
Diverse actions of ouabain and its aglycone ouabagenin in renal cells
13
Diverse actions of ouabain and its aglycone ouabagenin in renal cells Raphael C. Valente & Luiz S. Capella & Márcia M. M. Oliveira & Luciana T. Nunes-Lima & Fernanda C. M. Cruz & Roberta R. Palmieri & Aníbal G. Lopes & Márcia A. M. Capella Received: 6 March 2009 / Accepted: 20 August 2009 / Published online: 9 September 2009 # Springer Science + Business Media B.V. 2009 Abstract The cellular actions of ouabain are complex and involve different pathways, depending on the cell type and experimental conditions. Several studies have reported that Madin–Darby canine kidney (MDCK) cellular sensitivity to ouabain is not related to Na-K-ATPase inhibition, and others showed that some cell types, such as Ma104, are resistant to ouabain toxicity albeit their Na-K-ATPase isoforms possess high affinity for this glycoside. We describe here that the effects of ouabain and ouabagenin also diverge in MDCK and Ma104 cells, being MDCK cells more resistant to ouabagenin, while Ma104 cells are resistant to both molecules. This feature seems to correlate with induction of cell signaling, since ouabain, but not ouabagenin, induced an intense and sustained increase in tyrosine phosphorylation levels in MDCK cells. Moreover, ouabain-induced phos- phorylation in Ma104 cells was approximately half than that observed in MDCK cells. The proportion between α and β subunits of Na-K-ATPase was similar in MDCK cells, though Ma104 cells presented more α subunits, located mainly at the cytoplasm. Furthermore, a fluorescent ouabain-analog labeled mainly the cytoplasm of Ma104 cells, the opposite of that seen in MDCK cells, corroborating the results using anti-Na-K-ATPase antibodies. Hence, the results suggest that ouabain and ouabagenin differ in terms of Na-K-ATPase inhibition and cell signaling activation in MDCK cells. Additionally, MDCK and Ma104 cell lines respond differently to ouabain, perhaps due to an intrinsic ability of this glycoside to selectively reach the cytoplasm of Ma104 cells. Keywords Ouabain . Ouabagenin . Na-K-ATPase . MDCK . Ma104 Abbreviations OUA Ouabain OBG Ouabagenin OUABDP Ouabain-Bodipy PTyr Phosphorylated tyrosine Introduction Na-K-ATPase is essential for both the maintenance of the membrane potential and for the transport of organic solutes in epithelial cells. It is presently known that this protein plays an essential role in the Cell Biol Toxicol (2010) 26:201–213 DOI 10.1007/s10565-009-9136-8 R. C. Valente : M. A. M. Capella Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brazil L. S. Capella : M. M. M. Oliveira : L. T. Nunes-Lima : F. C. M. Cruz : R. R. Palmieri : A. G. Lopes : M. A. M. Capella (*) Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, CCS—Bloco G, 21949-900 Rio de Janeiro, Rio de Janeiro, Brazil e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diverse actions of ouabain and its aglycone ouabagenin in renal cells

Diverse actions of ouabain and its aglycone ouabageninin renal cells
Raphael C. Valente & Luiz S. Capella & Márcia M. M. Oliveira &
Luciana T. Nunes-Lima & Fernanda C. M. Cruz & Roberta R. Palmieri &Aníbal G. Lopes & Márcia A. M. Capella
Received: 6 March 2009 /Accepted: 20 August 2009 /Published online: 9 September 2009# Springer Science + Business Media B.V. 2009
Abstract The cellular actions of ouabain are complexand involve different pathways, depending on the celltype and experimental conditions. Several studieshave reported that Madin–Darby canine kidney(MDCK) cellular sensitivity to ouabain is not relatedto Na-K-ATPase inhibition, and others showed thatsome cell types, such as Ma104, are resistant toouabain toxicity albeit their Na-K-ATPase isoformspossess high affinity for this glycoside. We describehere that the effects of ouabain and ouabagenin alsodiverge in MDCK and Ma104 cells, being MDCKcells more resistant to ouabagenin, while Ma104 cellsare resistant to both molecules. This feature seems tocorrelate with induction of cell signaling, sinceouabain, but not ouabagenin, induced an intense andsustained increase in tyrosine phosphorylation levelsin MDCK cells. Moreover, ouabain-induced phos-phorylation in Ma104 cells was approximately halfthan that observed in MDCK cells. The proportion
between α and β subunits of Na-K-ATPase wassimilar in MDCK cells, though Ma104 cells presentedmore α subunits, located mainly at the cytoplasm.Furthermore, a fluorescent ouabain-analog labeledmainly the cytoplasm of Ma104 cells, the oppositeof that seen in MDCK cells, corroborating the resultsusing anti-Na-K-ATPase antibodies. Hence, theresults suggest that ouabain and ouabagenin differ interms of Na-K-ATPase inhibition and cell signalingactivation in MDCK cells. Additionally, MDCK andMa104 cell lines respond differently to ouabain,perhaps due to an intrinsic ability of this glycosideto selectively reach the cytoplasm of Ma104 cells.
Keywords Ouabain . Ouabagenin . Na-K-ATPase .
MDCK . Ma104
AbbreviationsOUA OuabainOBG OuabageninOUABDP Ouabain-BodipyPTyr Phosphorylated tyrosine
Introduction
Na-K-ATPase is essential for both the maintenance ofthe membrane potential and for the transport oforganic solutes in epithelial cells. It is presentlyknown that this protein plays an essential role in the
Cell Biol Toxicol (2010) 26:201–213DOI 10.1007/s10565-009-9136-8
R. C. Valente :M. A. M. CapellaInstituto de Bioquímica Médica,Universidade Federal do Rio de Janeiro,Rio de Janeiro, Rio de Janeiro, Brazil
L. S. Capella :M. M. M. Oliveira : L. T. Nunes-Lima :F. C. M. Cruz : R. R. Palmieri :A. G. Lopes :M. A. M. Capella (*)Instituto de Biofísica Carlos Chagas Filho,Universidade Federal do Rio de Janeiro,CCS—Bloco G,21949-900 Rio de Janeiro, Rio de Janeiro, Brazile-mail: [email protected]

regulation of cell proliferation, differentiation, anddevelopment, as well as in cell attachment (for arecent review, see Aperia 2007).
It has been proposed that ouabain (OUA), acardiotonic steroid known as the most potent Na-K-ATPase inhibitor, is an endogenous hormone relatedto hypertension, since elevated plasma concentrationsof an endogenous analog of this glycoside have beendescribed in several patients with essential hyperten-sion (Manunta et al. 1999; Wang et al. 2003).Although this is a controversial issue, the fact thatthere is an endogenous analog of OUA in humanplasma increased the need for the comprehension ofthe cellular actions of this glycoside in both physio-logical and nonphysiological concentrations.
The sustained blockage of Na-K-ATPase by OUAwas initially thought to lead to cell death as a resultof hydroelectrolytic imbalance. However, severalreports suggested that ouabain-induced cell deathdoes not depend on the inhibition of the Na-K-ATPase-mediated ion fluxes (Contreras et al. 1995a;Ho et al. 2000; Pchejetski et al. 2003). In addition, ithas been shown that ouabain-induced sustainedphosphorylation of tyrosine residues is related tocell death (Contreras et al. 1999; Valente et al. 2003).Nevertheless, contrasting to its toxic effects, ouabaincan also protect cells from apoptosis induced byserum withdrawal (Orlov et al. 1999, 2000). Thus, itseems that cellular effects triggered by ouabain arean intricate matter, involving different pathways,which can cause or avoid cell death, depending onthe cell type and experimental conditions. Due tothis complexity, some studies have hypothesized theexistence of other functional receptors (Manunta etal. 2000, 2001; Ward et al. 2002), although the αsubunit of the Na-K-ATPase is so far the onlyrecognized receptor for ouabain.
It has been suggested that the activity and potencyof ouabain rely on the sugar moiety. Studies attempt-ing to elucidate the specific components of cardio-tonic steroids essential for Na-K-ATPase binding andinhibition demonstrated that the interaction of cardiacglycosides with Na-K-ATPase initiates with thebinding of the genin, which consequently unlocks abinding site for the sugar moiety (Yoda et al. 1973;Yoda 1974). The subsequent binding of rhamnosethen anchors the ouabain molecule tightly to thesodium pump (Yoda and Yoda 1977). On the otherhand, the dissociation of the glycoside seems to begin
with the dissociation of the sugar moiety, followed bya conformational change that closes the sugar bindingsite and facilitates unbinding of the aglycone moiety(Yoda 1973). Therefore, the sugar moiety appears tobe important in uptake, distribution, and bindingstability, since when it is cleaved from the glycoside,the genin remained retains a diminished affinity to thesodium pump (Ozaki et al. 1984; Hallaq et al. 1991;Manunta et al. 2001).
The effects of cardiotonic steroids on survival ofdifferent cell types may be caused by cell type-specific expression of intermediates of cell signalinginvolved in triggering and progression of the celldeath machinery, or to unidentified ouabain receptors.Another possibility is that the differences observedamong ouabain effects in diverse cell lines (Contreraset al. 1995a, 1999; Ho et al. 2000; Pchejetski et al.2003; Valente et al. 2003; Orlov et al. 1999, 2000)might be related to unusual interactions of Na-K-ATPase with the sugar moiety, which could lead tothe activation of different signaling pathways.
If this hypothesis is correct, then ouabain and itsaglycone ouabagenin (OBG) would probably triggerdiverse responses in different cell lines. Therefore, inthe present study, we aimed to evaluate the effects ofouabain and ouabagenin in Ma104 and Madin–Darbycanine kidney (MDCK), two cell lines previouslydescribed as having different sensitivities to ouabain,although their Na-K-ATPases have the same affinityfor this glycoside (Contreras et al. 1995a, b, 1999;Valente et al. 2003).
Materials and methods
Cells and culture conditions
The epithelial cell lines Ma104 (obtained frommonkey kidney) and MDCK (derived from caninekidney) were grown in Dulbecco’s modified Eaglemedium (GIBCO, USA) with penicillin and strepto-mycin (GIBCO, USA) and supplemented with 10%fetal bovine serum (GIBCO, USA) in disposableplastic bottles, at 37°C, until reach confluence.
Treatments and analysis of cellular viability
Ma104 and MDCK cells were seeded in 96-wellplates, at a concentration of 4×104 cells per milliliter.
202 Cell Biol Toxicol (2010) 26:201–213

All experiments were performed 24 h after seeding, toensure uniform attachment of the cells at the onset ofthe experiments. OUA or OBG was added (concen-trations ranging from 10−7 to 10−5 M), and the cellswere incubated for 48 h. After incubation, the cellularviability was measured using the thiazolyl blue(MTT) assay (Mosmann 1983) on a microplate reader(absorbance at 490 nm). Each experiment wasrepeated at least four times using triplicate samples.
Immunofluorescence: labeling of phosphotyrosine
MDCK cells were seeded at 1×105 cells per milliliterin coverslips and after 24 h were treated with OUA orOBG (10−6 M) for 3 h. After incubation, cells werewashed three times with phosphate buffered salinesolution (PBS), fixed and permeabilized with coldmethanol for 3 min, followed by incubation with 10%fetal bovine serum for 30 min at room temperature.After this time, cells were incubated overnight inhumidified chamber with the mouse monoclonalantibody PTyr (PY20), which specifically detectsphosphotyrosine-containing proteins and do notcross-react with phosphoserine or phosphothreonine(Santa Cruz Biotechnologies, USA). After washingwith PBS, cells were incubated with Alexa Fluor 546antimouse secondary antibody (Molecular Probes,USA) for 1 h at room temperature, in the dark,washed three times with PBS and once with water,and then the coverslips were mounted in glass slideswith 50% glycerol in PBS. The fluorescence wasvisualized under an Axiovert 100 microscope (Zeiss,Germany) using a 40× fluar oil objective (Zeiss,Germany).
Immunofluorescence: labeling of Na-K-ATPase
Cells grown on coverslips at 1×105 cells per milliliterwere washed three times with PBS and fixed in coldmethanol for 3 min. After new washing, cells wereincubated with 10% fetal bovine serum for 30 min atroom temperature, followed by incubation with ananti-α1 isoform antibody of Na-K-ATPase (SantaCruz Biotechnology, USA). Rabbit antigoat AlexaFluor 546-conjugated secondary antibody was usedfor detection (Molecular Probes, USA). After washingthree times with PBS and once with water, thecoverslips were mounted in glass slides with antifad-ing kit (Molecular Probes, Invitrogen, USA), and the
fluorescence was visualized under an Axiovert 100microscope (Zeiss, Munich, Germany), using a 40×fluar oil objective (Zeiss). Quantification of fluores-cence levels was analyzed using Image Tool softwareversion 3.00 (UTHSCSA, USA) and expressed inarbitrary units.
Ouabain binding fluorescence detection
Ma104 and MDCK cells were grown at 1×105 cellsper milliliter in special Petri dishes (Scharfstein et al.2000) and further incubated with 10−6 M ouabain-Bodipy (OUABDP; Molecular Probes, Invitrogen,USA) in serum-free medium for 30 min. After that,cells were washed with PBS to remove the excess ofOUABDP in the medium, and the fluorescence wasimmediately observed under an Axiovert 100 micro-scope (Zeiss, Germany) using a 40× fluar oilobjective (Zeiss, Germany).
Flow cytometry
Flow cytometry was employed to quantify the bindingof the fluorescent ouabain analog OUABDP. Ma104and MDCK cells were plated on 24-well plates (TPP,Germany) at a concentration of 1×105 cells permilliliter and the experiments were performed 24 hafter seeding. These cells were pre-incubated witheither ouabain or ouabagenin, more specifically 10−6
and 10−5 M (MDCK cells) and 10−6, 10−5, and 10−4M(Ma104 cells), for 30 min, and then OUABDP wasadded in a concentration of 10−6 M. After 30 min ofincubation, cells were washed with PBS, harvested inHanks solution containing ethylenediaminetetraaceticacid (EDTA), and the fluorescence was measured byflow cytometry in a FACSCalibur device (Becton andDickinson, USA), equipped with an air-cooled ArgonLaser tuned to emit 15 mW in 488 nm. Thefluorescence was measured through a 530-nm long-pass filter.
Measurement of ATPase activity
Confluent monolayers of the two cell lines grown onplastic bottles (TPP, Germany) were removed by cellscrapper and solubilized in a solution containingdeoxycholic acid 0.1%, EDTA 1 mM, 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES)–Tris 20 mM, and sucrose 250 mM. All reagents were
Cell Biol Toxicol (2010) 26:201–213 203
obtained from SIGMA, USA. The ATPase activitywas measured as described elsewhere (Jorgensen andSkou 1971; Grubmeyer and Penefsky 1981) in thecellular extracts incubated with different concentra-tions of ouabain (10−9 to 10−3 M). The composition ofthe standard assay medium used to measure Na-K-ATPase activity was 10 mM MgCl2, 5 mM [γ-32P]ATP, 20 mM HEPES–Tris (pH 7.0), 5 mM azide,120 mM NaCl, and 30 mM KCl. Protein concen-trations were determined by the Folin phenol method(Lowry et al. 1951) using bovine serum albumin as astandard. Each experiment was performed six timesusing independent preparations. After a sigmoidaldose–response curve fitting, the I50 values weredetermined as the concentration of drug that inhibited50% of the maximum Na-K-ATPase activity.
Western blot analysis
Expression of Na-K-ATPase α and β subunits inMDCK and Ma104 cells was assessed by immuno-blotting using specific antibodies (Santa Cruz Bio-technology, USA). After aspiration of the culturemedia, confluent monolayers were washed three timeswith PBS (pH 7.4) at room temperature, scraped, andcentrifuged at 8,000×g for 90 s. Whole-cell extractswere prepared by diluting the cell pellets directly insample buffer, consisting of 1.5% sodium dodecylsulfate (SDS), 10 mM Tris[hydroxymethyl]amino-methane (Tris-Cl), pH 6.8, 0.6% dithio-L-threitol,0.5% β-mercaptoethanol, and 6% glycerol. Theproteins were immediately subjected to SDS-polyacrylamide gel electrophoresis and transferred toa polyvinylidene fluoride membrane (Bio-Rad, Her-cules, CA, USA). The membranes were blocked withWestern Breeze blocking solution (Invitrogen, USA)and incubated with specific antibodies against α1 andβ1 isoforms of Na-K-ATPase (Santa Cruz, USA),which are the major isoforms found in kidney. Thephosphatase alkaline Western Breeze Kit (Invitrogen,USA) was used to visualize the bands in themembranes.
Statistical analysis
Each experiment was repeated from three to six times.Data are expressed as means ± standard errors of themeans (SEM) and were analyzed using one-wayanalysis of variance followed by Dunnet or Bonfer-
roni test, or Student’s t test for comparison of thedifferences. Values of P<0.05 were consideredstatistically significant.
Results
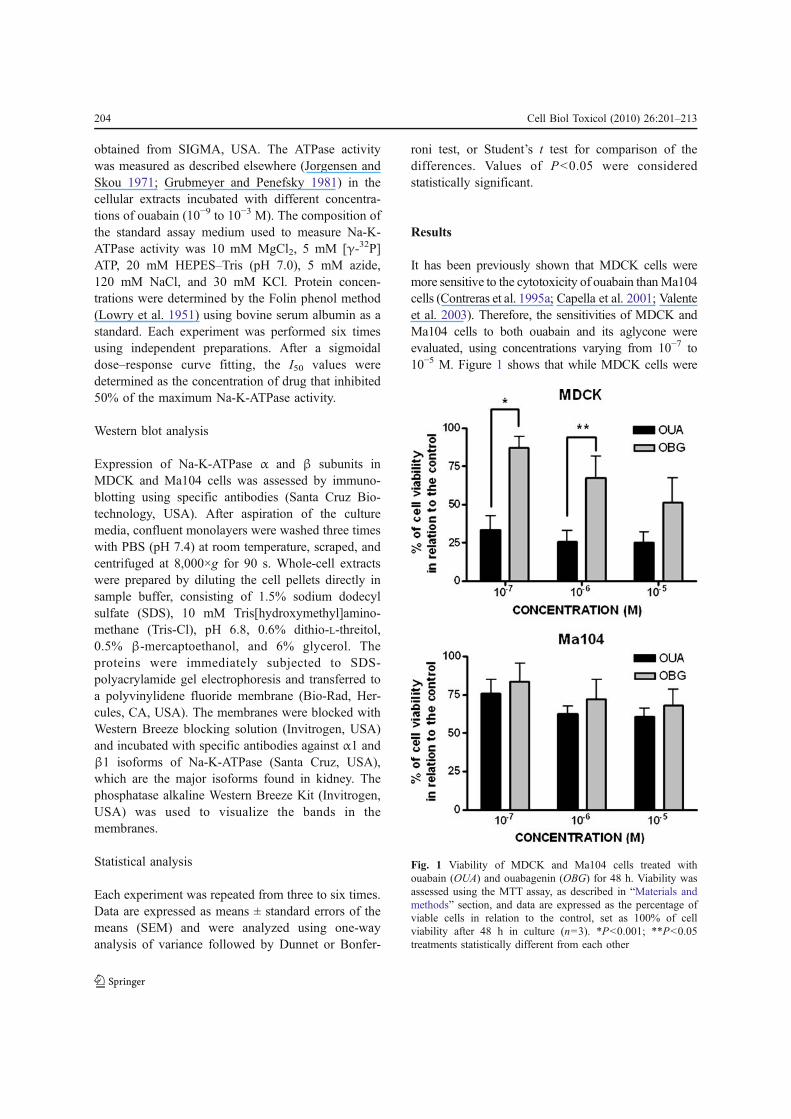
It has been previously shown that MDCK cells weremore sensitive to the cytotoxicity of ouabain than Ma104cells (Contreras et al. 1995a; Capella et al. 2001; Valenteet al. 2003). Therefore, the sensitivities of MDCK andMa104 cells to both ouabain and its aglycone wereevaluated, using concentrations varying from 10−7 to10−5 M. Figure 1 shows that while MDCK cells were
Fig. 1 Viability of MDCK and Ma104 cells treated withouabain (OUA) and ouabagenin (OBG) for 48 h. Viability wasassessed using the MTT assay, as described in “Materials andmethods” section, and data are expressed as the percentage ofviable cells in relation to the control, set as 100% of cellviability after 48 h in culture (n=3). *P<0.001; **P<0.05treatments statistically different from each other
204 Cell Biol Toxicol (2010) 26:201–213
indeed more sensitive to the cytotoxicity of OUA thanOBG (cells treated with 10−7 and 10−6 M of thesesubstances), Ma104 cells were equally resistant to bothmolecules. In addition, these two cell lines were equallyresistant to 10−6 M OBG (viability in both cases nearly70% in relation to control, which was considered to be100% of cell viability).
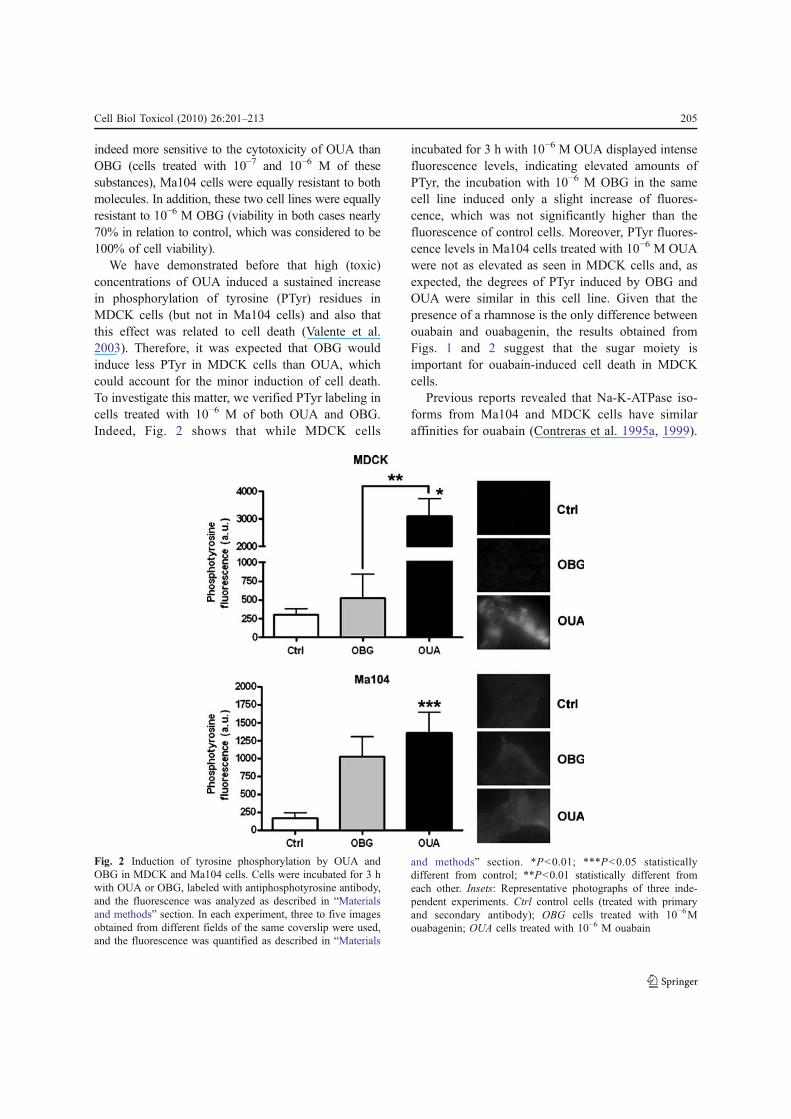
We have demonstrated before that high (toxic)concentrations of OUA induced a sustained increasein phosphorylation of tyrosine (PTyr) residues inMDCK cells (but not in Ma104 cells) and also thatthis effect was related to cell death (Valente et al.2003). Therefore, it was expected that OBG wouldinduce less PTyr in MDCK cells than OUA, whichcould account for the minor induction of cell death.To investigate this matter, we verified PTyr labeling incells treated with 10−6 M of both OUA and OBG.Indeed, Fig. 2 shows that while MDCK cells
incubated for 3 h with 10−6 M OUA displayed intensefluorescence levels, indicating elevated amounts ofPTyr, the incubation with 10−6 M OBG in the samecell line induced only a slight increase of fluores-cence, which was not significantly higher than thefluorescence of control cells. Moreover, PTyr fluores-cence levels in Ma104 cells treated with 10−6 M OUAwere not as elevated as seen in MDCK cells and, asexpected, the degrees of PTyr induced by OBG andOUA were similar in this cell line. Given that thepresence of a rhamnose is the only difference betweenouabain and ouabagenin, the results obtained fromFigs. 1 and 2 suggest that the sugar moiety isimportant for ouabain-induced cell death in MDCKcells.
Previous reports revealed that Na-K-ATPase iso-forms from Ma104 and MDCK cells have similaraffinities for ouabain (Contreras et al. 1995a, 1999).
Fig. 2 Induction of tyrosine phosphorylation by OUA andOBG in MDCK and Ma104 cells. Cells were incubated for 3 hwith OUA or OBG, labeled with antiphosphotyrosine antibody,and the fluorescence was analyzed as described in “Materialsand methods” section. In each experiment, three to five imagesobtained from different fields of the same coverslip were used,and the fluorescence was quantified as described in “Materials
and methods” section. *P<0.01; ***P<0.05 statisticallydifferent from control; **P<0.01 statistically different fromeach other. Insets: Representative photographs of three inde-pendent experiments. Ctrl control cells (treated with primaryand secondary antibody); OBG cells treated with 10−6Mouabagenin; OUA cells treated with 10−6 M ouabain
Cell Biol Toxicol (2010) 26:201–213 205
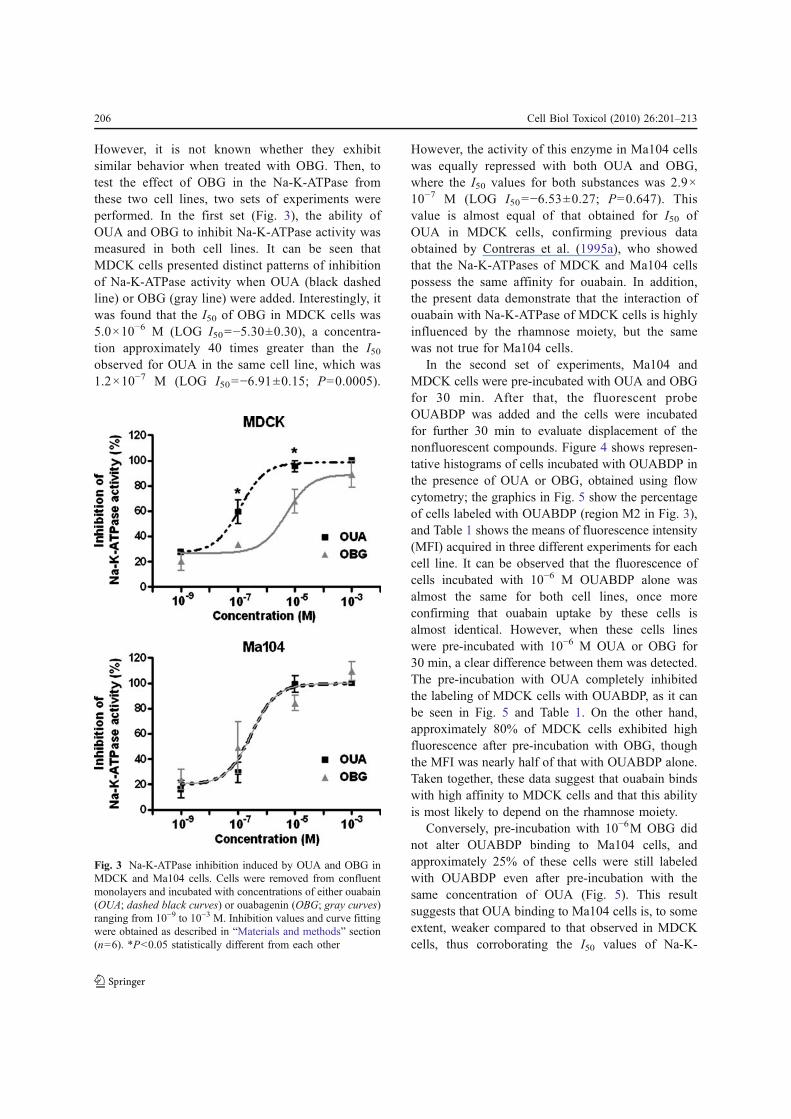
However, it is not known whether they exhibitsimilar behavior when treated with OBG. Then, totest the effect of OBG in the Na-K-ATPase fromthese two cell lines, two sets of experiments wereperformed. In the first set (Fig. 3), the ability ofOUA and OBG to inhibit Na-K-ATPase activity wasmeasured in both cell lines. It can be seen thatMDCK cells presented distinct patterns of inhibitionof Na-K-ATPase activity when OUA (black dashedline) or OBG (gray line) were added. Interestingly, itwas found that the I50 of OBG in MDCK cells was5.0×10−6 M (LOG I50=−5.30±0.30), a concentra-tion approximately 40 times greater than the I50
observed for OUA in the same cell line, which was1.2×10−7 M (LOG I50=−6.91±0.15; P=0.0005).
However, the activity of this enzyme in Ma104 cellswas equally repressed with both OUA and OBG,where the I50 values for both substances was 2.9×10−7 M (LOG I50=−6.53±0.27; P=0.647). Thisvalue is almost equal of that obtained for I50 ofOUA in MDCK cells, confirming previous dataobtained by Contreras et al. (1995a), who showedthat the Na-K-ATPases of MDCK and Ma104 cellspossess the same affinity for ouabain. In addition,the present data demonstrate that the interaction ofouabain with Na-K-ATPase of MDCK cells is highlyinfluenced by the rhamnose moiety, but the samewas not true for Ma104 cells.
In the second set of experiments, Ma104 andMDCK cells were pre-incubated with OUA and OBGfor 30 min. After that, the fluorescent probeOUABDP was added and the cells were incubatedfor further 30 min to evaluate displacement of thenonfluorescent compounds. Figure 4 shows represen-tative histograms of cells incubated with OUABDP inthe presence of OUA or OBG, obtained using flowcytometry; the graphics in Fig. 5 show the percentageof cells labeled with OUABDP (region M2 in Fig. 3),and Table 1 shows the means of fluorescence intensity(MFI) acquired in three different experiments for eachcell line. It can be observed that the fluorescence ofcells incubated with 10−6 M OUABDP alone wasalmost the same for both cell lines, once moreconfirming that ouabain uptake by these cells isalmost identical. However, when these cells lineswere pre-incubated with 10−6 M OUA or OBG for30 min, a clear difference between them was detected.The pre-incubation with OUA completely inhibitedthe labeling of MDCK cells with OUABDP, as it canbe seen in Fig. 5 and Table 1. On the other hand,approximately 80% of MDCK cells exhibited highfluorescence after pre-incubation with OBG, thoughthe MFI was nearly half of that with OUABDP alone.Taken together, these data suggest that ouabain bindswith high affinity to MDCK cells and that this abilityis most likely to depend on the rhamnose moiety.
Conversely, pre-incubation with 10−6M OBG didnot alter OUABDP binding to Ma104 cells, andapproximately 25% of these cells were still labeledwith OUABDP even after pre-incubation with thesame concentration of OUA (Fig. 5). This resultsuggests that OUA binding to Ma104 cells is, to someextent, weaker compared to that observed in MDCKcells, thus corroborating the I50 values of Na-K-
Fig. 3 Na-K-ATPase inhibition induced by OUA and OBG inMDCK and Ma104 cells. Cells were removed from confluentmonolayers and incubated with concentrations of either ouabain(OUA; dashed black curves) or ouabagenin (OBG; gray curves)ranging from 10−9 to 10−3 M. Inhibition values and curve fittingwere obtained as described in “Materials and methods” section(n=6). *P<0.05 statistically different from each other
206 Cell Biol Toxicol (2010) 26:201–213
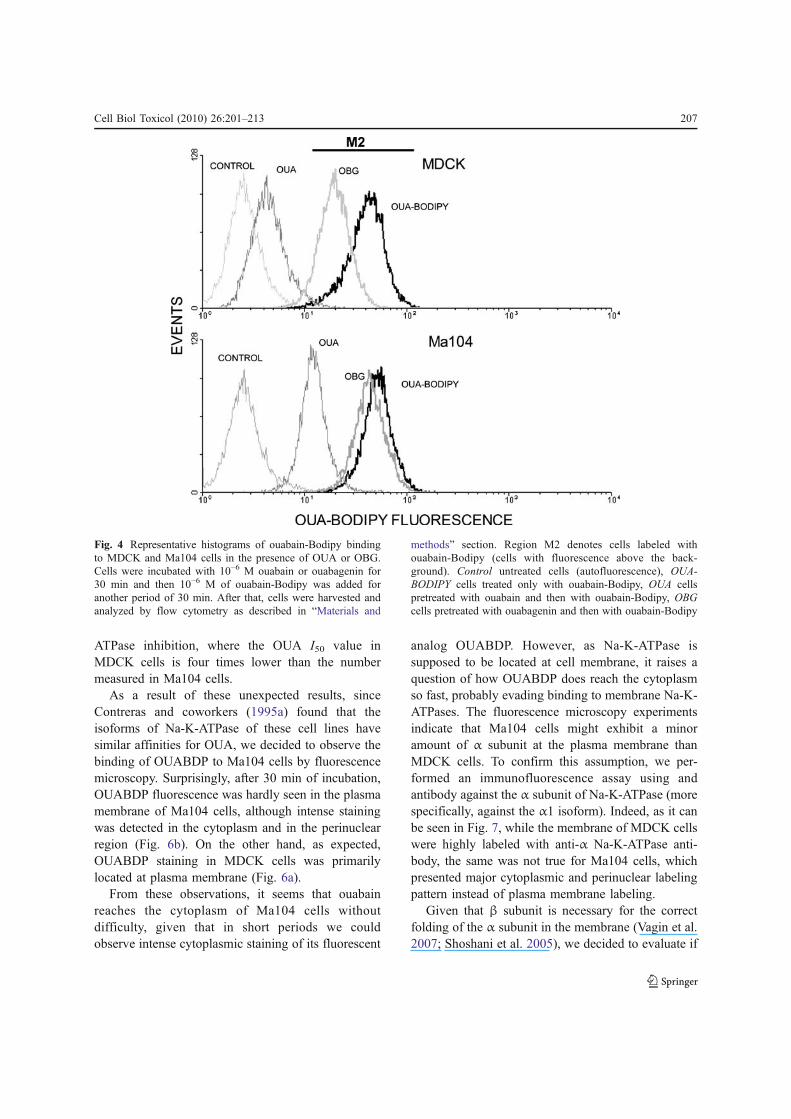
ATPase inhibition, where the OUA I50 value inMDCK cells is four times lower than the numbermeasured in Ma104 cells.
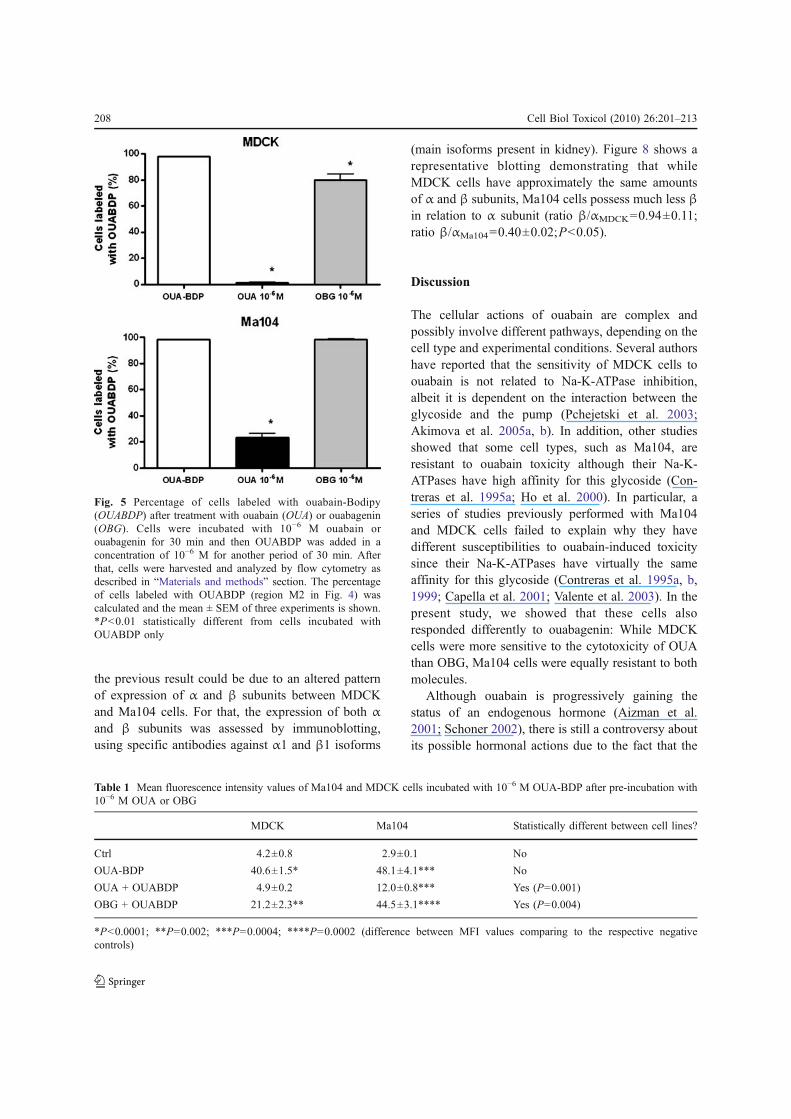
As a result of these unexpected results, sinceContreras and coworkers (1995a) found that theisoforms of Na-K-ATPase of these cell lines havesimilar affinities for OUA, we decided to observe thebinding of OUABDP to Ma104 cells by fluorescencemicroscopy. Surprisingly, after 30 min of incubation,OUABDP fluorescence was hardly seen in the plasmamembrane of Ma104 cells, although intense stainingwas detected in the cytoplasm and in the perinuclearregion (Fig. 6b). On the other hand, as expected,OUABDP staining in MDCK cells was primarilylocated at plasma membrane (Fig. 6a).
From these observations, it seems that ouabainreaches the cytoplasm of Ma104 cells withoutdifficulty, given that in short periods we couldobserve intense cytoplasmic staining of its fluorescent
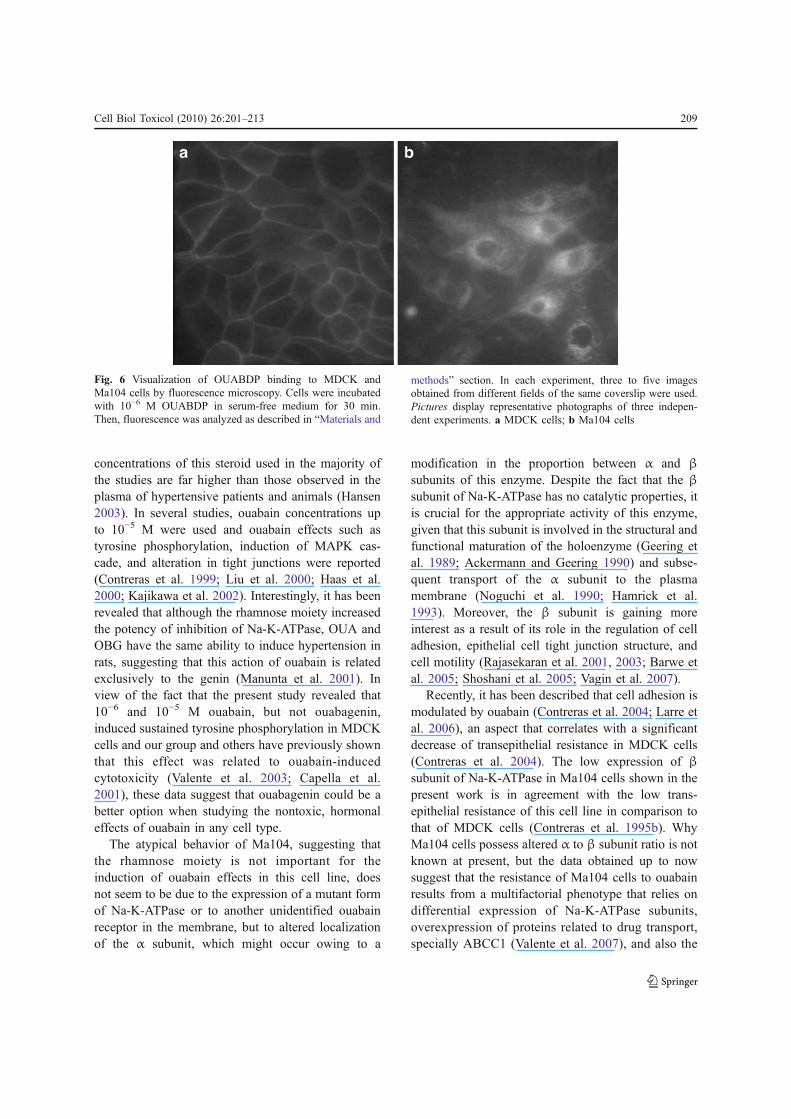
analog OUABDP. However, as Na-K-ATPase issupposed to be located at cell membrane, it raises aquestion of how OUABDP does reach the cytoplasmso fast, probably evading binding to membrane Na-K-ATPases. The fluorescence microscopy experimentsindicate that Ma104 cells might exhibit a minoramount of α subunit at the plasma membrane thanMDCK cells. To confirm this assumption, we per-formed an immunofluorescence assay using andantibody against the α subunit of Na-K-ATPase (morespecifically, against the α1 isoform). Indeed, as it canbe seen in Fig. 7, while the membrane of MDCK cellswere highly labeled with anti-α Na-K-ATPase anti-body, the same was not true for Ma104 cells, whichpresented major cytoplasmic and perinuclear labelingpattern instead of plasma membrane labeling.
Given that β subunit is necessary for the correctfolding of the α subunit in the membrane (Vagin et al.2007; Shoshani et al. 2005), we decided to evaluate if
Fig. 4 Representative histograms of ouabain-Bodipy bindingto MDCK and Ma104 cells in the presence of OUA or OBG.Cells were incubated with 10−6 M ouabain or ouabagenin for30 min and then 10−6 M of ouabain-Bodipy was added foranother period of 30 min. After that, cells were harvested andanalyzed by flow cytometry as described in “Materials and
methods” section. Region M2 denotes cells labeled withouabain-Bodipy (cells with fluorescence above the back-ground). Control untreated cells (autofluorescence), OUA-BODIPY cells treated only with ouabain-Bodipy, OUA cellspretreated with ouabain and then with ouabain-Bodipy, OBGcells pretreated with ouabagenin and then with ouabain-Bodipy
Cell Biol Toxicol (2010) 26:201–213 207
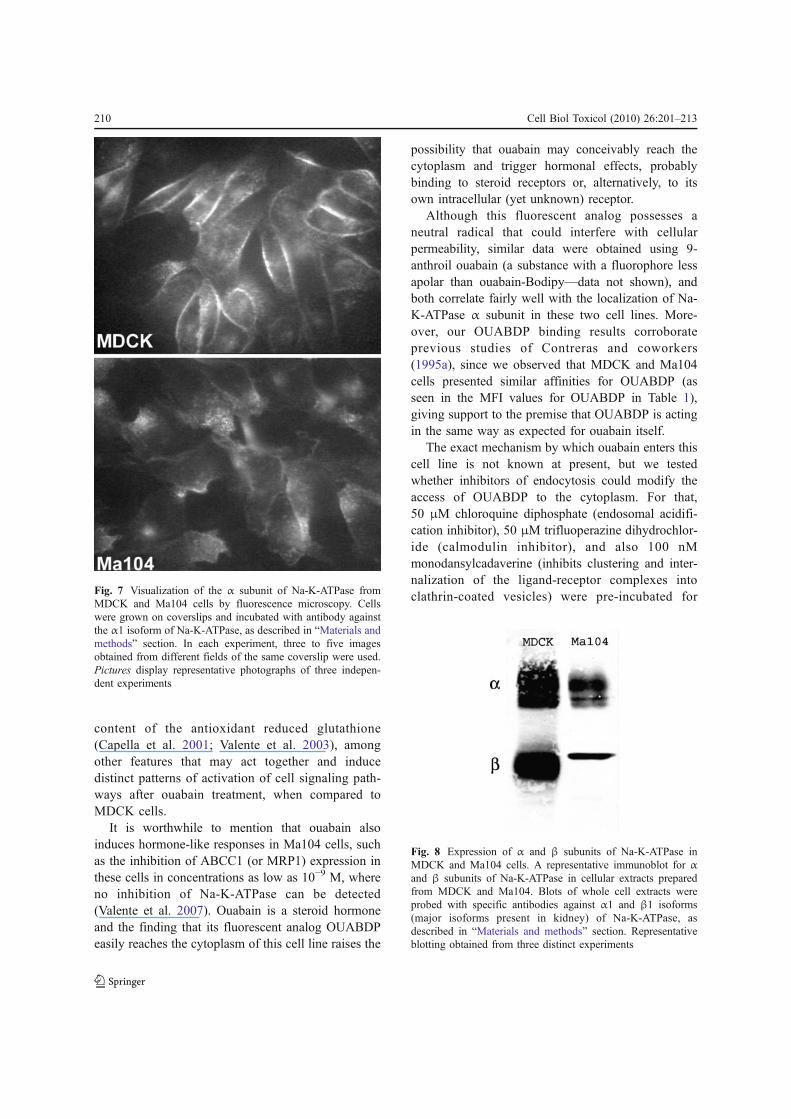
the previous result could be due to an altered patternof expression of α and β subunits between MDCKand Ma104 cells. For that, the expression of both αand β subunits was assessed by immunoblotting,using specific antibodies against α1 and β1 isoforms
(main isoforms present in kidney). Figure 8 shows arepresentative blotting demonstrating that whileMDCK cells have approximately the same amountsof α and β subunits, Ma104 cells possess much less βin relation to α subunit (ratio β/αMDCK=0.94±0.11;ratio β/αMa104=0.40±0.02;P<0.05).
Discussion
The cellular actions of ouabain are complex andpossibly involve different pathways, depending on thecell type and experimental conditions. Several authorshave reported that the sensitivity of MDCK cells toouabain is not related to Na-K-ATPase inhibition,albeit it is dependent on the interaction between theglycoside and the pump (Pchejetski et al. 2003;Akimova et al. 2005a, b). In addition, other studiesshowed that some cell types, such as Ma104, areresistant to ouabain toxicity although their Na-K-ATPases have high affinity for this glycoside (Con-treras et al. 1995a; Ho et al. 2000). In particular, aseries of studies previously performed with Ma104and MDCK cells failed to explain why they havedifferent susceptibilities to ouabain-induced toxicitysince their Na-K-ATPases have virtually the sameaffinity for this glycoside (Contreras et al. 1995a, b,1999; Capella et al. 2001; Valente et al. 2003). In thepresent study, we showed that these cells alsoresponded differently to ouabagenin: While MDCKcells were more sensitive to the cytotoxicity of OUAthan OBG, Ma104 cells were equally resistant to bothmolecules.
Although ouabain is progressively gaining thestatus of an endogenous hormone (Aizman et al.2001; Schoner 2002), there is still a controversy aboutits possible hormonal actions due to the fact that the
Fig. 5 Percentage of cells labeled with ouabain-Bodipy(OUABDP) after treatment with ouabain (OUA) or ouabagenin(OBG). Cells were incubated with 10−6 M ouabain orouabagenin for 30 min and then OUABDP was added in aconcentration of 10−6 M for another period of 30 min. Afterthat, cells were harvested and analyzed by flow cytometry asdescribed in “Materials and methods” section. The percentageof cells labeled with OUABDP (region M2 in Fig. 4) wascalculated and the mean ± SEM of three experiments is shown.*P<0.01 statistically different from cells incubated withOUABDP only
Table 1 Mean fluorescence intensity values of Ma104 and MDCK cells incubated with 10−6 M OUA-BDP after pre-incubation with10−6 M OUA or OBG
MDCK Ma104 Statistically different between cell lines?
Ctrl 4.2±0.8 2.9±0.1 No
OUA-BDP 40.6±1.5* 48.1±4.1*** No
OUA + OUABDP 4.9±0.2 12.0±0.8*** Yes (P=0.001)
OBG + OUABDP 21.2±2.3** 44.5±3.1**** Yes (P=0.004)
*P<0.0001; **P=0.002; ***P=0.0004; ****P=0.0002 (difference between MFI values comparing to the respective negativecontrols)
208 Cell Biol Toxicol (2010) 26:201–213
concentrations of this steroid used in the majority ofthe studies are far higher than those observed in theplasma of hypertensive patients and animals (Hansen2003). In several studies, ouabain concentrations upto 10−5 M were used and ouabain effects such astyrosine phosphorylation, induction of MAPK cas-cade, and alteration in tight junctions were reported(Contreras et al. 1999; Liu et al. 2000; Haas et al.2000; Kajikawa et al. 2002). Interestingly, it has beenrevealed that although the rhamnose moiety increasedthe potency of inhibition of Na-K-ATPase, OUA andOBG have the same ability to induce hypertension inrats, suggesting that this action of ouabain is relatedexclusively to the genin (Manunta et al. 2001). Inview of the fact that the present study revealed that10−6 and 10−5 M ouabain, but not ouabagenin,induced sustained tyrosine phosphorylation in MDCKcells and our group and others have previously shownthat this effect was related to ouabain-inducedcytotoxicity (Valente et al. 2003; Capella et al.2001), these data suggest that ouabagenin could be abetter option when studying the nontoxic, hormonaleffects of ouabain in any cell type.
The atypical behavior of Ma104, suggesting thatthe rhamnose moiety is not important for theinduction of ouabain effects in this cell line, doesnot seem to be due to the expression of a mutant formof Na-K-ATPase or to another unidentified ouabainreceptor in the membrane, but to altered localizationof the α subunit, which might occur owing to a
modification in the proportion between α and βsubunits of this enzyme. Despite the fact that the βsubunit of Na-K-ATPase has no catalytic properties, itis crucial for the appropriate activity of this enzyme,given that this subunit is involved in the structural andfunctional maturation of the holoenzyme (Geering etal. 1989; Ackermann and Geering 1990) and subse-quent transport of the α subunit to the plasmamembrane (Noguchi et al. 1990; Hamrick et al.1993). Moreover, the β subunit is gaining moreinterest as a result of its role in the regulation of celladhesion, epithelial cell tight junction structure, andcell motility (Rajasekaran et al. 2001, 2003; Barwe etal. 2005; Shoshani et al. 2005; Vagin et al. 2007).
Recently, it has been described that cell adhesion ismodulated by ouabain (Contreras et al. 2004; Larre etal. 2006), an aspect that correlates with a significantdecrease of transepithelial resistance in MDCK cells(Contreras et al. 2004). The low expression of βsubunit of Na-K-ATPase in Ma104 cells shown in thepresent work is in agreement with the low trans-epithelial resistance of this cell line in comparison tothat of MDCK cells (Contreras et al. 1995b). WhyMa104 cells possess altered α to β subunit ratio is notknown at present, but the data obtained up to nowsuggest that the resistance of Ma104 cells to ouabainresults from a multifactorial phenotype that relies ondifferential expression of Na-K-ATPase subunits,overexpression of proteins related to drug transport,specially ABCC1 (Valente et al. 2007), and also the
Fig. 6 Visualization of OUABDP binding to MDCK andMa104 cells by fluorescence microscopy. Cells were incubatedwith 10−6 M OUABDP in serum-free medium for 30 min.Then, fluorescence was analyzed as described in “Materials and
methods” section. In each experiment, three to five imagesobtained from different fields of the same coverslip were used.Pictures display representative photographs of three indepen-dent experiments. a MDCK cells; b Ma104 cells
Cell Biol Toxicol (2010) 26:201–213 209
content of the antioxidant reduced glutathione(Capella et al. 2001; Valente et al. 2003), amongother features that may act together and inducedistinct patterns of activation of cell signaling path-ways after ouabain treatment, when compared toMDCK cells.
It is worthwhile to mention that ouabain alsoinduces hormone-like responses in Ma104 cells, suchas the inhibition of ABCC1 (or MRP1) expression inthese cells in concentrations as low as 10−9 M, whereno inhibition of Na-K-ATPase can be detected(Valente et al. 2007). Ouabain is a steroid hormoneand the finding that its fluorescent analog OUABDPeasily reaches the cytoplasm of this cell line raises the
possibility that ouabain may conceivably reach thecytoplasm and trigger hormonal effects, probablybinding to steroid receptors or, alternatively, to itsown intracellular (yet unknown) receptor.
Although this fluorescent analog possesses aneutral radical that could interfere with cellularpermeability, similar data were obtained using 9-anthroil ouabain (a substance with a fluorophore lessapolar than ouabain-Bodipy—data not shown), andboth correlate fairly well with the localization of Na-K-ATPase α subunit in these two cell lines. More-over, our OUABDP binding results corroborateprevious studies of Contreras and coworkers(1995a), since we observed that MDCK and Ma104cells presented similar affinities for OUABDP (asseen in the MFI values for OUABDP in Table 1),giving support to the premise that OUABDP is actingin the same way as expected for ouabain itself.
The exact mechanism by which ouabain enters thiscell line is not known at present, but we testedwhether inhibitors of endocytosis could modify theaccess of OUABDP to the cytoplasm. For that,50 μM chloroquine diphosphate (endosomal acidifi-cation inhibitor), 50 μM trifluoperazine dihydrochlor-ide (calmodulin inhibitor), and also 100 nMmonodansylcadaverine (inhibits clustering and inter-nalization of the ligand-receptor complexes intoclathrin-coated vesicles) were pre-incubated for
Fig. 8 Expression of α and β subunits of Na-K-ATPase inMDCK and Ma104 cells. A representative immunoblot for αand β subunits of Na-K-ATPase in cellular extracts preparedfrom MDCK and Ma104. Blots of whole cell extracts wereprobed with specific antibodies against α1 and β1 isoforms(major isoforms present in kidney) of Na-K-ATPase, asdescribed in “Materials and methods” section. Representativeblotting obtained from three distinct experiments
Fig. 7 Visualization of the α subunit of Na-K-ATPase fromMDCK and Ma104 cells by fluorescence microscopy. Cellswere grown on coverslips and incubated with antibody againstthe α1 isoform of Na-K-ATPase, as described in “Materials andmethods” section. In each experiment, three to five imagesobtained from different fields of the same coverslip were used.Pictures display representative photographs of three indepen-dent experiments
210 Cell Biol Toxicol (2010) 26:201–213

20 min before the addition of OUABDP and main-tained during the period of uptake of the fluorescentanalog (30 min at 37°C). However, none of thesesubstances altered the pattern of cytoplasmic andperinuclear labeling of OUABDP in Ma104 cells, afeature that was only observed when OUABDPincubation was performed at 4°C, where the accessof this fluorescent analog to the cytoplasm wasremarkably diminished (data not shown).
It has been postulated that adrenocortical cellspossess another binding site with high affinity forouabain, which was not detected in skeletal muscleor liver membranes (Ward et al. 2002). Recently,these new binding sites were associated with atransport of ouabain into sealed vesicles that couldpossibly be involved in the regulation or secretion ofendogenous ouabain (Liu et al. 2008). Consequently,it is possible that ouabain could reach the cytosoland perinuclear region of Ma104 cells through aspecific transporter. However, nothing is knownabout a similar transport system neither in MDCKor Ma104 cells, and this matter needs furtherexamination to be fully understood.
It is not known so far which of the effects ofouabain or ouabagenin could be attributable toalterations in membrane permeability between MDCKand Ma104 cells. This is a very difficult subject toaddress, since transport of compounds in culturedcells can fluctuate owing to many circumstances, suchas culture conditions, the density and age of themonolayer, and also stage of differentiation (Volpe2008). Another issue that remains unsolved is whyouabagenin diverged in actions in these cell lines,either in terms of cell viability or inhibition of Na-K-ATPase. MDCK cellular viability decreased in aconcentration-dependent fashion with this molecule,though Ma104 cells were almost indifferent to theincrease of its concentration. Moreover, the ability ofOBG to inhibit Na-K-ATPase activity was lower thanof that observed for OUA in MDCK cells, but theinhibition curves of both molecules were identical inMa104 cells. Although the experiments analyzing thedisplacement of OUA and OBG from their bindingsites using the fluorescent OUABDP indicated thatOBG possesses distinct potentials to bind to MDCKand Ma104 cells (Fig. 4; Table 1), it was not possibleto perfectly comprehend how OBG induces its effectsin both cell lines with the results obtained in thepresent study.
Since the discovery of an endogenous analog ofouabain in human tissues and plasma, several studieshave been performed with the purpose of determiningthe physiological role of this glycoside. The firstobserved ouabain function was the inhibition of Na-K-ATPase, and it was initially suggested that endog-enous ouabain could act as an electrolyte-controllingmolecule, governing Na-K-ATPase activity (Hamlynet al. 1991). However, subsequent studies observedouabain actions that did not fully correlate with Na-K-ATPase impairment, such as its hypertensinogenicactivity (Manunta et al. 2000, 2001; Saunders andScheiner-Bobis 2004) or the activation of cellsignaling pathways related with cell growth ordifferentiation (Haas et al. 2000; Liu et al. 2000).Although using concentrations of ouabain above thephysiological levels found in human plasma, thepresent work highlights a new element that couldaccount for the understanding of cellular responses toouabain, which corroborates previous observationsthat some cells present a second binding site forouabain (Ward et al. 2002; Liu et al. 2008): thepossibility that this hormone induces at least part ofits effects by entering the cells and reaching thecytoplasm, by means of binding to an intracellularsteroid or, alternatively, to its own receptor.
Acknowledgments This work was supported by grants fromCoordenação de Aperfeiçoamento de Pessoal de Nível Superior(CAPES), Fundação Carlos Chagas Filho de Amparo àPesquisa do Estado do Rio de Janeiro (FAPERJ), Programade Oncobiologia (FECD/FAF/ONCO II), PRONEX, and Con-selho Nacional de Desenvolvimento Científico e Tecnológico(CNPq). The authors are indebted to Dr. Elizabeth Giestal deAraujo for the kind donation of monodansylcadaverine.
References
Ackermann U, Geering K. Mutual dependence of Na, K-ATPase alpha- and beta-subunits for correct posttransla-tional processing and intracellular transport. FEBS Lett.1990;269:105–8.
Aizman O, Uhlen P, Lal M, Brismar H, Aperia A. Ouabain, asteroid hormone that signals with slow calcium oscilla-tions. Proc Natl Acad Sci USA. 2001;98:13420–4.
Akimova OA, Lopina OD, Hamet P, Orlov SN. Search forintermediates of Na+, K+-ATPase-mediated [Na+]i/[K+]i-independent death signaling triggered by cardiotonicsteroids. Pathophysiology. 2005a;12:125–35.
Akimova OA, Bagrov AY, Lopina OD, Kamernitsky AV,Tremblay J, Hamet P, et al. Cardiotonic steroids differen-tially affect intracellular Na+ and [Na+]i/[K+]i-indepen-
Cell Biol Toxicol (2010) 26:201–213 211

dent signaling in C7-MDCK cells. J Biol Chem.2005b;280:832–9.
Aperia A. New roles for an old enzyme: Na, K-ATPase emergesas an interesting drug target. J Intern Med. 2007;261:44–52.
Barwe SP, Anilkumar G, Moon SY, Zheng Y, Whitelegge JP,Rajasekaran SA, et al. Novel role for Na, K-ATPase inphosphatidylinositol 3-kinase signaling and suppression ofcell motility. Mol Biol Cell. 2005;16:1082–94.
Capella LS, Gefé M, Silva EF, Morales MM, Affonso-MitidieriO, Lopes AG, et al. Reduced glutathione protect cells fromouabain toxicity. Biochim Biophys Acta. 2001;1526:293–300.
Contreras RG, Lazaro A, Mujica A, Gonzalez-Mariscal L,Valdes J, Garcia-Villegas MR, et al. Ouabain resistance ofthe epithelial cell line (Ma104) is not due to lack ofaffinity of its pumps for the drug. J Membr Biol.1995a;145:295–303.
Contreras RG, Lazaro A, Bolivar JJ, Flores-Maldonado C,Sánchez SH, Gonzalez-Mariscal L, et al. A novel type ofcell–cell cooperation between epithelial cells. J MembraneBiol. 1995b;145:305–10.
Contreras RG, Shoshani L, Flores-Maldonado C, Lazaro A,Cereijido M. Relationship between Na(+), K(+)-ATPaseand cell attachment. J Cell Sci. 1999;112:4223–32.
Contreras RG, Flores-Maldonado C, Lázaro A, Shoshani L,Flores-Benitez D, Larré I, et al. Ouabain binding to Na+,K+-ATPase relaxes cell attachment and sends a specificsignal (NACos) to the nucleus. J Membr Biol. 2004;198(3):147–58.
Geering K, Theulaz I, Verrey F, Häuptle MT, Rossier BC. Arole for the beta-subunit in the expression of functional Na+-K+-ATPase in Xenopus oocytes. Am J Physiol.1989;257:C851–8.
Grubmeyer C, Penefsky HS. The presence of two hydrolyticsites on beef heart mitochondrial adenosine triphosphatase.J Biol Chem. 1981;256:3718–27.
Haas M, Askari A, Xie Z. Involvement of Src and epidermalgrowth factor receptor in the signal-transducing functionof Na+/K+-ATPase. J Biol Chem. 2000;275:27832–7.
Hallaq H, Heller M, Panet R, Eilam Y. Binding properties andbiological effects of oxidized-ouabain on culturedneonatal-rat cardiac myocytes. Implications on the mech-anism of action of the digitalis-glycosides. BiochemPharmacol. 1991;41:509–19.
Hamlyn JM, Blaustein MP, Bova S, DuCharme DW, HarrisDW, Mandel F, et al. Identification and characterization ofa ouabain-like compound from human plasma. Proc NatlAcad Sci USA. 1991;88:6259–63.
Hamrick M, Renaud KJ, Fambrough DM. Assembly of theextracellular domain of the Na,K-ATPase beta subunitwith the alpha subunit. Analysis of beta subunit chimerasand carboxyl-terminal deletions. J Biol Chem.1993;268:24367–73.
Hansen O. No evidence for a role in signal-transduction of Na+/K+-ATPase. Eur J Biochem. 2003;270:1916–9.
Ho CK, Chen SM, Wang SY. Ouabain resistance of a humantrophoblast cell line is not related to its reactivity toouabain. Eur J Cell Biol. 2000;79:765–70.
Jorgensen PL, Skou JC. Purification and characterization of(Na+ + K+)-ATPase. I. The influence of detergents on the
activity of (Na+ + K+)-ATPase in preparations from theouter medulla of rabbit kidney. Biochim Biophys Acta.1971;233:366–80.
Kajikawa M, Fujimoto S, Tsuura Y, Mukai E, Takeda T,Hamamoto Y, et al. Ouabain suppresses glucose-inducedmitochondrial ATP production and insulin release bygenerating reactive oxygen species in pancreatic islets.Diabetes. 2002;51:2522–9.
Larre I, Ponce A, Fiorentino R, Shoshani L, Contreras RG,Cereijido M. Contacts and cooperation between cellsdepend on the hormone ouabain. Proc Natl Acad SciUSA. 2006;103:10911–6.
Liu J, Tian J, Haas M, Shapiro JI, Askari A, Xie Z. Ouabaininteraction with cardiac Na+/K+-ATPase initiates signalcascades independent of changes in intracellular Na+ andCa2+ concentrations. J Biol Chem. 2000;275:27838–44.
Liu Y, Ward SC, Hamilton BP, Hamlyn JM. Novel ouabainreceptors in bovine adrenal cortex: evidence for ouabaintransport. FASEB J. 2008;22:936.19.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Proteinmeasurement with the Folin phenol reagent. J Biol Chem.1951;193(1):265–75.
Manunta P, Stella P, Rivera R, Ciurlino D, Cusi D, Ferrandi M,et al. Left ventricular mass, stroke volume and ouabain-like factor in essential hypertension. Hypertension.1999;34:450–6.
Manunta P, Hamilton J, Rogowski AC, Hamilton BP, HamlynJM. Chronic hypertension induced by ouabain but notdigoxin in the rat: antihypertensive effect of digoxin anddigitoxin. Hypertens Res. 2000;23:S77–85.
Manunta P, Hamilton BP, Hamlyn JM. Structure–activityrelationships for the hypertensinogenic activity of ouabain:role of the sugar and lactone ring. Hypertension.2001;37:472–7.
Mosmann T. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicityassays. J Immunol Methods. 1983;65:55–63.
Noguchi S, Higashi K, Kawamura M. A possible role of thebeta-subunit of (Na, K)-ATPase in facilitating correctassembly of the alpha-subunit into the membrane. J BiolChem. 1990;265:15991–5.
Orlov SN, Thorin-Trescases N, Kotelevtsev SV, Tremblay J,Hamet P. Inversion of the intracellular Na+/K+ ratioblocks apoptosis in vascular smooth muscle at a siteupstream of caspase-3. J Biol Chem. 1999;274:16545–52.
Orlov SN, Taurin S, Thorin-Trescases N, Dulin NO,Tremblay J, Hamet P. Inversion of the intracellular Na(+)/K(+) ratio blocks apoptosis in vascular smoothmuscle cells by induction of RNA synthesis. Hyperten-sion. 2000;35:1062–8.
Ozaki H, Nagase H, Urakawa N. Involvement of the sugarmoiety in the inhibitory action of the cardiac glycosides onthe palytoxin-induced responses in vascular smoothmuscles. J Pharmacol Exp Ther. 1984;231:153–8.
Pchejetski D, Taurin S, Sarkissian SD, Lopina OD, PshezhetskyAV, Tremblay J, et al. Inhibition of Na+, K+-ATPase byouabain triggers epithelial cell death independently ofinversion of the [Na+]i/[K+]i ratio. Biochem Biophys ResComm. 2003;30:735–44.
Rajasekaran SA, Palmer LG, Moon SY, Peralta-Soler A,Apodaca GL, Harper JF, et al. Na, K-ATPase activity is
212 Cell Biol Toxicol (2010) 26:201–213

required for formation of tight junctions, desmosomes, andinduction of polarity in epithelial cells. Mol Biol Cell.2001;12:3717–32.
Rajasekaran SA, Hu J, Gopal J, Gallemore R, Ryazantsev S,Bok D, et al. Na, K-ATPase inhibition alters tight junctionstructure and permeability in human retinal pigmentepithelial cells. Am J Physiol Cell Physiol. 2003;284:C1497–507.
Saunders R, Scheiner-Bobis G. Ouabain stimulates endothe-lin release and expression in human endothelial cellswithout inhibiting the sodium pump. Eur J Biochem.2004;271:1054–62.
Scharfstein J, Schmitz V, Morandi V, Capella MAM, Lima AP,Morrot A, et al. Host cell invasion by Trypanosoma cruziis potentiated by activation of bradykinin B 2 receptors. JExp Med. 2000;192:1289–300.
Schoner W. Endogenous cardiac glycosides, a new class ofsteroid hormones. Eur J Biochem. 2002;269:2440–8.
Shoshani L, Contreras RG, Roldán ML, Moreno J, Lázaro A,Balda MS, et al. The polarized expression of Na+, K+-ATPase in epithelia depends on the association betweenbeta-subunits located in neighboring cells. Mol Biol Cell.2005;16:1071–81.
Vagin O, Sachs G, Tokhtaeva E. The roles of the Na, K-ATPasebeta 1 subunit in pump sorting and epithelial integrity. JBioenerg Biomembr. 2007;39:367–72.
Valente RC, Capella LS, Monteiro RQ, Rumjanek VM, LopesAG, Capella MA. Mechanisms of ouabain toxicity.FASEB J. 2003;17:1700–2.
Valente RC, Capella LS, Nascimento CR, Lopes AG, CapellaMA. Modulation of multidrug resistance protein (MRP1/
ABCC1) expression: a novel physiological role forouabain. Cell Biol Toxicol. 2007;23:421–7.
Volpe DA. Variability in Caco-2 and MDCK cell-basedintestinal permeability assays. J Pharm Sci. 2008;97(2):712–25.
Wang J, Staessen JA, Messaggio E, Nawrot T, Fagard R,Hamlyn JM, et al. Salt, endogenous ouabain and bloodpressure interactions in the general population. J Hyper-tens. 2003;21:1475–81.
Ward SC, Hamilton BP, Hamlyn JM. Novel receptors forouabain: studies in adrenocortical cells and membranes.Hypertension. 2002;39:536–42.
Yoda A. Structure–activity relationships of cardiotonic steroidsfor the inhibition of sodium- and potassium-dependentadenosine triphosphatase. I. Dissociation rate constants ofvarious enzyme–cardiac glycoside complexes formed inthe presence of magnesium and phosphate. Mol Pharma-col. 1973;9:51–60.
Yoda A. Association and dissociation rate constants of thecomplexes between various cardiac monoglycosides andNa, K-ATPase. Ann N Y Acad Sci. 1974;242:598–616.
Yoda A, Yoda S. Association and dissociation rate constants ofthe complexes between various cardiac aglycones andsodium- and potassium-dependent adenosine triphospha-tase formed in the presence of magnesium and phosphate.Mol Pharmacol. 1977;13:352–61.
Yoda A, Yoda S, Sarrif AM. Structure-activity relationships ofcardiotonic steroids for the inhibition of sodium- andpotassium-dependent adenosine triphosphatase. II. Asso-ciation rate constants of various enzyme–cardiac glycosidecomplexes. Mol Pharmacol. 1973;9:766–73.
Cell Biol Toxicol (2010) 26:201–213 213




















