
Diversity and distribution of Neotyphodium -infected grasses in Argentina
11
ORIGINAL ARTICLE Diversity and distribution of Neotyphodium-infected grasses in Argentina Leopoldo J. Iannone & James F. White Jr. & Liliana M. Giussani & Daniel Cabral & María Victoria Novas Received: 4 December 2009 / Revised: 10 March 2010 / Accepted: 19 March 2010 # German Mycological Society and Springer 2010 Abstract In the present work, we studied the presence of “epichloë/neotyphodium endophytes” in native grasses from Argentina. An extensive area of this country, representative of several different environments, was studied. Nine new host species of the genera Briza, Bromus and Poa were observed to be infected with asexual seed- borne endophytes. Epichloë stromata were not observed on any grass species. The incidence of infection in natural populations was highly variable among host species and among populations of the same host species. Morphological characterization revealed differences among the endophytes of different host species and among endophytes of different populations of the same host species. We also summarize the previous knowledge and present unpublished data on host diversity and distribution of these fungi in Argentina. Our results are discussed and compared with previous studies on endophytes in Argentina. This work supports the hypothesis that sexual species (Epichloë) are not present in the southern hemisphere, and suggests the existence of a high diversity of asexual endophytes in South America. Keywords Neotyphodium . Endophytes . Diversity . South America Introduction Many cool-season grasses are infected by epichloë/neotypho- dium endophytes (sexual species in the genus Epichloë Tul., Clavicipitaceae, Hypocreales; and their asexual relatives in the genus Neotyphodium Glenn, Bacon and Hanlin). These endophytes grow asymptomatically in the intercellular spaces of the aerial tissues of the host plants. Epichloë species produce perithecia in stromata that choke the inflorescence, causing total or partial sterility of the host plant. The ascospores produced in these stromata are responsible for the contagious dispersal to other plants (Chung and Schardl 1997). In contrast, Neotyphodium species colonize the developing flowers and are vertically transmitted through the seeds of infected mother plants to the daughter plants (Schardl 1996; Schardl et al. 1997). The symbiosis between Neotyphodium and their hosts has been considered a mutualism, since host plants may benefit from endophyte infection by increasing plant growth and providing resistance to herbivores, pathogens and abiotic stressors (Clay 1990; Malinowski and Belesky 2000; Novas et al. 2003; Schardl et al. 2004; Clarke et al. 2006; Iannone and Cabral 2006). Because of the beneficial effects on its host, endophytes have been used in grass breeding programs (Funk and White 1997). However, in L. J. Iannone : D. Cabral : M. Victoria Novas Lab. de Micología, Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires & PROPLAME-PRHIDEB-CONICET, Av. Intendente Güiraldes 2160. Pab. II, 4º piso, Cdad. Universitaria, C1428EHA Buenos Aires, Argentina L. J. Iannone (*) Departamento de Ingeniería Química, Facultad de Ingeniería, Universidad de Buenos Aires, Buenos Aires, Argentina e-mail: [email protected] J. F. White Jr. Department of Plant Biology & Pathology, Rutgers University, New Brunswick, New Jersey, USA L. M. Giussani Instituto de Botánica Darwinion—CONICET, San Isidro, Pcia., Buenos Aires, Argentina Mycol Progress DOI 10.1007/s11557-010-0669-2
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Diversity and distribution of Neotyphodium -infected grasses in Argentina

ORIGINAL ARTICLE
Diversity and distribution of Neotyphodium-infected grassesin Argentina
Leopoldo J. Iannone & James F. White Jr. &Liliana M. Giussani & Daniel Cabral &María Victoria Novas
Received: 4 December 2009 /Revised: 10 March 2010 /Accepted: 19 March 2010# German Mycological Society and Springer 2010
Abstract In the present work, we studied the presence of“epichloë/neotyphodium endophytes” in native grassesfrom Argentina. An extensive area of this country,representative of several different environments, wasstudied. Nine new host species of the genera Briza, Bromusand Poa were observed to be infected with asexual seed-borne endophytes. Epichloë stromata were not observed onany grass species. The incidence of infection in naturalpopulations was highly variable among host species andamong populations of the same host species. Morphologicalcharacterization revealed differences among the endophytesof different host species and among endophytes of differentpopulations of the same host species. We also summarizethe previous knowledge and present unpublished data onhost diversity and distribution of these fungi in Argentina.
Our results are discussed and compared with previousstudies on endophytes in Argentina. This work supports thehypothesis that sexual species (Epichloë) are not present inthe southern hemisphere, and suggests the existence of ahigh diversity of asexual endophytes in South America.
Keywords Neotyphodium . Endophytes . Diversity .
South America
Introduction
Many cool-season grasses are infected by epichloë/neotypho-dium endophytes (sexual species in the genus Epichloë Tul.,Clavicipitaceae, Hypocreales; and their asexual relatives inthe genus Neotyphodium Glenn, Bacon and Hanlin). Theseendophytes grow asymptomatically in the intercellularspaces of the aerial tissues of the host plants. Epichloëspecies produce perithecia in stromata that choke theinflorescence, causing total or partial sterility of the hostplant. The ascospores produced in these stromata areresponsible for the contagious dispersal to other plants(Chung and Schardl 1997). In contrast, Neotyphodiumspecies colonize the developing flowers and are verticallytransmitted through the seeds of infected mother plants to thedaughter plants (Schardl 1996; Schardl et al. 1997).
The symbiosis between Neotyphodium and their hostshas been considered a mutualism, since host plants maybenefit from endophyte infection by increasing plantgrowth and providing resistance to herbivores, pathogensand abiotic stressors (Clay 1990; Malinowski and Belesky2000; Novas et al. 2003; Schardl et al. 2004; Clarke et al.2006; Iannone and Cabral 2006). Because of the beneficialeffects on its host, endophytes have been used in grassbreeding programs (Funk and White 1997). However, in
L. J. Iannone :D. Cabral :M. Victoria NovasLab. de Micología, Departamento de Biodiversidad y BiologíaExperimental, Facultad de Ciencias Exactas y Naturales,Universidad de Buenos Aires& PROPLAME-PRHIDEB-CONICET,Av. Intendente Güiraldes 2160. Pab. II,4º piso, Cdad. Universitaria,C1428EHA Buenos Aires, Argentina
L. J. Iannone (*)Departamento de Ingeniería Química, Facultad de Ingeniería,Universidad de Buenos Aires,Buenos Aires, Argentinae-mail: [email protected]
J. F. White Jr.Department of Plant Biology & Pathology, Rutgers University,New Brunswick, New Jersey, USA
L. M. GiussaniInstituto de Botánica Darwinion—CONICET,San Isidro, Pcia.,Buenos Aires, Argentina
Mycol ProgressDOI 10.1007/s11557-010-0669-2

some cases, endophytes produce alkaloids that cause livestocktoxicosis in animals feeding on tall fescue (Schedonorusarundinaceus (Schreb.) Dumort.) and perennial ryegrass(Lolium perenne L) (Bacon 1995; Fletcher and Harvey1981). Because of this, one of the strategies used in order toobtain non-toxic improved grasses is the inoculation of non-toxic or ‘animal-friendly’ endophytes into selected cultivars(Funk and White 1997). Endophyte-grass symbiota holdgreat promise for their use in agriculture, turf and remedi-ation projects (Kuldau and Bacon 2008), and knowledge ofthe diversity of endophytes is needed for their use inmanagement programs.
Although the abundance and diversity of epichloë/neo-typhodium endophytes has been widely investigated inNorth America and Europe (White and Cole 1985, 1986;White 1987; Clay and Leuchtmann 1989; White andBaldwin 1992; Leuchtmann 1992; Saikkonen et al. 2000;Leyronas and Raynal 2001; Bazely et al. 2007), few studieshave been carried out in Argentina and, to our knowledge,there is no information about its presence elsewhere inSouth America.
Argentina extends from latitude 21°50’S to the SouthPole with a surface of approximately 3,000,000 km2. Thediversity in vertical and horizontal biogeographical regionsis of extraordinary magnitude, including subtropical rainforests, central temperate, fertile mollisol soils in the HumidPampas, extensive western arid and semi-arid regionsbordering the Andes, and cold subantartic zones at thesouth of the country. About two-thirds of continentalArgentina are associated with arid and semi-arid rangelandecosystems (Fernandez and Busso 1997).
The presence of fungal endophytes in native grassesfrom Argentina has been known from the beginning of thetwentieth century, when they were associated with thelivestock toxicity of Festuca hieronymi Hack. (Rivas andZanolli 1909). Several surveys on both field and herbariummaterial have reported endophyte infections in 25 nativegrass species (Bertoni et al. 1993; Lugo et al. 1998). Theendophytes of 10 host species have been isolated andcharacterized by means of morphological and phylogeneticstudies (Cabral et al. 1999; Gentile et al. 2005; Iannone etal. 2009). Based on morphological and phylogeneticapproaches, 2 asexual endophyte species have beendescribed for Argentina: Neotyphodium tembladerae Cabralet White, with a wide range of hosts in the genera Poa,Festuca, Bromus, Phleum and Melica (Cabral et al. 1999;Gentile et al. 2005), and N. pampeanum Iannone et Cabral,associated with some populations of Bromus auleticus Trin.ex Nees (Iannone et al. 2009).
In agreement with what has been observed in other partsof the world (Schulthess and Faeth 1998; Saikkonen et al.2000; Wei et al. 2006; Bazely et al. 2007), the incidence ofendophytes varies among grass populations. In the central
Andes, the incidence of endophytes in populations ofBromus setifolius Presl. correlates with the abundance ofleaf-cutting ants (White et al. 2001), whereas that inpopulations of Phleum alpinum L., Bromus setifolius andPoa spiciformis (Steud.) Hauman and Parodi from southPatagonia correlates with environmental factors and theplant community (Novas et al. 2007).
Although our knowledge on endophytes in nativegrasses from Argentina has increased in recent years(Cabral et al. 1999; White et al. 2001; Gentile et al. 2005;Novas et al. 2007, 2009; Iannone et al. 2009), theirmorphological diversity, distribution and incidence hasbeen poorly studied in most of the hosts.
In this work, we isolated the Neotyphodium endophytes ofnew hosts from Argentina, and we also studied thegeographical distribution of the populations with endophytesand the incidence of endophytes in populations of the newhosts. In addition, we summarized previous research on thehost ranges and distribution of endophytes in this region ofSouth America.
Materials and methods
Plant sampling and surveyed area
Plant samples
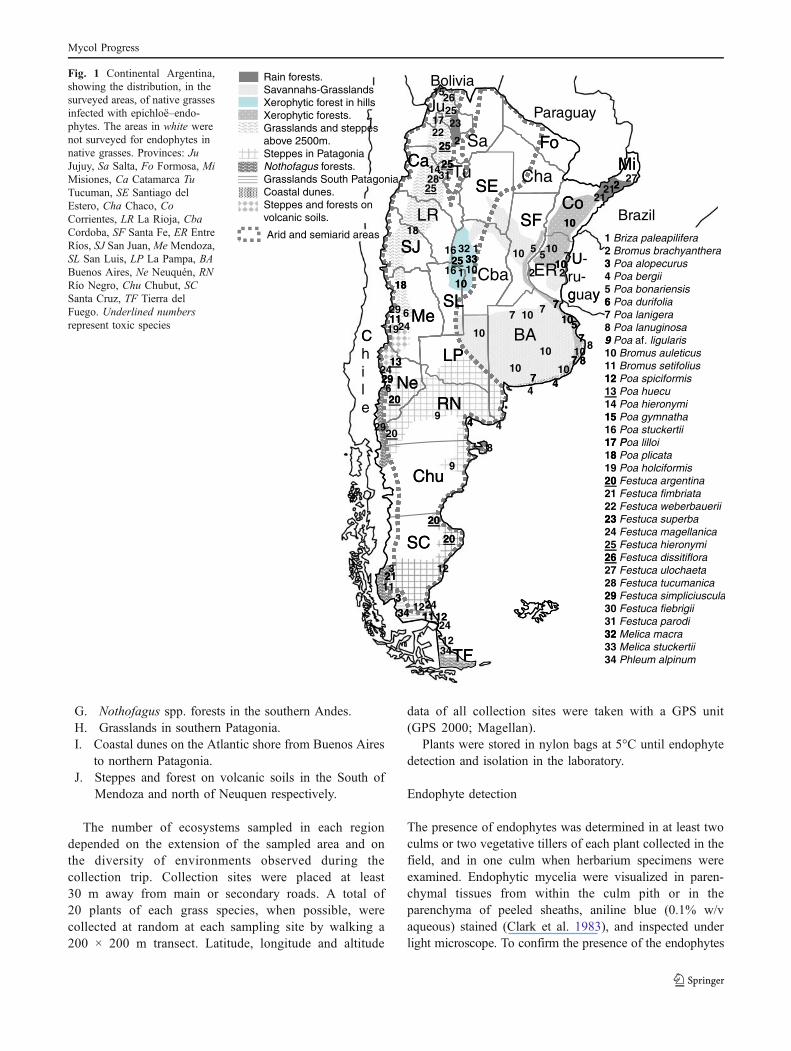
During spring and summer (of the years) 2004–2007 and2009, collection trips were made for endophyte detection inseveral regions not previously screened in Argentina. Thesurveyed area included large areas of continental Argentina(Fig. 1) in different phytogeographical regions (Cabrera1976).
A total of 205 collection sites were established in orderto sample different environments in each region. Theenvironments were characterized as follows:
A. Subtropical rain forests in Misiones province, andYungas forests in mountains of Salta province,and gallery forests in the margins of UruguayRiver and delta of Parana river.
B. Savannahs and grasslands in Humid Pampas andMesopotamia.
C. Xerophytic forests and grasslands between 500 and1,800 m a.s.l. in hills of Córdoba and San Luisprovinces.
D. Prosopis spp. and Celtis spp. forests in the meso-potamic region (Corrientes and Entre Ríos provinces)and shelly ribs along the coast of Rio de la Plata.
E. Highland grasslands and steppes of the Andean regionbetween 2,500 and 4,900 m a.s.l.
F. Shrubby and grassy steppes in Patagonia.
Mycol Progress
G. Nothofagus spp. forests in the southern Andes.H. Grasslands in southern Patagonia.I. Coastal dunes on the Atlantic shore from Buenos Aires
to northern Patagonia.J. Steppes and forest on volcanic soils in the South of
Mendoza and north of Neuquen respectively.
The number of ecosystems sampled in each regiondepended on the extension of the sampled area and onthe diversity of environments observed during thecollection trip. Collection sites were placed at least30 m away from main or secondary roads. A total of20 plants of each grass species, when possible, werecollected at random at each sampling site by walking a200 ! 200 m transect. Latitude, longitude and altitude
data of all collection sites were taken with a GPS unit(GPS 2000; Magellan).
Plants were stored in nylon bags at 5°C until endophytedetection and isolation in the laboratory.
Endophyte detection
The presence of endophytes was determined in at least twoculms or two vegetative tillers of each plant collected in thefield, and in one culm when herbarium specimens wereexamined. Endophytic mycelia were visualized in paren-chymal tissues from within the culm pith or in theparenchyma of peeled sheaths, aniline blue (0.1% w/vaqueous) stained (Clark et al. 1983), and inspected underlight microscope. To confirm the presence of the endophytes
Rain forests. Bolivia1526
Paraguay1725Ju
Xerophytic forests.
225
23
Fo22Grasslands and steppes
above 2500m. 25
MiFoSa
Ca 25Steppes in Patagonia
2 271428
MiChaSE
TuCa
3125Nothofagus forests.
Grasslands South Patagonia 221
25
CoSE 21Coastal dunes.
10 BrazilSFCo
LRSteppes and forests on volcanic soils. 10SF
SJ18
1 Briza paleapiliferaArid and semiarid areas
55710 1032 1
33SJ
U-16
25 102 Bromus brachyanthera3
18101633
2CbaUru-
25
101 ER10
23 Poa alopecurus4 Poa bergii
18
SL guay10
75 Poa bonariensis6
57 7
101061129 Me
SL g y7 6 Poa durifolia
7 Poa lanigera510
10C 1911
7BAMe
24 8 Poa lanuginosa9
10Ch 13
107
87 8LP
9 Poa af. ligularis10 Bromus auleticus
10i 2413
29
7 810
7 4Ne
LP11 Bromus setifolius12
4l20
629 7 4
RNNe 12 Poa spiciformis
13 Poa huecu
49
e20 RN 14 Poa hieronymi
1544
202915 Poa gymnatha16 Poa stuckertii17 P8 17 Poa lilloi18 Poa plicata
9Chu
819 Poa holciformis20Chu 20 Festuca argentina21 Festuca fimbriata
2022 Festuca weberbauerii2320
20SC23 Festuca superba24 Festuca magellanica
20SC 25 Festuca hieronymi26
12321
26 Festuca dissitiflora27 Festuca ulochaeta
1121
328 Festuca tucumanica29
121224
11343 29 Festuca simpliciuscula
30 Festuca fiebrigii12113424 31 Festuca parodi
32
3412
TF32 Melica macra33 Melica stuckertii
TF 34 Phleum alpinum
Savannahs-GrasslandsXerophytic forest in hills
Fig. 1 Continental Argentina,showing the distribution, in thesurveyed areas, of native grassesinfected with epichloë–endo-phytes. The areas in white werenot surveyed for endophytes innative grasses. Provinces: JuJujuy, Sa Salta, Fo Formosa, MiMisiones, Ca Catamarca TuTucuman, SE Santiago delEstero, Cha Chaco, CoCorrientes, LR La Rioja, CbaCordoba, SF Santa Fe, ER EntreRíos, SJ San Juan, Me Mendoza,SL San Luis, LP La Pampa, BABuenos Aires, Ne Neuquén, RNRío Negro, Chu Chubut, SCSanta Cruz, TF Tierra delFuego. Underlined numbersrepresent toxic species
Mycol Progress

in seeds, caryopses were soaked for 8 h in a 5% v/vaqueous solution of sodium hydroxide at room temper-ature (20–22°C), and then rinsed and stained with anilineblue as mentioned.
A plant or seed was considered as endophyte-infected ifeither typical unbranched intercellular mycelia weredetected in parenchymal tissues or when twisted hyphaewere observed to be associated with the layer of aleuronecells in seed squash preparations.
Endophyte isolation
Endophyte-infected plants were stored in a refrigerator innylon bags until endophyte isolation. Depending on thenumber of infected plants of each new host speciescollected in each site, three to five plants of each host werechosen for endophyte isolation. Caryopses, and approxi-mately 5-mm-long pieces of leaf sheaths or culm segmentsof endophyte-infected plants, were surface-sterilized bywashing 1 min in 70% v/v ethanol, followed by a 15-minwash in 2.5–3% sodium hypochlorite (50% v/v commercialbleach), and then a 1-min wash in 50% ethanol. Aftersterilization, caryopses and leaf sheaths were placed ontopotato dextrose agar (PDA) plates, incubated in the darkat 23°C, and checked regularly for fungal growth for upto 2 months. Acremonium spp. and other fungal endo-phytes were occasionally isolated, but were not consideredin this study. The isolated endophytes were identified asNeotyphodium by morphological characterization of thecolonies, conidia and conidiogenous cells. In many cases,several morphologically homogenous Neotyphodium-likecolonies were observed growing from tissue pieces of thesame plant, but only one isolate per plant was consideredin this study.
Endophyte distribution and incidence
The incidence of endophytes per population of the newhosts was established as infected plants over examinedplants (White et al. 2001; Novas et al. 2007). In terms ofendophyte incidence, each grass species was considered asa local population in each collection site. Because stainingculms or sheaths from infected plants could have missedhyphae, our estimation of endophyte frequency should beconsidered to be underestimated.
Endophyte characterization
To examine the macroscopic characteristics of endophytecultures, small (2 ! 2 mm) agar blocks were cut from themargins of actively growing colonies and transferred toPDA plates to make two replicate cultures of each isolate.Cultures were incubated for 30 days at 23°C in the dark,
and then characteristics of the colony such as color, marginsand mycelium aspect were recorded. The average diameterof each colony was established by measuring two perpen-dicular diameters (±1 mm) on each replicate and thesevalues were used to calculate the average growth rate (mm/day). For microscopic characterization, mycelium fromPDA-grown cultures was directly mounted on glass slidesand stained with either lactophenol blue or calcofluor whiteM2R. In addition, pieces of actively growing myceliumwere ground in sterile water and 100-!l aliquots werespread on 2% water agar (WA) plates. These cultures wereallowed to grow for 20–30 days at 23°C in the dark aspreviously described (Moon et al. 2002, 2007; Gentile et al.2005). Blocks of agar (1 cm2) were cut and mounted onslides without cover slips, in order to keep the arrangementand disposition of fungal structures unaltered, and observedand photographed at magnifications up to !400. Thereafter,material was stained with either lactophenol or calcofluorwhite M2R. At least 20 conidia and conidiogenous cells perisolate were measured and photographed at !1,000–1,600magnifications, using a Karl Zeiss Axioskope microscope.
The isolates were grouped in morphotypes according tothe combination of characteristics they exhibit. Isolates thatshare common characteristics were ascribed to in onemorphotype, and those isolates that differ in one or moreof the characteristics were ascribed to other morphotypes.
Results
More than 4,500 plants of approximately 80 species of coolseason grasses were examined in a total of 205 collectionsites in an approximately 1.5 million km2 area that includesrepresentative environments from Argentina. Nine new hostspecies were detected in 42 of the collection sites (Table 1).A total of 34 epichloë/neotyphodium endophyte-infectedgrass species (Table 1) (including previously reportedhosts) were observed in 80 of the 205 collection sitesstudied between 2004 and 2009. No plants with chokesymptoms (Epichloë state) were observed at any of thecollection sites.
New hosts
Characteristic epichloë/neotyphodium endophyte-like hyphae,were observed in culm pith, leaf sheaths and cariopses of:Briza paleapilifera Parodi, Bromus brachyanthera Döll, Poaalopecurus Kunth. Poa bergii Hieron., P. bonariensis (Lam.)Kunth., P. durifolia Giussani, Nicora and Roig, P. lanigeraNees, P. lanuginosa Poir. and P. aff. ligularis Nees (Table 1).Festuca fimbriata Nees, previously reported as endophyte-infected by herbarium material examination (Bertoni et al.1993), was collected for the first time for endophyte isolation.
Mycol Progress
Incidence of endophytes and distribution of infectedpopulations
The incidence of endophytes in field populations of the hostspecies was highly variable among host species and amongpopulations within one species (Table 1). In populations ofBriza paleapilifera, endemic in hills of central Argentina,Bromus brachyanthera, inhabiting shaded tropical forests,Festuca fimbriata, enedemic in marshes in the northeastern
Corrientes province, Poa bergii, endemic in coastal dunesand Poa durifolia growing on volcanic soils, the incidenceof Neotyphodium endophytes was in general higher than80% for all the populations studied. In other species withwider distribution areas, such as Poa lanigera, Poalanuginosa, Poa bonariensis and P. ligularis that inhabitdifferent environments, including coastal dunes, shellyridges, grasslands, xerophytic forests or steppes, theincidence ranged from 0 to 100%. The geographical
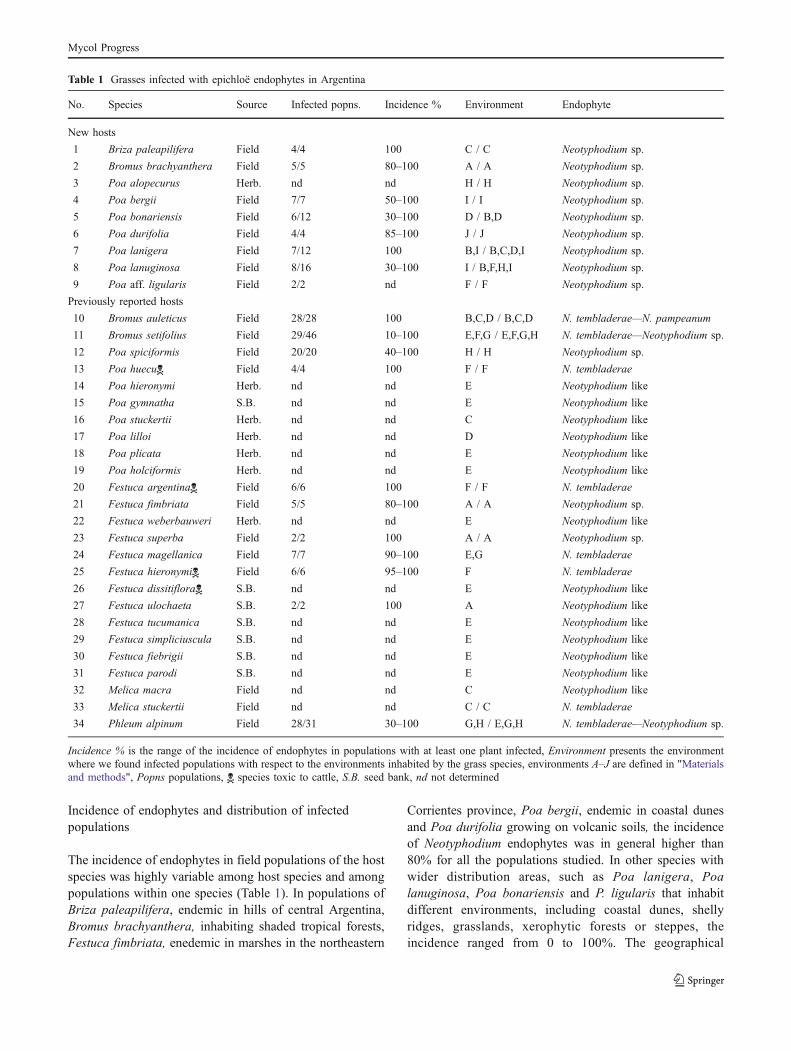
Table 1 Grasses infected with epichloë endophytes in Argentina
No. Species Source Infected popns. Incidence % Environment Endophyte
New hosts
1 Briza paleapilifera Field 4/4 100 C / C Neotyphodium sp.
2 Bromus brachyanthera Field 5/5 80–100 A / A Neotyphodium sp.
3 Poa alopecurus Herb. nd nd H / H Neotyphodium sp.
4 Poa bergii Field 7/7 50–100 I / I Neotyphodium sp.
5 Poa bonariensis Field 6/12 30–100 D / B,D Neotyphodium sp.
6 Poa durifolia Field 4/4 85–100 J / J Neotyphodium sp.
7 Poa lanigera Field 7/12 100 B,I / B,C,D,I Neotyphodium sp.
8 Poa lanuginosa Field 8/16 30–100 I / B,F,H,I Neotyphodium sp.
9 Poa aff. ligularis Field 2/2 nd F / F Neotyphodium sp.
Previously reported hosts
10 Bromus auleticus Field 28/28 100 B,C,D / B,C,D N. tembladerae—N. pampeanum
11 Bromus setifolius Field 29/46 10–100 E,F,G / E,F,G,H N. tembladerae—Neotyphodium sp.
12 Poa spiciformis Field 20/20 40–100 H / H Neotyphodium sp.
13 Poa huecu Field 4/4 100 F / F N. tembladerae
14 Poa hieronymi Herb. nd nd E Neotyphodium like
15 Poa gymnatha S.B. nd nd E Neotyphodium like
16 Poa stuckertii Herb. nd nd C Neotyphodium like
17 Poa lilloi Herb. nd nd D Neotyphodium like
18 Poa plicata Herb. nd nd E Neotyphodium like
19 Poa holciformis Herb. nd nd E Neotyphodium like
20 Festuca argentina Field 6/6 100 F / F N. tembladerae
21 Festuca fimbriata Field 5/5 80–100 A / A Neotyphodium sp.
22 Festuca weberbauweri Herb. nd nd E Neotyphodium like
23 Festuca superba Field 2/2 100 A / A Neotyphodium sp.
24 Festuca magellanica Field 7/7 90–100 E,G N. tembladerae
25 Festuca hieronymi Field 6/6 95–100 F N. tembladerae
26 Festuca dissitiflora S.B. nd nd E Neotyphodium like
27 Festuca ulochaeta S.B. 2/2 100 A Neotyphodium like
28 Festuca tucumanica S.B. nd nd E Neotyphodium like
29 Festuca simpliciuscula S.B. nd nd E Neotyphodium like
30 Festuca fiebrigii S.B. nd nd E Neotyphodium like
31 Festuca parodi S.B. nd nd E Neotyphodium like
32 Melica macra Field nd nd C Neotyphodium like
33 Melica stuckertii Field nd nd C / C N. tembladerae
34 Phleum alpinum Field 28/31 30–100 G,H / E,G,H N. tembladerae—Neotyphodium sp.
Incidence % is the range of the incidence of endophytes in populations with at least one plant infected, Environment presents the environmentwhere we found infected populations with respect to the environments inhabited by the grass species, environments A–J are defined in "Materialsand methods", Popns populations, species toxic to cattle, S.B. seed bank, nd not determined
Mycol Progress

distribution and the environments where the infectedpopulations of these new hosts were found are shown inFig. 1 and Table 1.
Previously described hosts
The 25 species of grasses, previously reported as endophyte-infected by Bertoni et al. (1993), Lugo et al. (1998) andWhite et al. (2001), are listed in Table 1, and theirdistributions are shown in Fig. 1. In the surveys conductedbetween 2004 and 2009, new populations of some previouslyreported hosts, including the species previously reported astoxic for cattle, Festuca hieronymi in the mountains of Jujuyand the hills of Cordoba provinces, as well as twopopulations of F. argentina in Santa Cruz, were detected.The incidence of endophytes in these populations was 100%.New populations of Phleum alpinum were detected in SantaCruz province and the incidence of endophytes rangedbetween 0 and 80%. This species inhabits stream banks inthe central Andes between 2,800 and 4,500 m.a.s.l., and iscommon in humid grasslands and Southern Beech (Notho-fagus spp.) forests in southern Patagonia and in Tierra delFuego. In this species, endophytes had previously beendetected only in Tierra del Fuego.
Non-host species
Among the species examined in the present work that werenot associated with epichloë/neotyphodium endophytes, wecan highlight the species toxic to cattle, Stipa leptostachyaGriseb. (Parodi) (Parodi 1950), and the non-toxic speciesBriza erecta Lam., Briza subaristata Lam., Bromus brevisNees, Bromus japonicus Thumb., Bromus madritensis L.,Bromus pellitus Hack., Deschampsia anctartica E.Desv.,Elymus scabrifolius J.H.Hunz., Festuca gracillima Hook, F.pallescens Parodi, F. scirpifolia Kunth, Glyceria multifloraSteudel., Melica argyrea Hack., M. macra Nees, M.sarmentosa Nees, Poa calchaquiensis Hack, P. dollicophylaHack, P. iridifolia Hauman, and several species of thegenera Agrostis L., Hordeum L., Piptochaetium J. Presl.,Stipa L., Trisetum Pers. and Vulpia C.C. Gmel..
Phleum pratense L. and Dactylis glomerata L., twocommonly occurring naturalized species, have been foundto be infected by Epichloë typhina in the northernhemisphere (Sampson 1933). However, the plants analyzedin this work were consistently endophyte-free.
Characterization of the endophytes of the new hosts
Epichloë/neotyphodium endophytes were isolated from 117out of 195 plants of the new host species.
Considerable morphological variability was observedbetween endophytes isolated from different host species
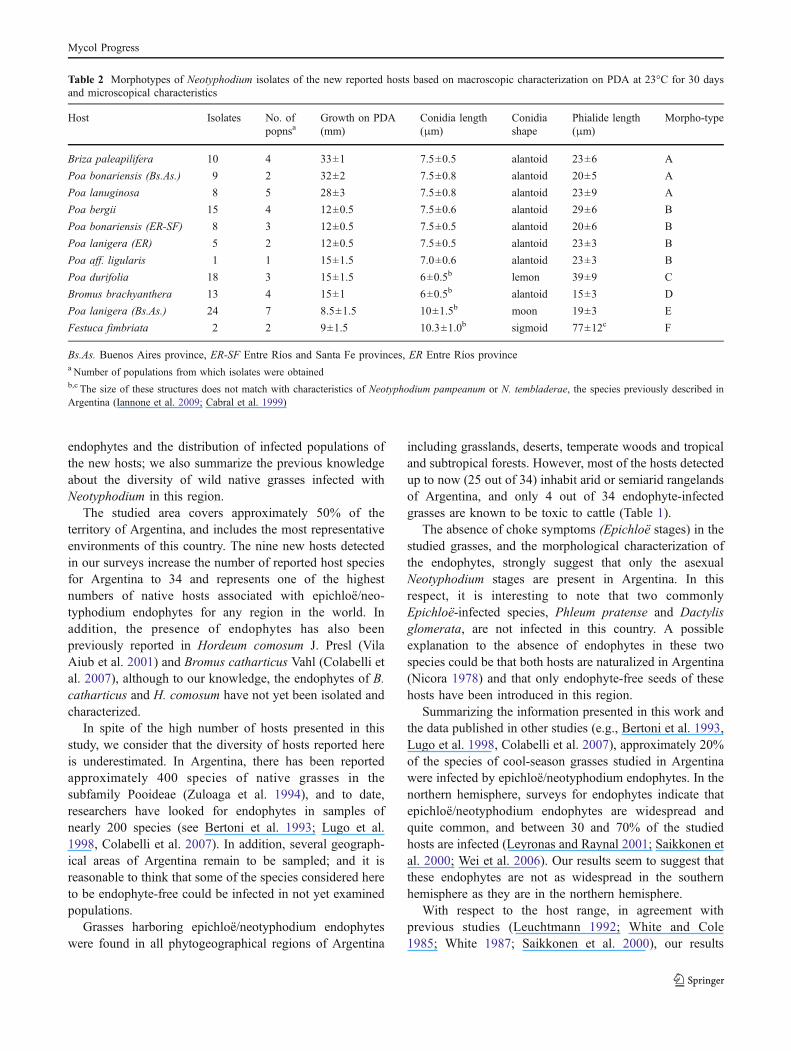
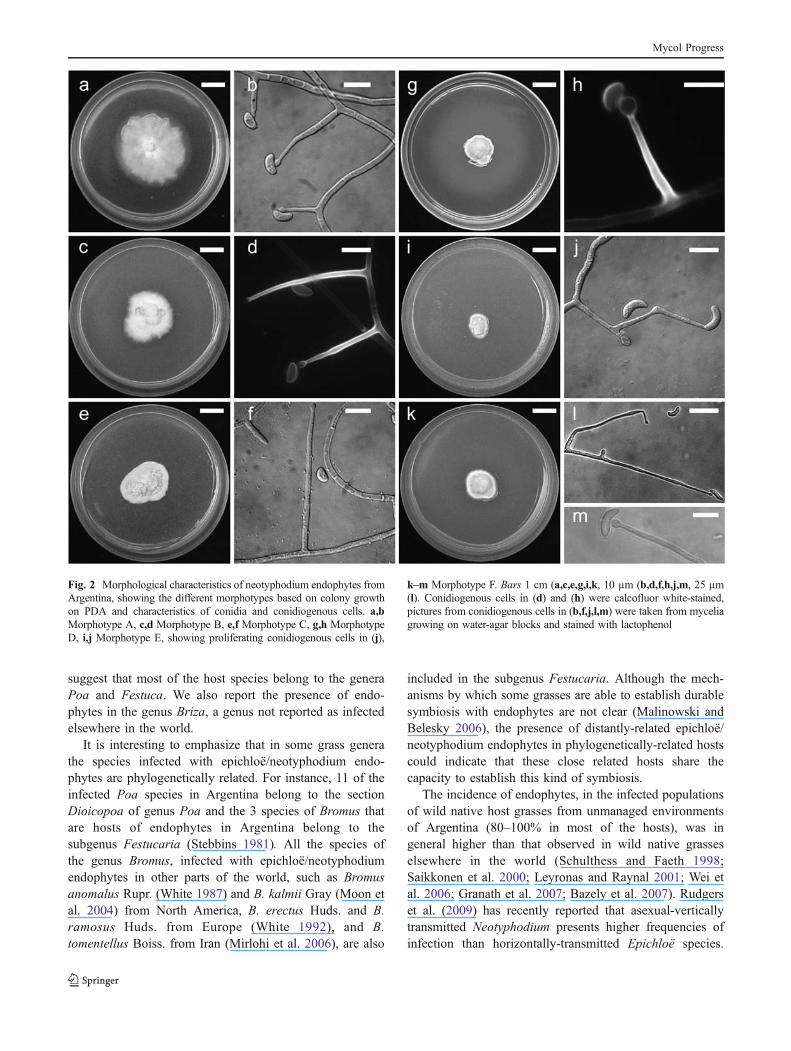
and from different populations of a same host species (Table 2;Fig. 2). Colonies on PDA were white, felted to velvety, andthe rate of growth on PDA ranged from 0.2–1.1 mm/day.Conidiogenous cells were solitary or sympodially proliferat-ing, sometimes branched (Fig. 2j), smooth, 8–100 !m longfrom base to tip, tapering gently from 1.5–2.5 !m at the baseto 0.5–1.0 !m at the tip. A basal septum was rarely present.Conidial ontogeny was generally enteroblastic and two(rarely three) conidia were produced by each conidiogenouscell (Fig. 2b; Table 2). Conidia were variable in shape, butmostly alantoid (reniform). However, moon-shaped conidiawere observed in isolates of Poa lanigera, sigmoid conidiawere observed in isolates of Festuca fimbriata, and lemon-shaped conidia were observed in P. durifolia isolates.Conidia measures ranged from 4.5 to 14 !m long and from2 to 4 !m wide (Fig. 2 b, d, f, h, j, l, m; Table 2). Based onthese morphological characteristics, the absence ofEpichloë stromata on hosts and our previous knowledgeon epichloë/neotyphodium endophytes from Argentina,we consider that the isolates obtained belong to the genusNeotyphodium.
The combination of these macro- and microscopiccharacters allowed us to differentiate the endophytesinto six morphotypes (Morphotypes A–F; Table 2).Morphotype A (Fig. 2a, b; Table 2) included fast-growing endophytes isolated from Briza paleapilifera,Poa lanuginosa and plants of P. bonariensis frompopulations in Buenos Aires province. Morphotype B(Fig. 2c, d; Table 2) included moderately slow-growingendophytes of P. bergii, P. ligularis and isolates of P.bonariensis in populations from Entre Rios and Santa Feprovinces and isolates of P. lanigera collected in pop-ulations from Entre Rios province. The endophytes of P.durifolia with small conidia and very long conidiogenouscells were included in Morphotype C (Fig. 2e, f; Table 2).The isolates of B. brachyanthera exhibited small conidia(4.5–7 !m in length) and small conidiogenous cells (12–18 !m long) and were thus grouped in Morphotype D(Fig. 2g, h; Table 2). Morphotype E (Fig. 2i, j; Table 2)included only slow-growing isolates of P. lanigera frompopulations from Buenos Aires province, which presentlarge conidia (8.5–13 !m long). The two isolates from F.fimbriata presented extremely long (60–90 !m) conidiog-enous cells and large conidia (9–13 !m in length) andwhere included in Morphotype F (Fig. 2k–m).
Discussion
In this article is reported for the first time the presence andcharacteristics of Neotyphodium in nine new host grassesdistributed in a wide range of environments of Argentina.We present preliminary analyses of the incidence of
Mycol Progress
endophytes and the distribution of infected populations ofthe new hosts; we also summarize the previous knowledgeabout the diversity of wild native grasses infected withNeotyphodium in this region.
The studied area covers approximately 50% of theterritory of Argentina, and includes the most representativeenvironments of this country. The nine new hosts detectedin our surveys increase the number of reported host speciesfor Argentina to 34 and represents one of the highestnumbers of native hosts associated with epichloë/neo-typhodium endophytes for any region in the world. Inaddition, the presence of endophytes has also beenpreviously reported in Hordeum comosum J. Presl (VilaAiub et al. 2001) and Bromus catharticus Vahl (Colabelli etal. 2007), although to our knowledge, the endophytes of B.catharticus and H. comosum have not yet been isolated andcharacterized.
In spite of the high number of hosts presented in thisstudy, we consider that the diversity of hosts reported hereis underestimated. In Argentina, there has been reportedapproximately 400 species of native grasses in thesubfamily Pooideae (Zuloaga et al. 1994), and to date,researchers have looked for endophytes in samples ofnearly 200 species (see Bertoni et al. 1993; Lugo et al.1998, Colabelli et al. 2007). In addition, several geograph-ical areas of Argentina remain to be sampled; and it isreasonable to think that some of the species considered hereto be endophyte-free could be infected in not yet examinedpopulations.
Grasses harboring epichloë/neotyphodium endophyteswere found in all phytogeographical regions of Argentina
including grasslands, deserts, temperate woods and tropicaland subtropical forests. However, most of the hosts detectedup to now (25 out of 34) inhabit arid or semiarid rangelandsof Argentina, and only 4 out of 34 endophyte-infectedgrasses are known to be toxic to cattle (Table 1).
The absence of choke symptoms (Epichloë stages) in thestudied grasses, and the morphological characterization ofthe endophytes, strongly suggest that only the asexualNeotyphodium stages are present in Argentina. In thisrespect, it is interesting to note that two commonlyEpichloë-infected species, Phleum pratense and Dactylisglomerata, are not infected in this country. A possibleexplanation to the absence of endophytes in these twospecies could be that both hosts are naturalized in Argentina(Nicora 1978) and that only endophyte-free seeds of thesehosts have been introduced in this region.
Summarizing the information presented in this work andthe data published in other studies (e.g., Bertoni et al. 1993,Lugo et al. 1998, Colabelli et al. 2007), approximately 20%of the species of cool-season grasses studied in Argentinawere infected by epichloë/neotyphodium endophytes. In thenorthern hemisphere, surveys for endophytes indicate thatepichloë/neotyphodium endophytes are widespread andquite common, and between 30 and 70% of the studiedhosts are infected (Leyronas and Raynal 2001; Saikkonen etal. 2000; Wei et al. 2006). Our results seem to suggest thatthese endophytes are not as widespread in the southernhemisphere as they are in the northern hemisphere.
With respect to the host range, in agreement withprevious studies (Leuchtmann 1992; White and Cole1985; White 1987; Saikkonen et al. 2000), our results
Table 2 Morphotypes of Neotyphodium isolates of the new reported hosts based on macroscopic characterization on PDA at 23°C for 30 daysand microscopical characteristics
Host Isolates No. ofpopnsa
Growth on PDA(mm)
Conidia length(!m)
Conidiashape
Phialide length(!m)
Morpho-type
Briza paleapilifera 10 4 33±1 7.5±0.5 alantoid 23±6 A
Poa bonariensis (Bs.As.) 9 2 32±2 7.5±0.8 alantoid 20±5 A
Poa lanuginosa 8 5 28±3 7.5±0.8 alantoid 23±9 A
Poa bergii 15 4 12±0.5 7.5±0.6 alantoid 29±6 B
Poa bonariensis (ER-SF) 8 3 12±0.5 7.5±0.5 alantoid 20±6 B
Poa lanigera (ER) 5 2 12±0.5 7.5±0.5 alantoid 23±3 B
Poa aff. ligularis 1 1 15±1.5 7.0±0.6 alantoid 23±3 B
Poa durifolia 18 3 15±1.5 6±0.5b lemon 39±9 C
Bromus brachyanthera 13 4 15±1 6±0.5b alantoid 15±3 D
Poa lanigera (Bs.As.) 24 7 8.5±1.5 10±1.5b moon 19±3 E
Festuca fimbriata 2 2 9±1.5 10.3±1.0b sigmoid 77±12c F
Bs.As. Buenos Aires province, ER-SF Entre Ríos and Santa Fe provinces, ER Entre Ríos provincea Number of populations from which isolates were obtainedb,c The size of these structures does not match with characteristics of Neotyphodium pampeanum or N. tembladerae, the species previously described inArgentina (Iannone et al. 2009; Cabral et al. 1999)
Mycol Progress
suggest that most of the host species belong to the generaPoa and Festuca. We also report the presence of endo-phytes in the genus Briza, a genus not reported as infectedelsewhere in the world.
It is interesting to emphasize that in some grass generathe species infected with epichloë/neotyphodium endo-phytes are phylogenetically related. For instance, 11 of theinfected Poa species in Argentina belong to the sectionDioicopoa of genus Poa and the 3 species of Bromus thatare hosts of endophytes in Argentina belong to thesubgenus Festucaria (Stebbins 1981). All the species ofthe genus Bromus, infected with epichloë/neotyphodiumendophytes in other parts of the world, such as Bromusanomalus Rupr. (White 1987) and B. kalmii Gray (Moon etal. 2004) from North America, B. erectus Huds. and B.ramosus Huds. from Europe (White 1992), and B.tomentellus Boiss. from Iran (Mirlohi et al. 2006), are also
included in the subgenus Festucaria. Although the mech-anisms by which some grasses are able to establish durablesymbiosis with endophytes are not clear (Malinowski andBelesky 2006), the presence of distantly-related epichloë/neotyphodium endophytes in phylogenetically-related hostscould indicate that these close related hosts share thecapacity to establish this kind of symbiosis.
The incidence of endophytes, in the infected populationsof wild native host grasses from unmanaged environmentsof Argentina (80–100% in most of the hosts), was ingeneral higher than that observed in wild native grasseselsewhere in the world (Schulthess and Faeth 1998;Saikkonen et al. 2000; Leyronas and Raynal 2001; Wei etal. 2006; Granath et al. 2007; Bazely et al. 2007). Rudgerset al. (2009) has recently reported that asexual-verticallytransmitted Neotyphodium presents higher frequencies ofinfection than horizontally-transmitted Epichloë species.
Fig. 2 Morphological characteristics of neotyphodium endophytes fromArgentina, showing the different morphotypes based on colony growthon PDA and characteristics of conidia and conidiogenous cells. a,bMorphotype A, c,d Morphotype B, e,f Morphotype C, g,h MorphotypeD, i,j Morphotype E, showing proliferating conidiogenous cells in (j),
k–m Morphotype F. Bars 1 cm (a,c,e,g,i,k, 10 µm (b,d,f,h,j,m, 25 µm(l). Conidiogenous cells in (d) and (h) were calcofluor white-stained,pictures from conidiogenous cells in (b,f,j,l,m) were taken from myceliagrowing on water-agar blocks and stained with lactophenol
Mycol Progress

Thus, these differences could be explained by the fact thatmany of the studies conducted in the northern hemispherehave been performed on Epichloë-infected grasses and onlyNeotyphodium seems to be present in South America.
Our data indicate that the high incidence of endophytes(>90%) in populations of the toxic species Festucahieronymi, F. argentina and Poa huecu, which cattle avoideating, is clearly associated with an advantage for infectedplants. High levels of infection were also observed in thosenon-toxic species that inhabit particular environments,including shady and moist forests (Festuca superba andBromus brachyanthera), swamps (F. fimbriata), or volcanicsoils (Poa durifolia). Taken together, these observationssuggest that, in these environments, endophytes couldprovide some benefit to the infected plants. On the otherhand, in most of the host species that inhabit a wide rangeof environments, endophyte incidence ranges from 0 tonearly 100%. Bromus auleticus seems to be the only non-toxic to cattle host that inhabits a wide range of environ-ments and presents a high incidence of endophytes in all thepopulations. However, is interesting to note that this speciesis an excellent forage grass that grows in a region that hasbeen historically subjected to high pressure by grazing bylivestock. In addition, greenhouse studies, conducted in ourlaboratory, indicated that endophyte-infected plants of B.auleticus present a higher regrowth rate and are moreresistant to defoliation than endophyte-free plants (Iannoneand Cabral 2006).
Some authors assert that the incidence of endophytesin populations does not necessarily reflect the advantagesor disadvantages that endophytes confer to the hosts(Saikkonen et al. 2002) because endophytes could be lostalong the life cycle of the host (Afkhami and Rudgers2008). Environmental conditions can alter the efficiencyof the transmission of the endophytes through seeds ortillers (Welty et al. 1987; Afkhami and Rudgers 2008; Hilland Roach 2009). Mathematical models predict that inannual grasses any existing spatial or temporal variabilityin transmission efficiency should be expected to drive thepatterns of endophyte infection frequency (Gundel et al.2008). However, in perennial grasses, as most of the speciesexamined in this work, the effect of the efficiency oftransmission via seeds on endophyte infection frequencycould be less important than in annual grasses. There-fore, the effect of environmental conditions on endophytetransmission or on the detrimental or beneficial effect ofnon toxic endophytes on the host plant should be furtherexplored.
The characterization of the endophytes isolated from thenew hosts suggests existence of a wide diversity ofendophytes. The isolates of some hosts seemed to bemorphologically similar to Neotyphodium tembladerae.However, the isolates of Festuca fimbriata and some
isolates from Poa lanigera and Poa durifolia could not bemorphologically ascribed to the previously describedspecies, N. tembladerae or N. pampeanum. Thus, theendophytes of these hosts could represent new species orvarieties of the species previously described.
The existence of a high diversity of asexual endophytesin Argentina has been previously detected by molecularstudies performed in previously isolated endophytes(Cabral et al. 1999; Gentile et al. 2005). In addition,phylogenetic studies have indicated a hybrid asexual state(Neotyphodium) for the previously analyzed endophytes often host species (Cabral et al. 1999; Gentile et al. 2005;Iannone et al. 2009). The conidia of all the endophytesisolated from new hosts presented in this paper exceed4.5 !m in length. Conidia of this size are comparable to theconidia of other hybrid asexual Neotyphodium endophytes(Kuldau et al. 1997), suggesting a possible hybrid origin forthe new isolates. Based on the diversity detected in theendophytes isolated from the newly reported hosts, anextensive phylogenetic analysis should be conducted inorder to establish the genotypic diversity of endophytes inSouth America and their phylogenetic relationships.
Asexual endophytes offer a tool for their utilization inbiological control and plant improvement programs (Kuldauand Bacon 2008). Most of the hosts presented in this workhave not been reported as toxic to cattle and some of them(Bromus auleticus, B. setifolius, Poa bonariensis and P.lanigera) are known to have good quality as forage (Burkart1969; Cabrera 1970; Millot 2001). Studies in order toevaluate the effect of the endophytes of some hosts on plantproductivity are being conducted in our laboratory.
Acknowledgements This paper is dedicated to the memory of Dr.Daniel Cabral, our friend, teacher and colleague, and one of the firstscientists to perceive the biological relevance of these grass-fungusinteractions in South America. This research was supported byUniversity of Buenos Aires (UBACyT X833), CONICET (PIP5311,PIP 1482), and ANPCyT, PAE-PICT N°58. PRHIDEB-CONICET :Publication 179
References
Afkhami ME, Rudgers JA (2008) Symbiosis lost: imperfect verticaltransmission of fungal endophytes in grasses. Am Nat 172:405–416
Bacon CW (1995) Toxic endophyte-infected tall fescue and rangegrasses: historic perspectives. J Anim Sci 73:861–870
Bazely RD, Ball JP, Vicari M, tanentzap AJ, Bérenger M, RakocevicT, Kok S (2007) Broad-scale patterns in the distribution ofvertically transmitted, asexual endophytes in four naturally-occurring grasses in Sweden. Ecography 30:367–374
Bertoni MD, Cabral D, Romero N, Dubcovsky J (1993) Endofitosfúngicos en especies sudamericanas de Festuca (Poaceae). BolSoc Argent Bot 29:25–34
Burkart A (1969) Gramíneas: la familia de los pastos. Parte II. In:Flora Ilustrada de Entre Ríos (Argentina). INTA, Buenos Aires
Mycol Progress

Cabral D, Cafaro MJ, Saidman B, Lugo M, Reddy PV, White JF Jr(1999) Evidence supporting the ocurrence of a new species ofendophyte in some South American grasses. Mycologia 91:315–325
Cabrera AL (1970) Gramíneas. In: Flora de la Provincia de BuenosAires. Parte II. INTA Buenos Aires
Cabrera AL (1976) Regiones fitogeográficas argentinas. In: KuglerWF (ed) Enciclopedia Argentina de agricultura y jardinería.Tomo 2, 2nd edn. Acme, Buenos Aires, pp 1–85
Chung KR, Schardl CL (1997) Sexual cycle and horizontaltransmission of the grass symbiont, Epichloë typhina. MycolRes 101:295–301
Clark EM, White JF, Patterson RM (1983) Improved histochemicaltechniques for the detection of Acremonium coenophialum in tallfescue and methods of in vitro culture of the fungus. J MicrobiolMethh 1:149–155
Clarke BB, White JF, Hurley RH, Torres MS, Sun S, Huff DR (2006)Endophyte-mediated suppression of dollar spot disease in finefescues. Plant Dis 90:994–998
Clay K (1990) Fungal endophyte of grasses. Annu Rev Ecol Syst21:275–297
Clay K, Leuchtmann A (1989) Infection of woodland grasses byfungal endophytes. Mycologia 81:805–811
Colabelli M, Torres MS, Clausen AM, Fernandez ON, Peretti A (2007)Neotyphodium spp.—grass association in natural grassland inArgentina. In Proceedings of the 6th International symposium onFungal Endophytes of Grasses, pp 95–99
Fernandez OA, Busso CA (1997) Arid and semi-arid rangelands: twothirds of Argentina. RALA Report 200:41–60
Fletcher LR, Harvey IC (1981) An association of a Lolium endophytewith ryegrass staggers. N Z Vet J 29:185–186
FunkCR,White JF (1997) Use of natural and transformed endophytes forturf improvement. In: BaconCW,Hill NS (eds)Neotyphodium/grassinteractions. Plenum, New York, pp 229–239
Gentile A, Rossi MA, Cabral D, Craven KD, Schardl CL (2005)Origin, divergence, and phylogeny of Epichloë endophytes ofnative Argentine grasses. Mol Phylogenet Evol 35:196–208
Granath G, Vicari M, Bazely DR, Ball JP, Puentes A, Rakocevic T(2007) Variation in the abundance of fungal endophytes in fescuegrasses along altitudinal and grazing gradients. Ecography30:422–430
Gundel PE, Batista WB, Texeira M, Martinez-Ghersa MA, OmaciniM, Ghersa CM (2008) Neotyphodium endophyte infectionfrequency in annual grass populations: relative importance ofmutualism and vertical transmission efficiency. Proc R Soc LondB 275:897–905
Hill NS, Roach PK (2009) Endophyte survival during seed storage:endophyte—host interactions and heritability. Crop Sci 49:1425–1430
Iannone LJ, Cabral D (2006) Effects of the Neotyphodium endophytestatus on plant performance of Bromus auleticus, a wild nativegrass from South America. Symbiosis 41:61–69
Iannone LJ, Cabral D, Schardl CL, Rossi MS (2009) Phylogeneticdivergence, morphological and physiological differences distinguisha new Neotyphodium endophyte species in the grass Bromusauleticus from South America. Mycologia 101:336–347
Kuldau G, Bacon C (2008) Clavicipitaceous endophytes: their abilityto enhance resistance of grasses to multiple stresses. Biol Control46:57–71
Kuldau GA, Tsai HF, Scharld CL (1997) Molecular systematics ofClavicipitaceae supporting monophyly of genus Epichloë and forgenus Ephelis. Mycologia 89:431–441
Leuchtmann A (1992) Systematics, distribution, and host specificityof grass endophytes. Nat Toxins 1(150–1):62
Leyronas C, Raynal G (2001) Presence of Neotyphodium-likeendophytes in European grasses. Ann Appl Biol 139:119–127
Lugo MA, Anton AM, Cabral D (1998) Micofilas en gramíneasSudamericanas. An Jard Bot Madr 56:15–22
Malinowski DP, Belesky DP (2000) Adaptations of endophyte-infected cool-season grasses to environmental stresses: mech-anisms of drought and mineral stress tolerance. Crop Sci40:923–940
Malinowski DP, Belesky DP (2006) Ecological importance ofNeotyphodium spp. grass endophytes in agroecosystems. Grass-land Science 52:1–14
Millot JC (2001) Bromus auleticus: una nueva especie domes-ticada. In PROCISUR-IICA (ed) Diálogo LVI: los recursosfitogenéticos del género Bromus en el cono sur. Montevideo,pp 3–5
Mirlohi A, Sabzalian MR, Sharifnabi B, Nekoui MK (2006) Widespreadoccurrence of Neotyphodium-like endophyte in populations ofBromus tomentellus Boiss.in Iran. FEMS Microbiol Lett 256(1):126–131
Moon CD, Miles CO, Järlfors U, Schardl CL (2002) The evolutionaryorigins of three new Neotyphodium endophyte species fromgrasses indigenous to the southern hemisphere. Mycologia94:694–711
Moon CD, Craven KD, Leuchtmann A, Clement SL, Schardl CL(2004) Prevalence of interspecific hybrids amongst asexualfungal endophytes of grasses. Mol Ecol 13:1455–1467
Moon CD, Guillaumin J-J, Ravel C, Li C, Craven KD, Schardl CL(2007) New Neotyphodium endophyte species from the grasstribes Stipeae and Meliceae. Mycologia 99:895–905
Nicora EG (1978) Graminea. Parte III. In: Flora Patagónica. INTA,Buenos Aires
Novas MV, Gentile A, Cabral D (2003) Comparative study of growthparameters on diaspores and seedlings between populations ofBromus setifolius from Patagonia, differing in Neotyphodiumendophyte infection. Flora 198:421–426
Novas MV, Collantes M, Cabral D (2007) Environmental effects ongrass-endophyte associations in the harsh conditions of southPatagonia. FEMS Microbiol Ecol 61:164–173
Novas MV, Iannone LJ, Godeas A, Cabral D (2009) Positiveassociation between mycorrhiza and foliar endophytes in Poabonariensis, a native grass. Mycol Prog 8:75–81
Parodi LR (1950) Las gramíneas tóxicas para el ganado en laRepública Argentina. Rev Arg Agronomia :163–229
Rivas H, Zanolli M (1909) La tembladera. Enfermedad propia de losanimales herbívoros de las regiones andinas. Rev Fac Agron VetPlata 5:5–35
Rudgers JA, Afkhami ME, Rúa MA, Davitt JA, Hammer SA, HuguetVM (2009) A fungus among us: broad patterns of endophytedistribution in the grasses. Ecology 90:1531–1539
Saikkonen K, Ahlholm J, Helander M, Lehtimäki S, Neimeläinen O(2000) Endophytic fungi in wild and cultivated grasses inFinland. Ecography 23:360–366
Saikkonen K, Ion D, Gyllenberg M (2002) The persistence ofvertically transmitted fungi in grass metapoplations. Proc R SocLond B 269:1397–1403
Sampson K (1933) The systemic infection of grasses by Epichloëtyphina (Pers.) Tul. Trans Br Mycol Soc 18:30–47
Schardl CL (1996) Epichloë species: fungals symbionts of grasses.Phytopathology 34:109–130
Schardl CL, Leuchtmann A, Chung KR, Penny D, Siegel MR (1997)Coevolution by common descent of fungal symbionts (Epichloëspp.) and grass hosts. Mol Biol Evol 14:133–143
Schardl CL, Leuchtmann A, Spiering MJ (2004) Symbiosis of grasseswith seed-borne fungal endophytes. Annu Rev Plant Biol55:315–340
Schulthess FM, Faeth SH (1998) Distribution, abundances, andassociations of the endophytic fungal community of Arizonafescue (Festuca arizonica). Mycologia 90:569–578
Mycol Progress

Stebbins GL (1981) Chromosomes and evolution in the genus Bromus(Gramineae). Bot Jahrb Syst 102(1–4):359–379
Vila Aiub MM, Demartin EB, Maseda P, Gundel PE, Ghersa CM(2001) Exploración de la presencia de hongos endofíticos empastos de la Estepa Patagónica. Actas de la I Reunión Binacionalde Ecología
Wei YK, Gao YB, Xu H, Su D, Zhang X, Wang YH, Lin F, Chen L,Nie LY, Ren AZ (2006) Occurrence of endophytes in grassesnative to northern China. Grass Forage Sci 61:422–429
Welty RE, Azevedo MD, Cooper TM (1987) Influence of moisturecontent, temperature, and length of storage on seed germinationand survival of endophytic fungi in seeds of tall fescue andperennial ryegrass. Phytopathology 77:893–900
White JF (1987) Widespread distribution of endophytes in thePoaceae. Plant Dis 71:340–342
White JF (1992) Endophyte-host associations in grasses. XVII.Ecological and physiological features characteriszing Epichloë
typhina and some anamorphic varities in England. Mycologia84:431–441
White JF, Baldwin NA (1992) A preliminary enumeration of grassendophytes in west central Europe grasslands. Sydowia 44:78–84
White JF, Cole GT (1985) Endophyte-host associations in foragegrasses. I. Distribution of fungal endophytes in some species ofLolium and Festuca. Mycologia 77:323–327
White JF, Cole GT (1986) Endophyte-host associations in foragegrasses. V. Occurrence of fungal endophytes in certain species ofBromus and Poa. Mycologia 78:846–850
White JF, Sullivan RF, Balady GA, Gianfagna TJ, Yue Q, Meyer WA,Cabral D (2001) A fungal endosymbiont of the grass Bromussetifolius: distribution in some Andean populations, identificationand examination of beneficial properties. Symbiosis 31:241–257
Zuloaga FO, Nicora EG, Rúgolo de Agrazar ZE, Morrone O, PensieroJ, Cialdella AM (1994) Catálogo de la familia Poaceae en laRepública Argentina. Missouri Botanical Garden
Mycol Progress




















