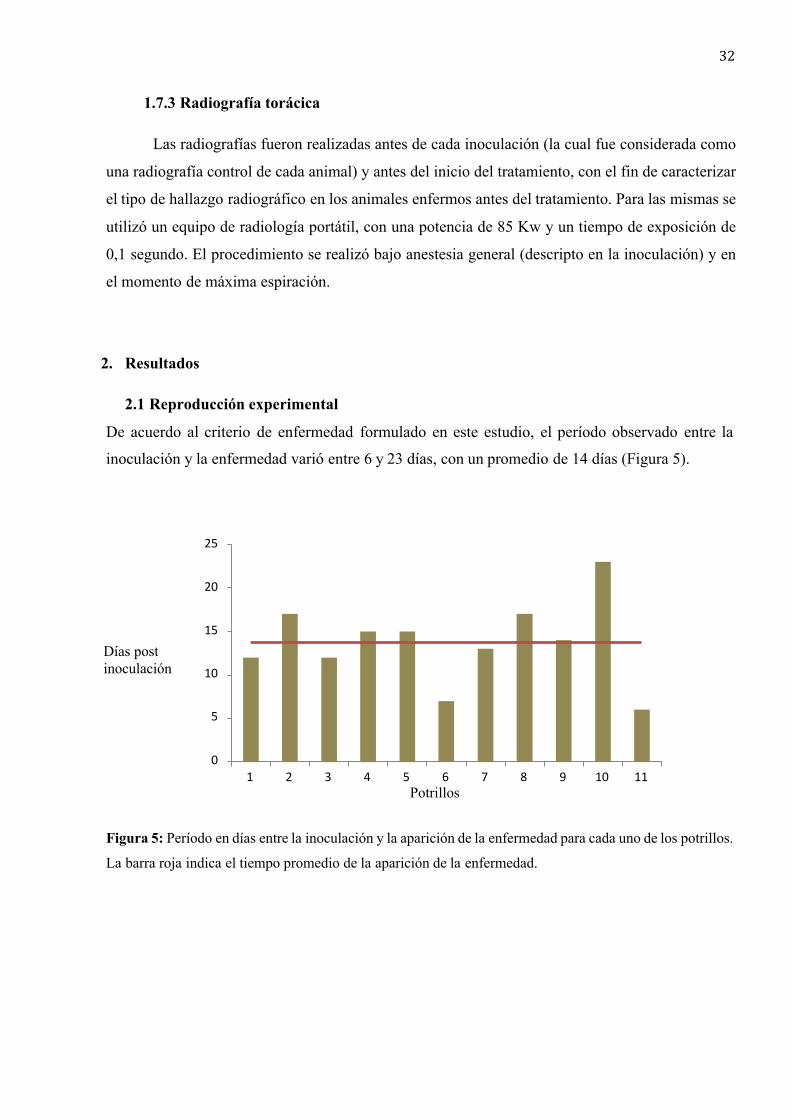
32
1.7.3 Radiografía torácica
Las radiografías fueron realizadas antes de cada inoculación (la cual fue considerada como
una radiografía control de cada animal) y antes del inicio del tratamiento, con el fin de caracterizar
el tipo de hallazgo radiográfico en los animales enfermos antes del tratamiento. Para las mismas se
utilizó un equipo de radiología portátil, con una potencia de 85 Kw y un tiempo de exposición de
0,1 segundo. El procedimiento se realizó bajo anestesia general (descripto en la inoculación) y en
el momento de máxima espiración.
2. Resultados
2.1 Reproducción experimental
De acuerdo al criterio de enfermedad formulado en este estudio, el período observado entre la
inoculación y la enfermedad varió entre 6 y 23 días, con un promedio de 14 días (Figura 5).
25
20
15
Días post inoculación 10
5
0 1 2 3 4 5 6 7 8 9 10 11
Potrillos
Figura 5: Período en días entre la inoculación y la aparición de la enfermedad para cada uno de los potrillos.
La barra roja indica el tiempo promedio de la aparición de la enfermedad.

33
El período observado entre la inoculación y la muerte varió entre 15 y 40 días, con un
promedio de 24 días (Figura 6). El 72% (8/11) de los potrillos enfermos murió de forma espontánea
a pesar del tratamiento, y a los restantes 3 animales (28%) se les practicó la eutanasia debido a la
severidad de los signos clínicos observados.
45
40
35
30
25
Días post 20 inoculación
15
10
5
0 1 2 3 4 5 6 7 8 9 10 11
Potrillos
Figura 6: Período entre la inoculación y la muerte. La barra roja indica el período promedio entre
inoculación y muerte.
2.2 Estudio bacteriológico
En el 91% (10/11) de las muestras de BAL de los potrillos muertos se aisló la cepa 103+ de
R. equi y el 91% (10/11) de los animales presentó aislamiento positivo proveniente de la muestra
de pulmón obtenida en la necropsia (Tabla 2).

34
Tabla 2: Detalle de los aislamientos obtenidos in vivo (BAL) y post mortem (pulmón).
Potrillos BAL Pulmón
1 + + 2 + + 3 - + 4 + + 5 + + 6 + + 7 + + 8 + + 9 + - 10 + + 11 + +
Promedio 91% 91%
Una vez que los potrillos se consideraron enfermos se comenzó el tratamiento antibiótico, aunque
dichos tratamientos tuvieron una duración variable; el promedio de los mismos fue de 10 días
(Figura 7).
35
30
25
20
15
10
5
01 2 3
4 5 6 7 8
9 10 11
Figura 7: Detalle de la duración de los tratamientos antibióticos en días para cada uno de los potrillos y el
promedio de duración de los mismos (barra azul).

35
2.3 Estudio clínico
Con respecto al porcentaje de aparición de los signos clínicos, el 63% (7/11) de los potrillos
considerados enfermos desarrolló signos clínicos que incluyeron (de mayor a menor severidad y
frecuencia de presentación) tos, taquipnea, disnea, hipopión/hipema, depresión y adinamia. El 100%
de los animales presentó aumento de temperatura rectal. El aumento de fibrinógeno sanguíneo
(superior a 400 mg/dL) se observó en el 81% (9/11) de los potrillos, con un rango de entre 600 y
800 mg/dL, y un promedio de 636 mg/dL. Las áreas ecogénicas por ultrasonografía, interpretadas
como piogranulomas pulmonares, se observaron en el 100% de los potrillos enfermos (Figura 8).
100
90
80
70
60% 50
40
30
20
10
0
Signos clínicos Aumento de temperatura
Fibrinógeno Ultrasonografía
Figura 8: Porcentaje de potrillos con cada uno de los 4 parámetros clínicos evaluados para diagnosticar la
enfermedad, luego de la infección experimental por R. equi.
El período promedio transcurrido desde la inoculación hasta la aparición de los signos
clínicos fue de 9 días. El aumento de temperatura rectal se observó a los 13 días promedio post
inoculación. El aumento de fibrinógeno sanguíneo fue detectado, en promedio, a los 12 días post
inoculación. La observación ultrasonográfica positiva se detectó entre los 6 y 18 días post
inoculación, con un promedio de 8 días (Figura 8). El período promedio para la detección de la
enfermedad fue de 10 días (Figura 9).

36
14,0
12,0
Días
10,0
8,0
6,0
4,0
2,0
0,0Signos clínicos Aumento de
temperatura
Fibrinógeno Ultrasonografía
Figura 9: Promedio de días desde la inoculación hasta la aparición de cada uno de los parámetros evaluados.
La barra azul indica el promedio de días desde la inoculación hasta la detección de la enfermedad.
La Figura 10 muestra la evolución de la temperatura rectal de cada uno de los potrillos
luego de la inoculación experimental.
41,5
41,0
40,5
40,0
39,5
39,0
38,5
38,0
37,5
0 2 4 6 8 10 12 14 16 18 20 22
Días
Figura 10: Evolución de la temperatura rectal post inoculación de cada potrillo. La barra roja señala el
valor de temperatura límite por encima del cual se consideró una temperatura anormal.
Temp
eratur
a

37
Días
2.4 Medición de fibrinógeno sanguíneo
La totalidad de los potrillos presentaron valores que no superaron los 0,2 mg/dL de fibrinógeno
sanguíneo preinoculación. La figura 11 muestra la evolución de los valores de fibrinógeno
sanguíneo de cada uno de los potrillos luego de la inoculación experimental.
1,4
1,2
1,0
0,8
0,6
0,4
0,2
0,0
-0,2
-0,4
-0,6
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
Días
Figura 11: Evolución del fibrinógeno sanguíneo postinoculación de cada potrillo. La barra negra punteada
señala el valor de fibrinógeno por encima del cual se consideró anormal.
2.5 Ultrasonografía torácica
En la ultrasonografía torácica de un equino sano es posible observar 2 líneas hiperecoicas
paralelas, que desde el exterior al interior corresponden a: i) la piel y ii) la superficie pleural
(incluyendo pleura visceral y parietal) y al espacio pleural, respectivamente (Figura 12).
Fibr
inog
eno
(mg/
dl)

38
Figura 12: Ultrasonografía torácica de un equino sano. Es posible observar desde el exterior hacia el interior:
la primera línea hiperecoica correspondiente a la piel ( ), la segunda línea hiperecoica correspondiente a la
superficie pleural y al espacio pleural ( ) y una región hipoecoica difusa correspondiente al parénquima
pulmonar ( ).
Los hallazgos ultrasonográficos observados en los pulmones derecho e izquierdo de los
potrillos enfermos se resumen en la Tabla 3. En todos los potrillos enfermos se observaron lesiones
ultrasonográficas en el pulmón derecho. Aunque la inoculación fue realizada en el bronquio
derecho, el 36% (4/11) de los potrillos considerados enfermos presentó áreas ecogénicas de
calificación 1 en el pulmón izquierdo y solo un potrillo presentó áreas ecogénicas de calificación 4.

39
Tabla 3: Detalle de la calificación de las lesiones ultrasonográficas observadas en los pulmones
derecho ( ) e izquierdo ( ).
Potrillos Calificación de las áreas ecogénicas
1 2 3 4
1
2
3
4
5 6
7
8
9
10
11
En las ultrasonografías torácicas se observaron áreas ecogénicas con calificación variable de
1 a 4 y sus posibles combinaciones, entre las que se encuentran: múltiples (> 20) áreas ecogénicas de
calificación 1 (Figura 13), y áreas ecogénicas extensamente focales de calificación 4, que en los
casos más severos formaron grandes áreas de cavitación con bordes hiperecoicos de forma irregular
(Figura 14) y en un potrillo se observó engrosamiento de las pleuras (visceral y parietal) y del
espacio pleural (Figura 15).

40
Figura 13: Esta ultrasonografía del potrillo 5 muestra una zona con múltiples áreas ecogénicas de
calificación 1 (círculo negro), que afecta al parénquima de forma coalescente.
Figura 14: Esta ultrasonografía del potrillo 2 muestra un área ecogénica de calificación 4, caracterizada por
un centro hipoecoico de una cavidad como consecuencia del acúmulo de exudado inflamatorio de
piogranulomas coalescentes ( ) y múltiples zonas hiperecoicas en la periferia de la cavitación, resultado de la
formación de un piogranuloma pulmonar ( ).
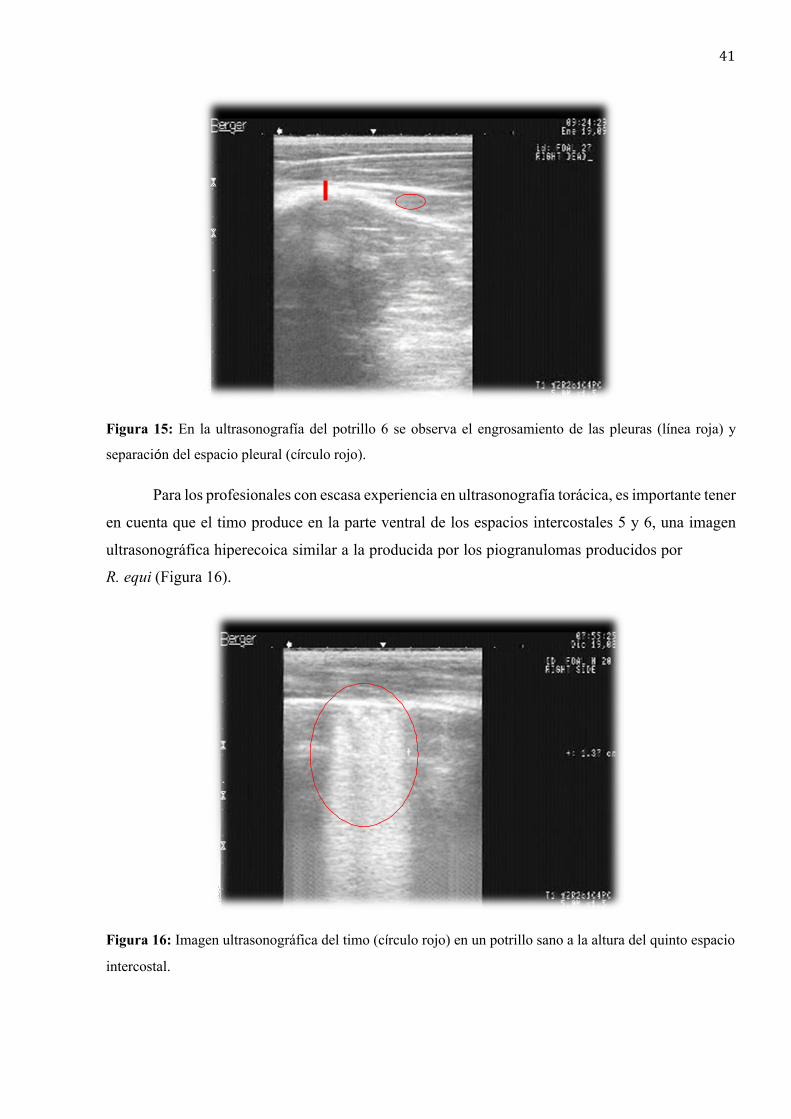
41
Figura 15: En la ultrasonografía del potrillo 6 se observa el engrosamiento de las pleuras (línea roja) y
separación del espacio pleural (círculo rojo).
Para los profesionales con escasa experiencia en ultrasonografía torácica, es importante tener
en cuenta que el timo produce en la parte ventral de los espacios intercostales 5 y 6, una imagen
ultrasonográfica hiperecoica similar a la producida por los piogranulomas producidos por
R. equi (Figura 16).
Figura 16: Imagen ultrasonográfica del timo (círculo rojo) en un potrillo sano a la altura del quinto espacio
intercostal.

42
2.6 Radiografía torácica
Todos los potrillos incluidos en este estudio presentaron algún tipo de hallazgo radiográfico
compatible con la infección por R. equi (Tabla 4).
Tabla 4: Detalle de los hallazgos radiográficos observados para cada potrillo.
Potrillos Consolidación
parenquimatosa
Broncograma
aéreo
Patrón intersticial Patrón alveolar
1 + + - -
2 + + - -
3 + + - +
4 + + - -
5 + + - -
6 + + - -
7 + + - -
8 + + + -
9 + + - -
10 + + - -
11 + + + -

43
En la radiografía torácica de un potrillo sano es posible reconocer distintas estructuras
anatómicas, incluyendo la vista frontal del bronquio principal derecho, la aorta, la arteria pulmonar,
la vena cava caudal, el corazón, el diafragma y las costillas (Figura 17).
Figura 17: Radiografía torácica de un potrillo sano que muestra la vista frontal del bronquio principal
derecho, la cual produce una imagen hiperclara redonda (negra) por encima del corazón. La aorta y la arteria
pulmonar producen una imagen radiopaca elongada (blanco) y el corazón produce una imagen radiopaca
globosa en la región ventral.
El 100% de los potrillos presentó consolidación parenquimatosa y broncograma aéreo
positivo (Figura 18), el 18% (2/11) desarrolló un patrón intersticial (Figura 19), el cual consiste en
un engrosamiento del intersticio peribronquiolar, y el 9% (1/11) presentó un patrón alveolar, el cual
fue interpretado como un hallazgo previo al desarrollo de broncograma aéreo.
Costilla
Aorta
Arteria pulmonar
Vena cava caudal
Bifurcación del bronquio principal
derecho Diafragma
Corazón

44
Figura 18: El potrillo 1 muestra consolidación del parénquima pulmonar (*) y broncograma aéreo positivo
en la región caudodorsal (†).
Figura 19: El potrillo 8 muestra un patrón intersticial en la región caudodorsal (*).
†

45
3. Discusión y conclusión
Este estudio demostró que es posible reproducir experimentalmente bronconeumonía en
potrillos con la inoculación de una dosis total de 5x103 UFC/mL. Otros estudios, los cuales tuvieron
un objetivo diferente al de esta tesis, incluyendo lo publicado por Giguère et al. (1999) y Uhl et al.
(2002), utilizaron una dosis de inóculo 400 000 veces mayor al inóculo utilizado en este estudio.
Hooper-McGrevy et al. (2001) utilizaron una dosis 10 000 veces mayor y Horohov et al. (2011)
utilizaron una dosis 300 000 veces mayor. En ambos estudios se produjeron lesiones pulmonares
severas en un período más corto que el observado en este trabajo (Tabla 5) (Giguère et al., 1999;
Hooper-McGrevy et al., 2001; Horohov et al., 2011; Uhl et al., 2002).
Con respecto a los resultados bacteriológicos del BAL, creemos que el resultado negativo
obtenido en el potrillo 3 pudo haberse debido a una dilución de la muestra obtenida. A pesar de que
todos los potrillos considerados enfermos fueron tratados con antibióticos, en la mayoría de los
animales se logró un aislamiento bacteriano de la cepa 103+ de R. equi. Especulamos que esto
podría haberse debido a que la concentración de antibiótico no fue la suficiente para eliminar las
bacterias inoculadas, a excepción del potrillo 9.
El intervalo entre la inoculación y la aparición de signos clínicos respiratorios, aumento de
temperatura rectal, incremento del fibrinógeno sanguíneo y abscesos pulmonares por
ultrasonografía, coinciden con los trabajos realizados por Giguère et al. (1999), Hooper-McGrevy
et al. (2001), Horohov et al. (2011) y Uhl et al. (2002) (Tabla 5). Sin embargo, no es posible
comparar el intervalo entre la inoculación y la muerte causada por la infección experimental, debido
a que con la excepción de este estudio, el resto de los trabajos utilizó un tiempo fijo de eutanasia
(Giguère et al., 1999; Hooper-McGrevy et al., 2001; Horohov et al., 2011; Uhl et al., 2002).
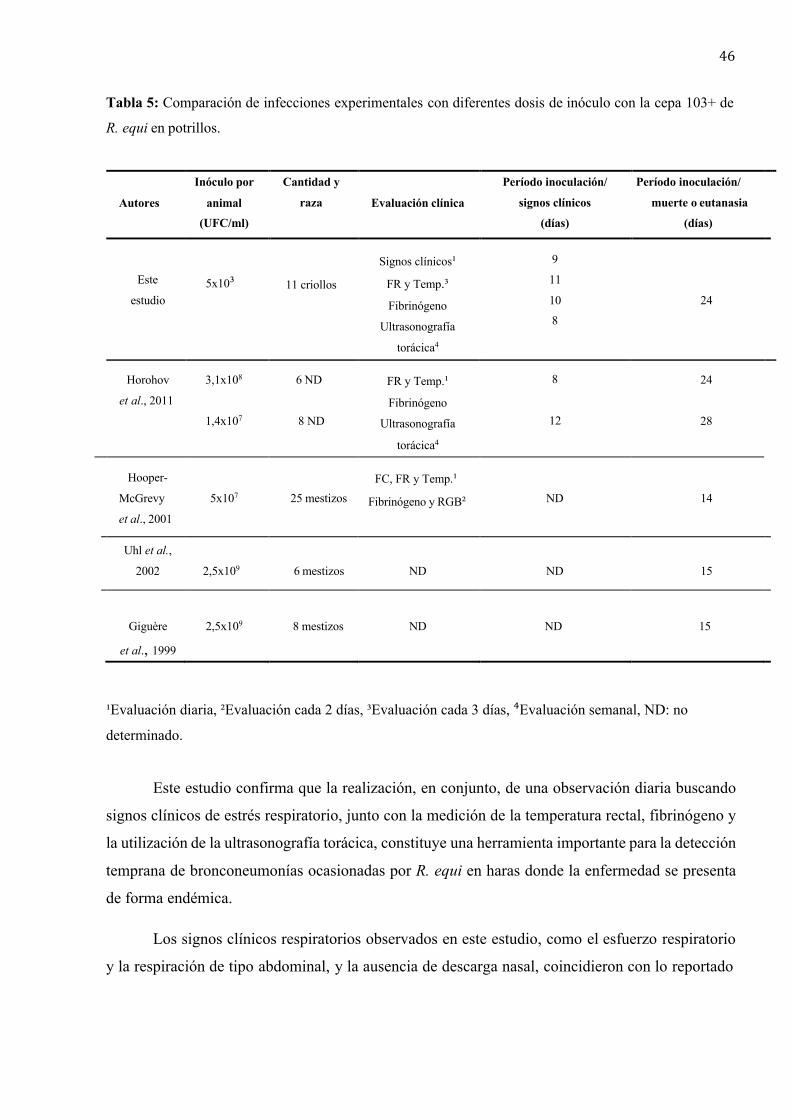
46
Tabla 5: Comparación de infecciones experimentales con diferentes dosis de inóculo con la cepa 103+ de
R. equi en potrillos.
Autores
Inóculo por
animal
(UFC/ml)
Cantidad y
raza
Evaluación clínica
Período inoculación/
signos clínicos
(días)
Período inoculación/
muerte o eutanasia
(días)
Este
estudio
5x10³
11 criollos
Signos clínicos¹
FR y Temp.³ Fibrinógeno
Ultrasonografía
torácica4
9
11
10
8
24
Horohov
et al., 2011
3,1x108
1,4x107
6 ND
8 ND
FR y Temp.¹ Fibrinógeno
Ultrasonografía
torácica4
8
12
24
28
Hooper-
McGrevy
et al., 2001
5x107
25 mestizos FC, FR y Temp.¹
Fibrinógeno y RGB²
ND
14
Uhl et al.,
2002
2,5x109
6 mestizos
ND
ND
15
Giguère
et al., 1999
2,5x109
8 mestizos
ND
ND
15
¹Evaluación diaria, ²Evaluación cada 2 días, ³Evaluación cada 3 días, ⁴Evaluación semanal, ND: no
determinado.
Este estudio confirma que la realización, en conjunto, de una observación diaria buscando
signos clínicos de estrés respiratorio, junto con la medición de la temperatura rectal, fibrinógeno y
la utilización de la ultrasonografía torácica, constituye una herramienta importante para la detección
temprana de bronconeumonías ocasionadas por R. equi en haras donde la enfermedad se presenta
de forma endémica.
Los signos clínicos respiratorios observados en este estudio, como el esfuerzo respiratorio
y la respiración de tipo abdominal, y la ausencia de descarga nasal, coincidieron con lo reportado

47
por Horohov et al., (2011) y Cohen (2014). En el resto de los trabajos no se determinó el tipo de
signo clínico ni el período de aparición de los mismos.
En coincidencia con Horohov et al., (2011), todos los animales inoculados desarrollaron un
aumento de temperatura y valores elevados de fibrinógeno, aunque nuestro estudio detectó un
aumento de temperatura 3 días más tarde y valores de fibrinógeno elevados 2 días más tarde que lo
reportado por Horohov et al., (2011).
En este estudio, al igual que lo observado por Giguère et al., (1999), todas las lesiones
pulmonares fueron detectadas a través de la ultrasonografía torácica. Los hallazgos
ultrasonográficos más característicos de las lesiones pulmonares descriptos en la literatura son áreas
ecogénicas producidas por los piogranulomas. En este estudio también se detectó engrosamiento
pleural. Posiblemente este hallazgo es debido no solo a la acumulación de células inflamatorias sino
también de fibrina. En potrillos es importante tener en cuenta que en la región torácica apical
ventral, a nivel de la quinta costilla y por delante del corazón, el timo puede producir una imagen
hiperecoica similar a las lesiones piogranulomatosas producidas por R. equi; esto puede ser difícil
de diferenciar de lesiones pulmonares. El timo adquiere su máximo tamaño a los 2 meses de edad,
disminuyendo progresivamente a medida que el potrillo crece (Farrow, 2007).
El broncograma aéreo observado en la totalidad de los potrillos es un signo radiológico que
indica que dentro de un área de consolidación existe un bronquio que todavía contiene aire. Es
posible observar esta imagen en focos de bronconeumonías, no así en tumores. Es importante tener
en cuenta que los patrones radiográficos de enfermedad pulmonar (intersticial, alveolar, bronquial)
tienen poca correlación con las lesiones histopatológicas. Por ejemplo, los equinos con un patrón
pulmonar intersticial difuso a la radiografía, presentan lesiones peribronquiales en el estudio
histopatológico (Farrow, 2007). También los patrones radiográficos reflejan la severidad de la
enfermedad; así, patrones intersticiales y bronquiales indican una lesión menos severa que el patrón
alveolar (Farrow, 2007).
Los resultados de este capítulo sugieren que es posible mejorar el modelo equino de
infección pulmonar por R. equi, acercándolo hacia lo que ocurre en condiciones naturales,
utilizando una dosis de inóculo sustancialmente menor a la utilizada hasta el momento.
Actualmente no existe un método de diagnóstico único con la suficiente especificidad y
sensibilidad para el diagnóstico in vivo de bronconeumonía causada por R. equi (Muscatello, 2012;
Slovis, 2007). Por este motivo, este estudio sugiere -coincidiendo con lo reportado por Cohen

48
(2014)- la utilización del criterio de enfermedad aquí propuesto, en un intento de mejorar la
detección temprana de la infección por R. equi y mejorar la elección de los animales para el
tratamiento. Esto se debe a que solo el 21% de los potrillos con evidencia ultrasonográfica de
consolidación pulmonar puede desarrollar los signos clínicos de bronconeumonía.
4. Bibliografía
1. Aisworth, D.M.; Eicker, S.W.; Yeagar, A.E.; Sweeney, C.R.; Viel, L.; Tesarowski, D.;
Lavoie, J.P.; Hoffman, A.; Paradis, M.R.; Reed, S.M.; Erb, H.N.; Davidow, E.; Nalevanko,
M. (1998). Associations between physical examination, laboratory, and radiographic
findings and outcome and subsequent racing performance of foals with Rhodococcus equi
infection: 115 cases (1984-1992). J Am Vet Med Assoc. 213, 510-515.
2. Campero, C.M.; Camezzana, O.R.; Elbusto, C.A.; Igarza, L.M. (1981). Bronconeumonía a
Corynebacterium equi en potrillos pura sangre de carrera. Gac Vet. 43, 775-781.
3. Caston, S.S.; McClure, R.S.; Martens, J.R.; Chaffin, K.M.; Miles, G.K.; Griffith, W.R.;
Cohen, D.N. (2006). Effect of hyperimmune plasma on the severity of pneumonia caused
by Rhodococcus equi in experimentally infected foals. Vet Therap. 7, 361-376.
4. Cohen, N.D. (2014). Rhodococcus equi foal pneumonia. Vet Clin Equine. 30, 609-622.
5. Cortez, J.; Aguilar, J.J.; Lagiogia, M.; Fernández, R.; Losino, L. (2008). Use of
ultrasonography to detect pulmonary lesions in thoroughbred foals in Argentina. Eq Vet Ed.
20, 154-158.
6. Farrow C.S. (2007). The equine lung, pp. 463-475. En: Thrall D.E. Vet Diag Radiology.
Saunders, Philadelphia.
7. Giguère, S.; Holandus, M.K.; Yager, J.A.; Darrah, P.; Mosser, D.M.; Prescott, J.F. (1999).
Role of the 85-kilobase plasmid and plasmid-encoded virulence-associated protein A in
intracellular survival and virulence of Rhodococcus equi. Infect Immun. 67, 3548-3557.
8. Hooper-McGrevy, K.E.; Giguère, S.; Wilkie, B.N.; Prescott, J.F. (2001). Evaluation of
equine immunoglobulin specific for Rhodococcus equi virulence-associated protein A and
C for use in protecting foals against Rhodococcus equi-induced pneumonia. AJVR. 62,
1307-1313.
9. Horohov, D.W.; Loynachan, A.T.; Page, A.E.; Hughes, K.; Timoney, J.F.; Fettinger, M.;
Hatch, T.; Spaulding, J.G.; McMichael, J. (2011). The use of streptolysin O (SLO) as an
adjunct therapy for Rhodococcus equi pneumonia in foals. Vet Microbiol. 154, 156-162.

49
10. Jacks, S.; Giguère, S.; Crawford, P.C.; Castleman, W.L. (2007). Experimental infection of
neonatal foals with Rhodococcus equi triggers adult-like gamma interferon induction. Clin
Vaccine Immunol. 14, 669-677.
11. Muir, W.W. (2008). Procedimientos y técnicas anestésicas en el caballo, pp. 391-393. En:
Manual de Anestesia Veterinaria. Elsevier, Madrid.
12. Muscatello, G. (2012). Rhodococcus equi pneumonia in the foal –part 2: diagnosis,
treatment and disease management. Vet J. 192, 27-33.
13. Noah, D.C.; Chaffin, K.; Martens, R.J. (2002). How to prevent and control pneumonia
caused by Rhodococcus equi at affected farms. Proceedings of the Annual Convention of
the AAEP. 48, 295-299.
14. Prescott, J.F. (1991). Rhodococcus equi: an animal and human pathogen. Clin Microbiol
Reviews. 4, 20-34.
15. Slovis, N. (2007). Rhodococcus equi pneumonia: new perspectives on screening.
Proceedings of the NAVC. January 13-27. Orlando, USA.
16. Takai, S.; Yamatoda, N.; Kimura, A.; Sasaki, Y.; Kakuda, T.; Tsubaki, S.; Bemis, D.; Brace,
J.J. (2004). Prevalence of virulent Rhodococcus equi in soil environment on a horse-
breeding farm in Tennessee, U.S.A. J Eq Scien. 15, 75-79.
17. Uhl, E.W.; Giguère, S.; Jack, T.J.; Hodge, T. (2002). Increased pulmonary activation of
nuclear factor-κB (NF-κB) in foals inoculated with Rhodococcus equi is associated with
increased expression of inflammation cytokines. Vet Pathol. 39, 132-136.
18. VonBargen, K.; Haas, A. (2008). Molecular and infection biology of the horse pathogen
Rhodococcus equi. FEMS. 33, 870-891.
19. Wada, R.; Kamada, M.; Anzai, T.; Nakanishi, A.; Kanemaru, T.; Takai, S.; Tsubaki, S.
(1997). Pathogenicity and virulence of Rhodococcus equi in foals following intra-tracheal
challenge. Vet Microbiol. 56, 301-312.

50
CAPÍTULO III Estudio anatomopatológico e inmunohistoquímico de lesiones pulmonares inducidas
experimentalmente por la cepa 103+ de Rhodococcus equi en potrillos
Introducción
Rhodococcus equi es un cocobacilo gram positivo, intracelular facultativo que causa
bronconeumonía en potrillos menores de 6 meses y también en humanos inmunocomprometidos o
bajo tratamiento inmunosupresivo (Darrah et al., 2000). Esta bacteria produce enzimas solubles
(fosfolipasa y colesterol oxidasa) llamadas “factores equi”. Sin embargo, su virulencia está asociada
a la presencia de un plásmido de virulencia de entre 85 y 90 kb que codifica para las proteínas
asociadas a la virulencia llamadas proteínas Vap-A, C, D, E, G y H, siendo la proteína Vap-A la
más importante para la virulencia de este microorganismo (Giguère, 2001; Harrington et al., 2005;
Muscatello, 2012).
Cuando R. equi es inhalado, se deposita en la mucosa del sistema de conducción respiratorio
(tráquea, bronquios y bronquiolos) debido a la fuerza centrífuga y turbulencia del aire inspirado
(Muscatello et al., 2007). Si este microorganismo logra alcanzar la unión bronquioloalveolar, el
mismo ya no puede ser eliminado por el barrido mucociliar y es fagocitado, previa opsonización de
R. equi, con anticuerpos (Ac) o con la fracción C3b del complemento, por los macrófagos
alveolares. Además, es captado por las células dendríticas mediante endocitosis, a través de
seudópodos que penetran entre los estrechos espacios del epitelio respiratorio (Zachary, 2012). Los
macrófagos procesan los antígenos y los presentan asociados al CMH II, a los linfocitos T
colaboradores sensibilizados. Las células dendríticas transportan la bacteria hacia los linfonódulos
regionales ingresando a través de los vasos linfáticos aferentes, donde presentan los antígenos
asociados al CMH II, a los linfocitos T colaboradores vírgenes (Zachary, 2012). Usualmente,
durante la fagocitosis por parte de los macrófagos, este microorganismo es envuelto por una porción
de la membrana plasmática y es internalizado, formando una nueva organela llamada fagosoma, la
cual posteriormente se fusiona con vesículas lisosomales y forma el fagolisosoma. Normalmente,
en el fagolisosoma la bacteria es expuesta a numerosos factores bactericidas como pH ácido (4,0-
5,0), enzimas hidrolíticas (proteasas, lipasas, ADNasas, ARNasas, entre otras) y metabolitos
reactivos del oxígeno (radicales superóxidos, peróxido de hidrógeno y óxido nítrico) que
contribuyen a la destrucción y eliminación de la bacteria (Demmers et al., 2001; Wilkins, 2009).
Sin embargo, en los macrófagos que han fagocitado cepas virulentas de R. equi no se produce la
maduración del fagosoma y como consecuencia hay ausencia o disminución de la formación del
fagolisosoma, lo que permite la replicación intracelular de R. equi (VonBargen y Haas, 2009).

51
Es sabido que la respuesta inmune adaptativa mediada por células de tipo Th-1 es necesaria
para eliminar la infección por R. equi. Esta respuesta se caracteriza por la presencia de macrófagos
y células dendríticas que producen citoquinas tales como IL-2, IL-12, IL-18, γ-IFN y FNT-α que
estimulan a linfocitos T indiferenciados a diferenciarse hacia una respuesta Th-1 (Dawson et al.,
2010).
Desde el punto de vista patológico, la inflamación granulomatosa puede ser dividida en una
forma nodular, que es el resultado de una respuesta inmunológica Th-1 y una forma lepromatosa o
difusa, la que se considera consistente con una respuesta inmunológica Th-2. También puede ser
producida por reacciones alérgicas en mucosas y piel (Zachary, 2012). La forma granulomatosa
nodular puede o no tener un centro necrótico y puede ser de forma redonda, oval, irregular o
multinodular y de tamaño variable. Así, en un granuloma clásico es posible distinguir tres zonas:
una zona central con necrosis rodeada por una zona intermedia de macrófagos epitelioides y células
gigantes multinucleadas y una zona externa con linfocitos T y B, células plasmáticas y fibroblastos
(Zachary, 2012). El granuloma lepromatoso es una inflamación pobremente delimitada con
numerosos macrófagos dispuestos en láminas, poca cantidad de linfocitos y células plasmáticas, y
sin la presencia de fibroblastos. Cuando la lesión granulomatosa incluye abundante cantidad de
neutrófilos mezclados con las demás células inflamatorias, se denomina piogranuloma; esta es la
lesión típica producida por R. equi. El piogranuloma está compuesto por un centro de necrosis
caracterizado por restos celulares entremezclados con gran cantidad de neutrófilos viables y
degenerados, y macrófagos, incluyendo macrófagos epitelioides. Esta área central está rodeada por
una zona periférica de infiltración con linfocitos, células plasmáticasy células gigantes
multinucleadas tipo Langhans. Por último, la capa externa está formada por bandas de tejido fibroso.
La lesión macroscópica pulmonar producida por R. equi está caracterizada por áreas firmes
multifocales a coalescentes consolidadas de color blanco amarillento, con un tamaño variable de
entre 1 y 10 cm de diámetro (Zachary, 2012). Estas lesiones suelen ser erróneamente interpretadas
como abscesos; sin embargo, estos no poseen una cápsula fibrosa (Caswell y Williams, 2007).
Microscópicamente, la lesión pulmonar se clasifica como una bronconeumonía piogranulomatosa
licuefactiva (Caswell y Williams, 2016).
En este estudio se llevó a cabo la caracterización de las lesiones pulmonares macroscópicas
y microscópicas de potrillos infectados experimentalmente con una dosis baja de R. equi (5x10³
UFC/mL) y se utilizó de la técnica de inmunohistoquímica para la detección de linfocitos T (CD3)
y linfocitos B (CD79) para confirmar la respuesta inmune Th-1.

52
Hipótesis
La inoculación experimental intrabronquial con una dosis de 5x10³ UFC/mL de la cepa 103+ de R.
equi produce lesiones pulmonares macroscópicas y microscópicas, con una respuesta inmune Th-1
similar a las descriptas en infecciones naturales.
Objetivo
Identificar y describir las lesiones pulmonares macroscópicas y microscópicas producidas por la
inoculación experimental de R. equi a baja dosis y confirmar, mediante técnicas de
inmunohistoquímica, una respuesta inmune Th-1.
1. Materiales y métodos
1.1 Animales
Se analizaron 11 potrillos descriptos en el capítulo II de esta tesis (p. 25), de los cuales el
72% (8/11) murió de forma espontánea y el 28% (3/11) restante fue sacrificado debido a la severidad
de los signos clínicos observados. La eutanasia se realizó mediante la inducción de una anestesia
general con xilacina (1,1 mg/Kg) y ketamina (2,2 mg/Kg) y posterior inyección endovenosa de 150
mL de solución hipertónica de cloruro de potasio (Muir, 2008). Los procedimientos de eutanasia
fueron aprobados por la Comisión de Bienestar Animal de la Facultad de Ciencias Veterinarias
U.N.C.P.B.A. (aval número: 9/2018).
1.2 Inoculación experimental
La preparación del inóculo (p. 26), el procedimiento de inoculación (p. 28) y el criterio de
enfermedad (p. 29) fueron descriptos en el capítulo II de esta tesis.

53
1.3 Estudio anatomopatológico
Las necropsias se efectuaron entre las 2 y 8 horas de ocurrida la muerte, y se realizó tanto
un registro fotográfico de las lesiones macroscópicas como la caracterización de las lesiones
macroscópicas pulmonares y extrapulmonares. Se tomaron muestras de pulmón, linfonódulos
mediastínicos y mesentéricos, corazón, hígado, bazo, glándula adrenal, riñón, intestino delgado e
intestino grueso, las cuales fueron fijadas por inmersión en formol bufferado al 10%, pH 7,4 durante
24-48 horas. Posteriormente se deshidrataron en alcohol y xileno, antes de ser embebidos en
parafina. Las muestras procesadas fueron cortadas en secciones de 5 micras de espesor y teñidas
con hematoxilina-eosina, de acuerdo al procedimiento estándar para su observación microscópica.
1.4 Tinciones especiales
Cortes seleccionados de pulmón fueron coloreados con las técnicas especiales de tinción de
hematoxilina ácida fosfotúngstica (PTAH), tricrómica de Masson y Gram.
1.5 Evaluación inmunohistoquímica
Muestras seleccionadas de pulmón de todos los potrillos fueron procesadas mediante
técnicas de inmunohistoquímica, para la detección de componentes de antígeno-receptor de
linfocitos T (CD3) y antígeno-receptor de linfocitos B (CD79a). La técnica de inmunohistoquímica
se realizó utilizando el método de inmuno-marcado con un conjugado avidina-biotina (ABC)
siguiendo las instrucciones del fabricante (Dako North America, Carpinteria, CA) y el
procedimiento estándar del Veterinary Medical Teaching Hospital de la Universidad de California–
Davis. Para la recuperación del antígeno se utilizó vapor de citrato bufferado, seguido de un
anticuerpo secundario específico de la especie equina. Contra el receptor CD3-12 se utilizó un
anticuerpo primario monoclonal de rata (Dr. Peter Moore, Department of Pathology, Microbiology
and Immunology, School of Veterinary Sciences, University of California, CA) y contra el receptor
CD79a-HM57 se utilizó un anticuerpo monoclonal de ratón (Dako North America, Carpinteria,
CA). Se utilizaron linfonódulos de potrillos sanos como controles positivos. Cortes de los mismos
tacos procesados sin el anticuerpo primario se utilizaron como controles negativos. En cada sección
de pulmón procesada para la identificación de linfocitos B y T se evaluaron 10 campos a 400x y se
le asignó un rango subjetivo de 0 a 5, de acuerdo al porcentaje de linfocitos positivos. Se consideró
grado 0 cuando no se observaron linfocitos, grado 1 cuando se observó un 20%, grado 2 cuando se
observó un 40%, grado 3 cuando se observó un 60%, grado 4 cuando se observó un 80% y grado 5
cuando se observó un 100%. Los resultados para cada potrillo fueron volcados en la Tabla 4 de
abundancia celular relativa (linfocitos T y B).

54
1.6 Evaluación de la respuesta inmune
Para dicha evaluación fue necesario identificar los distintos tipos celulares encontrados en
las lesiones pulmonares de los potrillos afectados. La identificación de macrófagos, neutrófilos,
eosinófilos, mastocitos y fibroblastos fue realizada teniendo en cuenta la morfología celular con la
tinción hematoxilina-eosina. La identificación de los linfocitos T y B fue realizada mediante la
técnica de inmunohistoquímica anteriormente descripta. Se evaluó de forma subjetiva el porcentaje
de las células inflamatorias identificadas mediante morfología celular e inmunohistoquímica en 10
campos de 400x. A cada porcentaje observado se le asignó un rango subjetivo de 0 a 5. Así, se
consideró grado 0 cuando no se observaron esas células inflamatorias, grado 1 cuando se observó
un 20% del tipo celular evaluado, grado 2 cuando se observó un 40%, grado 3 cuando se observó
un 60%, grado 4 cuando se observó un 80% y grado 5 cuando se observó un 100% del tipo celular
evaluado. Posteriormente, el resultado total de dicha evaluación expresado en porcentaje y en
grados, fue volcado en la Tabla 4 de abundancia celular relativa.
2. Resultados
2.1 Patología macroscópica
La totalidad de los potrillos desafiados presentaron lesiones pulmonares macroscópicas
similares. A la apertura de la cavidad torácica los pulmones no colapsaron. Los pulmones
presentaban aspecto marmóreo caracterizado por áreas de color blanco amarillento de forma
irregular intercaladas con áreas rojo oscuro, afectando en promedio entre el 40 y 80% de la región
caudoventral (Figura 1).
Figura 1: La región caudoventral del pulmón derecho muestra un área de aspecto marmóreo (círculo blanco).
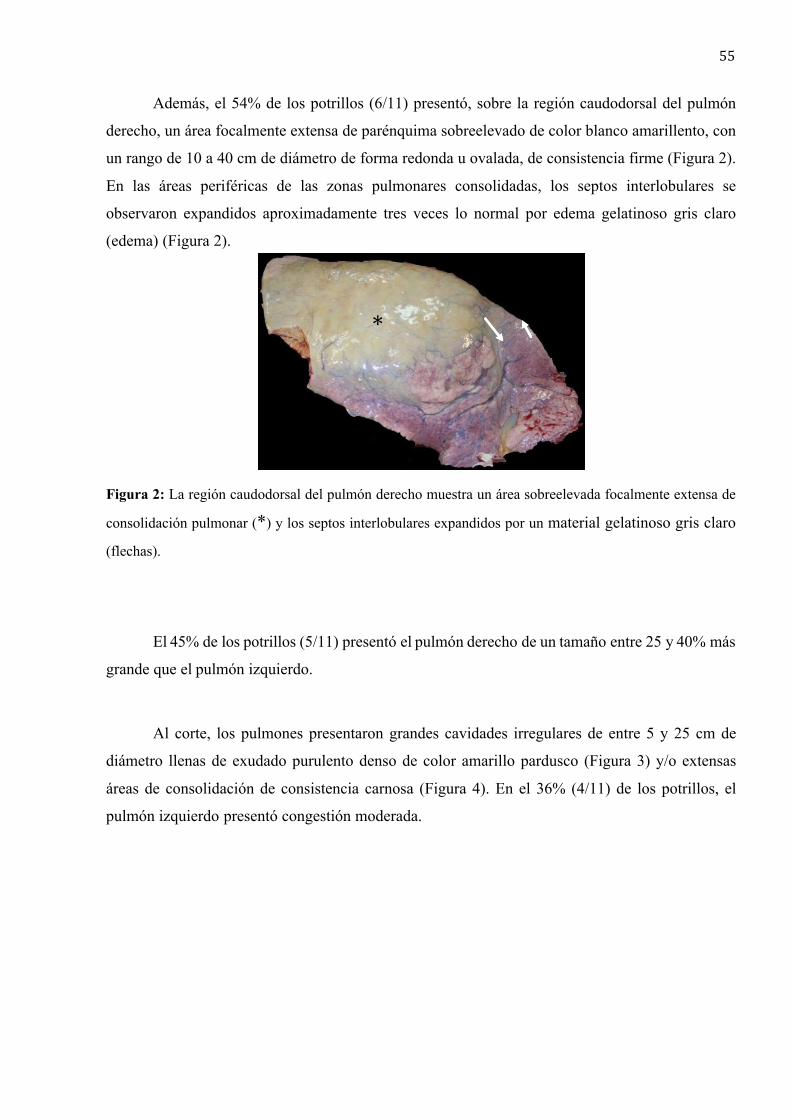
55
Además, el 54% de los potrillos (6/11) presentó, sobre la región caudodorsal del pulmón
derecho, un área focalmente extensa de parénquima sobreelevado de color blanco amarillento, con
un rango de 10 a 40 cm de diámetro de forma redonda u ovalada, de consistencia firme (Figura 2).
En las áreas periféricas de las zonas pulmonares consolidadas, los septos interlobulares se
observaron expandidos aproximadamente tres veces lo normal por edema gelatinoso gris claro
(edema) (Figura 2).
Figura 2: La región caudodorsal del pulmón derecho muestra un área sobreelevada focalmente extensa de
consolidación pulmonar (*) y los septos interlobulares expandidos por un material gelatinoso gris claro
(flechas).
El 45% de los potrillos (5/11) presentó el pulmón derecho de un tamaño entre 25 y 40% más
grande que el pulmón izquierdo.
Al corte, los pulmones presentaron grandes cavidades irregulares de entre 5 y 25 cm de
diámetro llenas de exudado purulento denso de color amarillo pardusco (Figura 3) y/o extensas
áreas de consolidación de consistencia carnosa (Figura 4). En el 36% (4/11) de los potrillos, el
pulmón izquierdo presentó congestión moderada.

56
Figura 3: Lesión pulmonar cavernosa rellena con abundante cantidad de exudado purulento.
Figura 4: Áreas focalmente extensas y coalescentes de consolidación pulmonar de aspecto carnoso (*).
En la tráquea torácica, se observó la presencia de exudado mucopurulento de color amarillo
intenso (Figura 5), mezclado ocasionalmente con abundante cantidad de espuma estable.
Figura 5: La luz de la tráquea muestra abundante cantidad de exudado mucopurulento.

57
Las lesiones macroscópicas extrapulmonares se consignan en la tabla 1 y se describen a
continuación. El 36% de los potrillos muertos (4/11) desarrolló hipopión (Figura 6) o hipema
bilateral (Figura 7).
Figura 6: Hipopión en el ojo izquierdo. Figura 7: Hipema en el ojo derecho.
El 27% (3/11) de los potrillos presentó un puntillado rojo oscuro (patrón centrolobulillar)
afectando el 80% del parénquima hepático (Figura 8).
Figura 8: El hígado muestra un puntillado rojo oscuro distribuido de forma difusa.
Un potrillo presentó en los riñones congestión severa y grandes áreas irregulares de color
pardo diseminadas en la superficie (Figura 9).

58
Figura 9: La superficie renal presenta un aspecto marmóreo, con áreas de color pardo y otras de color rojo
oscuro distribuidas en forma difusa.
Un potrillo presentó hemorragias subserosas multifocales diseminadas en el duodeno y
yeyuno, con abundantes coágulos de sangre libres en el lumen cubriendo la mucosa congestiva
(Figura 10). La mucosa del ciego de este animal presentó múltiples petequias y en la luz del colon
mayor ventral se encontró abundante cantidad de materia fecal semiblanda de color rojo oscuro
(Figura 11).
Figura 10: Segmentos de duodeno y yeyuno con hemorragias multifocales en la serosa y contenido
hemorrágico de forma difusa.
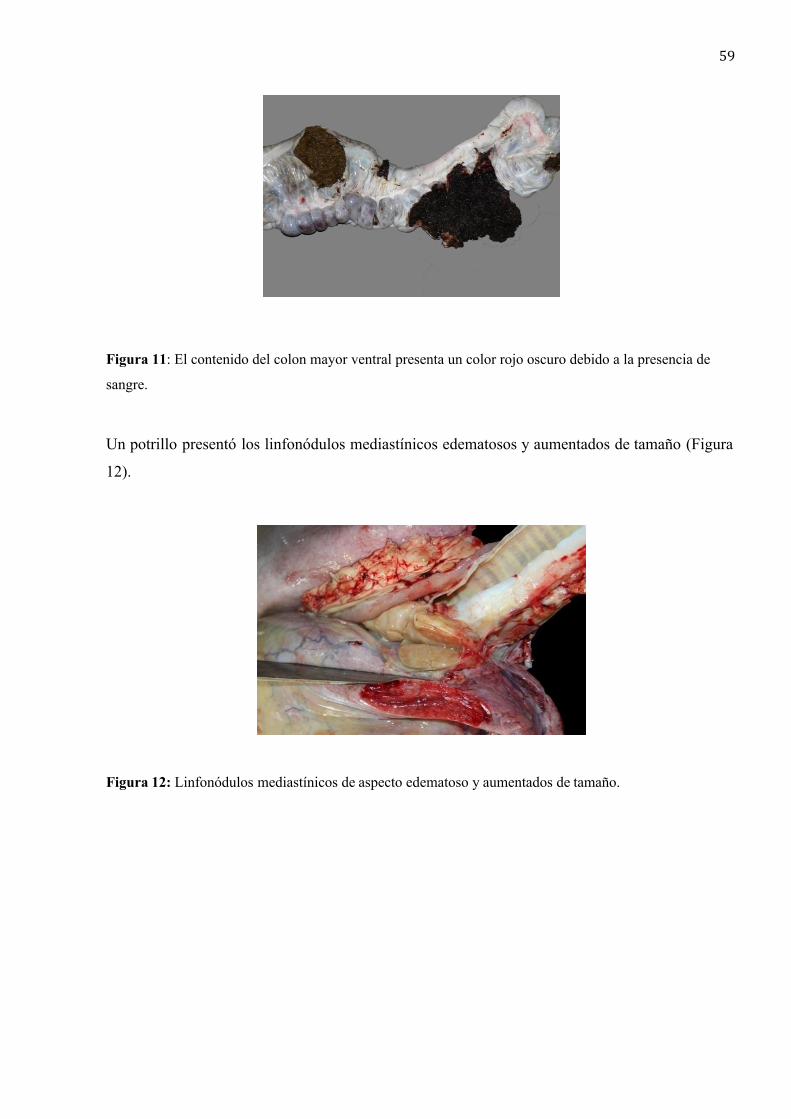
59
Figura 11: El contenido del colon mayor ventral presenta un color rojo oscuro debido a la presencia de
sangre.
Un potrillo presentó los linfonódulos mediastínicos edematosos y aumentados de tamaño (Figura
12).
Figura 12: Linfonódulos mediastínicos de aspecto edematoso y aumentados de tamaño.
60
Tabla 1: Gráfico de las lesiones macroscópicas extrapulmonares observadas en los potrillos
inoculados con la cepa 103+ de R. equi.
Potrillo Ojo Hígado Riñón Intestino Linfonódulos
mesentéricos
1 Hipopión SLA SLA SLA SLA 2 SLA Patrón
centrolobulillar
SLA SLA SLA
3 SLA SLA SLA SLA SLA 4 Hipopión Patrón
centrolobulillar
SLA SLA SLA
5 SLA SLA SLA SLA SLA 6 SLA SLA Congestión SLA SLA 7 Hipopión SLA SLA Hemorragia SLA 8 SLA Patrón
centrolobulillar
SLA SLA Edema
9 SLA SLA SLA SLA SLA 10 SLA SLA SLA SLA SLA 11 Hipema SLA SLA SLA SLA
SLA: Sin lesiones aparentes.
2.2 Patología microscópica y tinciones especiales
Las lesiones microscópicas pulmonares se consignan en la tabla 2 y se describen a
continuación. El 100% (11/11) de los potrillos presentó bronconeumonía piogranulomatosa. El
espacio alveolar y el lumen de los bronquios y bronquiolos estaban ocupados por abundante
cantidad de neutrófilos viables y degenerados, y macrófagos espumosos con ocasionales
cocobacilos intracitoplasmáticos basofílicos gram positivos, de entre 2 y 3 µm de largo. Además,
se observó una moderada cantidad de células gigantes multinucleadas del tipo Langhans y una
moderada cantidad de linfocitos y células plasmáticas que frecuentemente se encontraron
mezclados con fibrina, restos celulares y células epiteliales necróticas descamadas. Los septos
alveolares estaban engrosados principalmente por una gran cantidad de neutrófilos viables y
degenerados; macrófagos y una menor cantidad de linfocitos y células plasmáticas. El parénquima
pulmonar periférico a los piogranulomas estaba comprimido (atelectasia) y/o con grandes áreas
claras irregulares (enfisema) (Figura 13).

61
Figura 13: Una sección de pulmón muestra el parénquima periférico comprimido (atelectasia) (
de enfisema compensatorio (. ) GH&E. 40x.
y áreas
El 20% (2/11) de los potrillos presentó, en las lesiones pulmonares, macrófagos alveolares
con cocobacilos intracitoplasmáticos basofílicos gram positivos (Figura 14).
Figura 14: Se observa un macrófago alveolar con abundante cantidad de bacterias intracitoplasmáticas
gram positivas dentro del citoplasma (flecha). Tinción de Gram. 400x.
)

62
El 54% (6/11) de los potrillos presentó, en los espacios alveolares de las áreas periféricas,
una moderada a severa cantidad de un material amorfo eosinofílico (edema proteináceo).
En el 63% (7/11) de los potrillos se observó que las paredes alveolares se encontraban
revestidas, de forma multifocal, con células cuboides con un citoplasma que contenía
microvacuolas claras (neumocitos tipo II), lo que se interpretó como un fenómeno de epitelización
(Figura 15).
Figura 15: Se observa un espacio alveolar ocupado por abundante cantidad de neumocitos degenerados y
necróticos descamados mezclados con macrófagos. Las paredes interalveolares se encuentran revestidas
por células cuboidales grandes interpretadas como neumocitos tipo II (flechas), fenómeno conocido como
epitelización. H&E. 400x.

63
El 45% (5/11) de los potrillos presentó focos de inflamación piogranulomatosa licuefactiva
pulmonar severa caracterizada por una gran cantidad de células inflamatorias viables y
degeneradas, y restos celulares (Figura 16).
Figura 16: Área extensa de inflamación piogranulomatosa licuefactiva ( ) bien delimitada, caracterizada
por abundante cantidad de restos celulares mezclados con células inflamatorias viables y degeneradas.
H&E. 100x.
En el 36% (4/11) de los potrillos se observó, en el lumen de algunas venas y arterias
pulmonares de tamaño mediano, un material de color rosa claro amorfo frecuentemente adherido
a las paredes vasculares, ocluyendo de forma parcial la luz de las mismas (trombosis) (Figura 17).
En todos los animales que presentaron trombosis pulmonar, dicho material resultó ser positivo a
la Tinción PTAH (Figura 18).

64
Figura 17: Se observa una vena de calibre mediano que contiene, en el lumen, un material de color rosa
claro amorfo (flecha) interpretado como trombosis. H&E.100x.
Figura 18: La luz de una arteria presenta una oclusión parcial, con hebras de fibrina de color azul (flecha).
Tinción PTAH. 200x.
En el 27% (3/11) de los potrillos se observó, alrededor de las zonas de bronconeumonía
piogranulomatosa, un tejido denso caracterizado por una cantidad moderada de fibroblastos y
fibras colágenas e infiltrado por una cantidad moderada de linfocitos y células plasmáticas,
interpretado como fibroplasia (Figura 19). En el 72% (8/11) de los potrillos afectados se observó,
con la tinción de hematoxilina-eosina, proliferación de fibroblastos en las lesiones pulmonares. En
todas esas lesiones se resaltó, de forma más especifica, la abundancia de fibras colágenas mediante
la tinción tricrómica de Masson (Figura 20).
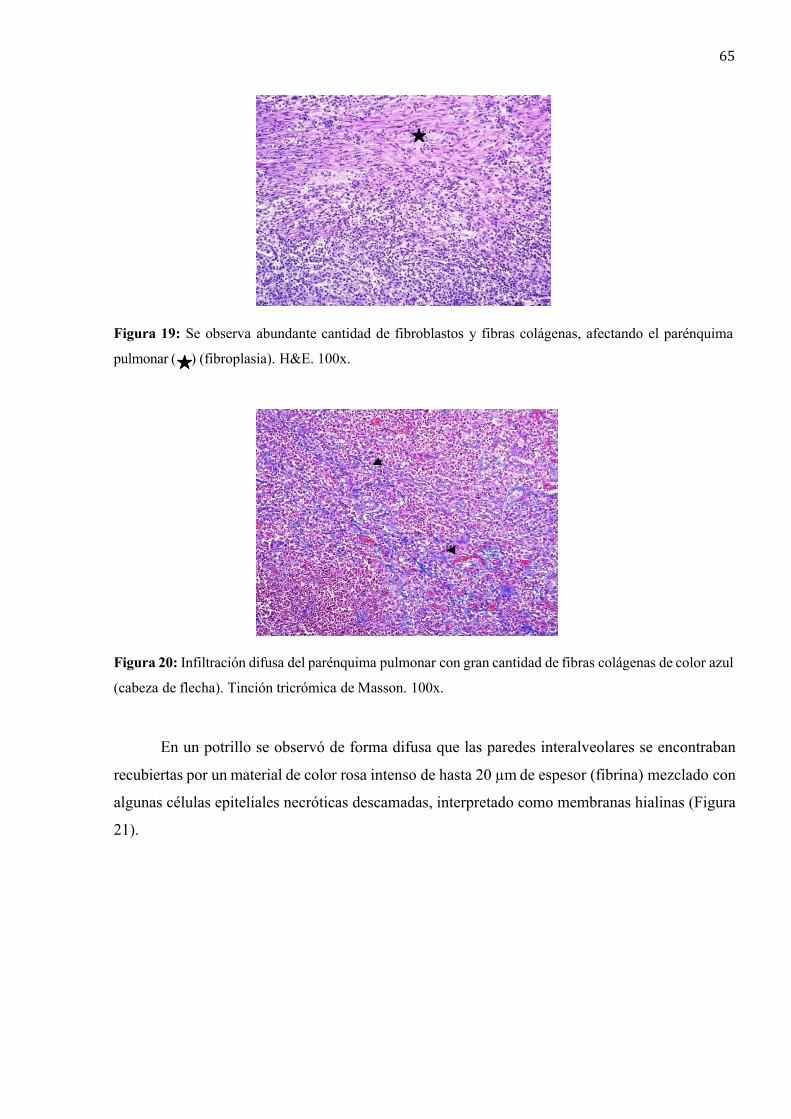
65
Figura 19: Se observa abundante cantidad de fibroblastos y fibras colágenas, afectando el parénquima
pulmonar ( ) (fibroplasia). H&E. 100x.
Figura 20: Infiltración difusa del parénquima pulmonar con gran cantidad de fibras colágenas de color azul
(cabeza de flecha). Tinción tricrómica de Masson. 100x.
En un potrillo se observó de forma difusa que las paredes interalveolares se encontraban
recubiertas por un material de color rosa intenso de hasta 20 µm de espesor (fibrina) mezclado con
algunas células epiteliales necróticas descamadas, interpretado como membranas hialinas (Figura
21).

66
Figura 21: Las paredes alveolares están recubiertas por una capa de un material de color rosa intenso, de
hasta 20 µm de espesor (flechas) (membranas hialinas). H&E. 200x.
Tabla 2: Lesiones microscópicas pulmonares observadas en los potrillos inoculados con la cepa
103+ de R. equi.
Potrillo Nº
Epitelización
Edema
Necrosis
Trombosis
Fibroplasia Membranas
hialinas
1 + + - - + -
2 + - + - - -
3 + + + + - -
4 - + - + - +
5 + + + + - -
6 + - - + - -
7 - + + - + -
8 + - - - - -
9 + - + - + -
10 - - - - - -
11 - + - - - -
Las lesiones microscópicas extrapulmonares se consignan en la Tabla 3 y se describen a
continuación. El 54% (6/11) de los animales presentó hepatitis y/o necrosis hepática
centrolobulillar y mediozonal. La hepatitis se caracterizó por la infiltración de una cantidad
moderada de células inflamatorias (principalmente neutrófilos y macrófagos), mientras que las
áreas necróticas presentaron desorganización y pérdida de hepatocitos con hemorragia moderada
(Figura 22).

67
Figura 22: El área hepática mediozonal (*) presenta desorganización del parénquima con pérdida de
hepatocitos. H&E. 100x.
El 45% (5/11) de los potrillos presentó linfadenitis y necrosis subcapsular moderada en los
linfonódulos mediastínicos con grandes áreas irregulares con linfocitos degenerados o necróticos
(linfolisis) mezclados con macrófagos espumosos y edema (Figura 23). Las áreas afectadas se
caracterizaron por la presencia de restos celulares mezclados con abundante cantidad de
neutrófilos viables y degenerados, macrófagos espumosos, células gigantes multinucleadas tipo
Langhans, en las cuales ocasionalmente se observaron vacuolas intracitoplasmáticas claras
conteniendo cocobacilos basofílicos y escasa cantidad de linfocitos y células plasmáticas.
*

68
Figura 23: El tejido linfoide de la paracorteza de un linfonódulo muestra áreas con linfocitos viables ( )
y áreas con abundante cantidad de linfocitos degenerados y necróticos (linfolisis) y restos celulares
(flechas), los cuales están separados por una moderada cantidad de un material granular de color rosa claro
(edema). H&E. 400x.
El 18% (2/11) de los potrillos presentó nefritis intersticial (Figura 24), caracterizada por un
infiltrado con macrófagos espumosos y linfocitos. Los glomérulos se encontraron rodeados por
hemorragia y los capilares se encontraron con numerosos émbolos bacterianos basófilos,
mezclados con un material amorfo eosinofílico interpretado como fibrina. Ocasionalmente, el
lumen de algunos túbulos contorneados proximales se encontró lleno de un material eosinofílico
amorfo interpretado como cilindros de proteína. Los vasos sanguíneos de pequeño calibre de la
corteza renal exhibieron necrosis fibrinoide transmural severa, mezclada con neutrófilos
degenerados y trombosis.
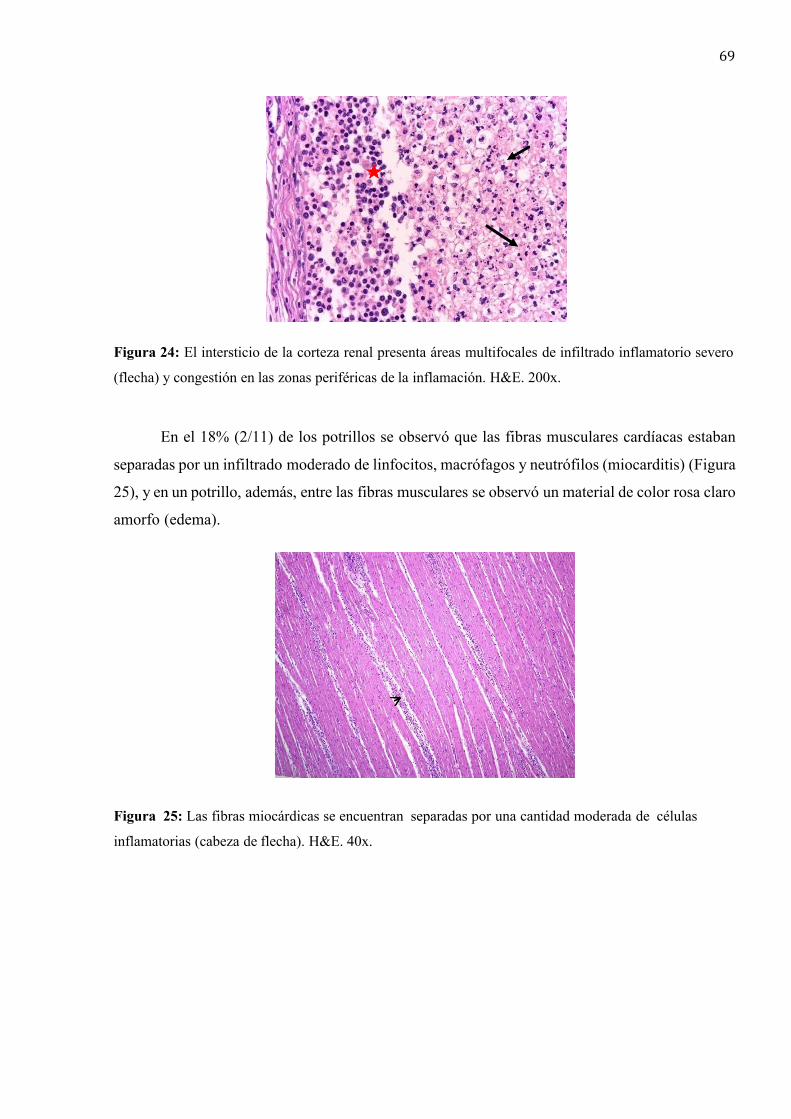
69
Figura 24: El intersticio de la corteza renal presenta áreas multifocales de infiltrado inflamatorio severo
(flecha) y congestión en las zonas periféricas de la inflamación. H&E. 200x.
En el 18% (2/11) de los potrillos se observó que las fibras musculares cardíacas estaban
separadas por un infiltrado moderado de linfocitos, macrófagos y neutrófilos (miocarditis) (Figura
25), y en un potrillo, además, entre las fibras musculares se observó un material de color rosa claro
amorfo (edema).
Figura 25: Las fibras miocárdicas se encuentran separadas por una cantidad moderada de células
inflamatorias (cabeza de flecha). H&E. 40x.

70
En el 9% (1/11) de los potrillos se observó que de forma multifocal, el lumen del intestino
delgado estaba ocupado por sangre (Figura 26), mezclada con células epiteliales descamadas desde
la mucosa (autolisis). En la lámina propia se observó un infiltrado inflamatorio difuso
caracterizado por una moderada cantidad de macrófagos, linfocitos y células plasmáticas y una
escasa cantidad de neutrófilos (Figura 27). Ocasionalmente algunos vasos sanguíneos de la mucosa
estaban ocluidos por trombos de fibrina (Figura 28). No se observaron cambios microscópicos en
la mucosa del intestino grueso.
Figura 26: El intestino delgado muestra la mucosa cubierta de forma focal por abundante cantidad de
hemorragia (flecha). H&E. 40x.
Figura 27: La lámina propia del intestino delgado se encuentra expandida por un infiltrado inflamatorio
(flechas) caraterizado por una moderada cantidad de macrófagos, linfocitos, células plasmáticas y una
menor cantidad de neutrófilos. H&E. 400x.

71
Figura 28: La mucosa del intestino delgado muestra el acortamiento moderado de las vellosidades,
desprovistas de células epiteliales (círculo negro) y, ocasionalmente, la luz de algunos vasos sanguíneos
pequeños de la lámina propia presenta trombosis fibrinosa (flecha). H&E. 100x.
Tabla 3: Lesiones microscópicas extrapulmonares observadas en los potrillos inoculados con la
cepa 103+ de R. equi.
Casos Hígado Linfonódulos Riñón Corazón Intestino Delgado
1 Necrosis Necrosis SL Miocarditis SL 2 Necrosis SL SL SL SL 3 SL Necrosis SL SL SL 4 Necrosis SL SL SL SL 5 Necrosis SL SL SL SL 6 SL SL Nefritis
intersticial SL SL
7 SL SL SL SL Hemorragia/trombosis Enteritis
8 Necrosis Necrosis SL Miocarditis SL 9 Necrosis Necrosis SL SL SL 10 SL SL SL SL SL 11 SL Necrosis Nefritis
intersticial SL SL
SL: Sin lesión.

72
2.3 Evaluación inmunohistoquímica y de la respuesta inmune
El resultado de la inmunohistoquímica realizada y la evaluación de las distintas células
inflamatorias se presentan en la Tabla 4 y se describen a continuación. Dentro de las lesiones
pulmonares, el 90% (10/11) de los potrillos presentó células CD3 positivas correspondientes a
linfocitos T (Figura 29) y el 45% (5/11) de los potrillos presentó células CD79 positivas
correspondientes a linfocitos B (Figura 30).
En el 100% de los potrillos se observaron macrófagos y neutrófilos.
Tabla 4: Porcentaje de las células inflamatorias observadas en las lesiones pulmonares de cada
potrillo.
Los grados se encuentran indicados entre paréntesis.
Potrillos
Células
inflamatorias
1
2
3
4
5
6
7
8
9
10
11
Porcentaje
total de
células
inflamatorias
Linfocitos T
10%
(0,5)
0%
16%
(0,8)
10%
(0,5)
10%
(0,5)
30%
(1,5)
20%
(1)
20%
(1)
10%
(0,5)
20%
(1)
16%
(0,8)
14,72%
Linfocitos B 0% 0% 4%
(0,2)
0% 0% 0% 0% 10%
(0,5)
10%
(0,5)
10%
(0,5)
4%
(0,2)
3,45%
Macrófagos 30%
(1,5)
60%
(3)
30%
(1,5)
60%
(3)
40%
(2)
60%
(3)
60%
(3)
70%
(3,5)
40%
(2)
50%
(2,5)
60%
(3)
50,90%
Neutrófilos 50%
(2,5)
40%
(2)
40%
(2)
20%
(1)
40%
(2)
10%
(0,5)
10%
(0,5)
0% 30%
(1,5)
20%
(1)
10%
(0,5)
24,54%
Eosinófilos y
mastocitos
0% 0% 0% 0% 0% 0% 0% 0% 0% 0% 0% 0
Fibroblastos 10%
(0,5)
0% 10%
(0,5)
10%
(0,5)
10%
(0,5)
0% 10%
(0,5)
0% 10%
(0,5)
0% 10%
(0,5)
6,36%
Total 100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%
(5)
100%

73
Figura 29: Linfocitos T inmunomarcados de forma positiva (flecha). El recuadro superior derecho muestra
el control negativo. IHC CD3. 400x.
Figura 30: Linfocitos B inmunomarcados de forma positiva (flecha). El recuadro superior derecho muestra
el control negativo. IHC CD79. 400x.
3. Discusión y conclusión
Las lesiones pulmonares macroscópicas y microscópicas reproducidas en este estudio
fueron compatibles con las lesiones pulmonares producidas por R. equi en infecciones naturales
descriptas en la bibliografía (Caswell y Williams, 2016; Lakritz et al., 1993; Zachary, 2012).
Con respecto a las lesiones pulmonares macroscópicas, el color blanco amarillento observado en
las mismas es atribuible a la infiltración con macrófagos y linfocitos del parénquima pulmonar, y
su textura firme a la formación de tejido conectivo fibroso (Zachary, 2012). Las lesiones

74
caudodorsales observadas, probablemente se hayan debido a la profundidad del lugar de la
inoculación, facilitando la evasión del barrido mucociliar y la llegada de R. equi a la región
caudodorsal de los pulmones.
Con respecto a las lesiones microscópicas pulmonares, aunque la hiperplasia de neumocitos tipo
II no se encuentra entre los hallazgos histopatológicos más importantes producidos por R. equi
descriptos en la bibliografía, en este estudio más de la mitad de los potrillos inoculados presentaron
en el pulmón el fenómeno de epitelización. Dicho fenómeno es producido cuando los neumocitos
tipo I sufren un daño celular irreversible dejando desnuda la membrana basal. Si luego de los tres
días de ocurrida la lesión la membrana basal permanece intacta, los neumocitos tipo II (que son
células precursoras y más resistentes a las lesiones) experimentan mitosis proveyendo gran
cantidad de células indiferenciadas que cubren las membranas basales desnudas, para finalmente
diferenciarse en neumocitos tipo I (Zachary, 2012). Especulamos que la inflamación inducida por
las bacterias de R. equi inoculadas y el aumento de sobrevida de los potrillos debido al tratamiento
antibiótico y de soporte suministrado, hayan sido factores importantes en el desarrollo del
fenómeno de epitelización.
También el daño celular de los neumocitos tipo I permite el movimiento de proteínas
plasmáticas y fibrinógeno hacia el lumen alveolar, y en algunos casos (como consideramos que
ocurrió en el potrillo 4) la filtración excesiva de proteínas se mezcla con restos celulares y
surfactante pulmonar formando una capa eosinofílica amorfa a lo largo de los septos
interalveolares. Este material fue interpretado como membranas hialinas (Zachary, 2012). La
observación de membranas hialinas no es una lesión característica producida por R. equi descripta
en la bibliografía internacional y puede ser producida por otras causas, como por ejemplo:
septicemia, síndrome de distrés respiratorio agudo (SDRA), endotoxemia, reacciones de
hipersensibilidad y falla cardíaca izquierda crónica (Lakritz et al., 1993; Zachary, 2012).
Aunque en la totalidad de los potrillos se observaron macrófagos alveolares con
cocobacilos basofílicos intracitoplasmáticos, solo 2 potrillos presentaron resultados positivos a la
tinción de Gram. Esto pudo deberse a la alta concentración de glucolípidos con ácido micólico que
contiene la pared celular de R. equi, los cuales afectan la tinción de Gram y protegen a esta bacteria
de las enzimas intracitoplasmáticas de los macrófagos (Ishino et al., 1992). Por esta razón, algunos
autores recomiendan realizar la técnica de Ziehl-Neelsen modificada en paralelo a la tinción de
Gram (Hermoso de Mendoza y Piriz, 2003).
Especulamos que la trombosis, confirmada mediante la tinción de PTAH en este estudio,
se haya debido a un aumento en la coagulabilidad sanguínea debido a la inflamación pulmonar, ya
sea por un daño directo de las células endoteliales, un aumento en la concentración de proteínas

75
hemostáticas activadas y/o una disminución de la degradación de estas proteínas por una inhibición
de las proteínas fibrinolíticas (Zachary, 2012).
La fibroplasia leve a moderada observada en 3 potrillos, y confirmada mediante la tinción
tricrómica de Masson, puede ser producida por una inflamación persistente y severa, la cual
estimula la proliferación de fibroblastos y miofibroblastos (Zachary, 2012). El proceso de
fibroplasia es asociado a la secreción de citoquinas (IL-1, IL-4, IL-10, IL-13), factores de
crecimiento (factor de crecimiento de fibroblastos y poliaminas) e inhibidores de la matriz de
metaloproteinasas por parte de los macrófagos, los cuales incrementan la síntesis de colágeno, la
proliferación de fibroblastos y disminuyen la actividad de la matriz de metaloproteinasas sobre los
fibroblastos, respectivamente (Zachary, 2012).
Las lesiones extrapulmonares macroscópicas observadas en este estudio fueron el hipopión
y el hipema, y se presentaron de forma consistente aproximadamente dos días antes de la muerte.
Esto coincide con lo observado por Reuss et al., (2009) quien sugiere que existe una fuerte
asociación entre la uveítis e hipopión y la posibilidad de muerte, debido posiblemente a que la
uveítis e hipopión aparezcan en respuesta a una infección sistémica más severa. A pesar de que en
nuestro estudio no se realizaron análisis histopatológicos del ojo para confirmar uveítis, la
presencia de hipopión e hipema es considerada una manifestación clínica de uveítis anterior
(Zachary, 2017). Los tejidos del globo ocular usualmente funcionan como una unidad integrada,
por lo que una lesión en una parte del globo ocular puede afectar la salud de otros tejidos del globo
ocular (Zachary, 2017). Particularmente el estroma del iris posee libre comunicación con el humor
acuoso, por lo tanto cualquier mediador químico de la inflamación secretado hacia el humor acuoso
produce la reacción inflamatoria de esta porción del tracto uveal (Zachary, 2017). En oposición a
lo mencionado en la bibliografía internacional consultada, en la que se consigna que
aproximadamente una tercera parte de los potrillos infectados desarrolla polisinovitis y un menor
porcentaje desarrolla artritis sépticas y osteomielitis, en las necropsias realizadas en este estudio
no se observaron lesiones articulares ni óseas (Giguère et al., 2011; Morresey et al., 2011; Reuss
et al., 2009).
La prevalencia de lesiones microscópicas extrapulmonares de este estudio fue del 81%
(9/11). Este resultado coincide aproximadamente con la prevalencia del 74% observada por Reuss
et al., (2009). A pesar de la diferencia con respecto a la cantidad de animales analizados, la
prevalencia de nuestro estudio fue más alta que la encontrada por Zink et al., (1986), quien en las
necropsias identificó lesiones extrapulmonares en el 54% de 131 potrillos; y de la observada por
Takai et al., (2000) con el 59% de prevalencia sobre 108 potrillos afectados. Especulamos que las
lesiones histopatológicas del hígado (principalmente la necrosis mediozonal) pueden ser

76
producidas por hipoxia debido al gran porcentaje del parénquima pulmonar afectado (> 50%). El
edema y proliferación de macrófagos en los linfonódulos puede ser atribuible al drenaje de las
áreas inflamadas del pulmón y como respuesta a la colonización bacteriana, respectivamente. En
muchas enfermedades la nefritis intersticial -como la observada en dos de los potrillos inoculados-
es considerada un hallazgo incidental, cuando solamente es visible microscópicamente y no está
asociada a una falla renal (Zachary, 2012).
Coincidiendo con lo reportado hasta el momento, sumado a que solamente un animal
presentó lesiones renales macroscópicas y dos animales lesiones renales microscópicas, creemos
que estos hallazgos pueden ser considerados como incidentales (Zachary, 2012). Los hallazgos
microscópicos hepáticos y renales difieren de los descriptos por Reuss et al., (2009),
puntualizando, principalmente, lesiones microscópicas de hepatitis y nefritis piogranulomatosas
con formación de microabscesos.
A pesar de que no se descartaron causas de origen viral, especulamos que la miocarditis
observada en este estudio pudo haber sido producida por una infección bacteriana diseminada por
vía hematógena hacia el miocardio (Zachary, 2012).
Contrariamente a lo reportado por Cohen (2014), quien observó que el signo clínico y la
lesión intestinal extrapulmonar más común fueron diarrea y enterotiflocolitis ulcerativa, en este
estudio se observó enteritis, hemorragia y trombosis intestinal. Consideramos que el color rojo
oscuro del contenido del colon pudo deberse a la sangre que provino del intestino delgado. La
hemorragia, la enteritis, y la trombosis intestinal observadas en el potrillo 7 son compatibles con
varias causas infecciosas, incluyendo Clostridium difficile, Clostridium perfringens tipo C
y Salmonella sp., las cuales no pudieron ser confirmadas. La infección por C. difficile es asociada,
de forma secundaria, con tratamientos antibióticos (Baverud, 2002; Giguère et al., 2003; Uzal et
al., 2012; Weese et al., 2001; Zink et al., 1986).
En coincidencia con la mayor parte de la bibliografía consultada, en este estudio no se
observaron lesiones extrapulmonares sin la presencia de lesiones pulmonares. En un estudio
realizado por Zink et al., (1986), sobre 131 casos se observó que un 4% presentó lesiones
intestinales sin bronconeumonía.
Los resultados de los análisis inmunohistoquímicos y morfológicos de las células
inflamatorias en el pulmón son compatibles con una respuesta inflamatoria Th-1, la cual
histológicamente está caracterizada principalmente por la presencia de macrófagos, una cantidad
variable de linfocitos T, células dendríticas y la ausencia de eosinófilos. Mediante técnicas de IHC,
nuestro estudio demostró una mayor proporción de linfocitos T, con respecto a los linfocitos B
(Tabla 4). La respuesta Th-1 está inducida por IL-12, 18, 23 y 27, liberadas por las células

77
dendríticas que producen el reclutamiento de linfocitos T colaboradores (CD4), los cuales liberan
IL-2 que induce la proliferación de linfocitos T; estos producen interferón gamma y factor de
necrosis tumoral β que desencadenan la activación de macrófagos (Zachary, 2012).
Este estudio demostró que la inoculación de una dosis baja de la cepa 103+ de R. equi es
capaz de producir lesiones pulmonares similares a las producidas por la infección natural de R. equi.
Se debería explorar el efecto de una dosis aun menor (por debajo de 5x103 UFC/mL), que quizás
podría producir lesiones severas pero con un intervalo de tiempo mayor entre la inoculación y la
enfermedad, asemejándose más a una infección real, lo que podría ser beneficioso para estudios
de vacunas y tratamientos contra esta enfermedad.
4. Bibliografía
1. Baverud, V. (2002). Clostridium difficile infections in animals with special reference to the
horse. A review. Vet Quart. 24, 203-219.
2. Caswell, J.L.; Williams, K.J. (2007). Pathology of domestic animals, ed. Saunders Elsevier,
5th ed., p. 630. Philadelphia, PA.
3. Caswell, J.L.; Williams, K.J. (2016). Pathology of domestic animals, ed. Elsevier, 6th ed., p.
514. St. Louis, MO.
4. Cohen, N.D. (2014). Rhodococcus equi foal pneumonia. Vet Clin North Am Equine Pract.
30, 609-622.
5. Darrah, P.A.; Holandus, M.K.; Chen, Q.; Ischiropoulus, H.; Mosser, D.M. (2000).
Cooperation between reactive oxygen and nitrogen intermediates in killing of Rhodococcus
equi by activated macrophages. Infect Immun. 68, 3587-3593.
6. Dawson, T.; Horohov, D.W.; Meijer, W.G.; Muscatello, G. (2010). Current understanding of
the equine immune response to Rhodococcus equi. An immunological review of R. equi
pneumonia. Vet Immunol and Immunophatol. 135, 1-11.
7. Demmers, S.; Johannisson, A.; Grondahl, G.; Jensen-Waern, M. (2001). Neutrophil functions
and serum IgG in growing foals. Equine Vet J. 33, 676-680.
8. Giguère, S. (2001). Rhodococcus equi pneumonia. Proceedings of the Annual Convention of
the AAEP. 47, 456-465.
9. Giguère, S.; Cohen, N.D.; Chaffin, K.M.; Hines, M.K.; Holandus, M.K; Prescott J.F.; Slovis
N.M. (2011).Rhodococcus equi: clinical manifestation, virulence, and immunity. J Vet Intern
Med.25, 1221-1230.

78
10. Giguère, S.; Hernandez, J.; Gaskin, J.; Miller, C.; Bowman, J.L. (2003). Evaluation of white
blood cell concentration, plasma fibrinogen concentration, and an agar gel immunodiffusion
test for early identification of foals with Rhodococcus equi pneumonia.
JAVMA. 222, 775-781.
11. Harrington, J.R.; Golding, M.C.; Martens, R.J.; Halbert, N.D.; Cohen, N.D. (2005).
Evaluation of a real-time quantitative polymerase chain reaction assay for detection and
quantitation of virulent Rhodococcus equi. AJVR 66, 755-761.
12. Hermoso de Mendoza, J.; Piriz, S. (2003). Manual de microbiología veterinaria, ed.
Interamericana, 1 ed., pp. 524-525. Madrid.
13. Ishino, S.; Kumagai, K.; Kuniyoshi, S.; Nakazawa, M.; Matzuda, I.; Oka, M. (1992).
Immunohistochemical observation on pneumonic lesions caused by Rhodococcus equi in
foals. J Vet Med Scien. 54, 509–515.
14. Lakritz, J.; Wilson, W.D.; Berry, C.R.; Schrenzel, M.D.; Carlson, G.P.; Madigan, J.E.
(1993). Bronchointerstitial pneumonia and respiratory distress in young horses: clinical,
clinicopathologic, radiographic, and pathological findings in 23 cases (1984-1989). J Vet
Intern Med. 7, 277-288.
15. Morresey, P.R.; Garrett, K.S.; Carter, D. (2011). Rhodococcus equi occipital bone
osteomyelitis, septic arthritis and meningitis in a neurological foal. Eq Vet Edu. 23, 398-402.
16. Muir, W.W. (2008). Eutanasia, pp. 579. En: Manual de anestesia veterinaria. Elsevier,
Madrid.
17. Muscatello, G. (2012). Rhodococcus equi pneumonia in the foal –part 2: diagnostics,
treatment and disease management. Vet J. 192, 27-33.
18. Muscatello, G.; Leadon, D.; Ocampo-Sosa, A.; Lewis, D.; Fogarty, U.; Buckley, T.;
Gilkerson, J.; Meijer, W.; Vazquez-Boland, J. (2007). Rhodococcus equi infection in foals:
the science of rattles. Eq Vet J. 39, 470-478.
19. Reuss, S.M.; Chaffin, M.K.; Cohen, N.D. (2009). Extrapulmonary disorders associated with
Rhodococcus equi infection in foals: 150 cases (1987-2007). JAVMA 235, 855-862.
20. Takai, S.; Higuchi, T.; Matsukura, S. (2000). Some epidemiological aspects of Rhodococcus
equi infection in foals in Japan: a review of 108 cases in 1992-1998. J Equine Sci. 11, 7-14.
21. Uzal, F.A.; Diab, S.S.; Blanchard, P.; Moore, J.; Anthenill, L.; Shahriar, F.; Garcia, J.P.;
Songer, J.G. (2012). Clostridium perfringens type C and Clostridium difficile co-infection in
foals. Vet. Microbiol. 156, 395-402.

79
22. VonBargen, K.; Haas, A. (2009). Molecular and infection biology of the horse pathogen
Rhodococcus equi. FEMS Microbiol Lett. 33, 870–891.
23. Weese, J.S.; Staempfli, H.R.; Prescott, J.F. (2001). A prospective study of the roles of
Clostridium difficile and enterotoxigenic Clostridium perfringens in equine diarrhea. Equine
Vet J. 33, 403-409.
24. Wilkins, P.A. (2009). Large animal internal medicine, ed. Mosby Elsevier, 4th ed., p. 512.
St. Louis, MO.
25. Zachary, J.F. (2012). Pathologic basis of veterinary disease, ed. Mosby Elsevier, 5th ed.,
p. 179. St. Louis, MO.
26. Zachary, J.F. (2017). Pathologic basis of veterinary disease, ed. Elsevier, 6th ed., p. 1278.
St. Louis, MO.
27. Zink, M.C.; Yager, J.A.; Smart, N.L. (1986). Corynebacterium equi infections in horses,
1958-1984: a review of 131 cases. Can J Vet Res. 27, 213-217.




















